(Diplostraca Gerstaecker, 1866) Отряд Диплостраки, Водяные блохи, Order Diplostraca Gerstaecker, 1866 (Clam shrimps, Water fleas) 19 семейств
Синонимы:
Cladocera Latreille, 1829
Eucladocera
Gymnomera G.O. Sars, 1865
Onychocaudata Olesen & Richter, 2013
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ПОДОТРЯД КОНХОСТРАКИ (CONCHOSTRACA)
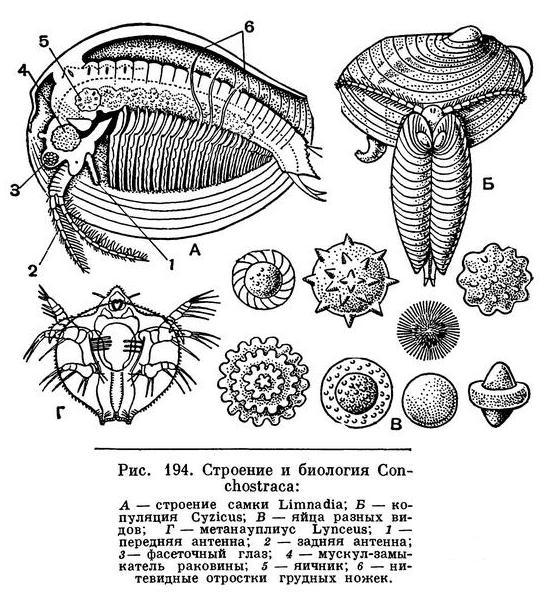
В мелких, иногда пересыхающих водоемах как средней, так и северной части нашей страны, а нередко и высоко в горах обитают рачки, менее заметные и не достигающие столь крупных размеров, как щитни. Они обычно окрашены в буроватые, розоватые или зеленоватые тона. Все тело их покрыто двустворчатой хитиновой раковинкой, которая у большинства видов имеет концентрические полосы прироста, несколько напоминающие кольца на раковинах двустворчатых моллюсков, например беззубки и перловицы. Эти полосы у рачков возникают после каждой линьки, поскольку при линьке наружная часть старой раковины сохраняется, а затем раковина начинает расти по периферии. Число полос соответствует числу линек.
Обе створки скреплены одним поперечным сильным мускулом-замыкателем. Рачки реагируют на опасность, сокращая этот мускул, причем створки раковины плотно сжимаются и рачок, как мертвый, остается неподвижно лежать на дне.
Раскроем створки раковины, чтобы рассмотреть строение тела рачка. Голова многих видов уплощена сверху вниз. Сидячие фасеточные глаза сближены и расположены на лобной ее части, а у одного рода (Cyclestheria) даже соединяются между собой. Перед ними находится простой науплиальный глазок. Передние антенны развиты слабо и не расчленены. Зато задние антенны длинные, двуветвистые, причем обе ветви снабжены многочисленными щетинками. Рачки плавают преимущественно за счет взмахов задних антенн, действующих как весла. Лишь у очень обычного рода Lynceus, способного широко раскрывать створки своей шаровидной раковины, не только задние антенны, но и грудные ножки совершают веслообразные движения.
Число сегментов, составляющих грудной отдел, колеблется от 10 до 32, и соответственно этому изменяется число пар грудных ножек. Они имеют приблизительно такое же строение, как у щитней, и их размеры также постепенно уменьшаются по направлению спереди назад. У самцов часть внутренних лопастей первой, а иногда и второй пары грудных ног преобразована в крючковидные придатки, которыми самец удерживает самку при спаривании (рис. 194, Б). У самок наружные лопасти нескольких средних ног превращены в длинные, загнутые вверх нитевидные отростки, приспособленные для удерживания яиц на спинной стороне тела под раковиной. Однако основные функции грудных ног — это подача пищи вперед ко рту и дыхание. Жабрами служат ближайшие к основанию наружные лопасти всех грудных ног. Для обеспечения этих функций грудные ножки все время находятся в движении, благодаря чему вода между створками раковины беспрерывно обновляется. Тело заканчивается крупным хитинизированным тельсоном, который или сжат с боков и в таком случае снабжен парой крупных когтей, либо сплющен в спинно-брюшном направлении и несет короткие коготки.
Большинство Conchostraca — донные животные. Наиболее крупные из них — Limnadia, достигающие в длину 17 мм, лежат боком на дне, Cyzicus роется в грунте, взмучивая ил движением брюшка и отчасти ножек. Представители этих и других родов плавают лишь в период спаривания. Только сравнительно мелкий (4—5 мм) Lynceus brachyurus, отличающийся шаровидной раковиной без полос прироста, обычно плавает спиной кверху, раскрыв створки раковины, и лишь изредка опускается на дно. Он питается главным образом планктонными организмами, в то время как Cyzicus ест грунт с находящимися в нем организмами, a Limnadia объедает микроскопические водоросли, находящиеся на дне и на поверхности водных растений. Крупные несъедобные частицы Limnadia склеивает специальным секретом, выделяемым железами ножек, и выбрасывает через заднюю щель своей раковины.
