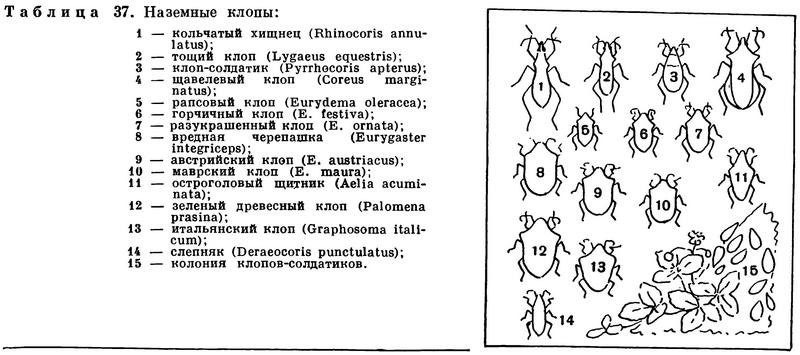
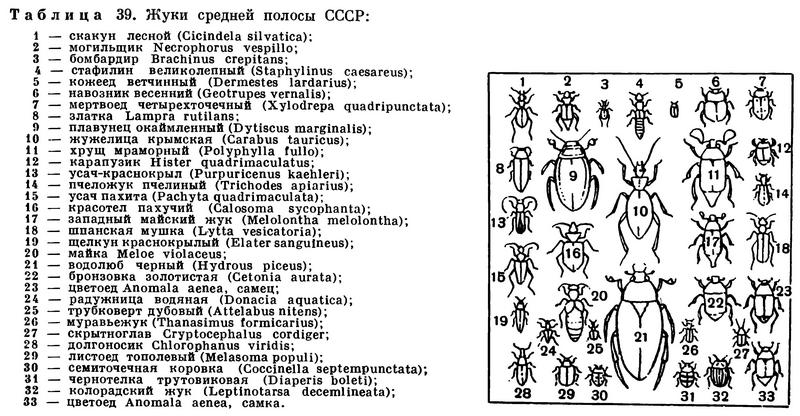
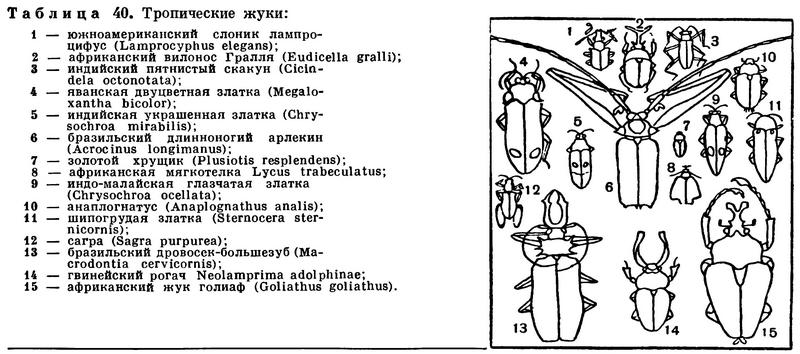
(Insecta Linnaeus, 1758) Класс Инсекты, Насекомые, Class Insecta Linnaeus, 1758 (Insects) 30 отрядов
Синонимы:
Ideloblattidae Zalessky, 1929
Жизнь животных. Том 3. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1969 г.
КЛАСС НАСЕКОМЫЕ (INSECTA, или HEXAPODA)
В класс насекомых включают всех трахейнодышащих членистоногих, имеющих 3 пары ног. Их принято разделять на первичнобескрылых и крылатых. Однако детальные исследования особенностей строения, развития и биологии первичнобескрылых, т. е. таких шестиногих, предки которых явно тоже не имели крыльев, показали, что эта группа очень неоднородная.
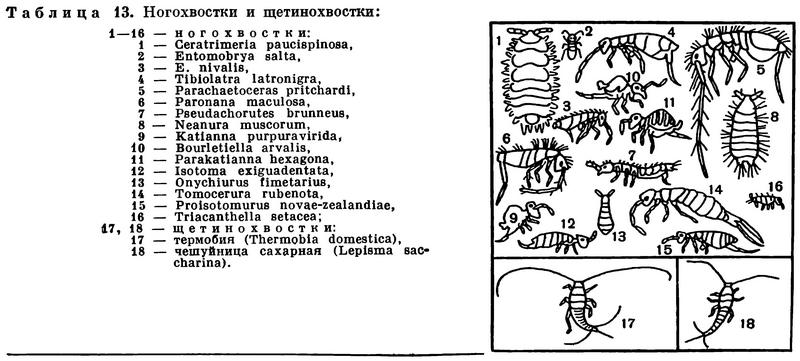
Один отряд таких шестиногих — так называемые щетинохвостки (Thysanura) — очень близок ко всем «настоящим» крылатым насекомым (подкласс открыточелюстные — Insecta-Ectognatha). Остальные же отряды — бессяжковых (Protura), ногохвосток (Collembola), вилохвосток (Diplura) — по многим важным признакам совершенно специфичны и заслуживают выделения в подкласс скрыточелюстных (Insecta-Entognatha), как это заключили специалисты, обсуждавшие вопрос о низших насекомых на Международном энтомологическом конгрессе в Монреале в 1956 году.
Класс насекомых кратко можно охарактеризовать следующим образом. Это наземные членистоногие, у которых тело явно разделено на голову, грудь и брюшко, а основные, служащие для передвижения конечности находятся в числе 3 пар на грудном отделе. Дыхание осуществляется с помощью трахейной системы или кожное — всей поверхностью тела.
Те насекомые, которые дышат с помощью всей поверхности тела и имеют проницаемые для дыхательных газов и для испарения покровы, ограничены в своем распространении только влажными средами. Они встречаются в основном в почве и в гниющих остатках организмов.
Те высшие насекомые, у которых покровы непроницаемы и развита трахейная система, могут обитать и в сухих средах. Эти насекомые широко расселились по нашей планете.
Именно такие, ведущие открытый образ жизни и часто ярко окрашенные высшие насекомые и известны большинству читателей.
Класс насекомых разделяется на 2 подкласса — насекомые скрыточелюстные (Insecta-Entognatha) и насекомые открыточелюстные (Insecta-Ectognatha).
ПОДКЛАСС ОТКРЫТОЧЕЛЮСТНЫЕ, ИЛИ НАСТОЯЩИЕ, НАСЕКОМЫЕ (INSECTA-ECTOGNATHA)
Основные сведения о насекомых. Из общего числа видов животных, населяющих Землю, на долю насекомых приходится около 70%. Число уже описанных видов приближается к миллиону, но ежегодно специалисты открывают и описывают все новые и новые тысячи видов, и, по принятым в настоящее время подсчетам, на земном шаре обитает более 2 миллионов различных видов насекомых.
Основная масса насекомых — обитатели суши, и здесь насекомые распространены необычайно широко. Исключительно богат и разнообразен мир насекомых в тропиках. В тропических лесах богатство видами многих групп насекомых, в том числе наиболее бросающихся в глаза, например бабочек, так велико, что легче встретить новый вид, чем другой экземпляр уже встреченного!
В более умеренных широтах число видов насекомых не так велико, но зато общая их масса огромна. Академик В. И. Вернадский, создатель науки биогеохимии, образно показал, что, например, вес одной стаи саранчи, однажды перелетавшей через Красное море в начале нашего века, превышал вес всех цветных металлов, включая свинец и медь, выплавленных человечеством за всю историю до этого дня!
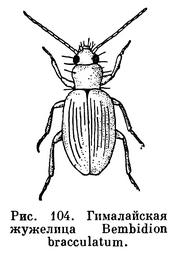
Насекомые заходят высоко в горы. Их можно встретить на огромных высотах. Например, мелкая жужелица Bembidion bracculatum в Гималаях населяет только пояс от 4300 до 5000 м, приспособившись к жизни в этих суровых условиях, а всего из зоны вечного снега в Гималаях описано 25 видов только этого рода жужелиц!
Насекомые населяют и абсолютно безводные пустыни, такие, как, например, пустыня Намиб в Южной Африке, где никогда не бывает ни капли дождя, где нет никаких растений и где живут жуки-чернотелки (Gonopus, Syntyphlus subterraneus), питающиеся только приносимыми за сотни километров знойным ветром остатками растений, отлагающимися у оснований барханов. Необходимую для жизни влагу чернотелки получают биохимическим путем, используя воду, которая образуется при окислении углеводов сухой, недоступной другим организмам пищи!
Много различных насекомых обитает в пещерах. Там, где нет, кажется, никакого поступления органических веществ, в обстановке вечной тьмы, встречаются слепые представители этой группы, чаще всего жуки, использующие те ничтожные запасы пищи, которые удается отыскать.
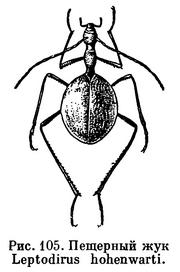
Первое пещерное насекомое было открыто в пещере Постойна на территории Югославии — жук Leptodirus hohenwarti — в 1831 году, а с тех пор из разных стран описана богатая и удивительная фауна насекомых-троглобионтов.
Очень далеко заходят насекомые и в высокие широты: за полярным кругом, на почти голых скалах, покрытых большую часть года снегами и льдом и лишь на очень короткий период освобождающихся от зимних оков, среди лишайников и мхов можно встретить мелких представителей этого класса.
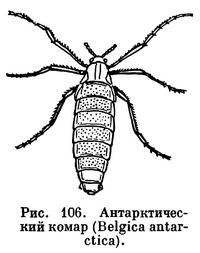
Даже на островах Антарктики недавно найдены бескрылые комары-дергуны, как антарктический комар (Belgica antarctiса) — родич нашего мотыля, красные личинки которого хорошо известны рыболовам и любителям аквариумных рыб.
Приспособившиеся к полету крылатые насекомые завоевали воздушную среду — лучшие летуны среди них активно поднимаются на высоту нескольких сотен метров (двукрылые, бабочки), но можно встретить насекомых и выше, на высоте нескольких километров. Сюда они заносятся восходящими токами нагревающегося у поверхности земли воздуха и затем ветром переносятся на огромные расстояния. В высоких слоях атмосферы чаще встречаются не хорошо летающие, а легко парящие в воздухе насекомые — мелкие мушки, в основном злаковые мушки (Agromyzidae), тли, трипсы. Но не только крылатых насекомых можно встретить на высоте нескольких десятков и сотен метров. Легкие плоские маленькие личинки щитовок (Coccidae, рис. 217, 5), сдуваемые порывами ветра с ветвей деревьев, тоже поднимаются с пылью высоко вверх и так расселяются. Высокой способностью к расселению насекомых объясняется то, что на всяком океаническом островке, удаленном за тысячи километров от остальной суши, можно найти насекомых. А на поверхности суши они заселяют все растения, поднимаясь к вершинам самых высоких деревьев. В тропических лесах Африки, например, есть такие насекомые, которые еще недавно считались исключительно редкими, а оказались обычными, но постоянно обитающими высоко в кронах гигантских деревьев; они недоступны для сборов без пожарных лестниц или вертолетов. Все участки крон деревьев, весь ковер травянистых растений, мхов и лишайников, вся почва на глубину того слоя, куда проникают корни растений, и даже глубже осваиваются насекомыми. В Туркмении, например, закаспийский термит (Anacanthotermes ahngerianus) прокладывает свои ходы на глубину до 12 метров!
Некоторые насекомые приспособились к жизни в тканях растений, к развитию в теле различных животных — от губок до млекопитающих и человека, к использованию всевозможных очень специфических местообитаний, самых разных источников пищи.
По способности жить на суше в самых различных условиях класс насекомых не имеет себе равных и именно благодаря тому, что отдельные виды приспособились к жизни в самых разных, иногда невероятных условиях (например, личинки некоторых мух в горячих источниках или в насыщенных солевых растворах), насекомые в процессе своей бурной эволюции, продолжающейся уже во всяком случае более 200 миллионов лет, дали то исключительное богатство видов, которое обеспечивает современное процветание этой группы членистоногих.
От жизни на суше многие насекомые перешли к жизни в воде. Уже в каменноугольном периоде палеозойской эры в воде развивалось большое количество видов насекомых, и в частности вымерший в конце этого периода отряд палеодиктиоптер (Palaeodictyoptera, рис. 185), некоторые представители которого были много крупнее, чем самые крупные из ныне живущих насекомых (до 60 см).
Завоевали пресные водоемы и стали водными в полном смысле слова, т. е. дышащими за счет того кислорода, который растворен в воде, только личинки насекомых, взрослая окрыленная стадия которых способна оставлять воду. Если же насекомое и во взрослом состоянии живет в воде, то оно дышит кислородом воздуха, поднимаясь к поверхности. Только некоторые, как например мелкие (0,2 мм) наездники-карафрактусы (Caraphractus) и прествичии (Prestwichia aquatica, рис. 371, 3,4), развивающиеся в яйцах жуков-плавунцов, ведут во взрослом состоянии совершенно подводный образ жизни.
Некоторые насекомые, как например клопы-водомерки (Hydrometridae и Gerridae), живут на поверхности воды, бегая по ней, отталкиваясь длинными, как у пауков, ногами от пленки поверхностного натяжения.
Близкие к нашим водомеркам представители семейства Halobatidae — морские водомерки — бегают по поверхности морей, a Halobates micans в тропическом поясе обитает в океанах за сотни километров от берега. Правда, нельзя морских водомерок назвать настоящими водными насекомыми — они живут на поверхности воды, питаются всплывающими трупами животных, а яйца откладывают на плавающие водоросли, перья птиц и даже на себе подобных. Настоящих морских насекомых, развивающихся в морской воде, очень мало — немногим более десятка, все из отряда двукрылых. Только один вид комаров-долгоножек (Limonia monostromia), обитающий у берегов Японских островов, в течение всего цикла развития не оставляет моря.
Насекомые как класс сформировались на суше и происходят от уже наземных многоножкообразных животных, этим и объясняется малое количество среди них морских форм. При этом все морские насекомые — представители наиболее высокоорганизованных отрядов.
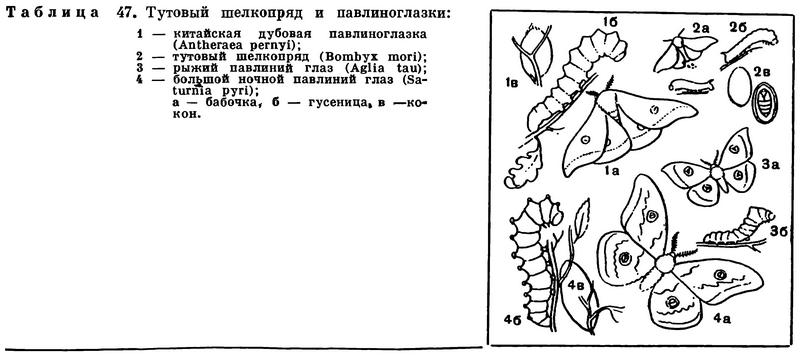
Размеры насекомых варьируют в очень широких пределах: самые крупные превосходят по длине тела самых мелких примерно в 1000 раз. Самые крупные представители современных насекомых — обитатели тропиков. Среди жуков жук-геркулес (Dynastes hercules, рис. 262) и жук-слон (Megasoma elephas) достигают длины 12 см; палочник Phybalostoma cervicornis имеет длину 26 см; хищный водный клоп Belostoma grande — 11,5 см; шелкопряд Attacus atlas в размахе крыльев достигает 24 см, а бабочка Thysania agrippina (рис. 328) — 26 см. Даже в фауне Европейской части СССР есть крупные насекомые. Наш большой дубовый усач (Cerambyx cerdo, рис. 284, 2) достигает длины 5 см, большой ночной павлиний глаз (Saturnia pyri, табл. 47, 4) — 15 см в размахе крыльев. Самые же мелкие из насекомых имеют длину меньше 0,25 мм (жуки-перистокрылки — сем. Ptiliidae, наездники-яйцееды — сем. Mymaridae). Таким образом, самые мелкие насекомые мельче, чем самые крупные простейшие, а самые крупные — больше, чем самые мелкие млекопитающие.
Несмотря на огромное разнообразие условий и образа жизни, на исключительные различия размеров и обилие видов, чаще всего, увидев насекомое, можно с уверенностью сказать, что это насекомое.
Для насекомых характерно то, что тело их разделено на 3 отдела — голову, грудь и брюшко, каждый из которых выполняет определенную функцию. На голове находятся основные органы чувств, позволяющие ориентироваться в пространстве, и ротовые органы — органы захватывания пищи. В грудном отделе у насекомых сосредоточены основные органы передвижения — двигательные конечности (ноги), а у взрослых высших насекомых — крылья. Брюшко — отдел, в котором сосредоточена основная масса внутренних органов, включая среднюю и заднюю кишку, жировое тело, выделительные и половые органы, совокупительный аппарат или яйцеклад и т. д. Такое деление тела на отделы (рис. 107) хорошо прослеживается у большинства взрослых насекомых и бывает не выражено либо у личинок, либо у таких взрослых насекомых, строение которых вторично изменилось под влиянием неподвижного или паразитического образа жизни.
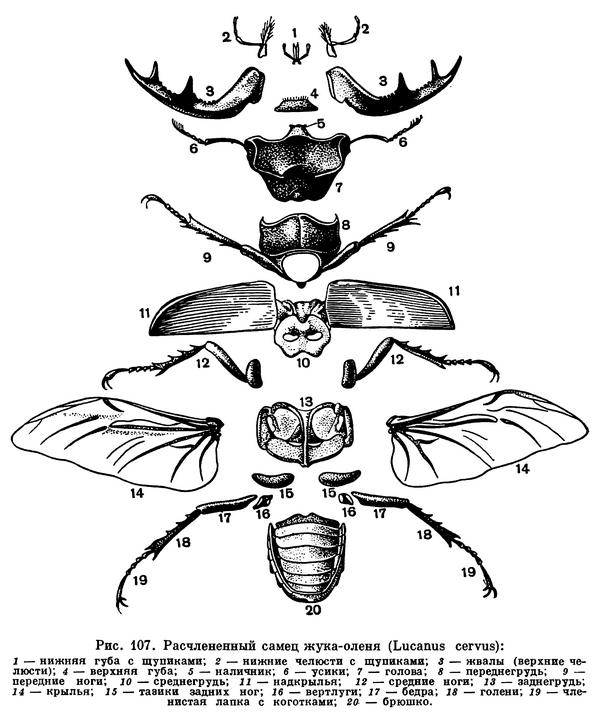
Голова образуется из 6 сегментов, закладывающихся у зародыша, но у сформировавшегося насекомого границы сегментов головы не видны — сегменты полностью сливаются.
Грудных сегментов у насекомых всегда 3. Каждый грудной сегмент несет по паре ног, а у многих насекомых ко второму и третьему сегментам причленяются органы полета — крылья, представляющие не конечности, а специализированные складки. В грудном отделе сосредоточена основная двигательная мускулатура насекомых.
Брюшко у насекомых образуется за счет закладки не более 12 зародышевых сегментов, у большинства высших десятый сегмент является последним. В некоторых случаях передние брюшные сегменты сливаются с грудным отделом, явственно отделяясь от остального брюшка перехватом. Такой случай наблюдается у жалоносных перепончатокрылых, например у пчел. Еще реже встречается слияние грудных сегментов с головой, как это имеет место у самок веерокрылых (Strepsiptera, рис. 307, 2; 309, 2), но в этом случае слияние связано с общим регрессом в развитии.
Наружный покров у насекомых представлен сложной кутикулой, выделяемой слоем расположенных под ней клеток — гиподермой. Кутикула покрывает не только открытые участки тела, но выстилает и передний отдел кишечника (рот, глотку, пищевод), заднюю кишку, ближайшие к дыхальцам участки трахейных трубок. У наземных насекомых в кутикуле различаются прилегающий к живым клеткам внутренний слой — эндокутикула, очень эластичная, составляющая основную часть толщи панциря, следующая за ней механически прочная экзокутикула и самая наружная, содержащая много жироподобных и воскоподобных веществ эпикутикула, имеющая ничтожную толщину — обычно менее 1 мк. Эпикутикула не пропускает воду. Она не развивается у большинства личинок насекомых, обитающих во влажной почве, в воде или в тканях растений. Чем суше та среда, где живет насекомое, тем яснее у него выражена эпикутикула. Естественно, что в кутикуле, выстилающей кишечник, нет непроницаемого слоя — эпикутикулы.
На теле насекомых кутикула развита неравномерно. Особенно сильно она выражена на сегментах; между сегментами находится нежная перепонка, состоящая из гораздо более тонких эндокутикулы и экзокутикулы. Так как складки перепонки обычно бывают скрыты под более уплотненными щитками, на ней эпикутикулы иногда нет.
Уплотненные толстые щитки на сегментах называются склеритами. Они развиты сильнее там, где больше требуется защита от повреждений, и там, где прикрепляются мышцы. Склериты туловища у насекомых расположены симметрично с правой и левой сторон, но на спинной стороне правый и левый склериты часто сливаются, образуя единый щиток — тергит. Склериты по бокам тела называются плейритами, а на брюшной стороне — стернитами.
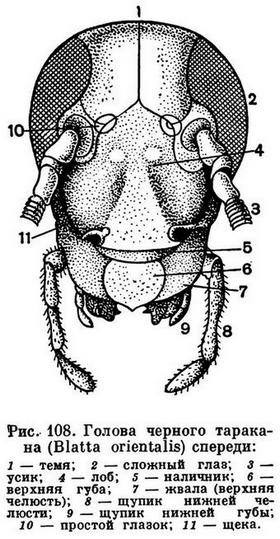
Голова у насекомых почти всегда сильно склеротизована, часто сильнее, чем другие отделы. Это объясняется тем, что прочная сплошная головная капсула у насекомых служит местом прикрепления мощной челюстной мускулатуры. Вспомним, что возникновение черепной коробки и у позвоночных было связано с этой же функцией. Слияние сегментов, из которых состоит голова, настолько велико, что у сформировавшегося насекомого никаких следов границ сегментов нет. Швы, которые наблюдаются на голове (рис. 108), — это не границы сегментов, а либо места прикрепления мышц, либо те линии, по которым идет растрескивание черепной капсулы при линьке. Но при изучении эмбрионального развития насекомых установлено, что голова у них образована за счет 6 зародышевых сегментов. Первому сегменту соответствуют глаза, второй сегмент несет усики, третий сегмент («вставочный») не несет придатков и сливается со вторым, на четвертом, пятом и шестом сегментах развиваются парные части ротового аппарата — верхние челюсти (жвалы, или мандибулы), нижние челюсти (максиллы) и нижняя губа. Эти ротовые части у зародыша закладываются точно так же, как закладываются ноги на следующих сегментах тела. Это дает основание считать ротовые части насекомых конечностями, вошедшими в состав ротового аппарата. Что конечности могут входить в состав ротового аппарата, хорошо видно на примере губоногих многоножек, у которых первая пара туловищных ног, сохраняя еще типичное для ног расчленение, стала действовать в сущности как мандибулы. То, что головная капсула имеет значение как место прикрепления мышц, доказывается тем, что, например, у личинок мух, питающихся уже полностью переваренной пищей, головы нет.
Глаза у насекомых бывают двух типов — фасеточные, как и у высших ракообразных, и простые. Фасеточные глаза всегда располагаются по бокам головы и свойственны большинству взрослых насекомых. Иногда они занимают большую часть поверхности головы и у многих хорошо летающих насекомых число фасеток в них огромно. У стрекоз, например, в сложном глазу бывает до 28 000 фасеток, у бабочек — до 17 000, у комнатной мухи — 4 000.
У видов насекомых, разные особи которых ведут неодинаковый образ жизни, можно наблюдать зависимость сложности фасеточного глаза от биологических особенностей. Например, у муравья-вора (Solenopsis fugax) бескрылые рабочие особи, живущие в ходах почвы, имеют глаза, состоящие из 6—9 фасеток. В глазах самок, летающих во время роения, фасеток около 200, а у самцов, которые во время полета должны найти и опознать самок своего вида, в глазу свыше 400 фасеток. У некоторых рабочих муравьев (Ponera punctissima) в каждом глазу по одной фасетке.
У личинок насекомых с полным превращением и у таких паразитических форм, как вши, а также у личинок щитовок (Coccidae) на месте сложных глаз развиваются боковые простые глазки. Иногда простых глазков бывает много (более 30 у личинок скорпионниц) и они выглядят как фасеточный глаз. Между фасеточным глазом и скоплением глазков основная разница заключается в том, что каждый простой глазок имеет свою кутикулярную роговицу, а в фасеточном глазу все омматидии, т. е. отдельные глазки сложного глаза, имеют общую роговицу.
Наряду со сложными глазами у многих хорошо летающих насекомых имеется по 3 или 2 простых глазка, расположенных между сложными глазами на лбу.
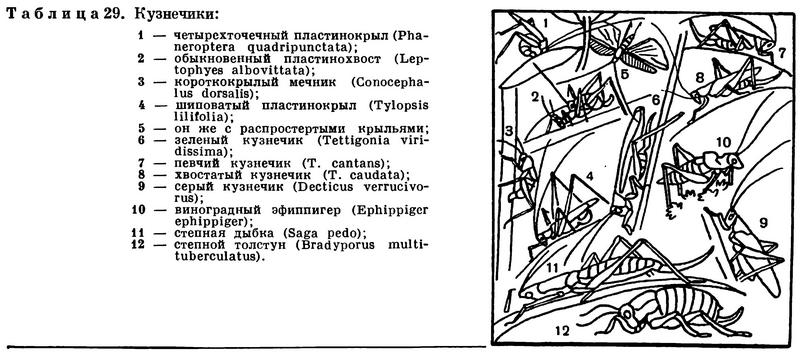
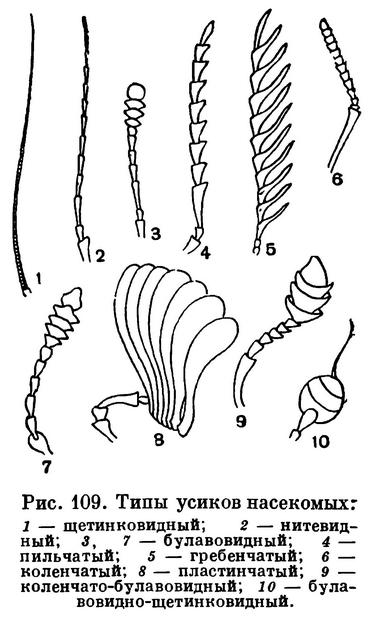
Усики (иначе называемые антеннами) у насекомых располагаются в передней части головы. Вариации строения усиков у насекомых невероятно широки, но, как правило, целые семейства, подотряды или отряды насекомых характеризуются той или иной определенной формой усиков. Обычно усики состоят из большого числа члеников, но иногда они короткие и члеников в них немного — у мух и стрекоз, например, в усиках всего по 3 членика, а у многих личинок, например у личинок долгоносиков и ручейников, вместо усиков имеются только едва заметные рудиментарные сосочки, у личинок мух усиков нет. У жуков усики обычно 11-члениковые, у а перепончатокрылых они состоят из 12—13 члеников. Иногда число члеников в усиках бывает очень велико, так как часто исходные членики распадаются на множество колец. У таких насекомых, как, например, кузнечики и некоторые жуки-дровосеки, усики бывают длиннее тела (рис. 154, 283, 284; табл. 29, 4; 40, 6, 43, 5).
Различают много форм усиков. Сужающиеся равномерно к вершине называются щетинковидными (как у таракана). Усики нитевидные, если все членики одинаковой толщины и каждый членик цилиндрический. Примером могут служить усики жужелиц (рис. 251). Если у члеников усиков середина усика толще, чем основание и вершина, усики четковидные, как у термитов. У таких жуков, как щелкуны, усики гребенчатые (рис. 268, 5). Если от каждого членика усиков в обе стороны отходит по тонкому выросту, усики перистые, как у ночных бабочек-шелкопрядов. Бывает, что усик на большей части своей длины более или менее одинаковой толщины, а у вершины расширяется — это булавовидный усик (у дневных бабочек). Если первый членик усика длинный, а последующие располагаются под углом к нему, усики коленчатые. Иногда у коленчатых усиков бывает булава, состоящая из веерообразно сложенных пластинок (пластинчатоусые жуки) или из гребешков (жук-олень). Бывают различной формы неправильные усики (рис. 247), иногда с причудливыми выростами, как у жука-нарывника из рода Cerocoma.
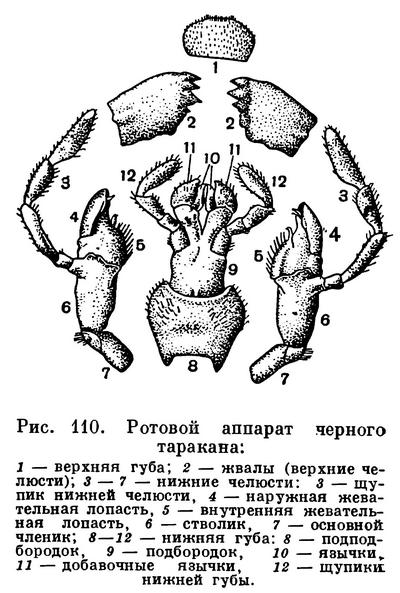
Ротовой аппарат включает 3 пары придатков, развивающихся как конечности. Наиболее хорошо выражены все ротовые части у представителей более древних групп насекомых, имеющих грызущий ротовой аппарат, при питании отгрызающих твердые кусочки пищи. Таковы тараканы, саранчовые и многие другие (рис. 110).
Первая пара челюстей — верхние челюсти — у всех насекомых состоит из цельных, нерасчлененных пластинок (жвал). По форме жвал бывает довольно легко установить, какой пищей питаются насекомые. У потребителей растительной пищи поверхность вершин жвал широкая, зазубренная, приспособленная к отрыванию твердых тканей, а у основания жвал широкая бугорчатая перетирающая поверхность — полная аналогия резцам и коренным зубам травоядных млекопитающих. Такие жвалы — у саранчовых, гусениц, многих жуков. А у хищников, таких, как жуки-скакуны, личинки златоглазки и др., жвалы серповидно изогнутые с острыми концами. Такие жвалы аналогичны серповидно изогнутым клыкам хищных млекопитающих — клыкам, приспособленным к пронзанию тела добычи и разрыванию его. Жуки-скакуны по-айглийски недаром даже называются tiger-beetles — «жуки-тигры» — и за хищный образ жизни, и за сходство их мандибул с клыками саблезубого тигра.
Так характер пищи накладывает отпечаток на строение верхних челюстей, играющих наиболее ответственную роль в грызущем ротовом аппарате. Мандибулы бывают очень прочными. Достаточно сказать, что известны случаи, когда личинки земляных усачей повреждали своими мандибулами свинцовую оболочку подземного электрического кабеля, а жуки-короеды и точильщики легко прокладывают ходы в твердой древесине дуба.
У хищных личинок жуков-плавунцов серповидные мандибулы служат не только для прокалывания добычи. В каждой мандибуле у них есть желобок, через который личинка сперва вводит пищеварительные соки в тело жертвы, а затем, когда внутри тела схваченного животного белки его мышц разжижаются и перевариваются впрыснутыми ферментами, личинка плавунца всасывает получающуюся переваренную кашицу через эти же каналы.
Следующая пара челюстей — нижние челюсти — у насекомых с грызущими ротовыми частями представляет членистые образования. Каждая челюсть состоит из основного членика, стволика и на вершине несет две жевательные лопастинки — внутреннюю и наружную. Часто они сливаются вместе или одна из них недоразвивается. От стволика отходит и членистый чувствующий придаток — щупик. Щупики иногда похожи по строению и функции на усики (например, у личинок жужелиц), иногда целиком редуцируются (у стрекоз, личинок сетчатокрылых).
Принципиально так же устроена и нижняя губа у насекомых, которая развивается из двух зачатков конечностей, срастающихся затем под ротовым отверстием так, что из частей, соответствующих основным членикам, образуется непарный подподбородок, а из стволиков — подбородок. Жевательные пластинки нижней губы не срастаются, а образуют язычки и придаточные язычки. Иногда нижняя губа становится основным хватательным аппаратом, выполняя такую роль, как направленные вперед мандибулы у многих хищных личинок. Это характерно для личинок стрекоз (рис. 182).
В ряде случаев ротовой аппарат сохраняет типичное для грызущего типа строение, а функционирует как сосущий, причем канал для введения в тело добычи фермента образуется не внутри мандибулы, а трубкообразным просветом между мандибулой и соответствующей максиллой. Такие каналы имеются у личинок муравьиного льва, личинок светляков.
В большинстве случаев в состав грызущего ротового аппарата входят не только 3 пары конечностей, но и прикрывающая ротовое отверстие сверху непарная складка покровов — верхняя губа, подвижно сочлененная с передним отделом головной капсулы. У личинок жуков, прокладывающих ходы направленной вперед головой, верхняя губа часто редуцирована (личинки щелкунов, жужелиц). В ротовой аппарат входят также непарные утолщенные части стенок ротовой полости — надглоточник, часто сливающийся с верхней губой, и подглоточник, подходящий к середине нижней губы.
У тех насекомых, которые добывают пищу высасывая соки из тканей растений или животных, ротовые части преобразуются в колюще-сосущий аппарат — в хоботок. У полужесткокрылых, например, вытянутая нижняя губа превращается в трубчатый футляр со спрятанными внутри способными выдвигаться колющими щетинками, в которые превратились верхние и нижние челюсти (рис. 221).
У комара колющими щетинками становятся и другие части ротового аппарата — подглоточник и верхняя губа с надглоточником (рис. 407, В). Жидкая пища (соки растений, кровь) поступает из ранки в рот насекомого по капиллярам, образованным соприкасающимися колющими щетинками, или по желобкам в них.
У бабочек, сосущих нектар (что не требует прокалывания твердых тканей), развивается хоботок, состоящий из сложенных, очень сильно вытянутых нижних челюстей. Верхние челюсти полностью исчезают, нижняя губа представлена маленькой пластинкой, от которой отходят хорошо развитые нижнегубные щупики, выполняющие функцию органов химического чувства (рис. 320).
Описание всех тех преобразований, которые характерны для ротового аппарата разных насекомых, требует нескольких специальных томов. Отметим только, что иногда ротовые части с утратой потребности в них сильно или целиком редуцируются. Последнее наблюдается у личинок многих мух, живущих в том веществе, которое служит им пищей (например, личинки мясных мух Calliphora в трупах животных); они разжижают пищу, отрыгивая пищеварительные соки наружу. Пища переваривается личинкой вне организма и всасывается в жидком легко усваиваемом виде. У таких личинок вместо 3 пар челюстей остается только пара ротовых крючков, помогающих при ползании. Не развиваются ротовые органы и у многих взрослых непитающихся крылатых насекомых (поденки, самцы щитовок, самцы бабочек-чехлоносок).
У всех насекомых за головой следует грудь. Иногда голова незаметно переходит в грудь, широко с ней соприкасаясь, иногда бывает отделена тонким перехватом. Большинство насекомых может двигать головой только сверху вниз или наклоняя ее в стороны, и только богомолы (Mantodea) могут поворачивать голову, как это делаем мы. Грудь состоит из передне-, средне- и заднегруди. У первичнобескрылых сегменты груди подвижны друг относительно друга, а в большинстве случаев происходит слияние средне- и заднегруди в единое целое. Развитие отдельных сегментов груди зависит от степени участия разных ее придатков в движении.
У взрослых насекомых каждый из сегментов груди несет по паре конечностей, состоящих из следующих отделов: тазик, вертлуг (у некоторых перепончатокрылых он двойной — есть еще вертлужок), бедро, голень и лапка. Число члеников лапки бывает 1—5, но не более 5. Если все пары ног развиты одинаково, конечности называются ходильными или бегательными (палочники, жуки-жужелицы, табл. 28, 39, 10). Такие конечности приспособлены к бегу по твердой поверхности. Иногда «твердая» поверхность может быть и очень зыбкой — вспомним водомерок, у которых на лапках выделяется жировой слой, препятствующий смачиванию и прорыву поверхностной пленки воды.
При обитании в траве или листве деревьев много преимуществ дает движение сильными прыжками. При прыжке наибольший эффект обеспечивает быстрое распрямление задних конечностей, толкающих тело вперед (это наблюдается и у четвероногих: лягушка, заяц, тушканчик имеют более развитые задние конечности, чем передние). У многих насекомых, передвигающихся таким образом, сильно удлиненные мощные бедра и длинные голени задних ног. Такие задние ноги, как у саранчовых, кузнечиков, жуков земляных блошек, клопов-прыгунов (Halticus), называются прыгательными (табл. 29, 31).
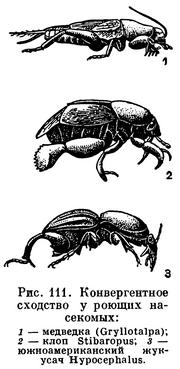
Жизнь взрослых насекомых в почве приводит к усилению значения передних ног при движении — к развитию роющих конечностей, характеризующихся расширением лопатообразных голеней (у медведок, у южноамериканских цилиндрохет, жуков-карапузиков и др., рис. 111).
У тех насекомых, которые передними ногами ловят добычу, бедра и голени передних ног удлиненные, с шипами на сторонах, сближаемых при складывании; такие ноги называются хватательными (у мантисп, богомолов, табл. 25, рис. 239).
У насекомых, перешедших к жизни в воде, ноги часто бывают с удлиненными плоскими голенями, покрытыми длинными волосками. Такие плавательные ноги работают как весла.
Редукция ног у взрослых насекомых наблюдается при переходе к неподвижному образу жизни и может быть полной (у настоящих щитовок — Diaspinae). У личинок многих насекомых утрата конечностей бывает связана с приспособлением к червеобразному передвижению в почве и в сходных субстратах (например, у личинок двукрылых, долгоносиков и др.). У насекомых с полным превращением (жуки, сетчатокрылые), если личинки даже имеют развитые бегательные ноги, они никогда не имеют такого полного расчленения, как у взрослых форм, — часто происходит слияние голени и лапки (образуется голенелапка), лапка всегда одночлениковая и имеет один коготок (иногда двойной). Иногда ноги у личинок сохраняются только в виде одночлениковых образований (например, у зерновок).
Для высших насекомых характерна способность к активному полету с помощью крыльев. Крылья развиваются полностью только при переходе насекомого во взрослое состояние, а закладываются как плоские листовидные выпячивания покровов средне- и заднегруди. Это выпячивание разрастается, его противоположные стенки плотно сближаются, в результате чего образуется эластичная пластинка, укрепленная жилками, представляющими собой затвердевшие участки тех путей, по которым во время формирования крыла в него нагнеталась гемолимфа, расправляя крыловую пластинку. Жилки располагаются так, что становятся механической опорой крыла и помогают наилучшим образом преодолевать сопротивление воздуха при полете. В частности, у хорошо летающих насекомых передний край крыла укреплен мощной жилкой. Расположение и число жилок — признаки, хорошо характеризующие каждую систематическую группу насекомых. Строение крыльев в значительной мере кладется в основу систематики крылатых насекомых, а так как в ископаемом состоянии от насекомых лучше всего сохраняются крылья, палеоэнтомология — наука об ископаемых насекомых — особенно широко использует признаки строения крыла. В крыле различаются продольные жилки, частично ветвящиеся, и поперечные, соединяющие продольные друг с другом. Просветы перепончатой части крыла, заключенные между жилками, называются ячейками. Каждая жилка соответственно ее расположению на крыловой пластинке имеет определенное наименование, так же как и ячейки.
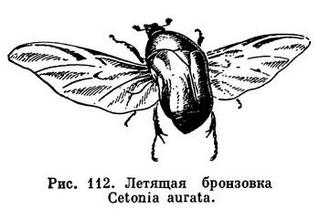
У более примитивных групп насекомых сеть жилок гуще, чем у представителей высших отрядов. Кроме того, у представителей высших отрядов передние и задние крылья обычно значительнее отличаются друг от друга, чем у более примитивных, у которых обе пары крыльев довольно сходны. Нередко развивается лишь одна пара крыльев, чаще передние (у мух, самцов щитовок, некоторых поденок), но бывает и наоборот — развиты только задние крылья (у веерокрылых). Нередко крылья, особенно передние, принимают на себя дополнительные функции, а часто такие дополнительные функции становятся основными, а летная функция утрачивается. Например, передние крылья у прямокрылых (саранча, кузнечик) более кожистые, чем задние; в сложенном состоянии более тонкие перепончатые задние крылья прикрыты передними, защищающими от повреждений при передвижениях в зарослях растений. У клопов защитная функция передних крыльев, превратившихся в надкрылья, выражена еще ярче: у них только самая вершина передних крыльев перепончатая (рис. 220). А для самого богатого видами отряда насекомых — жуков характерно то, что передние крылья целиком превратились в жесткие защитные надкрылья. При полете надкрылья жуков играют роль только планирующих выростов, не производящих собственных движений, летные движения совершают задние крылья. Вероятно, это наблюдали многие, кто видел, как летают божьи коровки, жуки-нарывники, жуки-олени и др. А иногда у жуков при полете надкрылья даже не расправляются — задние крылья только высовываются в стороны из-под надкрылий (у бронзовок, рис. 112).
Нередки случаи утраты летной функции крыльев и их исчезновение. Для многих жуков, живущих в сухих местностях, характерно, что у них надкрылья полностью срастаются друг с другом, образуя защитный «шлем» над брюшком. Естественно, что со срастанием надкрылий задние крылья не могут действовать и исчезают. Это наблюдается у многих долгоносиков-скосарей, жужелиц (хлебная жужелица, рис. 251, 2) и особенно часто у степных и пустынных чернотелок.
У некоторых широко распространенных видов чернотелок можно проследить разные степени редукции крыльев. Так, например, песчаный медляк (рис. 113) во Франции местами совершает перелеты, а на нашей территории это жук нелетающий, у которого можно проследить все переходы от развитых, но бездействующих крыльев под надкрыльями до жалких, почти полностью исчезнувших рудиментов.

