(Mesostigmata) Отряд Мезостигматы, Order Mesostigmata 21 семейство
Жизнь животных. Том 3. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1969 г.
ОТРЯД ПАРАЗИТИФОРМНЫЕ КЛЕЩИ (PARASITIFORMES)
Этот отряд по числу видов меньше предыдущего, насчитывает около 4000, но цифра эта также далека от действительной, так как изучение ряда семейств только начато. Сюда относятся три основные группы, рассматриваемые обычно как надсемейства: гамазоидные клещи (надсем. Gamasoidea), уроподы (надсем. Uropodoidea) и иксодовые клещи (надсем. Ixodoidea). Первые два надсемейства велики и разнообразны, наряду со свободноживущими формами здесь немало сожителей (комменсалов) насекомых и позвоночных, а среди гамазоидных клещей и паразитов последних. Иксодовые клещи представлены целиком кровососущими паразитами наземных позвоночных.
Выяснение происхождения этого отряда и его положения среди арахнид затрудняется тем, что даже наименее специализированные его представители значительно видоизменены, а палеонтологических материалов почти нет. Своеобразный тип строения и индивидуального развития этого отряда был выяснен автором в 1947 году при изучении наименее изменившихся почвенных гамазоидных клещей семейства Rhodacaridae. Так как среди паразитиформных клещей сегментированные формы не сохранились, пришлось прибегнуть к косвенным методам выяснения сегментального состава тела и его изменений в индивидуальном развитии. На теле этих клещей имеется сложный и постоянный набор щетинок и лировидных органов чувств. Изучение численного состава, топографии этих органов, а также порядка их появления по фазам развития показало, что они образуют поперечные ряды, соответствующие слившимся сегментам. В результате такого, как его теперь называют, хетологического анализа была получена теоретическая картина сегментации тела (в форме карты хетомерии, т. е. предполагаемых посегментных наборов щетинок), которая совпала с другими анатомическими данными (расположение мышц и др.) и, что особенно важно, с сегментацией зародыша. В дальнейшем среди низших уропод удалось найти формы, у которых частично сохранилась сегментация брюшка, также подтвердившая теоретическую картину. В результате оказалось возможным описать строение и развитие паразитиформных клещей в форме, сравнимой с другими арахнидами, причем выявилось большое своеобразие этого отряда и особенно резкие его отличия от акариформных клещей.
Туловище исходных форм (рис. 72) разделено на головогрудь и брюшко, причем граница между ними делит предполовой сегмент на часть, которая примыкает к головогруди, и часть, примыкающую к брюшку. В составе головогруди слились шесть сегментов, несущих конечности, и боковые участки головной лопасти, передняя часть которой отделена и входит в состав сложно устроенного ротового аппарата — гнатемы. В брюшке объединены шесть сегментов и два-три недоразвитых концевых.
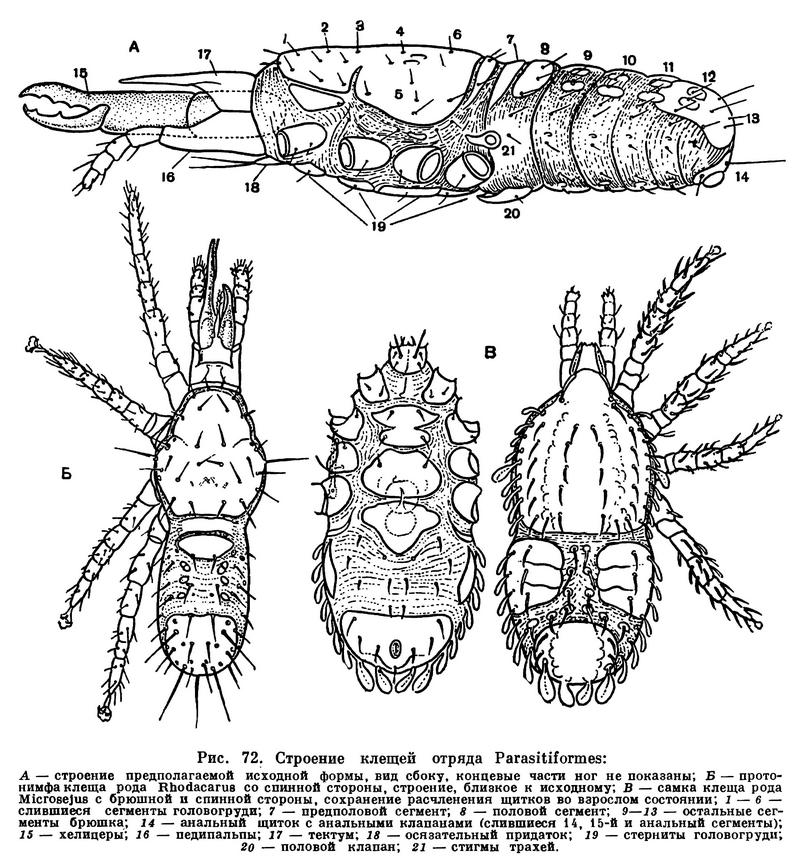
Для отряда в целом характерно образование сложного имагинального панциря, причем изучение индивидуального развития и эволюционных рядов форм позволяет понять, как он образуется. В исходном состоянии наружный скелет представлен многими посегментными склеритами. Головогрудь покрыта щитом, передний отдел которого соответствует трем передним сегментам и отделен швом. Снизу между подвижно причлененными тазиками ног расположены стерниты, сливающиеся в дальнейшем в цельную грудину. Судя по всему, ближайшие предки Parasitiformes имели членистое брюшко с небольшими парными тергитами и стернитами — местами прикрепления посегментных спинно-брюшных мышц. Слияние сегментов начинается с заднего конца брюшка, сливающиеся тергиты образуют здесь сначала небольшой пигидиальный щиток, который затем разрастается вперед, превращаясь в сплошной верхний брюшной щит (нотогастер). У многих форм последний сливается далее с головогрудным щитом в общий спинной щит и следы разделения головогруди и брюшка исчезают.