Размножение Conchostraca происходит по-разному. Все представители семейства Limnadiidae размножаются только партеногенетически, самцы среди них неизвестны. У других семейств самцы встречаются регулярно, причем, чем южнее обитают рачки, тем больше относительная численность самцов. При спаривании самец удерживает самку крючковидными отростками своих задних антенн и просовывает задний конец тела в заднюю щель между створками раковины самки. Яйца откладываются в пространство между спинной стороной тела самки и ее раковиной. У Cyclestheria часть задних грудных сегментов расширена таким образом, что перед ними образуется настоящая выводковая сумка. Проносив некоторое время яйца на спине под раковиной, самка линяет, и яйца выпадают на дно водоема.
Эти яйца окружены плотной и прочной оболочкой и способны переносить высыхание, замерзание и другие неблагоприятные условия. Нередко они имеют причудливую форму, у многих видов они снабжены выростами, способствующими их переносу ветром и различными животными (рис. 194, В). Яйца некоторых видов сохраняют жизнеспособность в течение 7 лет.
После того как яйцо попадет в благоприятные для его развития условия, из него выходит личинка, находящаяся на стадии науплиуса. Науплиус Conchostraca отличается очень маленькими передними антеннами. На следующей стадии — метанауплиуса — у личинки развивается зачаток раковины. У большинства Conchostraca эта зачаточная раковина уже двустворчатая и очень напоминает раковину ветвистоусых ракообразных (см. ниже). У Lynceus она имеет вид плоского щита, похожего на карапакс щитней (рис. 194, Г), и только на более поздних стадиях развития она перегибается пополам и становится двустворчатой. Все развитие происходит очень быстро. Так, например, Cyzicus достигает половозрелости через 19 дней после выхода личинки из яйца.
Современные Conchostraca не имеют никакого практического значения. Однако вымершие виды этих ракообразных привлекают к себе внимание геологов. В пресноводных отложениях, вообще очень бедных ископаемыми, хорошо сохраняющиеся раковинки Conchostraca встречаются довольно часто. По ним удается определять геологический возраст соответствующих слоев.
В предыдущие геологические периоды Conchostraca были, по-видимому, более многочисленны и шире распространены, чем ныне. Вымерших видов известно около 300, а современных вдвое меньше. Наиболее древние Conchostraca найдены в девонских отложениях. Замечательно, что среди древних вымерших видов, особенно триасовых, многие жили в море, где сейчас Conchostraca полностью отсутствуют.
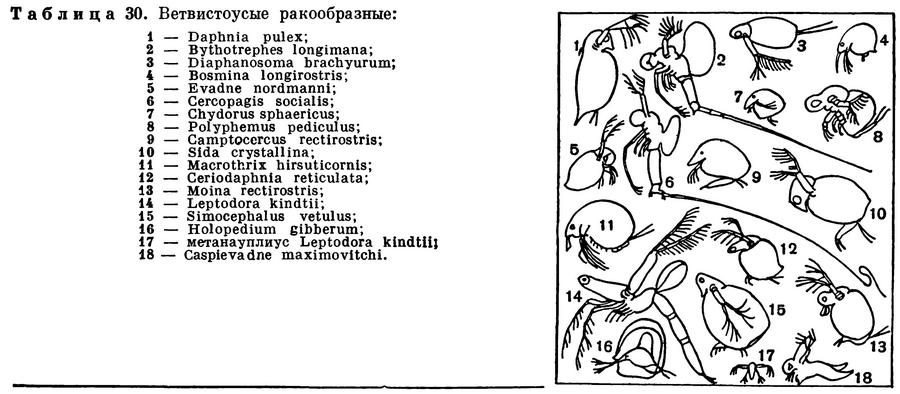
ПОДОТРЯД ВЕТВИСТОУСЫЕ РАКООБРАЗНЫЕ (CLADOCERA)
Почти в каждом пресноводном водоеме, начиная с огромных глубоких озер и кончая мелкими прудками, любой лов сачком или специальной сеткой из мельничного шелкового газа приносит десятки и сотни мелких рачков, часто называемых водяными блохами (табл. 30). Они оживленно снуют в толще воды, делая характерные взмахи своими задними антеннами, а некоторые из них передвигаются по дну или по подводным растениям. Их хорошо знают не только ученые-зоологи, но и все любители аквариумов, которые кормят ими, живыми или сушеными, своих рыбок. Их знают и рыбоводы, занимающиеся их разведением для кормления молоди рыб, выращиваемой на рыбоводных заводах. Многие из этих рачков всю жизнь проводят в толще воды и составляют значительную часть пресноводного планктона. Иногда они развиваются в таком количестве, что вода приобретает буроватый оттенок. Наряду с планктонными есть и донные виды. Есть и такие, которые то прикрепляются к растениям или опускаются на дно, то всплывают в толщу воды. Подавляющее большинство видов ветвистоусых обитает в пресных водах, но есть и несколько морских видов. Размеры этих рачков незначительны, обычно меньше 1 мм, редко до 5 мм и в исключительных случаях до 10 мм. Поэтому для изучения их строения приходится пользоваться микроскопом.