В разных группах насекомых можно столкнуться с бескрылыми формами, целиком утратившими крылья. Это относится и к тем группам, большинство представителей которых хорошо летает: среди двукрылых, бабочек, прямокрылых, клопов известны формы с исчезнувшими крыльями. В таких случаях бывает ясен факт утраты крыльев, существовавших у предков этих насекомых. И рассматривая черты внутреннего строения, и сравнивая бескрылые виды с рядом представителей той же группы, имеющих развитые и редуцированные органы полета, можно установить, что бескрылые формы произошли от крылатых предков. Такая утрата крыльев может быть вызвана разными причинами (переходом к паразитизму, к жизни в почве, при обитании на островах или в горах и т. д.).
Даже в тех случаях, когда целые отряды включают совершенно бескрылых насекомых (вши, пухоеды, блохи), изучение их строения и развития в сравнении с другими группами насекомых позволяет считать у них отсутствие крыльев следствием редукции и утраты. Вероятно, у предков, например, блох были крылья, исчезнувшие с приспособлением взрослой стадии к паразитизму.
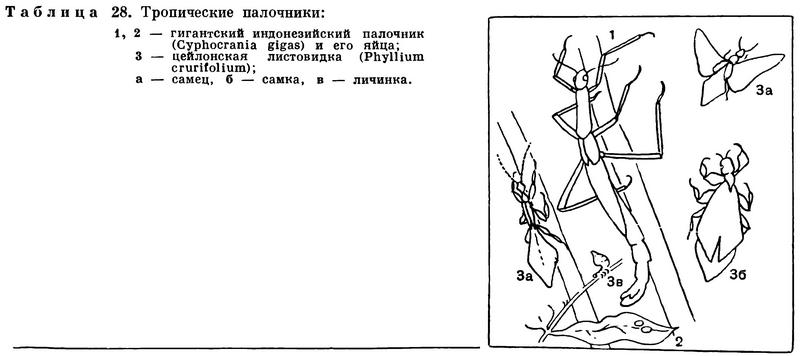
Но среди низших насекомых есть и такие отряды, представители которых никогда и не имели крыльев: отряд щетинохвосток, например, заслуживает названия первичнобескрылых. Все особенности представителей этого отряда дают основание считать его происшедшим от далеких бескрылых предков, бывших, однако, и предками крылатых насекомых.
У насекомых, имеющих более или менее сходные передние и задние крылья, снабженные густой сетью жилок, т. е. у более примитивных (стрекозы, сетчатокрылые, термиты и др.), передние и задние крылья при полете двигаются независимо друг от друга, а у бабочек, перепончатокрылых, цикад и других более высокоорганизованных форм переднее и заднее крылья сцепляются так, что образуют единую согласованно двигающуюся поверхность. У мух, имеющих только передние развитые крылья, задние крылья превращены в небольшие булавовидные придатки, богатые чувствующими волосками. Они работают как гироскопы, служат стабилизаторами при полете. Изменение направления при полете достигается изменением положения плоскости передних крыльев (у дневных бабочек) или амплитуды и плоскости вибрации (у пчел).
Скорость вибрации крыльев у разных насекомых значительно варьирует. Бабочка махаон (табл. 43, 2) при полете делает только 5—9 взмахов крыла в секунду, пчела около 200, а комары-звонцы — 1000. Такая скорость сокращений обеспечивается резко и густо исчерченными поперечнополосатыми мышцами, на долю которых у хорошо летающих форм приходится 15—25% веса тела.
Скорость полета у насекомых очень различна: у златоглазки — 0,5 м, у пчелы — 3 м, у бабочек бражников — 15 м в секунду. Самые сильные насекомые, такие, как бражники, без посадки летят на сотни километров. Известны, например, случаи залета олеандрового бражника (Deilephila nerii), распространенного в средиземноморских странах (где растет олеандр), в Ленинград и Эстонию.
На втором — девятом брюшных сегментах только у первичнобескрылых есть одночлениковые рудименты конечностей. Кроме того, на конце брюшка у низших насекомых бывают придатки (церки), выполняющие роль органов чувств (особенно это выражено у эмбий) или хватательных органов (у уховерток). Видоизмененные зачатки конечностей сегментов рядом с половым отверстием служат вспомогательными органами при оплодотворении (гоноподиями). У низших групп насекомых самки гоноподиями подхватывают пакет с семенной жидкостью — сперматофор.
Кутикула многих насекомых покрыта различной длины волосками, иногда очень густо. Те волоски, которые представляют просто выросты кутикулы, служат главным образом для того, чтобы при затоплении насекомого (во время дождя и. т. п.) создать вокруг тела слой воздуха, изолировать тело от соприкосновения с водой. Но кроме них, на теле насекомых много волосков и щетинок, подвижно сочлененных с ямкой в кутикуле, к основанию которых подходят окончания нервов — это основные осязательные элементы насекомого.
Нередко тело (у чешуйниц) и крылья (у жуков, бабочек и др.) бывают покрыты чешуйками. Волоски и чешуйки определяют у многих насекомых окраску. Например, у бабочек яркая пестрая окраска крыльев зависит не от присутствия какого-либо пигмента, а от явлений светового резонанса при преломлении световых лучей через чешуйки — сами чешуйки бесцветны. От структуры кутикулы зависит и металлическая окраска жуков, перепончатокрылых и других насекомых. Такая окраска, зависящая не от пигмента, а от структуры покровов, называется оптической. Она была расшифрована в 1902 году киевским физиком И. И. Косоноговым. Наряду с оптическими явлениями окраску многих насекомых определяют и отлагаемые клетками гиподермы пигменты. Черная окраска чаще всего зависит от отложений меланина. Распространенная у насекомых зеленая окраска (у саранчовых, палочников и др.) долгое время рассматривалась как связанная с хлорофиллом растений, но, по новым данным, она определяется особыми пигментами.
Окраска многих насекомых, особенно определяемая преломлением лучей в хитиновых частях покровов, сохраняется и у мертвых высушенных экземпляров. Так, бабочки, собранные и наколотые уже более 200 лет тому назад К. Линнеем, сохраняют такую же свежесть и яркость тонов, как живые. Прекрасно сохраняют металлический блеск и многие жуки, например бронзовки. А яркая окраска надкрылий божьих коровок, прямокрылых и других насекомых, у которых она зависит от органических пигментов, у собранных высушенных экземпляров тускнеет и выцветает. Выцветает иногда и металлическая окраска, например у жука щитоноски (Cassida nobilis).
Изумительно многообразие расцветок насекомых. Многие из них по окраске могут соперничать с пестрыми и яркими цветами, а многие имеют тусклую неяркую окраску, делающую их непривлекательными и незаметными. Чем же объясняется такое многообразие окрасок насекомых?
Окраска имеет огромное биологическое значение. Не защищенные другими способами насекомые, если их окраска напоминает тот фон, на котором они держатся, становятся незаметными для истребляющих их хищников, в первую очередь для птиц. Каждый собиратель насекомых знает, как трудно бывает обнаружить сидящую на вбитом в землю потемневшем деревянном столбике озимую совку, как незаметны бывают сидящие на покрытом лишайниками стволе дерева бабочки пяденицы, как легко пройти мимо зеленой гусеницы, сидящей на листе, как ускользает из поля зрения прыгающая на поверхности серой почвы серая саранча и т. п.
Итальянский ученый Чеснола делал такие опыты. Он собирал крупных богомолов, имеющих сероватую или зеленую окраску. Сероватые незаметны на фоне сухой травы, где они держатся, а зеленые — на зеленой. Он привязывал равное количество богомолов разной окраски к травинкам такого же цвета или к травинкам другого (половину сероватых к сухой траве, половину к зеленой, половину зеленых к сухой, половину к зеленой). Оказалось, что количество склеванных птицами богомолов было больше в тех случаях, когда они были привязаны к травинкам другого цвета, что больше выжило тех, которые были на «своем цвете». Опыты Чеснолы у нас были проверены М. М. Беляевым. Окраска, делающая насекомое незаметным на окружающем фоне, называется покровительственной (табл. 16).
Металлически блестящие места покровов насекомых в естественной обстановке отражают преобладающий окружающий фон и таким образом служат тоже своеобразной защитной окраской.
Часто покровительственной бывает не только окраска, но и форма тела — у тех же богомолов вытянутое тело напоминает стебелек травинки. Еще ярче выражена покровительственная окраска и форма у палочников, у гусениц пядениц, тело которых часто похоже на сучок, и т. д.
Интересна так называемая криптическая окраска у насекомых. Бывает, что причудливый узор, например на крыльях бабочки, не привлекает внимания хищника, а, наоборот, делает насекомое незаметным. Так, глазчатые пятна на крыльях большого ночного павлиньего глаза делают крылья похожими на поверхность коры с углублениями — рисунок имитирует, если смотреть издалека, неровности поверхности, создает стереоскопический эффект. Такие примеры «стереоморфизма» изучал ленинградский профессор Б. Н. Шванвич.
Но бывает, что окраска насекомого как бы нарочно привлекает внимание. В таких случаях насекомое обычно надежно защищено от хищников отталкивающим запахом или вкусом, ядовитыми железами и т. п. Так, яркую окраску имеют многие клопы, например солдатики; жуки-нарывники, божьи коровки выделяют едкие вещества, например кантаридин. Ярко окрашены и защищены ядовитым жалом осы, например, шершни. Такая яркая окраска носит название предупреждающей (табл. 16). Действительно, замечено, что птицы научаются не трогать таких защищенных насекомых.
Следует, правда, иметь в виду, что нередко окраска, воспринимаемая нами как пестрая, для хищника, истребляющего данный вид насекомого, может представляться однотонной и быть защитной. Известно, что способность различать цвета неодинакова у разных видов животных и, например, собака цвета не различает. Способность же разных видов птиц различать цвета специально не изучалась.
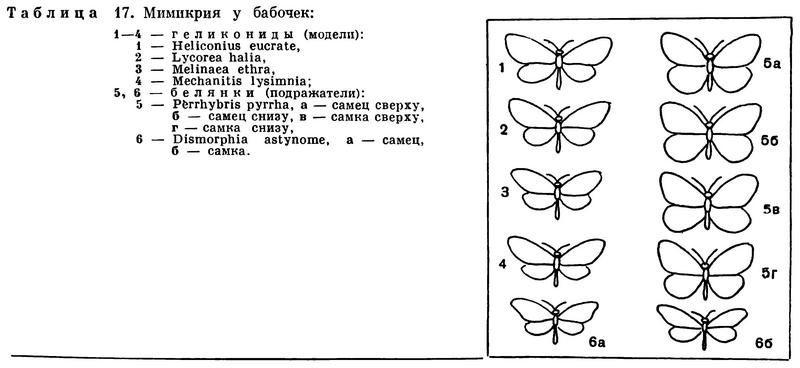
Большое распространение в мире насекомых имеет так называемая мимикрия (табл. 17) — сходство очертаний и окраски двух видов, из которых один характеризуется предупреждающей окраской и несъедобен или опасен для преследователей, а другой, незащищенный, имитирует защищенный. Неопытному наблюдателю бывает трудно отличить мух-сирфид от пчел и ос — настолько совершенно и расцветка, и характер движений этих беззащитных мух имитируют жалоносных ядовитых перепончатокрылых. Неискушенный собиратель насекомых нередко опасается взять в руки сирфиду, думая, что имеет дело с пчелой, осой или шмелем. Похожи на ос и бабочки-стеклянницы. Очень интересен пример мимикрии южноамериканских бабочек-белянок, «подражающих» ядовитым бабочкам-геликонидам, около 100 лет тому назад описанный Бэтсом.
Во всех таких случаях незащищенное насекомое меньше истребляется птицами, ящерицами и другими врагами, так как они принимают его за ядовитую «модель».
Иногда, конечно, окраска имеет и прямое приспособительное значение. Так, жуки-чернотелки в пустынях характеризуются либо белой окраской (парадокс — чернотелки, а белые!), отражающей лучи солнца, либо чаще (отсюда и название семейства) интенсивно черной — плотные черные покровы задерживают губительные короткие лучи спектра.
На поверхности тела у насекомых особыми канальцами открываются различные железы, выделяющие пахучие вещества, иногда воспринимаемые только другими насекомыми, иногда доступные и нашему обонянию (например, у различных клопов, жуков-медляков рода Blaps и т. д.). Часто пахучие выделения помогают особям одного вида находить друг друга (выделения самок привлекают самцов, запахи залегающих на зимовку клопов-черепашек или божьих коровок обеспечивают скопление зимующих особей и т. д.). Некоторые выделения желез имеют пищевую привлекательность для особей того же вида (например, выделения шейных желез самцов кузнечиков слизываются самками) или для других видов (выделения жуков, обитающих в муравейниках, охотно слизываются муравьями). Иногда железы выделяют ядовитые вещества. Поступающие в полые волоски ядовитые выделения служат средством защиты — известно, что ядовитые волоски многих гусениц, например златогузки, могут вызывать воспалительные процессы кожи человека. Едкие защитные вещества выделяют многие жуки (кантаридин — жуки-нарывники, педерин — некоторые стафилиниды). Такие яды могут вызывать явления воспаления и омертвения покровов, если попадают на кожу. Нередки у насекомых железы, выделяющие воск и воскоподобные вещества, часто служащие насекомым для защиты от потери воды (щитки у щитовок, восковые нити у червецов и подушечниц и т. д.).
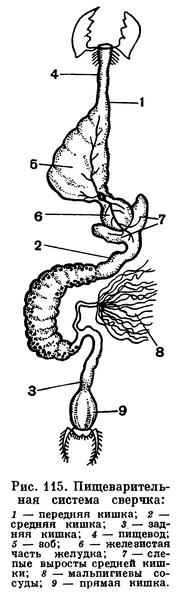
Пищеварительная система у насекомых, как и у всех членистоногих, состоит из передней, средней и задней кишки (рис. 115).
Передняя кишка начинается ротовой полостью, в которую впадают слюнные железы. Слюнные железы у разных насекомых вырабатывают разные ферменты. У растительноядных это чаще всего ферменты, переводящие дисахариды в моносахариды (инвертазы). У хищников, например у водных клопов, в слюне содержится фермент, расщепляющий белки на аминокислоты (протеаза). У кровососов часто в слюне есть фермент, препятствующий свертыванию крови (например, у малярийного комара).
На нижней губе у личинок бабочек, ручейников и ряда других насекомых открываются железы, секрет которых быстро затвердевает; это шелкоотделительные железки, продукт которых употребляется для изготовления различных защитных (коконы) или ловчих (тенета ручейников) образований. Трубчатый пищевод в задней части нередко расширяется в зоб, иногда приобретающий вид мешка, особенно развитый у тех насекомых, которые питаются жидкой пищей.
Средняя кишка является тем местом, где в основном переваривается пища. Железистые стенки среднего отдела кишечника (его расширенная часть называется желудком) вырабатывают ферменты, которые расщепляют и разжижают служащие пищей вещества.
Кроме ферментов (вырабатывающихся и у нас), расщепляющих белки, жиры, крахмал и сахар, многие насекомые соответственно роду пищи вырабатывают и другие энзимы. Многие личинки жуков-дровосеков вырабатывают целлюлазу, разрушающую клетчатку, личинки кожеедов выделяют кератиназу, действующую на роговые вещества, и т. и. Очень часто такие трудноразрушаемые соединения, как клетчатка, разрушаются с помощью симбиотических бактерий и простейших. Известно, что термиты потребляют в основном клетчатку, но своих разлагающих ее ферментов не выделяют — у них в кишечнике много симбиотических многожгутиковых (Trichonympha и др.), усваивающих целлюлозу. У тех насекомых, которые используют кишечную микрофлору для переваривания пищи, кишка имеет большие слепые выросты или достигает очень большой длины и пища задерживается в кишечнике на длительные сроки (например, у личинок майского хруща). Недавно было выяснено, что те насекомые, которые питаются бедными белками соками растений (например, тли), усваивают азот воздуха с помощью симбиотических азотфиксирующих бактерий.
В средней кишке пищевая масса обычно одевается особой легкопроницаемой белковой оболочкой, защищающей нежные клетки кишки от повреждений. Оболочка выделяется самым передним участком средней кишки. Средняя кишка не имеет кутикулы, а задняя, как и пищевод, покрыта проницаемой кутикулой.
В задней кишке происходит всасывание переваренной пищи и, главное, всасывание воды. Расположенные в конце пищеварительного тракта у многих насекомых так называемые «ректальные железы» служат для интенсивного всасывания воды.
Выделительными органами у насекомых служат мальпигиевы сосуды — тонкие трубчатые выросты кишечника на границе средней и задней кишки (рис. 115). Число их варьирует от 2 до 150, их свободные слепые концы лежат в полости тела, не сообщаясь с нею. Растворимые продукты обмена поглощаются стенками мальпигиевых сосудов и по их просветам проходят в заднюю кишку, обычно еще в них переходя в мелкие кристаллики мочевой кислоты. У тех более примитивных крылатых насекомых, которые живут во влажной среде, особенно у водных насекомых (стрекозы, поденки), или у тех, которые потребляют много сочной растительной пищи (прямокрылые), мальпигиевых трубочек много — они принимают на себя роль осморегуляторных образований, выводящих избыток воды. Слепые концы мальпигиевых трубочек у насекомых, живущих в сухих условиях, — у жуков и личинок чернотелок, гусениц, личинок муравьиных львов — прирастают к стенке задней кишки ниже того места, где эти трубочки впадают в кишечник. Просвета в месте присоединения нет, поглощение воды концом мальпигиева сосуда из задней кишки происходит путем диффузии. Этим обеспечивается непрерывное промывание мальпигиева сосуда одним и тем же небольшим объемом воды (рис. 116).
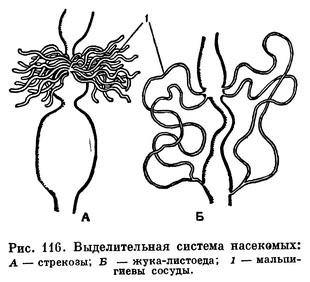
У некоторых первичнобескрылых насекомых (Machilis) наряду с мальпигиевыми трубочками некоторую роль в выделении играют открывающиеся на нижней губе железы — органы выделения, напоминающие максиллярные железы ракообразных.
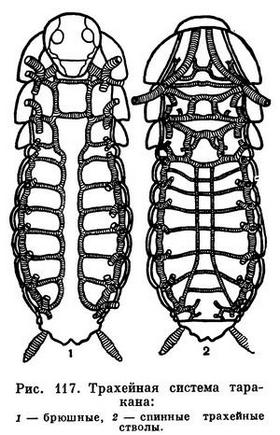
Основная дыхательная система у насекомых — трахеи. У живущих в воде, во влажной почве и в тканях растений личинок насекомых большую роль играет также кожное дыхание, от которого и произошло трахейное дыхание. У некоторых двукрылых вышедшая из яйца личинка еще не имеет трахей, которые развиваются только после линьки. Но чаще кожное дыхание существует наряду с трахейным, или, когда насекомые вторично переходят к кожному дыханию, у них сохраняется хорошо развитая система трахей, не имеющих сообщения с внешней средой.
Трахейная система у насекомых представляет сообщающуюся систему трахейных трубочек, открывающихся по бокам тела отверстиями — дыхальцами. Только у махилисов (Machilis) пучки трахей не сообщаются друг с другом.
Наибольшее число дыхалец у насекомых — 10 пар (по паре на первых 8 брюшных сегментах, 2 пары на грудных), но количество действующих дыхалец (у личинок насекомых, живущих в воде, во влажной земле и т. п.) может уменьшаться до 1 пары.
У насекомых, как и у других наземных членистоногих, кислород попадает от дыхалец к клеткам тела путем диффузии по просветам трахей и их тончайших веточек — трахеол. Но у многих высших насекомых имеется система вентиляции, проталкивания воздуха через крупные стволы трахей, что ускоряет газообмен. При движениях брюшка то расширяются, то сжимаются широкие трахейные стволы или связанные с ними воздушные мешки. А определенное направление тока воздуха в трахеях при таких дыхательных движениях обеспечено тем, что у таких насекомых дыхальца снабжены клапанами, то закрывающимися, то открывающимися. У тех личинок насекомых, которые живут в воде, во влажной земле и в других влажных средах, дыхальца всегда открыты для воздуха. Хорошо приспособившиеся к жизни в воде личинки поденок, веснянок, ручейников и других насекомых не имеют открытых дыхалец. Кислород у них проникает через поверхность всех участков тела, где покровы достаточно тонки, особенно через поверхность листовидных выростов, пронизанных сетью слепо заканчивающихся трахей. У таких личинок хорошо развита трахейная система, но механизм дыхания сложный — кислород проникает через покровы, растворяется в полостной жидкости, затем из полостной жидкости диффундирует в трахеи, затем уже по трахеям доходит до всех внутренних органов.
Многие живущие в воде насекомые (водные жуки и клопы, личинки и куколки комаров и др.) должны время от времени подниматься к поверхности, чтобы захватить воздуха, т. е. у них дыхание воздушное. Личинки комаров, водные личинки долгоножек и др. на время обновления запаса воздуха в своей трахейной системе подвешиваются с помощью венчиков несмачиваемых жирных волосков к поверхностной пленке воды. Водные жуки — водолюбы, плавунцы, — и клопы, например гладыши, обновив запас воздуха у поверхности, уносят его с собой вглубь — воздух уносится под надкрыльями и в виде слоя, приставшего к несмачиваемым волоскам на поверхности тела.
Оказалось, что, находясь под водой, такие насекомые тратят гораздо больше кислорода, чем содержится в запасе, унесенном ими из атмосферы. Дело в том, что между содержанием кислорода в соприкасающемся с водой слое воздуха на теле нырнувшего насекомого и окружающей водой устанавливается некоторое равновесие: по мере того как потребляется кислород из этого слоя, в него проникает кислород, находящийся в растворенном виде в окружающей воде!
Таким образом, слой воздуха на теле водного насекомого играет роль как бы особого приспособления к дыханию под водой (как водолазный колокол) и даже называется «физической жаброй». Вспомним, что несмачиваемые волоски на теле наземных насекомых имеют такое же значение в случае, например, затопления талыми водами в конце зимовки в почве: вокруг тела насекомого сохраняется слой воздуха — «физическая жабра».
У многих насекомых, например у личинок комаров-мотылей (Chironomus), нет трахейных жабр, но на заднем конце тела есть отростки с тонкими покровами, которые считались тоже жабрами («кровяные жабры»). А оказалось, что эти отростки — места поглощения солей из воды, а не жабры; дыхание у личинок мотылей кожное, всей поверхностью тела. Тело их кажется красным от просвечивающего гемоглобина. Но гемоглобин у таких личинок служит не для переноса кислорода, а для более прочного его связывания и запасания. Это бывает важно для живущих в илистом грунте личинок — там часто создаются анаэробные условия.
Кровяная жидкость — гемолимфа, циркулирующая в полости тела насекомых, — бесцветная. Гемолимфа не имеет большого значения для снабжения всех тканей кислородом — она служит для транспортировки растворенных в ней поступающих из кишечника питательных веществ, для выноса из клеток продуктов обмена, для распределения в теле гормонов. В гемолимфе есть клетки-фагоциты, помогающие насекомым справляться с попадающими в их тело микроорганизмами, а иногда и с более крупными паразитами. Фагоциты окружают и часто «инкапсулируют» паразита, попавшего в тело насекомого, что вызывает его гибель.
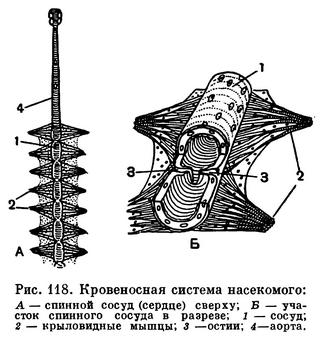
Циркуляция гемолимфы обеспечивается чередующимися расширениями и сокращениями трубчатого спинного сосуда — сердца. При растягивании стенок сосуда, осуществляемого крыловидными мышцами (рис. 118), гемолимфа, обогащенная питательными веществами, через боковые отверстия (остии) попадает в сердце, а при сокращении стенок сосуда клапаны в остиях закрываются и гемолимфа выталкивается через аорту к головному мозгу.
Нервная система у насекомых построена сходно с нервной системой низших членистоногих и даже кольчатых червей: есть головной мозг — надглоточные скопления нервных клеток — и брюшная нервная цепочка. Большого развития и усложнения достигает у насекомых головной мозг. Брюшная же цепочка состоит из подглоточного узла и обычно 10 груднык и брюшных ганглиев, причем соседние ганглии брюшной цепочки часто сближаются и полностью сливаются друг с другом. В головном мозге, всегда представляющем слитное образование, различают 3 части: «первичный» мозг, обычно самый большой отдел, связанный с органами зрения, «вторичный», связанный с усиками, и «третичный», дающий ветви к верхней губе и передней части кишечника. В мозге развиты в основном ассоциативные клетки, к которым подходят нервы от органов чувств. В центре «первичного» мозга есть «грибовидные тела», состоящие из скоплений ассоциативных клеток.
Чем сложнее поведение насекомого, тем сильнее у него развит головной мозг.
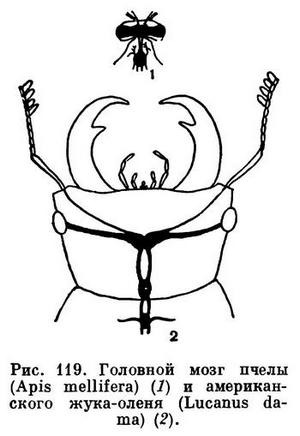
У пчелы он составляет 1/174 объема тела, а у жука-плавунца всего 1/420. У маленькой пчелы мозг даже абсолютно больше, чем у крупного жука-оленя (рис. 119).
Чем многообразнее раздражения, получаемые насекомыми, чем детальнее насекомое способно их различать и чем совершеннее возможности отвечать на раздражение многообразными движениями, тем сложнее поведение. Особенно сложным бывает поведение многих насекомых в период размножения.
Какие же раздражения ощущаются насекомыми? Они очень многообразны. У большинства ведущими органами чувств оказываются органы зрения, о чем можно судить и на основании особо сильного развития «первичного» мозга.
Конечно, не у всех насекомых глаза развиты одинаково, очень немногие виды подвергались изучению, но те материалы, которые есть, позволяют отметить, что насекомые (личинка стрекозы, пчела) воспринимают смену изображений как мелькание при несколько даже большей частоте, чем воспринимаем мы. Сложные глаза дают достаточно четкую картину близких предметов и позволяют различать силуэты отдаленных. Простые глазки, а у многих бабочек и сложные глаза хорошо воспринимают изменение интенсивности освещения. Для многих насекомых доказана способность различать цвета. Эта способность не всегда одинакова, но экспериментально показано, что тли распознают длинноволновую и коротковолновую части спектра (красный — зеленый участок отличают от синего — фиолетового), что шведскую мушку привлекают голубые оттенки на зеленом фоне, что муравьи различают ультрафиолетовые лучи, а пчелы хорошо различают четыре цвета, но иные, чем различаем мы. Один цвет для пчел — это красный, желтый и зеленый; в этом участке спектра пчелы, выражаясь нашими понятиями, дальтоники. Другой цвет — сине-зеленый, третий — сине-фиолетовый, а четвертый — недоступный нам ультрафиолетовый!
Удивительная способность многих членистоногих различать поляризованный свет и воспринимать плоскость поляризации хорошо показана на насекомых: в глазу мух есть структуры, примерно так же действующие, как в полярископе Николевы призмы! Выяснилось, что глаз насекомого легче различает двигающиеся предметы, чем неподвижные, что запоминание пчелой летка ее улья связано с видом не только улья и самого летка, но и с восприятием всего ландшафта. Если на земле передвинуть улей на 2 м, возвращающиеся в него пчелы испытывают замешательство, стремясь туда, где леток был до перестановки улья. Но если держать улей на плоту на озере и переплыть на километр, пчелы легко находят свой улей, так как на фоне водной глади плот с ульем — единственный ориентир.
Не так давно выяснили, что пчелы могут различать и форму предметов, причем легче они различают фигуры с изрезанными, как у цветка или снежинок, краями (сходные с венчиками цветов), а труднее, но все же различают геометрические фигуры, на распознавание которых пчел можно выдрессировать.
Реакции насекомых на зрительные раздражения очень быстрые — ориентируясь по виду летящего насекомого, стрекоза меняет направление полета и схватывает добычу на лету.
Еще не все выяснено в области изучения зрения насекомых, но и то, что известно, поразительно.
Зрение не единственный путь получения информации насекомыми.
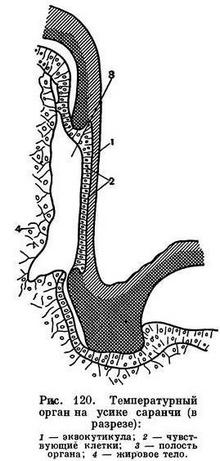
Все насекомые очень чувствительны к изменениям температуры, влияние которой на активность, длительность развития и длительность жизни насекомых исключительно велико. Каким образом насекомые, одетые в плотный панцирь, быстро воспринимают изменения температуры? Оказалось, что у насекомых есть особые теплочувствительные участки на поверхности тела, например у перелетной саранчи у основания усиков есть места с особо тонкой кутикулой, подстилаемой чувствительными клетками (рис. 120). У многих такими органами оказываются особые короткие толстостенные волоски. Такие волоски на лапке таракана воспринимают изменение температуры поверхности, по которой он ходит, в 1° — точность, труднодоступная нашему осязанию.
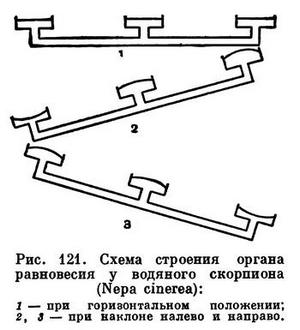
Живущие в воде насекомые хорошо реагируют на изменение давления, по величине которого они получают информацию о той глубине, на которой находятся, а также определяют положение тела. Например, у клопа, называемого водяным скорпионом (Nepa cinerea, табл. 38, 4), дыхальца на 3 сегментах ведут в камеру, выстланную очень тонкой кутикулой (мембраной), под которой лежат чувствительные клетки. В горизонтальном положении тела в воде мембраны на всех сегментах испытывают одинаковое давление, а при изменении положения на одни мембраны давление усиливается, а на другие ослабляется. Эта разница (равная всего 0,00015 am!) уже улавливается водным клопом!
Очень часто у насекомых бывают развиты органы слуха. Следует только оговориться, что слух у насекомых — это способность к восприятию не только звуковых колебаний, воспринимаемых нами, но и любых колебаний среды.
Насекомые — единственные беспозвоночные, для которых точно доказана способность различать звуки. Классическое доказательство — опыты со сверчками, когда в одном помещении перед микрофоном сажали стрекочущего самца, а в другом — самку перед телефоном. Когда включали микрофон, самка устремлялась к телефону. Не надо только думать, что органы слуха у насекомых похожи на наше ухо. Наше ухо воспринимает изменение давления воздуха, вызываемое источником звукового колебания, а у насекомых чаще воспринимается движение воздуха.
Наиболее соответствуют нашему уху тимпанальные органы,
состоящие из тонких перепонок, натянутых на склеротизованные кольца (вроде нашей барабанной перепонки), под которыми находятся пузыревидно расширенные участки трахей с подходящими к ним нервами. Такие органы есть у кузнечиков и сверчков на голенях передних ног, у саранчовых на боках первого брюшного сегмента (рис. 158), у совок на боках заднегруди, у огневок на первом брюшном сегменте, у цикад на втором и т. д.
Проще построенные хордотональные органы — натянутые эластичные волокна, к которым подходят нервные окончания, — видимо, есть у всех насекомых. Интересно, что по опытам, проведенным над несколькими видами бабочек совок, оказалось, что они реагируют на многие звуковые колебания (бабочки, улавливая звук, ускоряют полет и меняют его направление или «притворяются мертвыми»). Частота колебаний, на которую они реагируют, от 15000 до 175000 в секунду, причем сильнее всего реакция на колебания порядка 30000—80000, т. е. такого порядка, как частота ультразвуков, испускаемых летучими мышами. Известно теперь, что ориентировка летучих мышей при полете происходит по принципу радара: они испускают ультразвуки и улавливают их отражение от летящих насекомых. И у ночных бабочек выработалась защитная реакция — изменение полета — в ответ на эти «радарные лучи» их врагов. А некоторые ночные бабочки и сами издают ультразвуки, помогающие распознавать друг друга.
У насекомых органами слуха служат и специальные волоски, располагающиеся на всей поверхности тела или на отдельных участках. Точными опытами установлено, что, хотя у сверчка основные слуховые органы — тимпанальные органы находятся на голенях передних ног, ослабленный слух сохраняется у него и после их удаления. Волоски, расположенные на церках прямокрылых, улавливают звуки частотой 50—400 колебаний в секунду, а при синхронном колебании — до 800.
Подвижно причлененные волоски — это и органы для восприятия направления ветра, тока воздуха. У основания усиков крылатых насекомых есть сложно устроенные органы чувств — джонстоновы органы, с помощью которых насекомые контролируют скорость и направление полета.
Те группы насекомых, у которых наиболее хорошо развиты слух и органы слуха, способны и издавать звуки. Это относится к прямокрылым, у которых звуки издаются путем трения переднего края заднего крыла о нижнюю поверхность переднего крыла или о внутренний край бедер задних ног (у саранчовых) либо путем трения особых участков передних крыльев друг о друга (у сверчков, кузнечиков, рис. 149), к цикадам, у которых вибрирует особая мембрана, и т. д.
Явственно выражены сходные по строению «стридуляционные» органы у многих насекомых, например у личинок жуков (жуков-оленей, навозников, пассалид — рис. 248), которые живут в небольших объемах пищи. Их звуков мы не слышим, но они явно предупреждают друг друга о взаимно опасном сближении! До сих пор никто не изучал издаваемые ими не слышимые нами звуки. Следует учесть, что часто степень слышимости разных звуков неодинакова. Довольно много людей в общем с нормальным слухом, которые не слышат стрекота кузнечиков; правда, и кузнечики, вероятно, не слышат орудийных выстрелов, так как их стрекот при стрельбе не прекращается.
При восприятии звуков волосками трудно провести грань между слухом и осязанием. Многие насекомые воспринимают колебания той поверхности, на которой сидят. Полевой сверчок при частоте колебаний около 1500 в секунду воспринимает их даже при амплитуде в 0,1 миллимикрона (миллимикрон — одна миллионная миллиметра!). А садовая жужелица (Carabus hortensis) воспринимает колебания частоты 200—400 в секунду при амплитуде начиная с 5500 миллимикрон! Органы осязания насекомых — волоски, особенно чувствительные волоски на усиках и церках.
Исключительно сильно развиты у насекомых органы химического чувства. В большинстве случаев они разбросаны по всему телу (тонкие полые волоски, в полость которых заходят окончания чувствующих клеток), но основная их масса сосредоточена на усиках и щупиках. Чувствительность насекомых к запахам много выше, чем наша, например пчелы обнаруживают метилгептанон при концентрации в 40 раз меньшей, чем та, при которой его начинаем чувствовать мы.
Живущие в земле проволочники имеют обоняние в 10 раз более острое, чем наше, даже в отношении тех веществ, запах которых мы легко обнаруживаем. Именно по запаху обнаруживает и находит пищу большинство насекомых. Меченые самцы бабочки Actias selene прилетали на запах самки за 11 км, а самцы непарного шелкопряда — за 3,8 км.
Растворенные вещества могут восприниматься насекомыми не только ротовыми частями, но и лапками. Именно лапками передних ног мухи и бабочки могут пробовать, сладок ли раствор. При этом оказывается, что бабочки «ногами» чувствуют концентрации сахара в воде, в 2000 раз меньшие, чем те, начиная с которых мы распознаем сладковатый привкус! Выяснено, что насекомые могут различать сладкое, соленое, горькое и кислое. У большого водолюба, например, органы чувств на нижнечелюстных щупиках различают сладкий, горький и соленый вкус, а для распознавания кислого служат нервные окончания на вершине нижнегубных. Многие живущие в почве насекомые ориентируются по концентрации растворенных в почвенной влаге веществ, а недавно было показано, что они воспринимают и ничтожные изменения концентрации углекислоты: выделение углекислоты корнями растений привлекает личинок хрущей, проволочников и других живущих в почве вредителей.
На усиках находятся и удивительные по чувствительности органы восприятия влажности воздуха. Это небольшие бугорки и ямки (сенсиллы), которые позволяют, например, личинкам щелкунов (проволочникам) различать 0,5% относительной влажности воздуха (100% и 99,5%). Такие органы имеются и на щупиках.
Неясно, каким образом некоторые насекомые, например термиты, могут воспринимать магнитное поле, но индийские энтомологи установили, что самки термитов, находящиеся в подземных гнездах, располагаются в гнезде так, что ось тела проходит по магнитному меридиану.
Насекомые улавливают и воздействие электростатического поля, что также пока не может быть объяснено. А. Ф. Лебедев полагал, что основная функция покрова волосков и выростов на поверхности тела насекомого — отведение электрического заряда.
Приведенный краткий обзор показывает, что насекомые имеют очень развитую систему органов чувств — анализаторов. Поведение насекомых носит ярко выраженный рефлекторный характер: на раздражение, воспринимаемое тем или иным органом чувств (или их комплексом), насекомое реагирует теми или иными движениями, из совокупности которых формируется поведение насекомых. Иногда поведение имеет характер ярко выраженных реакций на раздражение — характер таксисов. Так, например, живущие в почве личинки насекомых стремятся с поверхности уйти вглубь (положительный геотаксис); активные днем насекомые, попав в темное помещение, стремятся к окну, к свету (положительный фототаксис); многие скрытоживущие насекомые, например чешуйницы, уходят от света (отрицательный фототаксис) и т. п. Часто в зависимости от условий, от физиологического состояния и т. д. меняется реакция на раздражитель. Чернотелки пимелии на севере ареала в нашем полушарии дневные, а на юге ночные. Чернотелки блапсы при более низкой температуре проявляют положительный фототаксис, а при более высокой — отрицательный. Мухи формии (Phormia) привлекаются альдегидом изовалериановой кислоты при концентрации его менее 1:50000, а при более высокой — отпугиваются им.
Очень часто определенный раздражитель является сигналом для начала той или иной активности насекомого. Например, перед заходом солнца снижение освещенности до определенной степени интенсивности является тем сигналом, по которому начинается лёт многих хрущей. В Средней Азии, где в мае часты безоблачные дни, можно с точностью до минуты предсказать, когда начнется лёт июньского хруща.
Реакции типа тропизмов у насекомых выражены недостаточно четко; сильнее выражены так называемые кинезисы. Как, например, происходит выбор насекомыми места с определенной температурой? Обычно насекомое не просто переходит, допустим, из более жаркого в более прохладное место. Находясь там, где температура выше оптимальной, насекомое быстро и беспорядочно бегает, пока не наткнется на место с более благоприятной температурой, где беспорядочные движения становятся замедленными. Если случайно насекомое выходит за пределы такой благоприятной зоны, оно снова начинает беспокойно передвигаться, пока не попадет в благоприятную зону, где постепенно замедляет движение и наконец, попав в наиболее благоприятную обстановку, останавливается. Итог — «выбрано» наиболее благоприятное место, но не путем прямого к нему устремления, а методом многочисленных «проб и ошибок».
Часто действия насекомого, которые кажутся простыми, представляют сложную цепь закономерно меняющихся друг за другом действий. Например, при движении жука-короеда к пригодному для откладки яиц дереву должно смениться в совершенно определенной последовательности не менее 7 реакций на различные раздражители (температура, влажность, цвет, запах и др.). Не только внешние, но и внутренние причины — состояние насекомого — определяют поведение. Самка жука, сегодня летающая, завтра, после оплодотворения, в точно таких же условиях среды не летает. Рабочая пчела, которая вчера не оставляла улей, несмотря на летную погоду, сегодня начинает летать за взятком.
Поведение насекомых носит характер в основном наследственно закрепленных инстинктов, представляющих определенную очень длинную и сложную цепь безусловных рефлексов. Инстинктивная деятельность многих насекомых (жуков-навозников, пчел, термитов) очень сложная, их поведение соответствует обычным условиям их жизни и производит впечатление «разумности». Однако такое наследственно закрепившееся поведение при изменении условий становится часто нецелесообразным и насекомое или его потомство погибает. Много примеров этому привел замечательный наблюдатель природы французский энтомолог Ж. А. Фабр, очерки которого («Жизнь насекомых») следует прочесть каждому, кто интересуется насекомыми.
Однако после работ Фабра долгое время недооценивалась роль индивидуального опыта, возможность выработки условных рефлексов у насекомых. При работах с пчелами выяснилась возможность выработки у них условных связей, «обучения» пчел. Оказалось возможным при дрессировке пчел научить их распознавать цвета и некоторые геометрические фигуры, брать пищу из сосудов определенной формы и цвета.
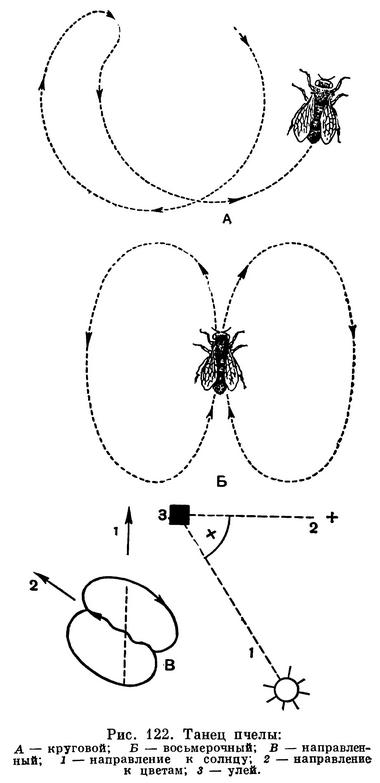
Кроме того, оказалось, что пчелы могут, общаясь, передавать друг другу индивидуальный опыт. Если научить пчелу узнавать чашки с сахарным сиропом определенной формы и цвета, а затем поместить эти чашки в районе облета пчелы, пчела, прилетев в улей, совершая движения определенным образом, указывает другим пчелам своего улья направление, по которому нужно лететь за пищей (рис. 122).
Некоторые исследователи, например у нас П. И. Мариковский, изучая поведение муравьев, нашли у них целый ряд жестов, которые служат сигналами, побуждающими других особей к определенному поведению («дай пищи», «опасность» и т. п.).
За последнее десятилетие были проведены исследования, показавшие, что у многих насекомых, особенно у общественных (муравьи, термиты и др.), выделяются биологически активные вещества, известные под общим названием «феромоны», которые влияют на развитие других особей и определяют их поведение. Так, например, насекомые могут оставлять определенные следы, по которым двигаются другие особи того же вида. Выделения матки пчелы в небольшом улье тормозят поведение рабочих особей, направленное на воспитание других маток, и т. п.
В данном случае речь идет об определенных химических соединениях, выделяемых одной особью и влияющих на поведение других. Такая химическая регуляция поведения представляет одну из очень интересных и еще малоисследованных особенностей поведения насекомых.
Очень интересна зависимость поведения от степени скученности насекомых данного вида. Поведение же в свою очередь, как оказалось, определяет и многие черты строения насекомого, что было впервые установлено для многих саранчовых Б. П. Уваровым. За последние 10 лет выяснилось, что влияние друг на друга особей одного вида у многих гусениц тоже приводит к различию и их поведения и их строения (как это, например, хорошо заметно на гусеницах ильмового ногохвоста, окраска которых при одиночном и скученном образе жизни различна). В этих случаях имеет значение уже не выделение каких-либо веществ, а воздействие особей друг на друга через нервную систему.
Особенно сложное поведение наблюдается у так называемых общественных насекомых, т. е. у тех, которые всегда живут колониями и иначе существовать не могут. В сущности, колония общественных насекомых — это одна семья, потомство одной самки (у термитов, пчел, муравьев). Для общественных насекомых характерно разделение функций, связанное с различными физиологическими особенностями и различиями в строении отдельных групп особей. В этом коренное отличие общественных насекомых от человеческого общества. В колониях общественных насекомых есть способные к размножению особи — самцы и самки (самок часто называют «матками» или «царицами») — и особи с недоразвитыми половыми железами, неспособные к размножению, но способные заготовлять пищу, выращивать личинок и т. д. Колония общественных насекомых существует именно благодаря такому разделению функций между различными группами особей. Естественно, что слаженная целостная жизнь колонии может обеспечиваться только при определенным образом координированном поведении всех входящих в нее особей. (Подробнее этот вопрос разобран в главе о перепончатокрылых)
Почти все насекомые раздельнополы. Только немногие, например обитающая в гнездах термитов муха Termitoxenia, гермафродиты. В некоторых местах в Калифорнии гермафродитом оказался и австралийский желобчатый червец: у него часть половых клеток превращается в сперматозоиды, часть в яйца, и самооплодотворение происходит внутри особи, имеющей вид самки; иногда часть яиц бывает не оплодотворена — из них развиваются самцы, утратившие биологическое значение. Но в общем для насекомых характерна раздельнополость.
У многих насекомых хорошо выражен половой диморфизм, т. е. самцы и самки сильно отличаются друг от друга. Иногда это признаки, связанные у самок с приспособлением к откладке яиц. Так, очень длинные яйцеклады, отсутствующие, конечно, у самцов, есть у самок наездников (рис. 364), рогохвостов (табл. 50, 5), кузнечиков (табл. 29) и других насекомых. У многих хрущей самки, зарывающиеся в землю для откладки яиц, имеют голени более расширенные, чем самцы. Таких примеров много. Нередко при большой массе откладываемых яиц самки бывают более грузными, чем самцы, и хуже летают, что в итоге приводит к тому, что у самок редуцируются крылья, и тогда самки нередко деградируют. Так, например, у бабочек-мешечниц (Psychidae) и у наших жуков-светляков (Lampyris noctiluca) самки личинкообразные, без всяких зачатков крыльев, у ложнощитовок (Lecaniinae) тело неподвижной самки утрачивает сходство с насекомыми вообще — кажется выступом коры (рис. 215), а у щитовок (рис. 214)— утрачивает все конечности. Самцы же у перечисленных насекомых имеют облик нормальных насекомых. Редко бывают обратные соотношения (например, у инжирного опылителя — рис. 363, 7,8), когда самец бескрылый, а самки крылаты.
Иногда различия в строении самцов и самок не имеют такого явного приспособительного значения. Так, у бабочек-голубянок (Lycaena) голубые крылья имеют только самцы, а у самок крылья коричневые, незаметные.
Иногда самцы и самки различаются по строению ротовых органов. Так, у самки жука-оленя жвалы развиты нормально, а у самца гипертрофированно изменены, превратясь только в «турнирное оружие», как и у настоящих оленей. У самца жука-носорога на голове рогообразный вырост, которого нет у самки, и т. п.
Нередко у насекомых наблюдается полиморфизм, когда, например, у общественных насекомых (пчелы, термиты) разные особи выполняют разные функции, обеспечивающие сохранение вида Так, у пчел есть способные к размножению самки (матки) и рабочие особи — самки с недоразвитыми половыми железами, обеспечивающие снабжение пищей всей колонии. По всему облику эти самки сильно разнятся друг от друга. Еще более разительны различия у муравьев, у которых самки крылаты, а рабочие совершенно бескрылы; у термитов много форм: рабочие, солдаты и др.
Резко выражен полиморфизм у тлей, у которых самки бывают бескрылыми или крылатыми, партеногенетическими или оплодотворяемыми в зависимости от условий освещения, питания, температуры.
Обычным способом размножения у насекомых является обычное половое размножение, но нередко наблюдается и партеногенез, чаще встречающийся у насекомых, самки которых бескрылы. Партеногенез обычен у тлей, щитовок, долгоносиков-скосарей, бескрылых эмбий, палочников и др.
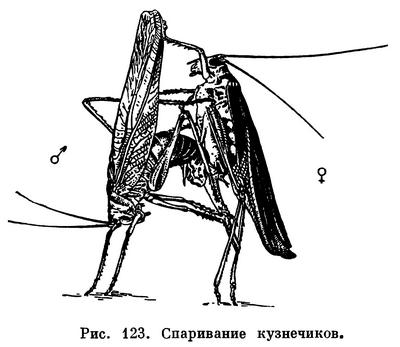
Оплодотворение у низших насекомых (чешуйницы) наружно-внутреннее: самец откладывает на землю сперматофор, а самка подбирает его с земли при помощи половых придатков. Близкий способ оплодотворения у низших групп крылатых насекомых как с неполным (таракановые, богомолы, кузнечики, сверчки), так и с полным (верблюдки, вислокрылки, сетчатокрылые) превращением. У них самка подхватывает находящийся ниже ее брюшка сперматофор своими половыми придатками в момент его откладки самцом (рис. 123). Этим достигается сокращение срока пребывания сперматофора во внешней среде. У более высокоорганизованных групп насекомых (бабочки, жуки, двукрылые) самец вводит сперматофор в половое отверстие самки с помощью совокупительного органа, благодаря чему семенная жидкость ни мгновения не остается во внешней среде, т. е. оплодотворение внутреннее. А так как при этом защитная роль оболочки сперматофора становится излишней, при внутреннем оплодотворении часто в половое отверстие самки вводится не сперматофор, а непосредственно семенная жидкость.
Большинство насекомых откладывает яйца. У разных видов форма яиц очень разнообразна (табл. 20). У насекомых яйца покрыты скорлупой, защищающей их от неблагоприятных воздействий, причем скорлупа образуется до оплодотворения яйца. Сперматозоид проникает в яйцо через особое отверстие в скорлупе, находящееся против ядра яйцеклетки, — через так называемое микропиле. При вылуплении личинка обычно прогрызает оболочку яйца.
В некоторых случаях можно заставить яйца развиваться и без оплодотворения у тех видов, у которых нормально требуется оплодотворение. Такой «искусственный партеногенез» был впервые получен московским зоологом А. А. Тихомировым на яйцах тутового шелкопряда в конце прошлого века. А недавно тоже в Москве Б. Л. Астауров добился развития шелкопряда только за счет ядра сперматозоида, убив в яйце рентгеновскими лучами ядро, полученное от материнского организма. Такой «андрогенез» представляет огромный теоретический интерес для определения роли ядра в наследственности. У шелкопряда роль ядра оказалась решающей, полученный андрогенетический шелкопряд имел все признаки отца и никаких признаков матери, хотя материнской плазмы в яйцеклетке много больше, чем отцовской в сперматозоиде.
Для представителей всех групп водных беспозвоночных, перешедших к жизни на суше (в почве), а также для всех групп низших наземных членистоногих, тоже тесно связанных с почвой, характерно то, что у них из яйца выходит молодой организм, очень похожий на взрослый. Это относится и к связанным с почвой низшим насекомым — щетинохвосткам, у которых рост молодого организма не сопровождается существенными изменениями строения. Вероятно, так развивались и предки насекомых, которые жили в таких же условиях. Такой тип развития условно можно назвать прямым развитием. Такое развитие возможно, если в течение всего периода после выхода из яйца животные ведут примерно однотипный образ жизни, обитая в одной и той же среде.
Правда, например, даже у чешуйниц наблюдается, что взрослые формы чаще покидают почву и другие укрытия, чем молодь. Выходы взрослой стадии из почвы на ее поверхность у предков современных насекомых были очень кратковременными, но биологическое значение их было велико — на открытой поверхности облегчается возможность встречи полов и легче происходит расселение. Взрослые формы, имеющие относительно меньшую поверхность тела, чем мелкие молодые, всегда легче выносят временный дефицит влаги (даже у самых влаголюбивых видов). От временных переползаний на поверхности почвы взрослая стадия предков насекомых перешла к более длительным выходам из почвы, к расселению сперва путем такого же переползания, как по ходам в почве, а затем прыжками (махилисы). Более подверженные иссушению спинные участки тела стали более плотными, над боками образовались боковые защитные выросты тергитов — «паранотальные выросты», характерные для многих наземных членистоногих; такие выросты могли дать начало зачаткам крыльев, с помощью которых взрослые насекомые могли сперва перейти к планирующему полету, а затем и к активному. На рисунке 85 приведена гипотетическая схема происхождения насекомых от червеобразных предков.
При переходе взрослой стадии насекомых к жизни в воздушной среде возможны были два пути эволюции.
В одном случае и личинки стали переходить к такому же образу жизни, как взрослые. Но жизнь на открытой поверхности требует более сложного строения. Поэтому у тех насекомых, у которых личинки ведут такой же образ жизни, как взрослые, но уже в открытой воздушной среде, они из яйца выходят на более высокой ступени развития, чем выходили их жившие в укрытиях предки. Это путь, по которому шло развитие насекомых с неполным превращением.
Другой путь эволюции был связан с тем, что личинки продолжали вести скрытый образ жизни в почве и в других укрытиях, приспосабливаясь к нему всё совершеннее, а взрослые насекомые вели открытый образ жизни, активно летая и отыскивая подходящие места для развития потомства. Приспособление ранних стадий к одному образу жизни, а взрослых к другому сделало невозможным переход из личиночного состояния во взрослое путем обычной линьки. Потребовалась стадия куколки — стадия перестройки. Это путь развития насекомых с полным превращением. У насекомых с полным превращением личинки часто имеют очень упрощенное строение; например, у личинок мух нет ни ног, ни головы, весь их облик скорее напоминает червя. Такая простота строения — своеобразное приспособление к развитию в запасах пищи. Более примитивные личинки насекомых с полным превращением похожи по внешнему виду на первичнобескрылых насекомых — у них 3 пары ног, а на конце брюшка бывают церки. Куколка насекомых — непитающаяся стадия, по общему облику более напоминающая взрослое насекомое, чем личинку. На стадии куколки происходит разрушение специфических личиночных органов, осуществляемое в процессе разрушения личиночных тканей (гистолиз), и образование из скоплений особых клеток (имагинальные диски) тканей и органов взрослого насекомого (гистогенез). Чем больше отличается строение личинки от строения взрослого насекомого, тем сильнее процессы гистолиза в теле куколки. Особенно сложные перестройки происходят у таких насекомых, как мухи, у которых в первые дни после окукливания основная масса внутренних органов куколки представляет жидкую кашицу.
Основные типы куколок — свободная (рис. 261, 266, 317) и покрытая (рис. 327, 412). Свободная куколка характеризуется тем, что у нее все придатки (ноги, усики, ротовые части, крылья) не только хорошо различимы, но и легко могут быть отделены иголочкой от поверхности тела, даже если они плотно прижаты. У покрытых куколок придатки, хотя и заметны, но сливаются с общей поверхностью тела и не могут быть отведены иголкой от тела без повреждения насекомого. Особый тип куколки — бочонковидный. Куколки этого типа характерны для высших мух, у которых роль оболочки куколки принимает последняя личиночная шкурка, которая перед окукливанием личинки сильно раздувается и затвердевает (рис. 423, 5).
Нередко стадия куколки называется «покоящейся». Такое определение неверно, так как, во-первых, в теле куколки идут интенсивные процессы перестройки организма, а во-вторых, у многих насекомых куколки производят энергичные движения. Только бочонковидные куколки совершенно неподвижны, остальные же способны двигать хотя бы брюшком. Развивающиеся в воде покрытые куколки ручейников и комаров превосходно и очень активно плавают. Куколки бабочки древесницы въедливой активно проползают из глубины хода, проложенного гусеницей в древесине, к тому месту, где находится отверстие, через которое выходит бабочка. У верблюдок куколка бегает не менее проворно, чем личинка, а у некоторых мошек, как показал английский ученый Хинтон, куколка даже способна прогрызать шелковый чехлик, который перед окукливанием сплетает личинка.
У некоторых насекомых, ведущих очень сложный образ жизни в стадии личинки, наблюдается так называемый гиперметаморфоз. В тех случаях, когда личинки разного возраста имеют резко различающееся строение, переход одной формы лпчинки в другую сопровождается прохождением стадии перестройки, обычно неподвижной («ложнокуколка»). Таким образом, при гиперметаморфозе имеются две стадии перестройки организма — ложнокуколка и куколка.
Рост насекомых, как и всех членистоногих, сопровождается линьками. Из всех открыточелюстных насекомых только у щетинохвосток линьки продолжаются и во взрослом состоянии, да у поденок есть одна линька уже окрылившихся насекомых. Остальные же насекомые во взрослом состоянии не линяют. В личиночном возрасте обычно бывает 4—5 линек, но иногда их не бывает вовсе (некоторые паразитические перепончатокрылые) или их более 20 (личинки некоторых щелкунов, цикад).
Как и у всех животных с непостоянной температурой тела, скорость развития у насекомых зависит от температуры. В пределах примерно от 6 до 36° С при повышении температуры на каждые 10° скорость развития удваивается. Опытным путем было установлено, что для каждого вида насекомых можно установить то количество «тепла», которое требуется для развития. Наименьшая температура, при которой уже происходит развитие насекомого, называется температурным порогом развития (у большинства насекомых это 6—10°). Среднесуточная температура выше этого «порога» называется эффективной. Для многих видов насекомых умеренной полосы определена та сумма среднесуточных эффективных температур, которая требуется для развития. Как правило, в пределах области распространения того или иного вида в зависимости от местных условий года меняется конкретное число дней, требуемых для развития данного насекомого, но сумма эффективных температур остается величиной более или менее постоянной. Поэтому в зависимости от длины теплого (вегетационного) периода и от среднесуточных температур (обе величины закономерно изменяются при переходе из одних широт в другие) многие виды имеют разное число поколений. Например, луговой мотылек (Loxostege sticticalis) — один из наших хорошо изученных широко распространенных видов вредителей — на севере области распространения (в нечерноземной зоне) дает одно поколение в году, в зоне лесостепи — два, на юге степной зоны — три, а в Закавказье даже четыре. Но многие виды дают меньше поколений в году, чем можно определить по сумме эффективных температур. Бывает, что при благоприятных температуре и влажности развитие насекомого вдруг останавливается. Такая остановка развития имеет приспособительное значение для насекомого. Например, на Украине гусеницы люцерновой совки (Chloridea dipsacea), развивающиеся из яиц, отложенных в начале лета, окукливаются и через 2 недели из куколок вылетают бабочки. Гусеницы, вышедшие из яиц, отложенных этими бабочками, развиваются примерно столько же времени, сколько развивались и гусеницы первого поколения, тоже окукливаются, но бабочки из этих куколок через 2 недели не выходят. Куколки второго поколения, несмотря на благоприятные условия, останавливаются в развитии и зимуют. Такая остановка развития носит название глубокого покоя или диапаузы.
Диапауза имеет явно приспособительный характер. Если бы у люцерновой совки из куколок второго поколения бабочки вылетали в тот же год, из отложенных ими яиц вышли бы гусеницы, которые не успели бы закончить свое развитие.
Уход в диапаузу связан с существенными изменениями обмена веществ насекомых — с накоплением избытка жира и т. п. «Сигналом», определяющим уход в диапаузу люцерновой совки, оказывается длина светового дня. Если гусениц держать при длинном световом дне, они дают быстро развивающихся куколок, а при сокращенном — диапаузирующих. Температура, влажность — факторы очень изменчивые, зависящие от погоды. А вот длина дня для каждой зоны — величина строго определенная для каждого дня в году. Поэтому естественно, что приспособление к уходу в зимовку в подготовленном состоянии определяется таким постоянно действующим фактором. Ну, а выход из диапаузы в разбираемом случае связан с теми изменениями в организме, которые определяются действием зимних температур. Только после морозного периода диапаузирующая куколка люцерновой совки снова способна к развитию. Так достигается то, что превращение куколки в бабочку осуществляется весной, когда ее потомству обеспечена обильная пища.
В состояние диапаузы разные насекомые могут впадать на разных стадиях развития. У саранчовых известна зародышевая диапауза (в стадии яйца), у желудевого долгоносика диапаузируют личинки, у люцерновой совки — куколки, у колорадского жука — взрослая стадия.
Таким образом, развитие насекомых регулируется теми изменениями их состояния, которые вызываются условиями среды. Но, кроме того, развитие насекомых управляется и внутренними факторами. За последние 10 лет установлен ряд гормонов, регулирующих развитие насекомых. Эти гормоны выделяются нервной системой (мозг, подглоточный ганглий) и специальными железами внутренней секреции — придаточными телами мозга. Для некоторых гормонов, например для гормона линьки (экдизона), уже известно химическое строение, тогда как другие гормоны, например «ювенильный гормон» (неотин), выделяемый придаточными телами, тормозящий созревание, известен только по физиологическому действию.
О влиянии на развитие «феромонов» (называемых также «телергонами») — веществ, выделяемых одними особями и влияющих на развитие других представителей того же вида, упоминалось ранее.
Насекомые очень быстро растут и очень плодовиты. Естественно, что и влияние полчищ насекомых на природу огромно. Многие насекомые — важные почвообразователи, разрушители растительного опада. Известно, что в основном насекомыми разлагается, например, опад в зоне хвойных лесов и что там, где нет насекомых, накапливаются волокнистые торфообразные неплодородные слои. В тропиках основными почвообразователями являются термиты, перерабатывающие в лесах всю отмирающую древесину. За свою жизнь насекомые перерабатывают количество пищи, в десятки раз превышающее их вес. Насекомые быстро уничтожают накапливающиеся на поверхности почвы остатки растений, трупы и экскременты животных.
Некоторые насекомые, питающиеся разлагающимися растительными и животными остатками, неразборчивы в пище, а некоторые очень специализированы; есть, например, жуки-навозники, развивающиеся только в заячьем помете. Осуществляемую насекомыми очистку почвы от скоплений остатков организмов трудно переоценить. Еще К. Линней подсчитывал, что в тропиках потомство трех мух съедает труп лошади быстрее, чем лев! В известных опытах почвоведа П. А. Костычева листья, подвергавшиеся разложению микроорганизмами, 3 года сохраняли свое строение, а когда на них отложил свои яйца грибной комарик, за 3 дня превратились в перегнойную труху!
Среди насекомых огромное количество видов приспособилось к питанию высшими (цветковыми) растениями. По ископаемым остаткам удалось установить, что наиболее прогрессивные группы насекомых развивались параллельно с цветковыми растениями. Цветковые растения — основной источник и пищи и влаги для большинства насекомых. В то же время большинство цветковых растений (около 80%) в свою очередь непосредственно зависит от насекомых. Оказывается, что привлекающие наш взор пестрота и многообразие цветов обязаны тому, что растения приспособились к опылению насекомыми. Посещающие цветы насекомые обеспечивают перенос пыльцы с пыльников одних цветков на рыльца других, обеспечивая перекрестное опыление. Особенно велика роль как опылителей растений представителей трех высших отрядов насекомых с полным превращением — перепончатокрылых, двукрылых и бабочек — тех групп, в отношении которых и по геологическим наслоениям прослеживается связь с цветковыми растениями.
Многие растения имеют такое строение цветка, какое требует обязательного участия в опылении какого-либо определенного вида или группы видов насекомых.
Насекомые посещают цветки для сбора пыльцы (некоторые жуки, многие перепончатокрылые) или ради сладкого выделения — нектара (мухи, бабочки, многие перепончатокрылые). Иногда насекомые используют цветки как убежище или забираются в цветки и соцветия для пристраивания там своего потомства. И все типы посещения цветков насекомыми, оказывается, используются растениями для обеспечения перекрестного оплодотворения.
Часто цветы бывают ярко окрашенные, но без особого запаха. Они привлекают насекомых, которые ориентируются в основном с помощью зрения. Цветки с мелкими венчиками, как правило, бывают собраны в соцветия, чтобы привлекать насекомых, которые их лучше в таком случае замечают. Иногда рядом с невзрачным цветком для привлечения насекомых развиваются яркие прицветные или кроющие листья, как у белокрыльника, иван-да-марьи. Для привлечения насекомых, летающих ночью или в сумерки, особое значение имеет запах; обычно открывающиеся в темноте цветки имеют белую окраску, даже для нас более заметную при недостатке освещения. Не всегда запах цветов бывает приятен для человека — цветы, привлекающие падальных мух, обладают трупным запахом и иногда имеют окраску венчиков мясного цвета (например, тропическая раффлезия). У многих цветков нектар находится вблизи поверхности и достать его нетрудно (бузина, липа, селезеночник). Такие цветки опыляются случайными посетителями, нередко мухами и даже мелкими жуками. Цветки с полускрытым нектаром, такие, как у различных крестоцветных, у которых нектарники находятся на дне короткой трубочки венчика или защищены волосками, как у розоцветных (яблоня, земляника), опыляются мухами и некоторыми перепончатокрылыми, имеющими короткий хоботок. Обычно такого типа цветки правильные (с радиально расположенными лепестками венчиков) и имеют белую, желтую или розовую окраску. Есть же такие цветки, и их много, у которых нектар спрятан глубоко в недрах сросшихся лепестков венчика. Такие цветки чаще бывают неправильными (двустороннесимметричными), и окраска таких цветков чаще бывает синих, пурпурных и фиолетовых тонов. Нектар таких цветков доступен не для любых насекомых, а только для определенных, имеющих длинный хоботок и нередко достаточно сильных, чтобы раздвинуть лепестки при доставании нектара. Такая форма цветка заставляет определенный вид насекомого, высасывающего нектар, занять на цветке определенное положение, которое обеспечит покрытие пыльцой тех частей тела крылатого гостя, которые войдут в контакт с рыльцем следующего посещаемого цветка.
Приспособления к переносу пыльцы бывают различными. Иногда пыльца просто легко прилипает к телу насекомого, посещающего цветок, как например у лилии, мака. В этих случаях почти любое насекомое, привлекаемое цветком и переходящее с него на другой, обеспечивает опыление. Но не так легко достать нектар из цветка, например, шалфея, доступ к нектару которого закрыт особыми пластинчатыми выростами оснований тычинок. Только сильное насекомое, например пчела, садясь на цветок, может раздвигать лепестки шалфея. Устремляясь к нектару, пчела головой нажимает на вырост тычинки, пыльник которой ударяет пчелу по спинке. При посадке на следующий цветок шалфея пчела именно тем местом, на которое попала пыльца, соприкасается с рыльцем и переносит таким образом на него пыльцу (рис. 124). Сходно приспособление к опылению у дельфиниума.
Обычно привлечение насекомых цветком вызывается пищевыми раздражителями, но не всегда. Например, некоторые орхидеи опыляются самцами шмелей. Было выяснено, что цветок таких орхидей напоминает очертания самки шмеля и привлекательность цветка связана с половым инстинктом насекомого.
Очень своеобразные отношения сложились между американским растением юккой и юкковой молью (Pronuba juccasella). Самка юкковой моли по вечерам посещает свисающие цветки юкки и собирает комок пыльцы с помощью специально видоизмененных челюстей, в которых переносит этот комок на другой цветок юкки. Своим яйцекладом она прокалывает завязь, откладывает в нее яйца, а после этого спускается по столбику и засовывает комок пыльцы в рыльце (рис. 125). Таким образом моль обеспечивает опыление юкки, а тем самым и развитие своего потомства — выходящие гусеницы питаются завязывающимися семенами, часть которых съедают. Не менее удивительна взаимная приспособленность фигового дерева и его опылителя — хальциды-бластофаги.
Нередко недостаток опылителей определяет неурожай многих растений. Это хорошо известно тем, кто имел дело с плодовыми деревьями — яблони, груши без достаточного количества пчел не дают плодов. Некоторые культурные растения зависят не от домашней пчелы, а от диких опылителей. Например, клевер опыляется в основном шмелями. Не всякая домашняя пчела обеспечивает опыление клевера. Хоботок нашей среднерусской пчелы недостаточно длинен, чтобы доставать нектар из глубоких нектарников цветков клевера; только длиннохоботковая кавказская раса пчелы может опылять клевер. Но, кроме того, пчел на клевер нужно дрессировать. Известно, что пчела, раз начав посещать цветки определенного вида растений, продолжает посещать преимущественно их весь период цветения. Подкормка пчел сиропом, изготовленным на цветках клевера, повышает посещаемость клевера пчелами. Иногда приходится дрессировать пчел и на посещение таких цветов, у которых нектар легкодоступен. Например, чтобы получить хороший урожай семян рапса, недостаточно расположить вблизи посева этой культуры пасеку; надо предварительно провести дрессировку пчел на рапс.
Есть много культурных растений, например люцерна, опыляющихся дикими пчелами, целиком заменить которых домашней пчелой нельзя.
Не могут опыляться пчелами и такие цветы, которые открываются в темное время суток. Такие цветы, особенно имеющие глубоко расположенные нектарники, как, например, душистый табак, опыляются ночными бабочками, преимущественно бражниками и совками.
Если питание нектаром и избыточной пыльцой растений насекомых-опылителей приносит пользу растениям, то поедание насекомыми других частей растений причиняет им в большинстве случаев явный ущерб.
Характер повреждений различными насекомыми тканей растений очень многообразен. Он зависит от того, какие органы растения повреждаются насекомым и где находится насекомое в момент повреждения, и от строения ротового аппарата насекомого. Для вредящих растениям насекомых характерны 2 типа ротового аппарата — грызущий и колюще-сосущий. Насекомые с грызущим ротовым аппаратом грубо откусывают участки тканей растения. Если грызущие насекомые питаются листьями растений, они либо «неправильно» объедают лист, съедая кусок за куском, как это делают крупные жуки, например свекловичный долгоносик на свекле, либо оставляют только толстые жилки («скелетируют» лист), как это делает тополевый листоед на тополе, либо прогрызают в листьях окошечки, как жуки-щитоноски и их личинки на лебеде, либо обгрызают края листовой пластинки, как жуки клубеньковые долгоносики на листьях гороха. Всех способов нанесения повреждений (рис. 126) не перечислить, но по характеру повреждений, которые бывают обычно очень типичны для каждого вредителя, питающегося на данном виде растения, определить вид насекомого бывает даже легче, чем по признакам строения самого вредителя. Те насекомые, которые питаются твердыми или мясистыми частями растений (стволы, стебли, плоды), чаще вгрызаются в ткани, прокладывая там ходы. Такие ходы тоже бывают настолько индивидуальны, что специалисту спутать, какому виду принадлежит ход, например, короеда, невозможно.

