(Pogonophora Johansson, 1937) Класс Погонофоры, Class Pogonophora Johansson, 1937 (Pogonophoras, Beard worms) 3 отряда
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ТИП ПОГОНОФОРЫ (POGONOPHORA)
Погонофоры интересны во многих отношениях. Открытие их явилось одним из самых неожиданных событий в современной зоологии. Еще недавно казалось, что время описания новых классов и типов давно миновало, что все группы животных высокого систематического ранга давно хорошо известны. История изучения погонофор опровергла это представление. Этот тип, объединяющий несколько десятков своеобразных морских животных, был установлен лишь в середине нашего столетия, а детально изучен лишь в последние 10 лет.
Погонофоры относятся ко вторичноротым (Deuterostomia) — большой эволюционной ветви животного мира, к которой принадлежат также иглокожие, полухордовые и хордовые (в том числе позвоночные). О родстве погонофор со всеми этими типами особенно убедительно говорит их эмбриональное развитие. Многие примитивные особенности погонофор позволяют по-новому оценивать первые этапы эволюции вторичноротых.
Погонофоры интересны также отсутствием кишечника, который в процессе эволюции исчез у них полностью. Этим они резко отличаются от всех остальных свободноживущих многоклеточных животных (не считая очень примитивных губок). Известные случаи полной утраты пищеварительного канала касаются исключительно паразитических животных (скребни, ленточные черви, некоторые эндопаразитические брюхоногие моллюски и ракообразные). Как только отсутствие кишечника у погонофор было доказано, встал сложный вопрос о том, как они питаются.
Погонофоры интересны своим образом жизни. Это животные, ведущие почти неподвижную жизнь на дне моря внутри длинной защитной трубки. Подобным же образом в трубках обитают многие многощетинковые черви (Polychaeta, Sedentaria), относящиеся к типу кольчатых червей. Одинаковый образ жизни этих двух неродственных друг другу групп животных привел к выработке у них сходных, но возникших независимо приспособлений. Благодаря этому погонофоры и живущие в трубках полихеты приобрели большое внешнее сходство, настолько смущавшее некоторых зоологов, что они ошибочно считали погонофор своеобразно измененными кольчатыми червями (Ливанов и Порфирьева, 1962). Таким образом, погонофоры дают возможность изучить важные вопросы о закономерностях развития у неродственных животных сходных приспособлений (конвергенций) под влиянием одинаковых условий жизни.
Наконец, любопытна и история изучения погонофор. Эти своеобразные обитатели океанических глубин, еще недавно казавшиеся большой зоологической редкостью, в действительности принадлежат к самым обычным элементам морской фауны. Не удивительно поэтому, что погонофоры уже давно попадали в руки ученых. Однако они не привлекали к себе внимания или причислялись к давно известным группам животных. В одном из музеев естественной истории в США многие десятки лет хранились неопознанные погонофоры, собранные еще в прошлом столетии в Тихом океане американскими океанографическими экспедициями. Лишь теперь, когда мы уже знаем, что такое погонофоры, эти уникальные экземпляры «дождались» своих исследователей. Недавно английский зоолог Д. Карлайл (Carlisle) писал: «Когда в 20-х годах английское исследовательское судно «Дискавери II» работало в Антарктике и Субантарктике, одна из обычнейших неприятностей при добывании донных животных с больших глубин заключалась в том, что после 10 или 12 часов работы драга поднималась, забитая массой каких-то волокон. Очень часто мешок драги был настолько заполнен этой противной массой «gubbins», что исключалась возможность получения какого бы то ни было материала, интересного в биологическом отношении, и весь улов после быстрого и грубого осмотра выбрасывался за борт. Это случалось так часто, что биологи на борту судна употребляли шуточное наименование «Gubbinidae» в качестве названия «семейства» для этого досадного материала».
Вспоминая этот период на борту «Дискавери II», профессор сэр Алистер Харди сказал мне, что «Gubbinidae», несомненно, были погонофоры. Таким образом, некоторые из ведущих морских биологов того времени выбросили за борт много тонн погонофор. Пикантность этой истории состоит также в том, что на «Дискавери II» было строгое правило — не выбрасывать никакого собранного биологического материала. Он должен быть сохранен целиком и привезен в Англию. Это правило существовало для того, чтобы предотвратить как раз подобные случаи уничтожения материала первостепенного научного интереса только потому, что он не был сразу правильно определен. Совершенно очевидно, что биологи того времени не подозревали о биологической природе «Gubbinidae».
