(Annelida Lamarck, 1802) Тип Аннелиды, Кольчатые черви, Кольчецы, Phylum Annelida Lamarck, 1802 (Segmented worms) 3 класса
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ТИП КОЛЬЧАТЫЕ ЧЕРВИ (ANNELIDA)
К кольчатым червям принадлежат первичные кольчецы, многощетинковые и малощетинковые черви, пиявки и эхиуриды. В типе кольчатых червей насчитывают около 8 тыс. видов. Это наиболее высокоорганизованные представители группы червей. Размеры кольчецов колеблются от долей миллиметра до 2,5 м. Преимущественно это свободноживущие формы. Тело кольчецов подразделено на три части: голова, туловище, состоящее из колец, и анальная лопасть. Такого четкого деления тела на отделы нет у стоящих ниже по своей организации животных.
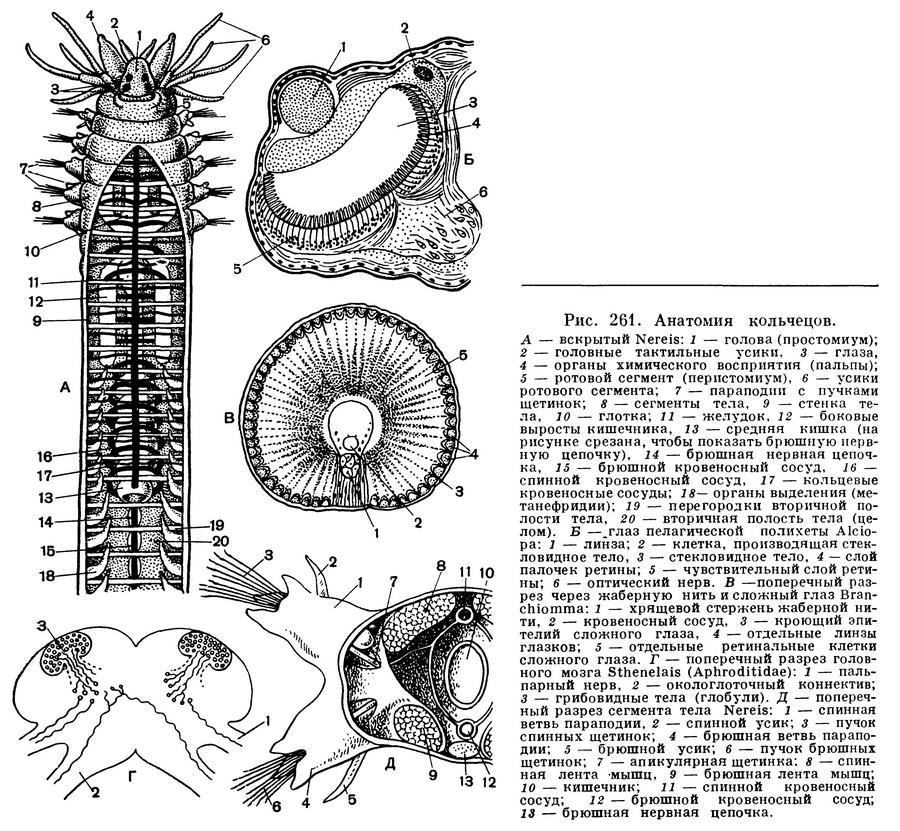
Голова кольчецов снабжена различными органами чувств. У многих кольчецов хорошо развиты глаза. Некоторые имеют особо острое зрение, и их хрусталик способен к аккомодации. Правда, глаза могут быть расположены не только на голове, но и на щупальцах, на теле и на хвосте. У кольчецов развиты и вкусовые ощущения. На голове и щупальцах у многих из них есть особые обонятельные клетки и ресничные ямки, которые воспринимают различные запахи и действие многих химических раздражителей. Хорошо развиты у кольчецов органы слуха, устроенные по типу локаторов. Недавно у морских кольчецов эхиурид открыты органы слуха, очень похожие на органы боковой линии у рыб. С помощью этих органов животное тонко различает малейшие шорохи и звуки, которые в воде слышны гораздо лучше, чем в воздухе.
Тело кольчецов состоит из колец, или сегментов. Число колец может достигать нескольких сотен. Другие кольчецы состоят всего из нескольких сегментов. Каждый сегмент до некоторой степени представляет самостоятельную единицу целого организма. Каждый сегмент включает части жизненно важных систем органов.
Очень характерны для кольчецов особые органы движения. Они располагаются по бокам каждого сегмента и называются параподиями. Слово «параподии» означает «похожие на ноги». Параподии — это лопастевидные выросты тела, из которых наружу торчат пучки щетинок. У некоторых пелагических полихет длина параподии равна диаметру тела. Параподии развиты не у всех кольчецов. Есть они у первичных кольчецов и многощетинковых червей. У малощетинковых остаются только щетинки. Примитивная пиявка акантобделла имеет щетинки. Остальные пиявки обходятся в движении без параподий и щетинок. У эхиурид параподий нет, а щетинки есть только на заднем конце тела.
Параподии, узлы нервной системы, органы выделения, половые железы и, у некоторых полихет, парные карманы кишечника планомерно повторяются в каждом сегменте. Эта внутренняя сегментация совпадает с наружной кольчатостью. Многократное повторение сегментов тела названо греческим словом «метамерия». Метамерия возникла в процессе эволюции в связи с удлинением тела предков кольчецов. Удлинение тела вызвало необходимость многократного повторения сначала органов движения с их мускулатурой и нервной системой, а затем и внутренних органов.
Чрезвычайно характерна для кольчецов сегментированная вторичная полость тела, или целом. Эта полость находится между кишечником и стенкой тела. Полость тела выстлана сплошным слоем эпителиальных клеток, или целотелием. Эти клетки образуют слой, покрывающий кишечник, мышцы и все другие внутренние органы. Полость тела разделена на сегменты поперечными перегородками — диссепиментами. По средней линии тела проходит продольная перегородка — мезентерий, которая делит каждый отсек полости на правую и левую части.
Полость тела заполнена жидкостью, которая по своему химическому составу очень близка к морской воде. Жидкость, заполняющая полость тела, находится в непрерывном движении. Полость тела и полостная жидкость выполняют важные функции. Полостная жидкость (как и всякая жидкость вообще) не сжимается и поэтому служит хорошим «гидравлическим скелетом». Движением полостной жидкости могут переноситься внутри тела кольчецов различные питательные продукты, выделения желез внутренней секреции, а также кислород и углекислый газ, участвующие в процессе дыхания.
Внутренние перегородки защищают организм при тяжелых ранениях и разрывах стенки тела. Например, дождевой червь, разрезанный пополам, не погибает. Перегородки не позволяют полостной жидкости вытекать из тела. Внутренние перегородки кольчецов, таким образом, предохраняют их от гибели. Морские корабли и подводные лодки тоже имеют внутренние герметические перегородки. Если борт пробит, то вода, хлынувшая в пробоину, заливает только один поврежденный отсек. Остальные отсеки, не залитые водой, сохраняют плавучесть поврежденного корабля. Так и у кольчецов нарушение одного сегмента их тела не влечет за собой гибели всего животного. Но не все кольчатые черви имеют хорошо развитые перегородки в полости тела. Например, у эхиурид полость тела не имеет перегородок. Прокол стенки тела эхиуриды может привести ее к гибели. Кроме дыхательной и защитной роли, вторичная полость выполняет роль вместилища для половых продуктов, которые вызревают там, прежде чем выводятся наружу.