Большие систематические группы насекомых приспособились к развитию в лубе древесных пород (лубоеды), в древесине (многие усачи, златки, рогохвосты), в плодах (долгоносики-ринхиты), в желудях и орехах (долгоносики-плодожилы), в головках клеверов (долгоносики-семееды), в соломинах злаков (хлебные пилильщики), в толстых стеблях травянистых растений (горбатки, долгоносики-фрачники), в клубеньках на корнях бобовых (долгоносики-ситоны) и т. д.
Многие мелкие насекомые поселяются в листовых пластинках, проделывая в них ходы — мины, которые снаружи хорошо заметны и на зеленом фоне листа кажутся белыми. Есть целые семейства минирующих мух, минирующих молей и др., именно таким образом вредящих растениям.
Развитие внутри растительных тканей создает очень благоприятные условия для многих насекомых — внутри растения воздух насыщен водяным паром, как и в почве. Поэтому обитающие внутри растения личинки насекомых избавлены от самого неблагоприятного воздействия, которому могут подвергаться наземные насекомые, — им не угрожает высыхание. Поэтому, например, в пустынях, где воздух очень сух, большинство растительноядных насекомых либо обитает в почве, повреждая корни растений, либо развивается внутри растительных тканей.
Не надо думать, однако, что в сухих местностях нет открыто живущих вредных для растений насекомых, — их очень много и они в этих условиях особенно опасны. Теряя влагу, они стремятся восполнить ее запасы за счет сочных тканей растений, и потому именно в сухих местностях вредные насекомые наносят особо сильный ущерб растениям. Достаточно напомнить, что именно в сухих местностях распространены и сильно вредят различные саранчовые, съедающие в годы массовых размножений всю зелень, встречающуюся на их пути.
Те насекомые, которые высасывают соки из растений, не причиняют таких бросающихся в глаза нарушений повреждаемым органам, однако в результате их деятельности растения тоже очень страдают.
Если вредитель, например тля, поселяется на листьях, листья начинают сперва светлеть, затем желтеть, а потом и завядать. Нередко происходит скручивание листьев — это наиболее простая деформация листа, вызываемая сосущими насекомыми. При массовом поселении сосущих насекомых на корнях (например, кровяной тли на яблоне) или на стволах (например, запятовидной щитовки на саженцах яблони) растения могут засохнуть. Нередко поселение на растении сосущих, а также развитие в тканях растений и некоторых грызущих насекомых вызывает разрастание тканей растений, которое либо имеет вид неправильных наростов, либо напоминает какие-либо плоды.
Такие разрастания тканей растений под влиянием механических раздражений и химических воздействий, причиняемых насекомыми (или другими организмами), носят название галлов (рис. 127, табл. 58). Так, галлы, вызываемые орехотворками на листьях дуба, напоминают плод типа костянки: внутри одетого румяной кожуркой мягкого, похожего на яблоко шаровидного галла вокруг тела личинки даже образуется затвердение тканей листа, напоминающее косточку. Галлы, вызываемые хермесами на молодых побегах ели, несколько напоминают шишку, а на листьях береста, на которых поселяется берестовая тля, вокруг тли образуются орешкоподобные или стручкообразные галлы, которые растрескиваются, когда тля закончит свое развитие и должна вылетать. Образование галлов — результат длительной эволюции, приспособления друг к другу растения и насекомого. Растение «создает» благоприятные условия для существования насекомого, но как бы изолирует насекомое, защищая этим ткани, удаленные от места сосания, от его воздействия.

Важным моментом, определяющим галлообразование, является выделение насекомыми ростовых веществ типа ауксинов. Известно, что, применяя обработку растений гетероауксином, можно вызвать усиленный рост некоторых органов, например органов плодоношения у томатов, что применяется в нашей практике. Воздействие насекомых на формообразовательные процессы у растений гораздо тоньше и гораздо эффективнее — изучение галлов может дать много нового для понимания процессов развития растений.
Те насекомые, которые питаются культурными растениями, причиняют огромный вред народному хозяйству. Достаточно указать, что по подсчетам, сделанным 10 лет назад, только сельское хозяйство нашей страны несет ежегодно потери порядка 4,5 миллиарда рублей (в современном исчислении) от вредителей и болезней. Это без учета потерь от вредителей леса! Только от вредных насекомых (не считая потерь от болезней растений) потери сельского хозяйства в США составляют 4 миллиарда долларов в год. Эти поистине чудовищные цифры в большой мере объясняют то, что на изучение насекомых направлены усилия огромного числа специалистов, работающих во всех странах мира. Достаточно сказать, что каждые несколько минут из печати выходит научная работа по энтомологии — разделу теоретической и прикладной зоологии, который посвящен насекомым.
Но никогда не следует забывать, что, если бы провести точные подсчеты той пользы и того вреда, который приносится насекомыми, польза бы значительно перевесила. Достаточно вспомнить, что без насекомых не давали бы урожая многие важные культурные растения — плодовые, гречиха, крестоцветные, подсолнечник и др. и многие просто бы погибли. Если бы не было жуков-навозников, погибли бы пастбища.
Что это не досужее предположение, показывает такой интересный случай из истории сельского хозяйства Австралии. В Австралии некоторые пастбища стали погибать из-за того, что на поверхности почвы стали скапливаться неразлагающиеся и затрудняющие рост травы кучи коровьего навоза. Оказалось, что там не было навозников. Завоз навозников и их акклиматизация быстро привели к разложению скоплений навоза и к большому повышению урожайности начавших приходить в запустение пастбищ.
Когда мы говорим о насекомых-вредителях, мы имеем в виду не тот вред, который насекомые причиняют растениям, а тот урон, который они наносят нашей экономике. Например, когда в Австралии в массе размножились на пастбищах колючие кактусы, мешавшие пастьбе скота, туда были завезены насекомые, истребляющие кактусы, которые рассматривались как полезные насекомые. А когда были выведены бесколючковые формы кактусов, которые охотно поедаются скотом, виды насекомых, истребляющих кактусы, из полезных стали вредителями.
Из насекомых, дающих человеку непосредственно используемые им продукты, только медоносная пчела приносит пользу и растениям. Кстати сказать, польза от пчелы как от опылителя культурных растений во много раз превышает ту пользу, которую мы получаем, используя мед и еще более ценный продукт, широко применяемый в технике, — воск. Другие непосредственно полезные для нас насекомые повреждают те растения, на которых питаются. Их не так много. Это тутовый шелкопряд, гусеницы которого выделяют при постройке кокона шелк. Несмотря на замечательные достижения современной химии синтетических волокон, шелк оказывается непревзойденным еще материалом. Шелк по степени полимеризации и по зависящим от нее механическим качествам нити превосходит лучшие сорта капрона, нейлона и других синтетических волокон. Поэтому для наиболее ответственных, требующих высокой прочности изделий, — для парашютов, для герметических прокладок —натуральный шелк представляет пока лучший материал. Другой шелкопряд — китайский дубовый шелкопряд — дает очень прочную, но более грубую нить, из которой изготавливается чесуча.
Очень ценный продукт — шеллак дают обитающие на ряде древесных пород на юго-востоке Азии лаковые червецы. Воскоподобные вещества, выделяемые этими червецами, обладают исключительными изоляционными свойствами и находят широкое применение в электротехнике, особенно в радиотехнике.
Другие червецы дают краску: как источник ценной краски кармина известны обитающие на средиземноморском дубе червецы кермесы (Kermes), значение которых было особенно велико в древности, но снизилось, когда стали добывать кармин из живущих в Мексике на кактусе опунции кошенилей (Coccus cacti). Теперь с развитием химии красителей значение этой краски несколько уменьшилось.
Некоторое значение в медицине имеет кантаридин, содержащийся в жуках «шпанских мушках», обгрызающих листья ясеня. Кантаридин — активное вещество нарывного пластыря. Вот в сущности и все те непосредственно полезные для нас насекомые, которые дают используемые человеком вещества.
Медоносная пчела, тутовый и дубовый шелкопряды, лаковые червецы и кошениль — немногие примеры полуодомашненных насекомых, разводимых человеком ради доставляемых ими продуктов.
Среди остальных питающихся растениями видов насекомых мы сталкиваемся с более или менее безразличными для нас, которые питаются не используемыми человеком видами растений, и с вредителями, которые повреждают культурные растения, становясь нашими конкурентами.
Среди вредных для культурных растений насекомых можно выделить две биологические группы. Одна включает многоядных вредителей, которые малоразборчивы в выборе пищи и повреждают целый ряд культурных (полевых, огородных и др.) растений. Таковы, например, личинки щелкунов, большинство саранчовых, гусеницы лугового мотылька, озимой совки и многие другие. Другая группа — специализированные вредители, питающиеся одним или немногими видами культурных растений. Это, например, такие насекомые, как вредящий сахарной свекле свекловичный долгоносик, вредящая зерновым культурам шведская мушка, повреждающая виноградную лозу филлоксера и др.
Все вредители культурных растений перешли на них с дикорастущих. Как правило, многоядные вредители переходят на посевы культурных растений быстрее. Когда, например, распахиваются целинные земли, развивающиеся в почве и питающиеся корнями разных растений личинки щелкунов принимаются за питание корнями посеянного по нови растения. Переход специализированных вредителей с дикорастущих растений на родственные культуры иногда совершается очень медленно. Так, например, в Красноярском крае долгоносик льняной скрытнохоботник стал вредить льну только лет через сто после введения его там в культуру, а до этого существовал за счет дикорастущего желтого льна.
Мальвовая моль в Армении недавно перешла с дикорастущей мальвы на хлопчатник и стала опасным вредителем его коробочек, а в Средней Азии она хотя и встречается на мальве, но на хлопчатник не переходит. А, например, при широком введении в культуру кок-сагыза в нашей стране в 30-х годах с первых же лет на его посевах в массе встречалось несколько видов вредителей, перешедших с родственных кок-сагызу дикорастущих одуванчиков. Большинство вредителей культурных растений — представители местной фауны, ранее питавшиеся за счет местных дикорастущих растений.
Часто новые культурные растения, которые не имеют близкородственных дикорастущих видов среди местной флоры, оказываются почти неповреждаемыми вредителями. Например, эвкалипты на родине, в Австралии, страдают от многих вредителей, а в Европе и Азии практически не повреждаются вредителями. В таких случаях очень важно для защиты посевов не допустить вредителя из удаленных местностей на территорию, где вредителя нет. Это достигается путем применения карантинных мер, контроля за тем, чтобы с посадочным материалом и с разными грузами вредитель не последовал за растением. История знает случаи завоза очень опасных вредителей, которые в массе размножились и стали причинять большие опустошения там, где их ранее не было. Так, в Северную Америку был завезен из Европы стеблевой, или кукурузный, мотылек, ставший там бичом кукурузы. Еще ранее из Европы туда попала сильно вредящая пшенице гессенская мушка. Около 100 лет назад в Америку был завезен непарный шелкопряд, объедающий там леса и сады сильнее, чем на родине, в Европе. В Европу из Америки тоже около 100 лет назад завезли виноградную филлоксеру, разорившую виноградное хозяйство Франции, и широко распространившуюся кровяную тлю, губящую яблони у нас на юге.
В нашей стране нет многих опасных вредителей, таких, например, как «розовый червь» (гусеница) хлопчатника, уничтожающий хлопковые семена и волокна в коробочках (плодах).
Строгий карантинный контроль, облегчаемый у нас монополией внешней торговли, позволяет нашей стране не допускать многих опасных вредителей, в других странах причиняющих большой ущерб.
Следует иметь в виду, однако, что и активные перелеты насекомых-вредителей, и расширение транспортных связей делают задачи карантина все более сложными. И некоторые вредители, еще недавно у нас отсутствовавшие, проникают на нашу территорию. Примером может служить колорадский жук. У нас картофель не знал серьезных вредителей. Около 100 лет тому назад в Америке в Скалистых горах с дикого картофеля на культурный перешел колорадский жук, быстро приспособившийся к новому для себя виду растений и распространившийся по всей территории США до Атлантического океана уже к 1874 году. В начале XX века неоднократно его завозили случайно в Европу, но быстро уничтожали в местах появления. Во время первой мировой войны он был завезен во Францию, но обратили на него внимание только тогда, когда он размножился на значительной территории. За период с 1922 года (когда его заметили) до второй мировой войны колорадский жук достиг границ Германии, а во время второй мировой войны он проник далеко на восток. Фашистская оккупация способствовала его завозу в Чехословакию, Югославию, Польшу, даже на запад УССР. В СССР его очаги были ликвидированы, но постепенное распространение жука (несмотря на строгие мероприятия по борьбе с ним) в ГДР и Польше привело к тому, что он продвинулся на восток и проник на нашу территорию на Украине, в Белоруссии, Прибалтике (рис. 128). Так колорадский жук стал объектом не только внешнего, но внутреннего карантина в нашей стране.
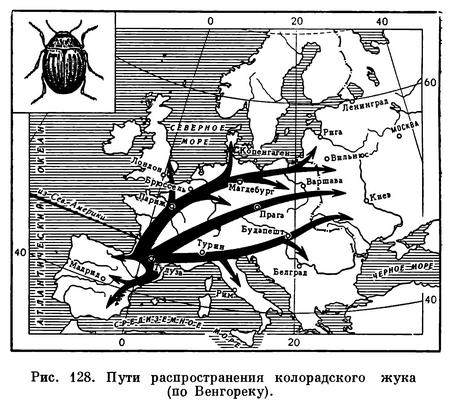
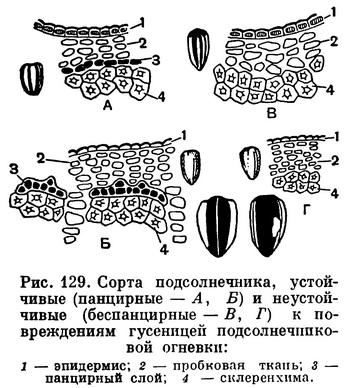
Чтобы стать вредителем культурного растения, насекомое должно не только питаться им, но и приспособиться к существованию в тех условиях, которые создаются при его возделывании. А эти условия в значительной мере в руках человека. Правильно организуя севообороты, выбирая приемы агротехники, которые благоприятны для растения, но неблагоприятны для вредителя, можно подавить развитие вредителя и даже свести на нет наносимый им вред.
Замена бессменной культуры пшеницы правильными севооборотами на юге УССР и Северном Кавказе привела к снижению численности хлебного жука-кузьки, к тому, что перестали вредить земляные усачи, пырейные огневки и другие виды. Распашка меж, связанная с переходом от единоличных форм землепользования к коллективным, привела к ликвидации вреда от жука-кравчика. Зяблевая вспашка и ликвидация послеуборочных остатков уничтожают зимующие яйца капустной тли, хлебных пилильщиков и других вредителей. Занятые пары сдерживают размножение озимой совки и т. д. Большое значение имеет выведение устойчивых к вредителям сортов растений — подсолнечника к подсолнечниковой огневке (рис. 129), яблони к кровяной тле и т. д. Комплекс агротехнических мероприятий, меняя среду обитания насекомого и растения, является решающим звеном в деле защиты растений.
Большую роль в снижении численности вредителей играют их естественные враги — хищники, паразиты, возбудители болезней.
Среди насекомых большое число видов — хищники, а многие развиваются паразитируя в других насекомых. Использование таких естественных врагов насекомых-вредителей позволяет не допускать потери от них урожая. Эти мероприятия составляют основу биологического метода борьбы с вредителями.
Особенно хорошие результаты дает биологический метод против тех вредителей, которые были завезены и не встретили эффективных местных врагов. Показательны такие примеры, как кровяная тля. Этот вредитель корней яблони — американского происхождения, и, например, в СССР не было такого естественного врага тли, который бы сдерживал ее размножение. Ввоз ее паразита наездника афелинуса (Aphelinus mali) дал во многих районах прекрасные результаты. Блестящий пример успешной биологической борьбы дает история австралийского желобчатого червеца. Но бывает и так, что ввоз естественных врагов не дает эффекта. Например, в США для борьбы с непарным шелкопрядом, завезенным из Европы, были завезены все главные враги этого вредителя, сдерживающие его размножение. Но на новой родине хищники и паразиты либо не прижились, либо перешли на питание другими насекомыми — биологическая борьба с непарным шелкопрядом не удалась. Были такие примеры и у нас. Например, против калифорнийской щитовки в Краснодарский край были завезены с Дальнего Востока ее враги — хищные божьи коровки хилокорусы. Хилокорусы прижились и размножились, но... перешли к питанию другими щитовками, а не калифорнийской.
Для борьбы с вредителями можно использовать и местных хищных и паразитических насекомых. Например, для борьбы с тлями в предгорных равнинах используют местных божьих коровок — хищников, в массе уничтожающих тлей. Божьи коровки в горах зимуют большими скоплениями. Их собирают и держат в холодильниках, пока не появятся на культурных растениях тли, а затем выпускают.
В некоторых случаях в борьбе с многими вредными гусеницами хороший результат дает использование наездника-яйцееда трихограммы, развивающейся в яйцах многих чешуекрылых. В лабораторных условиях разводят трихограмму на яйцах зерновой моли, а в период массовой яйцекладки вредителей (озимой совки, яблонной плодожорки) яйца моли с находящейся в них трихограммой выносят на защищаемые участки: трихограмма вылетает и заражает яйца вредителя. Очень важно для подавления численности вредителей не только разводить его паразитов, но и создавать для них благоприятные условия. Например, большинство наездников, личинки которых развиваются во вредных насекомых, убивая их, во взрослом состоянии нуждаются в питании нектаром. Поддерживание вблизи полей и садов цветущих круглое лето растений повышает число наездников.
Конечно, важное звено в системе мер борьбы с вредными насекомыми представляет привлечение насекомоядных птиц. Обнадеживающие результаты дает применение культур микроорганизмов, вызывающих заболевания и гибель вредных насекомых. К ним относится препарат «энтобактерин», с успехом испытанный против ряда вредных гусениц, культура грибка боверии (Beauveria bassiana), опыливание которой во влажные годы вызывает гибель колорадского жука, и др.
Однако часто ни агротехнических мероприятий, ни использования естественных врагов насекомых не бывает достаточно для защиты культурных растений. В практике все шире применяется химическпй метод борьбы с вредными насекомыми, чаще всего основанный на том, что защищаемые растения и находящиеся на них вредители покрываются тончайшим слоем ядовитого для насекомых вещества. Яды наносятся на растения либо путем опрыскивания, обычно в водном растворе или в виде растворов в маслах и эмульсиях, либо, теперь чаще, в виде мельчайшего порошка (опыливание). А за последние 10—15 лет внедряется и использование ядов в виде мельчайшей взвеси в воздухе (аэрозоли).
Для химической борьбы с вредными насекомыми либо применяют специальные наземные машины (опрыскиватели), либо яды распыляют с помощью самолетов — метод, особенно распространенный у нас для борьбы с вредителями лесов, полевых культур и садов. Авиаметод в борьбе с вредителями был впервые введен в практику в СССР. Для борьбы о вредителями, имеющими грызущий ротовой аппарат, например против саранчовых, применяются яды кишечного действия, попадающие в организм насекомого вместе с пищей. Это в основном различные неорганические соединения — соли мышьяковой и мышьяковистой кислот, фтористые соединения и др.
За последние десятилетия было синтезировано много хлорорганических соединений, очень эффективных в борьбе с вредителями. Это прежде всего ДДТ (дихлордиэтилтрихлорэтан), ГХЦГ (гексахлорциклогексан), алдрин, диэлдрин, гептахлор и др. Они действуют не только попадая в кишечник, но и контактно, попадая в организм насекомого через покровы. На многих насекомых эти яды действуют в совершенно ничтожных количествах и могут применяться и против грызущих, и против сосущих насекомых. Другая группа синтетических ядов — фосфорорганические (меркаптофос, тиофос, вофатокс и др.). Многие из них впитываются растением (не вызывая его гибели) и отравляют насекомое, высасывающее из него соки, действуя как «системный яд», а не только при контакте с вредителем. Это позволяет бороться даже со скрытоживущими вредителями.
Фосфорорганические яды очень ядовиты и для человека, что ограничивает их применение. Некоторые хлорорганические яды (ДДТ) сначала считались безвредными для теплокровных организмов. Однако теперь выяснилось, что ДДТ, например, накапливается в организме человека, вызывая тяжелые изменения в печени и в нервной системе, что заставляет ограничивать применение таких препаратов.
Хлорорганические и фосфорорганические препараты отличаются тем, что действуют не только на вредителей, но и на других насекомых, в том числе и полезных — опылителей и энтомофагов (хищников и паразитов вредителей), а также на многих почвообразователей.
Поэтому, несмотря на высокую эффективность и широкое применение химических мер борьбы с вредителями, в последние годы все больше специалистов высказывается за ограничение применения современных синтетических ядов. Нередко в местностях, где регулярно проводится борьба с вредителями химическими методами, оказываются уничтоженными естественные враги и того вредителя, с которым боролись, и враги тех вредителей, борьба с которыми не предполагалась. Например, в плодовых садах, регулярно обрабатываемых ДДТ против плодожорки и других насекомых, в массе размножаются паутинные клещики, не погибающие от ДДТ, а даже становящиеся от него более плодовитыми. Это связано с тем, что ДДТ убивает тех хищных насекомых, которые сдерживают размножение клещиков.
Неизбирательное уничтожение большинства насекомых при химической борьбе приводит к гибели насекомоядных птиц — в итоге уничтожаются факторы естественной регулировки численностн вредителей. На последнем Международном энтомологическом конгрессе в 1964 году в Лондоне многие крупнейшие специалисты высказывались за разработку той системы мероприятий, которая исключала бы массовое применение химических мер борьбы с вредителями. Такую интегральную борьбу с вредителями приветствовали и участники II Международного конгресса по защите растений в 1965 году в Неаполе. Основой в деле борьбы с вредителями должно быть то правило, что легче не допускать размножения вредителя, чем потом его уничтожать на больших площадях. Надо создать такой комплекс агротехнических (а в лесу — лесохозяйственных) условий и такой биологический режим, при котором бы не было предпосылок для массовых размножений вредителей. Химическую борьбу надо проводить не на больших площадях, когда вредитель размножился, как это часто во всем мире делают сейчас, а в самом начале вспышек размножения, на тех небольших участках, где вредитель только начинает появляться в больших количествах. Это важно и потому, что для успешного уничтожения вредителя химическим методом нужно, чтобы мероприятие было проведено в определенный срок, когда вредитель наиболее доступен для уничтожения и наиболее чувствителен к ядам. Своевременное проведение борьбы на ограниченном участке — выполнимая задача. На больших же площадях, даже при использовании самолетов, борьба растягивается и может совпасть с периодами, когда яд на вредителя не действует. При таком синтезе биологических и химических мер борьбы следует ожидать хороших результатов. Но организовать такую борьбу можно только при глубоком знании особенностей и вредителей, и их паразитов, и хищников, хорошо зная жизнь животных!
Нередко применение химических мер борьбы с вредителями встречается с неожиданным затруднением. Вредители, прежде чувствительные к яду, становятся устойчивыми к нему. Например, во многих местностях чувствительность комнатной мухи к ДДТ уменьшилась в 10 раз. В Западной Европе во многих местностях резко снизилась чувствительность колорадского жука к ДДТ и линдану. Такие изменения наследственных особенностей насекомых под влиянием регулярно действующего нового фактора естественного отбора представляют не только серьезную практическую проблему (время от времени приходится менять применяемые яды), но и большой биологический интерес, так как показывают, что у насекомых за короткое время могут измениться наследственные свойства. Большая плодовитость и высокая скорость их размножения недаром сделали многих насекомых теми объектами, на которых решаются закономерности наследственности (дрозофила, мучной хрущак, шелкопряды и др.).
Конечно, вредители растений и борьба с ними — серьезная экономическая проблема, стоящая перед человечеством. Но не менее, а часто более важны те насекомые, биология которых связана с теплокровными животными и непосредственно с самим человеком.
Например, только от кожных оводов ежегодная потеря продукции крупного рогатого скота в СССР составляет около 0,5 миллиарда рублей. Огромные убытки животноводству и снижение работоспособности людей причиняет в осваиваемых таежных и других лесных районах так называемый «гнус» — совокупность различных кровососущих насекомых, наносящих болезненные зудящие уколы. Это слепни, мошки, мокрецы — двукрылые разных размеров, связанные с неосвоенными территориями. Убытки от гнуса огромны, не говоря о том угнетающем действии, которое он производит на работающих, например в тайге, людей — лесозаготовителей, геологов, строителей. При строительстве, например, Братской ГЭС много усилий и средств пришлось затратить на борьбу с насекомыми-кровососами.
Еще опаснее те кровососы, которые переносят возбудителей различных заболеваний. Тут уж причиняемый ими вред исчисляется не в денежном выражении.
а человеческими жизнями. До приобретения независимости в Индии от малярии, возбудитель которой передается малярийным комаром (род Anopheles), ежегодно умирало около 200 тысяч человек.
Большое снижение работоспособности людей вследствие заболевания малярией было у нас в рисосеющих районах Средней Азии до того, как малярия как массовое заболевание в СССР была ликвидирована. В тропической Америке не поддается учету количество человеческих жизней, уносимых передаваемой комарами желтой лихорадкой. В Западной Африке бичом населения является передаваемая мухой це-це сонная болезнь.
В наших широтах огромное эпидемиологическое значение имеют вши, передающие сыпной и возвратный тифы, и блохи, передающие чуму. Если наша страна не знает теперь вспышек этих болезней, то это потому, что налажен строгий контроль за этими заболеваниями и созданы санитарно-гигиенические условия, препятствующие распространению насекомых-паразитов. Огромна роль мух в переносе различных кишечных инфекций, в том числе и таких, как брюшной тиф, дизентерия и др. Только насекомыми передаются менее опасные, но тяжелые болезни, такие, как пендинская язва, лихорадка папатачи и др.
Приведенные данные показывают, насколько велика роль насекомых в природе, в экономике общества, в жизни людей, насколько необходимо всестороннее изучение огромной армии наших шестиногих друзей, союзников и врагов.
РАЗДЕЛ ПЕРВИЧНОБЕСКРЫЛЫЕ НАСЕКОМЫЕ (APTERYGOTA)
ОТРЯД ЩЕТИНОХВОСТКИ (THYSANURA)
Среди насекомых много таких, у которых нет крыльев. Бывает, что у насекомых нет даже зачатков крыльев, особенно у разных паразитов (вши, пухоеды, блохи). Однако изучение внутреннего строения и развития таких насекомых и сравнение их с другими, крылатыми насекомыми убеждает в том, что они происходят от каких-то крылатых форм, что они утратили крылья в процессе эволюции, приспособившись к каким-то специальным условиям жизни, сделавшим крылья ненужными.
Изучение щетинохвосток показывает другое: оказывается, это такие насекомые, у которых нет ни малейших признаков исчезновения крыльев, крыльев нет не только у них, но не было и у их предков. Поэтому они и выделяются в группу первичнобескрылых и могут рассматриваться как примитивные насекомые, сохранившие много тех черт, которые были присущи и многоножкообразным предкам этого обширного класса.
Щетинохвостки характеризуются тем, что у них более или менее веретеновидное тело (табл. 13, 17). На голове находятся фасеточные глаза, между которыми располагаются три простых глазка. Усики у них длинные, нитевидные. На грудных сегментах — хорошо развитые ноги, заканчивающиеся 2 коготками. Грудь плавно переходит в сужающееся кзади 11-члениковое брюшко, на конце которого находится пара длинных членистых церков и между ними еще более длинная хвостовая нить (рис. 130).
На брюшных сегментах у щетинохвосток находятся нерасчлененные придатки, занимающие такое же положение, как грудные конечности. На эти грифельки щетинохвостки опираются при движении; грифельки — действующие рудименты ног заднего отдела туловища.
Спинная поверхность тела щетинохвосток покрыта нежными тонкими чешуйками, придающими щетинохвосткам серебристый блеск. В этих чешуйках есть непроницаемый для испарения слой, и благодаря этому щетинохвостки могут находиться некоторое время в сухой атмосфере, хотя в общем щетинохвостки — влаголюбивые насекомые. Интересно они возобновляют запас воды в своем теле. У них у основания грифельков находятся выпячивающиеся пузырьки с очень тонкими покровами. Этими пузырьками щетинохвостки приникают к влажной поверхности скалы, почвы и т. п. и впитывают воду.
Живут щетинохвостки долго, некоторые до 3 лет, и линяют не только пока растут, но и во взрослом состоянии. Развитие щетинохвосток проходит без превращения: из яйца выходит личинка, похожая на взрослое насекомое, отличающаяся лишь меньшим числом члеников усиков и хвостовых нитей.
Среди щетинохвосток различается 2 подотряда. Более примитивные махилисы (подотряд Machiloidea) живут преимущественно среди скал и нагромождений крупных камней, покрытых лишайниками, которыми эти насекомые и питаются. Махилисы довольно крупные насекомые, до 1,5 см длиной. Большую часть времени они ведут скрытый образ жизни, держась между камнями и под ними, но временами, особенно по утрам, когда воздух влажен, они выходят погреться на освещенную солнцем поверхность скал.
Махилисы передвигаются либо бегая, либо совершая прыжки. Прыгают они на расстояние, раз в 10 превышающее длину их тела. В прыжке участвуют брюшко, которое они резким движением сокращают, грифельки, которые они при этом отгибают, и ноги. Они могут передвигаться и по поверхности воды, плавая, как тюлени, и даже выпрыгивая из воды на высоту до 2 см.
Распространены махилисы в подходящих местах по всему земному шару, кроме самых холодных, у нас на севере доходят, например, до Финляндии и Урала. Много их в горных местностях, особенно близ морей (у нас приморский махилис — Machilis maritimus встречается на скалах Южного берега Крыма). В горах махилисы встречаются на высоте до 4000 м, причем интересно, что северные виды, например, представители родов Lepismachilis, Dila, доходящие до Финляндии, в Альпах не держатся ниже 2000 м.
Любопытно происходит размножение у махилисов. В период спариваиия самец выпускает паутинку, на которую выделяет капельки семенной жидкости, а затем заставляет самку стать рядом с паутинкой. После этого самец у одних видов усиками, у других хвостовой нитью собирает с паутинки капельки семенной жидкости и переносит их к половому отверстию самки. Таким образом, у махилисов оплодотворение похоже на оплодотворение у пауков, а по типу наружно-внутреннее, как у многоножек.
Яйца махплисы откладывают во влажные места: без доступа воды из окружающей почвы яйца не развиваются.
Многим знакомы юркие чешуйницы (подотряд Lepismoidea), некоторые виды которых, например сахарная чешуйница (Lepisma saccharina, табл. 13), встречаются в домах, в более сырых местах. Многие видели это проворное изящное отливающее серебром насекомое, которое на многих языках даже называют «серебряной рыбкой». Веретеновидно сужающееся тело чешуйницы достигает длины около 1 см.
Чешуйницы — теплолюбивые насекомые и держатся в домах в более теплых местах, но там, где могут найти влажные укрытия. Они боятся света и выходят только ночью. Войдя в помещение, где есть чешуйницы, и включив свет, легко увидеть их паническое бегство.
Чешуйницы — вегетарианцы, хотя иногда питаются трупами насекомых. Онн питаются гифами и спорами грибков, одноклеточными водорослями, в домах — скоплениями пыли, в которой всегда находится что-либо для них съедобное. Охотно питаются они крахмалом клея, которым приклеены обои, но могут питаться и клетчаткой, например влажной бумагой, которую переваривают с помощью населяющих их кишечник бактерий. Пища задерживается в кишечнике чешуйниц долго, неделю-две.
Не надо думать, что чешуйницы обитают только в домах. На Южном берегу Крыма встречается гребенчатая чешуйница (Ctenolepisma lineata), живущая в почве на известняковых склонах. А некоторые чешуйницы приспособились к жизни в гнездах муравьев, где не только питаются всяким мусором, по даже получают пищу от муравьев. Муравей отрыгивает из зоба пищу голодному сочлену своего муравейника, а в это время «квартирант» — чешуйница муравьиная (Atelura formicaria) приближается к муравьям и слизывает передаваемую каплю, что не вызывает протеста у муравьев (рис. 131).
Чешуйницы довольно надежно защищены от высыхания чешуйками и потому могут забираться и в сухие места, но в периоды своих частых линек они не защищены от потери воды и на это время забираются в щели, под камни, в трещины почвы — в такие места, где воздух насыщен водяным паром.
Размножение чешуйниц — с очень типичным наружно-внутренним оплодотворением. Встретившись с самкой, самец откладывает на землю грушевидный сперматофор. Затем он толкает самку на сперматофор и самка сама подбирает его половыми придатками. Этот способ оплодотворения чешуйниц позволяет понять происхождение сперматофорного размножения у прямокрылых и других крылатых насекомых.
Интересно, что уже в мезозое, в триасовый период, жили махилисы, очень похожие на современных. А. Г. Шаровым описан из триаса Triassomachilis, лишь мелкими деталями отличающийся от современных. А из пермских отложений палеозоя этот же ученый описал близкий к махилисам новый отряд ископаемых насекомых — однохвосток (Monura), имевших одну среднюю хвостовую нить, но не имевших церков.

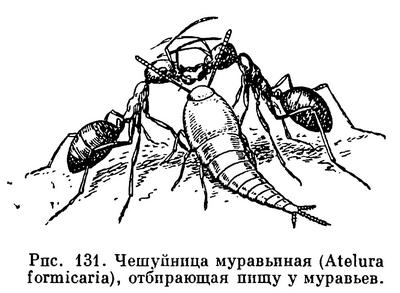
РАЗДЕЛ КРЫЛАТЫЕ НАСЕКОМЫЕ (PTERYGOTA)
У представителей этого огромного раздела во взрослом состоянии имеются развитые или редуцированные крылья, а в некоторых случаях, когда у перешедших к неподвижному или паразитическому образу жизни насекомых, относимых к этой группе, крыльев даже нет совсем, но по строению их других органов можно заключить, что их предки были крылатыми.
Во взрослом окрыленном состоянии крылатые насекомые не линяют — исключение представляют только поденки.
Крылатых насекомых разделяют на две большие группы — на два отдела.
К насекомым с неполным превращением (Hemimetabola) относятся те, у которых развитие после выхода из яйца проходит без непитающейся стадии куколки (за исключением самцов щитовок и червецов, у которых куколка есть). Для насекомых с неполным превращением типично то, что у выходящих из яиц личинок расчленение тела и конечностей такое же, как у взрослого насекомого, и глаза, как правило, фасеточные, если онн фасеточные у взрослых насекомых.
На первых этапах развития личинки таких насекомых (если взрослое насекомое и имеет крылья) — бескрылые, но по мере роста и линек у них появляются наружные зачатки крыльев.
У тех видов, у которых взрослые насекомые утратили крылья, развитие приближается к прямому. Личинок насекомых с неполным превращением теперь принято называть «нимфами», но иногда этот термин применяется только к личинкам старших возрастов, уже имеющим видимые зачатки крыльев. Личинки тех групп насекомых с неполным превращением, которые обитают в воде, называются «наядами». Но термином «личинка» можно пользоваться и в отношении «нимф», и в отношении «наяд».
К насекомым с полным превращением (Holometabola) относятся те, которые в своем развитии после личиночной стадии обязательно проходят покоящуюся (но не всегда неподвижную или малоподвижную) стадию куколки. Для личинок насекомых с полным превращением типично то, что их облик непохож на облик взрослого насекомого, разделение тела на отделы и число сегментов брюшка, а часто и строение ротового аппарата отличны от этих же признаков взрослого насекомого; ноги у личинок насекомых с полным превращением имеют иное расчленение, чем у взрослых, — лапка у личинок не бывает членистой; глаза у личинок насекомых с полным превращением никогда не бывают фасеточными.
Несмотря на резкие различия личинок насекомых с полным превращением и с неполным, абсолютно непроходимой грани между этими типами развития нет.
Однако все же деление на эти 2 группы более естественно, чем другие деления, поскольку оно основано на учете признаков всего развития насекомого.
В литературе сейчас принято деление всех крылатых насекомых на два отдела — «древнекрылых», неспособных складывать крылья на спине, и «новокрылых», складывающих крылья. Это деление было предложено А. В. Мартыновым, изучившим богатый ископаемый материал.
Объединение далеких друг от друга стрекоз и поденок в общую группу древнекрылых искусственно, и в этой книге принимается более старое, но более естественное деление на Hemimetabola и Holometabola.
ОТДЕЛ НАСЕКОМЫЕ С НЕПОЛНЫМ ПРЕВРАЩЕНИЕМ (HEMIMETABOLA)
ОТРЯД ТАРАКАНОВЫЕ (BLATTODEA)
Слово «таракан» вызывает чаще всего неприятные ассоциации: с ним связано представление о некоторых весьма докучливых сожителях человека, широко распространившихся вместе с ним по всему земному шару. А между тем это очень интересная группа животных, жизнь которых богата любопытными фактами.
Таракановые обладают плоским овальным телом с головой, обращенной ротовым отверстием книзу и почти или совсем прикрытой большой щитообразной переднеспинкой. Усики у них многочлениковые, щетинковидные; ноги бегательного типа с уплощенными бедрами и 5-члениковыми лапками. В тех случаях, когда имеется летательный аппарат, он всегда очень характерен: передние крылья плотные, роговые или кожистые, с обильным жилкованием; крылья перепончатые, в спокойном состоянии спрятаны под надкрыльями. Однако и надкрылья и крылья могут быть укорочены или полностью отсутствовать. Брюшко удлиненное, 8—10-члениковое, с мягкими покровами и на конце несет длинные, обычно членистые церки; у самцов на последнем стерните имеется 1 или 2 грифелька. Большинство таракановых имеет светлую окраску тела и надкрылий желтовато-коричневых оттенков, реже распространена темная или черная окраска.
Весь облик таракана с его уплощенным телом и плотными наружными покровами как нельзя лучше соответствует образу жизни этих насекомых. Среди них преобладают ночные животные, активные в темноте, а на день заползающие под камни или опавшие листья, в трещины на поверхности почвы, в норы грызунов и под кору пней и отмирающих деревьев. Однако есть и уклонения от этого типичного образа жизни. Во всех таких уклоняющихся случаях постоянно наблюдаются и изменения внешнего вида таракана.
В настоящее время известно свыше 2500 видов таракановых, из которых большинство (не менее 80%) тоже связано с лесами тропической зоны. Так, многие индийские и индонезийские виды встречаются в сырых местах по краям протекающих в джунглях ручейков. Живущие в таких условиях тараканы могут даже плавать. В этом отношении интересен саравакский таракан (Bhicnoda natatrix) с острова Калимантан (Борнео). Его личинки прячутся по краям луж под листьями, осыпающимися с деревьев. Если личинку потревожить, она бросается к воде и ныряет на дно. Не менее существенную группу тропических тараканов составляют виды, живущие в разлагающейся древесине отмирающих деревьев.
По сравнению с тропической фауной фауна СССР очень бедна представителями этого отряда: в ней насчитывается немногим больше 50 видов, включая сюда и виды иноземного происхождения, акклиматизировавшиеся в жилищах человека. И в то же время эта фауна очень поучительна. Ее изучение дает возможность восстановить ход истории развития такой оригинальной группы насекомых, какой являются таракановые, на всей территории Палеарктики.
Начавшееся здесь к концу третичного периода изменение климата привело к созданию неблагоприятных условий для существования тропических лесных видов. Это, с одной стороны, обусловило вымирание ряда широко распространенных третичных форм, а с другой стороны, явилось причиной формирования новых современных видов, приспособлявшихся к жизни в пустынях, степях и таежных лесах. Поэтому в каждой ландшафтной зоне Советского Союза имеются специфические таракановые, весь облик которых и образ жизни отражают особенности условий, в которых они сформировались и обитают сейчас.
Широко известны синантропные виды тараканов, связанные в своем распространении с человеком, особенно рыжий, или прусак (Blattella germanica), и черный (Blatta orientalis). По форме тела и надкрылий первый из них напоминает лапландского таракана. Он буровато-рыжего цвета, с 2 темными полосками на переднеспинке, длиной 10—13 мм. Родина его, по-видимому, Южная Азия, откуда он был в XVIII столетии завезен в Европу, в странах которой очень быстро распространился. Живет в теплых, отапливаемых зимой помещениях, особенно в старых банях и пекарнях, а также на кухнях. Теплолюбив и не выносит пониженных температур. При 22°С развивается в течение 172 дней; при более высоких температурах, порядка 30°С, развитие его сокращается до 75 дней. Температура ниже 5°С для него уже смертельна: при —5°С погибает через 30 минут, а при — 7°С — через минуту. Живя в помещениях, может питаться остатками хлеба, овощей, сахара, масла и других продуктов. Кроме того, повреждает бумагу, переплеты книг, а также обувь и другие кожаные предметы. В условиях теплого климата, например в Крыму, в южной части Средней Азии и на Дальнем Востоке, живет и на воле.
Черный таракан — более крупный вид, 18—30 мм длиной, с черным или черновато-бурым, как будто лакированным телом. У самца надкрылья немного короче брюшка, а у самки они короткие, в виде небольших чешуевидных лопастей. Своеобразный неприятный запах, исходящий от этого насекомого, зависит от быстро испаряющихся выделений особых кожных желез, находящихся в шестом сегменте брюшка. Родина черного таракана точно не установлена; род, к которому он относится, включает немного видов, распространенных в Африке и Австралии. В Европе появился не менее чем 300 лет тому назад; на Балканском полуострове и в Крыму может обитать и на воле, прячась днем в трещинах старых глиняных стен и под камнями по соседству с жильем. В постройках встречается обычно в таких же местах и питается такими же продуктами, как и прусак. Ведет ночной образ жизни. Личинки растут очень медленно, и их развитие растягивается до 4 лет.
Синантропные тараканы опасны для человека не только тем, что они портят и загрязняют продукты. Помимо того, они разносят различные бактерии и яйца паразитических червей. Бактерии, вызывающие дизентерию и другие кишечные заболевания человека, переносятся ими как на поверхности усиков и ног, так и внутри кишечного канала, из которого выходят вместе с экскрементами. В прямой кишке черного таракана обнаружены яйца власоглава и острицы, а в кишечнике прусака, помимо этих паразитов, еще яйца лентеца широкого. Поэтому борьба с домовыми тараканами представляет важное звено в системе санитарно-эпидемиологических мероприятий.
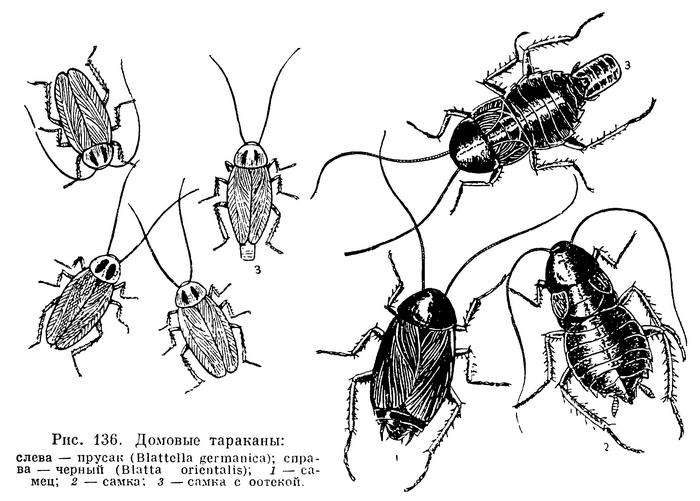
ОТРЯД ТЕРМИТЫ (ISOPTERA)
Термитов часто называют «белыми муравьями». Это название термиты получили из-за того, что они, как и муравьи, ведут «общественный» образ жизни, нередко сооружают конические постройки, как и муравьи, характеризуются полиморфизмом (кстати, у термитов полиморфизм сильнее выражен, чем у муравьев), причем основную роль в поддержании жизни колонии у термитов, как и у муравьев, играют недоразвитые в половом отношении особи. Но этими аналогиями, определяемыми сходными условиями жизни, и ограничивается сходство термитов и муравьев. Термиты — отряд насекомых с неполным превращением, а муравьи представители не только другого отряда (перепончатокрылых), но и другого отдела насекомых — Holometabola.
Термиты почти незнакомы жителям местностей с умеренным климатом: основная их стихия — тропики и субтропики, особенно тропики. Правда, отдельные виды распространены и шире и доходят, например, у нас до юга УССР, а в больших городах, приспособившись к жизни в отапливаемых зданиях, термиты могут встречаться и севернее: много термитов в Гамбурге, у нас термиты отмечены в Днепропетровске. Но в общем термиты — обитатели тропиков.
Всего известно около 2500 видов термитов.
Термиты — это средней величины насекомые. Размеры особей у одного вида и даже у одной касты сильно варьируют (у Bellicositermes natalensis — всеразрушающего термита Южной Африки — половые особи имеют длину 1,5 см, рабочие — 0,5—0,8 см, солдаты — до 1,5 см).
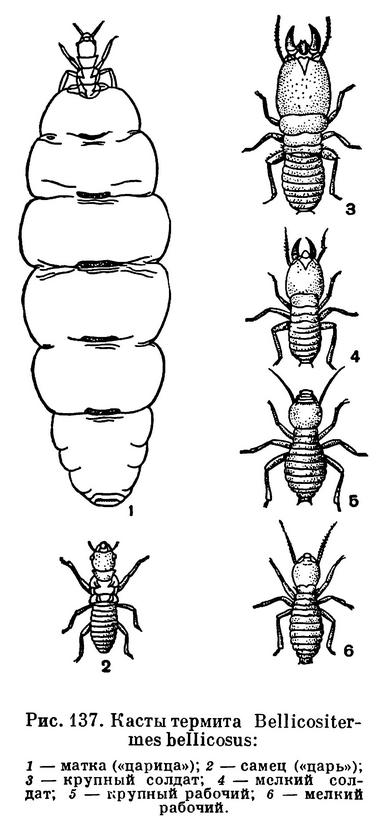
Обычно в семье, насчитывающей от нескольких сотен до сотен тысяч и даже миллионов особей, есть одна яйцекладущая самка («царица») и оплодотворяющий ее самец («царь»). Это половозрелые особи, сбросившие крылья. Кроме того, в термитнике в определенные периоды (перед роением) бывает довольно много выведшихся в нем крылатых самцов и самок, которые при подходящей погоде и в определенный срок оставляют гнездо, чтобы основать новые колонии.
Крылатые особи характеризуются наличием двух пар одинаково развитых, сходных по жилкованию длинных сетчатых крыльев; крылья настолько длинны, что, когда сложены на спине, далеко выступают за конец брюшка. По строению крыльев отряд и получил свое название (Isoptera — «одинаковокрылые»). Сегменты груди и брюшка у крылатых термитов довольно сильно склеротизованы.
Основная масса населения термитника — рабочие особи. Рабочие — недоразвитые в половом отношении самцы и самки. В этом отношении термиты сильно отличаются от муравьев, у которых, как и у других общественных перепончатокрылых, рабочие — всегда самки. Рабочие особи напоминают личинок термитов — в сущности развитие рабочих термитов после выхода из яйца прямое. У рабочих особей покровы мягкие, непигментированные, что связано с их постоянным обитанием в укрытиях, в атмосфере, насыщенной водяным паром. В этом отношении среди термитов есть только одно исключение. Некоторые южноафриканские термиты (Hodotermes) имеют рабочих, открыто обитающих; у них покровы темно-коричневые или черные. Но, как правило, покровы термитов бывают мягкими, тонкими, а у разводящих грибы Bellicositermes natalensis прозрачна даже головная капсула и сквозь покровы видны все внутренние органы насекомого.
Для рабочих характерна округлая голова, слабо развитой грудной отдел. На заднем конце брюшка — чувствующие 2—5-члениковые церки — признак, характерный для скрытоживущих форм. Глаза у рабочих особей недоразвиты, а часто и вовсе отсутствуют.
Солдаты — особая категория специализированных рабочих особей, характеризующихся очень сильно развитой головной капсулой и мощными длинными жвалами. Эти челюсти пускаются в ход против врагов — термитов других видов, а главное, против муравьев. У некоторых «носатых» солдат (рис. 138) в отростке головы проходит канал железы, через который на противника выбрызгивается клейкая жидкость, связывающая движение насекомого.
Термиты питаются в основном растительной пищей. К самостоятельному питанию у термитов способны только рабочие особи. Солдаты из-за непомерного развития жвал и слабого развития остальных частей ротового аппарата сами не питаются: их кормят рабочие особи либо выделениями изо рта, либо экскрементами непосредственно из заднепроходного отверстия — в них еще достаточно питательных веществ для солдат. Половые особи после основания колонии выкармливаются выделениями слюнных желез рабочих или личинок. Самых мелких личинок тоже кормят рабочие, давая им выделения своих слюнных желез или разжеванные споры грибов.
По мере роста колонии заметно меняется самка. У нее атрофируется и крыловая мускулатура, и мускулатура конечностей, даже мускулатура ротовых частей — идет «обратное развитие». Зато прогрессивно растет переполненное яйцами брюшко. Самка становится неподвижной, всецело зависящей от кормящих ее рабочих особей (рис. 141), она все время откладывает яйца, а рабочие выкармливают личинок, превращающихся в новых рабочих. Самка термитов выделяет какие-то вещества, слизываемые облизывающими ее рабочими особями. Эти вещества содержат телергоны (иначе, феромоны), влияющие на развитие личинок. Только когда колония разрастется или самка ослабеет, начинают появляться и крылатые особи: очевидно, в этом случае часть личинок не подвергается воздействию тормозящих развитие телергонов.
Нам трудно даже представить себе ту роль, которую термиты играют в жизни тропической природы, в быту жителей жарких стран.
В тропических лесах термиты — основные разрушители всех растительных остатков. Образование почвы в тропиках, перемешивание ее слоев, круговорот веществ в тропическом лесу — процессы, определяемые деятельностью термитов. Других почвенных животных в тропических лесах часто и не бывает, но термиты кишат. За редким исключением, термиты питаются только отмершей древесиной и в девственных лесах в большой мере определяют плодородие почвы. Но когда с термитами сталкиваются интересы человека, положительная роль их отступает перед тем вредом, который они нам причиняют.
Все деревянные сооружения подвергаются разрушительной деятельности термитов. Деревянный дом стоит всего несколько лет. Но и каменные фундаменты не спасают деревянные конструкции зданий от термитов. Эти влаголюбивые и избегающие света насекомые сооружают на поверхности каменных частей зданий крытые галереи, склеивая их из глинистых частиц так, чтобы они сообщались с почвой. Внутреннюю поверхность таких ходов термиты обрызгивают выделяемой ими жидкостью, чтобы поддерживать в галереях необходимую влажность.
По таким галереям термиты проникают к деревянным перекрытиям и буквально изрешечивают их, в результате чего обваливаются потолки, проваливаются полы и т. д. В доме, пустовавшем несколько месяцев, нередко мебель разваливается от легкого прикосновения — термиты выгрызают в деревянных предметах свои ходы, так что остается только тонкая пластинка на поверхности, защищающая от открытого воздуха, который термиты не выносят, и губчатые перемычки внутри досок, поддерживающие ставшие легкими источенные предметы. В Южной Америке редко можно найти уцелевшую от них книгу, изданную более 50 лет тому назад. В Африке, Индии, Юго-Восточной Азии известно много случаев, когда из-за термитов приходилось переносить целые поселки и даже города — настолько большой вред они причиняют. Иногда термиты способствуют ускорению гибели фруктовых деревьев.
В Индии ежегодные убытки от термитов определяются в 280 миллионов рупий.
У нас термиты наиболее распространены в Средней Азии: в Каракумах, Кызылкуме, в Голодной степи в массе встречаются делающие подземные гнезда закаспийский термит (Anacanthotermes ahngerianus) и туркестанский термит (Anacanthotermes turkestanicus). Поселения закаспийского термита узнаются по чуть выпуклому округлому широкому холмику, по окраске почвы, несколько отличающейся от окружающего фона. А туркестанского термита можно обнаружить по земляным галереям, проложенным по стволикам и стеблям сухих пустынных кустарников.
В городах и других поселениях эти термиты сильно повреждают постройки. Они разрушают саман (необожженный кирпич из глины с соломой), из которого легко и удобно строить в сухих местностях. Разрушают они и деревянные перекрытия зданий, хотя обычно в природных условиях почти не оставляют почву. Так, был случай обвала перекрытий одной из фабрик в Фергане, а после сильного землетрясения в Ашхабаде оказалось, что потолочные балки многих зданий были сильно изъедены термитами.
В местностях, где много термитов, перед закладкой зданий проводят затравку почвы, здание строят на бетонированном фундаменте, деревянные части зданий пропитывают противотермитными составами, деревянные шпалы заменяют железобетонными, за фундаментами домов ведут регулярные наблюдения, разрушая галереи поселяющихся термитов.
В умеренном климате термиты не так вредны. У нас в причерноморских местностях (Кавказ, юг УССР, Молдавия) распространен средиземноморский термит (Reticulitermes lucifugus), который нигде не причиняет опасных повреждений.
В СССР отмечено 5 видов термитов.
Термиты — древняя группа насекомых. Ископаемые формы, близкие к термитам, известны из пермского периода палеозойской эры.