Честь открытия первого представителя погонофор принадлежит французскому зоологу М. Коллери (Coullery). В 1914 г., изучая кольчатых червей, добытых с большой глубины в индонезийских морях голландской морской зоологической экспедицией на судне «Зибога», он обнаружил нитевидное животное, обитающее в тонкой кольчатой трубочке. Коллери исследовал его и назвал Siboglinum weberi (Siboga — название судна, linum — в переводе с латинского нить; weberi — видовое название в честь зоолога Вебера, начальника экспедиции). Хотя Коллери в 1944 г. переисследовал Siboglinum, выяснить полностью план строения этого животного и его систематическое положение не удалось. Вторая находка была сделана почти 20 лет спустя советским зоологом П.В.Ушаковым в Охотском море. В трале, поднятом с глубины 3500 м, оказалось несколько длинных бурых трубок, содержащих животных с пучком щупалец на переднем конце. Ушаков описал их в 1933 г. под названием Lamellisabella zachsi (от латинского lamella — пластинка, Sabella — родовое название многощетинкового червя; zachsi — видовое название в честь русского зоолога И. Г. Закса) и отнес к многощетинковым червям (Polychaeta) — классу уже давно и хорошо известному, объединяющему множество видов, широко распространенных во всех морях.
Однако шведский зоолог К. Э. Иоганссон (Johansson), исследовав Lamellisabella на разрезах, установил, что это животное ничего общего с многощетинковыми червями не имеет, и в 1937 г. выделил его в новый класс, названный Pogonophora (от греческого pogon — борода и phoro — несу). Так в системе животного мира среди примерно семидесяти классов появился еще один, в то время установленный для одного вида. Природа этого класса оставалась темной, а родственные связи и происхождение неясными, поскольку ни строение, ни эмбриональное развитие Lamellisabella не были еще изучены. Но интерес был возбужден, и в специальной зоологической литературе началось оживленное обсуждение систематического положения нового класса, которое, однако, без нового фактического материала не могло привести к определенным результатам. Интересно, что долгое время никому не приходило в голову сравнить животных, описанных Ушаковым и Коллери, и только в 1951 г. была доказана принадлежность Siboglinum к классу Pogonophora.
С 1949 г. начался новый период изучения погонофор — в морях Дальнего Востока приступил к исследованиям океанических глубин «Витязь», флагман советского исследовательского флота, экспедиционное судно Академии наук СССР. Первые же зоологические сборы «Витязя», сделанные на больших глубинах в Охотском и Беринговом морях и в Курило-Камчатской глубоководной впадине, содержали множество различных погонофор. За 15 следующих лет почти непрерывной экспедиционной работы «Витязь» совершил 35 исследовательских рейсов, участники экспедиций изучили фауну Тихого и Индийского океанов и собрали обильный уникальный материал по погонофорам. Эти животные, против всяких ожиданий, оказались весьма широко распространенными во многих океанах и морях. Новый материал дал возможность советским зоологам открыть более 60 новых видов, построить классификацию погонофор, изучить строение и эмбриональное развитие и сделать наблюдения над образом жизни и географическим распространением.
Сравнение погонофор позволило В. Н. Беклемишеву еще в 1944 г. высказать убеждение, что эта группа животных заслуживает ранга типа. Однако обоснованная характеристика нового типа погонофор и доказательства принадлежности его к эволюционной ветви вторичноротых были даны лишь в 1955 и 1957 гг. А. В. Ивановым, изучившим главные особенности строения и эмбрионального развития этих животных. К числу других существенных открытий относится объяснение способа питания погонофор, сделанное на основе изучения их щупалец и кровеносной системы (Иванов, 1955), установление особенностей раннего развития зародыша (Иванов, 1957), выяснение химической природы трубки и пигмента крови (Brunet a. Carlisle, 1958; Е. Southward a. A. Southward, 1963) и, наконец, обнаружение особого заднего отдела тела (Webb, 1964). Последнее открытие явилось совсем неожиданным. Оказалось, что на заднем конце длинного тела животного имеется особый, снабженный щетинками участок, который до сих пор оставался неизвестным, несмотря на то что через руки исследователей прошли тысячи погонофор. Это объясняется тем, что этот отдел тела помещается в заднем, очень нежном, тонкостенном участке трубки, который очень легко обрывается и или теряется при промывке улова трала, или вообще не попадает в трал, оставаясь на дне моря в глубоких слоях ила.