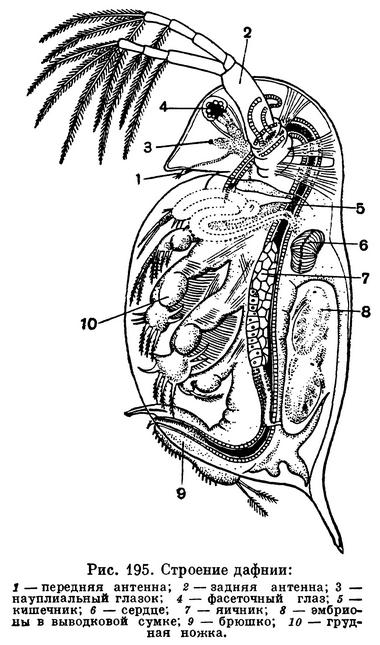
Тело большинства ветвистоусых ракообразных, например дафнии (рис. 195), заключено в хитиновую полупрозрачную двустворчатую раковинку, а голова вытянута в направленный вниз клюв, или рострум. На лобной части головы расположен непарный фасеточный глаз, возникший в результате слияния двух боковых глаз, а перед ним — простой науплиальный глазок, отсутствующий у некоторых видов.
Число фасеток, образующих сложный глаз, различно у разных видов и зависит от способа их питания. В глазу отфильтровывающей мелкие частицы дафнии насчитывается всего 22 фасетки, а у хищных Evadne и Leptodora соответственно 80 и 300. При нападении на мелких животных глаза хищных ветвистоусых имеют важное значение. Под рострумом прикрепляются палочковидные обыкновенно относительно очень маленькие передние антенны. Зато задние антенны непропорционально велики по сравнению с телом. Они двуветвисты, причем каждая ветвь снабжена длинными перистыми щетинками. Задние антенны служат основным органом передвижения ветвистоусых. Одновременно взмахивая обеими задними антеннами, рачки отталкиваются ими от воды и таким образом плывут короткими скачками. На нижней поверхности головы находится рот, окруженный спереди верхней губой, с боков жвалами и сзади челюстями.
Грудной отдел ветвистоусых укорочен и, судя по количеству пар ножек, состоит из 4—6 сегментов. Он, как уже указывалось, помещается внутри двустворчатой раковины, но у самок между спинной поверхностью тела и спинным краем раковины имеется обширная полость, выполняющая роль выводковой сумки. В эту сумку откладываются яйца, и при благоприятных условиях там же они развиваются. У хищных ветвистоусых раковина уменьшена в размерах, оставляет непокрытым почти все туловище и прикрепляется к его спинной стороне (табл. 30, 2, 5, 6, 8, 14, 18); в таких случаях она служит только для вынашивания яиц.
Строение грудных ножек различно у разных видов и зависит от характера их питания. Эти ножки в общем напоминают листовидные ножки жаброногов, щитней и Conchostraca. У большинства ветвистоусых ракообразных грудные ножки служат для отфильтровывания мелких взвешенных в воде частиц, которыми рачки питаются. В соответствии с этим лопасти грудных ножек снабжены многочисленными перистыми щетинками, образующими вместе фильтрационный аппарат. Ножки совершают до 300—500 взмахов в 1 минуту.
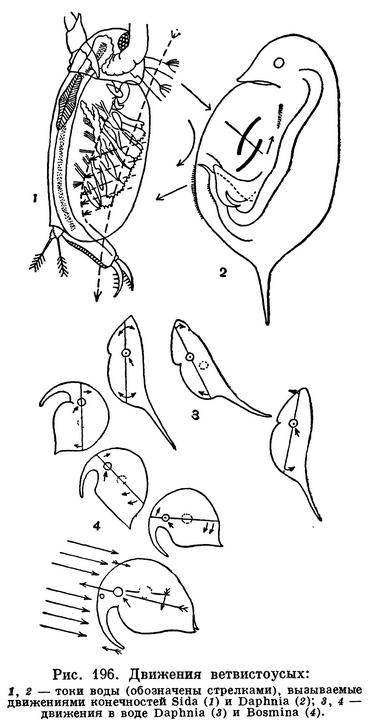
В простейшем случае, например у Sida (рис. 196, 2), вода входит между створками в передней части раковины. При одновременном движении всех 6 пар ножек вперед между ними возникает как бы ряд всасывательных камер, куда просачивается вода. При этом щетинки отфильтровывают всю мелкую взвесь. Движение ножек назад сжимает камеры и выгоняет воду через заднюю часть раковины. Отфильтрованные частицы направляются при помощи специальных щетинок в продольный желоб, расположенный на нижней стороне туловища между основаниями грудных ножек. По этому желобу находящимися на его стенках щетинками пищевой комок проталкивается вперед, по направлению ко рту.