ОТРЯД ЗОРАПТЕРЫ (ZORAPTERA)
Этот небольшой (один род Zoratypus, около 20 видов) отряд тропических насекомых стал известен зоологам сравнительно недавно — он был установлен итальянским энтомологом Ф. Сильвестри в 1913 году.
Это мелкие насекомые с грызущим ротовым аппаратом, с крупной переднегрудью, и с короткими, часто асимметричными церками. Для зораптер характерен полиморфизм (рис. 144): есть и крылатые, и бескрылые особи, причем бескрылые часто бывают непигментированными. Зораптеры ведут скрытый образ жизни (часто в заброшенных ходах термитов). Питаются, вероятно, мицелием грибов и иногда хищничают, поедая клещей. Зораптеры очень влаголюбивы и избегают света. Биология зораптер изучена слабо. Ближе всего они к термитам, и сближение их с сеноедами, проводившееся ранее, не подтверждается. Это самостоятельный своеобразный отряд, представители которого практического значения не имеют.
ОТРЯД ГРИЛЛОБЛАТТИДЫ (GRYLLOBLATTIDA)
Около пятидесяти лет тому назад, в 1914 году, в провинции Альберта в Западной Канаде было найдено интересное насекомое, описание которого вызвало большой интерес среди энтомологов всего мира. Трудно было определить, к какому отряду его можно отнести. По одним признакам это прямокрылое, ближе всего стоящее к сверчкам, по другим оно напоминает таракановых. Этим объясняется своеобразное родовое название (Grylloblatta), которое было присвоено этой форме (Gryllus — сверчок, Blatta — таракан).
В настоящее время известно 10 видов гриллоблаттид, заслуженно выделенных в особый отряд, который объединяет среднего размера бескрылых насекомых, отличающихся крупной головой, несущей многочлениковые нитевидные усики, узким, удлиненным телом, снабженным длинными, мягкими членистыми церками, а у самок также яйцекладом.
Семь видов встречается в США и Канаде, два — в Японии и один — в СССР. Столь удаленные места их нахождения характеризуются сходными условиями жизни — прохладным и влажным климатом, с обильными осадками и туманами. Обитают гриллоблаттиды под камнями или в пнях и гниющих стволах деревьев.
Один из североамериканских видов (Grylloblatta barberi) найден в штате Калифорния, близ озера Игл, на высоте около 1500 м. Здесь в глубоких расщелинах гор снег сохраняется все лето. В этих условиях гриллоблаттиды встречаются на сырых местах около льда, иногда прямо на нем. Наш вид (Grylloblattina djakonovi) известен из Приморского края; найдены самки и личинки, самцы не обнаружены. Самка достигает длины 22 мм, буровато-рыжая; церки 5-члениковые, яйцеклад равен по длине бедрам задних ног.
В Приморском крае найдены также представители рода Galloisiana, известного ранее из Японии.
Наличие у гриллоблаттид длинных церков — признак, характерный для примитивных насекомых. Гриллоблаттид заслуженно называют «живыми ископаемыми».
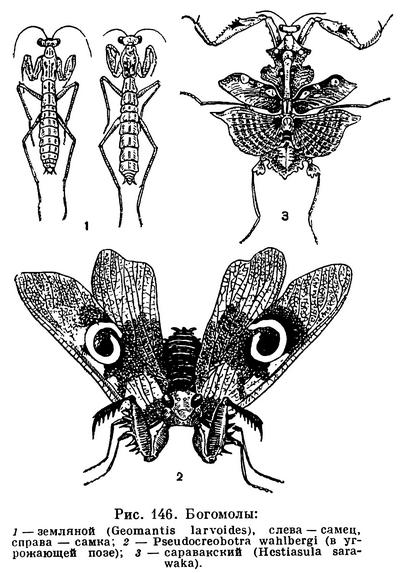
ОТРЯД БОГОМОЛОВЫЕ (MANTOPTERA, ИЛИ MANTODEA)
Богомоловые — преимущественно крупные насекомые с удлиненным телом весьма своеобразного строения. Их очень подвижная голова с большими выпуклыми глазами имеет почти треугольную форму и своим ротовым аппаратом грызущего типа направлена вниз и немного назад. Усики с большим количеством члеников обычно нитевидные, но у некоторых видов перистые или гребенчатые. Над основанием усиков располагаются 3 простых глазка. Из всех отделов груди особенно сильно развит передний, несущий характерные для богомолов хватательные конечности. Их структура настолько своеобразна, что уже по одной передней ноге можно легко отличить представителей этого отряда от всех остальных насекомых. В передней ноге богомола сильно удлинен тазик: он немного короче бедра. Бедро длинное и по нижнему краю усажено 3 рядами сильных шипов. Голень короче бедра и по нижнему краю тоже усажена острыми шипами; лапка тонкая, 5-члениковая, не отличается от лапок на остальных ногах. Бедро и голень, складываясь вместе, образуют мощный хватательный аппарат, функционирующий по типу ножниц.
У большинства богомолов хорошо развиты обе пары крыльев, причем передние более узкие и более плотные; они выполняют функцию надкрылий. Задние крылья широкие, перепончатые, в спокойном состоянии складываются веерообразно и прикрываются надкрыльями. Однако есть и совершенно бескрылые, напоминающие личинок виды, такие, как, например, обитающий в странах, окружающих Средиземное море, земляной богомол (Geomantis larvoides). Брюшко у богомолов удлиненное, 10-члениковое, мягкое, обычно слегка уплощенное, с многочлениковыми длинными церками.
Такая характерная структура тела богомола наглядно свидетельствует об очень тонких приспособлениях к хищному образу жизни, который ведут эти насекомые. По своему поведению они представляют собой классический пример «подстерегающих хищников», иначе называемых «западниками».
Подстерегающий добычу богомол сидит в толще травы или среди листьев кустарника, приподняв кверху переднеспинку и передние ноги, голени которых вложены в бедра, как лезвие перочинного ножа (табл. 25). Этой характерной позе богомол обязан своим названием. Насекомое сидит неподвижно, оно как бы замерло в «молитвенном экстазе». Только голова его находится в движении: она все время поворачивается из стороны в сторону, высматривая добычу. Но вот жертва обнаружена; богомол начинает свою охоту. Он медленными, едва заметными движениями подкрадывается к ней и затем, быстро выбрасывая вперед передние ноги, захватывает ее, зажимая между бедром и голенью. Когда добыча съедена, богомол опять принимает «молитвенную позу» в ожидании следующей жертвы.
Пищей богомолам служат самые разнообразные насекомые, и состав ее зависит главным образом от того, какие насекомые встречаются чаще во время охоты; только у молодых личинок в кормовом рационе наблюдается преобладание определенного вида корма, а именно тлей. Богомолы очень прожорливы и за время своей жизни поедают огромное количество насекомых. Так, личинка самца древесного богомола (Hierodula tenuidentata) за 85 дней своего развития съела 147 люцерновых тлей, 41 плодовую мушку и 266 комнатных мух. Будучи неприхотливыми в отношении выбора пищи, богомолы проявляют склонность к каннибализму, который наблюдается даже у молодых личинок при совместном их воспитании. Известны случаи, когда самки богомолов, которые вообще более прожорливы, поедали самцов во время спаривания или сразу после него.
Богомола, сидящего в засаде во время охоты, незаметным делает не только его неподвижная поза, но также сама форма тела и окраска. Среди богомолов встречаются виды, живущие на деревьях и напоминающие по внешнему виду кору; другие имеют палочковидную форму. Наблюдаются различные случаи имитации лишайников, листьев или цветов.
Окраска тела обычно носит защитный характер. Преобладают зеленый, желтый и бурый тона. В пределах одного и того же вида могут встречаться особи, отличающиеся по цвету. У обыкновенного богомола (Mantis religiosa) наблюдаются три цветовые формы: зеленая, желтая и бурая. Наблюдения в природе показывают, что зеленые особи обычно встречаются на вегетирующей растительности, а бурые — на растениях, выгорающих от солнца после вегетации. Для выяснения полезной роли таких окрасок у обыкновенного богомола ставилось много специальных опытов, в которых различно окрашенные особи помещались на разных фонах — соответствующих и несоответствующих их окраске. Эти опыты показали, что богомолы, находящиеся на фоне, не подходящем к их окраске, поедаются в большем количестве птицами, чем содержащиеся на фоне, соответствующем их окраске.
ОТРЯД ПРИВИДЕНЬЕВЫЕ, ИЛИ ПАЛОЧНИКИ (PHASMOPTERA, ИЛИ PHASMODEA)
Нет ни одной книги о жизни насекомых, в которой бы не говорилось об изумительной приспособленности привиденьевых к окружающей их обстановке. Полностью оценить меткость названия, данного этим насекомым, может только тот, кто сам имел удовольствие наблюдать их в природе или воспитывать дома. Если даже вы знаете, что на кусте растения находится палочник, то даже и тогда вы с трудом отыщете его среди ветвей и листьев, настолько хорошо он оказывается замаскированным. Когда же посчастливится обнаружить палочника, его причудливое тело появится перед вами бесшумно и внезапно, как это полагается сказочным привидениям. Это привидение обнаружится в виде сучка, сухой ветки или листа и меньше всего будет походить на живое насекомое.
Такая изумительная способность имитировать различные предметы объясняется особенностями строения тела привиденьевых, разнообразием их окраски и специфичностью поведения. Как правило, привиденьевые — крупные насекомые; некоторые тропические виды являются гигантами в мире насекомых, достигая в длину, вместе с вытянутыми вперед ногами, 30—35 см. Правда, есть и карликовые формы, самцы которых не превышают 2 см. Своеобразна структура отделов тела у привиденьевых: голова небольшая, шаровидная с округлыми глазами, нитевидными или щетинковидными усиками и направленным вперед грызущим ротовым аппаратом; из трех отделов груди самый маленький переднеспинка, имеющая почти квадратную форму; два остальных грудных отдела сильно удлинены, особенно средний. Что касается 10-членикового брюшка, то оно тоже сильно вытянуто или, наоборот, очень расширено и имеет листообразную форму. Вследствие этих особенностей строения тело у привиденьевых может принимать или палочковидную, или листовидную форму. Отсюда говорят о палочниках и листовидках.
В настоящее время известно около 2500 видов привиденьевых, распространенных преимущественно в тропических странах. Преобладающее большинство из них имеет палочковидную форму. Их сходство со стеблевыми частями растений достигается не только благодаря удлиненному телу. Характерно и строение их конечностей. Ноги у них обычно длинные, тонкие, с 5-члениковыми лапками и присоской между коготками. Только у калифорнийского рода Timema лапки 3-члениковые. Весьма любопытно строение бедер передних ног, которые в основной части с внутренней стороны имеют выемки (рис. 147). Потревоженное насекомое вытягивает вперед передние ноги, складывая их вместе так, что они имитируют заостренный конец сучка. При этом голова с вытянутыми вперед усиками плотно вкладывается в выемки на бедрах.
Надкрылья у палочников чаще всего укорочены или совсем отсутствуют; крылья иногда бывают хорошо развиты, веерообразной формы, иногда укорочены, а чаще тоже отсутствуют. На конце брюшка находятся небольшие одночлениковые церки удлиненной формы. Самки обычно крупнее самцов; яйцеклад у них короткий, располагается на конце брюшка снизу и не выступает за него.
Установлено, что решающее влияние на окраску палочников оказывают световые раздражения, поступающие в определенную область глаза.
В некоторых случаях у палочников наблюдается очень грубая скульптура покровов, причем на теле могут быть острые шипообразные выросты. Все это, вместе взятое, — окраска, конфигурация тела и скульптура покровов — создает огромное разнообразие приспособительных форм, которые оказываются специфичными у разных видов.
Так, гигантский индонезийский палочник (Cyphocrania gigas) имитирует крупную ветку (табл. 28, 1). Многие виды напоминают сучки или стебли кустарников и травянистых растений, куски коры, покрытые лишайниками, или даже мох.
Для сидящего в криптической (защитной) позе палочника характерна так называемая каталепсия, при которой придатки тела находятся в состоянии «восковой гибкости». В это время насекомому можно придать любую неестественную позу, например поставить головой вниз, причудливо изогнув брюшко и ноги, и оно на продолжительное время останется в таком состоянии (рис. 147, 5). Даже ампутация ноги или члеников брюшка не выведет его из каталепсии.
У некоторых палочников наряду с защитной окраской, маскирующей все видимые части неподвижно сидящего животного, наблюдается и демонстративная на тех частях тела, которые в покое скрыты (рис. 148). Ярко окрашенными оказываются части веерообразно складывающихся задних крыльев, находящихся под прикрытием надкрылий. В тех случаях, когда надкрылья полностью или почти полностью отсутствуют, та часть заднего крыла, которая в сложенном состоянии находится сверху, образует особую крыловую пластинку, имеющую покровительственную окраску, и, таким образом, выполняет защитную фунцию надкрылий.
В ряде случаев окраска может носить отпугивающий характер, как это имеет место у обитающего на островах Новые Гебриды палочника Cnipsus rhachis. Этот вид имеет крайне укороченные надкрылья и крылья и не способен к полету. В спокойном состоянии он совершенно незаметен благодаря серо-бурой окраске покрытого шипами тела. Но стоит только потревожить это насекомое, как оно сразу принимает угрожающую позу, поднимая свои маленькие надкрылья и демонстрируя крошечные задние крылья, окрашенные в яркий карминный цвет.
Среди привиденьевых особое место занимают листовидки, составляющие особое, хотя и немногочисленное, но очень оригинальное семейство Phylliidae, в своем распространении не выходящее за пределы тропиков.
У листовидок широкое, уплощенное тело, а форма и жилкование надкрылий у самок таковы, что делают их похожими на лист растения (табл. 28, 3). Это защитное сходство усиливается благодаря приспособительным окраскам и движениям. У цейлонской листовидки (Phyllium crurifolium) окраска надкрылий изменчива, вследствие чего разные самки напоминают листья различных оттенков — красновато-желтые, рыжевато-бурые или ярко-зеленые. Иногда листовидки висят под веткой или черешком листа, прицепившись к ним только двумя или тремя ногами. При этом они медленно вращают свое тело то в одну, то в другую сторону, вследствие чего становятся похожими на висящий «на ниточке» лист, колеблющийся от дуновения ветра и готовый вот-вот упасть.
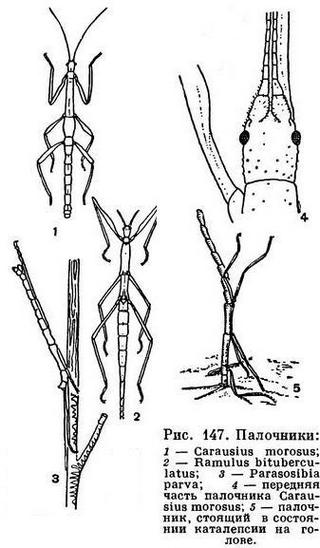
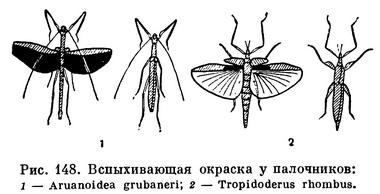
ОТРЯД ПРЯМОКРЫЛЫЕ (ORTHOPTERA)
Среди насекомых с неполным превращением прямокрылые занимают особое место. Это очень крупная группа, включающая свыше 20 000 видов, из которых более 700 встречаются в СССР. Прямокрылые широко распространены по всему земному шару — от тропиков до Заполярья — и отличаются большим разнообразием морфологических структур и физиологических приспособлений к весьма разнообразным условиям среды. Однако не только этим привлекают к себе внимание прямокрылые насекомые. С давних времен человек узнал среди них многих врагов, беспощадно уничтожающих плоды его труда. Слово «саранча» до сих пор вызывает представление о большой надвигающейся опасности.
К прямокрылым относят насекомых с удлиненным телом, грызущими ротовыми органами и характерным строением груди, летательного аппарата и задних конечностей. Голова у них с крупными, обычно овальными, сложными глазами и большей частью 3 глазками; находящиеся на ней усики могут быть длинными, превышающими длину тела (кузнечиковые, сверчковые), или короткими — короче половины тела (триперстовые, саранчовые). На этом различии в строении усиков основано деление прямокрылых на два подотряда — длинноусых и короткоусых.
Специфично строение груди прямокрылого: в ней сильно развита и подвижна переднегрудь, причем боковые части переднеспинки свешиваются вниз, образуя широкие лопасти, прикрывающие переднегрудь с боков. Остальные два отдела груди плотно слиты друг с другом. Крылья по большей части развиты нормально, хотя есть формы с укороченными крыльями и даже совсем бескрылые. Крылья передней пары более плотные и узкие и представляют собой надкрылья. Задние крылья, или просто крылья, широкие, перепончатые с хорошо развитым продольным жилкованием. При посадке насекомого они веерообразно складываются и прикрываются надкрыльями. Задние ноги прыгательного типа с утолщенными и удлиненными бедрами и длинными голенями. Поэтому прямокрылых иногда называют прыгающими насекомыми (Saltatoria). Брюшко 10-члениковое, удлиненное, с церками; снизу оно кажется 8- или 9-члениковым, так как один или два стернита оказываются редуцированными. Прямокрылые могут издавать и воспринимать звуки, так как имеют особые звуковые и слуховые аппараты, структура которых различна в разных подотрядах.
ПОДОТРЯД ДЛИННОУСЫЕ ПРЯМОКРЫЛЫЕ (DOLICHOCERA, ИЛИ ENSIFERA)
Длинноусые прямокрылые отличаются прежде всего длинными щетинковидными антеннами, превышающими обычно длину тела; орган слуха, в тех случаях когда он имеется, расположен на голенях передних ног; яйцеклад всегда длинный. В подотряде различают два надсемейства — кузнечиковых и сверчковых.
Кузнечиковых легко отличить от остальных прямокрылых по строению их лапок и характерной форме яйцеклада самки: лапки на всех ногах у них 4-члениковые, а яйцеклад длинный, обычно сжатый с боков, изогнутый в виде серпа или сабли или же мечевидный. Надкрылья и крылья у кузнечиковых большей частью развиты хорошо и тогда достигают вершины брюшка или даже заходят за него. При этом в состоянии покоя надкрылья сложены по длине тела крышеобразно и прикрывают спрятанные под ними крылья. У некоторых видов надкрылья и крылья в разной степени укорочены, есть и совершенно бескрылые формы.
Звуковой аппарат у кузнечиковых располагается на надкрыльях. В основании правого надкрылья находится так называемое «зеркальце» в виде округлой тонкой прозрачной перепонки, окруженной толстой жилкой, образующей рамку. На левом надкрылье тоже имеется перепоночка в виде зеркальца, но оно обычно непрозрачное, матовое и довольно плотное (рис. 149). Окружающая его жилка толстая и снабжена на нижней стороне надкрылья мелкими зубчиками. Эта характерная жилка называется стридуляционной; она выполняет роль смычка, в то время как «зеркальце» служит при стрекотании резонатором. Стрекотание осуществляется следующим путем. Левое надкрылье в спокойном состоянии всегда лежит над правым. Во время стрекотания кузнечик несколько приподнимает и раздвигает надкрылья, а затем приводит их в вибрирующее движение из стороны в сторону, в результате чего зубчики «смычка» трутся о рамку «зеркальца», находящегося на правом надкрылье. Благодаря «зеркальцу» звук значительно усиливается и потому слышен издалека. У каждого вида кузнечиков издаваемые им звуки специфичны. В преобладающем большинстве случаев звуковой аппарат имеется только у самцов, однако есть виды кузнечиков, у которых стрекочут и самки.
Слуховой аппарат находится на голенях передних ног; снаружи его легко обнаружить благодаря двум овальным перепонкам, расположенным по обеим сторонам голени и выполняющим функцию барабанных перепонок (рис. 149). В одних случаях эти перепонки открыты, хотя и помещаются в небольшом углублении; у большинства же видов они прикрыты хитиновыми покрышками, так что снаружи видны только две продольные щели, ведущие к перепонкам. Внутренняя часть слухового аппарата имеет сложную структуру, состоящую из окончаний нервов, чувствительных клеток, мышц и двух ветвей трахей, из которых каждая подходит к своей барабанной перепонке. Благодаря давлению воздуха в трахеях перепонки всегда натянуты.
В летнее время стрекотание кузнечиков раздается повсюду: на лугах, опушках лесов, лесных полянах и в садах. Оно не смолкает даже вечером и ночью. Однако обнаружить самих стрекочущих кузнечиков далеко не такая уж легкая задача: слишком хорошо они оказываются замаскированными в зелени кустов или в толще густой травы. Эту маскировку обусловливает сочетание формы тела и его защитной окраски. Виды, живущие в траве, часто обладают вытянутым телом и длинными узкими надкрыльями. Распространенный на Малайском архипелаге кузнечик Elimaea poaefolia имеет настолько удлиненное тело, что напоминает стебель растения, на котором он сидит. Среди тропических кузнечиков большое количество видов обнаруживает сходство с листьями деревьев. Это сходство достигается благодаря сильному расширению надкрылий, а также их специфическому жилкованию. В зависимости от окраски они имитируют или здоровые, или отмирающие и мертвые листья (рис. 150). Так, например, на листоподобных крыльях Cycloptera elegans имеются бурые пятна, напоминающие повреждения листьев паразитическими грибами. Еще более своеобразна форма и окраска надкрылий у видов Tanusia: имеющаяся на них пятнистость имитирует начало разложения листа, а неровные края создают впечатление, что лист объеден или обломан. Сильно объеденными кажутся листоподобные крылья у акридоксены (Acridoxena hewaniana). Некоторые индо-малайские кузнечики, живущие на деревьях, весьма успешно имитируют лишайники. Так, например, яванская сатрофилия (Satrophyllia femorata), сидя неподвижно с вытянутыми вперед усиками и передними ногами на ветке дерева, совершенно сливается с общим фоном покрывающих эту ветку лишайников.
Хотя у кузнечиков, так же как у всех прямокрылых, задние ноги прыгательного типа, т. е. с утолщенными бедрами, обычно они передвигаются по веткам или в траве, быстро ползая при помощи всех шести ног. Если же кузнечика потревожить, он упирается концами голеней задних ног, а затем быстро распрямляет ноги и делает прыжок вперед. Крылатые формы при этом могут распускать крылья, которые в данном случае служат парашютом и позволяют значительно увеличить прыжок.
Кузнечиковые представляют собой группу древнего происхождения. Их предками, по-видимому, были так называемые первичные прямокрылые (Protorthoptera), существовавшие в каменноугольном периоде. Они имели много черт сходства с кузнечиковыми, отличаясь от них уплощенным телом и задними конечностями непрыгательного типа. В пермском периоде появляются уже типичные представители кузнечиковых. В третичном периоде формируются некоторые современные роды, такие, как зеленые кузнечики (Tettigonia), серые (Decticus) и скачки (Platycleis).
В настоящее время около 70% кузнечиковых распространено в тропических и субтропических странах. На территории Советского Союза обитает свыше 170 видов, относящихся к 3 семействам.

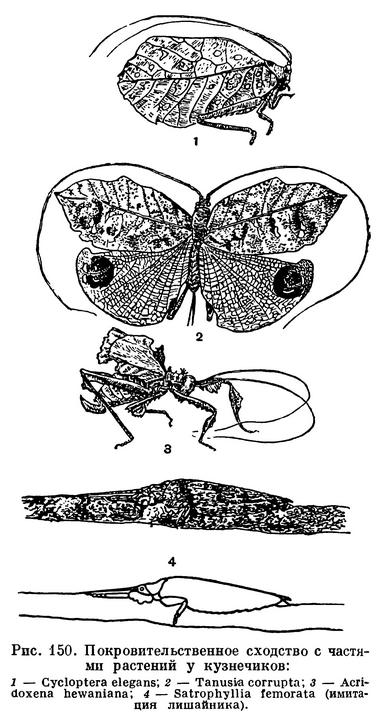
Сверчковые, в отличие от кузнечиковых, имеют 3-члениковые лапки, длинные гибкие церки и тонкий прямой яйцеклад (у самок), который на конце обычно копьевидно расширен. Только у медведки (сем. Gryllotalpidae) яйцеклад отсутствует.
Надкрылья у сверчковых в покое лежат плоско на спинной поверхности тела, причем левое всегда прикрыто правым. С этим связаны особенности звукового аппарата, который хотя и сходен по структуре с звуковым аппаратом кузнечиков, но отличается от последнего большей сложностью и иным расположением компонентов: стридуляционная жилка, выполняющая функцию смычка, располагается на правом надкрылье, а жилка, о которую она трется, воспроизводя звук, — на левом. Вообще весь этот аппарат развит сильнее и занимает значительно большую площадь на надкрыльях, чем у кузнечиков. Имеется он только у самцов, причем у каждого вида он отличен по строению и воспроизводимым звукам. Различают 6 типов стрекотания сверчков: призывное, брачное, стрекотание после спаривания, агрессивное стрекотание при встрече самцов, стрекотание, связанное с внезапным исчезновением самки, и у некоторых видов — «опознавательный» сигнал. Однако «репертуар» вида состоит обычно из меньшего количества песен. Наиболее древний тип песни, вероятно, брачное пение.
У некоторых видов сверчковых как надкрылья, так и крылья могут недоразвиваться или даже совсем отсутствовать. Нередки случаи, когда степень развития летательного аппарата неодинакова у различных особей одного и того же вида и пола. В тех случаях, когда задние крылья развиты нормально, они складываются веерообразно и в покое в виде жгутов выдаются из-под надкрылий.
Слуховой аппарат сверчковых такого же типа, как у кузнечиков, и также располагается на голенях передних ног.
В СССР встречается около 45 видов сверчковых, большинство которых относится к семейству сверчков (Gryllidae). Представителем этого семейства может служить широко распространенный в центральных и южных частях Европы, в Средней Азии и в Северной Африке полевой сверчок (Gryllus campestris). Это один из самых крупных наших видов, длиной 2—2,6 см. Он черного цвета с бурыми надкрыльями, при основании которых находится по одному оранжевому пятну; бедра задних ног внизу ярко-рыжие. На юге его стрекотание можно слышать уже в начале лета. Поющий самец сидит у входа в свою норку; если его потревожить, он сейчас же прячется в ней. Норка представляет собой наклонный ход, не шире пальца и примерно в палец длиной. Вход в норку закрыт пучком травы. Если к норке подходит другой самец, то между ними начинается драка. Оба бросаются друг на друга, ударяются своими толстыми головами, и каждый старается укусить своего противника. Побежденный противник поедается победителем, несмотря на то что обычно полевой сверчок питается растительной пищей.

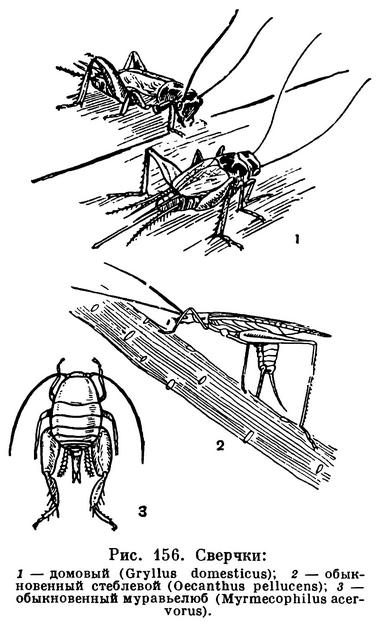
Подземный образ жизни ведут медведки (семейство Gryllotalpidae), которые резко отличаются от всех остальных сверчковых очень большой переднеспинкой, сравнительно короткими усиками, лишь немного заходящими за переднеспинку, и сильно измененными передними ногами, отлично приспособленными к копанию и передвижению в земле. По совершенству приспособления передняя конечность медведки не уступает передней ноге крота; отсюда происходит латинское название этого животного: Gryllotalpa — «сверчок-крот». В копательной конечности медведки сильно расширены бедро и голень, лапка же укорочена и прикрепляется сбоку голени, на которой имеется 4 черных зубца. Из встречающихся в СССР 3 видов медведок широко распространена в Палеарктике обыкновенная медведка (Gryllotalpa glyllotalpa). Ее крупное тело, длиной 3,5—5 см, сверху темно-бурого цвета, а снизу буро-желтое, кажется шелковистым благодаря густому покрову из мелких золотистых волосков; крылья развиты нормально и в спокойном состоянии выступают из-под укороченных надкрылий в виде загнутых книзу жгутов.
В естественных условиях медведка селится обычно в поймах рек, где почва всегда достаточно увлажнена. Однако ее часто можно находить на огородах и в садах, и тут она приносит большой вред, повреждая корневую систему многих культурных растений, в том числе кукурузы, картофеля, огурцов, а в Средней Азии — хлопка и риса.

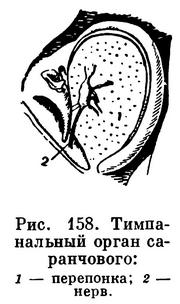
ПОДОТРЯД КОРОТКОУСЫЕ ПРЯМОКРЫЛЫЕ (BRACHYCERA, ИЛИ CAELIFERA)
В отличие от длинноусых прямокрылых представители этого подотряда обладают относительно короткими антеннами, никогда не достигающими длины тела. В подотряде различают два надсемейства — триперстовых и саранчовых.
Триперстовыми называют мелких насекомых, длиной 4—9 мм, напоминающих по внешнему виду маленьких медведок. Это сходство выражается в однотипной конфигурации тела и в строении передних ног, которые, так же как у медведок, приспособлены к копанию. Однако достаточно посмотреть на этих насекомых немного повнимательней, чтобы убедиться в том, что триперсты относятся к другой систематической группе, а именно к короткоусым прямокрылым. Об этом свидетельствует строение головы, которая несет четковидные усики, состоящие всего из 10 члеников.
Надкрылья у триперстов короткие, роговые; крылья могут быть или длиннее, или короче надкрылий, с непрозрачным передним краем и многочисленными радиально расходящимися жилками. Весьма своеобразно строение задних ног: они прыгательные, с широкими уплощенными бедрами и тонкими, слегка изогнутыми голенями. По обеим сторонам голени располагаются шипики, имеющие вид лопастинок, а на ее вершине сидят две короткие и две длинные шпоры. Лапка задней ноги состоит всего из одного более или менее вытянутого пальцевидного членика, располагающегося между длинными шпорами голени, что и дало основание называть этих насекомых «триперстами». Брюшко 9—10-члениковое, несет на вершине две пары удлиненных придатков, из которых верхняя, 2-членистая, представляет собой церки. У триперстов нет ни звукового, ни слухового аппарата; у самок наших видов отсутствует также и яйцеклад.
В СССР встречается всего 4 вида, относящихся к одному роду (Tridactylus), распространенному преимущественно в субтропиках. Наши виды известны с Кавказа, из Средней Азии и Казахстана, а также из степных районов Европейской части СССР и Западной Сибири. Обитают они обычно на песчаных берегах рек и других водоемов. В солончаковых пустынях встречаются около западин, где долго сохраняется вода.
Примером может служить обыкновенный триперст (Tridactylus variegatus), широко распространенный в степях и пустынях Евразии. Его можно встретить на песчаных отмелях, где с помощью передних ног и челюстей он выкапывает в сыром песке норки. В таких условиях триперстов можно находить иногда в массе. Они быстро перепрыгивают с одного места на другое на освещенной солнцем поверхности песка или в разреженном травянистом покрове. Триперсты — хищники, питающиеся мелкими насекомыми и дождевыми червями.
Саранчовые составляют самую многочисленную группу среди прямокрылых. От кузнечиков и сверчков они легко отличаются короткими усиками, не превышающими длины половины тела, коротким яйцекладом самки и весьма специфичными органами звука и слуха. В отличие от триперстовых у саранчовых лапки задних ног 3-члениковые, число члеников усиков может доходить до 28, церки не членистые, в виде конических выступов, а у самки всегда имеется яйцеклад.
Подобно кузнечикам и сверчкам саранчовые — признанные музыканты в мире насекомых. Чтобы убедиться в этом, достаточно летом выйти на любой луг или опушку леса; в дневное время их стрекотание явно забивает «пение» кузнечиков в силу того, что одновременно может стрекотать сразу большое число экземпляров.
Звуковой аппарат саранчовых располагается на бедрах задних ног и надкрыльях. Обычно вдоль внутренней поверхности бедра тянется длинный ряд бугорков или головчатых шишечек, а одна из жилок надкрылья оказывается толще остальных. Быстро двигая бедром, насекомое проводит этими бугорками по жилке, в результате чего раздаются отрывистые стрекочущие звуки. Эти звуки можно воспроизвести даже на мертвом насекомом, если быстро двигать его бедра, прижимая их к надкрыльям. Расстояние между бугорками может быть неодинаковым: у некоторых видов бугорки у основания бедра располагаются чаще, чем в его верхней части. Этим достигается разная высота издаваемого звука. В ряде случаев, как это имеет место у трескучей огневки (Psophus stridulus), на бедре находится гладкий кант, а бугорки располагаются на передней жилке крыла. Этот вид и некоторые другие могут издавать на лету звуки иного характера. При полете они громко трещат, воспроизводя звук, напоминающий треск деревянной трещотки. Механизм этого звучания не совсем ясен, однако, очевидно, он связан с особенностями жилкования крыльев: обычно у «трещоток» сильно уплощены несколько продольных жилок крыльев.
Тимпанальные органы помещаются у саранчовых по бокам первого брюшного сегмента, позади находящегося на нем дыхальца (рис. 158). Они представляют собой округлые или щелевидные отверстия, затянутые прозрачной барабанной перепонкой. Барабанная перепонка обрамлена хитиновым утолщением, слегка налегающим на нее спереди и сзади. Изнутри к перепонке прилегает большой воздушный мешок трахейной системы, который поддерживает ее в натянутом состоянии. По-видимому, в натяжении перепонки принимают участие также две специальные мышцы.
У саранчовых форма тела и окраска его покровов находится в очень тесном соответствии с поведением насекомого и отражает особенности той среды, в которой формировался и обитает в настоящее время тот или иной вид. Такое сочетание морфологических, биологических и физиологических свойств вида, находящееся в соответствии с окружающей средой, называют жизненной формой (табл. 30). Жизненные формы саранчовых могут быть объединены в два класса: обитатели растений, или фитофилы, и обитатели открытых участков на поверхности почвы, или геофилы.
Среди фитофилов различают хортобионтов и тамнобионтов. Настоящие хортобионты (табл. 30, 1—3) обитают в толще травянистого покрова и питаются преимущественно злаками. У них стройное удлиненное тело, нередко со скошенным лбом; соотношение между шириной и высотой тела в наиболее широкой его части (индекс ш/в) всегда ниже единицы. В некоторых случаях тело может быть настолько сильно удлинено и к тому же лишено крыльев, что приобретает палочковидную форму, как это имеет место у южноамериканской палочковидной кобылки (Cephalocoema lineata). Покровы тела хортобионтов гладкие, лишены грубой скульптуры и окрашены под цвет зеленой или сухой травы, часто с продольными полосками вдоль тела. Крылья обычно бесцветные, реже окрашены в бледные розоватые или зеленоватые тона. Хортобионты широко распространены на лугах, в степях и саваннах, т. е. там, где преобладают травянистые растения. Типичным представителем хортобионтов может служить степной конек (Euchorthippus pulvinatus), широко распространенный в ковыльных степях Европы и Азии. Среди хортобионтов многие виды являются вредителями сельского хозяйства.
Некоторые хортобионты предпочитают питаться не злаками, а широколиственными травянистыми растениями; их так и называют травоядными хортобионтами. Сюда относятся виды бескрылых кобылок конофим (Conophyma, табл. 30, 4), широко распространенных в горах Средней Азии, и целый ряд других форм. У травоядных хортобионтов туловище более коренастое, а голова с отвесным лбом. Характерно, что у них верхняя челюсть приспособлена к перетиранию и пережевыванию мягких листьев.
В ряде случаев встречаются виды, которые хотя и обитают в толще травы, но не избегают и открытых пространств на поверхности почвы, как это делают некоторые крестовички (Dociostaurus). Такие саранчовые называются факультативными хортобионтами. По своему поведению и форме тела они являются переходными к геофилам.
В отличие от хортобионтов, связанных с травянистой растительностью, тамнобионты (табл. 30, 5,6) живут на кустарниках и деревьях. Приспособления к такому образу жизни прежде всего выражаются в специфичной структуре конечностей: у них наблюдается асимметрия в вооружении голеней задних ног, на внутренней стороне которых шипы длиннее шипов наружного ряда; асимметричными могут быть и находящиеся на лапках коготки, между которыми находится сильно развитая присоска. Некоторые тамнобионты тесно связаны с определенными видами растений. Например, большая саксауловая горбатка (Dericorys albidula), как и некоторые другие виды этого рода, связана с саксаулом. В песчаных пустынях Средней Азии в годы массового размножения этого насекомого оно может сильно повреждать саксаул.
Второй класс жизненных форм — геофилы — заключает в себе саранчовых, обитающих обычно на поверхности почвы, забирающихся на растения только во время питания. В нем различают открытых геофилов и скрытоживущих геофилов (герпетобионтов). Открытые геофилы (табл. 30, 7,8), в отличие от хортобионтов, имеют более уплощенное тело с индексом ш/в, поднимающимся до единицы и выше; у них всегда слабо развиты, а иногда и совсем отсутствуют присоски между коготками. Покровы тела у открытых геофилов плотные, морщинистые, часто покрыты бугорками. Окраска их тела и надкрылий всегда защитная и соответствует тому общему фону, на котором они встречаются. Задние крылья нередко ярко окрашены, с темными, бросающимися в глаза перевязями. Такое сочетание окрасок надкрылий и крыльев стоит в тесной связи с поведением открытых геофилов. Можно подойти совсем близко к сидящему насекомому и не заметить его, так хорошо оно замаскировано. И вдруг совершенно неожиданно оно вылетает прямо из-под ног, привлекая к себе внимание яркими флагами раскрытых крыльев. Быстро перелетев на некоторое расстояние, насекомое так же внезапно садится на землю и вновь исчезает из глаз. Некоторые геофильные виды способны во время полета издавать трескучие звуки; так делают различные трещотки (Bryodema, Angaracris), трескучая огневка, некоторые пустынницы (Sphingonotus) и др. Летящие ярко окрашенные геофильные саранчовые, привлекают к себе внимание движением, окраской и звуком, но, как только садятся, они исчезают из глаз преследователя, явно дезориентируя его.
Высокая криптичность окраски наружных покровов легко объясняется явлением гомохромии, выражающимся в том, что при развитии личинок одного и того же вида на различных фонах их окраска будет сильно варьировать и в конечном счете в каждом конкретном случае в той или иной мере соответствовать фону. Это доказано специальными экспериментами, проводившимися с разными саранчовыми, и в том числе с голубокрылой кобылкой (Oedipoda coerulescens). Воспитанные в условиях опыта экземпляры этого вида имитировали окраску почвы, на которой они выросли, приобретая соответственно черный, красноватый или серый оттенок.
Открытые геофилы преимущественно обитатели пустынь и полупустынь; представители этой группы, встречающиеся в степях или в лесной зоне, предпочитают открытые, хорошо прогреваемые места с разреженной растительностью на обнажениях горных пород, солончаках, галечниках по берегам рек и т. д.
Среди открытых геофилов особо отличают петробионтов и псаммобионтов. Петробионты, или камнелюбы, обитают на каменистых участках, чаще всего на склонах гор. У них более резко, чем у остальных геофилов, выражена плотность и скульптурность покровов. Тело очень широкое и укороченное, вальковатое; переднеспинка обычно высокая, в ряде случаев с сильно приподнятым килем, как это имеет место, например, у саксетаний (Saxetania, табл. 30, 9), широко распространенных в северных горах Ирана и Афганистана, а у нас в Копет-Даге и некоторых других горных системах Средней Азии. У петробионтов обычно редуцирован летательный аппарат, а движения очень замедленны. Зато криптическая окраска выражена очень хорошо и всегда отражает специфику фона каменистых склонов.
Иные приспособления у псаммобионтов (табл. 30, 10,11) — обитателей песчаных массивов, особенно ярко представленных в песчаных пустынях. К ним в первую очередь нужно отнести виды песчанок (Hyalorrhipis) и тонкошпоров (Leptopternis). Их окраска хорошо имитирует общий тон и специфичную зернистость песчаного субстрата. Своеобразны их приспособления к своему местообитанию: у псаммобионтов сильно удлинены шпоры на голенях задних ног, что облегчает отталкивание на сыпучем песке припрыжке.
В отличие от открытых геофилов герпетобионты (табл. 30, 12,13) представляют собой формы, обитающие на поверхности почвы, занятой разреженным травянистым покровом, опадом листвы или другими растительными остатками. Их можно также встретить на увлажненном песке по берегам водоемов. Они очень требовательны к повышенному увлажнению. Обычно это мелкие формы с веретеновидным телом, суженным от заднегруди вперед и назад, окрашенным в защитные, часто сероватые тона. Типичными представителями герпетобионтов являются виды специфичного семейства прыгунчиков, или тетригид (Tetrigidae).
Все распространенные в СССР саранчовые имеют в году одну генерацию, причем большинство из них зимует в фазе яйца. Однако существуют виды, дающие в течение года 2—3 поколения. Для саранчовых, так же как и для других прямокрылых, характерно сперматофорное оплодотворение; сперматофоры имеют вид или пузыревидных резервуаров с длинной выводной трубчатой частью, или округлых баллонов. При спаривании, которое продолжается в течение нескольких (до 20) часов, самец вводит сперматофоры со спермиями в половые пути самки.

Всем саранчовым в той или иной мере свойственны различного типа миграции, выражающиеся в виде переходов или перелетов. Хортобионты регулярно совершают вертикальные миграции, передвигаясь по растениям вверх и вниз в утренние и вечерние, а нередко и в дневные часы суток. Геофилы все время совершают днем горизонтальные миграции, перелетая с одного места на другое. Мигрируют саранчовые и с участков, где они кормятся, в места, удобные для откладки яиц. Однако миграционные инстинкты особенно развиты у стадных видов, у которых активные переходы и перелеты осуществляются кулигами и стаями на большие расстояния. Как личинки, так и взрослые саранчовые ночуют обычно на верхних частях растений, где они пребывают в ночное время в состоянии холодового оцепенения. После восхода солнца по мере прогрева воздуха личинки становятся активными и концентрируются на освещенных участках почвы или иногда на растениях, образуя так называемые «солнечные кулижки». Это плотные скопления неподвижно сидящих личинок, в которых каждая особь тесно прижимается к соседним. Первичные «солнечные кулижки» образуются личинками, отрождающимися из одной кубышки. Как только температура тела находящихся совместно особей поднимается выше 40°С, кулижки рассеиваются и начинается пешая миграция, которая прекращается вечером, когда личинки опять впадают в состояние покоя. В жаркое время дня, в случае если температура тела насекомого поднимается выше 50°С, наступает дневной период тепловой депрессии, в котором также наблюдается состояние относительного покоя.
В годы массового размножения образуются настолько крупные кулиги, что занимаемая ими площадь может достигать размеров сотен и тысяч гектаров. В период личиночного развития кулиги все время находятся в состоянии миграционной активности, передвигаясь с мест отрождения в сторону участков с более благоприятными условиями температуры, влажности и питания. В результате за период личиночной жизни стадные саранчовые могут проходить очень большие расстояния — до 30 км и больше, как это имеет место, например, у азиатской саранчи. Характерно, что движение кулиги имеет определенное направление и осуществляется обычно в сторону, противоположную направлению ветра. Оптимальные условия для движения создаются при небольшом ветре (3—5 м/сек); при порывистом или сильном ветре, превышающем 10 м/сек, движение кулиг прекращается.
После окрыления саранчи ее передвижения принимают вид миграционных полетов, когда стаи перелетают за день расстояние в несколько десятков километров. Характерно, что направление движения стаи стоит в очень тесной связи с направлением ветра. Так, при слабом ветре (до 1 м/сек) стаи азиатской саранчи летят обычно по ветру; при повышении силы ветра до 5 м/сек саранча взлетает с большим трудом, а сам полет осуществляется под острым углом к направлению ветра. Полеты могут осуществляться не только днем, но в некоторых случаях и ночью, однако скорость полета при этом резко снижается: днем азиатская саранча пролетает 30—40 км, а ночью не больше 15 км. За весь период движения стая может улететь от своего гнездилища за 200—300 и больше километров. Еще дальше залетает пустынная саранча: в 1954 году небольшие стаи этого вида из Северо-Западной Африки достигли Британских островов, пролетев более 2400 км над открытым морем. Значительно меньшая миграционная способность у итальянского пруса и мароккской саранчи, которые отлетают от мест отрождения на более короткие расстояния, измеряемые десятками километров.
Виды саранчовых, имеющие широкий ареал, охватывающий различные ландшафтные зоны, в разных частях области своего распространения заселяют разные места обитания (стации). Выявлена следующая закономерность: такие виды в северной части своего ареала населяют более сухие (ксерофитные) стации, по мере продвижения на юг они последовательно перемещаются в более влажные места обитания — сначала в мезофитные (со средним увлажнением), а затем в гигрофитные (сильно увлажненные). Так, например, перелетная саранча в центрально-черноземной области и на юге Западной Сибири обитает на песчаных участках, а в пустынях Средней Азии и Казахстана — на побережьях рек, озер и морей в зарослях тростника. Итальянский прус на юге лесной зоны поселяется на меловых обнажениях с разреженным растительным покровом, а в Средней Азии — в долинах рек, оазисах и предгорьях, почему его и называют там оазисным прусом. Это явление получило название принципа зональной смены стаций (принцип Г. Я. Бей-Биенко). Оно объясняется в основном спецификой температурного режима и режима влажности в различных зонах.
Преобладающее большинство саранчовых — обитатели жарких стран: свыше 50% известных видов населяют тропики и субтропики. В СССР известно до 485 видов, относящихся к 5 семействам.
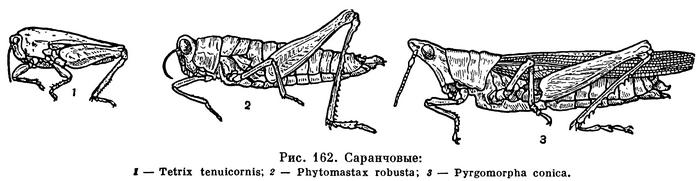
Настоящих саранчовых (семейство Acrididae) легко отличить от всех рассмотренных выше семейств. От тетригид и эумастацид они прежде всего отличаются наличием звукового аппарата, помимо отсутствия тех признаков, которые специфичны для этих семейств. В то же время у них совершенно иная, чем у памфагид и пиргоморфид, скульптура наружной поверхности бедер задних ног — в виде правильно расположенных между килями перистых площадок. Это семейство богато видами: в фауне СССР на его долю приходится около 80% известных в настоящее время видов.
Форма тела у настоящих саранчовых сильно варьирует в зависимости от принадлежности вида к той или иной жизненной форме. По существу, в этом семействе выражены почти все жизненные формы, исключая только герпетобионтов и петробионтов.
Среди распространенных в СССР видов свыше 100 в той или иной мере могут вредить сельскохозяйственным культурам. Из них наиболее опасны стадные формы — перелетная саранча, мароккская саранча, прус итальянский и туранский, а также залетающая в некоторые годы из Ирана и Афганистана пустынная саранча (табл. 31, 1—6).
Стая взрослой саранчи летит со скоростью 10—15 км в час и за день пролетает 80—120 км. Поэтому, несмотря на ограниченность размеров постоянных гнездилищ, азиатская саранча может распространяться на большие расстояния и в течение нескольких лет подряд жить в новых заселенных ею местах, проходя здесь свойственное ей развитие. Из всех наших саранчовых азиатская саранча самая прожорливая. Подсчитано, что каждый экземпляр ее в течение своей жизни съедает 300 г зеленого корма. Потомство одной самки саранчи за лето уничтожает столько корма, сколько хватило бы для двух овец. Кулиги и стаи этого насекомого в течение 1—2 часов могут уничтожить сотни и тысячи гектаров посевов. Излюбленной пищей азиатской саранчи в местах ее отрождения является тростник. Во время движения она вредит многим растениям, предпочитая культурные и дикорастущие злаки. Чем старше саранча, тем она становится более многоядной и, помимо злаков, может наносить большой вред капусте, арбузам, подсолнечнику и другим растениям.
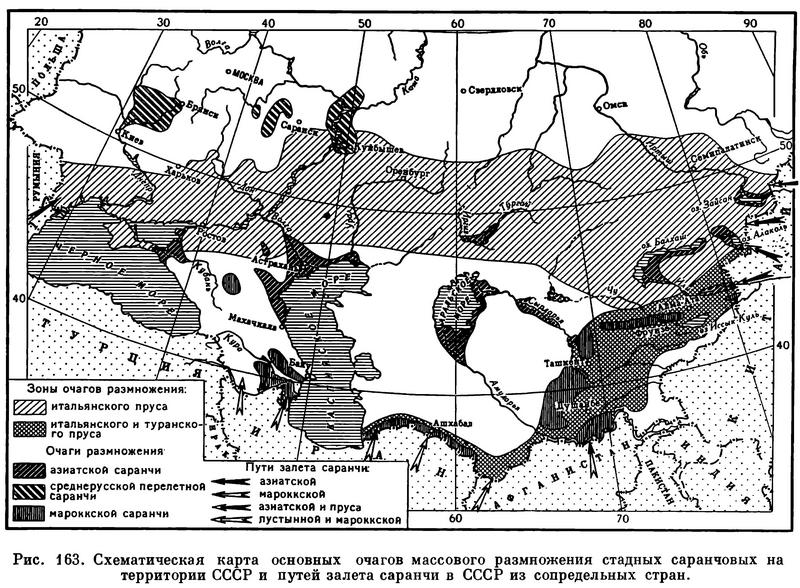
ОТРЯД УХОВЕРТКИ (DERMAPTERA)
Уховертки отличаются весьма своеобразным обликом. Это сравнительно небольшие насекомые, длиной 4—40 мм, с сильно уплощенным и удлиненным очень гибким телом, несущим на вершине брюшка два длинных хитинизированных отростка — клещи. Голова уховертки почти сердцевидной формы с направленными вперед жующими ротовыми органами; находящиеся на ней нитевидные усики у разных видов имеют различное число члеников — от 8 до 50. Наружные покровы тела всегда очень плотные, сильно хитинизированные.
Одной из характерных особенностей строения уховерток является их летательный аппарат. Передняя пара крыльев у них сильно укорочена и превращена в твердые, кожистые надкрылья. Мягкие, перепончатые задние крылья очень широкие, веерообразной формы; на их переднем крае имеется роговая пластинка. В спокойном состоянии крылья складываются наподобие веера вдоль жилок, а затем — еще два раза поперек и скрываются под надкрыльями, из-под которых наружу выступают только концы твердых роговых пластинок. Ноги обычно бегательного типа, относительно короткие, с 3-члениковыми лапками.
Относительно удлиненное брюшко состоит из 10 члеников; находящиеся на его конце «клещи» представляют собой измененные церки; они не разделены на членики и часто вооружены различными зубцами и выступами. У самцов клещи развиты значительно сильнее, чем у самок. Форма и размеры клещей очень изменчивы. В пределах одного и того же вида встречаются самцы с короткими и с длинными клещами.
Клещи, прежде всего, представляют собой орган защиты и нападения. Если потревожить уховертку, она принимает защитную позу. При этом брюшко изгибается кверху и вперед, вследствие чего концы клещей располагаются над головой и грудью. Характерно, что у некоторых видов, например у нашей обыкновенной уховертки (Forficula auricularia), двигательные мышцы клещей настолько сильно развиты, что она может проколоть ими кожу человека до крови. Клещи используются также для удерживания захваченной добычи при поедании.
Наиболее древняя форма уховерток известна из юрских отложений в горном хребте Каратау (южный Казахстан) и описана под названием Protodiplatys fortis (рис. 164). Обладая целым рядом типичных для современных уховерток признаков, в том числе характерным строением брюшка и укороченными жесткими надкрыльями, этот вид имел некоторые примитивные черты строения. Так, число члеников на лапках не три, а четыре-пять; церки не имеют форму клещей, они тонкие, короткие и к тому же поделены на членики. Нахождение данной формы в юре свидетельствует о большой древности отряда, а примитивные черты ее строения указывают на то, что она является промежуточной формой между современными уховертками и их палеозойскими тараканообразными предками, имевшими также 5-члениковые лапки задних ножек и короткие членистые церки. Интересно подчеркнуть, что последний признак (многочлениковые церки) сохранился у личинок некоторых тропических видов уховерток из подсемейства Diplatyinae, распространенного в Африке, Индо-Малайской области и Центральной Америке. Взрослые формы этих видов имеют настоящие нечленистые церки — клещи.
Современная фауна уховерток комплектуется приблизительно из 1200—1300 видов. Преобладающее большинство их распространено в тропических странах, где богатая лесная растительность в сочетании с теплым и влажным климатом создает благоприятные условия для их существования. В СССР обитает всего только 26 видов; из них широкоизвестна обыкновенная уховертка (Forficula auricularia) ржаво- или смоляно-бурого цвета с грязно-желтыми ногами и надкрыльями, длиной около 1,5 см. Это насекомое пользуется незаслуженной дурной славой. До сих пор еще можно слышать рассказы о том, что уховертка заползает в ухо спящего человека и, прорывая барабанную перепонку, выводит из строя слуховой аппарат. Все это не соответствует действительности, и боязнь вреда, якобы причиняемого этим человеку, ни на чем не основана.
Образ жизни, сходный с образом жизни обыкновенной уховертки, ведут многие представители этого отряда. Однако в ряде случаев некоторые виды уховерток характеризуются специфическими особенностями биологии.
Весьма своеобразны 2 индо-малайских вида уховерток — ариксений (Arixenia), обитающих в пещерах, заселенных летучими мышами. Благодаря отсутствию надкрылий и крыльев ариксении скорее напоминают личинок, чем взрослых насекомых. Их тело покрыто густыми волосками; глаза, как у большинства пещерных насекомых, сильно редуцированы; одночлениковые церки образуют очень несовершенные, слабые клещи. Один из видов (Arixenia esau) является наружным паразитом летучих мышей (Cheiromeles torquatus), другой (Arixenia jacobsoni) питается насекомыми, населяющими гуано летучих мышей. Вследствие своеобразия строения этих насекомых их выделяют в особый подотряд (Arixenina), противопоставляя всем остальным современным уховерткам.
Еще более оригинальны гемимеры (Hemimerida), представленные 7 видами одного рода (Hemimerus), распространенного в тропической Африке. Это уже настоящие паразиты, встречающиеся на коже хомяковой крысы (Cricetomys gambianus). У них уплощенное бескрылое тело с короткими ногами и тонкими длинными нечленистыми церками; глаза отсутствуют. Характерно, что развитие яиц происходит в теле матери, которая «рождает» довольно крупных личинок.
Положение гемимеров в системе до сих пор еще точно не определено: одни ученые относят их к уховерткам, выделяя в особый подотряд, другие же выделяют в самостоятельный отряд, отличный от отряда уховерток.