В настоящее время погонофоры найдены почти во всех морях, даже там, где донная фауна хорошо изучена. Так, недавно они обнаружены в Баренцевом море близ Мурманского побережья, в проливе Скагеррак у берегов Швеции, в западной части Атлантического океана, в фиордах Норвегии. Около 15 лет назад мы знали лишь 3 вида погонофор, ныне описано около 100 видов. Судя по тому, что все время в новых местах открываются все новые виды, и учитывая слабую изученность фауны Мирового океана, естественно предположить, что действительное количество видов погонофор выражается четырехзначным числом.
Географическое распространение погонофор имеет некоторые особенности. Они очень обычны в глубоководных районах Тихого, Индийского и Атлантического океанов, более или менее прилегающих к материкам, а также в окраинных морях, широко сообщающихся с океанами. Напротив, в большом удалении от островов и материков на обширных пространствах океанического дна они отсутствуют. Нет их также во внутренних морях, т. е. в морях, изолированных от Мирового океана или связанных с ним лишь сравнительно мелководными проливами. Так, например, очень обильной и разнообразной фауной погонофор отличаются Охотское и Берингово моря, тогда как соседнее с ними Японское море лишено погонофор. Отсутствуют они и в сильно опресненном Балтийском море, а также в Каспии, в Азовском и Черном морях. Недавно погонофоры обнаружены в Средиземном море.
Погонофоры в большинстве обитатели больших глубин. Почти половина известных видов живет глубже 3000 м. Исключительно глубоководными являются лишь 20 видов (25%), большей частью это эндемичные обитатели глубоководных впадин — Курило-Камчатской, Японской, Бугенвильской в Тихом океане и Яванской впадины в Индийском океане. Двадцать пять видов (около 30%) ни разу не встретились глубже 1000 м, а 7 видов (около 10%) живут на мелководье не глубже 300 м. Исключительно широким вертикальным распространением отличается Siboglinum caulleryi — форма, обычная в Охотском море, где она встречается даже на глубине около 20 м, но найденная также в пучине Курило-Камчатской впадины на глубине 8100 м.
Необходимыми условиями существования погонофор являются низкая температура (от —1,3 до +13°) и нормальная соленость воды. По-видимому, «глубоководность» погонофор обусловливается прежде всего тем, что в море наиболее холодная вода занимает более глубокие горизонты. С другой стороны, мелководные виды погонофор распространены в северных областях, где прибрежные воды подвержены сильному охлаждению. Таковы Siboglinum caulleryi в Охотском море, Nereilinum murmanicum в Баренцевом море, Galathealinum arcticum и Siboglinum hyperboreum в северном полярном бассейне.
Погонофоры всегда встречаются на илистом грунте. Нередко на твердых и длинных трубках Polybrachia, Lamellisabella и некоторых других поселяются различные сидячие беспозвоночные: фораминиферы, губки, гидроидные полипы, актинии, многощетинковые черви, мшанки, асцидии и даже мелкие стебельчатые морские лилии. Расположение этих «обрастателей» показывает, что верхняя часть трубки свободно торчит над поверхностью грунта, располагаясь более или менее вертикально, тогда как остальная, нижняя, часть ее глубоко погружена в ил.
Впрочем, у многих мелких форм трубка настолько нежна и эластична, что, несомненно, стелется своей верхней частью на поверхности грунта. Вероятно, в поселениях погонофор особи расположены довольно тесно. Необычайное обилие особей было отмечено в некоторых хорошо изученных точках северо-западной части Тихого океана, в Северном море, в Скагерраке, в антарктических водах и в других местах. Тралы приносят здесь массу населенных и пустых трубок погонофор, забивающих иногда мешок трала и даже повисающих на раме и на тросе. Если судить по обилию пустых и разлагающихся трубок погонофор, часто примешивающихся к илу, они играют важную роль в формировании грунта во многих местах океана.