На нижней стороне брюшка у исходных форм имеется небольшой анальный щиток с анальным отверстием, прикрытым клапанами, а на восьмом сегменте — половой щиток, прикрывающий половое отверстие. В дальнейшем нижняя поверхность брюшка уплотняется двумя способами: либо анальный щиток разрастается вперед, образуя так называемый вентро-анальный щит, либо половой щиток разрастается назад, образуя генито-вентральный щит. У самцов в связи с выработкой своеобразного способа копуляции половое отверстие смещается вперед, к основанию гнатемы, и принимает вид небольшой вороночки. У самок ряда гамазоидных клещей в связи с относительно крупными размерами яиц и особенно при живорождении половой щиток увеличивается, занимает пространство между третьей и четвертой парами ног, а под ним вокруг крупной поперечной половой щели образуется сложный внутренний скелет, широко раздвигающийся при выходе молоди. У иксодид и уропод, откладывающих яйца, более мелкие относительно самки, половое отверстие невелико.
По бокам тела образуются лентовидные или округлые так называемые перитремальные склериты, на которых открываются дыхальца трахей, а нередко еще набор боковых (плейральных) склеритов.
У многих свободноживущих форм во взрослом состоянии части скелета сливаются в сплошной твердый панцирь, что характерно для ряда гамазид и особенно уропод. У паразитических клещей, насасывающих много крови, напротив, наблюдается редукция скелета и освобождение тела от щитков.
Хелицеры и педипальпы входят в состав гнатемы, или «головки», строение которой у Parasitiformes иное, чем у акариформных клещей. Тазики педипальп срастаются не только друг с другом, но и с передней частью головной лопасти (так называемым тектумом), образуя склеритное кольцо (основание гнатемы), из которого выдвигаются хелицеры и в котором помещается сосательный желобчатый конус (эпистом), ведущий в глотку. По бокам основания ротового аппарата причленяются щупальца педипальп, а снизу видоизмененные жевательные лопасти. Гнатема причленяется перепончатой шейкой и весьма подвижна как целое. Хелицеры удлиненные, 3-члениковые, втягиваются в кожистые влагалища. У многоядных и хищных форм они клешневидные, нередко очень мощные (табл. 8, 12), у кровососущих паразитов они превращаются в колющие стилеты или более сложные образования, служащие для фиксации в коже хозяина. У ряда уропод, высасывающих свою добычу или соки растений, хелицеры также длинные, стилетовидные. У самцов хелицеры обычно служат копулятивными органами и в той или иной мере видоизменены. На пальцах клешни имеются щели или жгутовидные придатки — сперматодактили, служащие для переноса сперматофора в половое отверстие самки. У некоторых гамазид эти придатки пронизаны каналами, по-видимому, наполняемыми спермой, а у самок имеются семеприемники. Способ копуляции в этих случаях точно не выяснен, но не исключено, что он сходен с тем, что мы видели у пауков, только совокупительными органами служат не педипальпы, а хелицеры.
Ноги несут на конце лапок эластичный придаток с парой коготков и присоской между ними. У гамазид ноги обычно длинные, свободноживущие формы в большинстве быстро бегающие хищники. Передние ноги тоньше других и выполняют роль усиков, на конце лапок имеется сложно устроенный тарзальный орган, в составе которого, помимо длинных осязательных щетинок, имеются органы химического чувства. Ноги уропод короткие, вкладываются в специальные ямки при замирании. У постоянных паразитов ноги нередко толстые, с мощными коготками и шиловидными щетинками, служат для прикрепления к хозяину.
Дышат паразитиформные клещи с помощью трахей, которые открываются парой стигм по бокам тела над основаниями третьей-четвертой пар ног или позади последней. Трахеи хорошо развиты, ветвятся и имеют спиральные опорные утолщения. Стигмы открываются на перитремальных щитках, чаще лентовидных. По этим щиткам вперед от стигм обычно тянется трубка — перитрема, пронизанная порами, через которую, по-видимому, происходит дыхание при закрывании стигм. У иксодовых клещей этих трубок нет и стигмы открываются на округлых или запятовидных щитках, причем их запирательный аппарат также хорошо развит.
