(Branchiopoda Latreille, 1817) Класс Бранхиоподы, Жаброногие, Жаброногие ракообразные, Class Branchiopoda Latreille, 1817 (Branchiopods, Phyllopods) 3 отряда
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ПОДКЛАСС ЖАБРОНОГИЕ РАКООБРАЗНЫЕ (BRANCHIOPODA)
Жаброногие ракообразные считаются самыми примитивными представителями всего класса ракообразных. Их голова не срастается с грудными сегментами. Грудные ножки листовидные, слабо хитинизированные и служат одновременно для передвижения, дыхания и направления пищи ко рту. При их взмахах создается только один ток воды, который используется и для движения, и для питания, и для дыхания.
Число грудных сегментов непостоянно, строение половой, нервной и кровеносной систем примитивнее, чем у других ракообразных.
Жаброногие обитают преимущественно в пресных водах, морские виды немногочисленны и имеют пресноводное происхождение.
Все жаброногие ведут свободный образ жизни, среди них нет ни паразитов, ни сидяче-прикрепленных видов. Они не выработали приспособлений к существованию в подземных водах и на больших океанических глубинах.
В этот подкласс объединяются два отряда ныне живущих ракообразных — жаброноги (Anostraca) и листоногие (Phyllopoda).
ОТРЯД ЖАБРОНОГИ (ANOSTRACA)
Бурное таяние зимних снегов закончилось. На полях и в лесах еще остались лужи, заполненные талыми водами. Эти лужи недолговечны: через несколько дней они высохнут. Они существуют настолько непродолжительное время, что не успевают заселиться обычными пресноводными животными. Тем удивительнее наблюдать в некоторых из них сравнительно крупных (около 1 см длиной), изящных, полупрозрачных рачков, плавно двигающихся в воде брюшной стороной вверх. Это — жаброноги, принадлежащие к одной из наиболее примитивных групп современных ракообразных.
Жаброноги обитают не только во временных весенних водоемах. Условия их существования весьма разнообразны. Среди них есть виды, живущие круглый год в мелких холодных озерах Арктики, виды, населяющие слабосолоноватые водоемы степей и полупустынь, есть виды, приуроченные только к высокогорным водоемам, и, наконец, один вид замечателен тем, что в огромном количестве развивается в пересоленных, даже в самосадочных озерах; он выдерживает более высокую концентрацию солей, чем все остальные многоклеточные животные. Всего известно около 180 видов жаброногов.
Казалось бы, что при такой способности приспосабливаться к самым различным условиям среды, жить при высокой и низкой температуре и солености, жаброноги должны быть очень обычными и широко распространенными животными. В действительности это далеко не так. Они встречаются сравнительно редко и в очень немногих водоемах.
В чем же дело? По-видимому, основным условием возможности их существования надо считать отсутствие их врагов и конкурентов. Как правило, в водоемах, населенных жаброногами, не бывает ни рыб, ни других сильных хищников. В процессе эволюции жаброноги выработали тонкие приспособления к жизни в крайних, неблагоприятных для других животных условиях и только таким образом смогли выжить в жестокой борьбе за существование, избегая столкновений с более высокоорганизованными животными.
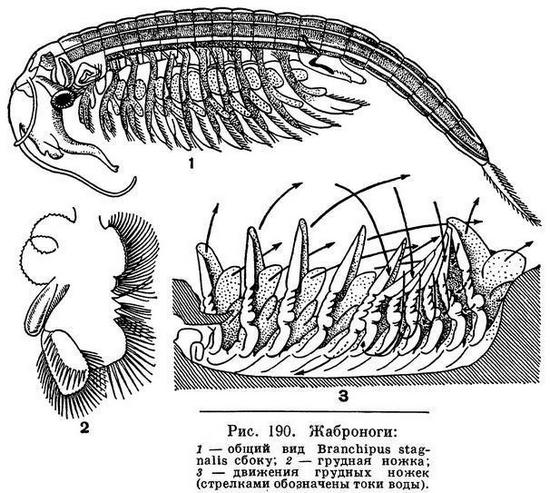
Тело жаброногов четко разделяется на голову, грудь и брюшной отдел. На голове можно заметить шов, отделяющий переднюю ее часть с антеннулами, антеннами и глазами от задней, несущей жвалы и челюсти. Передние антенны, или антеннулы, обычно довольно длинные, но не разделены на членики, задние гораздо более мощные, особенно у самцов. Самцы удерживают своими задними антеннами самок во время спаривания. У самцов некоторых жаброногов антенны снабжены многочисленными выростами и имеют весьма причудливую форму. По бокам головы располагается пара сидящих на стебельках сложных фасеточных глаз, а на лбу находится один непарный, так называемый науплиальный глаз.
Жвалы мощные, но лишены щупиков, а челюсти развиты сравнительно слабо. Перед жвалами находится имеющая форму козырька складка кожи — верхняя губа, задерживающая направляемый вперед грудными конечностями ток воды.
У большинства жаброногов грудь состоит из 11 и только у немногих из 17—19 сегментов. Каждый сегмент несет пару листовидных ножек. Все они устроены одинаково, и каждая имеет с наружной стороны 2—3 лопасти, служащие для дыхания, и одну лопасть, играющую роль весла, а с внутренней стороны 6 лопастей, служащих для направления тока воды вперед к ротовому отверстию (рис. 190, 2).
Между левыми и правыми ногами вдоль брюшной стороны тела проходит глубокая бороздка, по которой ток воды и устремляется ко рту.
Грудные ножки действуют последовательно. Каждая из них начинает изгибаться назад только после того, как это проделала предыдущая. Двигаясь назад, ножки отталкивают тело рачка от воды, а загибаясь вниз, они возбуждают ток воды, направляющийся между ногами к голове (рис. 190, 3). Таким образом, каждая ножка выполняет одновременно три функции: служит для передвижения рачка, направляет своими внутренними лопастями ток воды с пищевыми частицами ко рту и своими жаберными лопастями усваивает растворенный в воде кислород.
За грудью следует длинное цилиндрическое брюшко, в состав которого входят обычно 8 сегментов и концевая пластинка — тельсон. Первые два брюшных сегмента частично сливаются между собой. Здесь у самцов находятся два направленных вперед трубковидных выроста, играющих роль мужского копулятивного аппарата, а у самок к передним брюшным сегментам прикрепляется яйцевой мешок. С тельсоном сочленены ветви фурки, или вилочки, снабженные многочисленными щетинками.
Пищей жаброногам служат микроскопические водоросли и мелкие частицы разлагающейся растительности и другие органические частицы. Обитая всегда только в стоячих водоемах, жаброноги вынуждены весьма интенсивно фильтровать воду, пропуская ее через щетинки внутренних лопастей своих грудных ног и челюстей, с тем чтобы добыть себе достаточное количество пищи. Поэтому каждая грудная ножка совершает от 140 до 400 взмахов в минуту в зависимости от температуры. Собирающаяся перед верхней губой пища перетирается жвалами и только после этого поступает в рот. Железы верхней губы выделяют вещество, слепляющее мелкие пищевые частицы в пищевой комок.
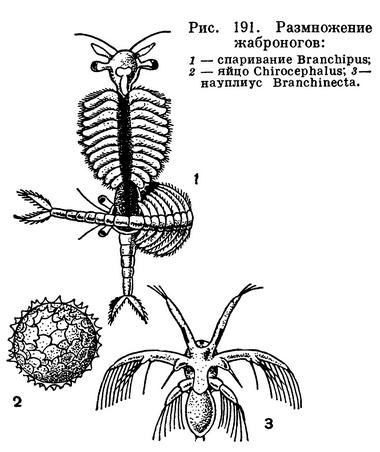
Когда наступает время спаривания, самец подплывает под самку и хватает ее задними антеннами. Удерживая ее этими: придатками, он изгибает свое тело так, чтобы его трубковидные отростки соприкасались с половым отверстием самки (рис. 191, 1). Сперма попадает в половые пути самки, где происходит оплодотворение яиц. Весь этот процесс продолжается несколько минут, после чего рачки расходятся. Яйца еще в половых путях самки окружаются желтком и покрываются очень плотной оболочкой, снабженной у некоторых видов шипами или бугорками (рис. 191, 2). Яйца выметываются самкой через 2—3 суток после спаривания и попадают в яйцевой мешок, но затем выбрасываются в воду, опускаются на дно и там развиваются.
Эти яйца отличаются необыкновенной стойкостью. Они переносят полное высыхание, резкие колебания температуры (в условиях опыта нагревание до 80°С). Они сохраняют жизнеспособность в течение 3—4 лет. Если водоем, в котором жили жаброноги, высох, яйца рачков могут остаться на месте в высохшем иле до следующей весны, когда это место снова заполнится водой. Вместе с пылью яйца могут переноситься ветром из водоема в водоем, что служит основным способом распространения жаброногов. Некоторые исследователи считали, что высыхание и промерзание необходимы для нормального развития жаброногов. Однако специально поставленные опыты не подтвердили этого предположения. В аквариумах, постоянно содержащих воду, отложенные жаброногами яйца развивались на следующий год после откладки вполне нормально.