Недавно было обнаружено, что некоторые погонофоры ведут сверлящий образ жизни.
Таковы описанный недавно Веббом (Webb) из одного фиорда в Норвегии Sclerolinum brattstromi и найденный в 1959 г. экспедицией на судне «Витязь» на дне Яванской впадины (около 6 тыс. м глубины) Sclerolinum javanicum. Тонкие буроватые трубочки этих видов, содержащие животных, пронизывают кусочки затонувшего дерева, например обломки древесных веток. У Sclerolinum javanicum трубочки проникают сквозь очень плотную древесину, так что нет никаких сомнений в способности животного каким-то образом сверлить дерево, но остается совершенно загадочным, как это происходит.
Погонофоры, отфильтровывающие из воды мелкие частицы детрита и планктонные организмы, зависят от количества этого взвешенного в морской воде пищевого материала и от развивающейся в нем бактериальной флоры. Поэтому они наиболее многочисленны именно в тех местах, где имеются более или менее постоянные скопления придонной органической взвеси, зависящей от направления и силы глубинных течений и от рельефа дна. По-видимому, этим и объясняется тот факт, что в морях и глубоководных впадинах фауна погонофор гораздо богаче, чем в очень удаленных от континента открытых пространствах океана.
Погонофоры не способны покидать свою трубку, так как не приспособлены к передвижению на открытом месте. Внутри трубки они, однако, могут быстро перемещаться, то высовывая наружу передний конец тела с щупальцами, то уходя глубоко в трубку. В этом отношении они напоминают живущих в трубках многощетинковых червей. По длине трубка всегда в несколько раз превосходит животное и не препятствует таким движениям. Для опоры на внутреннюю поверхность трубки служат хитиноидная уздечка, многочисленные прикрепительные сосочки, или папиллы, зубчатые щетинки поясков и щетинки заднего отдела туловища (см. ниже). Тело погонофор может сильно вытягиваться и сокращаться. Высовывание переднего конца тела из трубки наружу осуществляется путем сильного вытягивания туловища, причем задняя часть последнего в это время прочно прицепляется к стенкам трубки посредством щетинок поясков и заднего отдела. При раздражении или при опасности высунувшееся наружу животное мгновенно исчезает в глубине трубки вследствие простого сокращения мощной продольной мускулатуры, а прочное прикрепление заднего конца облегчает, конечно, это «движение бегства».
Погонофоры, поднятые на палубу с большой глубины, не обнаруживают никаких движений, хотя остаются некоторое время живыми, если их содержать в холодной воде. Английские зоологи супруги Саусворд (Southward) и датчанин Киркегаард (Kirkegaard) успешно содержали живых погонофор (представителей рода Siboglinum) в аквариуме при температуре в + 5° С. Однако животные не проявляли никакой активности в течение многих дней. Затем Карлайлу (Carlisle) удалось установить, что Siboglinum, подобно многим морским животным, активен лишь в темноте и что красный свет не препятствует его активности. Прибегнув к красному свету, Карлайл наблюдал движения животного и собирание пищи щупальцем.
Очень древнее происхождение погонофор не оставляет сомнений. Однако об ископаемых погонофорах до самого последнего времени ничего не было известно, хотя, казалось бы, их остатки в виде трубок должны встречаться начиная с нижнего палеозоя.
Погонофоры относятся к числу самых длинных животных. Их сильно вытянутое тело нитевидно или шнуровидно. Длина его обычно в 100—500 раз превышает ширину. Размеры тела варьируют очень широко. К числу миниатюрных форм относится, например, Siboglinum minutum из Берингова моря, у которого тело имеет не более 5,5 см в длину при ширине менее 0,1 мм. Таких мелких видов очень много, особенно в роде Siboglinum. К крупным погонофорам принадлежат Spirobrachia grandis, длиной не менее 25 см при диаметре туловища до 2,5 мм, и более тонкая Zenkevitchiana longissima, достигающая 36 см в длину. Однако самой большой формой является обитающий в Полярном бассейне у берегов Канады Galathealinum arcticum. Одна передняя часть самки этого вида имеет в длину 50 см и 1,33 мм в диаметре, все животное, вероятно, достигает 1 м. Сравнительно крупными являются и другие виды этого рода. Мелкие и среднего размера формы полупрозрачны, бесцветны или беловаты; сквозь стенку тела просвечивают наполненные красной кровью кровеносные сосуды, придающие щупальцам розовый или красноватый оттенок. У самцов в туловище обычно просвечивают беловатые половые органы, а у самок в средней части тела — желтоватые или красноватые яйца. Крупные Spirobrachia grandis и Galathealinum arcticum окрашены довольно ярко, первая в кирпично-красный, вторая форма в розовато-коричневый цвет.