По внутреннему строению Parasitiformes напоминают сенокосцев. Характерны несколько пар крупных слепых выростов средней кишки, мальпигиевы сосуды, сокращение коксальных желез до одной не всегда функционирующей пары, небольшое сердце, крайняя концентрация нервной системы и т. п.
Биология размножения своеобразна. О способе копуляции мы говорили. Обычно особи соприкасаются брюшными сторонами и самец с помощью хелицер вводит сперматофор в половое отверстие самки или приклеивает близ него. Большинство уропод и иксодовые клещи кладут яйца, среди гамазид наблюдаются все переходы от яйцекладущих к живородящим, причем есть формы, рождающие личинок и
нимф. В большинстве случаев яйца малы абсолютно и велики относительно самки, в связи с чем они вызревают и откладываются по одному. Но у иксодовых клещей, крупных кровососущих паразитов, все созревшие яйца находятся в половых путях самки и откладываются затем одной кладкой, обычно по нескольку тысяч.
Индивидуальное развитие чрезвычайно своеобразно (рис. 73). Выяснение сегментального состава тела и его изменений в послезародышевом развитии в сочетании с эмбриологическими данными позволяет понять наблюдаемые здесь явления. Эмбриональное развитие в этом отряде довольно однотипно, причем нет и следов тех примитивных черт, которые мы видели у акариформных клещей. Несмотря на малые размеры яиц, дробление поверхностное, и порядок сегментации зародыша вторично изменен, как у большинства арахнид. Наиболее характерно, что к концу эмбрионального развития зародыш приобретает строение, вполне типичное для арахнид вообще. Имеется не шесть, а восемь пар развитых расчлененных ног и полный состав сегментов брюшка, свойственный взрослым клещам этого отряда. Но далее у вполне развитого зародыша происходят необычайные явления. В течение нескольких часов (что удается наблюдать прижизненно, например, в прозрачных яйцах гамазоидных клещей) происходит сокращение нескольких сегментов в области соединения головогруди и брюшка. По мере роста зародыш как бы сжимается в средней части. При этом ноги четвертой пары съеживаются до небольших клеточных скоплений и уходят под покровы. В результате получается шестиногая личинка со «спрятанными» до времени ногами четвертой пары и укороченным брюшком. В таком виде она покрывается кутикулой и вскоре выходит наружу. Предличиночной фазы здесь вообще нет.
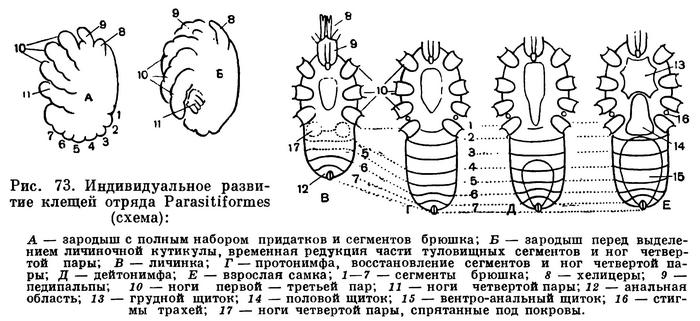
Послезародышевое развитие, помимо шестиногой личинки, включает две нимфальные фазы и взрослую. Нимф здесь называют прото- и дейтонимфой, но они ни в какой мере не соответствуют таковым акариформных клещей, и в их названии отражена лишь последовательность линек. При превращении личинки в протонимфу временно «спрятанные» сегменты и ноги четвертой пары как бы развертываются и нормальная организация клеща сразу восстанавливается. Протонимфа замечательна тем, что наиболее отчетливо олицетворяет предковое состояние этого отряда. Все, что временно извращено у личинки, у протонимфы восстановлено, но в то же время у нее еще минимальны вторичные изменения, наступающие на более поздних фазах по мере усложнения имагинального панциря. Поэтому протонимфа служит ключом, позволяющим понять организацию Parasitiformes. Расчлененный скелет, небольшой головогрудной щит и грудина, маленький пигидий на конце брюшка и посегментные склериты перед ним, стигмы еще без перитрем и т. д. — все это прототипические черты отряда (рис. 72, Б). При превращении в дейтонимфу в основных чертах оформляется вторичный наружный скелет, развиваются перитремы, посегментные наборы щетинок дополняются до окончательного состава. При превращении во взрослого клеща образуется половое отверстие, у самок — половой щиток, а у самцов — копулятивные органы хелицер. Происходит окончательное, нередко очень сильное уплотнение всего панциря.