Яйца испытывают полное дробление, и из них выходит первая личинка — науплиус (рис. 191, 3). В течение 2—3 недель происходит 7—8 линек личинки. С каждой линькой увеличивается количество сегментов тела рачка и относящихся к ним конечностей. Продолжительность жизни взрослых рачков зависит от температуры. Так, например, Chirocephalus grubei при 5,2°С живет 90 дней, при 7,9°С — 78 дней, а при 13,5°С — всего 5 дней.
По характеру размножения резко отличается от остальных жаброногов обитатель соленых озер артемия (Artemia salina). Во-первых, нередко мы находим в водоемах только самок артемий, которые откладывают яйца, способные развиваться без оплодотворения, т. е. партеногенетически. Наряду с этим столь же обычно присутствие самцов, спаривание их с самками и развитие оплодотворенных яиц, так же как у других видов. Во-вторых, в одних случаях самки артемий откладывают яйца подобно прочим жаброногам, а в других — рождают вышедших из яиц в яйцевых метках науплиусов. Способность выносить очень высокую концентрацию солей, изменчивость, связанная с жизнью в различных по солености водоемах, полиплоидность и важное практическое значение заставляют несколько подробнее остановиться на биологии этого рачка.
Артемия распространена очень широко, но приурочена к водоемам определенного типа и к определенным климатическим условиям. Она обитает в пределах степей и полупустынь Европы, Азии и Америки, в морских осолоненных лиманах и в соленых материковых озерах. Обычно она живет при солености от 40 до 230‰, но может встречаться и при более низкой (вплоть до полного опреснения), и при более высокой солености. Наиболее благоприятно для артемии преобладание в растворе ионов натрия, но необходимо присутствие и других солей.
Проф. Н. С. Гаевская содержала артемий в растворах химически чистых хлористого натрия, хлористого магния, сернокислого магния и некоторых других солей, причем оказалось, что в таких условиях рачки погибают через 16—44 часа, а в смеси этих солей живут гораздо дольше. В данном случае ядовитое действие какого-нибудь одного иона нейтрализуется действием его иона-антагониста.
В пресной воде большинство особей артемии погибает через 2—3 суток. Перед этим они прекращают размножаться.
Артемии, живущие при разной солености, заметно отличаются друг от друга по строению заднего конца тела. У рачков, существующих при очень высокой солености, ветви фурки совсем не выражены или если они есть, то очень маленькие, не отчлененные от тельсона и вооруженные всего 1—3 щетинками. Рачки из водоемов несколько меньшей солености имеют более сильно развитую и богатую щетинками фурку, и только у рачков из сравнительно слабо осолоненных водоемов фурка отчленена от тельсона и вооружена многочисленными щетинками (рис. 192). Кроме того, параллельно уменьшению солености изменяется соотношение длины грудного и брюшного отделов тела и степень развития задних антенн.
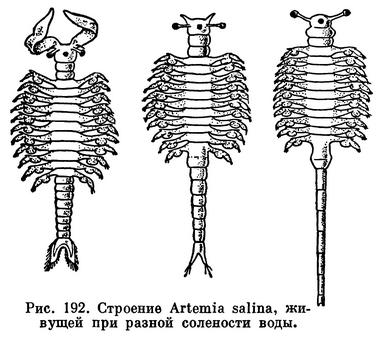
Раньше все эти формы считались особыми видами. Однако еще в 1875 и 1877 годах одесский биолог В. Шманкевич, воспитывая артемий при различной солености, показал, что именно соленость определяет эти морфологические особенности рачков. Он считал, что при уменьшении солености артемия приближается по своему строению к пресноводному роду Branchipus и даже переходит в него. В действительности такого перехода, конечно, не происходит. Просто артемии при различной солевой концентрации достигают половозрелости на разных стадиях развития — при более высокой солености на более ранних стадиях, чем при пониженной. Формы артемии, развивающиеся при пониженной солености, приобретают лишь поверхностное сходство с Branchipus.
Существуют ли в природе водоемы настолько высокой солености, что артемии не могут в них жить?
Некоторые наблюдения показывают, что, возможно, верхний солевой предел жизни артемии лежит около 300‰. Прежние исследователи Каспия, посещавшие огромный осолоненный залив этого моря Кара-Богаз-Гол, где в результате очень высокой концентрации солей идет выпадение мирабилита, были поражены обилием здесь артемий. Из-за массы рачков, кишевших в воде, она была красноватого цвета. После падения уровня Каспия обмелел пролив, соединяющий море с Кара- Богаз-Голом, и соленость залива еще повысилась. Живые рачки исчезли. Теперь на поверхности залива плавает масса яиц артемий, которые при такой высокой солености развиваться не могут.
Для выяснения особенностей физиологии артемии, позволяющих рачку переносить изменения солености от очень низких ее показателей до величины около 300‰, была изучена соленость крови артемии при помещении рачков в воду различной солевой концентрации. Обнаружилась удивительная картина: оказалось, что артемия сохраняет постоянную соленость крови независимо от колебаний солености во внешней среде. Эта неизменная концентрация солей в крови артемии такая же, как у других жаброногов, живущих в обычной пресной воде. Такое совпадение не случайно. Оно свидетельствует о пресноводном происхождении артемии и о вторичном характере ее приспособления к существованию в осолоненных водоемах.
Независимость солености крови артемии от солености внешней среды связана со способностью рачка выводить из организма поступающие в него соли, причем в разном количестве при жизни в разных концентрациях солей. Благодаря этому концентрация солей в крови артемии почти во всех населенных ею водоемах оказывается ниже, чем в окружающей среде. Таких совершенных способностей к регуляции солености крови мы не знаем у других беспозвоночных.
Основной пищей артемии служит несколько видов микроскопических водорослей, так же, как она, приспособленных к жизни в соленых водоемах, и, кроме того, бактерии грунта. В Сиваше, где артемий очень много, они поедают диатомовые (Amphora, Navicula) и зеленые (Dunaliella) водоросли. Если водорослей в воде недостаточно, рачки опускаются на дно, взмучивают придонную воду и отфильтровывают бактерий.
Есть указания на существование в некоторых соленых озерах в большом количестве только артемии и зеленой водоросли дуналиеллы. Рачки питаются дуналиеллой, а она, в свою очередь, усваивает некоторые продукты жизнедеятельности рачка, в частности соединения азота. Таким образом, в водоеме устанавливаются необыкновенно простые взаимоотношения между двумя организмами, составляющими единую пищевую цепь.
Дыхание артемий нередко бывает затруднено тем, что при высокой концентрации солей и малочисленности зеленых водорослей содержание кислорода в воде падает. Но в таких случаях артемия вырабатывает гемоглобин, растворенный в ее крови и связывающий значительное количество кислорода.
Период размножения артемий весьма продолжителен и определяется подходящей для этого температурой. Так, например, в Сиваше артемия размножается с мая до сентября — октября. При этом иногда она откладывает «зимние яйца», способные к развитию только на следующий год, а иногда рождает живых науплиусов, развивающихся без перерыва во взрослого рачка. Обычно рождение науплиусов происходит весной и летом, а откладка яиц осенью, поскольку такие яйца хорошо переносят неблагоприятные зимние условия, однако оба процесса могут происходить и одновременно. По наблюдениям А. Ф. Гунько, при резких изменениях температуры, солености и питания у артемии преобладает живорождение, а при постоянстве этих условий — откладка яиц. Замечательно, что оплодотворение не влияет на судьбу яйца: как оплодотворенные, так и неоплодотворенные, развивающиеся партеногенетически, яйца могут либо превращаться в зимние, либо сразу давать науплиусов. За один вымет самка производит до 70 яиц или науплиусов, а размножаться она может три раза в сезон, с промежутками от 6 до 28 суток. Через 10—16 суток, перелиняв за это время 7 раз, личинка превращается в молодого рачка, а через 20—29 суток после рождения, перелиняв еще 4—5 раз, рачок достигает половозрелости.
Замечательной особенностью артемии следует считать существование недавно открытых у нее полиплоидных рас. Полиплоидия, довольно обычное явление среди растений, в мире животных встречается очень редко. В Италии обнаружены ди-, три-, тетра-, пента- и октоплоидные расы артемии, т. е. расы, состоящие из рачков с двойным, тройным, четверным, пятерным и восьмерным набором хромосом. Среди диплоидных и тетраплоидных рас есть как способные к обоеполому размножению, так и состоящие только из размножающихся партеногенетически самок. Остальные расы включают исключительно партеногенетически размножающихся самок. В отличие от того что наблюдается у растений, расы артемии не различаются ни по размерам, ни по внешнему виду. К сожалению, это явление у артемии еще недостаточно изучено; в частности, остается неизвестным, отличаются ли эти расы артемии по жизнеспособности, отношению к солености и т. д.