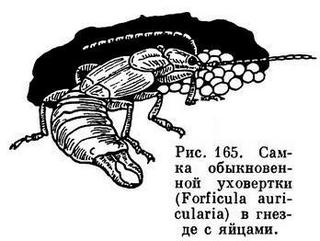
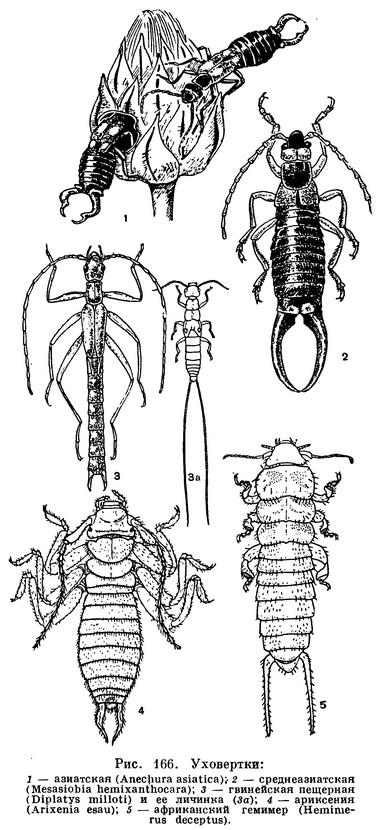
ОТРЯД ВЕСНЯНКИ (PLECOPTERA)
У веснянок, как и у поденок и стрекоз, личинки живут в воде и взрослые насекомые имеют две пары перепончатых укрепленных густой сетью жилок крыльев. Но в отличие от стрекоз и поденок они складывают крылья кровлеобразно на спине. Веснянки более родственны уховерткам и прямокрылым, чем стрекозам и поденкам.
Веснянки имеют уплощенное в спинно-брюшном направлении тело. Это сравнительно крупные насекомые с темными довольно жесткими покровами. Они хорошо бегают, если их потревожить, и бывают похожи на каких-то жуков. Задние крылья у них складываются веерообразно под защитой передних, которые, правда, лишь немного плотнее. Голова крупная с длинными нитевидными многочлениковыми усиками. Ротовые части мелкие — взрослые веснянки не питаются. Ноги у веснянок стройные, бегательные, с 3-члениковой лапкой. Брюшко 10-члениковое с парой членистых хвостовых нитей (церков) на конце.
Несмотря на то что веснянки — крупные насекомые и по берегам рек встречаются нередко, знают их далеко не все. Это связано с тем, что они редко и недолго летают, окрашены в темные тона и ведут скрытый или малоподвижный образ жизни, сидя на веточках тростника, на коре растущих у берега деревьев и в прибрежной траве.
Лёт бывает недолгим, после коротких взлетов самец подползает под самку и выделяет сперматофор, который самка тут же подхватывает подвижными придатками полового отверстия.
Яйца (рис. 168) веснянка некоторое время носит на конце брюшка, затем, летая над поверхностью воды, на короткое время погружает брюшко в воду обычно быстрых и чистых речек и ручьев. Яйца при этом быстро смываются, отделяются друг от друга и падают на дно, где застревают между камнями. Яйца веснянок овальные или имеют форму вазы и темные. У некоторых видов, например у настоящих веснянок (род Perla), яйца выводятся самкой целыми пачками, заключенными в слизистую массу.
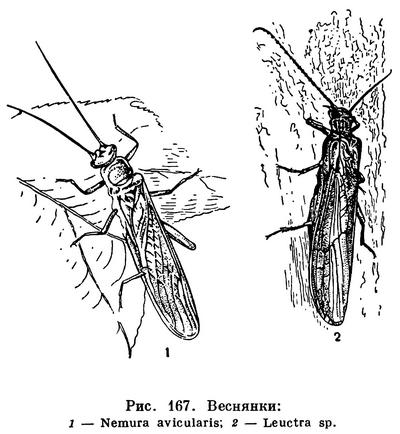
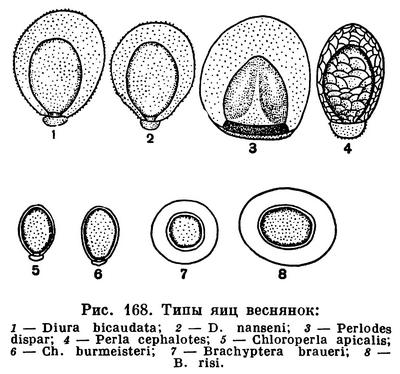
Личинки (рис. 169) по форме тела похожи на взрослых веснянок, но, конечно, сперва не имеют крыльев, зачатки которых появляются только в конце личиночного периода жизни. В отличие от взрослых веснянок личинки имеют сильные зазубренные челюсти. Личинки мелких веснянок питаются водорослями, но большинство — хищники, поедающие личинок комаров, мошек, поденок и других мелких беспозвоночных. Дыхание растворенным в воде кислородом осуществляется у них с помощью трахейных жабр, которые редко бывают листовидными (Nemura), а чаще представляют пучки пронизанных трахеями нитевидных выростов, расположенных на разных сегментах груди и брюшка. Ноги у личинок длинные и цепкие с большим количеством волосков. Личинки могут хорошо бегать по дну, неплохо плавают, но большую часть времени проводят уцепившись ногами за выступы дна, подкарауливая добычу.
Весь цикл развития у большинства веснянок продолжается год, у некоторых крупных видов в северных реках растягивается на 2—3 и даже 4 года. Зимовка происходит в стадии нимфы, имеющей уже явственные зачатки крыльев. За время личиночной жизни веснянки много раз линяют, например большая веснянка (Perla maxima), достигающая в длину 24 мм, линяет 22 раза.
Взрослые веснянки малоподвижны, у некоторых видов происходит утрата способности к полету. Интересно, что чаще не способны к полету самцы, например у появляющейся уже в феврале в предгорьях Кавказа и в Крыму черной веснянки (Capnia nigra) у самца крылья имеют вид небольших чешуек. А совсем недавно в Южной Америке в Андах были открыты такие веснянки (Megandiperla), у которых по общему облику взрослые насекомые и личинки совсем неотличимы — взрослые не имеют даже зачатков крыльев (рис. 170). Эти веснянки, в противоположность большинству других представителей отряда, ведут в стадии личинки не водный образ жизни, а наземный, ползая в подушках покрывающего скалы мха. Правда, места, где они обитают, находятся в горах, там, где ежедневно идет дождь и где мох всегда очень влажен. Интересно, что между личинками и взрослыми у этого вида сохраняется то различие, что у личинок трахейная система замкнута, хотя у них и нет наружных трахейных жабр, а у взрослых — открывается дыхальцами. В постоянно влажном воздухе для личинок веснянок более благоприятны условия снабжения кислородом, чем в водоемах, благодаря чему возможно кожное дыхание и без жаберных выростов за счет только поверхности тела. В воде тоже встречаются мелкие личинки веснянок, не имеющие жаберных нитей. У личинок крупных видов, таких, как большая веснянка, сперва имеются только слабые зачатки жабр (у большой веснянки в основании третьей пары ног), постепенно увеличивающиеся и в числе, и в размерах по мере линек. Любопытно, что рудименты жабр бывают и у некоторых взрослых веснянок.
Веснянки представлены 5 семействами, включающими немногим более 2000 видов, часть которых широко распространена, часть обитает только в северном, а часть — только в южном полушарии. В СССР известно около 200 видов.
Веснянки — древняя группа, ископаемые веснянки известны еще из пермских отложений, в которых встречались и родственные веснянкам Protopelaria.
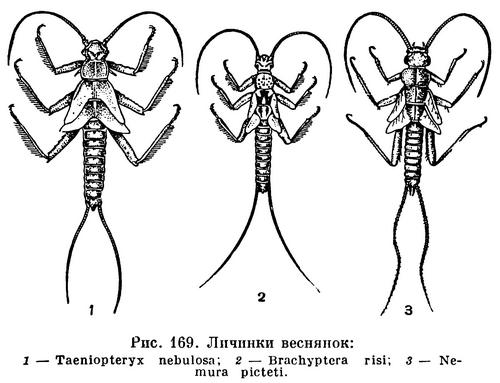
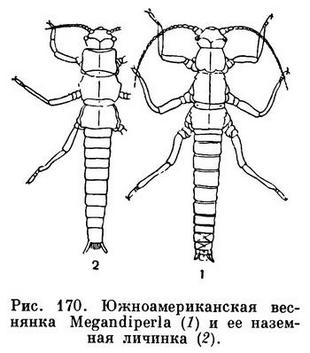
ОТРЯД ЭМБИИ (EMBIOIDEA)
Жители северных и средних районов нашей страны незнакомы с этими своеобразными насекомыми, встречающимися у нас только в Крыму, на Кавказе и в Средней Азии. Но мало кто обращает внимание на эмбий и в тех местностях, где они обычны. Это сравнительно небольшие насекомые, ведущие скрытый образ жизни, встречающиеся под камнями и в почве и редко только по ночам появляющиеся на поверхности.
Эмбии имеют удлиненное цилиндрическое, довольно мягкое, обычно серовато-коричневое тело. Голова крупная, с небольшими фасеточными глазами. Усики сравнительно короткие, нитевидные. Ротовые органы грызущего типа, но нижняя губа неясно расчлененная. Крылья, если имеются (Крылатыми бывают только самцы, а самки всегда бескрылы. Наши виды бескрылые.), небольшие, дымчатые, со слабым жилкованием (рис. 171). Их две пары, причем задние удалены от передних и никак с ними не соединяются. Из грудных сегментов переднегрудь самая короткая, среднегрудь самая длинная. Брюшко у эмбий удлиненное, 10-члениковое, и на заднем конце его находится пара 2-члениковых церков, которые у самцов бывают несколько несимметричными.
Эмбии могут довольно быстро бегать на своих коротких, но хорошо приспособленных к быстрому передвижению ногах, заканчивающихся 3-члениковыми лапками. Передние ноги интересны тем, что первый членик их лапок пузыревидно вздут и в нем находится железа, выделяющая шелковистую паутинку или жидкость для смазывания паутинки. Эмбии выстилают паутинкой ходы (рис. 172), целые системы ходов, по гладкой шелковистой выстилке которых быстро бегают.
Поразительна способность эмбий бегать с одинаковой скоростью и вперед и назад — их ноги сочленяются так, что движения их в обоих направлениях осуществляются одинаково легко. Церки их при беге «задом наперед» служат «усиками» — эмбии ощупывают ими путь, делая ловкие повороты, если встречают препятствия перед задним концом тела при таком необыкновенном способе движения. Особенно быстрыми, иногда даже стремительными бывают движения эмбий по их выстланным шелком ходам. Паутинку выделяют и взрослые эмбии, и личинки, причем сеть выстланных паутинкой ходов служит «домом», а вернее, целым городом для этих насекомых, ведущих полуколониальный образ жизни.
Питаются эмбии в основном всякими гниющими остатками растений. Но несмотря на кажущуюся слабость эмбий и мягкость их покровов, они иногда активно хищничают, расправляясь с такими нежными существами, как камподеи и ногохвостки, и даже мелкими жуками и клопами, которые в поисках влаги забираются под те камни или другие укрытия, под которыми устроили свое «разбойничье гнездо» эмбии. Среди эмбий известен также каннибализм — взрослые в период размножения иногда питаются собственной молодью, особенно самцы. Это не мешает эмбиям большую часть времени дружно обитать в совместных ходах.
В сухое жаркое время года эмбии уходят на довольно большую глубину, переходя в эти периоды от полуобщественного образа жизни к одиночному.
На Южном берегу Крыма во второй половине лета эмбии держатся на глубине около 1—1,5 м, причем встречаются только поодиночке, почти без паутинки, а ранней весной у поверхности почвы собираются взрослые перезимовавшие эмбии, потомство которых в течение влажной первой половины лета не расходится.
В тропиках эмбии встречаются не только в почве, но и на деревьях, где они делают свои паутинные гнезда под отстающей корой или поселяются в ходах, проложенных различными древоядными жуками.
В тропиках многие эмбии летающие, крылатые, тогда как в умеренных широтах больше бескрылых. В теплую безветренную погоду лёт эмбий происходит только по ночам.
Размножаются эмбии, откладывая яйца в своих шелковых галереях, весной; из яйца выходит маленькое насекомое, очень похожее на взрослое, — для бескрылых эмбий характерно почти прямое развитие.
В умеренных широтах весь цикл развития эмбий продолжается 1 год. Интересна хорошо выраженная способность эмбий к восстановлению утраченных конечностей.
Отряд эмбий небогат видами, их всего около 80.
На территории СССР отмечено всего 2 вида. Один из них — обитающая на Южном берегу Крыма, на Черноморском и Каспийском побережьях Кавказа и в Закавказье средиземноморская эмбия (Haploembia solieri). Наши крымская и кавказская эмбии описывались под разными названиями, но при сличении с экземплярами из Италии оказались относящимися к ранее описанной средиземноморской эмбии. Эта эмбия бескрыла, у нас этот вид, как и в других местах вблизи границ области распространения, представлен только самками, размножающимися партеногенетически, тогда как в окрестностях Рима встречаются оба пола.
Средиземноморская эмбия — небольшое, около 1 см, коричневатое насекомое. В Крыму и на Кавказе она служит надежным показателем («индикатором») средиземноморского климата, встречаясь только там, где и почвы, и растительность, и фауна почв имеют средиземноморский характер.
В Средней Азии в каменистых пустынях предгорий распространена туркестанская эмбия (Embia tartara). Она несколько крупнее (около 15 мм) и светлее — песочного цвета.
Самая крупная эмбия известна из Гималаев, она достигает длины около 2 см (рис. 171).


ОТРЯД СЕНОЕДЫ (COPEOGNATHA, ИЛИ PSOCOPTERA)
Сеноедами называют очень мелких насекомых, длиной от 1 до 5 мм, с нитевидными усиками и очень своеобразным строением головы. Сеноеда всегда легко отличить от других насекомых, если рассматривать его голову сбоку (рис. 173, А). Бросается в глаза, что между сильно развитым выпуклым наличником и верхней губой вклинивается в виде поперечной полоски своеобразное скелетное образование — наличничек, который не встречается больше ни в одном отряде насекомых. Характерен и ротовой аппарат сеноедов. Верхние челюсти у них массивные, типичные для грызущего ротового аппарата. В нижних же челюстях произошло очень сильное изменение: их внутренняя лопасть обособилась в виде длинной твердой палочки, далеко вдающейся внутрь черепа, где она погружена в особый плотный мешочек. На наружном конце палочка имеет небольшие зубцы. Богатая мускулатура придает большую подвижность этому органу, позволяя ему выбрасываться вперед, отклоняться вбок, вращаться и т. д. Палочка служит орудием, при помощи которого насекомое раздробляет пищу перед едой. В основном сеноеды питаются наземными зелеными водорослями, лишайниками, а также плесневыми и паразитическими грибами. Некоторые из них питаются органическими остатками, растительной трухой, откуда и происходит название отряда. Реже они употребляют в пищу живые ткани высших растений.
Большинство сеноедов — крылатые насекомые: у них две пары перепончатых крыльев (рис. 174), которые в покое крышеобразно складываются на брюшке. Крылья могут быть или бесцветными, или с темными пятнами и перевязями; вторая пара всегда меньше передних, с упрощенным жилкованием. У ряда видов крылья укорочены или даже совсем отсутствуют.
У сеноедов наблюдается половой диморфизм, выражающийся чаще всего в том, что у самцов усики более густо покрыты волосками, а глаза сильно увеличены, почти шаровидные. У некоторых крылатых видов самки могут быть короткокрылыми или бескрылыми.
То, что сеноеды приспособились к питанию мелкими организмами — лишайниками, водорослями и грибами, обеспечило их широкое распространение, особенно в тропических и субтропических странах. Известно свыше 1500 видов сеноедов, из которых в СССР обитает только около 60.
В природе благоприятные условия для существования сеноедов создаются в лесах, как лиственных, так и хвойных. Здесь их можно найти на стволах и ветках деревьев, а также на листьях. Особенно привлекательны для них стволы и ветки, покрытые лишайниками. Некоторые виды держатся в лесной подстилке, в сухой траве и даже под камнями. Иные встречаются в гнездах птиц и насекомых.
Большинство видов сеноедов имеет несколько поколений в году, при этом часто яйца могут развиваться без оплодотворения. Самки с помощью небольшого яйцеклада откладывают яйца или по одному, или небольшими кучками на листьях, хвоинках, в трещинах коры на стволах и ветках. В одних случаях кладки яиц бывают прикрыты сверху выделениями прямой кишки, которые, подсыхая, образуют небольшие чешуйки. В других случаях кладки покрываются легкой паутинкой. Паутинная нить выделяется у сеноедов особым прядильным аппаратом, проток которого лежит в желобке, образованном соответствующими углублениями на нижней губе и на прикрывающем ее язычке. Это единственный случай, когда у насекомых паутина выделяется изо рта. Отложенные осенью яйца зимуют. Весной из них вылупляются маленькие личинки, по внешнему виду напоминающие вшей, отчего их иногда называют «древесными вшами». Вылупившиеся из яиц личинки или расползаются в стороны и тогда ведут одиночный образ жизни, или же остаются на месте и держатся небольшими «кулигами», устраивая общее паутинное гнездо. Личинки несколько раз линяют и после каждой линьки увеличиваются в размерах. У крылатых форм после сбрасывания личиночной шкурки удлиняются зачатки крыльев.
Некоторые сеноеды встречаются в жилых домах, библиотеках и музейных помещениях. Одни из них обитают в горшках с домашними цветами и питаются обычно растительными остатками. Другие же проникают в книжные шкафы, гербарии и коллекции насекомых и в этих случаях могут повреждать музейные экспонаты. К таким видам относится так называемая книжная вошь (Liposcelis divinatorius) — очень мелкий сеноед, длиной около 1 мм, бледно-бурого или почти белого цвета, совершенно лишенный крыльев. Книжная вошь распространена почти по всему свету. В природе живет в гнездах птиц; в домах встречается в старых бумагах и книгах. Может проникать в коллекции насекомых и гербарии и, если они недостаточно хорошо охраняются, наносит им заметные повреждения.
Несколько крупнее (до 2 мм) пыльная вошь, или домовый сеноед (Trogium pulsatorium), который отличается тем, что у него имеются зачаточные крылья. Он светло-желтого цвета, причем на верхней стороне брюшка имеются красноватые пятна, образующие продольные полосы. Живет домовый сеноед в таких же условиях, как и книжная вошь; может встречаться в пыльных углах комнат. Интересно, что он издает слабые тикающие звуки, ударяя передней частью головы о бумагу или дерево.
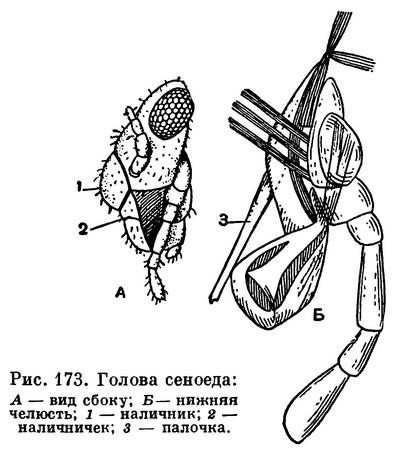
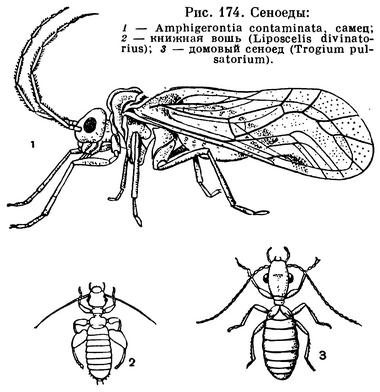
ОТРЯД ПОДЕНКИ (EPHEMEROPTERA)
На свет фонарей, зажигаемых на речных берегах, на огни пароходов в теплые тихие вечера иногда в огромных количествах слетаются нежные насекомые с прозрачными крыльями — так называемые поденки. Узнать их легко. У них две пары сетчатых прозрачных и очень тонких крыльев, причем передние всегда гораздо крупнее задних (у некоторых поденок задние крылья совсем не развиты), а на конце брюшка три или реже две длинные тонкие хвостовые нити. В тихую погоду вечером интересно наблюдать их характерный полет: быстро махая крыльями, они взмывают вверх, а затем замирают и благодаря большой поверхности крыльев и длинным хвостовым нитям, как на парашюте, спускаются вниз. Затем снова взлет, снова плавное падение. Такой «танец» совершают поденки в период размножения — самец подлетает к самке и тут же в воздухе снизу прицепляет сперматофоры к ее половым отверстиям, которых у поденок два — правое и левое.
Не случайно эти изящные насекомые получили название поденки или однодневки: некоторые из них во взрослом состоянии действительно живут один день, иногда даже меньше — несколько часов, хотя некоторые иногда живут и по нескольку дней, но всегда недолго.
Недолгий полет, при котором поденки могут попасть в новые благоприятные условия, и размножение — вот те биологические функции, которые осуществляются взрослой стадией этих насекомых.
Взрослые поденки не питаются, более того, они не могут принимать пищу. Ротовые органы у них недоразвитые, мягкие, а кишечник превращен в воздушный пузырь, облегчающий вес насекомого. Вот почему поденки так легко парят в воздухе и так плавно и медленно опускаются, когда прекращаются взмахи крыльев.
После спаривания самцы погибают, а самки откладывают яйца. Среди насекомых трудно найти другой отряд, в пределах которого были бы так разнообразны по своему внешнему виду яйца (рис. 176). У некоторых видов яйца откладываются кучками, у некоторых разбрасываются; яйца бывают с якорями, цепляющимися за подводные предметы. Редко, но бывает и живорождение, например у не имеющей задних крыльев двукрылой поденки (Cloeon dipterum).
Все личинки поденок развиваются в воде. Несмотря на большое разнообразие общей формы тела личинок поденок разных видов, по некоторым типичным особенностям их всегда удается отличить от других водных насекомых. Первое, что бросается в глаза, — длинные хвостовые нити. Обычно их три (рис. 177), реже только две, если развиты только церки, а срединная нить редуцирована. Главная же особенность личинок поденок та, что у них на первых 7 члениках брюшка находятся трахейные жабры. Правда, у только что вылупившихся из яиц личинок жабр обычно еще нет, например, у личинок Ephemera они появляются только на четвертый день.
Трахейные жабры бывают очень разной формы — в виде простых или перистых пластинок, часто с бахромой, иногда в виде пучка отростков, отходящих от боков сегментов. В жабры заходят трахеи, как заходят они и в основания хвостовых нитей. Хвостовые нити помогают личинкам поденок плавать, действуя по такому же принципу, как хвост у дельфина, т. е. двигаясь в вертикальной плоскости.
Большинство личинок поденок живет в быстрых ручьях и реках, хотя некоторые виды обитают в стоячих водоемах.
Личинки разных видов живут по-разному — некоторые ползают среди зарослей водных растений, другие сооружают норки в отвесных крутых берегах, есть формы, роющиеся в иле, а есть живущие под камнями, прикрепляющиеся к их нижней стороне в бурных потоках, — таких легко узнать по широкому сплющенному телу.
Питаются личинки поденок тоже по-разному. Часть их хищничает, многие кормятся разлагающимися остатками — детритом, илом и т. д. Личинки хиропектесов, обитающие на дне быстрых ручьев, интересны тем, что у них челюстные щупики и передние ноги покрыты длинными волосками, отфильтровывающими проплывающие пищевые частицы, и действуют как ловчая сеть.
Жизнь личинки длится нередко 2—3 года, и за этот срок личинка многократно линяет. Достоверно отмечено 23 линьки, но, вероятно, их число много больше. Когда личинка достигает своих окончательных размеров, она превращается во взрослое насекомое. Существует много способов превращения.
У личинок — обитателей спокойных вод к концу личиночной жизни вес тела облегчается благодаря тому, что между последней шкуркой личинки и новой шкуркой будущего крылатого насекомого образуется много пузырьков воздуха. Ставшая легкой, личинка всплывает к поверхности. После этого начинается появление пузырьков газа в ставшем пустым кишечнике. (Вспомним, что у взрослых поденок желудок превращается в воздушный пузырь!) Шкурка личинки лопается, взрослое насекомое, сидя на ней, как на лодке, довольно быстро расправляет крылья и улетает. В некоторых случаях всплывает огромное количество шкурок. Так, на озере Мичиган в Северной Америке бывает, что ветер сгоняет к берегу такое обилие плавающих шкурок, оставшихся после дружного выхода поденок Ephemera simulans, что они покрывают многие гектары береговой полосы слоем, достигающим толщины нескольких футов.
Личинки поденок, живущие в быстрых реках, вылезают на берег или на какой-нибудь торчащий из воды предмет и тоже очень быстро превращаются в крылатых насекомых, которые вскоре взлетают. Но оказывается, что уже окрылившаяся поденка еще не закончила своего развития. Вышедшая из последней личиночной шкурки поденка (субимаго), некоторое время полетав, садится на какую-нибудь травинку и за 2—3 минуты, а иногда и быстрее линяет (рис. 178). Шкурка крылатого насекомого лопается продольным швом на спине, и из сбрасываемой шкурки выходит новая крылатая стадия, на этот раз уже способная к размножению (имаго). Ни в одном другом отряде насекомых, кроме поденок, не бывает линьки окрылившихся особей, а у поденок есть две крылатые формы — неполовозрелая и половозрелая, разделенные линькой (рис. 179).
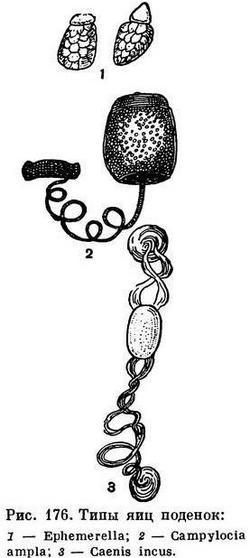
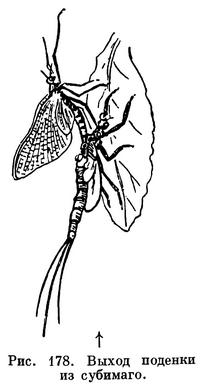
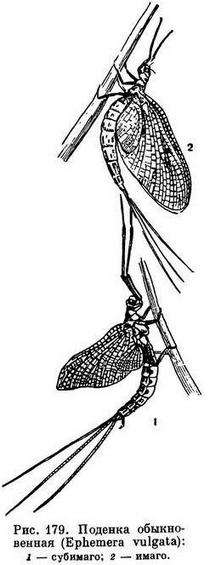
Личинки семейства прозопистом (Prosopistomidae) отличаются очень широким листовидно сжатым телом, их жабры скрыты под боковыми выростами слившихся сегментов, так что у них образуется жаберная полость. Хвостовые придатки этих личинок напоминают вилочку низших ракообразных (рис. 177, 4). Немудрено, что и описаны они были в свое время знаменитым зоологом Жоффруа Сент-Илером как ракообразные. Живут эти личинки в быстрых потоках под камнями.
У настоящих поденок (семейство Ephemeridae) личинки имеют вытянутое цилиндрическое тело с сильными роющими ногами. Эти личинки роют себе ходы в глинистом грунте медленно текущих рек.
Личинки гептагений (семейство Heptageniidae) имеют умеренно вытянутое тело, сплющенное в спинно-брюшном направлении. Ноги у них длинные, цепкие, хвостовые нити длинные — эти личинки ловко ползают среди камней и подводных растений.
Личинки маложильных поденок (семейство Oligoneuridae) отличаются крупными размерами и сжатым телом, а хвостовые нити у них короткие, короче длины тела.
Представители семейства баэтид (Baetidae) живут в основном в стоячих водоемах и характеризуются направленными в стороны ногами.
Личинки поденок чувствительны к присутствию в воде различных посторонних химических соединений, и количество их в тех реках, в которые поступают сточные воды заводов, резко сокращается.
Если сравнить степень разнообразия взрослых поденок и их личинок, окажется, что взрослые поденки разных видов куда меньше отличаются друг от друга, чем личинки. Это понятно, если вспомнить, что взрослые поденки живут так недолго и ведут примерно одинаковый образ жизни. А личинки развиваются иногда дольше года, в разных водоемах, в разных местах водоемов, отличающихся по грунту, течению и т. п., и у них, естественно, выработались разные приспособления.
Интересно, что по берегам равнинных рек взрослых поденок бывает огромное количество, они летают целыми тучами, а личинок их, зарывающихся в илистое дно, довольно трудно найти. Наоборот, в горных потоках бывает, что личинки поденок кишат под камнями, а взрослых встретить довольно трудно — они прячутся в прибрежной траве, ведут скрытый образ жизни.
Число видов поденок невелико — их известно немногим более 1500 видов.
Распространены поденки по всему земному шару, за исключением некоторых океанических островов. Так, например, их нет на Гавайских островах в Тихом океане, на острове Святой Елены в Атлантическом.
Палеонтологическая история поденок уходит в глубь каменноугольного периода палеозойской эры. По характеру развития, по строению хвостовых придатков, по линьке во взрослом состоянии и по многим другим признакам поденок можно считать более или менее близкими к щетинохвосткам. Поденки — очень древняя группа; если основываться на особенностях расположения крыловых жилок, на неспособности крыльев складываться, можно, как это сделал наш выдающийся специалист по ископаемым насекомым А. В. Мартынов, рассматривать поденок как «древнекрылых» насекомых. Вероятно, прямые предки нынешних поденок были крылатыми насекомыми, которые в личиночной стадии, как и щетинохвостки, жили во влажных местах на суше — под камнями, в трещинах скал и т. д. — и лишь постепенно перешли к развитию в воде (У личинок поденок, как показал чешский зоолог В. Ланда, есть рудиментарные дыхальца.).
Личинками поденок охотно питаются рыбы, в наших северных реках (Онеге, Печоре) поденки представляют основной вид пищи многих промысловых рыб.
У поденок есть своеобразные паразиты. Личинки некоторых комаров-дергунов, например Symbiocladius rithrogenae, паразитируют на личинках поденок, поселяясь в складках зачатков крыльев и высасывая из них кровь.
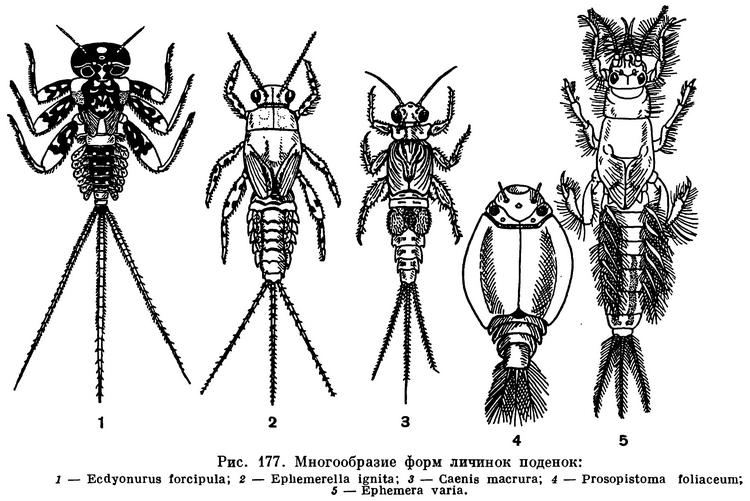
ОТРЯД СТРЕКОЗЫ (ODONATA)
Кто не знает стрекоз, кто не любовался их полетом — иногда стремительным, иногда почти парящим? Чьи взоры не привлекали блеск переливающих иногда всеми цветами радуги крыльев, яркая окраска их тела? У каждого народа есть свое название для стрекоз, и часто не одно! Это показывает, что издавна стрекозы привлекали к себе внимание. Многие поэты — В. Гёте, А. Толстой и другие, вдохновленные красотой и изяществом этцх насекомых, писали о них стихи.
Стрекозы характеризуются стройным, вытянутым, иногда ярко окрашенным или блестящим туловищем, крупной хорошо обособленной от него головой, большую часть поверхности которой составляют огромные глаза (табл. 32). Усики у стрекоз маленькие, малозаметные. У стрекоз две пары прозрачных крыльев, пронизанных густой сетью мелких жилок, в передней части крыльев близ вершины имеются темные пятнышки, служащие стабилизаторами, не дающими тонким крыльям вибрировать при полете.
Большинство стрекоз летает днем в самые жаркие часы. Их бывает особенно много по берегам водоемов, но нередко целые стаи их носятся по опушкам леса. На лету они ловят свою добычу — комаров-дергунов, настоящих кровососущих комаров и других мелких насекомых. Если булавкой отогнуть стрекозе нижнюю губу и раздвинуть челюсти, видно, какая огромная пасть у этих прожорливых хищников!
Хотя стрекозы прекрасно летают, крылья их совершают довольно простые движения, и нашему энтомологу Ю. М. 3алесскому удавалось заставить стрекоз летать на «протезах»: он обрезал стрекозе крылья, оставляя только основание крыла, к которому осторожно приклеивал крыло другой стрекозы или даже бабочки, и стрекоза на чужих крыльях, хоть и плохо, но летала!
Деятельны стрекозы в теплое время дня, на солнце. Вечером, когда темнеет и становится прохладно, стрекоза садится на камышинку или ветку и в застывшем, вялом состоянии проводит ночь, снова начинает летать, согретая ярким утренним солнцем.
Личинки мелких стрекоз питаются личинками насекомых, например поденок и комаров, мелкими водными червями и другими беспозвоночными. Личинки более крупных видов отваживаются нападать на мальков рыб и головастиков. В отличие от взрослых стрекоз у личинок (называемых, как и личинки поденок, наядами) усики более длинные, нитевидные. Ноги личинок стрекоз тоже длиннее и подвижнее, чем у взрослых.
Дыхание у личинок стрекоз, как и у настоящих водных животных, осуществляется за счет кислорода, растворенного в воде. Они не должны подниматься к поверхности воды или вылезать на берег, чтобы дышать. Растворенный в воде кислород проникает в организм личинки стрекозы не только через любой более тонкий участок покровов, но в основном через поверхность специальных дыхательных органов.
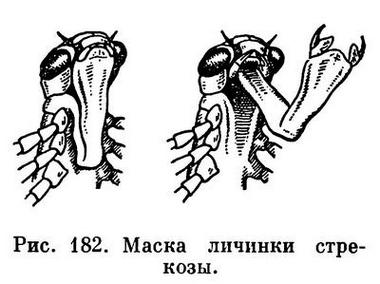
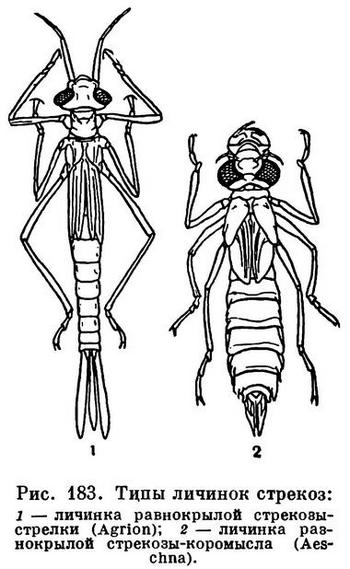
У личинок более мелких равнокрылых стрекоз (лютки, стрелки) дыхательными органами служат листовидные трахейные жабры, находящиеся на заднем конце брюшка (рис. 183, 2). Трахейными они называются потому, что их пронизывает густая сеть трахейных трубочек.
У личинок крупных стрекоз, таких, как коромысло и другие «неравнокрылые», вообще не бывает наружных трахейных жабр и дыхание осуществляется только путем «вдохов» и «выдохов» воды через анальное отверстие в заднюю кишку. С силой выталкивая воду из заднепроходного отверстия, личинка может быстрым толчком устремляться вперед, двигаясь по принципу действия реактивного двигателя. Такой тип движения наблюдается лишь у представителей немногих групп водных животных, например у медуз и головоногих моллюсков.
После выхода из воды личинка стрекозы обсыхает, вскоре на ее спине, а затем и голове появляется трещина и спустя некоторое время через нее из последней шкурки личинки вылезает взрослая стрекоза, вытаскивающая из оболочек ног личинки свои конечности (рис. 184). Вышедшая из шкурки наяды молодая стрекоза отползает от старой оболочки и снова затихает. Ее сморщенные крылья, сперва не превышающие зачатков, бывших у наяды, под давлением полостной жидкости (крови) расправляются и затвердевают. На расправление и затвердевание крыльев уходит около 6 часов.
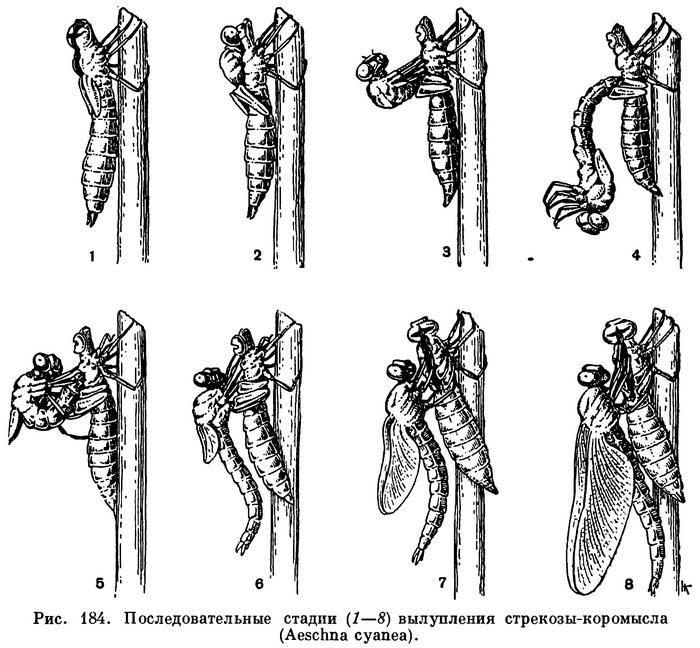
Как уже упоминалось, стрекозы — отличные летуны, и часто их можно встретить на далеком расстоянии от тех водоемов, где они вывелись. Нередко они перелетают целыми стаями, что дает им возможность быстрее заселить новые водоемы.
Способность к полету обеспечивает выживание части стрекоз и при высыхании водоемов. А при подсыхании временных луж личинки стрекоз, особенно не имеющие трахейных жабр, переползают по траве в соседние водоемы. Это тоже помогает выживанию видов.
Стрекоз известно более 3000 видов, больше их в теплых странах.
Стрекозы — очень древняя группа; хорошо сохранившиеся остатки ископаемых стрекоз известны еще из отложений каменноугольного периода палеозойской эры. В это время обитали и гитанты среди насекомых — палеодиктиоптеры (Palaeodictyoptera, рис. 185), родственные стрекозам. Некоторые из них, судя по ископаемым остаткам, в размахе крыльев достигали 90 см.
Практическое значение стрекоз невелико. В общем преобладает приносимая ими польза — взрослые стрекозы ловят различных мелких летающих насекомых, в том числе много кровососущих насекомых — комаров, мошек и др. Личинок стрекоз охотно поедают бентосоядные рыбы — карпы, лини и др. Но личинки крупных стрекоз (коромысла и др.) могут в рыбоводческих хозяйствах вредить, поедая мальков рыб.
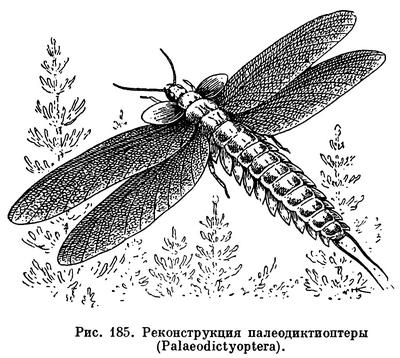
ОТРЯД ПУХОЕДЫ (MALLOPHAGA)
Пухоеды — мелкие наружные паразиты птиц (пухоеды и пероеды) и млекопитающих (власоеды) (рис. 187).
Так как вши в народе были известны с незапамятных времен, а на курах и других птицах паразитируют внешне похожие на них пухоеды, последних в просторечии часто называют вшами («куриная вошь» и т. д.), но это только следствие внешнего сходства, отражающегося во многих народных названиях животных (например, «земляной заяц» — распространенное название тушканчика и т. п.).
В отличие от вшей, на которых пухоеды несколько похожи, они не сосут кровь, а питаются перьями или волосами, а также чешуйками рогового слоя кожи и возникающими в местах расчесов струпьями, а некоторые заглатывают выступающие из расчесов капли крови.
Это очень подвижные, особенно в ранних возрастах, насекомые, отличающиеся плоским телом, огромной треугольной головой и свободной переднегрудью. Ротовые органы у пухоедов грызущие, смещенные на нижнюю поверхность головы. Покровы у большинства пухоедов плотные, желтоватые или темные.
Среди пухоедов, которых известно около 2500 видов, только около 60 паразитирует на млекопитающих (власоеды), в том числе и на сумчатых, но на человеке власоеды не паразитируют никогда.
Для большинства пухоедов характерна строгая приуроченность к хозяину — птице определенного вида. Заражение птиц пухоедами происходит при контакте хозяина с другими особями того же вида. Птенцы обычно заражаются пухоедами от родительских особей и часто слабые птенцы и молодые птицы бывают даже сильнее заражены, чем старые, которыо тщательно чистят себе перья, выбирая из них пухоедов. А у кукушки, как известно, откладывающей яйца в гнезда других птиц, имеющей своих специфических паразитов-пухоедов, на птенцах нет пухоедов — с «приемных родителей» других видов насекомоядных птиц пухоеды на кукушат не переходят, и только при спаривании взрослые кукушки заражаются друг от друга пухоедами. Когда птица погибает, погибают и почти всегда находящиеся на ней пухоеды, приспособленные к паразитическому образу жизни только при той высокой температуре, которая создается под покровом перьев хозяина. Но некоторые пухоеды могут уцелеть. Дело в том, что на птицах (и на млекопитающих) паразитируют и способные к полету мухи-кровохлебки (Hippoboscidae), которые после смерти птицы перелетают на другого хозяина. Иногда к ним прикрепляются и пухоеды и так в качестве пассажиров попадают на нового хозяина. Но кровохлебки не такие специализированные паразиты, как большинство пухоедов — они могут перелетать и на птицу другого вида. Доставленный кровохлебкой на птицу «своего» вида пухоед выживает, а если муха перелетит на птицу другого вида, специализированный паразит-пухоед некоторое время на ней просуществует, а затем погибнет. Такой способ расселения пухоедов помогает понять происхождение близких видов пухоедов, обитающих на близких видах птиц. Как правило, систематически близкие виды пухоедов паразитируют на родственных группах животных-хозяев. Пухоеды иногда позволяют орнитологам решать вопросы о систематическом родстве разных групп птиц. Так, сравнение пухоедов на фламинго с пухоедами других птиц показало, что они ближе к пухоедам утиных, чем к пухоедам аистов и разных других «голенастых», что дает дополнительный серьезный аргумент в пользу сближения фламинго с утиными.
Если на одном виде хозяина паразитирует несколько видов пухоедов, каждый из них приспособлен к жизни на определенном участке тела, на определенного типа перьях и т. д., т. е. разные виды не конкурируют друг с другом. Известны пухоеды, перешедшие к необычному образу жизни, — виды рода Piagetiella держатся в глоточном мешке пеликанов и в пищеводе бакланов. Эти пухоеды мало изучены, вероятно, они питаются кровью птиц.
Яйца пухоеды могут откладывать круглый год — для них всегда «теплый период» на теле теплокровного животного. Яйца у них с крышечками; иногда они прикрепляются к перьям целыми комочками. Развитие яиц длится около 2 недель; так как развитие у пухоедов прямое, выходящие личинки похожи на взрослых.
Хотя пухоеды, казалось бы, питаются в основном нечувствительными ороговевшими частями покровов животных (рис. 190), они причиняют сильный зуд, когда соскребают ороговевший эпителий или ползают по поверхности тела. Поэтому птицы часто чистят перья, выклевывая этих паразитов. А пухоеды очень подвижны, чтобы избегать клюва птицы. Известно, что пухоеды в массе размножаются на ослабленных птицах, которые не в состоянии постоянно очищать свои перья. А размножение пухоедов на птице в свою очередь ведет к ее истощению. Выражаясь языком кибернетики, между размножением пухоедов и состоянием хозяина устанавливаются отношения по принципу «обратной связи».
Настоящие пухоеды (подотряд Amblycera) имеют скрытые усики, часто с булавой, их жвалы направлены вперед. К настоящим пухоедам относится, например, бледный куриный пухоед (Menopon pallidum), поселяющийся на коже курицы и быстро бегающий.
Эта группа включает также 3 рода (Boopia и др.), паразитирующих на кенгуру, вомбатах и других сумчатых.
Перо- и власоеды (подотряд Ischnocera) характеризуются заметными нитевидными усиками (рис. 187) и жвалами, направленными вертикально. Сюда относятся такие виды, как очень узкотелый головной куриный пухоед (Lipeurus heterographus) и голубиный пухоед (Columbicola columbae). На утках паразитирует крупный утиный пухоед (Trinotumluridum) — 4—5 мм длины, а на орлах встречается достигающий 1 см в длину Laemobothrium titan — гигант среди пухоедов.
Из власоедов собачий (Trichodectes canis), овечий (Trichodectes sphaerocephalus), воловий (Trichodectes scalaris) и другие живут на млекопитающих. Власоеды часто бывают промежуточными хозяевами глистов: собачий — цепня, а некоторые птичьи — нематод-филарий. Борьба с пухоедами домашней птицы легко осуществляется путем обработки животных ДДТ при параллельной очистке и дезинсекции помещения.
Из свободноживущих насекомых пухоеды ближе всего к сеноедам, от которых, вероятно, и произошли.