Кольчецы, за немногими исключениями, имеют кровеносную систему. Однако сердца у них нет. Стенки крупных сосудов сами сокращаются и проталкивают кровь через тончайшие капилляры. У пиявок функции кровеносной системы и вторичной полости настолько совпадают, что эти две системы совмещаются в единую сеть лакун, по которым течет кровь. У одних кольчецов кровь бесцветная, у других окрашена в зеленый цвет пигментом, который называется хлоркруорин. Часто кольчецы имеют красную кровь, по составу сходную с кровью позвоночных. Красная кровь содержит железо, которое входит в состав пигмента гемоглобина. Некоторые кольчецы, зарываясь в грунт, испытывают острый дефицит кислорода. Поэтому у них кровь приспособлена к тому, чтобы особенно интенсивно связывать кислород. Например, у полихеты Magelona papillicornis развит пигмент гемэритрин, который содержит железа в пять раз больше, чем гемоглобин.
У кольчецов по сравнению с низшими беспозвоночными обмен веществ и дыхание протекают гораздо интенсивнее. У некоторых многощетинковых кольчецов развиваются специальные органы дыхания — жабры. В жабрах разветвляется сеть кровеносных сосудов, и сквозь их стенку кислород проникает в кровь, а затем разносится по всему телу. Жабры могут быть расположены на голове, на параподиях и на хвосте.
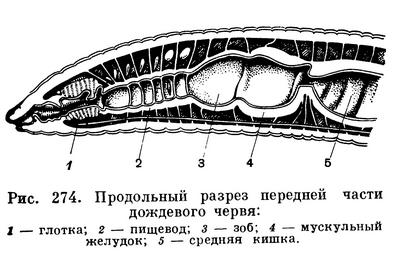
Сквозной кишечник кольчецов состоит из нескольких отделов. Каждый отдел кишечника выполняет свою особую функцию. Рот ведет в глотку. У некоторых кольчецов в глотке располагаются сильные роговые челюсти и зубчики, помогающие крепче схватывать живую добычу. У многих хищных кольчецов глотка служит мощным орудием нападения и защиты. За глоткой следует пищевод. Этот отдел часто снабжен мышечной стенкой. Перистальтические движения мышц медленно проталкивают пищу в следующие отделы. В стенке пищевода располагаются железы, фермент которых служит для первичной переработки пищи. За пищеводом следует средняя кишка. В отдельных случаях бывают развиты зоб и желудок. Стенка средней кишки образована эпителием, очень богатым железистыми клетками, которые вырабатывают пищеварительный фермент. Другие клетки средней кишки всасывают переваренную пищу. У одних кольчецов средняя кишка в виде прямой трубки, у других она изогнута петлями, третьи имеют с боков кишечника метамерные выросты. Задняя кишка заканчивается анальным отверстием.
Специальные органы — метанефридии — служат для выделения жидких продуктов обмена веществ. Часто они служат для выведения наружу половых клеток — сперматозоидов и яйцеклеток. Метанефридии начинаются воронкой в полости тела; от воронки идет извитой канал, который в следующем сегменте открывается наружу. В каждом сегменте располагаются два метанефридия.
Размножаются кольчецы бесполым и половым путем. У водных кольчецов часто встречается бесполое размножение. При этом их длинное тело распадается на несколько частей. Через некоторое время каждая часть восстанавливает голову и хвост. Иногда голова с глазами, щупальцами и мозгом образуется в середине тела червя еще до того, как он разделится на части. В этом случае отделившиеся части уже имеют голову со всеми необходимыми органами чувств. Полихеты и олигохеты сравнительно хорошо восстанавливают утраченные части тела.
Такой способностью не обладают пиявки и эхиуриды. Эти кольчецы утратили сегментированную полость тела. Отчасти поэтому, видимо, способность к бесполому размножению и восстановлению утраченных частей у них отсутствует.
Оплодотворение яиц у морских кольчецов происходит чаще всего вне тела материнского организма. В этом случае самцы и самки одновременно выбрасывают половые клетки в воду, где и происходит оплодотворение.
У морских многощетинковых кольчецов и эхиурид дробление оплодотворенных яиц приводит к развитию личинки, которая нисколько не похожа на взрослых животных и носит название трохофоры. Трохофора короткое время живет в поверхностных слоях воды, а затем оседает на дно и постепенно превращается во взрослый организм.
Пресноводные и наземные кольчецы чаще всего гермафродиты и имеют прямое развитие. Свободной личинки у пресноводных и наземных кольчецов нет. Это связано с тем, что пресная вода имеет солевой состав совсем иного свойства, чем морская. Для развития жизни морская вода более благоприятна. Пресная же вода содержит даже некоторые ядовитые ионы (например, магния) и для развития организмов менее пригодна. Поэтому развитие пресноводных животных почти всегда происходит под прикрытием специальных малопроницаемых оболочек. Еще более плотные оболочки — скорлупки — образуются у яиц наземных кольчецов. Плотные оболочки предохраняют здесь яйцеклетки от механических повреждений и от высыхания под палящими лучами солнца.
Практическое значение кольчатых червей все более возрастает в связи с развитием интенсивности биологических исследований.
У нас в СССР впервые в истории мировой науки осуществлена акклиматизация некоторых беспозвоночных для усиления кормовой базы моря. Например, полихета Nereis, акклиматизированная в Каспийском море, стала важнейшим пищевым объектом осетра и других рыб.
Дождевые черви не только служат наживкой при рыбной ловле и пищей для птиц. Они приносят огромную пользу человеку, разрыхляя почву, делая ее более пористой. Это благоприятствует свободному проникновению воздуха и воды к корням растений и увеличивает урожаи сельскохозяйственных культур. Роясь в земле, черви проглатывают кусочки почвы, измельчают их и выбрасывают на поверхность хорошо перемешанными с органическим веществом. Выносимое на поверхность червями количество грунта поразительно велико. Если бы распределить по всей поверхности суши почву, перепахиваемую дождевыми червями за каждые 10 лет, то получился бы слой плодородной земли толщиной в 5 см.
Пиявки применяются в медицинской практике при гипертонических заболеваниях и угрозе кровоизлияния. Они впускают в кровь вещество гирудин, которое препятствует свертыванию крови и способствует расширению кровеносных сосудов.
Тип кольчецов включает несколько классов. Наиболее примитивными являются морские первичные кольчецы — архианнелиды. Многощетинковые кольчецы и эхиуриды — обитатели моря. Малощетинковые кольчецы и пиявки — в основном обитатели пресных вод и почвы.
КЛАСС ПЕРВИЧНЫЕ КОЛЬЧЕЦЫ (ARCHIANNELIDA)
Архианнелиды — наиболее примитивный класс кольчатых червей. Кольчатость их тела имеет первичный и примитивный характер. Первые признаки кольчатости возникают в поверхностных слоях тела. Процесс возникновения кольчатости исторически, видимо, начинается с органов движения. На удлиненном теле первичных кольчецов возникает несколько ресничных колец, с помощью которых эти организмы плавают в воде или скользят в грунте.