Как уже указывалось, при благоприятных условиях артемия развивается в водоемах в массовом количестве. По подсчетам В. П. Воробьева, в Сиваше насчитывается в среднем 13,6 г артемии на 1 м3, что для всего залива составляет огромную величину — 14,8 тысячи тонн. Известно, что в отдельных случаях эти рачки служили пищей человеку. Так, например, американские индейцы, жившие на берегах Соленого озера в штате Ута (США), охотно ели артемию. Арабы, кочевавшие к западу от дельты Нила, вылавливали в соленых озерах артемий, приготавливали из них пасту, напоминавшую, по словам известного географа Петермана, соленую селедку, и охотно использовали эту пасту вместо мяса.
Широкое распространение получила артемия среди аквариумистов, которые выводят из яиц рачков, а затем скармливают их аквариумным рыбкам. Высушенные яйца артемий пересылаются по почте из страны в страну и служат предметом торговли. В последнее время и у нас начали их заготовку для экспорта за границу. Эти яйца массами выбрасываются волнами на берега Сиваша и других богатых артемией водоемов, и их сбор не представляет особых трудностей. Однако наиболее существенно значение артемии в рыбоводстве. Она представляет собой первоклассный, высокопитательный корм для молоди ценных промысловых рыб, разводимых на рыбоводных заводах, в первую очередь для осетровых.
Строительство электростанций на наших реках отрицательно влияет на запасы проходных рыб, т. е. таких, которые для нереста мигрируют в реки. Плотины преграждают рыбам путь на нерестилища. Для того чтобы сохранить запасы ценных промысловых проходных рыб, и прежде всего осетровых, необходимо в широких масштабах проводить их искусственное разведение на рыбоводных заводах. Самым трудным делом при этом оказалось снабжение вышедших из икры мальков живым кормом. Молодые рыбки должны некоторое время выкармливаться в специальных бассейнах, прежде чем они достаточно окрепнут, чтобы их можно было выпустить на волю. Они едят только живых животных, и артемия по своей калорийности и химическому составу оказалась одним из лучших видов корма для молоди осетровых.
В настоящее время разработаны и внедрены в производство методы разведения артемии в заводских масштабах. На Рогожкинском и Ачуевском рыбоводных заводах, в низовьях Дона, сооружены прямоугольные цементные бассейны размером 11,5х20х1 м каждый. В них налит рассол соленостью около 40—60‰, изготовленный из добытой на Сиваше соли. По мере испарения воды в бассейны подливается пресная вода. В качестве корма для рачков в бассейны периодически вносятся гидролизные дрожжи. В таких условиях артемии все летнее время производят живых науплиусов и развиваются в таком количестве, что их бывает достаточно для откорма молоди разводимых на этих заводах осетровых рыб. Осенью рачки дают покоящиеся яйца, а весной из них выходит новое поколение.
По мере необходимости артемий вылавливают из бассейнов с соленой водой и переносят в бассейны с пресной водой, в которых содержится молодь осетровых. В пресной воде рачки остаются живыми в течение нескольких часов и за это время усиленно поедаются осетрятами. Выкормленная на артемии молодь осетровых отличается быстрым темпом роста и высокими биохимическими показателями. Нет никакого сомнения в том, что разведение артемий будет организовано и на других многочисленных рыбоводных заводах нашей страны и что эти замечательные рачки сыграют важную роль в трудном деле сохранения запасов наших ценнейших промысловых рыб — осетровых.
ОТРЯД ЛИСТОНОГИЕ РАКООБРАЗНЫЕ (PHYLLOPODA)
Этот отряд отличается от отряда жаброногов присутствием головогрудного щита, или карапакса, который представляет собой разросшуюся назад двойную складку покровов заднего сегмента головы, покрытую хитином. У части листоногих карапакс имеет вид настоящего щита, прикрывающего тело сверху, у других этот щит как бы перегнулся так, что обе половины его превратились в створки двустворчатой раковины. Сложные глаза лишены стебельков и расположены непосредственно на поверхности головы. Грудные ножки листовидные и похожи на ножки жаброногов.
Листоногие обитают главным образом в пресных водах, морских видов очень мало. Отряд включает в себя три подотряда, различающихся строением, образом жизни и характером местообитания.
ПОДОТРЯД ЩИТНИ (NOTOSTRACA)
Вернемся к весенним лужам, в которых мы находили изящных полупрозрачных жаброногов. Если нам повезет, то в некоторых лужах мы обнаружим гораздо более крупных (до 5 см длиной) темно-коричневых рачков, то плавающих в воде, повернувшись брюшной стороной вверх, то опускающихся на дно и энергично взмучивающих поверхностный слой грунта. Нередко щитни попадаются в водоемах, казалось бы совсем не подходящих для жизни водных животных, например в заполненных грязной водой колеях проселочных дорог. Местные жители иногда наивно объясняют внезапное появление этих крупных рачков тем, что они падают из туч вместе с дождем.
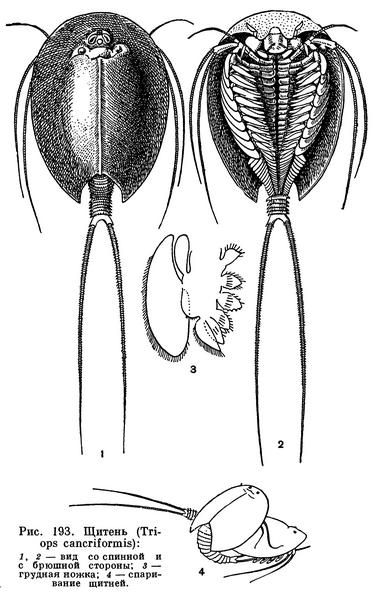
По своему строению щитни резко отличаются от жаброногов. Их плоский овальный щит покрывает голову, грудь и переднюю часть брюшка рачка. Близ переднего края щита на его поверхности находится глазной бугорок, на котором слева и справа расположено по темному, состоящему из многих фасеток сидячему (т. е. лишенному стебелька) глазу. Непосредственно перед сложными глазами между ними виден непарный науплиальный глаз, а за ними так называемый четырехклеточный орган. Функции этого органа еще не выяснены. Предполагается, что он служит органом внутренней секреции. По бокам головогрудного щита просвечивают крупные каналы выделительных желез, открывающихся при основании второй пары челюстей. Задний край щита имеет вид полукруглой выемки, оставляющей непокрытой заднюю часть брюшка. Оно заканчивается тельсоном, снабженным парой длинных членистых нитей — фуркой.
Рассматривая щитня с брюшной стороны, можно разобраться в строении его сегментов и конечностей. Головогрудной щит спереди загибается на брюшную сторону, и здесь с ним сочленяется большая, почти квадратная верхняя губа. Передние и задние антенны очень маленькие, зато жвалы крупные, с многочисленными зубцами. Позади верхней губы, между жвалами помещается рот, а позади жвал две пары маленьких пластинчатых челюстей (максилл). Далее следуют 10 грудных сегментов, каждый из которых несет по паре ножек. Эти ножки устроены приблизительно одинаково и во многом похожи на рассмотренные ранее ножки жаброногов. Они, так же как у жаброногов, имеют с внутренней стороны 6 лопастей, служащих для подачи пищи ко рту, а с наружной стороны — одну большую плавательную лопасть и одну (а не две, как у жаброногов) дыхательную лопасть, превращенную в жабру. Детальные исследования мускулатуры показали, что полного соответствия между листовидными ножками жаброногов и щитней нет, и возможно, что у тех и других листовидные ножки развились независимо, но функционируют они сходным образом, осуществляя одновременно дыхание, передвижение и подачу пищи ко рту.
Первая и в меньшей степени вторая пара грудных ножек щитня отличаются тем, что четыре внутренние их лопасти удлинены, расчленены и превращены в жгутики, выступающие наружу за края головогрудного щита. Эти жгутики принимают на себя функцию чувствительных органов и даже внешне напоминают жгуты антенн других ракообразных. Такое видоизменение внутренних лопастей передних грудных ножек щитней, несомненно, связано с редукцией их антенн.
На одиннадцатом сегменте туловища щитней открывается парное половое отверстие, и этот сегмент считается последним грудным. У самок его конечности имеют весьма своеобразное строение: большая наружная лопасть, служащая на других ножках для плавания, на одиннадцатой паре ног превращена в круглую яйцевую капсулу, в которой откладываются и вынашиваются яйца. У самцов ножки одиннадцатой пары не отличаются от остальных.
В строении щитня наблюдается еще одно удивительное явление. На каждом сегменте, начиная с тринадцатого, имеется не одна, а от 4 до 6 пар ног. Поэтому общее число пар ног рачка нередко достигает 70. Таким огромным количеством ног не обладает ни одно другое ракообразное.
Размеры ножек постепенно уменьшаются по направлению спереди назад, а задние сегменты тела совсем лишены ножек.