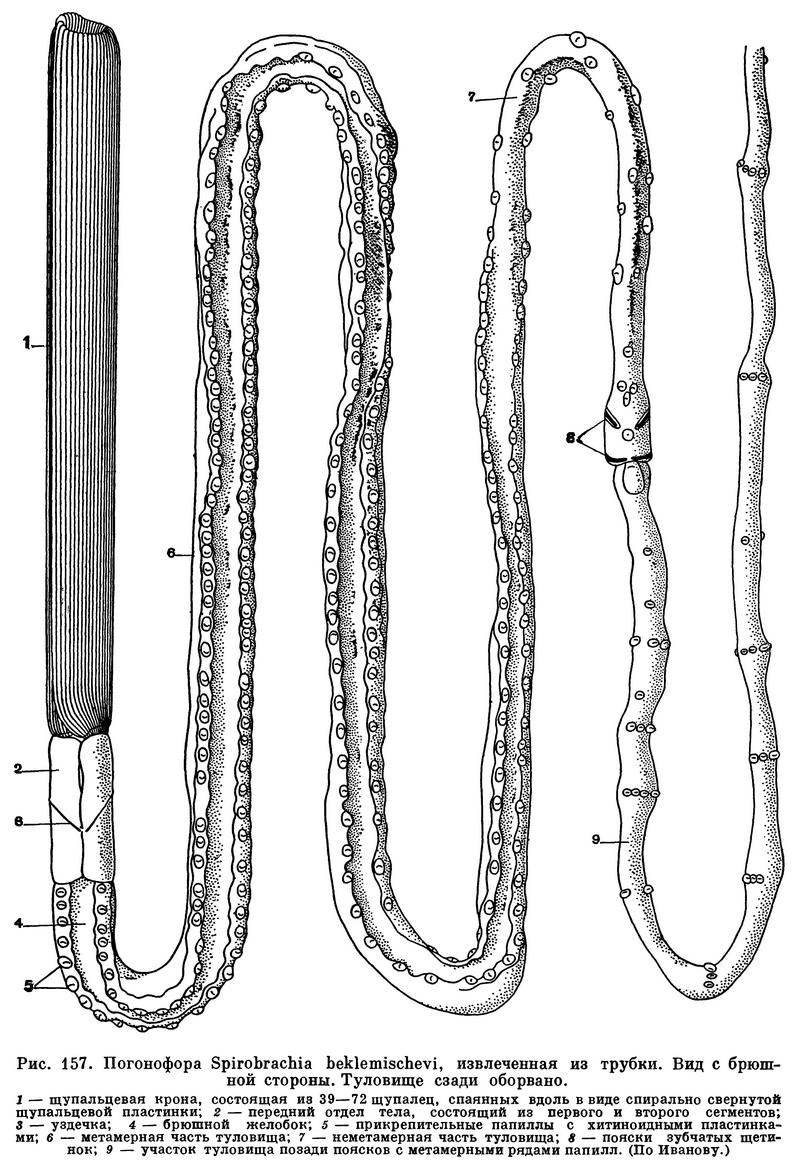
Внешний вид животных весьма своеобразен. Червеобразное цилиндрическое тело их состоит из трех главных отделов, или сегментов: двух передних, сравнительно коротких, и заднего, необычайно длинного. На самом коротком, первом сегменте располагаются очень длинные щупальца и спинная головная лопасть. Несколько более длинный второй сегмент несет пару косых хитиноидных ребрышек, или килей, называемых уздечкой; они служат для опоры передней части тела на стенку трубки. Третий сегмент, или туловище, значительно меняется от переднего конца к заднему. В нем ясно различимы два главных участка: передний — преаннулярный, простирающийся до особых поясков, и задний, лежащий позади поясков, — постаннулярный. Последний в свою очередь подразделяется на длинный (передний) участок, несущий прикрепительные папиллы, и на задний отдел, снабженный палочковидными щетинками.