Архианнелиды — чаще всего мелкие кольчецы. Некоторые из них очень малы и не превышают 2—3 мм. Число сегментов, или колец их тела, не превышает 7. Другие состоят из большего числа сегментов и достигают большей длины — до 2—3 см. Архианнелиды живут преимущественно в прибрежной части моря. Одни из них скользят по поверхности грунта или водорослей с помощью ресничек. Другие движутся в промежутках между частицами песка. В этом случае у них могут быть одновременно и ресничные кольца, и параподии со щетинками. Щетинки образуются из ресничных клеток как приспособление к жизни в грунте. Класс архианнелид состоит из нескольких семейств — Dinophilidae, Protodrilidae, Polygordiidae, Saccocirridae, Nerillidae.
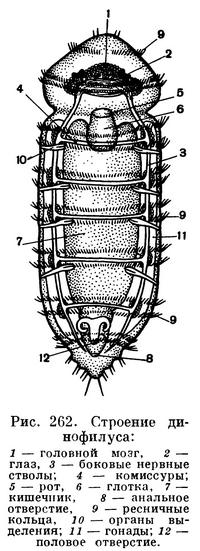
Наиболее примитивны среди первичных кольчецов представители семейства динофилид. Динофилиды очень малы и чаще всего окрашены в оранжевые или красноватые тона. Например, Dinophilus maris albi из Белого моря размером всего около 1 мм. Динофилиды живут среди нитчатых водорослей. Они питаются илом, богатым органическим веществом, а также одноклеточными водорослями, инфузориями и другими мельчайшими организмами.
Тело динофилид опоясано несколькими венчиками ресничек (рис. 262). Два венчика находятся на голове и еще пять располагаются на туловище. Нервная система состоит из головного мозга и двух широко расставленных брюшных нервных стволов. Соответственно пяти ресничным кольцам на каждом из нервных стволов имеются скопления нервных клеток — ганглии. В течение лета самки откладывают несколько десятков яиц, из которых выходят молодые динофилиды.
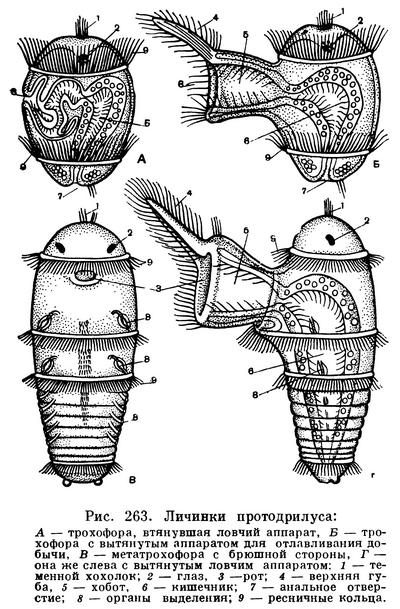
Другое семейство — протодрилид представлено мелкими кольчецами длиной от 1 до 20 мм. Тело их состоит из нескольких десятков сегментов, опоясанных ресничными кольцами. У некоторых видов наряду с ресничными кольцами развиты и щетинки. На голове пара длинных усиков. Протодрилиды живут в песке в прибрежной зоне моря. С помощью ресничной бороздки на брюшной стороне тела они пробираются в свободных промежутках между песчинками и головными усиками вылавливают ил и мелких животных, которыми питаются. Глаз у них нет. Зато имеются специальные сейсмические органы — статоцисты, которые отлично улавливают малейшие шорохи. У Protodrilus статоцисты расположены у основания головных щупалец. Воспринимая шорохи, эти кольчецы могут реагировать на присутствие поблизости жертвы или врага. Своей мускулистой глоткой они раздавливают добычу.
Взрослые протодрилиды откладывают между частицами песка маленькие коконы с несколькими яйцеклетками. Оплодотворенные яйцеклетки дробятся и постепенно превращаются в личинок — трохофор.
Сначала трохофоры имеют только один ресничный пояс. Тело их набито эмбриональным желтком. Позднее появляется второй ресничный пояс, глаза и ротовое отверстие. Когда через некоторое время сформируется сквозной кишечник, трохофоры покидают кокон и уплывают в море. Они имеют размер около 70 мк и не видны невооруженным глазом. Около ротового отверстия у них формируются огромные губы, которые могут выпячиваться далеко вперед и схватывать добычу. Они могут также складываться или совсем втягиваться внутрь (рис. 263). Личинки питаются очень мелкими одноклеточными водорослями и быстро растут. Тело личинок заметно вытягивается, и на нем появляются сначала третье, а затем четвертое ресничные кольца. В это время личинки напоминают взрослых динофилид с неполным числом ресничных колец. Личинки длиной около 0,5 мм оседают на дно, проникают в промежутки между частицами песка и постепенно превращаются во взрослых червячков. Число сегментов постепенно увеличивается.
Рассмотренный здесь цикл индивидуального развития одного из первичных кольчецов показывает возможный путь исторического развития всей группы кольчатых червей. Известно, что в индивидуальном развитии отражается в какой-то мере путь эволюционного развития той же группы животных. Эта закономерность носит название биогенетического закона.
Представители семейства полигордиусов ведут роющий образ жизни в песчаных грунтах. Обычно они живут в море на глубине от 10 до 20 м. Длина полигордиусов около 5 см.
Сильно вытянутое цилиндрическое тело состоит из многочисленных одинаковых сегментов. На голове развиты щупальца и две обонятельные ямки. С помощью этих органов животное воспринимает различные механические и химические раздражения. Глаза отсутствуют.
Если динофилиды и протодрилиды движутся главным образом с помощью ресничек и щетинок, то полигордиусы передвигаются в грунте только с помощью мышц. В стенке их тела хорошо развита продольная мускулатура, а кольцевая отсутствует. Нервная система состоит из головного и брюшного мозга. Головной мозг устроен очень сложно.
Питаются полигордиусы органическими частицами животного и растительного происхождения. Не брезгают они и мелкими животными. Специальной дыхательной системы у полигордиусов нет. Зато хорошо развиты кровеносные сосуды, по которым течет красная кровь.
Личиночное развитие полигордиусов протекает в толще воды. Трохофора описана в каждом учебнике зоологии.
Представители семейства саккоциррусов также обитают в морских песчаных грунтах. В прибрежной зоне Черного моря на каждый квадратный метр песчаного дна приходится в среднем несколько сотен тысяч этих кольчецов. Саккоциррусы имеют тело до 8 см длиной, с большим количеством сегментов. Сегменты с каждой стороны несут по одному пучку простых щетинок. Развитие этих кольчецов проходит с превращением. Трохофора сначала превращается в личинку с небольшим числом сегментов, количество которых постепенно увеличивается.
К семейству нериллид относятся архианнелиды не более 2 мм длиной, с небольшим числом сегментов. Сегменты снабжены пучками щетинок. Один из представителей этого семейства найден в пещерах Швейцарии. Называется он Troglochaetus beranecki, что в переводе означает «пещерный щетинконос беранека». Это исключительный случай приспособления архианнелиды к обитанию в пресных грунтовых водах пещер.
КЛАСС ОЛИГОХЕТЫ, ИЛИ МАЛОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ (OLIGOCHAETA)
Большинство олигохет — обитатели почвы: более крупные из них хорошо всем известны под названием земляных или дождевых червей. Есть, однако, и пресноводные и даже морские олигохеты. Из общего числа (около 3 тыс.) описанных до сих пор видов примерно 400 — обитатели рек, озер, болот и других пресных водоемов, несколько десятков видов встречается в солоноватых водоемах и прибрежной зоне морей, все остальные — обитатели почвы (но некоторые из них, встречающиеся преимущественно по берегам водоемов, отчасти в воде, ведут земноводный образ жизни). В пределах СССР пока зарегистрировано немногим более 300 видов; цифра эта, однако, далеко не отражает действительного положения вещей, так как некоторые семейства, и в частности одно из наиболее богатых видами — энхитреиды, остаются у нас еще почти не изученными.