Существенным отличием ножек щитней от ножек жаброногов следует считать присутствие у первых на основной части передних конечностей направленных внутрь шипов. Это связано со способностью щитней захватывать шипами сравнительно крупные пищевые частицы и затем передавать их от ноги к ноге вперед, ко рту. Фильтровать мелкие взвешенные в воде частицы, как это делают жаброноги, щитни, по-видимому, не могут. Их задние ножки совершают преимущественно дыхательные движения. Удается наблюдать, как во время недолгих остановок рачка задние его ножки продолжают действовать, а передние остаются неподвижными. При плавании щитня все его многочисленные ножки последовательно сгибаются и разгибаются так, как будто по ним пробегает волна. Шведский зоолог Лундблад пипеткой вносил около задних ног щитня немного воды, подкрашенной кармином. Эта вода медленно двигалась вперед по желобку между задними ножками рачка, но как только она достигала уровня десятой пары ног, она начинала струиться вперед, что говорит о важном значении передних ножек в процессе подачи пищи ко рту.
При плавании щитней важную роль играет зрение. Освещение дна аквариума в темной комнате заставляет рачков немедленно повертываться брюшной стороной вверх: при таком положении расположенные на спинной стороне тела глаза воспринимают свет. Тот же эффект получается даже после того, как сложные глаза покрывают непрозрачным лаком и рачок лежит брюшной стороной на дне аквариума. По-видимому, он реагирует на свет благодаря тому, что глазной бокал науплиального глаза обращен вниз и продолжается в прозрачный соединительнотканный канал, который заканчивается на брюшной стороне тела, перед верхней губой, лишенным пигмента «окошком». Благодаря этому щитень может одновременно воспринимать свет, направленный как сверху, так и снизу.
Поиски пищи осуществляются не с помощью зрения, а с помощью химического чувства, органами которого служат жгутовидные внутренние отростки первой пары грудных ног. Щитень легко находит в аквариуме и поедает куски тела земляного червя, но, если к такому куску добавлен хинин, рачок чувствует добавку жгутами и отказывается от невкусной пищи.
В естественных условиях щитни питаются частицами грунта, растениями и мелкими животными. Опускаясь на дно, они взмучивают грунт и при помощи внутренних лопастей передних ног гонят воду с частицами грунта по срединному желобку вперед ко рту. В пищу используются сравнительно крупные частицы, а мелкая взвесь обратным током воды направляется назад. Наряду с этим щитни нападают на жаброногов, которых истребляют беспощадно, на хирономид и других мелких насекомых, не имеющих твердых хитиновых покровов, и даже на головастиков лягушек и на мальков рыб. Наконец, они могут обгрызать нежные части растений.
С этим связаны сравнительно немногочисленные случаи нанесения щитнями вреда хозяйству человека. На рыбоводных заводах на Дону после заливания предназначенных для выращивания мальков осетровых бассейнов в них появлялись массы щитней, которые набрасывались на рыбок и поедали их. Есть также указания на то, что щитни размножались на залитых водой рисовых полях и объедали побеги риса.
В наших широтах, а также на севере щитни размножаются преимущественно при помощи неоплодотворенных яиц. Население наших прудов и луж состоит почти исключительно из самок щитней. На 1000 самок приходится не более 10 самцов, а иногда их не бывает совсем. По направлению с севера на юг относительное количество самцов увеличивается, и в тропиках самцы по числу особей превосходят самок.
Спаривание щитней из-за редкости самцов удалось наблюдать только дважды (рис. 193, 4). Самец сверху прикреплялся к самке и резко сгибал свое тело так, что значительная его часть оказывалась под телом самки. Своей одиннадцатой парой ног он удерживал одиннадцатую пару ног самки, и, вероятно, в этот момент его сперма попадала в половые пути самки.
Как оплодотворенные, так и неоплодотворенные яйца откладываются самкой в уже упоминавшуюся яйцевую камеру, образованную наружными лопастями одиннадцатой пары ног. Там яйца находятся короткий срок, а затем выпадают на дно водоема. Интересно, что никаких отличий в строении, жизнеспособности и развитии оплодотворенных и неоплодотворенных яиц заметить не удается.
Эти мелкие яйца снабжены очень прочной оболочкой. Они переносят высыхание, промерзание и сохраняют жизнеспособность в течение 7—9 лет, в сухом состоянии выдерживают повышение температуры до 80° С. Без всякого вреда для себя они проходят через кишечник лягушки. Они легко переносятся ветром, причем, по-видимому, даже на очень далекие расстояния. Благодаря таким качествам своих яиц щитни широко распространены по всему свету. Внезапное их появление в каком-нибудь мелком водоеме объясняется попаданием в него их покоящихся яиц, а вовсе не выпадением взрослых рачков из дождевых туч, как думают жители некоторых районов.
В случае попадания яйца в подходящие условия на следующий год из него выходит личинка. У широко распространенного щитня Triops cancriformis эта личинка — типичный науплиус, с тремя парами конечностей и без всяких следов сегментации. У других видов щитней личинка выходит из яйца на более поздней стадии — метанауплиуса, характеризующегося сегментацией заднего конца тела. Для достижения половозрелости Triops cancriformis должен слинять около 40 раз, причем эти линьки быстро следуют одна за другой и при летней температуре все развитие от науплиуса до взрослого рачка заканчивается в течение двух недель. Весенний щитенъ (Lepidurus apus) после 17 линек достигает в длину 12 мм, причем после 14-й линьки у него появляются половые различия.
Несмотря на очень широкое распространение в стоячих пресных водоемах всего земного шара от приполярных озер до жарких тропических луж, щитни представлены чрезвычайно малым количеством видов. Современные зоологи, учитывая изменчивость этих ракообразных, различают всего 9 видов щитней — 4 вида рода Triops и 5 видов рода Lepidurus. Некоторые из них, как, например, Triops cancriformis и Lepidurus apus, встречаются почти повсеместно, другие, как, например, Lepidurus arcticus, приурочены только к водоемам Крайнего Севера. Есть виды, обитающие только в Африке, и виды, живущие только в Австралии.
Чрезвычайно интересной особенностью щитней следует считать их необыкновенную геологическую долговечность. Современный вид Triops cancriformis был обнаружен в ископаемом состоянии в триасовых отложениях Германии. Детальный анализ многочисленных отпечатков хорошо сохранившихся рачков и их конечностей показал, что триасовый щитень вполне сходен с современными. При этом он жил в водоемах такого же типа, какие и сейчас населены щитнями. Современные африканские виды щитней также были найдены в ископаемом состоянии, но в юрских и в более молодых отложениях.
Мы не знаем других подобных случаев существования современных видов в столь отдаленные от нас геологические периоды. Щитней можно с полным правом называть «живыми ископаемыми».
Как можно представить себе неизменность вида на протяжении 200 миллионов лет, отделяющих нас от триасового периода? Английский ученый Лонгхерст предполагает, что это объясняется главным образом постоянным разносом покоящихся яиц щитней на огромные пространства, благодаря чему не создается изоляции отдельных поселений рачков. Однако следует учесть и другие не менее важные биологические особенности щитней, ставящие их в особое положение в борьбе за существование. В период их активной жизни в населяемых ими водоемах нет ни их врагов, ни их конкурентов. К тому же естественный отбор действует на активных рачков всего в течение каких-нибудь двух недель на протяжении целого года. Все остальное время рачки пребывают в стадии покоящихся яиц. Совершенно очевидно, что существование видов щитней зависит в первую очередь от стойкости таких яиц. Если допустить, что их яйца приобрели свои замечательные свойства еще за несколько миллионов лет до наших дней, а рачки в активном состоянии не подвергаются естественному отбору, то становится понятной поразительная консервативность видов этих удивительных ракообразных.
ПОДОТРЯД КОНХОСТРАКИ (CONCHOSTRACA)
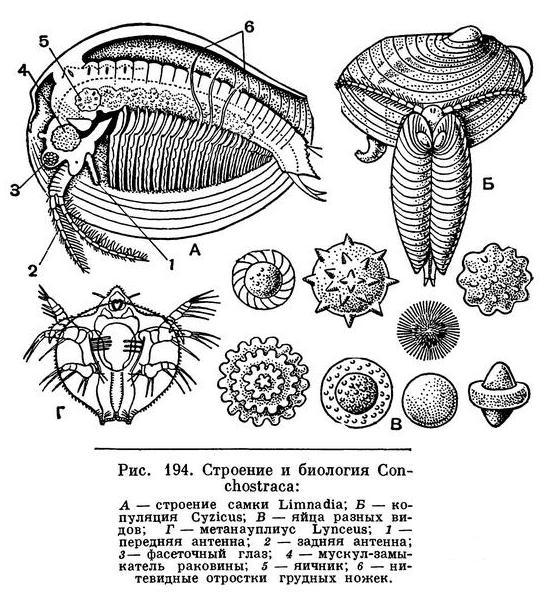
В мелких, иногда пересыхающих водоемах как средней, так и северной части нашей страны, а нередко и высоко в горах обитают рачки, менее заметные и не достигающие столь крупных размеров, как щитни. Они обычно окрашены в буроватые, розоватые или зеленоватые тона. Все тело их покрыто двустворчатой хитиновой раковинкой, которая у большинства видов имеет концентрические полосы прироста, несколько напоминающие кольца на раковинах двустворчатых моллюсков, например беззубки и перловицы. Эти полосы у рачков возникают после каждой линьки, поскольку при линьке наружная часть старой раковины сохраняется, а затем раковина начинает расти по периферии. Число полос соответствует числу линек.