У Daphnia этот аппарат усложнен и усовершенствован (рис. 196, 2). Из пяти пар ее грудных ножек фильтруют только третья и четвертая. При движении первой пары ножек вперед вода поступает в фильтрующую камеру. В это время третья и четвертая пары изгибаются вперед, обеспечивая дальнейший ток воды. Отфильтрованные пищевые частицы поступают в брюшной желобок. Пятая пара ног ограничивает сзади фильтрующую камеру. Делая взмах назад, она вместе с четвертой парой вызывает выводящий ток воды. Таким образом, между грудными ножками дафнии функции разделены, что говорит о большей их специализации.
Ближайшие к основанию наружные лопасти грудных ног фильтрующих ветвистоусых ракообразных превращены в жабры. По системе полостей кровь поступает в жабры и обогащается в них кислородом. В плазме крови ветвистоусых растворен гемоглобин, от которого в значительной степени зависит окраска рачков. Его концентрация меняется. Если содержание кислорода в воде низкое, кровь приобретает темно-красный цвет, если кислорода в воде много, кровь окрашена значительно светлее. При изменении содержания кислорода в воде меняется окраска рачков, но для этого требуется около 10 дней.
Дафнии с темно-красной кровью в воде с низким содержанием кислорода живут значительно дольше, чем дафнии, имеющие светлую, бедную гемоглобином кровь. Очевидно, гемоглобин, как и у позвоночных животных, связывает кислород.
Кровь приводится в движение мешковидным сердцем, взбалтывающим ее в полости тела (рис. 195, 6). Настоящих кровеносных сосудов нет. Сердце дафнии бьется очень часто. При температуре 10° С оно делает 150, а при температуре 20° С даже 500 ударов в минуту.
Совсем иначе устроены и действуют грудные ножки хищных ветвистоусых ракообразных (табл. 30). Их боковые лопасти недоразвиты или отсутствуют, жабр нет совсем, а хорошо развитые внутренние ветви снабжены сильными шипами и шиловидными щетинками. При помощи грудных ног рачки схватывают других мелких животных, служащих им пищей. Хищные ветвистоусые воспринимают кислород скоплением железистых клеток, находящихся на голове.
Задний брюшной отдел тела ветвистоусых укорочен, часто изогнут под прямым углом и может прятаться между створками раковины. Он обычно заканчивается двумя когтями. Однако у многих хищных видов задний отдел имеет форму трубки, сохраняет сегментацию и иногда продолжается в длинный шип, назначение которого остается непонятным.
Наблюдая за поведением ветвистоусых рачков в водоеме или в аквариуме, нетрудно убедиться в том, насколько различны повадки разных видов. Особенно многочисленны рачки, проводящие всю свою жизнь в толще воды. Планктонные дафнии и некоторые другие ветвистоусые после взмаха задними антеннами и вызванного этим движением скачка некоторое время неподвижно парят в воде. Благодаря тому что центр тяжести их тела находится ниже места прикрепления антенн, в момент удара антенн тело рачка наклоняется, а в период покоя снова выпрямляется. Распростертые антенны препятствуют погружению рачка (рис. 196, 3).
Планктонные ветвистоусые из рода Bosmina плавают иначе. Они беспрерывно машут своими задними антеннами, причем передняя часть их тела при каждом ударе несколько приподнимается. Однако длинные хоботообразные передние антенны служат балансиром, возвращающим тело в горизонтальное положение (рис. 196, 4). Если босмине отрезать передние антенны, она начинает беспомощно кувыркаться и теряет способность к плаванию.