Среди олигохет есть и карлики и гиганты. Длина тела самых мелких пресноводных измеряется долями миллиметра, так что это в полном смысле слова микроскопические животные; а у наиболее крупных тропических земляных червей она превышает 2,5 м. Но это исключение. Большинство же водных олигохет имеет размеры от нескольких миллиметров до 10—15 см, а почвенных — от полусантиметра до 30—40 см.
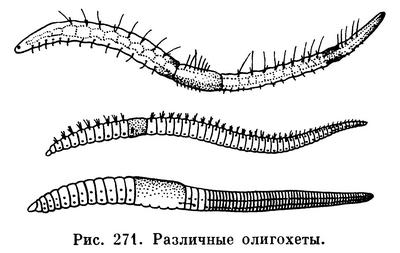
Важнейшей особенностью организации олигохет, как и полихет, является метамерия, т. е. правильная повторяемость органов вдоль оси тела животного. Внешне это выражается в том, что все червеобразное тело поделено перетяжками на отдельные участки — кольца (рис. 271), называемые сегментами или сомитами («члениками»). Число сегментов тела у разных видов олигохет различно, от 5—6 до 500—600; в отличие от полихет они никогда не имеют парных выростов — параподий, но несут по 4 пучка щетинок.
На переднем конце тела находится головная, или предротовая, лопасть; у некоторых водных видов она бывает вытянута в более или менее длинный щупальцевидный придаток («хоботок»), у немногих — почти исчезла, у большинства же представляет небольшой округленный выступ впереди рта. Далее следует первый, или ротовой, сегмент, никогда не несущий щетинок; на его нижней передней стороне находится рот. Все остальные сегменты, начиная со второго, как правило, снабжены четырьмя пучками щетинок каждый: двумя спинными (справа и слева) и двумя брюшными. У некоторых видов спинные пучки начинаются не вместе с брюшными, со второго сегмента, а несколько отступя от переднего конца тела — четвертого, пятого, шестого и даже двадцатого сегмента. Как исключение,
имеются виды лишь с брюшными щетинками и даже вовсе без них.
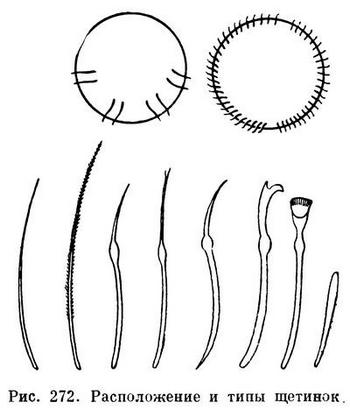
В каждом пучке часто бывает по две более или менее одинаковых щетинки (например, у большинства дождевых червей). У тропических червей рода Pheretima число щетинок на каждом сегменте значительно больше — несколько десятков, а у некоторых видов до 150 — и расположены они не пучками, а в один ряд вокруг всего сегмента. У водных олигохет, как правило, в каждом пучке по нескольку щетинок, от 3—4 до 10—15, причем эти щетинки бывают различной формы: игловидные, крючковидные с простым или двузубчатым концом, веерные, простые и перистые волосовидные и др. (рис. 272).
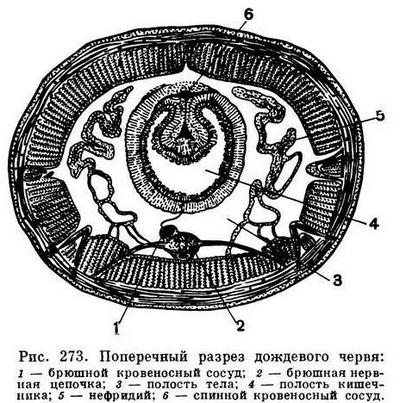
Стенка тела олигохет состоит из пяти основных слоев: тонкой неклеточной кутикулы, выделяемой кожей, кожного эпителия, состоящего из одного слоя клеток, двух слоев — мускулатуры наружного кольцевого и внутреннего продольного и, наконец, внутреннего слоя целомического эпителия, ограничивающего вторичную полость тела, или целом, в которой располагаются внутренние органы. Эта полость, заполненная более или менее прозрачной жидкостью, содержащей взвешенные в ней клетки, не тянется непрерывно вдоль всего тела, а разделена тонкими мускульными перегородками — диссепиментами — на отдельные отсеки, соответствующие наружной сегментации. Таким образом, в каждом сегменте находится свой обособленный участок полости тела, а в нем — пара своих выделительных органов — нефридиев, один справа, другой слева (рис. 273). Имеется замкнутая кровеносная система: продольные сосуды идут вдоль всего тела, связывающие их кольцевые сосуды имеются в каждом сегменте. Пищеварительный канал проходит вдоль всего тела от ротового отверстия до анального; обычно он подразделяется на отделы: ротовая полость, глотка, пищевод, иногда зоб, один или несколько желудков, средняя кишка (самая длинная часть), задняя кишка (рис. 274).
В каждом сегменте, в брюшной его части, находится двойной нервный узел (состоящий из сближенных и почти слившихся правого и левого) с отходящими от него нервами. Узлы всех сегментов соединяются продольными нервными тяжами — коннективами — в единую брюшную нервную цепочку. В головной лопасти или несколько позади нее, над глоткой, находится головной мозг (надглоточный ганглий), соединенный с первым узлом брюшной цепочки — подглоточным — двумя тяжами, охватывающими глотку, — окологлоточными коннективами. Глаз у большинства олигохет нет, они имеются только у части видов одного семейства водных олигохет — наидид; однако, как правило, олигохеты реагируют на освещение, что связано с наличием в коже, особенно на головном конце, особых светочувствительных клеток — фоторецепторов. Дыхание у подавляющего большинства кожное, но у некоторых пресноводных олигохет имеются жабры.
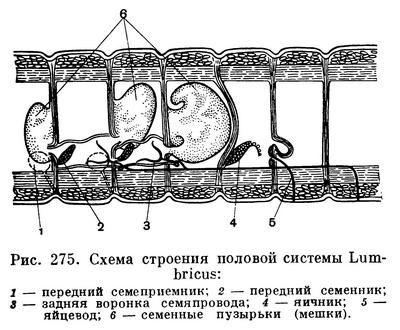
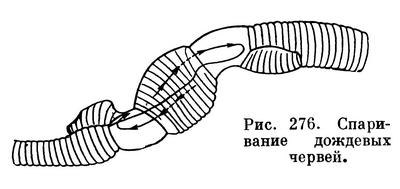
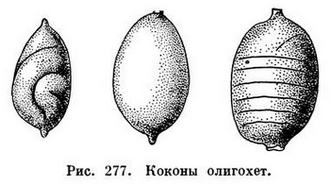
Все олигохеты гермафродиты, т. е. у каждого взрослого червя имеются одновременно и мужская и женская половые системы, строение которых довольно сложно (рис. 275). Оплодотворению предшествует спаривание, во время которого сперматозоиды каждого из двух червей переходят в семеприемники другого (рис. 276). После этого черви расходятся. Затем у каждого из них на пояске, представляющем железистое утолщение кожи нескольких определенных сегментов (у разных олигохет различных, но всегда ближе к головному концу), выделяется кокон, первоначально имеющий форму бочонка (или муфты), открытого с обоих концов. Образовавшийся кокон сокращениями мускулатуры тела сдвигается постепенно вперед и наконец сбрасывается «через голову», после чего концы его смыкаются и он принимает форму лимона (рис. 277). Но прежде чем кокон будет сброшен, пока он медленно скользит от пояска к головному концу, в него откладываются яйца (когда он продвигается над отверстиями яйцеводов), а затем из семеприемников поступают сперматозоиды, полученные при спаривании от другого червя. Таким образом, оплодотворение яиц происходит не в теле червя и не во внешней среде, а в коконе, внутри которого происходит и дальнейшее развитие. Из кокона выходят маленькие червячки, в общем уже похожие на взрослых (рис. 278). У некоторых олигохет описано самооплодотворение, а другие способны к партеногенезу, т. е. девственному, без оплодотворения, развитию отложенных яиц.