Обе створки скреплены одним поперечным сильным мускулом-замыкателем. Рачки реагируют на опасность, сокращая этот мускул, причем створки раковины плотно сжимаются и рачок, как мертвый, остается неподвижно лежать на дне.
Раскроем створки раковины, чтобы рассмотреть строение тела рачка. Голова многих видов уплощена сверху вниз. Сидячие фасеточные глаза сближены и расположены на лобной ее части, а у одного рода (Cyclestheria) даже соединяются между собой. Перед ними находится простой науплиальный глазок. Передние антенны развиты слабо и не расчленены. Зато задние антенны длинные, двуветвистые, причем обе ветви снабжены многочисленными щетинками. Рачки плавают преимущественно за счет взмахов задних антенн, действующих как весла. Лишь у очень обычного рода Lynceus, способного широко раскрывать створки своей шаровидной раковины, не только задние антенны, но и грудные ножки совершают веслообразные движения.
Число сегментов, составляющих грудной отдел, колеблется от 10 до 32, и соответственно этому изменяется число пар грудных ножек. Они имеют приблизительно такое же строение, как у щитней, и их размеры также постепенно уменьшаются по направлению спереди назад. У самцов часть внутренних лопастей первой, а иногда и второй пары грудных ног преобразована в крючковидные придатки, которыми самец удерживает самку при спаривании (рис. 194, Б). У самок наружные лопасти нескольких средних ног превращены в длинные, загнутые вверх нитевидные отростки, приспособленные для удерживания яиц на спинной стороне тела под раковиной. Однако основные функции грудных ног — это подача пищи вперед ко рту и дыхание. Жабрами служат ближайшие к основанию наружные лопасти всех грудных ног. Для обеспечения этих функций грудные ножки все время находятся в движении, благодаря чему вода между створками раковины беспрерывно обновляется. Тело заканчивается крупным хитинизированным тельсоном, который или сжат с боков и в таком случае снабжен парой крупных когтей, либо сплющен в спинно-брюшном направлении и несет короткие коготки.
Большинство Conchostraca — донные животные. Наиболее крупные из них — Limnadia, достигающие в длину 17 мм, лежат боком на дне, Cyzicus роется в грунте, взмучивая ил движением брюшка и отчасти ножек. Представители этих и других родов плавают лишь в период спаривания. Только сравнительно мелкий (4—5 мм) Lynceus brachyurus, отличающийся шаровидной раковиной без полос прироста, обычно плавает спиной кверху, раскрыв створки раковины, и лишь изредка опускается на дно. Он питается главным образом планктонными организмами, в то время как Cyzicus ест грунт с находящимися в нем организмами, a Limnadia объедает микроскопические водоросли, находящиеся на дне и на поверхности водных растений. Крупные несъедобные частицы Limnadia склеивает специальным секретом, выделяемым железами ножек, и выбрасывает через заднюю щель своей раковины.
Размножение Conchostraca происходит по-разному. Все представители семейства Limnadiidae размножаются только партеногенетически, самцы среди них неизвестны. У других семейств самцы встречаются регулярно, причем, чем южнее обитают рачки, тем больше относительная численность самцов. При спаривании самец удерживает самку крючковидными отростками своих задних антенн и просовывает задний конец тела в заднюю щель между створками раковины самки. Яйца откладываются в пространство между спинной стороной тела самки и ее раковиной. У Cyclestheria часть задних грудных сегментов расширена таким образом, что перед ними образуется настоящая выводковая сумка. Проносив некоторое время яйца на спине под раковиной, самка линяет, и яйца выпадают на дно водоема.
Эти яйца окружены плотной и прочной оболочкой и способны переносить высыхание, замерзание и другие неблагоприятные условия. Нередко они имеют причудливую форму, у многих видов они снабжены выростами, способствующими их переносу ветром и различными животными (рис. 194, В). Яйца некоторых видов сохраняют жизнеспособность в течение 7 лет.
После того как яйцо попадет в благоприятные для его развития условия, из него выходит личинка, находящаяся на стадии науплиуса. Науплиус Conchostraca отличается очень маленькими передними антеннами. На следующей стадии — метанауплиуса — у личинки развивается зачаток раковины. У большинства Conchostraca эта зачаточная раковина уже двустворчатая и очень напоминает раковину ветвистоусых ракообразных (см. ниже). У Lynceus она имеет вид плоского щита, похожего на карапакс щитней (рис. 194, Г), и только на более поздних стадиях развития она перегибается пополам и становится двустворчатой. Все развитие происходит очень быстро. Так, например, Cyzicus достигает половозрелости через 19 дней после выхода личинки из яйца.
Современные Conchostraca не имеют никакого практического значения. Однако вымершие виды этих ракообразных привлекают к себе внимание геологов. В пресноводных отложениях, вообще очень бедных ископаемыми, хорошо сохраняющиеся раковинки Conchostraca встречаются довольно часто. По ним удается определять геологический возраст соответствующих слоев.
В предыдущие геологические периоды Conchostraca были, по-видимому, более многочисленны и шире распространены, чем ныне. Вымерших видов известно около 300, а современных вдвое меньше. Наиболее древние Conchostraca найдены в девонских отложениях. Замечательно, что среди древних вымерших видов, особенно триасовых, многие жили в море, где сейчас Conchostraca полностью отсутствуют.
ПОДОТРЯД ВЕТВИСТОУСЫЕ РАКООБРАЗНЫЕ (CLADOCERA)
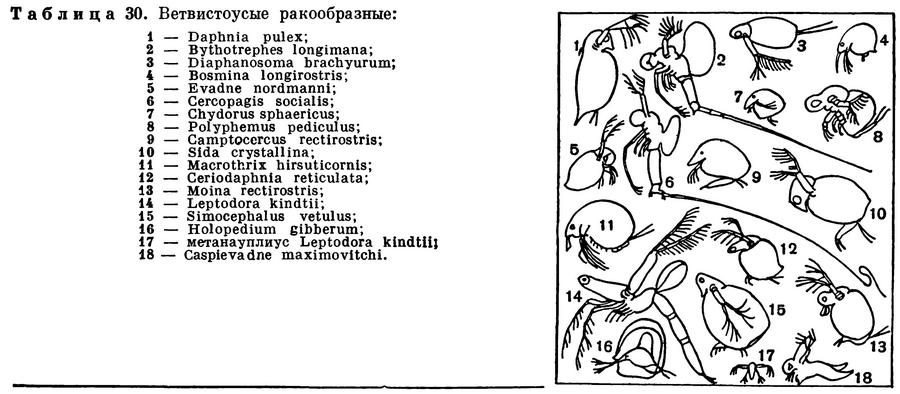
Почти в каждом пресноводном водоеме, начиная с огромных глубоких озер и кончая мелкими прудками, любой лов сачком или специальной сеткой из мельничного шелкового газа приносит десятки и сотни мелких рачков, часто называемых водяными блохами (табл. 30). Они оживленно снуют в толще воды, делая характерные взмахи своими задними антеннами, а некоторые из них передвигаются по дну или по подводным растениям. Их хорошо знают не только ученые-зоологи, но и все любители аквариумов, которые кормят ими, живыми или сушеными, своих рыбок. Их знают и рыбоводы, занимающиеся их разведением для кормления молоди рыб, выращиваемой на рыбоводных заводах. Многие из этих рачков всю жизнь проводят в толще воды и составляют значительную часть пресноводного планктона. Иногда они развиваются в таком количестве, что вода приобретает буроватый оттенок. Наряду с планктонными есть и донные виды. Есть и такие, которые то прикрепляются к растениям или опускаются на дно, то всплывают в толщу воды. Подавляющее большинство видов ветвистоусых обитает в пресных водах, но есть и несколько морских видов. Размеры этих рачков незначительны, обычно меньше 1 мм, редко до 5 мм и в исключительных случаях до 10 мм. Поэтому для изучения их строения приходится пользоваться микроскопом.
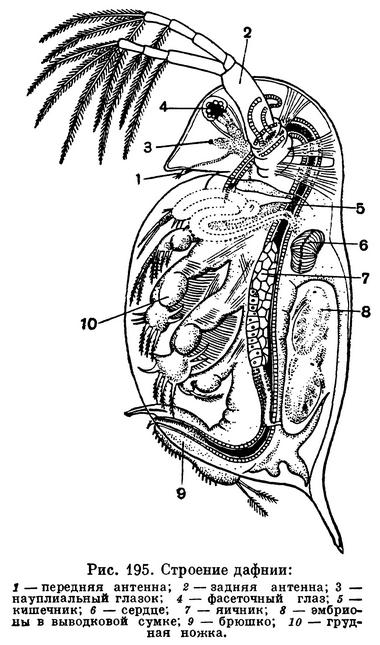
Тело большинства ветвистоусых ракообразных, например дафнии (рис. 195), заключено в хитиновую полупрозрачную двустворчатую раковинку, а голова вытянута в направленный вниз клюв, или рострум. На лобной части головы расположен непарный фасеточный глаз, возникший в результате слияния двух боковых глаз, а перед ним — простой науплиальный глазок, отсутствующий у некоторых видов.