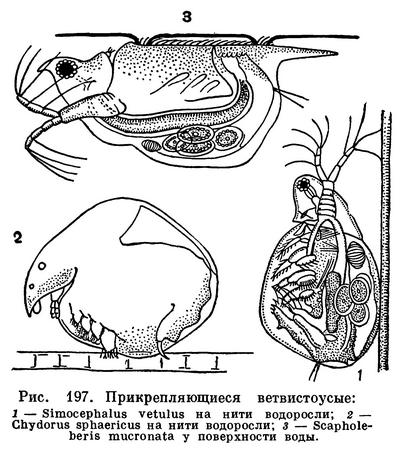
Среди зарослей подводных растений в прибрежной части озер и прудов часто попадается крупный (до 4 мм) ветвистоусый рачок Sida crystallina, который, как бы устав беспрерывно плавать, время от времени прикрепляется к растениям при помощи специальной присоски, находящейся на задней части его головы. Другой обитатель зарослей — Simocephalus — цепляется за растения щетинками задних антенн. Маленький шарообразный хидорус (Chydorus sphaericus), поплавав некоторое время, садится на водоросли и начинает ползать по ним, удерживаясь щетинками края раковины и отталкиваясь второй парой грудных ног и брюшком. Наиболее своеобразен способ передвижения Scapholeberis mucronata, который благодаря присутствию несмачиваемых щетинок на нижнем крае раковины может прикрепляться снизу к поверхностной пленке натяжения воды и быстро скользить по ней спинной стороной вниз (рис. 197).
Наряду с такими постоянно или временно плавающими формами существует большое количество видов, плавающих очень плохо и постоянно ползающих по дну. Некоторые из них способны даже зарываться в поверхностный слой ила.
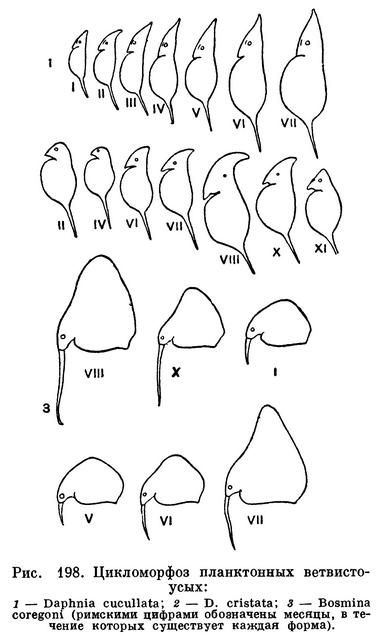
С жизнью многих ветвистоусых ракообразных в толще воды связано одно любопытное явление, получившее название цикломорфоза. Сущность его заключается в том, что форма и величина раковины определенных видов закономерно изменяются по сезонам года. В летнее время у некоторых видов дафнии на голове появляется так называемый «шлем», а у босмин на спинной стороне — «горб». Кроме того, у дафний удлиняется задний шип на раковине, а у босмин — передние антенны (рис. 198).
Эти изменения происходят параллельно изменению температуры воды. В данном случае важна, однако, не сама температура, а ее воздействие на плотность воды. Прогретая вода обладает меньшей плотностью, чем охлажденная. Для движения и парения в менее вязкой прогретой воде необходимы более длинные выросты, а иногда и иная форма тела. Поэтому летние формы дафний и босмин развивают специальные приспособления для передвижения и парения в таких условиях. Значение плотности воды для изменений формы тела рачков подтверждается наблюдениями Е. Ф. Мануйловой над дафниями озера Балхаш.
В этом озере соленость по направлению с востока на запад постепенно увеличивается и вместе с тем вода становится все более плотной. Местная планктонная дафния — Daphnia balchashensis, живущая при сравнительно высокой солености, имеет совершенно круглую голову, а живущая в опресненной части озера — голову, снабженную загнутым назад шлемом.
Большинство ветвистоусых ракообразных, как уже указывалось, питается, отфильтровывая мелкую, находящуюся в воде взвесь. Основной их пищей служат бактерии, одноклеточные водоросли и мелкие органические остатки — детрит. Наибольшее значение имеют бактерии. В опытах многих исследователей удавалось воспитывать ветвистоусых при кормлении их одними бактериями, а американский зоолог Банта получил в таких условиях около 1600 поколений дафний. В природных водоемах ветвистоусые прекрасно живут и размножаются при концентрации бактерий не менее 1 миллиона в 1 см3 воды. Если численность бактерий меньше 500 тысяч на 1 см3, наступление половозрелости дафний задерживается; если же численность бактерий падает ниже 200 тысяч на 1 см3, жизнь рачков становится невозможной. С другой стороны, избыточное количество бактерий (более 3—5 миллионов на 1 см3) также вредно влияет на рачков и вызывает прежде всего замедление или приостановку их размножения, а иногда даже их гибель. Это объясняется выделением бактериями продуктов обмена веществ, ядовитых для ветвистоусых.
От количества бактерий в водоеме зависит численность ветвистоусых. Неоднократно наблюдалось уменьшение числа ветвистоусых в период массового развития сине-зеленых водорослей. Некоторые исследователи предполагали вредное воздействие сине-зеленых на рачков. Однако, по данным Е. Ф. Мануйловой, бурное развитие сине-зеленых само по себе никакого влияния на рачков не оказывает. Оно подавляет развитие бактерий и именно таким образом косвенно воздействует на ветвистоусых. Часто наблюдающиеся периоды резкого увеличения численности ветвистоусых обычно следуют за периодами размножения бактерий.