По-видимому, все наиболее примитивные формы имеют только 4—6 очень длинных щупалец, но у большинства число щупалец в процессе эволюции увеличилось до 10, 20 и даже до 250 и более. С другой стороны, у некоторых форм (Siboglinum) произошла редукция многих щупалец и осталось лишь одно правое. На щупальцах обычно располагаются ряды очень нежных и тонких пиннул, или ворсинок.
У большинства погонофор все щупальца свободно прилегают друг к другу, но у Lamellisabella и Spirobrachia спаяны между собой. У Lamellisabella, которая имеет круговое расположение щупалец, при этом образуется полый щупальцевый цилиндр, а у Spirobrachia формируется щупальцевая пластинка, свернутая спирально наподобие рулона бумаги.
Чрезвычайно характерными образованиями туловища являются прикрепительные сосочки, или папиллы. Это небольшие округлые или грушевидные подушечки, обычно снабженные маленькими хитиноидными пластинками. Замечательно, что у наиболее примитивных погонофор они слабо развиты, представлены неопределенно большим числом и неправильно рассеяны на поверхности туловища. У более развитых родов и видов передние папиллы принимают правильное расположение, а именно лежат на брюшной стороне попарно и метамерно. Общее число папилл на туловище при этом сильно сокращается. В ряду высших форм количество передних метамерных папилл постепенно возрастает, и, наконец, почти все папиллы туловища становятся метамерными.
Преаннулярная область туловища отличается еще наличием широкой ресничной полоски на спинной стороне. Судя по связи со спинным нервным стволом, это орган чувств (может быть, орган химического чувства, или водного обоняния).
В средней части туловища находятся 2 или 3 пояска очень мелких зубчатых щетинок, которые, несомненно, служат для прикрепления тела к внутренней поверхности трубки. Позади поясков находится длинный участок туловища, отличающийся развитием метамерных папилл на брюшной стороне. За ним, наконец, следует небольшой задний отдел, несущий метамерные щетинки, которыми животное закрепляется в глубйне трубки. Характерные нарушения метамерии щетинок показывают, что первоначально они располагались на этом участке тела беспорядочно.
Покровы погонофор состоят из однослойного эпителия, богатого одноклеточными железами и почти сплошь покрытого кутикулой. Лишь незначительные участки на щупальцах и на спинной стороне туловища одеты мерцательным эпителием. Под кожным эпителием лежит тонкий слой кольцевых мускульных волокон, за которым располагается мощный продольный мускульный слой. Имеются многочисленные многоклеточные железы, открывающиеся протоками на поверхности тела; они выделяют вещество трубки.
Хорошо развитая полость тела имеет собственные эпителиальные стенки и, следовательно, представляет собой вторичную полость тела, или целом. Впереди, у основания щупалец, лежит непарный мешковидный или подковообразный целом первого сегмента. От него отходят целомические каналы щупалец. Кроме того, пара выделительных каналов открывается своими внутренними концами в этот целом, что указывает на принадлежность их к первому сегменту. Во втором и третьем сегментах имеются свои парные целомические мешки.
Нервная система крайне примитивна и целиком залегает в толще кожного эпителия. Она представлена почти сплошным нервным сплетением, клетки и волокна которого образуют сгущения на спинной стороне тела — мозг — в области первого сегмента и спинной нервный тяж, проходящий вдоль всего тела. От мозга отходят нервы щупалец.
Кровеносная система состоит из продольного брюшного сосуда, по которому кровь течет сзади наперед, и спинного сосуда, несущего кровь назад. Брюшной сосуд у основания щупалец снабжен мускулистым сердцем, сокращения которого проталкивают кровь в приносящие сосуды щупалец. У представителей отряда Athecanephria к сердцу со спинной стороны прилегает мешковидный участок целома — околосердечная сумка, или перикардий. В спинной сосуд кровь поступает из выносящих сосудов щупалец. В задней половине туловища оба продольных сосуда соединяются многочисленными поперечными ветвями. Кровь погонофор имеет красный цвет и содержит гемоглобин.