Число фасеток, образующих сложный глаз, различно у разных видов и зависит от способа их питания. В глазу отфильтровывающей мелкие частицы дафнии насчитывается всего 22 фасетки, а у хищных Evadne и Leptodora соответственно 80 и 300. При нападении на мелких животных глаза хищных ветвистоусых имеют важное значение. Под рострумом прикрепляются палочковидные обыкновенно относительно очень маленькие передние антенны. Зато задние антенны непропорционально велики по сравнению с телом. Они двуветвисты, причем каждая ветвь снабжена длинными перистыми щетинками. Задние антенны служат основным органом передвижения ветвистоусых. Одновременно взмахивая обеими задними антеннами, рачки отталкиваются ими от воды и таким образом плывут короткими скачками. На нижней поверхности головы находится рот, окруженный спереди верхней губой, с боков жвалами и сзади челюстями.
Грудной отдел ветвистоусых укорочен и, судя по количеству пар ножек, состоит из 4—6 сегментов. Он, как уже указывалось, помещается внутри двустворчатой раковины, но у самок между спинной поверхностью тела и спинным краем раковины имеется обширная полость, выполняющая роль выводковой сумки. В эту сумку откладываются яйца, и при благоприятных условиях там же они развиваются. У хищных ветвистоусых раковина уменьшена в размерах, оставляет непокрытым почти все туловище и прикрепляется к его спинной стороне (табл. 30, 2, 5, 6, 8, 14, 18); в таких случаях она служит только для вынашивания яиц.
Строение грудных ножек различно у разных видов и зависит от характера их питания. Эти ножки в общем напоминают листовидные ножки жаброногов, щитней и Conchostraca. У большинства ветвистоусых ракообразных грудные ножки служат для отфильтровывания мелких взвешенных в воде частиц, которыми рачки питаются. В соответствии с этим лопасти грудных ножек снабжены многочисленными перистыми щетинками, образующими вместе фильтрационный аппарат. Ножки совершают до 300—500 взмахов в 1 минуту.
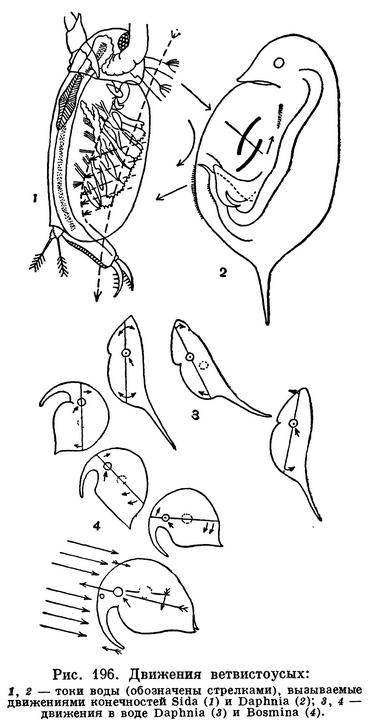
В простейшем случае, например у Sida (рис. 196, 2), вода входит между створками в передней части раковины. При одновременном движении всех 6 пар ножек вперед между ними возникает как бы ряд всасывательных камер, куда просачивается вода. При этом щетинки отфильтровывают всю мелкую взвесь. Движение ножек назад сжимает камеры и выгоняет воду через заднюю часть раковины. Отфильтрованные частицы направляются при помощи специальных щетинок в продольный желоб, расположенный на нижней стороне туловища между основаниями грудных ножек. По этому желобу находящимися на его стенках щетинками пищевой комок проталкивается вперед, по направлению ко рту.
У Daphnia этот аппарат усложнен и усовершенствован (рис. 196, 2). Из пяти пар ее грудных ножек фильтруют только третья и четвертая. При движении первой пары ножек вперед вода поступает в фильтрующую камеру. В это время третья и четвертая пары изгибаются вперед, обеспечивая дальнейший ток воды. Отфильтрованные пищевые частицы поступают в брюшной желобок. Пятая пара ног ограничивает сзади фильтрующую камеру. Делая взмах назад, она вместе с четвертой парой вызывает выводящий ток воды. Таким образом, между грудными ножками дафнии функции разделены, что говорит о большей их специализации.
Ближайшие к основанию наружные лопасти грудных ног фильтрующих ветвистоусых ракообразных превращены в жабры. По системе полостей кровь поступает в жабры и обогащается в них кислородом. В плазме крови ветвистоусых растворен гемоглобин, от которого в значительной степени зависит окраска рачков. Его концентрация меняется. Если содержание кислорода в воде низкое, кровь приобретает темно-красный цвет, если кислорода в воде много, кровь окрашена значительно светлее. При изменении содержания кислорода в воде меняется окраска рачков, но для этого требуется около 10 дней.
Дафнии с темно-красной кровью в воде с низким содержанием кислорода живут значительно дольше, чем дафнии, имеющие светлую, бедную гемоглобином кровь. Очевидно, гемоглобин, как и у позвоночных животных, связывает кислород.
Кровь приводится в движение мешковидным сердцем, взбалтывающим ее в полости тела (рис. 195, 6). Настоящих кровеносных сосудов нет. Сердце дафнии бьется очень часто. При температуре 10° С оно делает 150, а при температуре 20° С даже 500 ударов в минуту.
Совсем иначе устроены и действуют грудные ножки хищных ветвистоусых ракообразных (табл. 30). Их боковые лопасти недоразвиты или отсутствуют, жабр нет совсем, а хорошо развитые внутренние ветви снабжены сильными шипами и шиловидными щетинками. При помощи грудных ног рачки схватывают других мелких животных, служащих им пищей. Хищные ветвистоусые воспринимают кислород скоплением железистых клеток, находящихся на голове.
Задний брюшной отдел тела ветвистоусых укорочен, часто изогнут под прямым углом и может прятаться между створками раковины. Он обычно заканчивается двумя когтями. Однако у многих хищных видов задний отдел имеет форму трубки, сохраняет сегментацию и иногда продолжается в длинный шип, назначение которого остается непонятным.
Наблюдая за поведением ветвистоусых рачков в водоеме или в аквариуме, нетрудно убедиться в том, насколько различны повадки разных видов. Особенно многочисленны рачки, проводящие всю свою жизнь в толще воды. Планктонные дафнии и некоторые другие ветвистоусые после взмаха задними антеннами и вызванного этим движением скачка некоторое время неподвижно парят в воде. Благодаря тому что центр тяжести их тела находится ниже места прикрепления антенн, в момент удара антенн тело рачка наклоняется, а в период покоя снова выпрямляется. Распростертые антенны препятствуют погружению рачка (рис. 196, 3).
Планктонные ветвистоусые из рода Bosmina плавают иначе. Они беспрерывно машут своими задними антеннами, причем передняя часть их тела при каждом ударе несколько приподнимается. Однако длинные хоботообразные передние антенны служат балансиром, возвращающим тело в горизонтальное положение (рис. 196, 4). Если босмине отрезать передние антенны, она начинает беспомощно кувыркаться и теряет способность к плаванию.
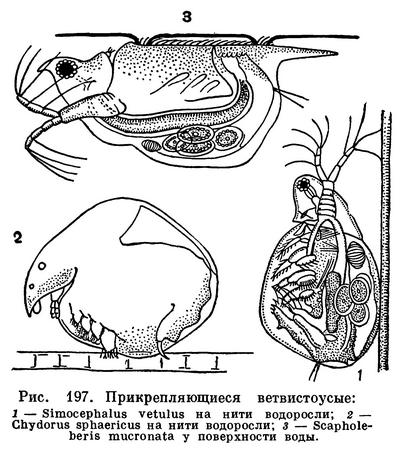
Среди зарослей подводных растений в прибрежной части озер и прудов часто попадается крупный (до 4 мм) ветвистоусый рачок Sida crystallina, который, как бы устав беспрерывно плавать, время от времени прикрепляется к растениям при помощи специальной присоски, находящейся на задней части его головы. Другой обитатель зарослей — Simocephalus — цепляется за растения щетинками задних антенн. Маленький шарообразный хидорус (Chydorus sphaericus), поплавав некоторое время, садится на водоросли и начинает ползать по ним, удерживаясь щетинками края раковины и отталкиваясь второй парой грудных ног и брюшком. Наиболее своеобразен способ передвижения Scapholeberis mucronata, который благодаря присутствию несмачиваемых щетинок на нижнем крае раковины может прикрепляться снизу к поверхностной пленке натяжения воды и быстро скользить по ней спинной стороной вниз (рис. 197).
Наряду с такими постоянно или временно плавающими формами существует большое количество видов, плавающих очень плохо и постоянно ползающих по дну. Некоторые из них способны даже зарываться в поверхностный слой ила.