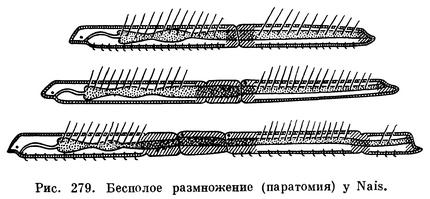
Кроме полового, у олигохет наблюдается и бесполое, вегетативное, размножение, но оно свойственно представителям только некоторых семейств водных червей. Бесполое размножение может происходить либо путем архитомии, либо путем паратомии. При архитомии тело червя самопроизвольно (иногда в результате внешнего раздражения) распадается на два или большее количество кусков — фрагментов, после чего у каждого куска развиваются недостающие части и он постепенно превращается в целого червя.
При паратомии (рис. 279) на теле червя, на некотором определенном расстоянии от головного конца намечается перетяжка, подразделяющая его на две части; после этого задняя часть, не отделяясь сначала от передней, образует свой головной конец, а передняя — свой задний. Теперь два образовавшихся таким образом червя могут разделиться окончательно, но часто, прежде чем это произойдет, каждый из них опять начинает делиться. Таким образом возникают «цепочки», состоящие из четырех-пяти, а иногда и большего числа особей.
Деление класса олигохет на отряды еще окончательно не установилось; мы рассмотрим последовательно важнейшие семейства.
КЛАСС ПИЯВКИ (HIRUDINEA)
Пиявки ... Это слово обычно вызывает неприятное чувство: воображение рисует длинных, темных червей, живущих в болотистых водоемах, нападающих на человека и сосущих его кровь. Многие люди знают только одну пиявку — медицинскую, применяемую при лечении некоторых заболеваний, нередко очень тяжелых. Между тем на земном шаре насчитывается около 400 видов пиявок, их строение разнообразно, они обитают не только в болотистых водоемах, но и в реках, озерах, горных ручьях и даже в морях и океанах. Пиявки-кровососы действительно составляют большинство этого класса типа кольчецов, но живут они за счет разных животных (представителей всех классов позвоночных, мягкотелых, ракообразных, водных насекомых, червей и др.), а не только млекопитающих и человека. И совсем уж мало кому известно, что многие виды пиявок не могут сосать кровь, а принадлежат к числу хищников, которые целиком или по частям заглатывают мелких животных.
Правда, хищные формы произошли от кровососущих и сохранили основные признаки своих предков, но по характеру питания они коренным образом отличаются от них. Что касается пиявок, сосущих кровь млекопитающих и человека, то в тропиках их изрядное количество, но в нашей стране таких видов всего два-три (из 50 пресноводных видов) и распространены они преимущественно в южных районах.
Принадлежность пиявок к типу кольчатых червей не вызывает никаких сомнений. Тело их сегментировано, центральная нервная система состоит из головных узлов, окологлоточных тяжей и брюшной цепочки; в кожно-муокульном мешке есть те же основные элементы, что и у других кольчецов, перенос веществ осуществляется при помощи кровеносной системы, органы выделения — метанефридии, кишечник сквозной, заканчивающийся анальным отверстием, мускулистый, обильно снабженный кровеносными сосудами, и т. д. В то же время для пиявок характерны многие особенности, которые позволяют легко отличить их от других групп типа.
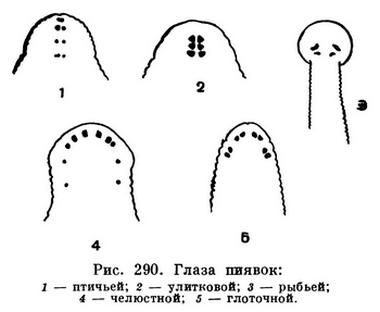
В связи с необходимостью прикрепляться к телу других животных для сосания крови у пиявок развились две присоски (передняя, окружающая рот, и задняя), тело стало более или менее уплощенным. Щетинок, за одним исключением, нет, так как ползают эти черви при помощи присосок. Число сегментов, или сомитов, в отличие от других кольчецов, постоянное и у всех видов, за исключением одного, равно 33, из которых последние семь образуют заднюю присоску. Сравнительно небольшое количество сегментов, вероятно, уменьшало гибкость тела, и у пиявок возникла очень характерная для них вторичная кольчатость тела: сомиты подразделяются на определенное для каждой группы видов число колец. У медицинской и других челюстных пиявок в сомите пять колец, у улитковой и подавляющего большинства видов семейства плоских пиявок — три и т. д. Вторичная кольчатость затрагивает только наружные покровы и на внутренние органы не распространяется. Центральным кольцом считается то, которое несет узел брюшной нервной цепочки.
Среди пиявок почти нет настоящих паразитов, т. е. таких видов, которые постоянно обитают на теле хозяина или внутри его, что всегда влечет за собой упрощение организации паразита. Наоборот, большинство их ведет активный образ жизни в поисках своих жертв, сопротивление которых они должны преодолеть. Поэтому у пиявок хорошо развита нервная система, и у всех червей, живущих на свету, имеются глаза, число и расположение которых характерны для групп видов или даже для отдельных видов. Тело очень мускулисто, и в этом отношении пиявки превосходят всех червей. Так, например, у дождевых червей мускулатура составляет 29,7% объема всего тела, а у пиявок — до 65,5%. Многие виды хорошо плавают. Промежутки между органами заполнены соединительной тканью, и полость тела (вторичная), характерная для всех кольчецов, у пиявок отсутствует. Тело их очень плотное, с трудом поддающееся разрыву. Кровеносная система, прекрасно развитая, претерпела существенные изменения. Изменились и метанефридии, лишившиеся в связи с отсутствием полости тела воронок.