Органами выделения служат целомодукты первого сегмента. Это длинные ресничные каналы, по степени развития и по локализации которых все погонофоры делятся на две группы. У представителей отряда Athecanephria они широко расставлены и прилегают к задним участкам боковых ветвей спинного продольного сосуда. У форм, принадлежащих к отряду Thecanephria, целомодукты очень длинны, извиты и сближены друг с другом медиально, причем прилегают к спинному сосуду, глубоко впячивая его брюшную стенку.


Погонофоры раздельнополы, при этом самцы и самки не отличаются по внешнему виду. У самцов сперматозоиды развиваются в паре очень длинных семенных мешков, лежащих в задней половине туловища. От семенных мешков отходит пара семяпроводов, тянущихся вперед до передней границы туловища, где они открываются наружу на брюшной стороне тела. В них формируются многочисленные уплощенные или веретеновидные перепончатые сперматофоры с очень длинной тонкой нитью. У самок яичники занимают переднюю половину туловища. Зрелые яйца сначала попадают в полость тела через разрыв стенки яичников, затем выводятся наружу по яйцеводам.
Самка откладывает яйца в переднюю часть своей собственной трубки, в которой и протекает все эмбриональное развитие. Дробление яйца происходит, как у низших вторичноротых, по радиальному типу. Образование стенок полости тела (мезодермы) осуществляется так называемым энтероцельным путем, при котором мешковидные зачатки вторичной полости тела сперва возникают как боковые выпячивания первичного кишечника, а потом отделяются от него. Этот способ развития целома чрезвычайно характерен для вторичноротых. У зародыша сперва развивается энтодермальиая кишка, ее крупные клетки переполнены запасными питательными материалами — желтком и жиром. При превращении зародыша в молодое животное первичная кишка, выполнив свою роль временного резервуара запасных питательных веществ, полностью рассасывается. Настоящей планктонной личинки погонофоры не имеют. Молодое животное выбирается из материнской трубки и после очень короткого периода свободной жизни оседает на дно и выделяет свою собственную трубку.
Для решения вопроса о том, как питаются погонофоры, эти неподвижные обитатели морского дна, полностью лишенные обычной пищеварительной системы и неспособные активно разыскивать пищу, рассмотрим строение их щупальцевого аппарата.
Несмотря на все разнообразие в числе и особенностях прикрепления щупалец, в щупальцевом аппарате погонофор всегда проявляются одни и те же характерные особенности. Прилегая друг к другу, щупальца ограничивают межщупальцевое пространство, у некоторых форм, как уже было сказано выше, щупальца даже соединяются друг с другом. При этом у одних (Lamellisabella) образуется полый цилиндр, а у других (Spirobrachia) спирально свернутая щупальцевая пластинка.
Таким образом, наличие временной (при свободных щупальцах) или постоянной (при спаянных щупальцах) межщупальцевой полости той или иной конфигурации очень характерно. Она образуется даже у Siboglinum вследствие штопорообразного закручивания единственного, но зато необычайно длинного щупальца.
Каждое щупальце является длинным выростом тела и содержит целомический канал. На внутренней поверхности щупальца у большинства погонофор расположены ряды пиннул — длинных и очень тонких выростов. Каждая пиннула представляет собой одну очень вытянувшуюся клетку эпителия щупальца, содержит одно ядро и пронизана двумя параллельными, соединяющимися у вершины пиннулы тончайшими кровеносными капиллярами — приносящим и выносящим. Пронизывая толщу эпителия щупальца, эти капилляры сообщаются с соответственными сосудами самого щупальца. У основания пиннул располагаются ряды мерцательных клеток. При соприкосновении соседних щупалец они образуют временные (у большинства форм), а при срастании щупалец — постоянные продольные ресничные бороздки. Между основанием пиннул и мерцательными клетками иногда имеются ряды железистых клеток. Характерно, что пиннулы обращены в межщупальцевое пространство, где образуют густое сплетение.