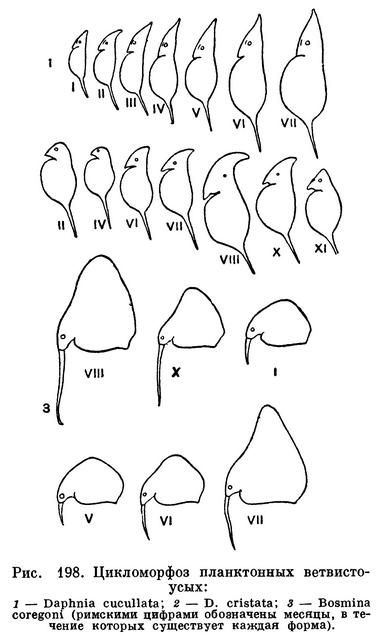
С жизнью многих ветвистоусых ракообразных в толще воды связано одно любопытное явление, получившее название цикломорфоза. Сущность его заключается в том, что форма и величина раковины определенных видов закономерно изменяются по сезонам года. В летнее время у некоторых видов дафнии на голове появляется так называемый «шлем», а у босмин на спинной стороне — «горб». Кроме того, у дафний удлиняется задний шип на раковине, а у босмин — передние антенны (рис. 198).
Эти изменения происходят параллельно изменению температуры воды. В данном случае важна, однако, не сама температура, а ее воздействие на плотность воды. Прогретая вода обладает меньшей плотностью, чем охлажденная. Для движения и парения в менее вязкой прогретой воде необходимы более длинные выросты, а иногда и иная форма тела. Поэтому летние формы дафний и босмин развивают специальные приспособления для передвижения и парения в таких условиях. Значение плотности воды для изменений формы тела рачков подтверждается наблюдениями Е. Ф. Мануйловой над дафниями озера Балхаш.
В этом озере соленость по направлению с востока на запад постепенно увеличивается и вместе с тем вода становится все более плотной. Местная планктонная дафния — Daphnia balchashensis, живущая при сравнительно высокой солености, имеет совершенно круглую голову, а живущая в опресненной части озера — голову, снабженную загнутым назад шлемом.
Большинство ветвистоусых ракообразных, как уже указывалось, питается, отфильтровывая мелкую, находящуюся в воде взвесь. Основной их пищей служат бактерии, одноклеточные водоросли и мелкие органические остатки — детрит. Наибольшее значение имеют бактерии. В опытах многих исследователей удавалось воспитывать ветвистоусых при кормлении их одними бактериями, а американский зоолог Банта получил в таких условиях около 1600 поколений дафний. В природных водоемах ветвистоусые прекрасно живут и размножаются при концентрации бактерий не менее 1 миллиона в 1 см3 воды. Если численность бактерий меньше 500 тысяч на 1 см3, наступление половозрелости дафний задерживается; если же численность бактерий падает ниже 200 тысяч на 1 см3, жизнь рачков становится невозможной. С другой стороны, избыточное количество бактерий (более 3—5 миллионов на 1 см3) также вредно влияет на рачков и вызывает прежде всего замедление или приостановку их размножения, а иногда даже их гибель. Это объясняется выделением бактериями продуктов обмена веществ, ядовитых для ветвистоусых.
От количества бактерий в водоеме зависит численность ветвистоусых. Неоднократно наблюдалось уменьшение числа ветвистоусых в период массового развития сине-зеленых водорослей. Некоторые исследователи предполагали вредное воздействие сине-зеленых на рачков. Однако, по данным Е. Ф. Мануйловой, бурное развитие сине-зеленых само по себе никакого влияния на рачков не оказывает. Оно подавляет развитие бактерий и именно таким образом косвенно воздействует на ветвистоусых. Часто наблюдающиеся периоды резкого увеличения численности ветвистоусых обычно следуют за периодами размножения бактерий.
Помимо бактерий, фильтрующие ветвистоусые используют в пищу одноклеточные водоросли. Они могут заглатывать только самые мелкие виды протококковых и зеленых водорослей, которые обычно развиваются в большом количестве в прудах, но не в крупных озерах. Обычные, особенно колониальные, планктонные водоросли слишком велики для того, чтобы быть проглоченными рачками. Во многих случаях водоросли приобретают большое значение для ветвистоусых не потому, что поедаются ими, а потому, что после отмирания вызывают массовое развитие бактерий — основного корма рачков.
При фильтрации пищи рачки не могут отсортировывать съедобные частицы от несъедобных. Они способны только отбирать частицы по их размеру, отбрасывая слишком крупные. С этим связаны, по наблюдениям В. М. Рылова, те изменения в составе планктона, которые наблюдаются при проникновении планктона стоячих водоемов в текучие. Эти изменения сводятся главным образом к резкому уменьшению численности ветвистоусых ракообразных, в то время как численность других планктонных животных, например коловраток, существенно не меняется. Оказывается, что речной поток несет мелкие минеральные частицы и попавшие в него планктонные ветвистоусые отфильтровывают их и заполняют ими свой кишечник. Рачки идут ко дну и погибают, а коловратки, захватывающие пищу иным способом, и притом еще более мелкую, остаются невредимыми.
Фильтрация пищи рачками происходит беспрерывно. Время, нужное для заполнения кишечника, колеблется от 10 до 240 минут и зависит от величины рачков, размеров пищевых частиц, их концентрации и температуры. Мелкая босмина в культуре водоросли Ankistodesmus наполняет кишечник в течение 10 минут, а в культуре бактерии Bacillus coli тратит на это 2—2,5 часа. В течение суток одна дафния поглощает от 4,8 до 40,8 миллиона бактерий (в зависимости от их концентрации). Даже после наполнения кишечника рачки продолжают фильтровать, и отфильтрованные частицы двигаются вперед по их брюшному желобку. Но в рот они не попадают: ротовые придатки и задний отдел тела выталкивают их снова в воду. Рачки начинают заглатывать пищу только после того, как кишечник хотя бы частично освободится от ее остатков.
Совсем иначе добывают себе пищу хищные виды. Самый крупный совершенно прозрачный ветвистоусый рачок Leptodora kindtii (табл. 30, 14) неподвижно парит в воде и своим громадным глазом высматривает добычу. Достаточно нескольких сильных взмахов его задних антенн для того, чтобы он настиг какого-нибудь более мелкого планктонного рачка или коловратку, схватил жертву своими далеко вынесенными вперед грудными ногами и вонзил в нее острые зазубренные жвалы. Приблизительно так же охотится и другой крупный хищник — Bythotrephes (табл. 30, 2). Оба рачка сначала размельчают жертву жвалами, а затем всасывают жидкую пищу. Они нападают почти на всех планктонных животных, включая собственную молодь, но не трогают сидячих и даже временно прикрепляющихся рачков, например сиду и симоцефалюса.
Лептодора и битотрефес весьма прожорливы. При температуре 20—25° С каждая лептодора съедает за сутки более 50, а битотрефес около 25 рачков. Соответствующие расчеты показывают, что в некоторых водоемах, например в Рыбинском водохранилище, хищные ветвистоусые выедают заметную часть зоопланктона (более 40% всех планктонных животных). Наносимый ими таким образом вред не так велик, поскольку они в свою очередь интенсивно потребляются многими рыбами.
В некоторых водоемах планктонные ветвистоусые ракообразные подобно другим планктонным животным совершают регулярные суточные вертикальные миграции. В темное время суток они поднимаются в поверхностные слои воды, а в светлое время уходят вниз. Однако во многих озерах и водохранилищах большая часть рачков круглые сутки остается близ поверхности.
До тех пор пока условия жизни ветвистоусых рачков благоприятны, в водоемах встречаются только самки, откладывающие и вынашивающие в своих выводковых камерах неоплодотворенные яйца. Число этих яиц различно не только у разных видов, но даже у различных особей одного вида. Разные виды дафний образуют до 50 или даже до 100 яиц на самку, босмины — до 16 яиц, а мелкие донные виды — не больше 2 яиц. Яйца развиваются в выводковой камере матери, где из них образуются сначала подвижные эмбрионы, а затем вполне сформировавшиеся маленькие рачки, покидающие выводковую камеру и приступающие к самостоятельному существованию. Летом, при достаточном количестве пищи, рождение молоди и появление в камере новых яиц происходят каждые 1—2 дня, благодаря чему численность ветвистоусых может очень быстро увеличиваться. Обыкновенно рождение молоди сопровождается линькой самки. У хищных ветвистоусых, например у лептодоры и битотрефеса, выводковая камера изолирована от окружающей среды и при выходе молоди каждый раз разрушается, а затем восстанавливается вновь.
Вышедшая на свободу молодь часто линяет и быстро растет. Первые три линьки происходят через 1—1,5 суток, а следующие через 2—3 суток. Большинство видов достигает половозрелости после третьей-четвертой линьки, т. е. через 2—6 дней после рождения. Так получаются новые поколения самок, продолжающие размножаться партеногенетически.