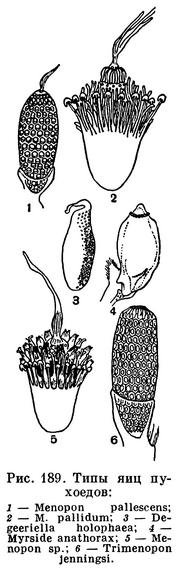
ОТРЯД ВШИ (ANOPLURA)
Отряд вшей объединяет мелких (длина тела от 0,4 до 6 мм) наружных паразитов млекопитающих.
Вши (рис. 191) — сосущие насекомые. Их ротовые органы приспособлены для прокалывания плотных покровов животного-хозяина и высасывания крови. Ротовые части преобразованы в колющие иглы, заключенные в выворачиваемую из ротовой полости мягкую трубку, края которой плотно прижимаются к прокалываемой стилетами коже животного-хозяина.
При сосании крови расширяется передний отдел пищевода вши, действующий как насос. Выделения мощно развитых слюнных желез, поступающие в ранку, препятствуют свертыванию крови. Когда вошь не питается, ротовые органы, образующие хоботок, втянуты в головную капсулу. Глаза у вшей либо представляют только маленькие пигментированные пятнышки, либо вовсе отсутствуют. Усики короткие, а каких-либо щупиков, связанных с ротовыми органами, нет вовсе. Грудь хорошо отделена от головы, все сегменты груди слитные.
Вши — бескрылые насекомые, ни у одного вида вшей нельзя обнаружить никаких рудиментов крыльев. Но тем не менее вши относятся к подклассу крылатых насекомых — ход ветвей трахейной системы и нервов у них такой, как у крылатых насекомых, и, вероятно, они произошли от перешедших к паразитизму и вторично утративших крылья предков. Ноги у вшей очень сильные, цепкие, приспособленные к удерживанию на шерстном и волосяном покрове животного-хозяина. Голени ног лапка одночлениковая с одним мощным серповидным коготком; на голени есть выемка, к которой коготок может подгибаться.
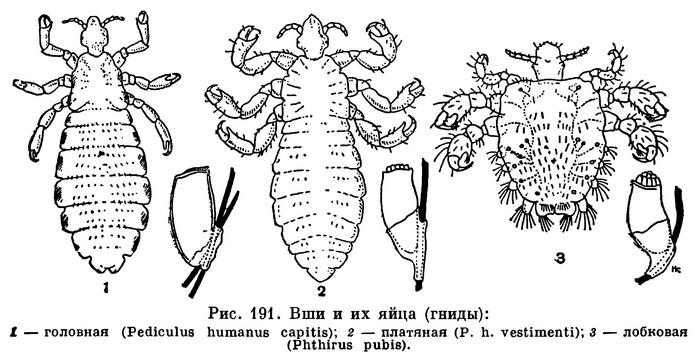
Захватывая волос коготком, прижимая коготок к голени, вши прочно удерживаются на теле хозяина (рис. 192). Интересно, что у Haematomyzus elephantis — довольно крупной слоновой вши (Слоновую вошь часто теперь выделяют в особый отряд.) (рис. 193), живущей в складках кожи этого громадного толстокожего животного, не имеющего густого волосяного покрова, ноги длинные, но слабые и без такого хватательного приспособления, и вошь удерживается на коже слона с помощью длинного хоботка, передний конец которого в толще кожи расширяется и не дает паразиту отвалиться. По-видимому, только на кенгуру и других сумчатых нет вшей.
Вши очень специализированные паразиты, обитающие только на одном или на немногих близких видах животных. Эта особенность вшей, как и некоторых других паразитов, позволяет использовать данные изучения паразитов для выяснения близости видов хозяев в некоторых трудных для зоологов случаях.
Среди вшей хорошо различаются 3 семейства. Семейство гематопид (Haematopidae) — слепые вши, обычно с покрытыми волосками покровами, — паразиты только наземных млекопитающих, кроме приматов. Сюда относится и упоминавшаяся слоновая вошь и свиная вошь (Haematopinus suis). На разных копытных живут другие виды этого рода, например, на крупном рогатом скоте — бычья вошь (Haematopinus eurystemus); заячья вошь (Haemodipsus ventricosus) — мелкое насекомое с вздутым волосистым телом паразитирует только на представителях семейства заячьих; оленьи вши (виды рода Cervophthirius) — только на оленях и т. д.
Любопытно семейство колючих вшей (Echinophthiridae), паразитирующих только на морских млекопитающих. У них шарообразное тело густо покрыто шипиками, как например у тюленьей вши (Echinophthirius horridus), живущей в полости около ноздрей тюленей. Когда тюлень ныряет, вошь оказывается в закрытой клапанами ноздре, т. е. в воздушной среде. Многие вши этой группы, живущие в волосяном покрове ластоногих, имеют на спинной поверхности чешуйки, под которыми задерживается слой воздуха, когда тюлень ныряет.
Третье семейство — педикулиды (Pediculidae) — паразиты только приматов — обезьян и человека. У этих вшей есть небольшие глазки. Представители подсемейства педицин паразитируют на обезьянах, а представители подсемейства педикулин — паразиты человека и некоторых человекообразных обезьян.
На покрытых волосами частях тела (кроме головы), чаще всего на лобке, под мышками, но бывает и в бороде, на бровях и даже ресницах, у человека паразитирует лобковая вошь, или площица (Phthirus pubis). Эта мелкая вошь (рис. 191, 3) имеет широкое тело, достигающее всего 1 мм у самца, 1,5 мм у самки, с длинной головой и неясно разграниченными грудью и брюшком, передвигается, часто ползая боком, как краб, а к волосам прикрепляется своими цепкими коготками очень прочно. В тех местах, где сосет эта вошь, появляются синеватые пятна, так как ее слюна разрушает гемоглобин крови. Никаких болезней эта вошь не переносит. Интересно, что два других вида этого рода — паразиты человекообразных обезьян: один паразитирует на горилле, а другой — на шимпанзе.
Родство таких специализированных паразитов, как вши, подтверждает несомненную близость человека и человекообразных обезьян!
Если лобковая вошь не представляет опасности как переносчик заболеваний человека, то другая вошь, живущая на человеке, — человечья вошь (Pediculus humanus) не только паразит, вызывающий мучительный зуд, но и переносчик многих опасных заболеваний. Человечья вошь дает нам интересный пример явного образования двух видов из одного. Она представлена двумя стойкими формами — платяной (Pediculus humanus vestimenti) и головной (Pediculus humanus capitis). Они обе резко отличаются от площицы большими размерами, удлиненной формой тела и короткой головой. Грудь и брюшко у них ясно разделены, причем брюшко шире, чем грудь. Головная вошь серее и короче (самец 2—3 мм, самка до 4 мм), платяная светлее и достигает 4,7 мм (самки).
Долгое время платяную и головную вшей считали за 2 разных вида. Но исследования, проведенные у нас в годы Великой Отечественной войны, показали, что, если воспитывать платяную вошь при более низкой температуре, через несколько поколений ее потомство приобретает признаки головной, и наоборот. Этого можно достичь и пересаживая головную вошь под повязку на тело человека, а платяных на голову. Кроме того,обе формы при скрещйвании дают плодовитое потомство. Но без насильственной пересадки платяные вши остаются по облику и образу жизни платяными, а головные — головными. Очевидно, обитание в разных условиях температуры и питания (на голове и под одеждой) приводит к расхождению признаков прежде единого вида.
На примере человечьей вши можно ознакомиться с особенностями биологии этих паразитов. Яйца, называемые «гнидами», вошь приклеивает к волосам (головная) или к волокнам ткани белья. За свою жизнь платяная вошь откладывает до 250—300 яиц; поскольку цикл развития длится 16 дней, а жизнь вши продолжается около 2 месяцев, потомство одной самки к концу ее жизни измеряется тысячами. Этим объясняются случаи завшивления людей, когда они лишены возможности мыться и менять белье.
Зрение у вши слабое — глаза позволяют только воспринимать свет и уходить от него, забираясь в гущу волос или складки одежды. Ориентируются вши по запаху, улавливаемому усиками. У вшей есть избирательность к тем или иным людям. Ставили такой опыт: за небольшой столик садилось четверо, на столик клали вошь, и она всегда безошибочно ползла к одному и тому же человеку.
Платяная вошь очень чувствительна к изменениям температуры, она хорошо чувствует себя при температуре поверхности тела здорового человека, при повышении температуры она начинает беспокойно ползать, а температура свыше 44° С для вшей уже гибельна. При понижении температуры тела (например, при остывании трупа) вши тоже начинают расползаться. При температуре 10—20°С вши могут около 10 дней обходиться без пищи. Чаще всего заражение вшами происходит при тесном контакте, при скученности людей, попадающих в тяжелые антисанитарные условия, лишающие возможности менять одежду, стирать ее, мыться. Особенно больших масштабов достигает зараженность населения вшами в периоды голода, войн и тому подобных бедствий.
Массовое размножение вшей и заражение ими большого числа людей приводит к жестоким эпидемиям — человечья вошь, особенно ее платяная форма, передает возбудителей таких опасных болезней, как сыпной тиф, возвратный тиф и ряд других. Особенную опасность представляет сыпной тиф — болезнь, вызываемая поселяющимися внутри клеток микроорганизмами — риккетсиями, передаваемая вшами при сосании крови. Известно, что во время русско-турецкой войны 1768—1774 годов от сыпного тифа в русской армии умерло больше людей, чем от ранений, в первую мировую и в гражданскую войны людские потери от сыпного тифа были огромными. Во время второй мировой войны в Египте, Алжире, Италии, в оккупированных немецко-фашистскими войсками балканских странах и в Польше были эпидемии сыпного тифа, и только четкая организация санитарной службы и в армии и в тылу предохранила нашу страну от эпидемии сыпного тифа.
Основой борьбы со вшами является личная гигиена. Зараженная или подозрительная на зараженность вшами и гнидами одежда подвергается обработке высокой температурой или инсектицидами в специальных дезинсекционных камерах. В случае появления вшей на белье и одежде, при обнаружении даже единичного экземпляра этого паразита необходимо тщательно выстирать, прокипятить и прогладить белье, вымыться горячей водой, прогладить одежду горячим утюгом. При появлении вшей в голове — вымыть голову, смочить волосы керосиново-мыльной эмульсией (взбить в теплой воде до состояния белого молока 50 г керосина и 30 г мыла на 1 л воды) и завязать смоченную эмульсией голову на 30 минут. Те меры, которые легко принять в нормальной обстановке, помогают полной ликвидации вшей, которые могут размножаться только на людях при несоблюдении ими правил гигиены.
Вши — небогатый видами отряд, всего известно немногим более 150 видов.
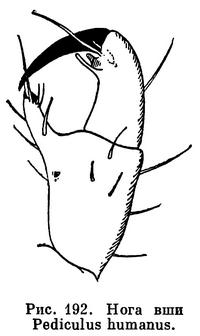
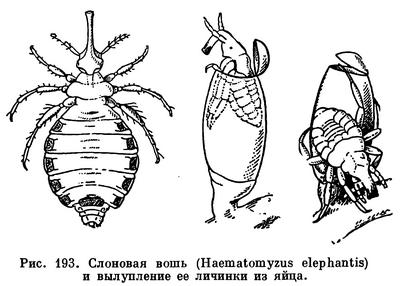
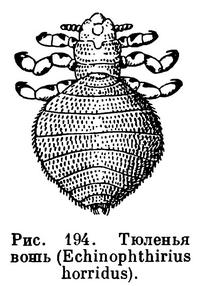
ОТРЯД РАВНОКРЫЛЫЕ ХОБОТНЫЕ (HOMOPTERA)
Представителей обширного отряда равнокрылых хоботных (описано более 30 000 видов) иногда объединяют вместе с полужесткокрылыми (клопами) в единый отряд хоботных (Rhynchota), так как и в строении ротового аппарата, и в особенностях анатомии внутренних органов у них много общего. Но несмотря на черты сходства и родства, обе группы хоботных в своей эволюции пошли разными путями, и правильнее их считать самостоятельными отрядами.
Равнокрылые хоботные — все без исключения сосущие насекомые, питающиеся только соками высших растений, преимущественно цветковых, — на папоротниках питается лишь немного видов. Наиболее характерные черты строения равнокрылых хоботных связаны с тем, что они используют в качестве источника воды и пищи соки растений.
Для высасывания соков служит хоботок, состоящий из расчлененной на 3—4 участка нижней губы, приобретающей вид желобчатой трубочки; сверху у основания желобок прикрыт вытянутой верхней губой, которая короче нижней губы. Внутри желобка такого хоботка находятся 4 длинные тонкие колющие щетинки, способные довольно далеко высовываться из хоботка. Эти щетинки, длина которых у некоторых равнокрылых превышает длину тела, — так сильно изменившиеся верхние и нижние челюсти. Щетинки прочные, но очень тонкие и гнущиеся. Именно благодаря тому что их охватывает, как трубка, нижняя губа, они, несмотря на гибкость, прокалывают оболочки клеток растения. Ротовой аппарат присоединяется к задней части направленной вниз головы.
Глаза у равнокрылых либо сложные, либо у малоподвижных форм упрощаются до скоплений небольших глазков, а у совсем сидячих представителей отряда и полностью исчезают. Грудные сегменты у летающих форм хорошо обособлены. Крыльев две пары, обе пары крыльев прозрачные, перепончатые или передние кожистые, но равномерно уплотненные по всей длине. Передние крылья развиты сильнее, а в некоторых случаях (у самцов щитовок) задние крылья совсем исчезают. Ноги с небольшим количеством члеников.
Для равнокрылых характерно внутреннее оплодотворение, у многих известно партеногенетическое размножение.
Трудно дать общую характеристику всех равнокрылых — они так разнообразны, что их делят на несколько подотрядов.
ПОДОТРЯД ЦИКАДОВЫЕ (AUCHENORRHYNCHA)
Представители этого подотряда (табл. 35, 36) включают в среднем более крупных насекомых, чем объединяемые в другие подотряды группы. Отличительные их признаки те, что голова снизу хорошо отделена от груди, а нижняя губа прикреплена впереди ног.
Усики 3-члениковые, толстые, первый с щетинистым придатком. Между сложными глазами 2—3 простых глазка. Ноги сильные, задние у многих видов удлиненные, прыгательные. Именно у цикадовых бывают плотные передние крылья.
Личинки у большинства цикад развиваются в почве на подземных частях растений, избегая таким образом сильной потери влаги, а те, которые перешли к развитию на надземных частях растений, выработали другие способы экономии воды (выделение пенистой жидкости или воскоподобных веществ).

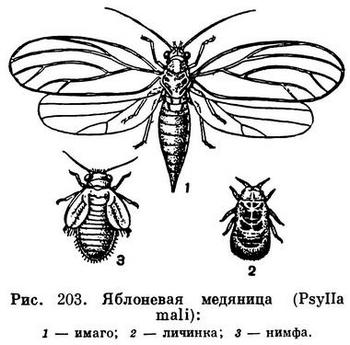
ПОДОТРЯД ЛИСТОБЛОШКИ (PSYLLOIDEA)
Известно около 1000 видов листоблошек. Это мелкие насекомые, по форме тела похожие на цикадок, а по размерам, нежности покровов и образу жизни они напоминают тлей. В СССР встречается около 100 видов листоблошек, некоторые из них вредят садовым культурам. Самые крупные листоблошки — австралийские крейсы (Creiis), достигающие всего 6 мм. А наши листоблошки меньше 3 мм.
Для листоблошек характерны многие особенности, сближающие их по образу жизни с тлями. Как и тли, листоблошки сосут так много соков из растений, что не усваивают весь содержащийся в пище сахар, и экскременты у них сладкие. Поэтому листоблошек часто посещают муравьи. Некоторые листоблошки, особенно в личиночной стадии, способны выделять восковые нити, защищающие от потери влаги; кроме того, этих насекомых от высыхания защищают и их сладкие выделения. В тропиках некоторые листоблошки вызывают образование галлов на листьях, а питание наших видов на листьях часто вызывает их скручивание.
Обычный у нас вид листоблошек — так называемая яблоневая медяница («медвяница» — Psylla mali, рис. 203). У этой медяницы зимуют яйца, отложенные у оснований почек на новых побегах яблонь. Личинки забираются в распускающиеся почки и бутоны и высасывают их — в результате почки не развиваются, а бутоны опадают. Личинки сперва оранжево-желтые, а затем зеленеют. Личинки выделяют много сахара, и побеги, поврежденные ими, неприятно липкие на ощупь. В начале лета происходит окрыление медяниц — превращение в зеленых крылатых насекомых. Если их много, то на дереве раздается шуршание, вызванное тем слабым стуком, который возникает, когда маленькое насекомое садится на лист. Несколько видов медяниц известно на груше, причем у грушевой медяницы (Psylla pyricola) перезимовывают и взрослые особи, и личинки. На ольхе ольховая листоблошка (Psylla alni) выделяет восковые нити, а живущая на ситнике и осоках ливия (Livia juncorum) делает галлы на цветах. С медяницами на культурных растениях приходится вести борьбу химическим путем, применяя контактные инсектициды.

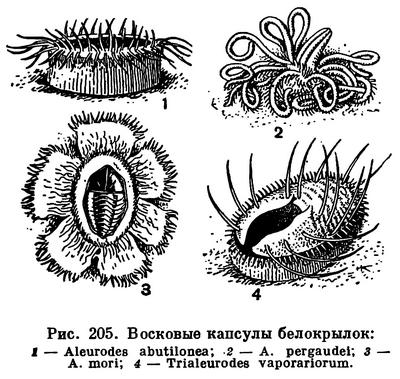
ПОДОТРЯД БЕЛОКРЫЛКИ (ALEURODOIDEA)
Белокрылки получили свое название потому, что их крылья, как и все тело, покрыты восковым порошковидным налетом, напоминающим муку. Эти нежные насекомые едва достигают 3 мм в длину. Взрослые самцы и самки складывают узкие, одинаково развитые крылья так, как это делают моли, что и делает их похожими на первый взгляд на мелких чешуекрылых (рис. 204). Если тронуть растение, на котором сидят крылатые белокрылки, они тут же легко взлетают, но стремятся скорее сесть где-нибудь на нижней поверхности листьев.
У белокрылок довольно сложное развитие. Яйца их с небольшим стебельком.
Выходящая подвижная плоская овальная 6-ногая личинка с двумя глазными пятнышками на голове отыскивает подходящее для питания место на растении и прижимается к его поверхности, поджимая под себя ноги. Восковые железы выделяют вокруг личинки восковой барьер, плотно прикрепляющийся к поверхности листа или коры. Уже после первой линьки у личинки сильно уменьшаются ноги и усики и она становится неподвижной. У личинки четвертого возраста меняется и форма тела — оно вздувается, находясь под защитой восковой капсулы (рис. 205). В это время личинка перестает питаться, в ее теле идет перестройка организации как в куколке, — закладываются крылья, длинные ноги, усики и другие органы взрослой белокрылки.
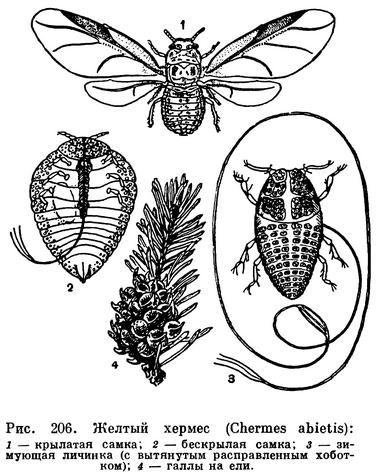
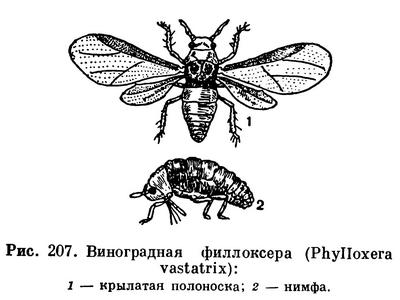
ПОДОТРЯД ТЛИ (APHIDODEA)
Тли, бесспорно, очень интересная группа насекомых и по своей исключительно разнообразной биологии, и по обилию видов, и по тому значению, которое они имеют как вредители. Это мелкие нежные насекомые, обычно сидящие большими колониями либо на надземных частях растений, либо на корнях. Многие тли вызывают болезненные изменения тех частей растений, на которых сидят, вызывая скручивание листьев, деформацию концов побегов, а иногда и образование полых внутри выростов — галлов. Многие тли выделяют сладкие экскременты, называемые медвяной росой, благодаря чему их охотно посещают муравьи, питающиеся этими выделениями. Не все тли выделяют медвяную росу. Ее не выделяют виды, тело которых покрыто восковым пушком, и обитатели галлов.
Причина выделения медвяной росы состоит в том, что покровы у тлей нежные и через них легко испаряется вода. Поэтому, чтобы не погибнуть от высыхания, тли много «пьют», поглощают много соков растений, на которых питаются. А питье тли — это и пища; сока тля поглощает больше, чем ей нужно для питания, избыток неусвоенных растворенных в соке растений сахаров выводится в непереваренном виде.
В жарких сухих районах, например в пустынях, тли выделяют гораздо больше медвяной росы, чем во влажных местностях. Поэтому понятно, что защищенные от высыхания восковым налетом или живущие внутри замкнутого галла тли не выделяют сахара — они меньше «пьют» сока, потребляя его лишь столько, сколько надо для питания.
У тлей очень сложный цикл развития. Наиболее типичный цикл развития с зимующими оплодотворенными яйцами. Весной из яиц выходят бескрылые самки, которые размножаются без оплодотворения (партеногенетически). Партеногенетически размножающиеся самки у большинства тлей яиц не откладывают, а рождают живых детенышей. Правда, это не настоящее живорождение. При настоящем живорождении (у млекопитающих, а также у скорпионов, перипатов) зародыш развивается за счет веществ, постепенно поступающих из организма матери. А у тлей наблюдается яйцеживорождение: партеногенетическая тля яиц не откладывает, а маленькая тля развивается из яйца за счет желтка, но только развитие протекает в организме матери.
Бывают тли, весь цикл развития которых протекает на одном виде растений (одноядные тли), бывают и такие, которые развиваются на разных видах растений (многоядные тли). Среди многоядных есть такие, для которых смена видов растений не обязательна, но у многих видов тлей происходит обязательная смена кормовых растений — это мигрирующие тли. В последнем случае тот вид растения, на котором перезимовывают яйца, называется основным кормовым растением. Вышедшие из яиц партеногенетические бескрылые самки называются основательницами. Иногда несколько бескрылых поколений потомства основательницы остается на том же растении, и лишь впоследствии среди них появляются крылатые тли (тоже партеногенетические самки), а иногда уже и первое поколение состоит из партеногенетических самок, перелетающих на другие растения. Это так называемые расселительницы. Переселяясь на новые кормовые растения, самки-эмигранты рождают бескрылых потомков, образующих новые колонии. Затем в потомстве бескрылых появляются снова крылатые особи, способные к дальнейшему расселению. Поколения тлей, развивающихся на дополнительных видах кормовых растений, т. е. на тех, на которые перекочевали расселительницы, называются переселенцами.
За лето многие тли дают несколько поколений. К осени среди переселенцев появляются так называемые полоноски, которые рождают самцов и самок. Оплодотворенные самки перелетают на те виды растений, на которых весной бывают основательницы, и откладывают зимующие, покрытые плотной оболочкой яйца.
Такой тип смены поколений у тлей очень обычен, но бывают и другие циклы. Разбор конкретных примеров может это хорошо иллюстрировать. Так как у тлей развивается несколько поколений в году, они размножаются с поразительной быстротой. Подсчитано, что в тех субтропиках, где около 300 теплых дней в году, потомство одной тли вырастает в 1013 раз! Конечно, такого обилия потомков никогда не бывает. Но практика борьбы с вредителями, например, садовых культур показывает, что, если не вести борьбу, тли могут почти сплошь покрывать растения, а от многих тлей происходит гибель возделываемых растений.
Одни из самых примитивных тлей — хермесы. У желтого хермеса (Chermes abietis), развивающегося на ели, крылатые особи появляются во второй половине лета и откладывают яйца на хвое у основания почек. Из яиц выходят зимующие основательницы, вызывающие образование галлов (рис. 206, 4). Этот вид, немигрирующий, партеногенетический, дает одно поколение в году. А очень близкий вид — зеленый хермес (Chermes viridis) — тоже перезимовывает в виде самок-основательниц на ели, но у него крылатые особи перелетают на лиственницу, где и откладывают яйца. Из них выходят тли, которые на коре лиственницы зимуют, а на следующий год кладут тут же яйца. Из яиц выходят крылатые тли-полоноски, а из яиц полоносок выходят самцы и самки. Оплодотворенные самки перелетают обратно на ель и кладут там яйца, из них выходят зимующие основательницы.
К хермесам близка виноградная филлоксера (Phylloxera vastatrix) — тля американского происхождения, которая у себя на родине вредного значения не имела, а попав в Европу, изменила свою биологию и стала опаснейшим вредителем, настоящим бичом виноградарства. Сперва она была завезена во Францию и разорила виноградное хозяйство этой страны, славящейся своим виноделием. Распространилась она и далее на восток. За 30 лет она погубила около 6 миллионов гектаров виноградников! У нас филлоксера есть не во всех районах возделывания винограда — только в Молдавии, на Правобережье УССР, на Северном Кавказе и местами в Закавказье.
Тли — очень богатая видами группа насекомых. Очень быстрое размножение, связанное с небольшой продолжительностью развития, способностью к партеногенезу, с живорождением, приводит к быстрым изменениям наследственных особенностей тлей. Отсюда и обилие их видов. В настоящее время описано около 20000 видов тлей, но их, несомненно, много больше.
У медяниц и тлей много черт регрессивного развития, связанных с большой плодовитостью, легкостью добывания пищи и малой подвижностью. Личинки тлей и бескрылые тли очень мало и неохотно передвигаются. Еще ярче эти черты выражены у червецов и щитовок.
ПОДОТРЯД КОКЦИДЫ (ЧЕРВЕЦЫ И ЩИТОВКИ) (COCCODEA)
Кокцид иногда довольно нелегко заметить, а человеку, незнакомому с их особенностями, трудно бывает признать в них насекомых. Действительно, самки многих щитовок выглядят как вздутия на коре, как мелкие чешуйки слущивающейся коры, как приставший к листу или коре комок мусора. Явно выраженная мимикрия самок кокцид — косвенный показатель их малой подвижности. Действительно, взрослые самки наиболее специализированных кокцид бывают совершенно неподвижны. Они прицепляются к кормовому растению длинным тонким хоботком, на всю жизнь втыкаемым в ткани растения, а глаза, усики, ноги у них полностью редуцируются; тело взрослой самки превращается в набитый яйцами мешок.
Очень многие самки кокцид бывают защищены восковыми выделениями, которые у таких совершенно неподвижных самок образуют плоский, похожий на чешуйку более или менее округлый или запятовидно вытянутый щиток (рис. 214). Отсюда и название таких кокцид — щитовки (сем. Diaspidae). Эти плоские, похожие на чешуйки щитки состоят не только из восковых выделений насекомого, но включают и многие шкурки личинки.
У некоторых кокцид воскового щитка нет, но все тело самки превращается в выпуклый щиток — под защитой плотных твердых покровов спинной поверхности насекомого созревают яйца, в них развиваются личинки. Самки таких кокцид бывают похожи на наросты или вздутия коры; они окрашены в тон коры. Это так называемые ложнощитовки (сем. Lecaniidae, сем. Kermococcidae, рис. 215). Иногда щиток не полностью прикрывает яйца, яйца откладываются под защитой мешочка из восковых нитей, с одной только стороны прикрываемого похожей на чешуйку постепенно высыхающей самкой. Это так называемые подушечницы (Pulvinaria, рис. 215). Самки некоторых кокцид тоже откладывают яйца в восковых мешках, но ползают, таская такой мешок (овисак) за собой. Тело самок таких кокцид бывает овальноудлиненное с эластичными, нетвердыми покровами, голова обычно с хорошо развитыми усиками, с явственными глазами. Ноги хорошо развитые, лапка из 1 или даже 2 члеников, но всегда с 1 коготком — признак, характерный для всех кокцид. Такие кокциды называются червецами (рис. 216). К ним принадлежат наиболее примитивные семейства (Ortheziidae, Margarodidae).
Кокциды — это единственная группа наземных животных, переходящая к совершенно неподвижному образу жизни. Только питание непрерывно приносимыми к хоботку соками растения обеспечивает на суше возможность прикрепленного образа жизни. Переход к неподвижности связан с утерей органов передвижения и органов ориентировки в пространстве. Это хорошо прослеживается на кокцидах.
Тело у них утрачивает свойственную членистоногим двустороннюю симметрию, становясь округлым, почти радиально-симметричным, утрачивает сегментацию.
У кокцид, как и всегда у сидячих форм, развиваются пассивные защитные приспособления — либо выделения воскового щитка (у щитовок), либо затвердевание спинных покровов самок (у ложнощитовок). Как всегда, с утратой подвижности развиваются и покровительственная форма и окраска.
Кокциды — группа насекомых, интересная своими биологическими особенностями и имеющая огромное практическое значение. Но эта группа очень трудна для изучения — большинство признаков, важных даже для определения вида насекомого (а это крайне существенно, так как из двух сходных видов один может быть опасным вредителем, а другой — безвредным для нас обитателем неиспользуемых растений), требует изготовления микроскопических препаратов и солидной подготовки.
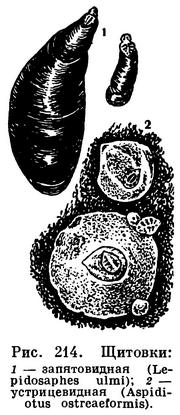
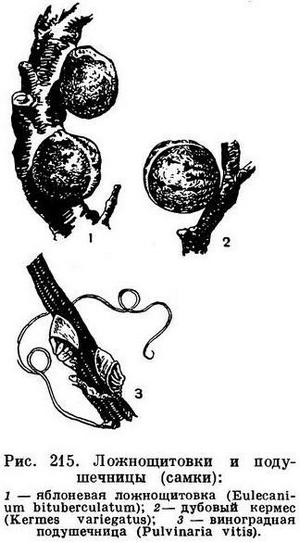
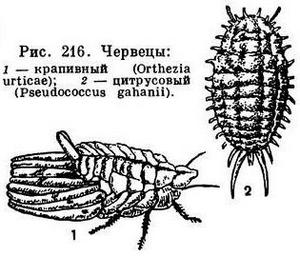
ОТРЯД ПОЛУЖЕСТКОКРЫЛЫЕ, ИЛИ КЛОПЫ (HEMIPTERA)
Полужесткокрылые, или клопы, представляют самый крупный отряд насекомых с неполным превращением. В настоящее время известно свыше 30000 видов клопов, распространенных по всему земному шару. Свое название полужесткокрылые они получили благодаря своеобразному строению передних крыльев, резко отличающихся от задних, имеющих вид прозрачных перепонок с небольшим количеством жилок. Передние крылья превращены в надкрылья, неоднородные по степени их хитинизации. Основная часть надкрылья состоит из твердого хитина, в то время как вершинная часть перепончатая и жилки на ней хорошо заметны.
Некоторым видам клопов свойствен так называемый крыловой диморфизм, сущность которого заключается в том, что наряду с полнокрылыми особями с вполне развитыми крыльями и надкрыльями у них имеются и короткокрылые особи. Так, у самцов соснового подкорного клопа (Aradus cinnamomeus) надкрылья развиты нормально, вторая пара крыльев отсутствует, самки же встречаются в виде двух форм — длиннокрылой и короткокрылой (рис. 219). У длиннокрылой самки развиты обе пары крыльев, и она способна к полету, короткокрылая же самка летать не может, так как у нее укорочены надкрылья и полностью отсутствуют крылья. Значительно реже случаи полной утраты крыльев клопами, как это имеет место, например, у постельного клопа (Cimex lectularius).
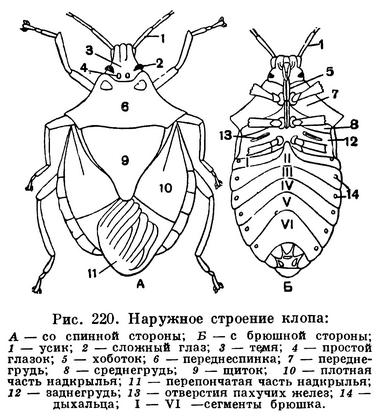
Помимо структуры надкрылий, характерным признаком, свойственным всем представителям этого отряда, является их колюще-сосущий ротовой аппарат, имеющий вид хоботка, отходящего от переднего края головы и не срастающегося с переднегрудью (рис. 220). Основную часть хоботка представляет сильно вытянутая членистая нижняя губа, образующая на внутренней стороне глубокий желобок, в котором помещаются сильно измененные верхние и нижние челюсти, имеющие вид тонких и длинных щетинок. Сверху хоботок прикрывается относительно недлинной верхней губой (рис. 221).
В связи с такой специфичной структурой ротового аппарата находится характер питания полужесткокрылых: прокалывая покровы тела животного или растения, клопы высасывают кровь или клеточный сок. При этом прокол делается верхними челюстями, а высасывание происходит при помощи нижнечелюстных щетинок, которые при погружении в тело жертвы, плотно смыкаясь, образуют два канала, располагающихся друг над другом. По верхнему каналу, более широкому, насасывается жидкая пища, а по нижнему стекает выделяемая клопом слюна. Нижняя губа в ткань не проникает и играет роль футляра, защищающего щетинки. У растительноядных клопов хоботок обычно длинный и тонкий, в спокойном состоянии он подогнут под тело и скрыт в особом желобке, расположенном на голове и груди; у хищных видов хоботок, наоборот, короткий, но толстый и сильный, дугообразно изогнутый в виде клюва.
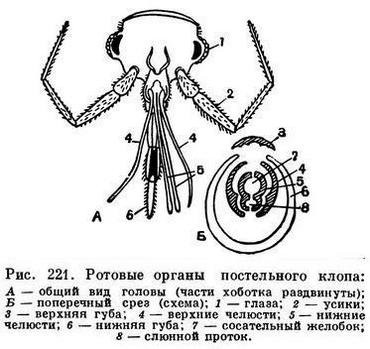
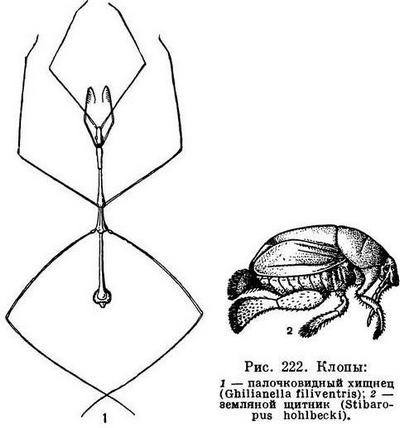
Размеры и форма тела у полужесткокрылых крайне изменчивы. Наряду с мелкими видами, длиной меньше 1 мм, есть очень крупные клопы, достигающие 10 см. Форма тела часто зависит от образа жизни клопов и характера тех условий среды, в которых они обитают. Обычно тело умеренно уплощенное, в той или иной мере округленное, с плоско сложенными на спине крыльями. Иногда оно принимает палочковидную форму, причем это имеет место в различных семействах. Южноамериканский палочковидный хищнец (Ghilianella filiventris) по внешнему виду скорее напоминает палочника, чем клопа. В то же время имеются виды с почти правильной шаровидной формой; последняя характерна преимущественно для клопов, обитающих в толще песка на дюнах или на бугристых песках в пустынях. У таких псаммофильных видов из семейства земляных щитников (Cydnidae) шаровидная форма тела сочетается с копательными конечностями, своеобразно приспособленными к передвижению в толще сыпучего песка (виды p. Stibaropus и др.). Сильное уплощение тела наблюдается у паразитических клопов (паразиты летучих мышей, птиц и человека), а также у видов, обитающих под корой деревьев, в ходах короедов и под другими укрытиями.
В окраске полужесткокрылых часто преобладают яркие цвета, находящиеся в контрастных сочетаниях (черный, желтый, красный и т. д.), что делает их заметными на окружающем фоне. Подобная окраска характерна, например, для тощего клопа (Lygaeus equestris, табл. 37, 2) и солдатика (Pyrrhocoris apterus, табл. 37, 3). Тощий клоп ярко-красного цвета с черным рисунком и белыми пятнами на верхней стороне тела, усики и ноги черные; длина 10—12 мм. Уже с ранней весны его можно встретить в больших количествах на стенах домов, на коре деревьев и на различных травянистых растениях. Солдатик приблизительно такого же размера, красного цвета с черным брюшком и черным рисунком на надкрыльях; надкрылья у него несколько укорочены, а крылья часто совсем отсутствуют.
Такая яркая окраска носит явно демонстрационный характер и хорошо сочетается с наличием у клопов пахучих грудных желез, выделяющих специфическую жидкость, с резким характерным «клоповым» запахом. Парные отверстия протоков этих желез открываются на заднегруди между местами причленения второй и третьей пар ног. Секрет, выделяемый железами, имеет кислую реакцию; его неприятный запах определяется цимициновой кислотой С15Н28O2. У клопов, обитающих в водной среде, а также у ряда хищных видов пахучие железы отсутствуют или слабо развиты.
Несмотря на то что многие виды полужесткокрылых встречаются в природе в больших количествах, плодовитость их самок в общем небольшая и колеблется в пределах от нескольких десятков яиц до 200. Яйца могут иметь овальную, цилиндрическую, веретенообразную или неправильно округлую форму. Их характерной особенностью является наличие на верхнем полюсе особой крышечки, открывающейся при выходе из яйца личинки первого возраста.
У некоторых клопов ярко выражена своеобразная забота о потомстве. В этом отношении интересен краевик-листовидка (Phyllomorpha laciniata), встречающийся на Кавказе и в Крыму. Его переднеспинка и брюшко расширены и несут листовидные полупрозрачные лопасти; усики, ноги и вся поверхность тела покрыты многочисленными длинными шипами. Охристо-желтая окраска и листовидная форма делают его похожим на высохший и растрескавшийся с краев листочек. Самка откладывает яйца на спинную поверхность самца, где они застревают между шипами и сохраняются там до вылупления личинок.
Число личиночных стадий у полужесткокрылых в преобладающем большинстве случаев пять, редко четыре. Постэмбриональное развитие продолжается от нескольких недель до двух лет. Личинки по внешнему виду похожи на взрослых, от которых отличаются прежде всего меньшими размерами и рядом морфологических признаков (отсутствием простых глазков, меньшим количеством члеников усиков и лапок и др.).
Зачатки крыльев появляются у личинок третьего возраста; с каждой линькой они увеличиваются в размерах, но даже и у личинок пятого возраста они занимают боковое положение и не соприкасаются по внутреннему краю. Личинки полужесткокрылых тоже имеют пахучие железы, в количестве 1—3 пар, однако протоки этих желез открываются на спинной поверхности брюшка.
Наиболее древние находки насекомых, идентичных современному отряду полужесткокрылых, относятся к мезозойской эре, где они в основном были представлены специфическими семействами, не встречающимися позднее ни в третичной, ни в современной фауне. В третичном периоде, по крайней мере уже с нижнего олигоцена, обособляется большинство современных семейств клопов.
В настоящее время насчитывают приблизительно 50 семейств, из которых около 40 (свыше 2000 видов) встречаются в СССР. Преобладающее же большинство видов населяет преимущественно тропические и субтропические области земного шара.
Среди клопов много видов хищных или со смешанным питанием, но преобладают растительноядные формы. Некоторые из них являются опасными вредителями сельскохозяйственных лесных растений (табл. 37).
Большой вред зерновым культурам наносят клопы-черепашки (семейство Scutelleridae). Представители этого семейства характеризуются сильно хитинизированной переднеспинкой, позади которой располагается выпуклый щиток, достигающий вершины брюшка и прикрывающий большую часть надкрылий. Внешнее сходство тела с панцирем черепах послужило основанием для русского названия этих клопов. Семейство включает около 600 видов, из которых в СССР встречается 55.
Много вредителей сельского хозяйства имеется в семействе щитников (Pentatomidae).
Среди серьезных вредителей огородов особо выделяется группа так называемых крестоцветных клопов (виды рода Eurydema, табл. 37, 5,6,7). Они сходны по биологии, но отличаются характером географического распространения. Рапсовый клоп (Eurydema oleracea) встречается в СССР почти повсюду, кроме северных областей; горчичный клоп (Eurydema festiva) не распространяется севернее 55° с. ш.; в Сибири его сменяет сибирский крестоцветный клоп (Eurydema gebleri); разукрашенный клоп (Eurydema ornata) вредит в степной и лесостепной зонах, а среднеазиатский капустный клоп (Eurydema maracandica) — в среднеазиатских республиках.
На юге, например на Украине, в Крыму и в Молдавии, часто можно встретить грушевую кружевницу (Stephanitis pyri), относящуюся к семейству кружевниц (Tingitidae). У нее черное тело длиной всего около 3 мм, с желтоватыми ногами и усиками. Характерны широкие прозрачные надкрылья этого клопа: их многочисленные жилки образуют тонкую сеточку. Этот вредитель обитает на яблоне, груше и других плодовых деревьях, а также на липе. После спаривания, которое происходит весной, самка откладывает яйца в ткань листьев с нижней стороны, кучками по 16—20 штук. Развитие яиц длится 20—30 дней, а развитие личинок — 20—25 дней. В течение года может давать 2—3 поколения. Поврежденные в результате сосания клопами и их личинками листья яблонь и груш усыхают, что приводит к недоразвитию плодов. Зимуют взрослые клопы под опавшими листьями или в щелях отмершей коры деревьев.
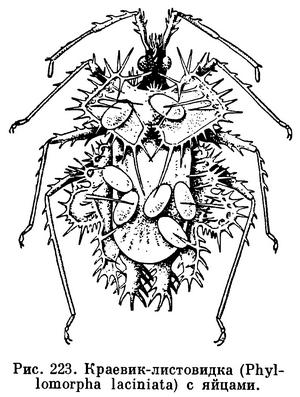

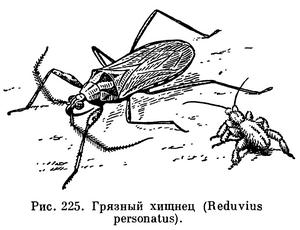
Типичными хищными полужесткокрылыми являются представители семейства хищнецов (Reduviidae). Это обычно крупные клопы, реже небольших размеров, с более или менее цилиндрической головой, вытянутой в длину и продолжающейся позади глаз в виде шеи; переднеспинка делится перетяжкой на более узкую переднюю и широкую заднюю части. Большинство видов имеет нормально развитые крылья, хотя встречаются также короткокрылые и бескрылые формы. Ноги у хищнецов, особенно задние, относительно длинные, однако они передвигаются медленно. Питаются различными насекомыми. Всего известно свыше 3000 видов, из которых в СССР встречается около 90.
Особое место среди полужесткокрылых занимают так называемые постельные клопы(семейство Cimicidae), характерным признаком которых является круглое или овальное, сильно уплощенное тело с резко укороченными надкрыльями. Название, данное этим клопам, не точно: собственно «постельных», т. е. тесно связанных с жильем человека, среди них не так много — всего несколько видов. Остальные же виды этого семейства (их около 30) — паразиты, сосущие кровь позвоночных животных (летучих мышей, пищух, птиц).
Широкому и быстрому распространению постельного клопа способствует и то, что, помимо человека, он нападает также на летучих мышей, некоторых мышевидных грызунов, разных птиц, в том числе на кур.
До сих пор мы знакомились с полужесткокрылыми, жизнь которых протекает в наземной обстановке. Но есть некоторые семейства этого отряда, которые в своем развитии связаны с водой и обладают изумительными приспособлениями к обитанию в водной среде. Именно поэтому они являются излюбленными объектами наблюдений на школьных экскурсиях.
Среди водных клопов (табл. 38) особенно богато видами семейство гребляков (Corixidae), которых в мировой фауне насчитывается около 600 видов. Некоторые из них в своем распространении заходят за северный полярный круг и встречаются в тундре. Гребляки — сравнительно мелкие клопы с широкой головой и относительно уплощенным телом. Самый крупный наш представитель этого семейства — Corixa dentipes, распространенный в лесной зоне, достигает в длину 13—16 мм; есть мелкие виды, длиной всего около 2 мм (Micronecta minutissima).
Яйца гребляков имеют более или менее шарообразную форму и слегка заострены на одном из полюсов (рис. 227). Самка прикрепляет их при помощи особого клейкого вещества к растениям и другим подводным предметам, располагая небольшими кучками.
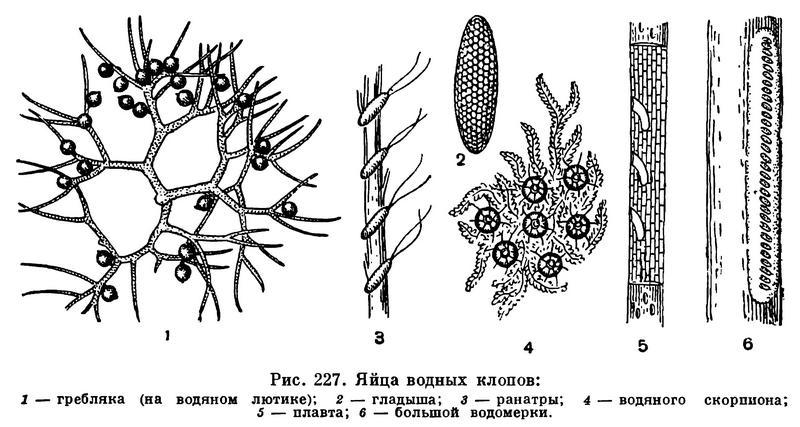
По внешнему виду гребляков напоминают гладыши (семейство Notonectidae), которые хотя и представлены у нас ограниченным числом видов (6—7), однако являются формами, обычными в наших пресных водоемах. Чаще всего встречается обыкновенный гладыш (Notonecta glauca, табл. 38, 1). Взрослый клоп достигает в длину около 1,5 см. Характерна форма тела гладыша: снизу оно плоское, а сверху выпуклое и благодаря этому немного напоминает лодку, перевернутую вверх дном. Тело зеленовато-белое с буроватыми или желтоватыми надкрыльями.
Наряду с гребляками и гладышами в стоячих, густо заросших подводными растениями водоемах часто встречается плавт (Naucoris cimicoides, табл. 38, 2) — представитель семейства плавтов (Naucoridae), распространенного преимущественно в тропиках. У плавта широкое, слабо выпуклое тело яйцевидной формы, длиной около 1,5 см; он зеленовато-желтого цвета с блестящей переднеспинкой, покрытой бурыми точками. У плавта сильно изменены передние конечности, представляющие собой очень подвижный хватательный аппарат, хорошо приспособленный для ловли жертв, какими могут быть личинки насекомых, моллюски и мелкие рыбки. Задние конечности плавта плавательные, покрытые волосками. Однако плавт не только хорошо плавает, но и ползает по растениям.
Весьма своеобразны клопы, относящиеся к семейству водяных скорпионов (Nepidae). У них, так же как у плавтов, передние ноги превращены в хватательные конечности, но две остальные пары являются ходильными. Это определяет характер их движения — водяные скорпионы медленно передвигаются, цепляясь коготками за растения. На заднем конце тела у них имеется длинный отросток, представляющий собой дыхательную трубку, состоящую из двух обращенных друг к другу желобков. Конец этой дыхательной трубки клоп выставляет из воды и при помощи ее набирает воздух под надкрылья. Яйца водяных скорпионов снабжены длинными нитевидными придатками, через которые внутрь яйца транспортируется запас воздуха. Водяные скорпионы известны уже из верхнеюрских отложений. В настоящее время насчитывают около 150 видов этих насекомых, распространенных преимущественно в тропических областях. В фауне СССР встречается 5—6 видов, из которых широко распространены водяной скорпион (Nepa cinerea, табл. 38, 4) и ранатра (Ranatra linearis, табл. 38, 7). Оба они встречаются в водоемах со стоячей или медленно текущей водой, заросших растениями.
К водяным скорпионам близко семейство белостоматид (Belostomatidae), которые отличаются тем, что задние ноги у них плавательные, а не ходильные. Хотя у них тоже имеются дыхательные трубочки, но последние едва выступают за конец брюшка или совсем скрыты. Белостоматиды — гиганты среди клопов: среди них есть формы, достигающие 10 см. Питаются они мальками рыб и головастиками. Своеобразна забота о потомстве у этих клопов: самка откладывает яйца на спинную поверхность самца, где они плотно приклеиваются к надкрыльям и таким образом сохраняются здесь в течение довольно долгого времени (10—12 дней) до вылупления личинок. Белостоматиды обитают в основном в тропических странах. В фауне СССР они представлены 2 видами, живущими на Дальнем Востоке.
Несколько семейств клопов, объединенных под общим названием «водомерок», приспособились к обитанию на поверхности водного зеркала, по которой они передвигаются с не меньшей легкостью, чем любые сухопутные насекомые по земле. Примером могут служить представители семейства настоящих водомерок (Gerridae).
С водоемами связаны и клопы-прибрежники (семейство Saldidae); они живут по берегам рек, ручьев, озер и морей. Их можно встретить около растений, растущих у воды, на крупных валунах рек, или бегающими по илу и грязи на отмелях. Это мелкие хищные формы, длиной 2—8 мм, обычно окрашенные в более или менее темные цвета, с широкой головой, несущей крупные выпуклые глаза. Они быстро двигаются, хорошо летают и делают большие прыжки. Одним из самых обычных видов, распространенных повсеместно в Европейской части СССР, является прыгающий прибрежник (Saldula saltatoria), черный со светлыми пятнами на надкрыльях, длиной 3,5—4,5 мм.

ОТРЯД ПУЗЫРЕНОГИЕ, ИЛИ ТРИПСЫ (THYSANOPTERA, ИЛИ PHYSOPODA)
Трипсов легко отличить от всех остальных насекомых вследствие своеобразного строения их головы, крыльев и ног. Это крошечные насекомые с удлиненным телом, обычно не превышающим 0,5—1,5 мм. Только в тропических странах известны трипсы длиной до 1 см. Голова трипса отличается сильно скошенным лбом и треугольной нижней губой, благодаря чему напоминает конус, обращенный своей вершиной вниз и назад. Усики короткие с небольшим количеством члеников (6—9).
Ротовой аппарат колюще-сосущий; в нем имеются три колющие щетинки, которые двигаются по желобку, находящемуся на внутренней стороне нижней губы. Эти щетинки представляют собой измененные внутренние лопасти двух нижних челюстей и верхнюю левую челюсть. Верхняя правая челюсть у них недоразвита. Щетинки являются не только колющим орудием; благодаря желобчатым углублениям на парных (нижнечелюстных) щетинках они представляют собой сосательную трубку. Такое строение ротового аппарата определяет характер питания трипсов. Большинство из них питается соками растений. При этом покровы растения прокалываются непарной щетинкой, после чего рана углубляется парными щетинками и по образуемой ими сосательной трубке высасываемые соки поступают в пищеварительную полость насекомого. Однако не все трипсы растительноядны, есть среди них и хищники, питающиеся соками других мелких насекомых и клещиков.
Весьма своеобразен летательный аппарат этих насекомых: обе пары крыльев у них узкие, длинные, усаженные по краю бахромой ресничек, отчего трипсов часто называют бахромчатокрылыми (Thysanoptera). Жилок на крыльях очень мало: кроме жилки, идущей по краю крыла, имеется всего две или одна продольная и несколько поперечных. У ряда видов крылья могут быть укороченными или даже совсем отсутствовать. Иногда один и тот же вид может встречаться в природе в виде трех форм: длиннокрылой (forma macroptera), короткокрылой (forma brachyptera) и бескрылой (forma aptera).
Есть у трипсов и еще одно название — пузыреногие (Physopoda), которое характеризует особенность строения их ног: ноги укорочены, с утолщенными бедрами, их лапки состоят всего из 1—2 члеников, коготки маленькие, в виде двух соединенных между собой пластинок, между которыми располагается пузыревидная присоска. Этот присасывательный аппарат приводится в движение особыми мышцами. При поднятой ноге насекомого оболочка присоски втянута внутрь и имеет форму бокала; при наступании ноги на субстрат оболочка расправляется и в присоску нагнетается из тела жидкость, туго наполняя ее. Такая конструкция ноги обеспечивает быстрое передвижение трипса по растению или по почве с фиксацией тела на субстрате в момент прекращения движения. Характерно, что некоторые трипсы могут делать довольно резкие прыжки, отталкиваясь при этом брюшком.
У трипсов хорошо выражен половой диморфизм. Как правило, самцы у них отличаются от самок менее крупными размерами и более стройным телом; могут они также отличаться и окраской тела. Что касается укорочения крыльев, то оно по-разному проявляется у разных видов: в одних случаях короткокрылыми или бескрылыми бывают самцы, в других — самки; иногда оба пола бывают частью с недоразвитыми, частью с вполне развитыми крыльями.
Благодаря своей тесной связи с растениями, на которых у большинства трипсов протекает их жизнь, ими заселена вся суша. Наибольшее количество представителей отряда падает на тропики и субтропики; по мере приближения к полюсам число видов этих насекомых значительно уменьшается. В настоящее время известно свыше 2000 видов трипсов, из которых в СССР встречается всего около 230.
ПОДОТРЯД ЯЙЦЕКЛАДНЫЕ ТРИПСЫ (TEREBRANTIA)
Характерной чертой, отличающей яйцекладных трипсов, является строение их яйцеклада. Он состоит из четырех сильно зазубренных лопастей, внутри которых имеется желобок для прохода яйца. Действуя яйцекладом, как пилой, насекомое пропиливает им покровную ткань растения и откладывает под нее свои яйца, имеющие характерную бобовидную форму.
У некоторых яйцекладных трипсов — представителей особого семейства Aeolothripidae, виды которого ведут хищный образ жизни, яйцеклад загнут кверху. Как взрослые, так и их личинки, нападая на тлей или других мелких насекомых, в том числе и на трипсов, прокалывают их своим хоботком и высасывают у них соки.
Представители другого семейства — Thripidae — преимущественно растительноядные формы; среди них много вредителей сельского хозяйства. Примером может служить табачный трипс (Thrips tabaci), являющийся серьезным вредителем табака, а также и других культурных растений (хлопок, картофель, лук и др.).
Многие другие виды трипсов, относящихся к семейству Thripidae, подсасывая листья, цветки или семена, наносят существенный вред, снижая урожайность свеклы, люцерны, клевера, гороха, льна, овса, ржи и других культурных растений.
ПОДОТРЯД ТРУБКОХВОСТЫЕ ТРИПСЫ (TUBULIFERA)
У трубкохвостых трипсов конец брюшка вытянут в трубку, а яйцеклад отсутствует. В связи с этими морфологическими особенностями они не могут откладывать яйца внутрь растительных тканей и поэтому приклеивают их, часто кучками, к той или иной части растения (на листья, на колосья и т. д.) или откладывают в трещины коры. Среди трубкохвостых трипсов преобладают обитатели коры деревьев, встречающиеся обычно либо под корой отмерших и загнивших сучьев, стволов и пней, либо на лишайниках, произрастающих на деревьях. Представителей этого подотряда можно находить также во мху и под опавшими листьями.
Среди трубкохвостых трипсов также есть целый ряд вредных видов. Так, в СССР почти всюду распространен пустоцветный трипс (Haplothrips aculeatus). Он встречается на разных растениях, в том числе на ржи и кукурузе. Повреждая эти культуры, он вызывает так называемую «череззерницу». В последнее время на посевах злаков в Узбекистане серьезное значение как вредитель приобрел пшеничный трипс (Haplothrips tritici), резкое нарастание численности которого было вызвано освоением под посевы злаков больших массивов целинных земель.
ОТДЕЛ НАСЕКОМЫЕ С ПОЛНЫМ ПРЕВРАЩЕНИЕМ (HOLOMETABOLA)
ОТРЯД ВЕРБЛЮДКИ (RHAPHIDIOPTERA)
Верблюдки — очень небольшой отряд, включающий несколько видов, относящихся к 2 семействам.
В профиль силуэт переднего конца тела верблюдки (рис. 232) и впрямь отдаленно напоминает шею и голову верблюда, откуда и произошло название этих своеобразных представителей древней группы насекомых. У этих насекомых удлиненное тело, вытянутая голова и особенно характерна очень длинная переднеспинка. У верблюдок две пары сетчатых крыльев, почти одинаковых размеров с небольшими темными пятнами близ вершин — так называемыми птеростигмами, играющими роль стабилизаторов при полете. Полет верблюдок порхающий, при полете иногда слышен легкий треск. У самок на заднем конце тела имеется длинный яйцеклад.
Верблюдки — прожорливые хищники; как это часто бывает у хищников, у них хорошо развиты большие фасеточные глаза; кроме того, на лбу между глазами у некоторых по 3 простых глазка. Усики довольно длинные, нитевидные, при поисках пищи вибрирующие. Верблюдки охотятся, ползая по веткам, на различных медленно двигающихся насекомых — в основном на тлей и гусениц.
Спариванию верблюдок предшествуют длительные «игры», во время которых они иногда друг друга сильно кусают, после чего самец вводит в половое отверстие самки сперматофор. Вскоре после спаривания самки откладывают яйца.
Верблюдки — лесные жители. В наших лесах средней полосы, например в окрестностях Брянска, они откладывают яйца под кору деревьев, а в сухих байрачных лесах и полезащитных лесных полосах в степной зоне — в поверхностный слой почвы. Недели через две из яиц выходят личинки. Как и у всех насекомых с полным превращением, у личинок мало общего со взрослой верблюдкой, но все же по очертаниям головы и удлиненной, блестящей, сильно склеротизованной переднеспинке можно угадать будущую верблюдку.
Тело личинки верблюдки удлиненное, веретеновидное, сверху сплющенное, спинная поверхность средне- и заднегруди и брюшко мягкие с коричневатыми узорами (рис. 232), благодаря чему личинки незаметны на фоне коры или опавших листьев, где они охотятся на личинок двукрылых, гусениц, тлей и других насекомых. Проникая в ходы короедов, они уничтожают их личинок.
Верблюдки — широко распространенная и в Старом и в Новом Свете группа насекомых, но везде они не слишком многочисленны, причем связаны только с довольно влажными местами.
Ископаемые остатки верблюдок известны из пермских отложений, а в миоцене уже были известны представители современных родов. У нас встречается 2 семейства верблюдок. У Rhaphidiidae {настоящие верблюдки) между фасеточными глазами на лбу по 3 простых глазка, а у Inocelliidae (безглазки) простых глаз нет.
Практическое полезное значение верблюдок в отдельные годы бывает велико — их личинки уничтожают в лесах в массе размножающихся вредителей, в частности кладки непарного шелкопряда.
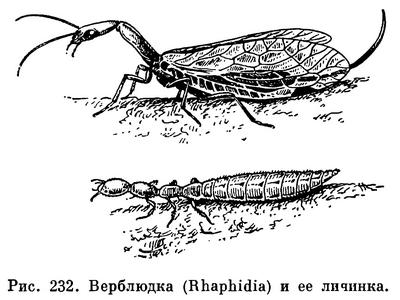
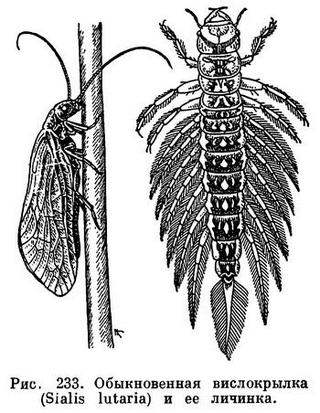
ОТРЯД ВИСЛОКРЫЛЫЕ (MEGALOPTERА)
Этот небольшой по числу видов отряд (всего несколько десятков видов) включает насекомых крупного или среднего размера с 4 сетчатыми однотипными крыльями; поперечные жилки крыльев немногочисленные. Усики щетинковидные, многочлениковые (более 40 члеников). Ротовые органы направлены вперед; фасеточные глаза крупные. Все лапки 5-члениковые. Личинки вислокрылок живут в воде и дышат растворенным в ней кислородом с помощью жаберных придатков брюшка.
В нашей стране встречаются представители семейства вислокрылок (Sialidae), относящиеся к роду Sialis. Обыкновенная вислокрылка (Sialis lutaria) черная с желтоватыми пятнами на голове и груди, с темными крыльями; в размахе крыльев достигает 30 мм. Другие роды этого семейства встречаются в Северной и Южной Америке, в Австралии, на Тасмании и Мадагаскаре — такое распространение, а также многие черты строения указывают на большую древность этого небогатого видами семейства.
В Северной Америке и в южном полушарии, особенно в тропиках, распространены представители другого семейства — коридалы (Corydalidae). Некоторые из них достигают очень крупных размеров. На Юнане и в других частях Юго-Восточной Азии встречается китайский коридал (Acanthocorydalis kolbei), в размахе крыльев достигающий 18 см. У коридалов замечателен половой диморфизм: у самок челюсти по длине такие же, как голова, а у самцов во много раз длиннее (рис. 234). У личинок коридалов и близких к ним хаулиодов (Chauliodes) жаберные выросты имеются на 8 члениках брюшка, а кроме того, у основания первых 7 выростов есть дополнительные пучки жаберных нитей.
Все личинки вислокрылых — хищники, в свою очередь охотно поедаемые рыбами, и могут рассматриваться как несомненно полезные насекомые.