Помимо бактерий, фильтрующие ветвистоусые используют в пищу одноклеточные водоросли. Они могут заглатывать только самые мелкие виды протококковых и зеленых водорослей, которые обычно развиваются в большом количестве в прудах, но не в крупных озерах. Обычные, особенно колониальные, планктонные водоросли слишком велики для того, чтобы быть проглоченными рачками. Во многих случаях водоросли приобретают большое значение для ветвистоусых не потому, что поедаются ими, а потому, что после отмирания вызывают массовое развитие бактерий — основного корма рачков.
При фильтрации пищи рачки не могут отсортировывать съедобные частицы от несъедобных. Они способны только отбирать частицы по их размеру, отбрасывая слишком крупные. С этим связаны, по наблюдениям В. М. Рылова, те изменения в составе планктона, которые наблюдаются при проникновении планктона стоячих водоемов в текучие. Эти изменения сводятся главным образом к резкому уменьшению численности ветвистоусых ракообразных, в то время как численность других планктонных животных, например коловраток, существенно не меняется. Оказывается, что речной поток несет мелкие минеральные частицы и попавшие в него планктонные ветвистоусые отфильтровывают их и заполняют ими свой кишечник. Рачки идут ко дну и погибают, а коловратки, захватывающие пищу иным способом, и притом еще более мелкую, остаются невредимыми.
Фильтрация пищи рачками происходит беспрерывно. Время, нужное для заполнения кишечника, колеблется от 10 до 240 минут и зависит от величины рачков, размеров пищевых частиц, их концентрации и температуры. Мелкая босмина в культуре водоросли Ankistodesmus наполняет кишечник в течение 10 минут, а в культуре бактерии Bacillus coli тратит на это 2—2,5 часа. В течение суток одна дафния поглощает от 4,8 до 40,8 миллиона бактерий (в зависимости от их концентрации). Даже после наполнения кишечника рачки продолжают фильтровать, и отфильтрованные частицы двигаются вперед по их брюшному желобку. Но в рот они не попадают: ротовые придатки и задний отдел тела выталкивают их снова в воду. Рачки начинают заглатывать пищу только после того, как кишечник хотя бы частично освободится от ее остатков.
Совсем иначе добывают себе пищу хищные виды. Самый крупный совершенно прозрачный ветвистоусый рачок Leptodora kindtii (табл. 30, 14) неподвижно парит в воде и своим громадным глазом высматривает добычу. Достаточно нескольких сильных взмахов его задних антенн для того, чтобы он настиг какого-нибудь более мелкого планктонного рачка или коловратку, схватил жертву своими далеко вынесенными вперед грудными ногами и вонзил в нее острые зазубренные жвалы. Приблизительно так же охотится и другой крупный хищник — Bythotrephes (табл. 30, 2). Оба рачка сначала размельчают жертву жвалами, а затем всасывают жидкую пищу. Они нападают почти на всех планктонных животных, включая собственную молодь, но не трогают сидячих и даже временно прикрепляющихся рачков, например сиду и симоцефалюса.
Лептодора и битотрефес весьма прожорливы. При температуре 20—25° С каждая лептодора съедает за сутки более 50, а битотрефес около 25 рачков. Соответствующие расчеты показывают, что в некоторых водоемах, например в Рыбинском водохранилище, хищные ветвистоусые выедают заметную часть зоопланктона (более 40% всех планктонных животных). Наносимый ими таким образом вред не так велик, поскольку они в свою очередь интенсивно потребляются многими рыбами.
В некоторых водоемах планктонные ветвистоусые ракообразные подобно другим планктонным животным совершают регулярные суточные вертикальные миграции. В темное время суток они поднимаются в поверхностные слои воды, а в светлое время уходят вниз. Однако во многих озерах и водохранилищах большая часть рачков круглые сутки остается близ поверхности.
До тех пор пока условия жизни ветвистоусых рачков благоприятны, в водоемах встречаются только самки, откладывающие и вынашивающие в своих выводковых камерах неоплодотворенные яйца. Число этих яиц различно не только у разных видов, но даже у различных особей одного вида. Разные виды дафний образуют до 50 или даже до 100 яиц на самку, босмины — до 16 яиц, а мелкие донные виды — не больше 2 яиц. Яйца развиваются в выводковой камере матери, где из них образуются сначала подвижные эмбрионы, а затем вполне сформировавшиеся маленькие рачки, покидающие выводковую камеру и приступающие к самостоятельному существованию. Летом, при достаточном количестве пищи, рождение молоди и появление в камере новых яиц происходят каждые 1—2 дня, благодаря чему численность ветвистоусых может очень быстро увеличиваться. Обыкновенно рождение молоди сопровождается линькой самки. У хищных ветвистоусых, например у лептодоры и битотрефеса, выводковая камера изолирована от окружающей среды и при выходе молоди каждый раз разрушается, а затем восстанавливается вновь.