Таковы в общих чертах особенности строения и развития этого отряда клещей. Сравнивая этот отряд с предыдущим (Acariformes), легко видеть, насколько существенны различия между ними. Их не только невозможно объединить, как это делается по традиции, но они оказываются стоящими на совсем разных эволюционных уровнях и в противоположных концах общей системы паукообразных. Сравнение Parasitiformes с другими арахнидами приводит к выводу, что они произошли от каких-то сенокосцеподобных форм, близких к некоторым ископаемым отрядам арахнид каменноугольного периода (Architarbi и др.), которые дышали легкими. Прежде Parasitiformes были более сходны с родственными им отрядами, но в дальнейшем благодаря измельчанию их строение изменилось (слияние сегментов, образование «головки» и др.), а индивидуальное развитие глубоко извратилось, внешне уподобившись развитию других клещей. Рассмотрение причин последнего явления увело бы нас в область специальных вопросов сравнительной эмбриологии. Заметим только, что, судя по картинам, наблюдаемым в эмбриональном развитии, ближайшие предки Parasitiformes не имели шестиногой личинки, из яйца выходила восьминогая форма, как у большинства арахнид (рис. 74, 1).
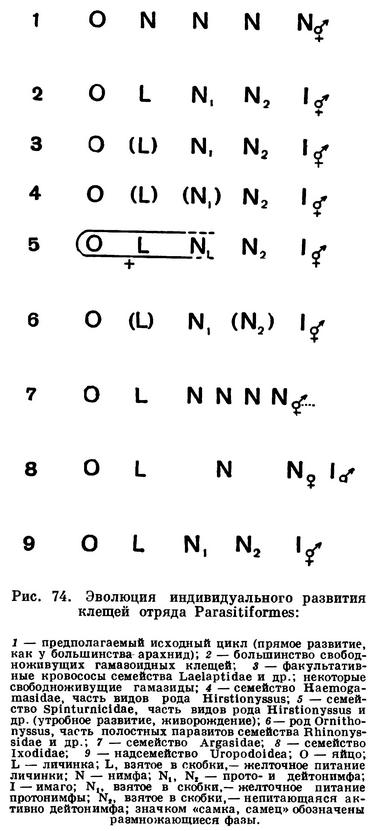
Малые размеры также открыли перед этим отрядом большие эволюционные возможности: заселение скважин почвы и всевозможных органических субстратов, переход к паразитизму. Однако характер эволюции Parasitiformes иной, чем у предыдущего отряда (Acariformes). Среди непаразитических форм господствуют хищники, разрывающие добычу хелицерами, причем хищничество сочетается с потреблением животных остатков и растительной пищи подобно тому, что мы видели у сенокосцев. Иной и характер паразитизма — переход от хищничества к сосанию крови наземных позвоночных. При этом разнообразие паразитов здесь создается не в силу разных источников питания, а благодаря различным взаимоотношениям с животными-хозяевами. В этом отряде можно различить несколько типов кровососущих паразитов, происходящих один от другого: подстерегающие временные паразиты — убежищные, т. е. нападающие на хозяина в гнезде, норе и иных укрытиях, и внеубежищные, или пастбищные, подстерегающие животных в открытой природе; постоянные паразиты — наружные, которые живут и размножаются на теле хозяина, и полостные паразиты, перешедшие к эндопаразитизму в дыхательных органах животных, и др. Как мы увидим, каждый из этих типов характеризуется целым комплексом приспособительных изменений строения, биологии и индивидуального развития, т. е. тем, что В. Н. Беклемишев в своих трудах по сравнительной паразитологии называет жизненной схемой вида. Специализация индивидуального развития идет при этом либо по линии живорождения, либо по линии увеличения или уменьшения числа нимфальных линек (рис. 74).
Питаясь кровью позвоночных, паразитиформные клещи вступили в тесные взаимоотношения с возбудителями многих болезней животных и человека — фильтрующимися вирусами, риккетсиями, бактериями, простейшими-кровепаразитами и др., и стали их специфическими переносчиками и хранителями. Наиболее важны в этом отношении иксодовые клещи, по числу переносимых болезней занимающие одно из первых мест среди кровососущих членистоногих. Они передают такие опасные заболевания, как энцефалиты, клещевые тифы, туляремию, чуму, бруцеллез, гемоспоридиальные болезни домашних животных и др. Эти клещи составляют важнейший объект медицинской и ветеринарной акарологии и паразитологии вообще. Их изучением и разработкой мер защиты человека и животных занимаются многие паразитологи, эпидемиологи и врачи во всех странах мира. Специальная литература по клещам — переносчикам болезней огромна. Советская наука по праву занимает в этой области ведущее место. Теория природной очаговости трансмиссивных болезней, т. е. передаваемых клещами и насекомыми, разработанная Е. Н. Павловским, и учение о жизненных схемах переносчиков болезней, созданное В.Н. Беклемишевым, служат основой исследовательской и практической работы в этой области и широко признаны у нас и во всем мире. Данные о кровососущих клещах изложены в целом ряде научных и учебных руководств: Е. Н. Павловский, Курс паразитологии человека, изд. АН СССР, т. I, 1945, т. II, 1949; «Учебник медицинской энтомологии», под ред. В. Н. Беклемишева, Медгиз, 1949; «Определитель членистоногих, вредящих здоровью человека», под ред. В. Н. Беклемишева, Медгиз, 1958, и др. Современный уровень знаний о кровососущих клещах хорошо отражают соответствующие главы в книге «Переносчики возбудителей природноочаговых болезней», составленной коллективом специалистов под руководством и при участии П. А. Петрищевой (Медгиз, 1962).