ОТРЯД СЕТЧАТОКРЫЛЫЕ (NEUROPTERA-PLANIPENNIA)
Сетчатокрылые — хищные насекомые, во взрослом состоянии имеющие 4 прозрачных сетчатых крыла, иногда бесцветных, часто с пятнами. Обе пары крыльев могут быть одинаковыми или разными.
Разные сетчатокрылые ведут неодинаковый образ жизни и в общем больше отличаются друг от друга по образу жизни и по строению в личиночной стадии, чем во взрослом состоянии (рис. 235).
Личинки сетчатокрылых имеют несколько характерных признаков. Между жвалой и нижней челюстью с каждой стороны образуется желобок, через который личинка всасывает пищу: у личинок сетчатокрылых наружное пищеварение.
Мы сперва проглатываем пищу, а затем ее перевариваем, личинки же сетчатокрылых поступают наоборот: они, схватив добычу (обычно какое-либо мелкое мягкое насекомое), сперва прокалывают ее покровы заостренными жвалами, затем отрыгивают в ранку пищеварительные соки, а затем, как через трубочки, всасывают уже разжиженную пищу.
У личинок сетчатокрылых, многие из которых напоминают по виду личинок жуков, никогда не бывает нижнечелюстных щупиков — по этому признаку даже неопытный собиратель легко может определить принадлежность найденной им личинки к отряду сетчатокрылых. Очень любопытна одна особенность внутреннего строения личинок сетчатокрылых: у них между средней и задней кишкой образуется непроницаемая перегородка.
Это связано с тем, что у личинок сетчатокрылых непереваренные остатки пищи не выводятся, а накапливаются в течение всего развития. Только при превращении куколки во взрослое насекомое происходит соединение средней и задней кишки, и поэтому непереваренные личинкой остатки пищи выбрасываются уже взрослым насекомым.
В задний отдел кишечника личинок открываются мальпигиевы сосуды, выделения которых имеют вид шелковистых нитей, используемых для изготовления кокона при окукливании.
Одно из самых древних семейств — Dilaridae — характеризуется тем, что у самцов усики гребенчатые, а у самок крылья слаборазвитые и имеется длинный вытянутый яйцеклад. Эти отдаленно напоминающие небольших неуклюжих стрекоз насекомые распространены во всех частях света в теплых местностях, но везде немногочисленны. У нас в Крыму и на северо-западе Кавказа встречается турецкий дилар (Dilar turcicus), длинные мелкие личинки которого (рис. 235, 1) живут в почве и охотятся на личинок долгоносиков и других нежных насекомых. Очень сходная личинка американского вида Nallachius americanus живет в ходах в древесине. Несколько видов обитает в почве Средней Азии.

В почве же развиваются личинки распространенных в песках Австралии итонид (семейство Itonidae), напоминающие по внешности личинок хрущей, которыми и питаются. Сходство с личинками хрущей объясняется сходным способом передвижения в почве этих неутомимых хищников.
От жизни в почве личинки некоторых представителей сетчатокрылых перешли к жизни по берегам водоемов. У берегов водоемов под камнями и даже просто в воде встречаются несколько похожие на личинок диларов личинки осмилов (Osmylus chrysops, семейство Osmylidae, рис. 235, 3).
Еще больше приспособились к жизни в воде личинки семейства сизирид (Sisyridae). Личинки сизиры (Sisyra fuscata, рис. 235, 2) живут в воде, в колониях пресноводных губок — бадяг. Эти темные личинки имеют очень длинные острые ротовые части, которыми прокалывают клетки бадяги, высасывая их содержимое. Личинки то появляются на поверхности тела бадяги, то скрываются в полости ее тела, никогда не поднимаясь к поверхности воды. Дышат личинки сизиры растворенным в воде кислородом, причем дыхание осуществляется через поверхность не только всего тела, но и особых жаберных выростов, попарно расположенных на передних 7 брюшных сегментах. Дыхалец у личинок сизиры нет. Когда личинка вырастает, она оставляет воду, сплетает на выступающих из воды стеблях растений кокон и окукливается. Из куколки вылетает темно-коричневое насекомое с четковидными усиками.
Другие сетчатокрылые приспособились к прохождению личиночной стадии в более сухих условиях. Многие, кому приходилось внимательно осматривать листья деревьев в саду или лесу, замечали иногда на нижней поверхности листьев тех деревьев, на которых много тлей, маленькие яйца на длинных изящных стебельках. Эти яйца принадлежат очень привлекательным светло-зеленым стройным насекомым с длинными нежными блестящими сетчатыми крыльями и золотыми глазами, поэтому и семейство получило название златоглазки (Chrysopidae). Чаще других у нас встречается обыкновенная златоглазка (Chrysopa perla), тело которой длиной всего около 1 см, а крылья, если их расправить, простираются в размахе на 3 см. Летают златоглазки мало и днем неохотно, но ночью часто прилетают на свет. При прикосновении к ним оказывается, что они издают неприятный запах.
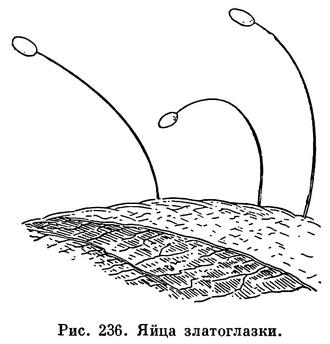
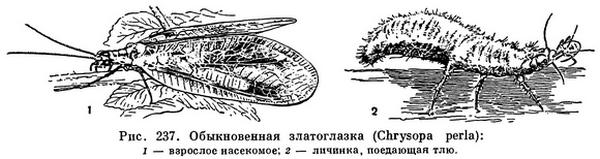
Что выходящие из яиц личинки (рис. 235, 6, 237, 2) свирепые хищники, видно сразу — у них длинные серповидно изогнутые и заостренные челюсти. Питаются они в основном малоподвижными мягкими насекомыми (тлями, медяницами и др.) и клещами.
Личинки ведут открытый образ жизни. С потерей влаги они борются двумя путями. Во-первых, высасывая много сочных насекомых, например быстро опустошая колонии тлей, они компенсируют расход испарившейся воды. А во-вторых, большинство личинок златоглазок покрывает свою спину шкурками высосанных тлей и другими частицами, что обеспечивает защиту от прямого действия солнечных лучей, сохраняя между чехликом и поверхностью тела прослойку влажного воздуха. Такой чехлик делает личинок и менее заметными для врагов.
Окукливаются личинки в белом паутинном коконе.
Удивителен образ жизни у так называемых муравьиных львов (семейство Myrmeleonidae). Муравьиные львы распространены в основном в тропиках и субтропиках, но такие представители, как обыкновенный муравьиный лев (Myrmeleo formicarius), довольно часто встречаются в степной зоне Европы и Азии и по песчаным участкам заходят много дальше на север.
Название свое эти насекомые получили благодаря своеобразному образу жизни хищной личинки. Личинки (рис. 235, 7) живут в сухом песке, вырывая в нем воронкообразную ямку, на дне которой они зарываются так, что только челюсти едва выступают из песка. Тело личинки расширенное и на спинной стороне покрыто бугорками, позволяющими личинке быстро закапываться своим задним концом в песок. Ямку личинка всегда делает в защищенном от дождя месте, с тем чтобы края ее всегда были покрыты сухим песом. Маленькое насекомое, пробегающее по сухому осыпающемуся краю воронкообразной ямки, например муравей, скатывается на дно и становится добычей «льва». Окукливаются личинки в песке, скрепляя шелковистыми паутинками отдельные песчинки и делая себе плотную колыбельку.
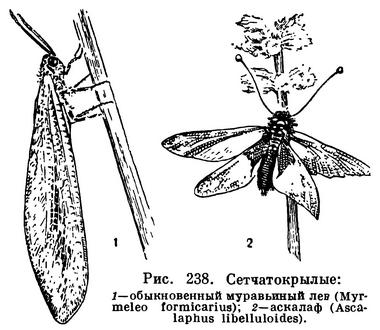
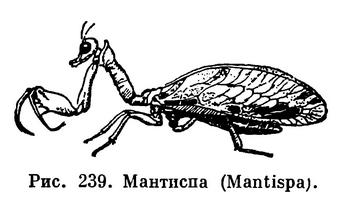
Не делая ловушек, охотятся и представители другого семейства сетчатокрылых, родственного муравьиным львам, — аскалафов (Ascalaphidae), распространенных в основном в тропиках и субтропиках. Аскалафы несколько похожи на бабочек, у них тело покрыто волосками, а булавовидные усики длиннее, чем у муравьиных львов. Все крылья у аскалафов примерно одинаковой величины, с яркими желтыми, черными или белыми пятнами. Личинка похожа на личинку муравьиного льва, но легко отличается наличием лопастевидных отростков по бокам брюшных сегментов. Личинки подкарауливают добычу, сидя под камнями, схватывая мощными челюстями проползающих насекомых.
Несколько видов аскалафов встречается и в СССР, например Ascalaphus macaronius в Крыму и на Кавказе.
Удивителен образ жизни у мантиспид (семейство Mantispidae). Взрослую мантиспу трудно спутать с каким-либо другим насекомым, так как у нее от длинной переднегруди отходят не обыкновенные ходильные ноги, а хватательные, похожие по своему строению на ноги богомолов, приспособленные к ловле различных насекомых. Самки мантиспы, например нашей Mantispa styriaca, осенью откладывают яйца, похожие на яйца златоглазки. Личинка вскоре выходит и не питаясь зимует. По виду личинка, вышедшая из яйца, похожа на личинок других етчатокрылых (рис. 235, 4). После зимовки, активно передвигаясь, личинка отыскивает кокон земляного паука (Lycosa), забирается в него, линяет, несколько изменяя свою форму, после чего начинает питаться паучьими яйцами. Ничего не подозревающая самка паука продолжает охранять свой кокон с яйцами, хотя потомство в нем энергично пожирается хищником. Через некоторое время личинка снова линяет, приобретая форму, отдаленно напоминающую личинку хруща. Такая С-образная личинка доедает в яйцевом коконе паука его яйца или вылупляющихся паучат, после чего плетет внутри его кокона свой кокон, в котором окукливается. Куколка находится сперва внутри последней личиночной шкурки, затем выбирается из кокона, заползает в трещины почвы и там превращается во взрослое насекомое. Подвижность куколки сближает этих сетчатокрылых с верблюдками. Некоторые виды мантисп, обитающие в тропиках, развиваются не в яйцах пауков, а в гнездах пчел и ос.
Очень мелких представителей сетчатокрылых включает семейство пыльнокрылов (Coniopterygidae), похожих по виду на белокрылок. Веретеновидные или овальные личинки (рис. 235, 5) пыльнокрылов (например, Conwentzia) поедают мелких сосущих насекомых и известны как истребители филлоксеры.
В тропиках и в южном полушарии встречается еще ряд семейств сетчатокрылых, некоторые виды которых иногда имеют причудливое строение и во взрослом и в личиночном состоянии. Так, очень изящны представители семейства Nemopteridae, у которых задние крылья превращены в направленные косо назад веслообразные пластинки (рис. 240, 1). У личинок некоторых видов голова соединяется с туловищем с помощью длинного трубкообразного выроста переднегруди (рис. 240, 2). Представители австралийских миодактилид (семейство Myiodactylidae) в стадии личинки живут на нижней поверхности листьев эвкалиптов и имеют листовидное тело (рис. 241).
В целом сетчатокрылые представляют очень своеобразный и древний отряд насекомых, хорошо известный с пермского периода и особенно распространившийся с триаса.
Число видов этих разнообразных по образу жизни насекомых невелико — всего около 5000.
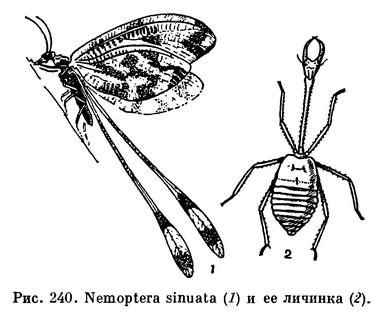
ОТРЯД СКОРПИОННИЦЫ (MECOPTERA)
Для представителей этого отряда, как и для сетчатокрылых, характерно наличие двух пар примерно одинаково развитых прозрачных сетчатых крыльев, чаще с неправильными пятнами.
От других насекомых скорпионниц довольно легко отличить по клювообразно вытянутому переднему концу головы, на вершине которого находятся очень мелкие ротовые части грызущего типа. У основания «клюва» помещаются довольно длинные нитевидные усики. По бокам головы имеются фасеточные глаза, а на лбу у большинства видов 3 простых глазка. Свое название «скорпионовые мухи», или «скорпионницы», эти безобидные для человека насекомые получили благодаря форме брюшка. Тонкое, вытянутое 10-члениковое брюшко у вершины загнуто кверху и у самцов утолщено, напоминая брюшко скорпиона.
Скорпионницы редко встречаются большими скоплениями. Распространены они во всех частях света.
Семейство настоящих скорпионниц (Panorpidae) представлено у нас панорпой (Panorpa communis), встречающейся в средней полосе повсюду, часто на полянках лиственных лесов или вблизи искусственных посадок. Эти средней величины насекомые (длина тела около 13 мм) держатся обычно в тени, во влажных местах. Тело панорпы темное, коричневое, крылья у сидящего насекомого плоско сложены на спине; их рисунок имеет вид неправильных темных поперечных полос. У самца утолщенный конец брюшка красного цвета.

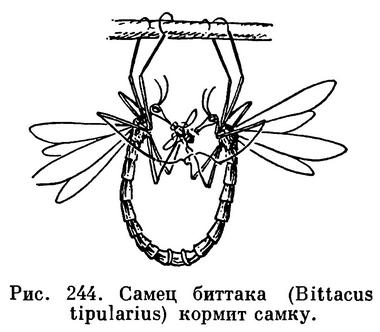
Представителей другого семейства скорпионниц — биттаков (Bittacidae) — легко узнать по очень тонкому телу и длинным ногам; общий облик их напоминает комаров-долгоножек, но наличие 2, а не 1 пары крыльев и «клюва» позволяет сразу распознать этих своеобразных насекомых.
Биттаки (Bittacus tipularius) встречаются во влажных лиственных лесах и на лугах. Они часто висят на растениях, зацепившись одной или двумя длинными ногами. Биттаки — хищники и в таком положении они подстерегают добычу, которую ловко схватывают своими, как ни странно, задними ногами, похожими на вывернутые передние ноги богомола. Добычей им служат разные мелкие насекомые и пауки.
Во время спаривания самец тоже кормит самку, но не слюной, как у панорп, а пойманной мушкой или другой мелкой добычей (рис. 244). Яйца у биттаков тоже откладываются в землю, а похожие на гусениц личинки (рис. 245) ползают по поверхности почвы под опавшими листьями, питаясь животной пищей.
В противоположность другим скорпионницам, представители семейства ледничников (Boreidae) — вегетарианцы, питаются мхами. Ледничники получили свое название потому, что взрослые насекомые встречаются ранней весной и поздней осенью, а во время оттепелей этих похожих на носатых кузнечиков насекомых можно встретить даже и на поверхности снега. Узнать их нетрудно — у самцов зимнего ледничника (Boreus hyemalis) характерны крючковатые зачатки недоразвитых крыльев (они не летают), а совсем бескрылые самки имеют довольно длинный вытянутый яйцеклад.
Отряд скорпионниц очень древний, близкие формы известны из пермских отложений палеозойской эры, а уже в мезозойскую эру встречались скорпионницы, похожие на современных.
Изучение ископаемых близких к скорпионницам форм и анатомии современных позволило установить черты их близости с ручейниками, чешуекрылыми и двукрылыми.
В настоящее время известно немногим более 300 видов современных скорпионниц, распространенных в разных частях света.
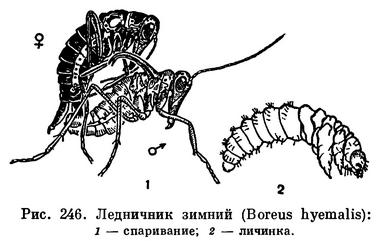
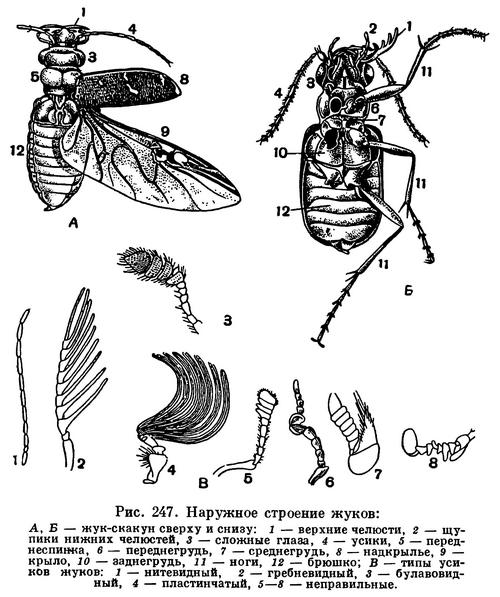
ОТРЯД ЖЕСТКОКРЫЛЫЕ, ИЛИ ЖУКИ (COLEOPTERA)
В названии отряда — жесткокрылые — отражен один из наиболее существенных признаков входящих сюда насекомых. Передние крылья, или надкрылья, у них очень жесткие и прочные; они прикрывают мягкую верхнюю сторону брюшка и расположенные здесь же перепончатые крылья второй пары. Именно эти перепончатые крылья служат для полета. Они намного длиннее надкрылий и в спокойном состоянии сложены и спрятаны под ними.
У некоторых групп жуков надкрылья иногда сильно укорачиваются и почти не прикрывают брюшко. Тогда перепончатые крылья либо еще сильнее складываются и все-таки прячутся под надкрыльями, либо лежат открыто на поверхности брюшка. Некоторые жуки, наоборот, утрачивают вторую пару крыльев и неспособны летать.
Этих жуков часто называют бескрылыми, хотя первая пара крыльев — надкрылья — у них всегда сохраняется. Надкрылья бескрылых видов иногда срастаются.
Фасеточные глаза отсутствуют лишь у тех видов, которые постоянно живут в темноте — в пещерах, термитниках, муравейниках. Впереди глаз прикрепляются усики, в огромном большинстве случаев состоящие из 11 члеников. Членики усиков могут быть однообразными, тонкими и длинными — тогда усики называются нитевидными. Иногда несколько последних члеников утолщаются и образуют округлую булаву (булавовидные усики). Иногда же, наоборот, они переходят в плоские боковые выросты, как это наблюдается у пластинчатоусых жуков. Если эти выросты не собраны в булаву, а имеются на всех члениках, усики напоминают гребешок и называются гребенчатыми.
Ротовые органы жуков грызущего типа. Основными орудиями размельчения пищи являются их верхние челюсти, называемые часто жвалами или мандибулами. Иногда они превращаются в украшения, достигая у самцов необычайного развития.
Надкрылья и крылья жуков прикреплены к средне- и заднегруди. Переднегрудь образует широкое кольцо, верхняя часть которого носит название переднеспинки. Снизу к трем грудным сегментам прикрепляются три пары ног, которые у жуков чрезвычайно разнообразны. Обычно они длинные, бегательные, у водных форм — плавательные, у обитающих в почве — копательные; иногда задние ноги увеличиваются в размерах, их бедра утолщаются — ноги становятся прыгательными. Заканчиваются ноги лапками, членики которых снизу несут подушечки, а у некоторых видов — присоски.
Обычно жуки ярко и красиво окрашены (табл. 39, 40), часто в металлически синие или зеленые тона. У многих видов имеются дополнительные скульптурные украшения в виде придатков на голове и усиках или своеобразных выступов и выростов на надкрыльях. Некоторые жуки формой тела и окраской подражают другим насекомым, чаще всего перепончатокрылым.
Тело личинок жуков состоит из 3 грудных и 10 брюшных сегментов, из которых десятый очень мал и нередко сдвинут на нижнюю сторону девятого. У них всегда хорошо развита голова, которая значительно сильнее хитинизирована, чем покровы тела, и окрашена в более темный цвет.
Окраска тела личинок (рис. 248) зависит от образа их жизни. Личинки, обитающие открыто на листьях растений или бегающие по поверхности почвы, окрашены в бурый, зеленый или черный цвета. Те же из них, которые живут в толще почвы или внутри тканей растений, имеют мясистое, белое тело. Подвижных уплощенных личинок жуков, снабженных длинными ногами и усиками, а также церками, называют камподеовидными, тогда как малоподвижных, толстых и внешне неуклюжих обитателей толщи различных твердых сред — эруковидными. Однако все многообразие личинок жуков не исчерпывается этими двумя примерами. Поэтому к отдельным группам личинок часто применяются названия, образованные от названий наиболее типичных семейств: куркулионоидные, скарабеоидные, церамбикоидные и т. д. личинки. Голова личинок снабжена парой сильных челюстей, которые могут быть как грызущими, так и колющими. У некоторых хищных личинок внутри колющих челюстей проходит канал, по которому в тело жертвы изливается пищеварительный сок. Разжиженные, полупереваренные ткани животного затем всасываются личинкой. Такой тип пищеварения называется внекишечным, хотя основные пищеварительные процессы всегда проходят в самом кишечнике.
Органы дыхания у личинок разнообразнее, чем у взрослых жуков. Некоторые обитатели водной среды дышат, например, при помощи трахейных жабр — специальных выростов тела, пронизанных трахеями.
За период развития личинки несколько раз линяют. Личинки разных возрастов обычно похожи друг на друга, но известны случаи сложного превращения, когда внешний облик личинок после линек совершенно меняется.
Наиболее своеобразна и сложна биология размножения маленького американского жука микромальтуса (Micromalthus debilis), выделяемого в особое семейство Micromalthidae. У него известно три личиночных возраста. Личинки первого возраста обладают длинными ногами и крупной головой. Они очень подвижны, но не питаются и, видимо, служат для расселения. После линьки они превращаются в мясистых безногих личинок второго возраста; основная их функция — питание; они грызут древесину и могут вредить. Накопив достаточно питательных веществ, эти личинки превращаются в личинок третьего возраста, которые не питаются, но зато способны к размножению и рождают множество личинок первого возраста. Известны у этого вида и взрослые жуки, которые развиваются из питающихся личинок второго возраста, хотя этот процесс изучен еще не полностью. У микромальтуса, таким образом, наблюдается единственный среди насекомых случай сочетания сложного гиперметаморфоза с размножением личинок — педогенезом (который встречается также у некоторых двукрылых из семейства галлиц).
Развитие личинок у жуков обычно завершается за несколько месяцев, реже растягивается на 3—5 лет. Куколки жуков, за редким исключением, свободного типа, их конечности не склеены с телом и сходны с конечностями взрослого насекомого.
Своеобразно проходит окукливание у некоторых личинок, имеющих длинное и тонкое тело. Живущие в гнилой древесине личинки древоедов (Eucnemidae) перед окукливанием готовят колыбельки, куда они могут поместить свое длинное тело, лишь сложившись вдвое. Такое странное положение личинок перед окукливанием характерно также для узкотелых златок. Постепенно все содержимое задней половины личинки перемещается в переднюю, и в конце концов из длинной и тонкой личинки образуется куколка, размеры которой точно соответствуют размерам приготовленной для нее колыбельки.
Жуки — самый разнообразный и богатый видами отряд насекомых. Уже сейчас их описано более 250000 видов, но ежегодно к этому числу добавляются тысячи ранее неизвестных форм. На территории СССР известно более 20000 видов жуков.
Отряд делится на два подотряда: плотоядные жуки (Adephaga) и разноядные жуки (Polyphaga).
Громадное большинство представителей отряда — обитатели суши, заселившие самые разнообразные местообитания; жуки встречаются в лесах и на лугах, в пустынях и болотах, в долинах и высоко в горах.
Трудно сказать, какая трофическая группа преобладает в отряде — потребители растительных тканей или хищники; паразитов среди жуков немного.
Основные запасы питательных веществ, обеспечивающих жизнь и размножение взрослой стадии, накапливаются личинкой. Однако для большинства жуков необходимо также дополнительное питание молодых самок и самцов, в процессе которого жуки окончательно созревают. Нередко вслед за откладкой первой партии яиц самка вновь начинает питаться, обеспечивая накопление необходимых запасов для дальнейшего размножения.
Необычайно велика приспособляемость жуков к самым различным условиям жизни.
Отсутствие воды в пустынях для многих животных является непреодолимым препятствием, однако и здесь жуки оказываются самой обильной группой среди обитателей этих пространств. Приспособления к строжайшей экономии воды позволяют пустынным жукам существовать за счет той воды, которая поглощается ими при питании живыми тканями растений, а также образуется в результате обмена веществ в их организме. Днем, спасаясь от палящих лучей солнца, жуки пустыни зарываются в почву, ночью же ведут активную жизнь.
В лесной зоне, наоборот, влаги вполне достаточно, но тепла не хватает, и поэтому солнечная погода является одним из наиболее важных условий дневной активности многих насекомых, в том числе и жуков. Лесные виды этого отряда примечательны в другом отношении — они смогли успешно приспособиться к развитию в тех средах, которые для большинства других насекомых оказались недоступными. Характерно, например, что в мертвой древесине, особенно на начальных этапах ее разложения, все ее обитатели почти исключительно личинки жуков. Интересно, что здесь встречаются не только те виды, чьи личинки способны питаться древесиной, основные компоненты которой — клетчатка и лигнин перевариваются с большим трудом. Многие жуки, обитающие в древесине, используют ее не как пищу, а только как среду для разведения грибков, которые специально заносятся в древесину самками при откладке яиц и в дальнейшем служат пищей личинкам.
У древоядных жуков выработались сложные инстинкты заботы о потомстве, которые по своему совершенству не уступают широко известным фактам из жизни жуков-навозников. Навозники запасают для своих личинок пищу в виде навозных колбас, шаров или груш строго определенной формы, которые изготовляются в специально выстроенных подземных камерах. Древоядные жуки разводят для личинок «грибные сады» на стенках ходов, специально выгрызаемых в древесине, а некоторые из них,например сахарные жуки (Passalidae), кормят личинок особым образом подготовленной древесиной, так как питаться сами личинки не могут.
Не менее совершенные инстинкты вырабатываются у жуков, приспособившихся к жизни за счет других животных, чаще всего за счет насекомых. Больше всего таких видов в муравейниках. Поведение мирмекофильных жуков таково, что муравьи не отличают их от других муравьев и часто выкармливают их личинок, как своих собственных. Они даже не препятствуют своим квартирантам тогда, когда жуки уничтожают яйца и личинок хозяев муравейника дополнительно к той пище, которую они получают непосредственно от муравьев.
У некоторых мирмекофильных жуков есть специальные железы, выделения которых поедаются муравьями. У представителей близкого к жужелицам семейства пауссид (Paussidae) повсюду на теле — на антеннах, лобной области, переднегруди, на надкрыльях и брюшке — расположены железы, выделяющие густую жидкость, привлекающую муравьев. Места, где открываются железы, покрыты густыми волосками, по которым секрет поднимается вследствие капиллярности и отсюда слизывается муравьями.
Нередко большой сложности достигают взаимосвязи жуков с растениями. Это выражается не только в высокой точности действий жуков при высверливании отверстия в орехе или при скручивании листа в «сигару» — все эти действия являются примерами заботы о потомстве, — но также и в способности личинок некоторых жуков стимулировать рост растительных тканей с последующим образованием галлов.
Галлы, вызываемые жуками, могут возникать в ответ на погрызы личинок, как это наблюдается у некоторых усачей, развивающихся в тонких ветвях деревьев к кустарников. Личинки слоников вызывают более крупные галлы, и, хотя образование этих галлов еще слабо изучено, их возникновение следует считать результатом не только механических, но и сложных биохимических воздействий личинок на растительную ткань.
Не менее интересны и многообразны приспособления жуков к защите от многочисленных врагов. Этой цели служат не только их прочные покровы и во многих случаях хорошо выраженная покровительственная окраска, маскирующая насекомое, но и биохимические средства защиты, которые также вырабатываются у многих жуков. Обычно ядовитые вещества, обладающие часто резким неприятным запахом, растворены в гемолимфе жуков и выделяются через сочленения тела тогда, когда жук бывает потревожен. Некоторые жуки, защищаясь от врагов, «выстреливают» едкой жидкостью, которая на воздухе быстро испаряется и отпугивает преследователя. У жуков-бомбардиров в этой жидкости содержатся окись азота и азотнокислые соли, и вся смесь при соприкосновении с воздухом взрывается.
Жуки, лишенные более эффективных средств защиты, при раздражении впадают в шоковое состояние, поджимают усики и лапки и падают вниз. Найти такого «притворившегося мертвым» жука нелегко.
Значение жуков в экономике природы и хозяйстве человека огромно. Ежегодно тратятся колоссальные средства на борьбу с вредителями сельского и лесного хозяйства. Гораздо меньше среди жуков полезных видов. Обычно это хищники, истребляющие вредных насекомых.
Коллекционирование жуков — одно из самых увлекательных занятий натуралистов. Вместе с бабочками эти оригинальные насекомые являются украшением коллекций и приносят любителям много приятных минут.
ПОДОТРЯД ПЛОТОЯДНЫЕ ЖУКИ (ADEPHAGA)
К этому подотряду относятся преимущественно хищные жуки, но есть и растительноядные формы. Важнейшие семейства подотряда — жужелицы, плавунцы и вертячки; кроме них, он включает несколько более мелких семейств.
ПОДОТРЯД РАЗНОЯДНЫЕ ЖУКИ (POLYPHAGA)
Этот подотряд гораздо обширнее первого. Как это отражено в названии подотряда, пищевые связи его представителей могут быть самыми разнообразными. Он включает основную массу жесткокрылых и делится на большое число семейств.
ОТРЯД ВЕЕРОКРЫЛЫЕ (STREPSIPTERА)
Веерокрылые — небольшой отряд мелких насекомых, сильно деградировавших в связи с переходом к расселению на стадии личинки и с паразитическим образом жизни.
Самцы веерокрылых (рис. 307, 1, 309, 1) заслуживают этого названия: у них задние крылья широкие, в расправленном состоянии веерообразные, а передние — редуцированы до небольших жужжалец. Ротовые органы у самцов атрофированы, лапки без коготков. Самки у веерокрылых иногда вообще непохожи на насекомых или напоминают личинок. Разные группы веерокрылых паразитируют на разных насекомых, но их циклы развития в общем довольно сходны.
Эоксены (Eoxenos) — паразиты живущих в муравейниках чешуйниц. Самки эоксенов остаются внутри личиночных и куколочной шкурок, образующих своеобразные коконы. Такие коконы встречаются под камнями в средиземноморских странах. Внутри кокона, вероятно, осуществляется и оплодотворение. Оплодотворенная самка зимует, а в середине следующего лета появляется масса своеобразных личинок первого возраста — триунгулинов (рис. 308, 1). Эти триунгулины как-то попадают на чешуйниц, внедряясь в них. В теле чешуйницы личинка становится червеобразной с зачаточными ногами, питается возле кишки. К августу личинки вырастают, линяют и оставляют чешуйниц, забираясь под камни. Осенью личинки окукливаются. Таким образом, у эоксенов самки часть времени проводят вне организма насекомого-хозяина.
Самки представителей семейства стилопсов (Stylopidae) никогда не оставляют тела насекомого-хозяина. Эти веерокрылые паразитируют в перепончатокрылых, клопах, цикадовых и прямокрылых.
Род Stylops паразитирует на диких пчелах, Xenos — на осах-полистах, Halictophagus и Elenchus — на цикадках.
В тропиках водятся «гигантские» веерокрылые — стихотремы (Stichotrema), паразитирующие на крупных (до 15 см) прямокрылых — сексавах на Адмиральских островах. Длина этих веерокрылых — 2—3 см.
По характеру развития и некоторым второстепенным признакам веерокрылые напоминают веероусых жуков (Bhipiphoridae). Поэтому раньше их сближали с жуками, а некоторые энтомологи даже считали веерокрылых семейством жуков.
Но за последние десятилетия ряд исследователей склонился к тому мнению, что этот специфический отряд ближе к перепончатокрылым, чем к жукам.
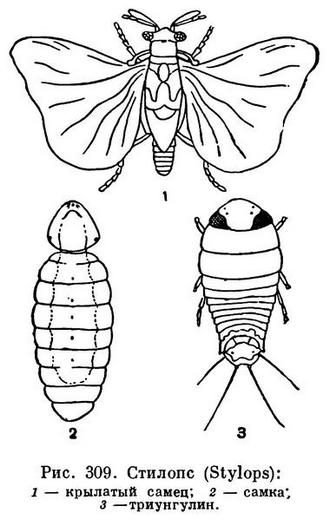
ОТРЯД РУЧЕЙНИКИ (TRICHOPTERА)
На дне многих пресных водоемов — чистых быстрых ручьев и заросших прудов — можно обнаружить удивительные существа, которые живут в трубчатых домиках, сооружаемых ими из различных мелких частиц, лежащих на дне. В зависимости от того, какие мелкие предметы лежат на дне, и в зависимости от вида насекомого домики могут быть построены из разного материала. У одних это сооружение из крупных песчинок, у других из камешков или раковинок мелких моллюсков, нередко это трубочка, состоящая из небольших обломков веточек или отмерших частей водных растении, и т. д. «Строительный материал» прочно скреплен паутинными нитями. Эти домики строят личинки ручейников.
Взрослые ручейники — довольно нежные насекомые, похожие на волосатых молей (рис. 310). Отличить ручейника от бабочки легче всего именно по крыльям — у бабочек крылья покрыты чешуйками, а у ручейников — волосками. В спокойном состоянии их темноокрашенные крылья складываются кровлеобразно на спине. Голова довольно крупная с фасеточными глазами и обычно с 3 простыми глазками между ними.
Усики длинные, нитевидные, ротовые органы редуцированы, в частности совсем нет жвал, а остальные ротовые части преобразованы в короткий хоботок с язычком. Взрослые ручейники не питаются, но могут пить воду. Ноги, заканчивающиеся 5-члениковыми лапками, довольно стройные. Эти в общем малозаметные невзрачные насекомые летают неохотно и вяло.
После спаривания самки ручейников откладывают в воду студенистые комочки яиц — «икру». Из яиц выходят личинки, которые у большинства видов с места в карьер начинают строить себе паутинный чехлик из шелковой нити, выделяемой видоизмененными слюнными железами. Чехлик инкрустируется подходящими мелкими частицами, лежащими на дне и доступными личинке. Включение в чехлик твердых предметов делает его прочнее и крепче. А надежная защита необходима личинке ручейника. Дело в том, что она никогда не выходит из воды и дышит всей поверхностью кожи всего удлиненного брюшного отдела тела. Брюшко личинок ручейников имеет не только очень тонкие, легко проницаемые (а раз так, то и легко ранимые) покровы, но нередко несет и многочисленные еще более нежные жаберные выросты, увеличивающие поверхность газообмена с водой. Пучки жабр бывают и на задних отделах груди.
Если вокруг все спокойно, личинка ползает по дну, таская на себе чехлик-домик. При движении личинка высовывает из чехлика голову и грудной отдел, на котором находится 3 пары довольно длинных и цепких выставленных вперед ног. Впрочем, передние ноги бывают нередко короче, чем остальные, а у некоторых личинок ручейников только две пары ног. Голова и грудные сегменты, выступающие из чехлика, имеют плотные покровы. Удивительна голова у личинок ручейников — на ней нет усиков. У личинок разных насекомых с полным превращением усики бывают разной длины, но редко они редуцируются до такой степени, что становятся совсем неразличимыми, как это бывает у личинок ручейников. Глаза у личинок имеют вид темных пятнышек и состоят из нескольких простых глазков (не более 6 с каждой стороны головы). Ротовой аппарат личинок в противоположность взрослым ручейникам хорошо развит, он грызущий. Личинки питаются и растительной пищей, соскабливая мягкие ткани зазубренными челюстями, и животной. Чехлик служит личинке ручейника не только постоянной броней, защищающей брюшко, но и убежищем: в случае опасности личинка вся втягивается в «домик», входное отверстие в который закрывает своей плотной и прочной гладкой головной капсулой. Задний конец тела личинки ручейника удерживается в чехлике с помощью пары направленных вперед мощных крючковидных отростков. Поэтому личинка может быстро спрятаться в чехлик. Удерживая домик крючками, личинка таскает его за собой, не теряя и только достраивая по мере роста.
Каких же личинок ручейников легко встретить в наших водоемах?
В быстрых ручьях с прохладной водой и каменистым дном под камнями легко обнаружить трубчатые домики стенофила (Stenophylax stellatus), сооруженные из аккуратно приделанных друг к другу крупных песчинок (рис. 311, 1). Личинка легко приподнимает свой домик, передний край которого капюшоном нависает над головой личинки, делая ее незаметной для проплывающих сверху рыб. Если повредить чехлик личинки, она сразу старается его починить, подбирая передними ногами песчинки нужной величины. Она прилаживает их к поврежденному краю чехлика, отбрасывает менее плотно прилегающие, пробуя и отбирая наиболее подходящие. Личинка приклеивает песчинки застывающей в шелковистую нитку слюной, многократно охватывает их нитями, связывая песчинки друг с другом, в результате чего чехлик оказывается очень прочным. После починки стенок домика личинка тщательно выстилает внутреннюю его поверхность несколькими слоями шелковой паутинки. Если личинку осторожно извлечь из чехлика и поместить в сосуд, на дно которого набросать вместо песка бисера, она сделает себе домик из мелких ярких бусинок. Питаются личинки стенофила и растительной и животной пищей.
В озерах, в которые впадают ручьи, в более открытых местах на дне живут личинки апатании (Apatania). Их домики по форме напоминают рог (рис. 311, 4). По бокам домика апатании вделаны более крупные песчинки.
В мелких песчаных местах делают свои построенные из песчинок домики личинки моланны (Molanna angustata). У моланны домик, если на него смотреть сверху, широкий и плоский. Центральная трубчатая часть, в которой сидит личинка, сделана из более крупных песчинок, но к ней по бокам прилажены как бы крылья из более мелких песчинок и такой же капюшон. В целом чехлик имеет вид довольно большого щитка, длина его больше 2 см (рис. 311, 5). Двигается личинка моланны со своим чехликом толчками.
В густых зарослях растений держатся личинки фриганей (Phryganea), делающие свои трубчатые домики из нагрызенных четырехугольных, как короткие дощечки, кусочков растений (рис. 311, 3). Часто такие домики сохраняют даже зеленую окраску — куски водных растений в воде долго сохраняют жизнеспособность. У фриганеи домик просторный и длинный, личинка в нем может свободно бегать. Задний конец такого домика-трубки открыт, и, если личинку вытолкнуть из чехлика, она быстро пробежит по его поверхности и ловко юркнет в него с заднего конца. Фриганея — крупное насекомое, длина взрослой личинки около 4 см. Хотя личинки фриганей, делая чехлики, откусывают кусочки растений и в случае необходимости, особенно летом и осенью, сидят в основном на растительной диете, они не вегетарианцы. Охотнее личинки фриганей едят личинок комаров и других мелких беспозвоночных.
На дне заросших прудов обычны личинки лимнофилов (Limnophilus). Домики некоторых видов лимнофилов довольно похожи друг на друга. Личинка строит домик из разных твердых мелких предметов, лежащих на дне. Тут могут быть и мелкие набухшие затонувшие палочки, и маленькие раковины моллюсков, и хвоинки, и другие растительные остатки, но камешки и песчинки лимнофилами не используются. Если личинку лимнофила выгнать из домика и домик убрать, она, выпуская липкие прядильные нити и беспокойно вертясь, сперва делает из чего попало временный домик, а затем, почувствовав, что брюшко как-то защищено, начинает делать постоянный домик, тщательно выбирая прочные частицы и хорошенько пригоняя их друг к другу.
В Северной Америке распространены улитчатые ручейники (семейство Helicopsychidae), делающие себе спирально извитые чехлики, настолько похожие на раковинки улиток (рис. 311, 6), что даже зоологи, прежде чем уверенно сказать, что им встретилось — раковина или домик ручейника, должны очень внимательно приглядеться.
Хотя личинки ручейников и очень хорошо приспособлены к жизни в воде, все же среди форм, строящих чехлики, есть и такие, которые оставили водную среду и перешли к жизни на суше. Таков сухопутный ручейник (Enoicyla pusilla), живущий в буковых лесах Западной Европы (рис. 312). Интересно, что у этого ручейника самки бескрылые. Личинки сухопутного ручейника живут в подстилке и среди мха, покрывающего стволы деревьев. Эта личинка избегает воды и, когда после сильных дождей слой опавших листьев сильно намокает, переселяется на стволы деревьев. Домик личинка делает из мелких кусочков опавших листьев.
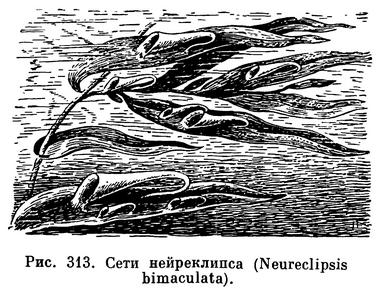
Хотя жизнь в чехликах и характерна для большинства личинок ручейников, представители некоторых семейств ведут иной образ жизни, несмотря на то, что имеют прекрасно развитые прядильные железы. В неглубоких и небыстрых речках в зарослях рдестов и других водных растений встречаются прикрепленные к водным растениям нежные, еле заметные прозрачные трубочки (рис. 313). Они колеблются струями мерно текущей воды. Обычно таких трубочек в одном месте бывает много — целое скопление. Делают их личинки нейреклипса (Neureclipsis bimaculata) из семейства полицентропид (Polycentropidae). Если перенести эти трубчатые образования в стоячую воду, например поместить в ведро с водой, они спадутся и станут невзрачными — течение воды надувало и поддерживало форму этих тонких подводных сачков. Если рассмотреть такую трубочку в бинокуляр, видно, что это и впрямь сеть — сеть, замечательно сплетенная, с мелкими однотипными ячейками. Плетут эти трубчатые сети узкие длинные личинки, живущие без чехла и не имеющие жабр. Личинки (рис. 314) строят себе в текучей воде не домики, а тенета — ловчие сети, в которые попадают проносимые течением мелкие ракообразные, личинки поденок и другие животные, становящиеся добычей нейреклипса. В воде хищная личинка этого ручейника ловит добычу так, как на суше это делают пауки-тенетники!
В больших равнинных реках — в водах Волги, Дона, Днестра — развивается много ручейников гидропсихид (семейство Hydropsychidae). Личинки гидропсихид делают тенета с прямоугольными ячейками, а сами сидят рядом в легком чехлике из тонких нитей (рис. 315). Стоит попасть в тенета мелкому рачку или насекомому, как хищные личинки (их размеры достигают примерно 2 см) выскакивают из укрытия и хватают добычу своими сильными челюстями! В виде мешочков (рис. 316) делают ловчие сети личинки плектрокнемий (Plectrocnemia). Интересно, что такие специализированные ловцы водной добычи, как гидропсихиды и плектрокнемии, могут выходить и на сушу. На расстоянии десятков метров от ручьев в лесной подстилке находили этих личинок, где они жили, конечно, не делая никаких ловчих сетей.
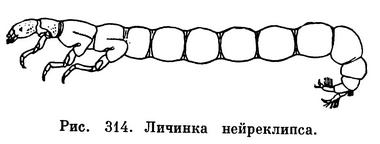
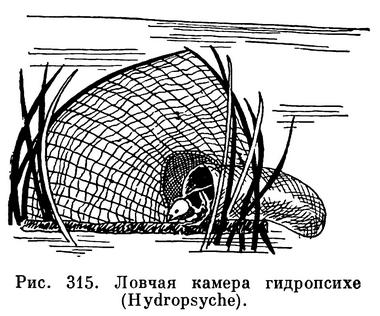
Впрочем, некоторые личинки ручейников (семейство Rhyacophilidae) и в воде не делают сложных сооружений. Ползающие по каменистому дну чистых холодных ручьев красивые зеленовато-синие личинки риакофил (Rhyacophila nubila), (рис. 311, 7), достигающие длины 2,5 см, только выпускают нить, которая удерживает личинку от сноса водой. Эти хищники цепляются за дно и за выделяемую ими нить ногами и прикрепительными крючьями на заднем конце брюшка и поджидают добычу. Быстрому схватыванию добычи личинкам риакофил помогает то, что у них сильные челюсти направлены прямо вперед, как у хищных личинок жужелиц.
Развитие ручейников длится обычно 1 год, но у крупных северных видов и 2—3 года.
Ознакомление даже с немногими представителями личинок ручейников показывает, насколько разнообразны их повадки и особенности. А взрослые ручейники не питаются, только размножаются, и все ведут сходный образ жизни. Поэтому понятно, что распознавать личинок ручейников сравнительно легко (у разных видов неодинаков не только образ жизни, но неодинаково и строение отдельных частей тела), а виды взрослых ручейников могут распознавать только энтомологи, специально их изучающие.
Куколка у ручейников свободная (рис. 317). Это в общем такая же приспособленная к жизни в воде стадия, как и личинка. Жизнь куколки легко проследить на примере стенофила, с рассмотрения которого было начато знакомство с личинками ручейников. Перед окукливанием личинка выбирает более спокойный участок водоема и, прикрепляя чехлик к камню, заплетает его концы так, чтобы в каждом было отверстие для свободного доступа воды. Когда личинка окуклится, куколка внутри чехлика делает все время колебательные движения, упираясь в стенку чехлика выростом на основании брюшка. Для прочистки отверстий у куколки на верхней губе имеются сильные щетинки и на заднем конце тела очистительные отростки. Ко времени созревания куколка прорывает своими мощными зубчатыми челюстями (непохожими на личиночные, а тем более на практически отсутствующие челюсти взрослых ручейников) передний конец чехлика и, выйдя из него, начинает быстро плавать на спине, как клопы-гладыши, делая гребные движения длинными, снабженными плавательными волосками средними ногами. Добравшись до камня, берега или растения, куколка цепляется за него и выползает из воды. Трудно назвать куколку ручейника «покоящейся стадией», как часто называют куколок насекомых!
Известно около 3000 видов ручейников — они распространены в основном в нежарких местностях. В СССР отмечено около 600 видов.
По системе крупного знатока этих насекомых А. В. Мартынова ручейники делятся на 2 подотряда. Подотряд цельнощупиковых (Integripalpia) назван так, поскольку у взрослых насекомых последний членик челюстных щупиков простой, не разделенный на кольца, этот подотряд включает ручейников, в основном делающих себе домики. Подотряд кольчатощупиковых (Annulipalpia) назван так по разделенным на колечки челюстным щупикам и включает, в частности, не делающих домиков гидропсихид и риакофил.
Всего в пределах отряда разные энтомологи выделяют от 13 др 16 семейств.
Ручейники — несомненно полезная группа насекомых, их личинками питаются промысловые рыбы наших рек. В горных ручьях личинками стенофилов питаются форели, поедая их, несмотря на прочные песчаные домики.
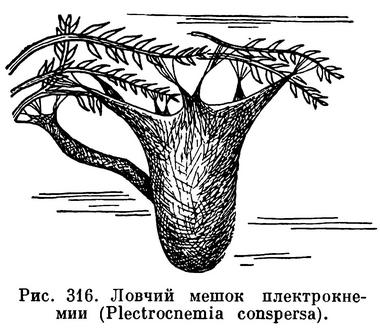
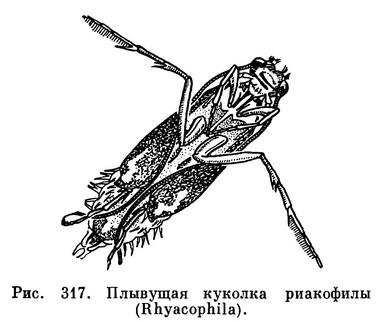
ОТРЯД ЧЕШУЕКРЫЛЫЕ, ИЛИ БАБОЧКИ (LEPIDOPTERA)
Из всех насекомых бабочки пользуются наибольшей известностью. Вряд ли найдется на свете человек, который не восхищался бы ими так же, как восхищаются красивыми цветами. Недаром в древнем Риме верили в то, что бабочки произошли от цветов, оторвавшихся от растений. Во всех уголках мира есть любители, коллекционирующие бабочек с не меньшей страстностью, чем другие коллекционеры собирают произведения искусства.
Красота бабочки в ее крыльях, в их разнообразных расцветках. В то же время крылья — важнейший систематический признак отряда: они покрыты чешуйками, от структуры и расположения которых зависит причудливость окраски. Поэтому бабочек и называют чешуекрылыми. Чешуйки представляют собой измененные волоски. В этом легко убедиться, если внимательно рассмотреть чешуйчатый покров бабочки аполлона (Parnassius apollo). По краю крыла находятся очень узкие чешуйки, почти волоски, ближе к середине они расширены, но концы их остры, и, наконец, еще ближе к основанию крыла сидят широкие чешуйки в виде сплюснутого, полого внутри мешочка, прикрепленного к крылу посредством тонкого короткого стебелька (рис. 318).
Чешуйки располагаются на крыле правильными рядами поперек крыла: концы чешуек обращены к боковому краю крыла, а их основания прикрыты черепицеобразно концами предыдущего ряда. Окраска чешуйки зависит от находящихся в ней пигментных зерен; наружная поверхность ее ребристая. Помимо таких пигментных чешуек, у многих видов, особенно тропических, крылья которых отличаются переливающейся металлической окраской, имеются чешуйки иного типа — оптические.
В таких чешуйках отсутствует пигмент, а характерная металлическая окраска возникает благодаря разложению белого солнечного луча на отдельные цветные лучи спектра при прохождении его через оптические чешуйки. Это разложение лучей достигается преломлением их в скульптуре чешуек, обусловливающим перемену цвета при изменении направления, по которому падают лучи. Особый интерес представляют пахучие чешуйки, или андроконии, встречающиеся преимущественно у самцов некоторых видов бабочек. Это измененные чешуйки или волоски, связанные с особыми железками, выделяющими пахучий секрет. Андроконии располагаются на разных частях тела — на ногах, крыльях, на брюшке. Распространяемый ими запах служит приманкой для самки, обеспечивая, таким образом, сближение полов; часто он бывает приятным, напоминающим в ряде случаев аромат ванили, резеды, земляники и т. д., но иногда он может быть и неприятным, например вроде запаха плесени. Следует подчеркнуть, что для каждого вида бабочек характерны и форма, и оптические и химические свойства находящихся на крыльях чешуек. В редких случаях чешуйки на крыльях отсутствуют, и тогда крылья кажутся совершенно прозрачными, как это имеет место у стеклянниц.
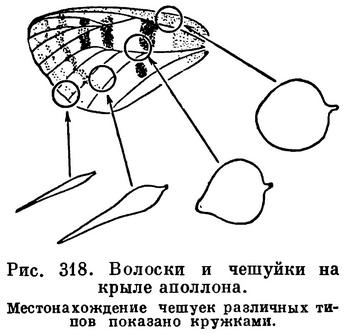
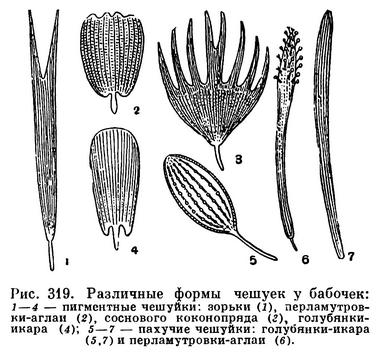
Обычно у чешуекрылых развиты все четыре крыла; однако у самок некоторых видов крылья могут недоразвиваться или совсем отсутствовать. Передние крылья всегда больших размеров, чем задние. У многих видов обе пары крыльев сцепляются друг с другом при помощи особой зацепки, или «уздечки», представляющей собой хитиновую щетинку или пучок волосков, одним концом прикрепленных на верхней стороне переднего края заднего крыла, а другим концом входящих в карманообразный придаток на нижней стороне переднего крыла. Могут быть и другие формы сцепочных механизмов, соединяющих переднее и заднее крыло.
Не менее характерным признаком, чем структура крыльев и покрывающих их чешуек, являются ротовые органы бабочек (рис. 320). В преобладающем большинстве случаев они представлены мягким хоботком, способным свертываться и развертываться наподобие часовой пружины. Основу этого ротового аппарата составляют сильно удлиненные внутренние лопасти нижних челюстей, которые и образуют створки хоботка. Верхние челюсти отсутствуют или представлены небольшими бугорками; сильной редукции подверглась и нижняя губа, хотя ее щупики развиты хорошо и состоят из 3 члеников. Хоботок бабочки очень эластичен и подвижен; он прекрасно приспособлен к питанию жидкой пищей, каковой в большинстве случаев служит нектар цветов. Длина хоботка того или иного вида обычно соответствует глубине расположения нектара в тех цветках, которые посещают бабочки. Так, на Мадагаскаре произрастает одна интересная орхидея (Angraecum sesquipedale) с глубиной венчика 25—30 см. Она опыляется длиннохоботным бражником (Macrosila morgani), имеющим хоботок длиной около 35 см. В некоторых случаях источником жидкой пищи чешуекрылых могут служить вытекающий сок деревьев, жидкие экскременты тлей и другие сахаристые вещества. У некоторых бабочек, которые не питаются, хоботок может быть недоразвит или совсем отсутствовать (тонкопряды, некоторые моли и др.).
Дневной образ жизни характерен прежде всего для так называемых дневных или булавоусых бабочек. Так называют комплекс (серию) семейств чешуекрылых, отличающихся булавовидными усиками (парусники, белянки, нимфалиды, геликониды, морфиды, голубянки). У них сильный и длинный хоботок, при помощи которого они высасывают нектар из цветков. Крылья широкие, в покое подняты вверх (за редким исключением), на задних крыльях отсутствует зацепка.
Вызывают восхищение изумительные окраски крыльев дневных бабочек; их верхняя сторона обычно окрашена ярко и пестро, в то время как расцветки нижней стороны часто имитируют окраску и рисунок коры, листьев и т. д. С особой любовью относился к дневным бабочкам создатель первой научной систематики животных знаменитый швед Карл Линней. Давая названия описываемым им видам, он искал их в мифах классической древности. Это вошло в традицию у лепидоптерологов, т. е. ученых, изучающих бабочек. Поэтому так часто среди названий дневных бабочек встречаются имена древнегреческих богов и любимых героев: Аполлон, Киприда, По, Гектор, Менелай, Лаэрт. Они как бы символизируют все яркое, сильное и прекрасное, что радует и восхищает человека.
Интересно биологическое значение ярких, пестрых окрасок верхней стороны крыльев, так часто наблюдающихся у булавоусых бабочек, особенно у нимфалид. Их основное значение заключается в распознавании особей своего вида на большом расстоянии. Наблюдения показывают, что самцы и самки таких пестроокрашенных форм привлекаются друг к другу издали окраской, а вблизи происходит окончательное распознавание по запаху, издаваемому андрокониями. Для проверки отрезали крылья у живых перламутровок и на их место приклеивали крылья белянок. Оперированные экземпляры выставлялись на лужайке и к ним вскоре прилетали белянки, большей частью самцы. Удавалось приманивать самцов бабочек на искусственные изображения самок их вида.
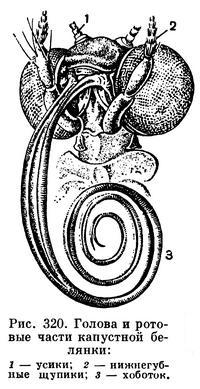
Если верхняя сторона крыльев у нимфалид всегда окрашена ярко, то иного типа окраски характерны для их нижней стороны: они, как правило, криптические, т. е. защитные. В этой связи интересны два типа складывания крыльев, широко распространенные у нимфалид, а также и в других семействах дневных бабочек. В первом случае бабочка, находясь в положении покоя, выдвигает передние крылья вперед так, что их нижняя поверхность, имеющая защитную окраску, открыта почти на всем протяжении (рис. 322, 1). По этому типу складываются крылья, например, у углокрыльницы С-белое (Polygonia C-album). У нее верхняя сторона буро-желтая с темными пятнами и наружной каймой; нижняя сторона серо-бурая с белой буквой «С» на задних крыльях, за что она и получила свое название. Неподвижно сидящая бабочка малозаметна также благодаря неправильному угловатому контуру крыльев.
Другие виды, например адмирал и репейница, прячут передние крылья между задними так, что видны только их кончики (рис. 322, 2). В этом случае на нижней поверхности крыльев бывают выражены два типа окрасок: та часть передних крыльев, которая в состоянии покоя оказывается спрятанной, ярко окрашена, вся остальная часть нижней поверхности крыльев носит явно криптический характер.
У многих нимфалид, особенно у тропических форм, наблюдается подражательное сходство с листьями, когда воспроизводится характерная окраска сухих или живых листьев, их контуры и специфическое жилкование. Классическим примером в этом отношении являются индо-малайские бабочки-листовидки рода каллима (Kallima). Верхняя сторона крыльев у каллимы окрашена ярко и пестро, а нижняя своей окраской и рисунком напоминает сухой лист. Сходство с листом у сидящей бабочки усиливается еще благодаря тому, что ее верхнее крыло заострено на вершине, а нижнее крыло имеет небольшой хвостик, имитирующий черешок листа (табл. 16, 4).
Во всех указанных случаях пестрота окраски зависит от распределения пигментов в чешуйках, покрывающих крыло. Как показали многочисленные опыты, отложение пигментов в большой мере зависит от температурного фактора, воздействующего на куколок. При воспитании куколок при пониженных температурах (от 0 до 10°С) можно получить взрослые формы с сильным развитием темного пигмента меланина. Так, у траурницы при воздействии низких температур на ее куколку темнеет общий фон крыла, уменьшаются голубые пятна и по всей желтой полоске, идущей вдоль наружного края крыльев, откладывается меланин в виде черных точек. Весьма характерно, что сходные изменения вызывает выдерживание куколок траурницы и при высокой температуре, порядка 35—37°С. Этим объясняется различная окраска у одного и того же вида в разных климатических условиях. В этой связи весьма интересна постоянная сезонная изменчивость у изменчивой пестрокрыльницы (Araschnia levana), развивающейся в двух поколениях, отличающихся друг от друга по окраске. У весеннего поколения крылья рыжевато-красные, со сложным черным рисунком и белыми пятнышками у вершины переднего крыла; летнее поколение имеет буровато-черные крылья с белыми или желтовато-белыми пятнами на переднем крыле и такой же перевязью на заднем.
Преобладающее большинство булавоусых бабочек распространено в тропических странах. Именно здесь встречаются виды, поражающие своей величиной и изумительной яркостью раскрасок.
Среди тропических видов особенно красивы и своеобразны морфиды (Morphidae), представленные всего только одним родом (Morpho). Это крупные бабочки, достигающие в размахе крыльев 15—18 см. Верхняя сторона их крыльев окрашена в синие или голубые сильно переливающиеся металлические цвета. Такая окраска зависит от того, что крыло покрыто оптическими чешуйками, причем нижняя часть оптических пластинок пигментирована; пигмент не пропускает света и тем самым придает большую яркость интерференционной окраске ребер. У самцов, например, у изображенной на цветной таблице 45 Morpho cypris, блеск крыла чрезвычайно силен и производит впечатление полированного металла. В соединении с крупными размерами морфид это приводит к тому, что при ярком солнечном освещении каждый взмах крыла виден за треть километра. Морфиды относятся к самым заметным насекомым, населяющим леса тропической части Амазонки. Особенно много их на прогалинах и освещенных солнцем дорогах. Летают они на большой высоте; некоторые из них вообще не опускаются к земле ближе чем на 6 м.
В ряде случаев у дневных бабочек ярко окрашенными бывают и верхняя и нижняя стороны крыльев. Подобная окраска обычно сочетается с несъедобностью обладающего ею организма, поэтому она получила название предостерегающей. Предостерегающая окраска характерна, например, для геликонид. Геликониды (Heliconidae) — это своеобразное семейство эндемичных булавоусых бабочек, в состав которого входит около 150 видов, распространенных в Южной Америке. Крылья у них окрашены очень пестро, преимущественно в оранжевый цвет с контрастным рисунком из черных и желтых полос и пятен (табл. 17). Многие из геликонид обладают противным запахом и неприятным вкусом, и потому их не трогают птицы. Бабочки изобилуют под сенью роскошных тропических лесов Амазонки. Своим поведением и повадками они как бы демонстрируют свою неуязвимость. Полет у них медленный, тяжелый; держатся всегда роями, причем не только в воздухе при полете, но и на отдыхе, когда рой опускается в крону дерева. Сильный запах, исходящий от скопления отдыхающих бабочек, в значительной мере предохраняет их от врагов.
Известный английский ученый Бэте, изучая поведение геликонид, открыл любопытное явление, получившее название мимикрии. Под мимикрией понимают сходство в окраске, форме и поведении между двумя или несколькими видами насекомых. Характерно, что мимикрирующие виды всегда имеют яркую предостерегающую (демонстрационную) окраску.
У бабочек мимикрия выражается в том, что одни из мимикрирующих видов оказываются несъедобными, в то время как другие лишены защитных свойств и только «подражают» своим защищенным моделям. Такими подражателями, для которых геликониды служат моделями, являются бабочки-белянки — дисморфия (Dismorphia astynome) и перхибрис (Perrhybris pyrrha). Они держатся в стаях летающих и отдыхающих геликонид, подражая им формой и окраской крыльев, а также полетом.
В дальнейшем выяснилось, что мимикрия довольно широко распространена среди чешуекрылых, причем формы ее проявления различны. Так, у одного из африканских видов парусников (Papilio dardanus) хорошо выражен половой диморфизм: самцы имеют хвостики на задних крыльях, общая окраска крыльев желтая с темными полосами; у самок задние крылья округленные, без хвостиков. При этом самки представлены несколькими формами, сильно отличающимися друг от друга (рис. 323); каждая форма воспроизводит определенный тип окраски, свойственный определенному виду несъедобных бабочек данаид (Danaidae). У формы hippocoon на обоих крыльях имеются голубые пятна, как у ее модели (Amauris niavius); у формы cenea голубые пятна только на передних крыльях, а основания задних крыльев желтого цвета, как у другой модели (Amauris echeria).