Вышедшая на свободу молодь часто линяет и быстро растет. Первые три линьки происходят через 1—1,5 суток, а следующие через 2—3 суток. Большинство видов достигает половозрелости после третьей-четвертой линьки, т. е. через 2—6 дней после рождения. Так получаются новые поколения самок, продолжающие размножаться партеногенетически.
Но вот условия существования ветвистоусых рачков ухудшились вследствие понижения температуры, уменьшения количества пищи или загрязнения водоема. Это ухудшение сейчас же сказывается на судьбе яиц, находящихся в половых путях самки. Из неоплодотворенных яиц после их откладки в выводковую камеру выходят не самки, а самцы. Кроме того, часть яиц в половых путях самки подвергается второму делению созревания, в результате которого количество хромосом в яйце уменьшается вдвое. Такие яйца могут развиваться только после оплодотворения. При слиянии яйца и сперматозоида восстанавливается полное количество хромосом.
Замечательно, что даже кратковременные отклонения от нормальных условий жизни могут прервать процесс партеногенетического размножения. Судьба яйца определяется за 15 минут до его выхода из половой системы самки. Если в этот момент рачки подвергнутся каким-нибудь неблагоприятным воздействиям, из их яиц выведутся самцы или эти яйца будут непременно нуждаться в оплодотворении.
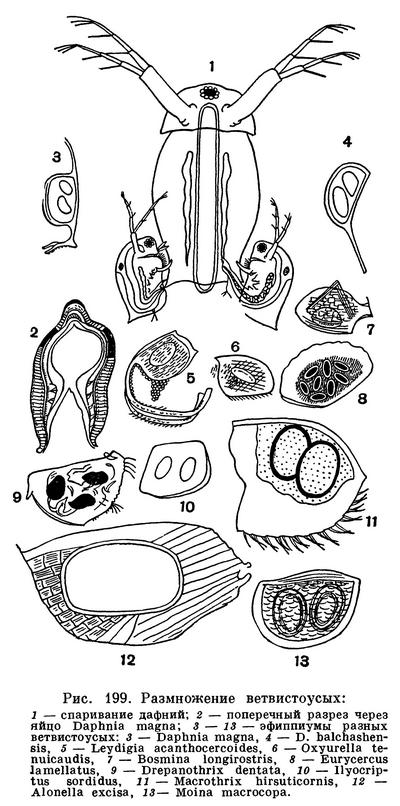
Самцы ветвистоусых ракообразных — карлики по сравнению с самками. Часто они в несколько раз меньше самок (рис. 199). Их передние антенны удлинены, а первая пара грудных ножек снабжена коготками, помогающими удерживать самку.
Обыкновенно самцы появляются в водоемах осенью, после значительного понижения температуры. Они прикрепляются к самкам при помощи коготков передних ног и при помощи передних антенн, причем у дафний нередко к одной самке прикрепляются два самца. Сперма проникает в выводковую камеру близ отверстий яйцеводов, а потом и в них.
Оплодотворенные яйца во многих отношениях отличаются от неоплодотворенных. Их гораздо меньше (редко больше двух), но зато они значительно богаче желтком и поэтому крупнее, чем партеногенетические. Вокруг них развиваются защитные оболочки. У Sida, Diaphanosoma, Holopedium и хищных видов оплодотворенные яйца снабжены двумя оболочками — плотной внутренней и клейкой наружной. Такие яйца откладываются в воду и опускаются на дно.
У большинства ветвистоусых оплодотворенные яйца откладываются в выводковую камеру, а в образовании их защитных оболочек принимает участие материнская раковина. Ее спинной край становится толще, под раковиной собираются клетки, плотным слоем окружающие яйцо или яйца. У дафний поверх этих клеток выделяется хитиновая оболочка и в выводковой камере возникает так называемое седлышко, или эфиппиум, содержащее обычно два яйца. Формирование эфиппиума продолжается 2—3 дня, и самка носит его в течение 1—2 дней. Затем она линяет, и эфиппиум оказывается на свободе. Благодаря присутствию воздухоносного слоя эфиппиумы дафний плавают на поверхности воды. У некоторых других ветвистоусых наружной оболочкой эфиппиума служит вся раковина или значительная ее часть, сброшенная при линьке (рис. 199).
Главная биологическая особенность оплодотворенных яиц заключается в том, что они могут развиваться только после более или менее продолжительного периода покоя и поэтому называются покоящимися. Такие яйца помогают рачкам переносить всевозможные неблагоприятные условия; они вмерзают в лед, высыхают, сохраняя в течение длительного времени жизнеспособность. В то же время они служат средством расселения ветвистоусых, легко переносясь из водоема в водоем ветром, вместе с илом, присохшим к лапкам водоплавающих птиц, и т. д. Благодаря своим покоящимся яйцам ветвистоусые широко распространены во всех стоячих водоемах нашей планеты.