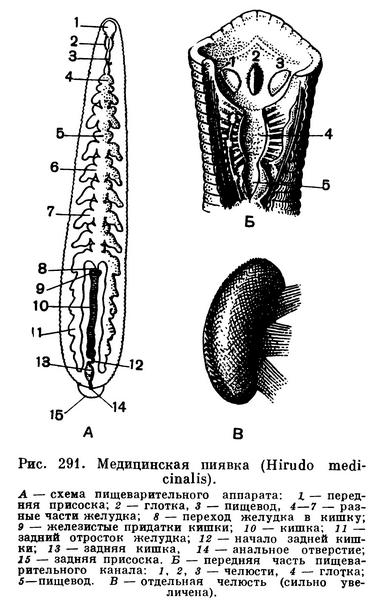
Существенным изменениям подвергся пищеварительный аппарат. Все пиявки, за исключением одного древнего вида, делятся на два отряда: хоботных и челюстных (бесхоботных). У первых в передней части пищеварительной трубки развился мускулистый хобот, у вторых — челюсти (их обычно три), усаженные зубчиками. При помощи хобота или челюстей кровососущие виды повреждают кожу или слизистые оболочки своих жертв. У хищных челюстных пиявок, целиком заглатывающих свою добычу, челюсти уменьшаются или даже совсем исчезают. За глоткой, служащей для сосания крови, и коротким пищеводом идет желудок, объем которого у кровососущих видов сильно увеличен благодаря парным боковым отросткам. У хищных видов отростки желудка полностью или частично исчезают. Усвоение пищи совершается в идущей за желудком кишке, которая у многих пиявок тоже имеет отростки. Кал удаляется через заднюю кишку и анальное отверстие, лежащее на спинной стороне, у задней присоски.
Хорошо известно, что после укусов пиявок раны долго кровоточат. Это объясняется тем, что в раны из слюнных желез, открывающихся в ротовую полость пиявок, попадает особое белковое вещество гирудин (от греческого слова «гирудо» — пиявка), которое препятствует свертыванию крови. Если бы гирудин не выделялся, то быстро образовались бы кровяные сгустки (тромбы) и сосание крови становилось бы невозможным. Благодаря гирудину и другим веществам, выделяемым слюнными железами, кровь месяцами сохраняется в желудке пиявок в жидком состоянии, не подвергаясь гниению.
Все пиявки гермафродиты (у взрослых червей развиты и мужские и женские половые органы) и размножаются только половым путем. На брюшной стороне этих червей, выше средней части тела, довольно хорошо заметны два половых отверстия: переднее, более крупное — мужское, заднее — женское. Расстояние между этими отверстиями, измеряемое количеством колец, у различных видов неодинаковое и имеет значение для определения видов пиявок. Оплодотворение, т. е. слияние живчиков и яйцеклеток, происходит внутри организма. Семя передается при половом акте двумя способами. У одних видов (в том числе у медицинской) оно вводится в женское половое отверстие при помощи совокупительного органа, имеющего вид тонкой нити, а у остальных живчики выделяются в особых мешочках (сперматофорах), которые прикрепляются к коже другой пиявки в разных местах. Живчики через ранки, образующиеся на коже после прикрепления сперматофоров, проникают внутрь тела, отыскивают зрелые яйцеклетки и оплодотворяют их.
Оплодотворенные яйца выделяются в коконах, строение которых будет описано дальше. У половозрелых пиявок, как и у малощетинковых червей, в области половых органов на коже образуется «поясок», часто хорошо заметный. Из выделений желез пояска образуются стенки коконов. По окончании развития, которое обычно длится несколько недель, из коконов выходят маленькие черви, в основном похожие на взрослых.
Пиявки распространены во всех частях света. Каждая зоогеографическая область характеризуется своим составом видов этих червей. Видов, обитающих в двух или более областях, очень мало. Морские пиявки разделяются на ряд групп, из которых каждая характерна для определенного района Мирового океана и прилегающих к нему морей. В Черном море пиявок нет совсем, так как его соленость (в два раза меньше солености океана) недостаточна для настоящих морских пиявок и слишком велика для видов, обитающих в пресных и солоноватых водах.
Пиявки имеют практическое значение, как полезное, так и вредное, о чем вкратце будет рассказано при описании отдельных видов.
ПОДКЛАСС ДРЕВНИЕ ПИЯВКИ (ARCHIHIRUDINEA)
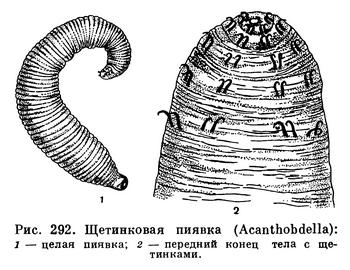
В сороковых годах прошлого столетия выдающийся русский натуралист А. Ф. Миддендорф во время путешествия по Енисею собрал с пеледи, или сырка (Coregonus peled — из сигов, семейство лососевых рыб), странных червей, разъедавших мягкие части спинного плавника. Уже поверхностное изучение этих червей, проведенное известным зоологом Э. Грубе, показало, что в их организации удивительным образом сочетаются признаки малощетинковых червей и пиявок. На переднем конце их тела имеются щетинки, которые выполняют роль передней присоски (отсутствующей у этих червей) и вместе с несовершенной задней присоской служат для прикрепления к телу хозяина. Грубе отнес описанных им червей к новому род, у Acanthobdella, что значит «вооруженная пиявка», а видовое название дал по имени рыб, с которых они были сняты. Впоследствии акантобделла, или щетинковая пиявка, была обстоятельно изучена известным русским зоологом Н. А. Ливановым, который выяснил, что во внутреннем строении ее тоже наблюдается сочетание признаков малощетинковых червей и пиявок, но признаки последних преобладают, и акантобделла была отнесена к особой, низшей группе класса пиявок. Сейчас щетинковых пиявок выделяют в особый подкласс древних пиявок, существование которых является блестящим подтверждением эволюционной теории.
Длина акантобделлы обычно не превышает 30 мм, а толщина — 3 мм. Цвет тела серый со слабым мясо-красным оттенком. Часто на теле заметны темные поперечные полоски. Эта пиявка распространена на крайнем севере Европы и Азии, от Норвегии до Колымы; самое южное ее местонахождение — Онежское озеро. Паразитирует щетинковая пиявка на разных лососевых рыбах, а также на хариусах и питается не только кровью, но и размягченными наружными тканями хозяев и, следовательно, еще не является специализированным кровососом. На каждой рыбе обычно обитает несколько акантобделл, которые причиняют существенный вред своему хозяину. Они появляются на рыбах поздней весной или в начале лета, когда вес их равен всего 5—10 мг; и в конце осени или в начале зимы достигают половой зрелости и веса больше 200 мг. Затем акантобделлы покидают хозяев и, по-видимому, размножаются и развиваются на мелких местах, заросших подводной растительностью. Но никто еще не ловил акантобделл в свободном состоянии и не наблюдал, как происходит размножение и развитие этих пиявок, хотя их удавалось содержать некоторое время в аквариумах. Можно только утверждать, что жизненный цикл этих интересных червей продолжается около года.
ПОДКЛАСС НАСТОЯЩИЕ ПИЯВКИ (EUHIRUDINEA)
В этот подкласс входят все виды пиявок, кроме щетинковой. Передняя присоска у них всегда хорошо развита, щетинок нет; все особенности, отличающие пиявок от малощетинковых червей, хорошо выражены. Подкласс делится на два отряда: отряд хоботных и отряд челюстных, или бесхоботных.
ОТРЯД ХОБОТНЫЕ ПИЯВКИ (RHYNCHOBDELLAE)
Название отряда говорит о главной особенности входящих в его состав видов: все они имеют хобот. Заметить у живой пиявки хобот очень трудно, у убитых червей он иногда торчит изо рта. Отряд в свою очередь подразделяется на два резко отличающихся между собой семейства: семейство плоских пиявок и семейство рыбьих пиявок.