Гамазоидные клещи (надсем. Gamasoidea) представлены свободноживущими и паразитическими формами и составляют большую часть отряда, до 30 семейств. Непаразитические гамазиды — хищники или многоядные формы, населяют почву, лесную подстилку, скопления всевозможных гниющих остатков. Многие живут в гнездах общественных насекомых, птиц и млекопитающих. Широко распространен паразитизм на наземных позвоночных, причем наблюдаются все стадии перехода от гнездового сожительства и хищничества к питанию кровью животных и различным способам паразитирования: подстерегающие гнездовые и открытоживутцие паразиты, живущие постоянно на теле хозяев, в дыхательных путях и т. п.
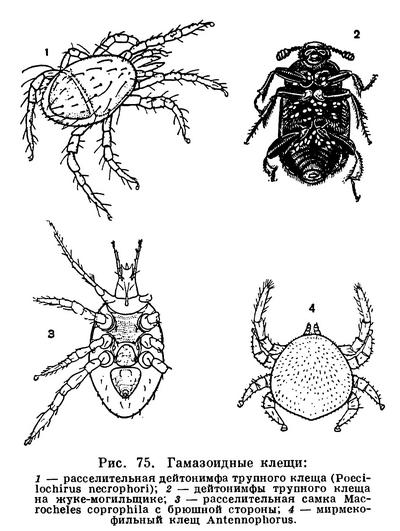
Среди непаразитических гамазид (рис. 75) распространено пассивное расселение (форезия) на насекомых и других животных, что особенно характерно для видов, размножающихся в недолго существующих, эфемерных, субстратах — помете, трупах и т. п. Расселяются чаще взрослые самки, но иногда дейтонимфы. Примером первого могут служить клещи семейства Macrochelidae, многие виды семейства Parasitidae и др. Эти сравнительно крупные (до 2,5 мм), быстро бегающие клещи размножаются в гниющих веществах, питаясь мелкими беспозвоночными — нематодами, другими клещами, яйцами и личинками мух, а также микрофлорой. Цикл развития непродолжительный, полторы-две недели. Личинки и нимфы влаголюбивы, живут в толще субстрата. На поверхность выходят расселительные самки, защищенные плотными щитками. Они прицепляются к насекомым, которые выплаживаются тут же или посещают гниющие вещества, разносятся насекомыми и попадают на свежий помет и трупы. Расселительные самки реагируют на малейшее сотрясение. Достаточно слегка постучать по подсохшей корочке навоза, заселенного клещами, как они высыпают из всех щелей, воспринимая стук как прилет транспортера. Есть виды, расселяющиеся на жуках-навозниках, могильщиках, стафилинах, на синантропных мухах. Последние заносят клещей в жилье, на пищевые продукты. На мясных мухах рода Calliphora бывает по два-три десятка клещей-макрохелид, и весь этот «десант» сбрасывается при посещении мухой пищевых продуктов, особенно мяса.
Трупный клещ (Poecilochirus necrophori) расселяется в фазе дейтонимфы. Он живет на падали, дейтонимфы держатся на поверхности и переносятся на новые трупы на жуках-могильщиках рода Necrophorus. Взрослые клещи, подобно личинкам и протонимфам, живут скрыто, питаясь тканями трупа и трупоядными членистоногими.
Интересно, что среди гамазид распространен партеногенез (девственное размножение), что, очевидно, имеет приспособительное значение, особенно при описанном способе расселения: достаточно попасть в субстрат одной самке, как она без помощи самцов дает начало колонии клещей. У разных видов партеногенез выражен в разной степени. В одних случаях самцы имеются всегда, в других появляются периодически, а иногда вообще отсутствуют. Один вид клещей семейства Aceosejidae в нашей лаборатории уже семь лет живет и размножается без самцов.
Гамазиды составляют значительную часть обильного населения гнезд общественных насекомых, птиц и млекопитающих. Взаимоотношения с хозяевами гнезд разнообразны — хищники, расселяющиеся на теле хозяев, нахлебники, паразиты. Например, мирмекофильные виды семейства Antennophoridae с длинными передними ногами, подобными усикам, держатся на нижней стороне головы муравьев, подхватывая капли пищи с их щупиков и челюстей.
Через гнездовое сожительство многие гамазоидные клещи перешли к паразитизму на мелких млекопитающих и птицах. Как возникли здесь кровососущие паразиты, позволяют понять семейства Laelaptidae и Haemogamasidae. В первом из них наряду с большим числом свободноживущих форм имеются гнездовые сожители, находящиеся на разных стадиях перехода к питанию кровью. Так, виды рода Hypoaspis, хищные или многоядные, охотно подсасывают кровь, вытекающую из поранений кожи животных. Виды родов Eulaelaps и Haemolaelaps (табл. 10, 2), так называемые факультативные кровососы, могут нормально размножаться уже только при смешанном питании мелкими членистоногими, микрофлорой и капельной кровью теплокровных, причем потребность в крови у разных видов различна. Одни еще неспособны повреждать кожу взрослых животных, но ранят кожу детенышей, например сосунков грызунов, другие прогрызают кожу взрослых и сосут вытекающую кровь. Хелицеры у большинства таких форм клешневидные и еще мало отличаются от хелицер хищников, но в семействе Haemogamasidae уже есть виды, например паразит грызунов Haemogamasus liponyssoides, у которых хелицеры превращены в стилеты и которые питаются только кровью — обязательные (облигатные) кровососы.