Многие виды зиму проводят в стадии покоящегося яйца, которое начинает свое развитие весной и дает начало самке, приступающей к партеногенетическому размножению. У всех ветвистоусых, за исключением лептодоры, из яйца выходит вполне сформировавшийся рачок. Лептодора — единственный представитель ветвистоусых, имеющий личинку; она вылупляется из покоящегося яйца на стадии метанауплиуса (табл. 30, 17).
Число известных видов ветвистоусых достигает 380, в СССР найдено 164 вида. Некоторые из них распространены очень широко и встречаются в самых разнообразных водоемах, другие приурочены к определенным районам или населяют водоемы какого-нибудь особого типа. Во всех странах света обитают обыкновенная дафния (Daphnia pulex), планктонная дафния (Daphnia longispina), босмина (Bosmina longirostris), хидорус (Chydorus sphaericus) и многие другие. В то же время существуют виды, живущие только на Крайнем Севере (Daphnia arctica, Eurycercus glacialis). Преимущественно в северной части СССР и Америки встречается своеобразный Holopedium gibberum, имеющий поверх раковины студенистую оболочку, придающую рачку вид прозрачного шарика (табл. 30, 16). Этот рачок изредка встречается и в средней части СССР, в водоемах, связанных с моховыми болотами. Наконец, есть и тропические виды, в нашей стране обитающие на залитых водой рисовых полях Узбекистана, в Закавказье, в водоемах дельты Волги. Высказывается предположение о том, что их покоящиеся яйца переносятся на колоссальные расстояния воздушными потоками.
Некоторые ветвистоусые рачки населяют только те водоемы, вода которых имеет кислую реакцию и бедна минеральными солями, но большинство видов способно существовать в самых разнообразных, в том числе и неблагоприятных условиях, например в условиях недостаточного содержания кислорода в воде.
Немногочисленные морские представители ветвистоусых ракообразных также распространены очень широко, но приурочены преимущественно к прибрежным водам. Только в Каспийском море обитает несколько чрезвычайно своеобразных видов (табл. 30, 6, 18), частично присущих только Каспию, частично встречающихся также в Азовском море и в опресненных районах Черного моря. Среди них особенно причудливы виды рода Cercopagis, снабженные необыкновенно длинной хвостовой иглой, в 5—6 раз превосходящей по длине их тело. Каспийские ветвистоусые близки к некоторым пресноводным северным видам (например, Bythotrephes); предполагается, что при таянии великого ледника эти виды были внесены в Каспийское море и, приспособившись к новым условиям, дали начало каспийским эндемикам.
Значение ветвистоусых ракообразных в жизни стоячих пресноводных водоемов очень велико. Они служат излюбленным и весьма питательным кормом для многих рыб, как взрослых, так и молоди, и истребляются рыбами в огромном количестве. В кишечнике рипуса из уральского озера Увильда оказалось 47 тысяч босмин и 8,5 тысячи дафний. Чудской сиг, снеток, ряпушка, молодь судака, амурского сазана и многих других рыб питаются главным образом ветвистоусыми ракообразными. Питательность этого корма очень высока: содержание белка в теле дафнии достигает 50%, а жира 11% по весу. Многие морские рыбы также в большом количестве поедают ветвистоусых.
Высокие пищевые качества ветвистоусых ракообразных в полной мере оценены рыбоводами. На рыбоводных заводах нашей страны осуществляется массовое разведение дафний и моин, которыми кормят молодь осетровых и лососевых рыб.
Перед рыбоводами, занимающимися этим трудным делом, стоит задача обеспечить беспрерывное партеногенетическое размножение разводимых рачков. Для этого необходимо создать в бассейнах, в которых разводятся рачки, постоянные благоприятные для них условия. В случае хотя бы временного ухудшения условий существования рачков их яйца, как было описано раньше, оплодотворяются и превращаются в покоящиеся. Подойдя к бассейну, рыбовод обнаруживает вместо массы живых копошащихся рачков плавающие на поверхности воды эфиппиумы, которых рыбки есть, конечно, не будут. Дело может кончиться гибелью от голода сотен и тысяч маленьких осетрят и севрюжат.
Дафний и моин разводят в цементированных или просто выкопанных в земле бассейнах, воду которых удобряют навозом, отваром с жиротопки и азотобактерином. Эти вещества благоприятствуют массовому развитию бактерий, используемых в пищу рачками. Иногда бассейны делают проточными, что позволяет регулировать их температурный и газовый режим. Регулярно следя за состоянием рачков, удается получать достаточное их количество для откорма молоди разводимых на заводах рыб.