ОТРЯД ЧЕЛЮСТНЫЕ, ИЛИ БЕСХОБОТНЫЕ, ПИЯВКИ (GNATHOBDELLEA, ARHYNCHOBDELLEA)
У видов этого отряда нет хобота, а развиты челюсти, которые у хищных форм менее развиты или совсем рудиментарны.
КЛАСС МНОГОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ (POLYCHAETA)
Полихеты названы так потому, что параподии у них включают пучки многочисленных щетинок.
По-гречески poly означает много, chaeta — щетинка. Полихеты — типичные морские животные и очень редко встречаются в пресной воде.
Всего насчитывается более 5 тыс. видов полихет. Основная масса их населяет соленые моря и океаны. Сравнительно немногие приспособились к жизни в солоноватых водоемах, к которым относятся, например, Азовское и Каспийское моря. Встречаются полихеты в солоноватой воде среди мангровых зарослей на побережьях Индии и Малайских островов. Еще меньше полихет в пресных водах. Такова, например, Manayunkia, обитающая в озерах Таймыра, реках Северной Америки, в озере Байкал. В пресной воде пещер Герцеговины обнаружена полихета Merifugia из семейства Serpulidae. Это единственный случай приспособления полихеты к пещерному существованию. В исключительных случаях полихеты приспосабливаются к наземному существованию. Известен один вид полихеты Lycastopsis catarractarum из семейства Nereidae, которая живет в тропиках, за многие километры от морского берега, среди пальм на плантациях бананов и какао.
Она питается гниющими листьями и сладкими плодами.
Как правило, полихеты живут свободно. Только в некоторых случаях они связывают место своего обитания с другими животными и делят с ними пищу. Таких партнеров называют сотрапезниками или комменсалами. Полихеты-комменсалы живут внутри губок, где находят дом и пищу, в раковинах раков-отшельников или на морских звездах.
В исключительных случаях полихеты паразитируют на рыбах. Например, Ichthyotomus sanguinarius, который ртом прикрепляется к плавникам морских угрей, путешествует с ними через Атлантический океан и сосет их кровь.
Морские полихеты встречаются во всех частях и на всех глубинах Мирового океана. Они обитают на дне, в толще грунта и в толще воды. Полихеты особенно обильны в прибрежной зоне моря и немногочисленны на больших глубинах. На больших глубинах обитают особые глубоководные (абиссальные) виды.
Наиболее богата и разнообразна фауна полихет в прибрежной зоне тропических морей. Но и в северных морях их довольно много. Иногда они встречаются большими скоплениями. Например, в Баренцевом море плотность поселения полихеты Maldane sarsi достигает 90 тыс. экземпляров на каждом квадратном метре морского дна. На больших площадях морского дна полихеты образуют живые ковры удивительных узоров, переливающиеся всеми цветами радуги.
Многие полихеты живут среди морских водорослей. Другие прячутся в укромных уголках среди зарослей коралловых рифов. Немалое количество полихет зарывается в песок или в мягкий ил. Очень часто полихеты строят себе роговые, песчаные или известковые трубки. Некоторые способны жить в очень трудных условиях. Полихета Capitella capitata, например, обитает в самых грязных гаванях, где нефть и мазут плавают на поверхности воды, покрывают все дно, все подводные предметы. В таких гаванях гибнут от нефти рыбы, тюлени и даже птицы. Только у берегов Англии погибает от нефти ежегодно четверть миллиона водоплавающих птиц. Трудно представить себе, как Capitella может существовать в таких неестественных условиях. Другая группа полихет приспособлена к обитанию в толще воды. Таких животных называют пелагическими. Весь свой жизненный цикл они совершают вдали от берегов и переносятся на большие расстояния морскими течениями. Пелагических полихет, например, много в теплом течении Гольфстрим.
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ТИП ПОГОНОФОРЫ (POGONOPHORA)
Погонофоры интересны во многих отношениях. Открытие их явилось одним из самых неожиданных событий в современной зоологии. Еще недавно казалось, что время описания новых классов и типов давно миновало, что все группы животных высокого систематического ранга давно хорошо известны. История изучения погонофор опровергла это представление. Этот тип, объединяющий несколько десятков своеобразных морских животных, был установлен лишь в середине нашего столетия, а детально изучен лишь в последние 10 лет.
Погонофоры относятся ко вторичноротым (Deuterostomia) — большой эволюционной ветви животного мира, к которой принадлежат также иглокожие, полухордовые и хордовые (в том числе позвоночные). О родстве погонофор со всеми этими типами особенно убедительно говорит их эмбриональное развитие. Многие примитивные особенности погонофор позволяют по-новому оценивать первые этапы эволюции вторичноротых.
Погонофоры интересны также отсутствием кишечника, который в процессе эволюции исчез у них полностью. Этим они резко отличаются от всех остальных свободноживущих многоклеточных животных (не считая очень примитивных губок). Известные случаи полной утраты пищеварительного канала касаются исключительно паразитических животных (скребни, ленточные черви, некоторые эндопаразитические брюхоногие моллюски и ракообразные). Как только отсутствие кишечника у погонофор было доказано, встал сложный вопрос о том, как они питаются.
Погонофоры интересны своим образом жизни. Это животные, ведущие почти неподвижную жизнь на дне моря внутри длинной защитной трубки. Подобным же образом в трубках обитают многие многощетинковые черви (Polychaeta, Sedentaria), относящиеся к типу кольчатых червей. Одинаковый образ жизни этих двух неродственных друг другу групп животных привел к выработке у них сходных, но возникших независимо приспособлений. Благодаря этому погонофоры и живущие в трубках полихеты приобрели большое внешнее сходство, настолько смущавшее некоторых зоологов, что они ошибочно считали погонофор своеобразно измененными кольчатыми червями (Ливанов и Порфирьева, 1962). Таким образом, погонофоры дают возможность изучить важные вопросы о закономерностях развития у неродственных животных сходных приспособлений (конвергенций) под влиянием одинаковых условий жизни.
Наконец, любопытна и история изучения погонофор. Эти своеобразные обитатели океанических глубин, еще недавно казавшиеся большой зоологической редкостью, в действительности принадлежат к самым обычным элементам морской фауны. Не удивительно поэтому, что погонофоры уже давно попадали в руки ученых. Однако они не привлекали к себе внимания или причислялись к давно известным группам животных. В одном из музеев естественной истории в США многие десятки лет хранились неопознанные погонофоры, собранные еще в прошлом столетии в Тихом океане американскими океанографическими экспедициями. Лишь теперь, когда мы уже знаем, что такое погонофоры, эти уникальные экземпляры «дождались» своих исследователей. Недавно английский зоолог Д. Карлайл (Carlisle) писал: «Когда в 20-х годах английское исследовательское судно «Дискавери II» работало в Антарктике и Субантарктике, одна из обычнейших неприятностей при добывании донных животных с больших глубин заключалась в том, что после 10 или 12 часов работы драга поднималась, забитая массой каких-то волокон. Очень часто мешок драги был настолько заполнен этой противной массой «gubbins», что исключалась возможность получения какого бы то ни было материала, интересного в биологическом отношении, и весь улов после быстрого и грубого осмотра выбрасывался за борт. Это случалось так часто, что биологи на борту судна употребляли шуточное наименование «Gubbinidae» в качестве названия «семейства» для этого досадного материала».