Большинство факультативных кровососов живет и размножается в гнезде, лишь на короткое время заползая на хозяина, чтобы напиться крови. Цикл развития этих форм мало изменен по сравнению с таковым непаразитических гамазид. Главное отличие в том, что личинка здесь кратковременная, непитающаяся, существует за счет желтка в кишечнике, а иногда не питается и протонимфа (сем. Haemogamasidae, рис. 74, 5,4). При этом обычно откладываются яйца, но иногда самка рождает личинок. Некоторые факультативные кровососы переходят к жизни в шерсти грызунов. Таковы клещи рода Laelaps (рис. 76, 2). В этом случае вырабатываются приспособления против механического воздействия хозяина — счесывания и раздавливания. Взрослые клещи защищены плотными щитками, ноги коренастые, с крепкими коготками, на теле крупные щетинки, удерживающие паразита в шерсти. Характерно живорождение, наиболее уязвимые молодые фазы — личинка, а иногда и протонимфа, — развиваются в теле самки под защитой ее покровов. Жизнь на теле хозяина — постоянный эктопаразитизм — приводит к специализации видов паразитов в отношении хозяев. Так, Laelaps muris живет на водяной полевке, Laelaps algericus — на домовой мыши, Laelaps echidninus — на крысе-пасюке, Laelaps jettmari — на хомячках и т. д.


Среди обязательных кровососов центральное место занимает семейство Dermanyssidae. Это семейство нередко делят на два: собственно Dermanyssidae и сем. Macronyssidae. Клещи этого семейства паразитируют на рептилиях, птицах и млекопитающих и по типу паразитизма представляют три направления специализации: гнездово-норовые паразиты, внеубежищные формы и постоянные эктопаразиты. Тип паразитизма проявляется в строении и биологии этих клещей.
Примером гнездовых дерманиссид служат птичьи клещи рода Dermanyssus. Наиболее известен куриный клещ (Dermanyssus gallinae, рис. 76, 4—6). Этот вид живет в птичниках, в гнездах голубейг в клетках певчих птиц, в дикой природе. Голодная самка 0,7 мм длиной, серо-желтоватая, очень подвижная. При сосании крови туловище клеща раздувается, просвечивающая кровь придает ему ярко-красный цвет, который по мере ее переваривания переходит в более темный. Щитки на теле небольшие, кутикула эластичная, растягивающаяся при наполнении кишечника кровью. Хелицеры превращены в длинные желобчатые стилеты, которые складываются в трубочку и служат для прокола кожи.
Клещи нападают на птиц обычно ночью, а днем прячутся в щелях насестов и стен, в сухом помете. Спариваются голодные клещи. Затем самка, насосавшись крови, прячется в щели и по мере ее переваривания, через сутки-двое начинает класть яйца, число которых зависит от количества поглощенной крови (обычно до 20). Из яиц выходят беловатые, непитающиеся личинки, которые, линяя, вскоре превращаются в протонимф. Последние сосут кровь и превращаются в дейтонимф, которые, снова нападая на птиц, питаются и превращаются во взрослых клещей. Цикл развития при +25°С занимает всего около полутора недель. Куриный клещ, размножаясь в птичниках в несметных количествах, особенно летом, сильно вредит в птицеводстве. Птицы становятся беспокойными, плохо растут, молодняк нередко гибнет. В сильно зараженном птичнике в насестах и под обшивкой стен можно найти сплошную копошащуюся массу клещей. Тут яйца, личинки, нимфы, взрослые клещи, голодные и с кровью, в разной степени переваренной, линечные шкурки. Клещи могут долго голодать, сохраняясь в щелях зараженных птичников, и нередко нападают на людей, вызывая сильное раздражение кожи. Зимой в летних птичниках, откуда птиц переводят в теплые помещения, клещи находятся в замерзшем состоянии, но, если принести их в тепло, они оттаивают и оживают. Куриного клеща уничтожают различными ядами. ДДТ малоэффективен, хорошо действует опрыскивание препаратами хлорофоса.
В гнездах диких птиц чаще встречается птичий клещ (Dermanyssus hirundinis), в массе размножающийся в период гнездования в гнездах закрытого типа, особенно в искусственных гнездовьях — скворечниках, синичниках. После вылета птенцов клещи остаются в гнездовьях и живут, не питаясь много месяцев; перезимовывают главным образом голодные протонимфы, которые нападают весной на прилетевших птиц. Некоторые виды переходят к жизни в оперении птиц, например, воробьиный клещ (Dermanyssus passerinus) летом встречается в гнездах, а зимой размножается на теле воробьев.
Гнездовых паразитов рода Dermanyssus напоминает по биологии мышиный клещ (Allodermanyssus sanguineus, табл. 10, 2), паразит домовой мыши, который встречается у нас в южных районах, размножается в жилье и может нападать на человека.