Одной из функций щупалец, несомненно, является дыхание. Однако перечисленные выше детали их строения показывают, что они служат также для собирания пищи, ее переваривания и всасывания продуктов пищеварения. Вероятно, мерцание ресничных бороздок на щупальцах гонит воду в межщупальцевое пространство. Вода входит между концами щупалец и выходит между их основаниями. Взвешенные в воде микроскопические организмы и детрит застревают в густой сети пиннул, которая, таким образом, играет роль фильтра. В межщупальцевом пространстве выделяются пищеварительные ферменты. Здесь же происходит переваривание пищи. Питательные вещества всасываются пиннулами, играющими роль своеобразных кишечных ворсинок, и поступают в кровь их сосудов, откуда с током крови попадают через выносящие сосуды в спинной сосуд и разносятся по всему телу. Возможно, фильтрация воды щупальцами и переваривание пищи происходят не одновременно, т. е. сперва накапливается некоторый запас пищи, потом животное переваривает ее, втянув крону щупалец в трубку.
Относительно способа питания Siboglinum, обладающего всего одним щупальцем, сначала на основании изучения его строения было сделано предположение (Иванов, 1960), что животное подбирает пищевые частицы с поверхности ила, «исследуя» окружающее пространство дна своим щупальцем. Затем щупальце обвивается вокруг пищевого комочка и вытягивается в трубку, где пища и переваривается. В самое последнее время Карлайлу удалось полностью подтвердить это предположение прямыми наблюдениями над животными, содержащимися в аквариуме.


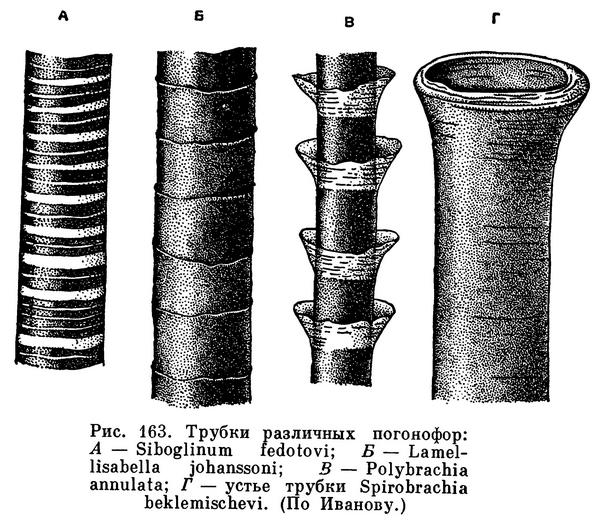
Трубки погонофор имеют почти цилиндрическую форму, так как очень постепенно расширяются от заднего конца к переднему. Длина их колеблется от нескольких сантиметров до 1,5 м у Zenkevitchiana longissima, а у Galathealinum arcticum, вероятно, достигает нескольких метров. Строение трубки в общем у всех погонофор сходно; однако у одних форм она гибка и нежна и обладает характерной кольчатостью, у других имеет беловатые пергаментообразные стенки, у третьих отличается прочностью и твердостью и темным бурым цветом. Детали строения трубок часто используются как хорошие видовые признаки. У большинства погонофор передний конец трубки имеет особое строение. Здесь стенки ее особенно тонки, обычно совсем прозрачны и легко спадаются. Напротив, у Lamellisabella и Spirobrachia передний конец трубки
обладает толстыми непрозрачными стенками, а устье трубки имеет форму воронки. Трубки погонофор состоят из хитина.
Тип погонофор содержит один класс, который делится на два отряда. К первому отряду — Athecanephria — относятся погонофоры со свободными щупальцами и с перикардиальным мешочком. Сперматофоры у представителей этого отряда веретеновидные, а передняя часть трубки всегда пленчатая, со спадающимися стенками. Второй отряд — Thecanephria — характеризуется свободными или спаянными щупальцами, отсутствием перикардия и листовидными сперматофорами. У некоторых относящихся сюда форм трубка с воронковидным устьем.

ОТРЯД ATHECANEPHRIA
К этому отряду, содержащему два семейства, относятся примитивные формы, иногда лишенные метамерии туловищных папилл.
ОТРЯД THECANEPHRIA
Из четырех семейств, принадлежащих к этому отряду, самыми примитивными являются полибрахииды. Они характеризуются хорошо отграниченным первым сегментом тела, не связанными друг с другом щупальцами и пленчатым передним участком трубки. Другие два семейства отличаются слиянием первых двух сегментов и срастанием щупалец, образующих более или менее сложный щупальцевый аппарат, а также имеют воронковидное устье трубки.