В настоящее время погонофоры найдены почти во всех морях, даже там, где донная фауна хорошо изучена. Так, недавно они обнаружены в Баренцевом море близ Мурманского побережья, в проливе Скагеррак у берегов Швеции, в западной части Атлантического океана, в фиордах Норвегии. Около 15 лет назад мы знали лишь 3 вида погонофор, ныне описано около 100 видов. Судя по тому, что все время в новых местах открываются все новые виды, и учитывая слабую изученность фауны Мирового океана, естественно предположить, что действительное количество видов погонофор выражается четырехзначным числом.
Географическое распространение погонофор имеет некоторые особенности. Они очень обычны в глубоководных районах Тихого, Индийского и Атлантического океанов, более или менее прилегающих к материкам, а также в окраинных морях, широко сообщающихся с океанами. Напротив, в большом удалении от островов и материков на обширных пространствах океанического дна они отсутствуют. Нет их также во внутренних морях, т. е. в морях, изолированных от Мирового океана или связанных с ним лишь сравнительно мелководными проливами. Так, например, очень обильной и разнообразной фауной погонофор отличаются Охотское и Берингово моря, тогда как соседнее с ними Японское море лишено погонофор. Отсутствуют они и в сильно опресненном Балтийском море, а также в Каспии, в Азовском и Черном морях. Недавно погонофоры обнаружены в Средиземном море.
Погонофоры, отфильтровывающие из воды мелкие частицы детрита и планктонные организмы, зависят от количества этого взвешенного в морской воде пищевого материала и от развивающейся в нем бактериальной флоры. Поэтому они наиболее многочисленны именно в тех местах, где имеются более или менее постоянные скопления придонной органической взвеси, зависящей от направления и силы глубинных течений и от рельефа дна. По-видимому, этим и объясняется тот факт, что в морях и глубоководных впадинах фауна погонофор гораздо богаче, чем в очень удаленных от континента открытых пространствах океана.
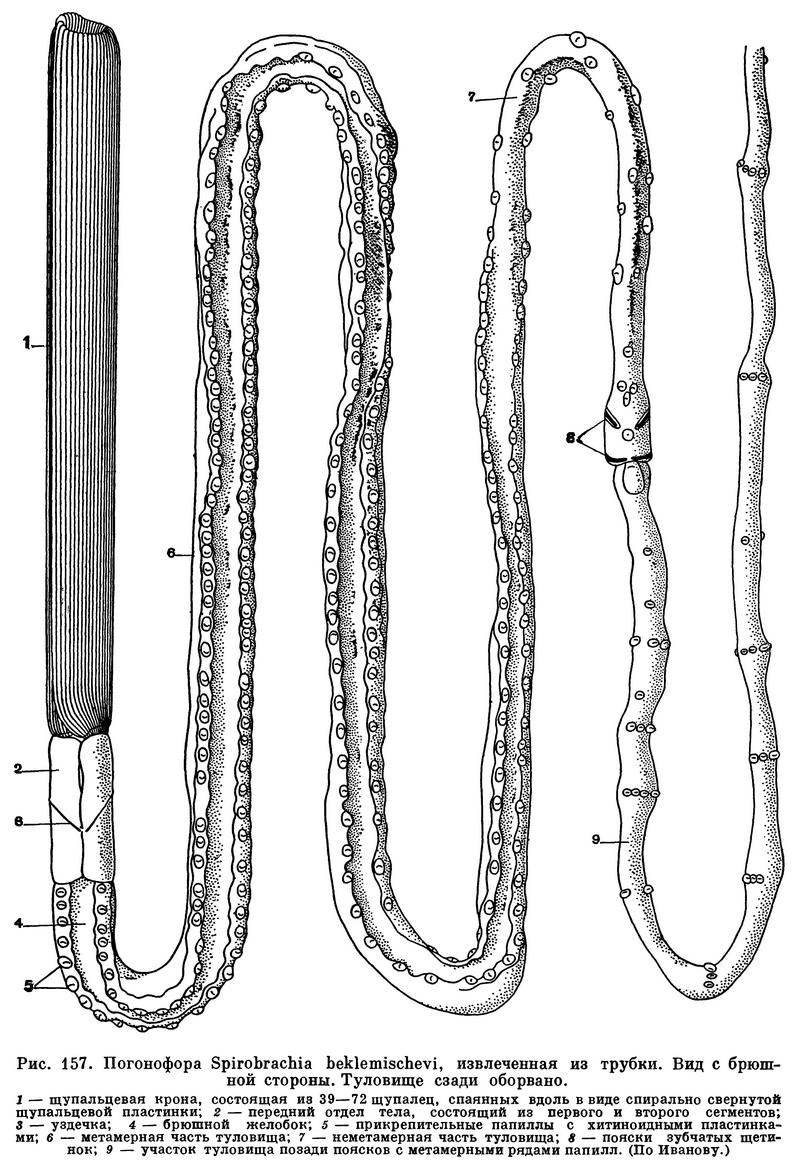
Погонофоры не способны покидать свою трубку, так как не приспособлены к передвижению на открытом месте. Внутри трубки они, однако, могут быстро перемещаться, то высовывая наружу передний конец тела с щупальцами, то уходя глубоко в трубку. В этом отношении они напоминают живущих в трубках многощетинковых червей. По длине трубка всегда в несколько раз превосходит животное и не препятствует таким движениям. Для опоры на внутреннюю поверхность трубки служат хитиноидная уздечка, многочисленные прикрепительные сосочки, или папиллы, зубчатые щетинки поясков и щетинки заднего отдела туловища (см. ниже). Тело погонофор может сильно вытягиваться и сокращаться. Высовывание переднего конца тела из трубки наружу осуществляется путем сильного вытягивания туловища, причем задняя часть последнего в это время прочно прицепляется к стенкам трубки посредством щетинок поясков и заднего отдела. При раздражении или при опасности высунувшееся наружу животное мгновенно исчезает в глубине трубки вследствие простого сокращения мощной продольной мускулатуры, а прочное прикрепление заднего конца облегчает, конечно, это «движение бегства».
Погонофоры относятся к числу самых длинных животных. Их сильно вытянутое тело нитевидно или шнуровидно. Длина его обычно в 100—500 раз превышает ширину. Размеры тела варьируют очень широко. К числу миниатюрных форм относится, например, Siboglinum minutum из Берингова моря, у которого тело имеет не более 5,5 см в длину при ширине менее 0,1 мм. Таких мелких видов очень много, особенно в роде Siboglinum. К крупным погонофорам принадлежат Spirobrachia grandis, длиной не менее 25 см при диаметре туловища до 2,5 мм, и более тонкая Zenkevitchiana longissima, достигающая 36 см в длину. Однако самой большой формой является обитающий в Полярном бассейне у берегов Канады Galathealinum arcticum. Одна передняя часть самки этого вида имеет в длину 50 см и 1,33 мм в диаметре, все животное, вероятно, достигает 1 м. Сравнительно крупными являются и другие виды этого рода. Мелкие и среднего размера формы полупрозрачны, бесцветны или беловаты; сквозь стенку тела просвечивают наполненные красной кровью кровеносные сосуды, придающие щупальцам розовый или красноватый оттенок. У самцов в туловище обычно просвечивают беловатые половые органы, а у самок в средней части тела — желтоватые или красноватые яйца. Крупные Spirobrachia grandis и Galathealinum arcticum окрашены довольно ярко, первая в кирпично-красный, вторая форма в розовато-коричневый цвет.