Примером внеубежищных паразитов служат некоторые дерманиссиды рептилий и крысиный клещ (Ornithonyssus bacoti, рис. 77). Последний, по происхождению тропический вид, расселился со своим хозяином крысой-пасюком по всему свету. Крысиный клещ теплолюбив и в умеренных широтах ведет себя как синантроп. Он поселяется в жилых и хозяйственных помещениях и держится в утепленных местах, у радиаторов отопления, печей и т. п. Подстерегание хозяина вне гнезда сильно снижает вероятность встречи с ним, в связи с чем у этих паразитов выработался ряд приспособлений. Цикл развития таков, что для его завершения клещу достаточно только дважды напасть на хозяина. Насосавшаяся крови самка откладывает яйца где-нибудь в укромном месте. Личинки, не питаясь, превращаются в протонимф, которые нападают на крыс, сосут кровь и превращаются в дейтонимф. Последние, подобно личинкам, не питаются и, перелиняв, становятся взрослыми клещами (рис. 74, 6). При этом женские протонимфы (будущие самки), попав на хозяина, сосут кровь дважды и уходят с него, а мужские протонимфы (будущие самцы) обычно сосут только раз и остаются в шерсти крысы вплоть до превращения в самцов. Последние отыскивают в шерсти животного упитанных женских дейтонимф, прицепляются к ним и сидят на них до тех пор, пока дейтонимфа не превратится в самку, после чего происходит оплодотворение. Оплодотворенная самка снова нападает на хозяина. Клещи могут долго голодать, а попав на животное, высасывают сразу много крови. Их хелицеры имеют откидной подвижный палец и фиксируются в коже животного, как гарпун. Питание и созревание яиц строго согласованы, полностью насосавшись, самка по мере переваривания крови откладывает до 20 яиц одной кладкой. Такая ритмическая согласованность питания и размножения у кровососущих членистоногих получила название гонотрофической гармонии, а повторные насасывания крови с последующей откладкой яиц называют гонотрофическими циклами. Редкость встреч с хозяином компенсируется у крысиного клеща и способностью сосать кровь не только крыс, но и других животных. Нередко этот клещ нападает и на человека.
Признаки внеубежищного паразитизма среди дерманиссид еще более выражены у паразитов рептилий — бездомных хозяев, которых клещи подстерегают в открытой природе: ящеричного клеща (Sauronyssus saurarum) и особенно змеиного клеща (Ophionyssus natricis, рис. 76, 7). Последний присасывается между брюшными чешуями у змей, самка за один прием высасывает количество крови, в десятки раз превышающее ее вес в голодном состоянии, и затем откладывает до 30 яиц. При содержании змей в неволе змеиные клещи нередко размножаются в несметном количестве и заедают змей насмерть. В ограниченном объеме садка, где змеям некуда деваться от паразитов, реализуется громадная потенциальная размножаемость этих внеубежищных по природе клещей.
Переход к постоянному паразитизму наблюдается в разных группах дерманиссид, например у клещей рода Hirstionyssus, паразитирующих на мелких млекопитающих. Часть видов этого обширного рода преобладает в гнездах, нападая на животное только на время сосания крови, другие задерживаются в шерсти или живут и размножаются на хозяине. При этом наблюдаются приспособления, сходные с теми, которые мы видели у клещей рода Laelaps, и направленные против механического воздействия хозяина: защитные щитки, сильные ноги с прицепными коготками, живорождение вплоть до дейтонимфы (рис. 74, 5) и др. В отличие от подстерегающих паразитов виды, живущие на хозяине, сосут кровь часто и небольшими порциями, не раздуваясь сильно, причем параллельно по одному развиваются зародыши и рождаются молодые формы, т. е. нет гонотрофической гармонии, о которой говорилось выше. Виды Hirstionyssus, подобно Laelaps, в большинстве приурочены к определенным хозяевам, например: Hirstionyssus sciurinus живет на белках, Hirstionyssus eusoricis — на землеройках, Hirstionyssus talpae — на кротах, Hirstionyssus criceti — на хомяках и т. д. Даже видовые названия этих паразитов даны по названиям хозяев: Sciurus — белка, Sorex — землеройка, Talpa — крот, Cricetus — хомяк и т. д.
Наиболее специализированные постоянные паразиты среди гамазид — это клещи семейства Spinturnicidae, живущие на рукокрылых, и полостные паразиты, обитатели дыхательных органов наземных позвоночных. Спинтурнициды (табл. 10, 3) — довольно крупные клещи с маленьким туловищем, большими плоско расставленными ногами, крепкими коготками и гарпуновидными хелицерами, фиксирующимися в коже летучей мыши. Они держатся обычно на летательной перепонке. Самка рождает дейтонимфу, которая похожа на взрослого клеща. Она тут же прицепляется к перепонке крыла, начинает сосать кровь и через некоторое время превращается во взрослую форму.