Но вот условия существования ветвистоусых рачков ухудшились вследствие понижения температуры, уменьшения количества пищи или загрязнения водоема. Это ухудшение сейчас же сказывается на судьбе яиц, находящихся в половых путях самки. Из неоплодотворенных яиц после их откладки в выводковую камеру выходят не самки, а самцы. Кроме того, часть яиц в половых путях самки подвергается второму делению созревания, в результате которого количество хромосом в яйце уменьшается вдвое. Такие яйца могут развиваться только после оплодотворения. При слиянии яйца и сперматозоида восстанавливается полное количество хромосом.
Замечательно, что даже кратковременные отклонения от нормальных условий жизни могут прервать процесс партеногенетического размножения. Судьба яйца определяется за 15 минут до его выхода из половой системы самки. Если в этот момент рачки подвергнутся каким-нибудь неблагоприятным воздействиям, из их яиц выведутся самцы или эти яйца будут непременно нуждаться в оплодотворении.
Самцы ветвистоусых ракообразных — карлики по сравнению с самками. Часто они в несколько раз меньше самок (рис. 199). Их передние антенны удлинены, а первая пара грудных ножек снабжена коготками, помогающими удерживать самку.
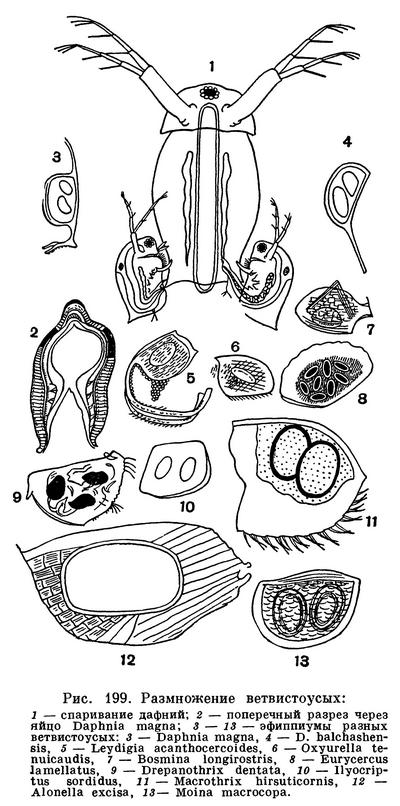
Обыкновенно самцы появляются в водоемах осенью, после значительного понижения температуры. Они прикрепляются к самкам при помощи коготков передних ног и при помощи передних антенн, причем у дафний нередко к одной самке прикрепляются два самца. Сперма проникает в выводковую камеру близ отверстий яйцеводов, а потом и в них.
Оплодотворенные яйца во многих отношениях отличаются от неоплодотворенных. Их гораздо меньше (редко больше двух), но зато они значительно богаче желтком и поэтому крупнее, чем партеногенетические. Вокруг них развиваются защитные оболочки. У Sida, Diaphanosoma, Holopedium и хищных видов оплодотворенные яйца снабжены двумя оболочками — плотной внутренней и клейкой наружной. Такие яйца откладываются в воду и опускаются на дно.
У большинства ветвистоусых оплодотворенные яйца откладываются в выводковую камеру, а в образовании их защитных оболочек принимает участие материнская раковина. Ее спинной край становится толще, под раковиной собираются клетки, плотным слоем окружающие яйцо или яйца. У дафний поверх этих клеток выделяется хитиновая оболочка и в выводковой камере возникает так называемое седлышко, или эфиппиум, содержащее обычно два яйца. Формирование эфиппиума продолжается 2—3 дня, и самка носит его в течение 1—2 дней. Затем она линяет, и эфиппиум оказывается на свободе. Благодаря присутствию воздухоносного слоя эфиппиумы дафний плавают на поверхности воды. У некоторых других ветвистоусых наружной оболочкой эфиппиума служит вся раковина или значительная ее часть, сброшенная при линьке (рис. 199).
Главная биологическая особенность оплодотворенных яиц заключается в том, что они могут развиваться только после более или менее продолжительного периода покоя и поэтому называются покоящимися. Такие яйца помогают рачкам переносить всевозможные неблагоприятные условия; они вмерзают в лед, высыхают, сохраняя в течение длительного времени жизнеспособность. В то же время они служат средством расселения ветвистоусых, легко переносясь из водоема в водоем ветром, вместе с илом, присохшим к лапкам водоплавающих птиц, и т. д. Благодаря своим покоящимся яйцам ветвистоусые широко распространены во всех стоячих водоемах нашей планеты.
Многие виды зиму проводят в стадии покоящегося яйца, которое начинает свое развитие весной и дает начало самке, приступающей к партеногенетическому размножению. У всех ветвистоусых, за исключением лептодоры, из яйца выходит вполне сформировавшийся рачок. Лептодора — единственный представитель ветвистоусых, имеющий личинку; она вылупляется из покоящегося яйца на стадии метанауплиуса (табл. 30, 17).
Число известных видов ветвистоусых достигает 380, в СССР найдено 164 вида. Некоторые из них распространены очень широко и встречаются в самых разнообразных водоемах, другие приурочены к определенным районам или населяют водоемы какого-нибудь особого типа. Во всех странах света обитают обыкновенная дафния (Daphnia pulex), планктонная дафния (Daphnia longispina), босмина (Bosmina longirostris), хидорус (Chydorus sphaericus) и многие другие. В то же время существуют виды, живущие только на Крайнем Севере (Daphnia arctica, Eurycercus glacialis). Преимущественно в северной части СССР и Америки встречается своеобразный Holopedium gibberum, имеющий поверх раковины студенистую оболочку, придающую рачку вид прозрачного шарика (табл. 30, 16). Этот рачок изредка встречается и в средней части СССР, в водоемах, связанных с моховыми болотами. Наконец, есть и тропические виды, в нашей стране обитающие на залитых водой рисовых полях Узбекистана, в Закавказье, в водоемах дельты Волги. Высказывается предположение о том, что их покоящиеся яйца переносятся на колоссальные расстояния воздушными потоками.
Некоторые ветвистоусые рачки населяют только те водоемы, вода которых имеет кислую реакцию и бедна минеральными солями, но большинство видов способно существовать в самых разнообразных, в том числе и неблагоприятных условиях, например в условиях недостаточного содержания кислорода в воде.
Немногочисленные морские представители ветвистоусых ракообразных также распространены очень широко, но приурочены преимущественно к прибрежным водам. Только в Каспийском море обитает несколько чрезвычайно своеобразных видов (табл. 30, 6, 18), частично присущих только Каспию, частично встречающихся также в Азовском море и в опресненных районах Черного моря. Среди них особенно причудливы виды рода Cercopagis, снабженные необыкновенно длинной хвостовой иглой, в 5—6 раз превосходящей по длине их тело. Каспийские ветвистоусые близки к некоторым пресноводным северным видам (например, Bythotrephes); предполагается, что при таянии великого ледника эти виды были внесены в Каспийское море и, приспособившись к новым условиям, дали начало каспийским эндемикам.
Значение ветвистоусых ракообразных в жизни стоячих пресноводных водоемов очень велико. Они служат излюбленным и весьма питательным кормом для многих рыб, как взрослых, так и молоди, и истребляются рыбами в огромном количестве. В кишечнике рипуса из уральского озера Увильда оказалось 47 тысяч босмин и 8,5 тысячи дафний. Чудской сиг, снеток, ряпушка, молодь судака, амурского сазана и многих других рыб питаются главным образом ветвистоусыми ракообразными. Питательность этого корма очень высока: содержание белка в теле дафнии достигает 50%, а жира 11% по весу. Многие морские рыбы также в большом количестве поедают ветвистоусых.
Высокие пищевые качества ветвистоусых ракообразных в полной мере оценены рыбоводами. На рыбоводных заводах нашей страны осуществляется массовое разведение дафний и моин, которыми кормят молодь осетровых и лососевых рыб.
Перед рыбоводами, занимающимися этим трудным делом, стоит задача обеспечить беспрерывное партеногенетическое размножение разводимых рачков. Для этого необходимо создать в бассейнах, в которых разводятся рачки, постоянные благоприятные для них условия. В случае хотя бы временного ухудшения условий существования рачков их яйца, как было описано раньше, оплодотворяются и превращаются в покоящиеся. Подойдя к бассейну, рыбовод обнаруживает вместо массы живых копошащихся рачков плавающие на поверхности воды эфиппиумы, которых рыбки есть, конечно, не будут. Дело может кончиться гибелью от голода сотен и тысяч маленьких осетрят и севрюжат.
Дафний и моин разводят в цементированных или просто выкопанных в земле бассейнах, воду которых удобряют навозом, отваром с жиротопки и азотобактерином. Эти вещества благоприятствуют массовому развитию бактерий, используемых в пищу рачками. Иногда бассейны делают проточными, что позволяет регулировать их температурный и газовый режим. Регулярно следя за состоянием рачков, удается получать достаточное их количество для откорма молоди разводимых на заводах рыб.