Полостные паразиты представлены тремя семействами: клещи семейства Entonyssidae живут в воздушных мешках змей, семейства Rhinonyssidae (рис. 76, 9) — в носовой полости птиц, семейства Halarachnidae (рис. 76, 8) — в трахеях и легких млекопитающих. Сходство жизненного цикла, наличие непитающейся дейтонимфы (рис. 74, 6) показывают, что эти клещи произошли от внеубежищных подстерегающих дерманиссид (Ornithonyssus и ближайшие роды). Среди последних есть формы, переходящие к постоянному наружному паразитизму, через который, очевидно, произошел переход к жизни в дыхательных путях. Скрытый образ жизни отразился в облике полостных паразитов. У взрослых клещей недоразвиты щитки, щетинки, перитремы дыхалец и другие имагинальные структуры, и взрослые клещи уподобляются протонимфам (педоморфоз). В то же время они обладают крепкими ногами с коготками, удерживающими клеща на слизистой оболочке; хелицеры колющие. Взрослые клещи и протонимфы сосут кровь, личинки и дейтонимфы не питаются. Самки кладут яйца или рождают молодь. Наиболее многочисленны ринониссиды птиц, изучение которых только начато, но уже известно до 300 видов. Обилие видов объясняется специфичностью этих паразитов. Каждый вид приурочен к одному или немногим обычно родственным видам птиц. Изучение распределения этих клещей по хозяевам может дать ценные сведения о родственных отношениях и эволюции пернатых. Клещи семейства Halarachnidae найдены в дыхательных органах обезьян, собак, ластоногих и некоторых грызунов.
В организме гамазоидных клещей не раз обнаруживались возбудители различных инфекционных болезней человека и животных, в связи с чем некоторые исследователи считают их переносчиками вирусов различных энцефалитов, геморрагических лихорадок, риккетсиозных тифов, туляремии, чумы (клещи родов Dermanyssus, Ornithonyssus, Hirstionyssus, Haemogamasus, Laelaps и др.). Однако роль гамазид как переносчиков этих болезней недостаточно изучена. Достоверно известно, что обитающие в жилье мышиный и крысиный клещи передают человеку от грызунов везикулезный риккетсиоз — острое лихорадочное заболевание, у нас встречающееся на Украине.
В связи с выясняющимся медицинским значением гамазиды интенсивно изучаются у нас и за рубежом. Число паразитических видов, обнаруженных в СССР, приближается к 200. Н. Г. Брегетовой, А. А. Земской, А. Б. Ланге составлены определители и написаны очерки по биологии и медицинскому значению этих клещей (в изданиях АН СССР, 1955, 1956; Медгиз, 1958, 1962 и др.).
Клещи-уроподы (надсем. Uropodoidea) — мелкие или более крупные формы (до 2 мм) с очень твердым имагинальным панцирем. Тело обычно щитообразное, выпуклое сверху и плоское снизу, овальное или грушевидное, с выступающим передним краем. Иногда панцирь покрыт сложной скульптурой (табл. 8, 14). Есть круглые в очертании совсем плоские формы, у которых край панциря образован слившимися уплощенными щетинками. Ротовые органы спрятаны в углублении панциря (камеростоме) и прикрываются снизу плоскими тазиками передних ног. Хелицеры обычно длинные, с небольшой колющей клешней. Ноги короткие, могут плотно прижиматься к телу и вкладываются в углубления на нижней стороне, что наблюдается при замирании (танатозе), очень характерном для этих медлительных клещей. Стигмы с небольшими искривленными перитремами. Половое отверстие расположено в средней части тела, у самца небольшое, зияющее, у самки прикрыто твердым щитком. Уроподы кладут яйца. У личинки и протонимфы на теле несколько щитков, которые у дейтонимфы сливаются; при превращении во взрослого клеща панцирь сильно уплотняется.
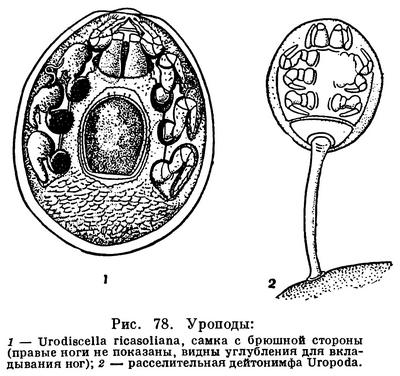
Уроподы живут в почве, лесной подстилке, навозе, много среди них мирмекофильных видов — специфических обитателей муравейников. Питание разнообразно: хищники, высасывающие почвенных нематод, личинок насекомых; формы, питающиеся органическими остатками; есть виды, сосущие сок растений и повреждающие рассаду в парниках. Некоторые имеют расселительных дейтонимф. Последние прикрепляются к насекомым, чаще жукам, с помощью секрета, выделяемого из анального отверстия и застывающего в виде гибкого стебелька (рис. 78, 2).
Уроподы изучены недостаточно. Насчитывается до 15 семейств этих клещей. Среди них наименее специализированные формы позволяют установить общность происхождения уропод и гамазоидных клещей. У представителей некоторых близких к уроподам семейств частично еще сохранилась членистость брюшка (сем. Microsejidae и др.).