Своеобразно проявление мимикрии у бабочек стеклянниц (Aegeriidae), которые своим обликом скорее напоминают перепончатокрылых насекомых или крупных мух, чем чешуекрылых. Это подражательное сходство достигается благодаря характерной структуре крыльев и общим контурам тела. Крылья стеклянниц почти лишены покрова из чешуек и потому прозрачные, стекловидные; задние крылья короче передних, и чешуйки на них сосредоточены только на жилках. Тело довольно стройное, с длинным брюшком, далеко выдающимся позади крыльев; усики нитевидные или слегка утолщены посредине.
В отличие от бабочек, летающих в дневные часы, виды, питающиеся нектаром в сумерках или ночью, обладают иным типом окраски. Верхняя сторона их передних крыльев всегда окрашена под цвет субстрата, на котором они сидят днем. В покое передние крылья складываются вдоль спины кровлеобразно или наподобие плоского треугольника, прикрывая нижние крылья и брюшко. Неподвижно сидящая бабочка становится незаметной.
Окраска задних крыльев чаще всего однотонная, неяркая. Однако в ряде случаев, например у совок, ленточниц, медведиц и бражников, она может быть яркой, предостерегающей. Так, у красной ленточницы (Catocala nupta, табл. 16, 11) задние крылья кирпично-красные с черными перевязями, у желтой (Catocala fulminea, табл. 16, 10) — охряно-желтые с черной срединной перевязью и таким же внешним краем, у голубой (Catocala fraxini, табл. 16, 9) — голубые с черной каймой и срединной перевязью. У обыкновенной медведицы (Arctia caja, табл. 16, 12) задние крылья красные с большими темно-синими, почти черными пятнами; брюшко с черными пятнами.
В спокойном состоянии днем бабочки сидят на стволах деревьев сложив крылья и потому незаметны; при угрозе нападения они раздвигают передние крылья и демонстрируют отпугивающий сигнал в виде ярко окрашенных нижних крыльев, а иногда и брюшка.
Своеобразна защитная окраска у серебристой лунки (Phalera bucephala). Передние крылья у нее серебристо-белые с большим желтым пятном в наружном углу; задние крылья серые. Днем бабочка сидит на дереве со сложенными кровлеобразно крыльями. В это время ее можно принять за обломок веточки. При этом желтые пятна на слегка вогнутых концах передних крыльев воспроизводят вид обнаженной древесины (табл. 16, 14).
Чешуекрылые — насекомые с полным метаморфозом. Их яйца очень разнообразны по форме, обычно окрашены, оболочка часто имеет сложную структуру. Личинок бабочек называют гусеницами (табл. 46, 1—16). В большинстве случаев они червеобразной формы; тело состоит из головы, 3 грудных и 10 брюшных колец. В отличие от взрослых чешуекрылых их гусеницы всегда имеют грызущий ротовой аппарат. Помимо трех пар грудных ножек, у гусениц имеются еще так называемые «ложные», или «брюшные», ножки, которых бывает до 5 пар; они помещаются обычно на третьем—шестом и девятом брюшных сегментах. Брюшные ножки не расчленены, и их подошвы усажены хитиновыми крючочками. Специфичной физиологической особенностью гусениц является наличие у них пары трубчатых прядильных, или шелкоотделительных, желез, открывающихся общим каналом на нижней губе. Они представляют собой измененные слюнные железы, у которых основная функция слюноотделения заменена выработкой шелка. Выделения этих желез быстро затвердевают на воздухе, образуя шелковую нить, при помощи которой одни гусеницы скрепляют свернутые в трубку листья, другие повисают в воздухе, спускаясь с ветки, третьи окружают себя и ветви, на которых они сидят, паутиной. Наконец, у гусениц шелковая нить используется для построек кокона, внутри которого происходит окукливание.
У преобладающего числа видов бабочек гусеницы являются фитофагами; значительно реже встречаются случаи питания продуктами животного происхождения: шерстью, воском, роговыми веществами. Есть среди гусениц хищники и даже паразиты.
По образу жизни гусениц можно разделить на две группы: 1) гусеницы, ведущие свободный образ жизни, которые живут более или менее открыто на растениях; 2) гусеницы, ведущие скрытый образ жизни. Свободноживущие гусеницы обитают как на травянистых, так и на древесных растениях, питаясь листьями, цветками и плодами.
Переход к скрытому образу жизни представляет обитание в переносных чехликах, которые гусеницы сплетают из шелковистых нитей. Передвигаясь по растению, гусеницы таскают на себе свой чехлик, прячась в него при опасности. Так поступают, например, гусеницы бабочек-мешечниц. Такое же промежуточное положение между указанными двумя биологическими группами занимают листоверты. Так называют гусениц, строящих убежища из листьев, свертывая их и скрепляя свернутые части шелковистой нитью. При постройке такого убежища используется один или несколько листьев. Для многих гусениц характерно свертывание листа в сигарообразную трубку.
Гусеницы, живущие «обществами», обычно устраивают специальные, иногда сложные гнезда, оплетая паутиной ветви, листья и другие части растений. Крупные паутинные гнезда образуют гусеницы яблонной горностаевой моли (Hyponomeuta malinellus), являющиеся опасными вредителями садов и лесов. Большими группами живут в паутинных гнездах гусеницы походных шелкопрядов (семейство Eupterotidae), отличающиеся своеобразным поведением: в поисках пищи они отправляются «в поход» стройными рядами, следуя гуськом друг за другом. Так ведут себя, например, гусеницы дубового походного шелкопряда (Thaumetopoea processionea, табл. 46, 2), изредка встречающегося в лесах Юго-Западной Украины.
Бабочка этого вида летает в августе и сентябре и откладывает яйца на кору дуба кучкой из нескольких прямых рядов, по 100—200 штук в кучке. Яйца зимуют, защищенные плотной прозрачной пленкой, образующейся из выделений самки. Вылупившиеся из яиц в мае гусеницы держатся группами в паутинном гнезде. Когда листья на дереве уже сильно объедены, они спускаются с него и в поисках пищи ползут по земле, всегда в определенном порядке: впереди ползет одна гусеница, за ней, касаясь ее своими волосками, следует другая. В середине колонны число гусениц в ряду увеличивается, сначала по 2, затем по 3—4 гусеницы ползут рядом. К концу колонна опять сужается. В июле — начале августа тут же в гнезде происходит окукливание, причем каждая гусеница плетет себе овальный кокон. Через две-три недели вылетают бабочки.
Скрытый образ жизни ведут все гусеницы, обитающие внутри различных органов растений. Сюда относятся минёры, плодожорки, бурильщики и галлообразователи.
Минёрами называют гусениц, живущих внутри листьев и их черешков и прокладывающих внутри хлорофиллоносных тканей внутренние ходы — мины. Некоторые минёры выедают не все содержимое листа, а ограничиваются или отдельными участками паренхимы, или же эпидермисом.
Форма мин весьма различна. В одних случаях мина прокладывается в виде округлого пятна (пятновидная мина); иногда такое пятно дает боковые отростки, напоминая звезду (звездовидные мины). В других случаях мина имеет вид галереи, очень узкой в основании, но затем сильно расширяющейся в верхней части (трубковидная мина). Встречаются также узкие длинные мины, но сильно извилистые (змеевидные мины) или спирально закрученные (спиральные мины).
Когда минирующие гусеницы живут внутри листа группами, могут возникать так называемые вздутые мины. Так, гусеницы сиреневой моли (Caloptilia syringella), относящейся к особому семейству молей-пестрянок (Gracillariidae), вначале живут по нескольку штук вместе в одной общей мине, имеющей форму широкого пятна, которое может занимать большую часть листа. Мины эти сильно вздуты от скопляющихся в них газов. Эпидермис, покрывающий мину, быстро желтеет. Позднее гусеницы выходят из своих мин и, скелетируя листья, свивают их в трубки. Перед окукливанием они уходят в землю. В течение лета бывает два поколения; зимует у сиреневой моли куколка.
Гусеницы-плодожорки живут внутри плодов различных растений. Одни из них повреждают мякоть плодов, другие питаются исключительно семенами. Гусеницы-бурильщики обитают в стеблях травянистых растений или внутри ветвей и стволов кустарников и деревьев. Среди бурильщиков особенно характерны стеклянницы (семейство Aegeriidae) и древоточцы (Cossidae).
Большинство видов стеклянниц развивается в стволах древесных растений, нанося им серьезные повреждения. К числу широко распространенных в Европе лесных вредителей нужно отнести большую тополевую стеклянницу (Aegeria apiformis).
Самки этого вида откладывают яйца на нижнюю часть стволов деревьев, преимущественно тополей. Гусеницы (табл. 46, 14) развиваются в течение двух лет, питаясь древесиной, в которой проделывают ходы. На третий год весной они окукливаются в колыбельке под корой в особом плотном коконе из опилок и экскрементов. Перед выходом бабочки куколка на 2/3 высовывается из лётного отверстия; даже после вылета бабочки куколочная шкурка продолжает сохранять такое положение.
Опасными для лесного хозяйства являются и некоторые виды древоточцев, например пахучий древоточец (Cossus cossus) и въедливая древесница (Zeuzera pyrina). Самка пахучего древоточца откладывает яйца кучками по 20—70 штук в трещины коры на стволах ив, тополей, ольхи, вяза и дуба. Развитие протекает в течение двух лет. Молодые гусеницы вгрызаются под кору, где делают общий ход неправильной формы, в котором они и зимуют. На следующий год гусеницы расходятся и каждая из них, углубляясь в древесину, выгрызает в ней широкий, преимущественно продольный ход. Гусеницы 16-ногие, с темно-бурой головой и розоватым телом, оттенок которого меняется в течение жизни; к концу развития достигают в длину 10—12 см (табл. 46, 15). Пахучим древоточец называется потому, что гусеница издает резкий, неприятный запах древесного спирта; такой же запах распространяет и поврежденная ею древесина. Хотя пахучий древоточец заселяет чаще всего старые и больные деревья, но он может быть опасен и для здоровых деревьев в тех случаях, когда образует небольшие, но устойчивые многолетние очаги.
Гусеницы въедливой древесницы (табл. 46, 16) многоядны: они повреждают более 70 древесных пород, в том числе ясени, ильмовые, яблоню, грушу и др. Самки этого вида откладывают яйца по одному на верхушки молодых побегов, в пазухи листьев и на листовые почки. По выходе из яйца гусеницы вгрызаются в молодые побеги и черешки листьев, отчего поврежденные листья засыхают и преждевременно опадают. К осени гусеницы переходят на молодые ветви, в древесине которых они выгрызают ходы. Здесь они зимуют. На следующий год, после перезимовки, гусеницы возобновляют свою вредную деятельность и по мере роста спускаются все ниже и ниже по дереву. Вторую зимовку они проводят в ходах, проложенных в средней и нижней частях дерева. Окукливание происходит в мае—июне, гусеница окукливается без кокона в верхней части хода, где она зимовала.
Настоящих галлообразователей среди гусениц очень немного. Больше всего их известно из семейства листоверток (Tortricidae). Наносимые ими повреждения чаще всего заключаются в уродливых вздутиях тех органов растения, внутри которых происходит развитие гусениц. Laspeyresia servillana вызывает вздутия стебля ивы, a Epiblema lacteana развивается в утолщенных стеблях полыни.
Весьма своеобразна жизнь чешуекрылых, гусеницы которых развиваются в водной среде. В середине лета по берегам водоемов, поверхность которых покрыта листьями белых лилий и желтых кувшинок, часто можно встретить небольшую бабочку с красивыми желтоватыми крыльями, сложный рисунок которых состоит из сильно изогнутых бурых линий и располагающихся между ними беловатых пятен неправильной формы (рис. 324). Это кувшинковая, или болотная, огневка (Hydrocampa nymphaeata). Она откладывает яйца на листьях различных водных растений, с нижней их стороны. Вылупляющиеся из яиц зеленоватые личинки сперва минируют ткани растений. В это время их дыхальца сильно редуцированы, поэтому дыхание происходит через поверхность кожи. После линьки гусеница оставляет мину и строит из вырезанных кусочков рдеста и кувшинок особый чехлик, причем дыхание пока остается прежним. В этом чехлике гусеница зимует, а весной покидает его и строит новый чехлик. Для этого она челюстями выгрызает из листа два овальных или круглых куска, которые скрепляет по бокам паутинкой. Такой чехлик всегда наполнен воздухом; на этой стадии у гусеницы имеются вполне развитые стигмы и трахеи, и она дышит теперь атмосферным воздухом. Ползая по водным растениям, гусеница таскает с собой чехлик так же, как это делают ручейники. Питается она, соскабливая челюстями кожицу и мякоть с листьев водных растений. Окукливание происходит в чехлике.
В чехликах под водой живет и серая гусеница рясковой огневки (Cataclysta lemnata), но строительным материалом в этом случае служит ряска, отдельные пластинки которой скрепляются паутинкой. Перед окукливанием гусеница обычно покидает свой чехлик и заползает в какую-нибудь камышовую или тростниковую трубочку.
Еще более приспособлена к водной жизни зеленоватая гусеница телорезной огневки (Paraponyx stratiotata), встречающаяся на листьях телореза, рдестов, роголистника и других растений. Она живет исключительно под водой в неправильных чехликах или вовсе без чехликов. Дышит трахейными жабрами, которые в виде длинных мягких разветвленных выростов расположены по 5 пар почти на каждом сегменте.
У подводной огневки (Acentropus niveus) самки встречаются в виде двух форм — крылатой и почти бескрылой, у которой сохраняются только небольшие зачатки крыльев. Бескрылые самки откладывают яйца под водой. Оливково-зеленая гусеница, живя на поверхности листьев рдеста и других растений, устраивает себе небольшую покрышку из отгрызенного кусочка. Окукливание происходит в коконе, прикрепленном к стеблям или нижней поверхности листа (рис. 326).
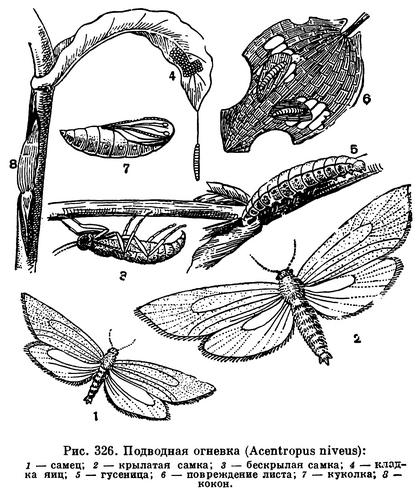
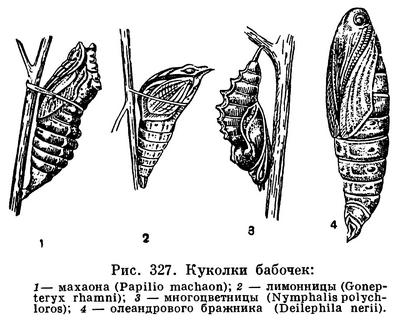
В тесной связи с образом жизни гусениц находятся форма и окраска их тела. Гусеницы, ведущие открытый образ жизни, часто имеют криптическую окраску, хорошо гармонирующую с окружающим фоном. Эффективность защитной окраски может повышаться благодаря особенностям рисунка. Так, у гусениц бражников по общему зеленому или серому фону проходят косые полосы, которые расчленяют тело на отрезки, делая его еще менее бросающимся в глаза. Покровительственная окраска, сочетаясь с характерной формой, часто приводит к возникновению покровительственного сходства с частями растений, на которых обитает гусеница. У пядениц, например, гусеницы бывают похожи на сухие сучки.
Наряду с криптической окраской у гусениц, ведущих открытый образ жизни, встречается и яркая демонстрационная окраска, свидетельствующая об их несъедобности. Эффект этой окраски зависит не только от цвета наружных покровов, но и от окраски волосяного покрова. Примером может служить гусеница античной волнянки (Orgyia antiqua), имеющая весьма причудливый вид; она серая или желтоватая с черными и красными пятнами и с пучками черных волос различной длины; на спинной стороне желтые волоски собраны в четыре густые щетки (табл. 46, 9). Некоторые гусеницы в момент опасности принимают угрожающую позу. К ним нужно отнести гусеницу большой гарпии (Cerura vinula), имеющую весьма своеобразную форму: у нее большая плоская голова, широкое в передней части тело сильно сужается к заднему концу, на вершине которого находится «вилка», состоящая из двух сильно пахучих нитей. Стоит потревожить гусеницу, как она сразу принимает угрожающую позу, приподнимая вверх переднюю часть тела и кончик брюшка с «вилкой» (табл. 46, 1).
Иного типа окраска гусениц, ведущих скрытый образ жизни: у них отсутствуют яркие сочетания цветов. Чаще всего для них характерны монотонные бледные окраски: беловатая, светло-желтоватая или розоватая.
Куколка у чешуекрылых имеет яйцевидно вытянутую форму, с заостренным задним концом (рис. 327). Ее плотные наружные покровы образуют твердую оболочку; все придатки и конечности спаяны с телом, в результате чего поверхность куколки становится сплошной, ноги и крылья не могут быть отделены от туловища без нарушения целостности покровов. Такая куколка носит название покрытой куколки. Передвигаться она не может, но у нее сохраняется некоторая подвижность последних сегментов брюшка. Очень причудливы куколки дневных бабочек: обычно угловатые, нередко с металлическим блеском, без кокона. Они прикрепляются к различным предметам, причем или висят головой вниз (висячая куколка), или опоясаны нитью, и тогда их голова обращена кверху (подпоясанная куколка).
У многих чешуекрылых гусеницы перед окукливанием плетут шелковистый кокон, в котором и происходит развитие куколки. У некоторых видов количество шелка в коконе настолько велико, что представляет большой практический интерес. С давних времен шелководство составляем очень важную отрасль промышленности.
Основной производитель натурального шелка в СССР — это тутовый шелкопряд (Bombyx mori), относящийся к семейству настоящих шелкопрядов (Bombycidae). В настоящее время в природе в диком состоянии этот вид не существует. Родина его, по-видимому, Гималаи, откуда он был завезен в Китай, где шелководство начало развиваться за 2500 лет до н. э. В Европе эта отрасль производства возникает приблизительно в VIII веке; более трехсот лет тому назад она проникла в Россию.
По внешнему виду тутовый шелкопряд невзрачная бабочка с толстым, сильно волосистым телом и белыми крыльями, достигающими в размахе 4—6 см (табл. 47, 2). Самцы отличаются от самок более тонким брюшком и перистыми усиками. Несмотря на наличие крыльев, бабочки в результате одомашнивания утратили способность летать.
Хотя тутовый шелкопряд нормально размножается путем спаривания самцов и самок, в некоторых случаях у него проявляется партеногенез. В 1886 году русский зоолог А. А. Тихомиров доказал возможность искусственного получения партеногенеза у тутового шелкопряда в результате стимуляции неоплодотворенных яиц различными механическими, термическими, химическими раздражителями. Это был первый случай получения искусственного партеногенеза. В настоящее время искусственный партеногенез получен у многих беспозвоночных (насекомых, иглокожих) и позвоночных животных (земноводных).
Гусеница тутового шелкопряда известна под названием шелковичного червя. Она крупная, длиной до 8 см, мясистая, беловатого цвета, с роговидным придатком на конце брюшка. Ползает сравнительно медленно. При окукливании гусеница выделяет одну цельную нить, длиной до 1000 м, которую она обматывает вокруг себя в виде шелковистого кокона.
Основные центры шелководства у нас находятся в Средней Азии и в Закавказье.
Их положение определяется распространением кормового растения, каковым является тутовое дерево (шелковица). Продвижению шелководства далее на север мешает отсутствие холодоустойчивых сортов шелковицы.
Помимо тутового шелкопряда, в шелководстве используются и другие виды бабочек, например китайская дубовая павлиноглазка (Antheraea pernyi), которая уже более 250 лет разводится в Китае. Получаемый из ее коконов шелк идет на изготовление чесучи. В Советском Союзе работа по акклиматизации этой бабочки ведется с 1924 года. Благоприятные условия для ее культуры у нас имеются в полесских районах Украинской и Белорусской ССР, где в поймах рек располагаются природные массивы низкорослых дубовых порослей. Китайская дубовая павлиноглазка (табл. 47, 1) — крупная бабочка (размах крыльев 12—15 см); самки более крупных размеров, красновато-палевой окраски, самцы серовато-палевые со слабым оливковым оттенком. Вдоль наружного края крыльев проходит светлая полоса; на каждом крыле находится крупный глазок с прозрачным окошечком.
Очень велико хозяйственное значение чешуекрылых как вредителей сельского и лесного хозяйства. На территории Советского Союза зарегистрировано свыше 1000 видов чешуекрылых, гусеницы которых повреждают полевые, садовые или лесные культуры. В преобладающем большинстве случаев комплекс вредителей формируется за счет представителей местной фауны, переходящих на культурные поля с дикорастущих растений. В этом отношении весьма любопытна история заселения подсолнечника подсолнечниковой огневкой (Homoeosoma nebulella). Родина этого растения — Северная Америка; в Россию оно попало только в XVIII столетии и долгое время считалось декоративным. Только с 60-х годов прошлого столетия подсолнечник стал у нас промышленной масличной культурой. В течение многих лет посевы его страдали от подсолнечниковой огневки, переходившей на него с дикних растений, преимущественно с чертополохов. Бабочки этого вредителя откладывают яйца на внутренние стенки пыльников; выходящие из яиц гусеницы вгрызаются в семянки и выедают в них зародыши. Современные панцирные сорта подсолнечника, выведенные советскими селекционерами, почти не повреждаются огневкой благодаря наличию в кожуре семянки особого панцирного слоя, который гусеница прогрызть не может.
Известны факты завоза вредных чешуекрылых из других стран. Совсем недавно в Европе широкую известность получила американская белая бабочка (Hyphantria cunea), родина которой — Северная Америка. На Европейском континенте она впервые была обнаружена в 1940 году в Венгрин, через несколько лет быстро распространилась в Австрии, Чехословакии, Румынии и Югославии. У бабочки белоснежные крылья (размах — 2,5—3,5 см), у некоторых особей на брюшке и на крыльях мелкие черные точки. Усики у самки нитевидные, у самца перистые, черные с белым налетом.
Гусеницы многоядны, могут питаться более чем на 200 видах растений. Характерно, что в Европе они предпочитают шелковицу, которую почти не трогают в Америке. Гусеницы сверху бархатисто-коричневые с черными бородавками, несущими длинные волоски; по бокам лимонно-желтые полосы с бородавками оранжевого цвета; длина 3,5 см. Зимуют куколки, которые находятся под корой деревьев, в развилинах ветвей и под опавшими листьями. Бабочка откладывает яйца на нижнюю сторону листьев, размещая в кладке от 300 до 800 яиц. Гусеницы развиваются в течение 35—45 дней. Молодые гусеницы живут в гнездах, сплетенных из шелковинок.
В распространении этих бабочек большую роль играют ветры, способствующие их перелетам. Новые очаги этого вредителя обнаруживаются вдоль железнодорожных магистралей и шоссейных дорог. Американская белая бабочка представляет важный карантинный объект государственного значения.
Среди других насекомых чешуекрылые представляют собой сравнительно «молодую» группу: ископаемые бабочки известны только начиная с третичных отложений. В то же время это второй по числу видов отряд насекомых, включающий в себя около 140 000 видов и уступающий по разнообразию форм только отряду жуков. Чешуекрылые распространены по всему миру; особенно много их в тропиках, где встречаются наиболее красивые и наиболее крупные формы, достигающие в ряде случаев в размахе крыльев почти 30 см, как это имеет место у одной из самых крупных в мире бабочек — совки агриппы (Thysania agrippina), распространенной в лесах Бразилии (рис. 328).
Отряд чешуекрылых делят на три подотряда — низших равнокрылых, высших равнокрылых и разнокрылых, отличающихся особенностями строения крыльев.
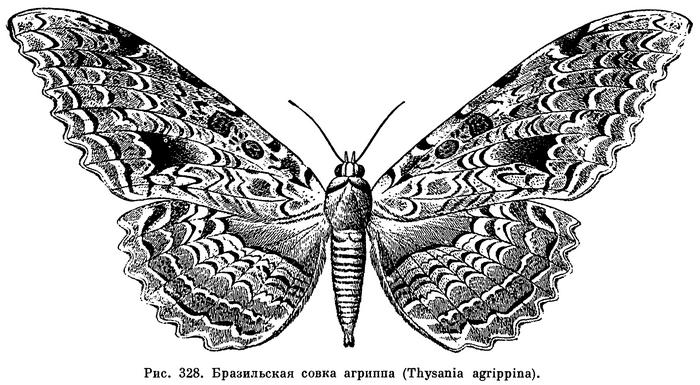
ПОДОТРЯД НИЗШИЕ РАВНОКРЫЛЫЕ БАБОЧКИ (MICROJUGATA)
Сюда относят небольшое число видов бабочек, имеющих примитивное строение крыльев: у них на обоих крыльях почти одинаковое жилкование; переднее крыло несет на внутреннем крае у основания узкую лопастинку, зацепляющуюся за заднее крыло. Хоботок или очень короткий, или совсем отсутствует; на голенях есть шпоры.
В нашей фауне низшие равнокрылые представлены двумя семействами — зубатых первичных молей (Micropterygidae) и первичных молей (Exiocraniidae). Это все мелкие виды, достигающие в размахе крыльев не более 1 см.
У зубатых первичных молей, или мелкокрылов, вполне развиты жующие челюсти, которые снабжены зубовидными отростками, хоботок отсутствует. Поэтому бабочки питаются пыльцой (рис. 329). Гусеницы живут свободно на мхах или лишайниках в сырых местах.
Иной образ жизни у первичных молей. Это типичные обитатели леса. У бабочек имеется короткий хоботок. Гусеницы у них мелкие, до 9 мм, безногие. Наши виды минируют листья березы, реже лещины, каштана и дуба. Мина имеет форму широкого вздутого пятна с характерными внутри нее экскрементами в виде сильно извитых нитей.

ПОДОТРЯД ВЫСШИЕ РАВНОКРЫЛЫЕ БАБОЧКИ (MACROJUGATА)
У высших равнокрылых бабочек на обоих крыльях жилкование тоже почти одинаковое. От представителей предыдущего подотряда их легко отличить благодаря отсутствию шпор на голенях. Кроме того, это средней или крупной величины насекомые, среди которых есть очень большие формы. Так, например, самка новозеландского тонкопряда (Zelotypia staceyi) достигает в размахе крыльев 24 см.
Тонкопряды — сумеречные бабочки; в состоянии покоя крылья у них складываются кровлеобразно. У самого крупного европейского вида — хмелевого тонкопряда (Hepialus humuli) — явно выражен половой диморфизм. У более крупной самки, достигающей в размахе крыльев 7,5 см, передние крылья желтые с красноватыми разводами; самец мельче (размах крыльев — 4,5 см), с серебристо-белыми передними крыльями без всякого рисунка. Желтоватая с черными точками и бурой головой гусеница, длиной до 5 см, живет на корнях или в корнях одуванчика, моркови, хмеля, щавеля и других травянистых растений.
Весьма любопытен кавказский эндемик тонкопряд шамилъ (Phassus schamyl). Это реликт древней тропической фауны, некогда существовавшей на Кавказе. Он относится к роду, включающему в себя около 40 видов, из которых только 4 заходят в Палеарктику. Тонкопряд шамиль — крупная бабочка (размах около 10 см) темно-вишневого цвета; на ее передних крыльях тончайший бурый узор с золотистыми пятнами и прожилками. В июне на закате солнца по опушкам тенистых балок или по окраинам вырубок можно наблюдать брачный полет этих красивых бабочек. Он продолжается всего 10—15 минут и прекращается с наступлением темноты. После брачного полета самец и самка садятся на ветку какого-нибудь куста и начинается спаривание. В это время они становятся похожими на сверток сухих листьев или поломанную веточку с засохшими побуревшими листьями. Светло-соломенные гусеницы тонкопряда достигают в длину 6—7 см. Вначале они держатся в прошлогодней подстилке и питаются полусгнившей листвой; более взрослые гусеницы заползают в почву, где живут в легких шелковистых чехликах — «рукавах», обычно открытых с обеих сторон. С этого времени они начинают питаться корнями различных травянистых растений. В последнее время установлено, что на Черноморском побережье они могут вредить виноградной лозе.
ПОДОТРЯД РАЗНОКРЫЛЫЕ БАБОЧКИ (FRENATA, ИЛИ HETEROPTERА)
К этому подотряду относится преобладающее большинство семейств чешуекрылых, у которых форма и жилкование передних и задних крыльев явно различны, а ротовые части обычно образуют типичный для бабочек хоботок. Передние крылья более или менее треугольной формы, а задние — округленные с менее густым жилкованием.

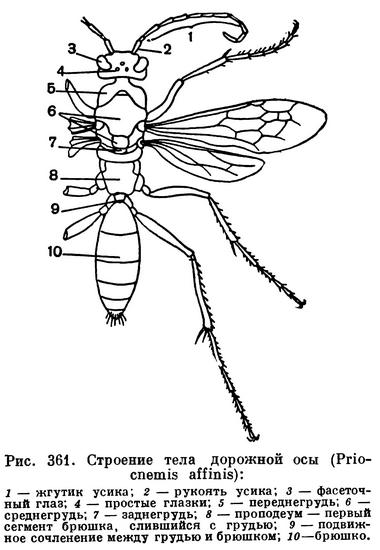
ОТРЯД ПЕРЕПОНЧАТОКРЫЛЫЕ (HYMENOPTERA)
Отряд перепончатокрылых насекомых насчитывает около 90000 видов и по числу видов уступает лишь жукам и бабочкам. К этому отряду относятся как довольно примитивные пилильщики, ложногусеницы которых, похожие на гусениц бабочек, питаются на растениях, так и насекомые с наиболее высокоорганизованной нервной системой и крайне сложной биологией — муравьи, пчелы и осы.
Самое крупное перепончатокрылое — яванская головастая сколия (Scolia capitata) — достигает в длину 6 см, а самое мелкое — наездник-яйцеед алаптус (Alaptus magnanimus) — 0,21 мм. Последний является вообще самым мелким из насекомых.
Взрослые насекомые имеют две пары перепончатых крыльев, покрытых сравнительно редкими жилками, а мелкие формы обычно почти или совершенно лишены жилкования. Задняя пара крыльев меньше и при полете имеет подчиненное значение. У живых насекомых обе пары крыльев обычно скрепляются при помощи крючков друг с другом и работают как одна плоскость. Некоторые виды (рабочие муравьи, самки дриинид, немок и некоторых бетилид и наездников) не имеют крыльев.
Ротовые части грызущие или лижущегрызущие. В последнем случае нижняя губа и нижние челюсти вытягиваются и образуют хоботок с язычком на конце. Такой ротовой аппарат служит для высасывания нектара из цветов. Жвалы хорошо развиты у всех видов и используются не только при питании, но и при строительстве гнезд, рытье почвы и т. д. У некоторых муравьев они имеют причудливую форму и превышают длину головы.
Усики простые, булавовидные, гребневидные, перистые, бывают как прямыми, так и коленчатыми. В последнем случае первый членик их удлинен и носит название рукояти, а остальные членики образуют жгутик. Число члеников усиков варьирует от 3 до 70. У большинства жалящих перепончатокрылых усики самок 12-, а самцов — 13-члениковые. На голове имеется пара сложных фасеточных глаз и 3 простых глазка, но некоторые муравьи совершенно слепы.
Ноги бегательные с 5-члениковой лапкой. Голень и лапка передней ноги иногда несут специальный аппарат для чистки усиков и лапок, образованный гребенчатой шпорой на конце голени и выемкой на первом членике лапки.
Брюшко причленяется к груди двумя способами: у паразитических и жалящих перепончатокрылых первый сегмент брюшка сужен и причленен к груди подвижно, а у сидячебрюхих первый сегмент брюшка широкий и причленен к брюшку всей поверхностью. У муравьев один или два первых членика отделены от брюшка и образуют узелок. Число члеников брюшка варьирует от 3 до 9 (не считая одного членика, который слился с грудью и неотличим от нее). Придатки двух последних сегментов образуют яйцеклад или (у жалящих) жало.
Интересной особенностью перепончатокрылых является то, что самки у них, как правило, откладывают яйца либо гаплоидные (т. е. с одинарным набором хромосом), либо диплоидные (т. е. с двойным набором хромосом). Из первых всегда развиваются самцы, все клетки тела которых гаплоидны, из вторых — только самки (или рабочие общественных перепончатокрылых). В типичных случаях гаплоидные яйца — это неоплодотворенные яйца, а диплоидные — оплодотворенные. В последнем случае диплоидный набор хромосом получается после слияния гаплоидного сперматозоида и гаплоидной яйцеклетки. Однако в ряде случаев наблюдается партеногенез. При этом при образовании яйцеклеток выпадает одно редукционное деление и неоплодотворенные яйца остаются диплоидными. Партеногенез отмечали у некоторых пилильщиков (у этих видов самцы отсутствуют или очень редки) и у рабочих некоторых видов муравьев.
Превращение полное. Личинки обычно имеют более или менее развитую голову. У личинок сидячебрюхих перепончатокрылых имеются грудные ноги, а у личинок пилильщиков также и брюшные. Личинки пилильщиков внешне очень похожи на гусениц и поэтому носят название ложногусениц. От настоящих гусениц бабочек они отличаются тем, что ложные ноги имеются у ложногусениц на 9—11 сегментах, тогда как у гусениц не более чем на 8 сегментах. Личинки паразитических и жалящих перепончатокрылых безногие, обычно белого или желтоватого цвета. Ротовые части у всех личинок грызущие. Куколки свободные, часто в коконе.
Половой диморфизм хорошо выражен. Часто имеется полиморфизм, при котором бывает несколько форм самок. У орехотворок наблюдается чередование поколений с крылатыми и бескрылыми самками. У общественных перепончатокрылых (муравьев, пчел, ос) развивается каста рабочих особей — бесплодных самок, выполняющих различные работы в гнезде. Наиболее резко полиморфизм выражен у муравьев, где рабочие всегда бескрылы. В пределах этой касты у некоторых муравьев наблюдается дальнейшее подразделение на подкасты солдат, «медовых бочек» и т. д. У некоторых видов число резко обособленных подкаст рабочих достигает 6. Все это связано со сложным разделением функций в муравьиной семье.
Образ жизни перепончатокрылых крайне разнообразен. Рогохвосты, как правило, развиваются в древесине деревьев. Личинки большинства пилильщиков питаются листьями растений, и вообще эта группа биологически сходна с бабочками, что отразилось на конвергентном сходстве личинок. Большинство паразитических и некоторые жалящие перепончатокрылые являются паразитами насекомых и некоторых других членистоногих. Среди жалящих перепончатокрылых мы находим огромное разнообразие сложной инстинктивной деятельности, связанной с заботой о потомстве, вершиной которой является «общественное» поведение муравьев, складчатокрылых ос и пчел.
В ископаемом состоянии перепончатокрылые известны с мезозоя. В настоящее время они распространены по всем материкам, кроме Антарктиды. Некоторые перепончатокрылые, например шмели, — одни из самых северных насекомых. Они же поднимаются в горы до вечных снегов.
Экономическое значение перепончатокрылых велико и разнообразно. Прежде всего следует сказать о медоносной пчеле — одном из немногих домашних насекомых, издавна дающем человеку мед, воск и пчелиный клей. Но сейчас стало известно, что наибольшую пользу это насекомое приносит, опыляя культурные растения. Важными опылителями являются и многие дикие пчелы. В последние годы началось одомашнивание еще одних насекомых — шмелей, причем разводятся они именно для того, чтобы с их помощью опылять красный клевер. Подробнее вопрос о роли насекомых в опылении растений рассматривался выше.
Среди сидячебрюхих перепончатокрылых имеется немало серьезных вредителей растений. Большой вред культурным зерновым злакам (ржи, пшенице, ячменю, овсу) причиняют стеблевые пилильщики (Cephidae), особенно хлебный (Cephus pygmaeus) и черный (Trachelus tabidus). Целый комплекс пилильщиков вредит плодовым садам. В плодах развиваются сливовый (Hoplocampa fulvicornis), грушевый (Hoplocampa brevis) и яблонный (Hoplocampa testudinea) пилильщики. Вишневый слизистый пилильщик (Caliroa cerasi) скелетирует листья вишен и других плодовых. Крыжовниковые пилильщики (Pristiphora pallipes, Pteronidea ribesii) объедают листья крыжовника. В садах Средней Азии немалый вред приносит урюковая толстоножка (Eurytoma samsonovi).
Немало среди перепончатокрылых и вредителей леса. В сосновых лесах серьезными вредителями являются сосновый (Diprion pini) и рыжий (Neodiprion sertifer) пилильщики, общественный пилильщик-ткач (Lyda erythrocephala) и др. В годы массовых размножений этих вредителей сосновые леса часто совершенно лишаются хвои.
Но, пожалуй, наибольшее значение перепончатокрылые имеют как объекты, применяющиеся в биологической борьбе с насекомыми-вредителями. Хотя основные сведения о биологическом методе борьбы даются в общем обзоре насекомых, нам кажется необходимым остановиться здесь подробнее на этом вопросе, поскольку паразитические и некоторые жалящие перепончатокрылые являются основными объектами, используемыми в биологической борьбе.
Биологическому методу борьбы с вредителями в последние годы в нашей стране уделяется все большее и большее внимание. Теперь уже стало ясно, что химический метод борьбы имеет целый ряд существенных недостатков, которые не позволяют только с помощью ядов избавиться от вредителей сельского и лесного хозяйства. И основной недостаток этого метода заключается в том, что яды в первую очередь убивают полезных паразитических и хищных насекомых, очень чувствительных к их действию, но не убивают нацело вредителей. В естественных условиях численность растительноядных насекомых сдерживается на низком уровне хищниками, паразитами и болезнями. Поэтому даже если 95% вредителей уничтожается ядами, оставшиеся 5% на следующий год или через год снова дадут вспышку массового размножения.
Вспышки массового размножения вредных насекомых возникают обычно как результат бесхозяйственного отношения к природе. Причинами их могут быть неправильное применение ядов, лесные пожары, сжигающие вместе с подстилкой зимующих в ней наездников, уничтожение подстилки и подлеска в лесах, неправильная агротехника и т. д. Кроме того, когда какое-нибудь животное или растение завозят в новый район, где у него нет естественных врагов, наблюдается его массовое размножение и вскоре такие животные и растения становятся настоящим бедствием. Примером может служить завоз непарного шелкопряда или японского жука в Америку или завоз колорадского жука в Европу.
Ведущее место в биологическом методе борьбы занимают перепончатокрылые. Достаточно сказать, что в 80 хорошо изученных случаях, когда ввоз и использование естественных врагов вредных насекомых привели к тому, что опасность массового размножения этих вредителей была ликвидирована полностью, было применено 4 вида вирусов, 1 вид бактерий, 1 вид паразитических грибов и 133 вида животных, из которых 75 видов — перепончатокрылые.
Существуют 3 направления биологической борьбы с вредными насекомыми: 1) интродукция и акклиматизация новых паразитов и хищников, 2) наводнение паразитами очагов массового размножения вредителей и 3) создание условий для лучшего развития местных энтомофагов.
Интродукция и акклиматизация новых видов используется главным образом для борьбы со случайно завезенными вредителями. Именно в этом направлении достигнуты наибольшие успехи, особенно в Северной Америке и на Гавайских островах. Суть метода заключается в следующем: предпринимается детальное изучение биологии вредителя у него на родине с тем, чтобы выяснить, какие паразиты и хищники являются там его основными врагами. Затем отлавливаются и перевозятся наиболее эффективные естественные враги вредителя. Обычно перевозят сравнительно небольшое число их, а затем их размножают в лаборатории и выпускают в природу.
Биологический метод борьбы во многих отношениях лучше химического метода. Эффективность его значительно выше, а стоимость, как правило, ниже. Однако сейчас еще невозможно применять его в широком масштабе, так как для этого необходимо очень хорошо знать образ жизни сотен вредителей и десятков тысяч паразитов и хищников. Предстоит еще много работы энтомологам, физиологам, биохимикам, агрономам и лесоводам. Но придет время, когда люди смогут заранее проектировать такие сады, леса, поля, в которых будет невозможно появление массовых вредителей. И знание биологии полезных перепончатокрылых будет одним из основных элементов, необходимых для составления таких проектов.
Отряд распадается на три подотряда: сидячебрюхие (Phytophaga), паразитические (Parasitica) и жалящие (Aculeata) перепончатокрылые. Последние два подотряда иногда объединяют в один подотряд стебелъчатобрюхих перепончатокрылых (Apocryta).
ПОДОТРЯД СИДЯЧЕБРЮХИЕ ПЕРЕПОНЧАТОКРЫЛЫЕ (PHYTOPHAGA)
Сидячебрюхие перепончатокрылые являются наиболее примитивными представителями отряда как по строению, так и по биологии. В фауне СССР насчитывается несколько сотен видов, относящихся к этому подотряду, т. е. не больше, чем в каком-нибудь крупном семействе жалящих или паразитических перепончатокрылых.
На примитивность этих насекомых указывает сложное строение их крыльев, имеющих, как правило, полный набор жилок и ячеек, и строение груди и брюшка. Брюшко их причленено к груди всей передней поверхностью и не отделено от груди ни перехватом, ни стебельком.
В отличие от жалящих и многих паразитических перепончатокрылых жизнь сидячебрюхих проходит в основном в личиночной фазе. Взрослые особи выходят из куколок с уже созревшими половыми продуктами, и самцы погибают после спаривания, а самки — после откладки яиц. Взрослые особи кормятся на цветах или вообще не питаются. Лишь у некоторых настоящих пилильщиков (Tenthredinidae) питание смешанное: взрослые насекомые кормятся на цветах и ловят малоподвижных насекомых.
Взрослые сидячебрюхие — довольно беззащитные насекомые: они малоподвижны и лишены жала. Поэтому у них часто встречаются подражательная и покровительственная окраски. Так, например, зеленый пилильщик (Rhogogaster viridis, табл. 50, 19), как видно из его названия, окрашен под цвет листьев, а многие виды родов Cimbex, Tenthredo и др. имеют черное тело с желтым и красным рисунком, как у ос или пчел.
Все сидячебрюхие имеют развитый яйцеклад и, как правило, откладывают яйца внутрь субстрата, которым будет питаться личинка. Лишь у паутинных пилильщиков (Pamphiliidae) самки откладывают яйца на поверхность листьев кормового растения, да и то не совсем свободно: яйцекладом самка делает надрез, в который погружает один конец яйца. По строению яйцеклада, характеру питания личинок и ряду других признаков сидячебрюхие перепончатокрылые делятся на 3 биологические группы: пилильщиков, рогохвостов и оруссид.
Яйцеклад пилильщиков развит слабо. Самка этих насекомых «пропиливает» в сочных частях растений (листьях, реже стеблях) отверстие, в которое и откладывает яйцо. Часто после откладки наблюдается увеличение размеров яйца, возможно, за счет усвоения соков растения. Вокруг таких яиц образуется небольшое вздутие тканей растения. У некоторых мелких настоящих пилильщиков (Tenthredinidae) из этого вздутия образуется галл — болезненное разрастание тканей растения, в котором развиваются личинки. Но у большинства видов пилильщиков личинки после вылупления выходят на поверхность растения.
Личинки большинства пилильщиков (табл. 46, 17—23) очень сходны как внешне (см. характеристику отряда), так и по образу жизни с гусеницами бабочек.
ПОДОТРЯД ПАРАЗИТИЧЕСКИЕ ПЕРЕПОНЧАТОКРЫЛЫЕ (PARASITICA)
Большинство представителей этого подотряда являются паразитами насекомых и некоторых других беспозвоночных и по-русски называются наездниками. Свое название они получили за присущую большинству видов характерную позу при откладке яиц: насекомое садится верхом на жертву и изгибает брюшко вниз, причем жертва часто продолжает при этом двигаться. Сравнительно небольшая часть паразитических перепончатокрылых (орехотворки) паразитирует на растениях, вызывая образование галлов, в том числе «чернильных орешков» на листьях дуба.
У паразитических и жалящих перепончатокрылых, в отличие от сидячебрюхих, первый сегмент брюшка при основании сужен (брюшко стебельчатое) и соединен с грудью подвижным сочленением. Такое сочленение обеспечивает большую подвижность брюшка, и многие из этих насекомых способны даже подгибать конец брюшка к голове. Целесообразность этого становится понятной, если учесть, что откладка яиц в тело живых насекомых требует большой подвижности конца брюшка, на котором помещается яйцеклад.
От жалящих перепончатокрылых паразитические перепончатокрылые отличаются 2-члениковыми вертлугами.
Взрослые наездники и орехотворки питаются, как правило, нектаром цветов, сладким соком растений или падью тлей, а также слизывают гемолимфу, выступающую из ранки, образовавшейся при откладке яйца в тело хозяина. Некоторые наездники даже специально колют хозяина яйцекладом, не откладывая яиц, только для того, чтобы напиться его гемолимфы. Без этого невозможно развитие яиц. А у заражающих скрытоживущих наездников — птеромалид (сем. Pteromalidae) и некоторых браконид (сем. Braconidae) — самки даже имеют специальные приспособления для питания соками хозяина. Самки этих паразитов вводят через стенку укрытия (например, кокона) яйцеклад в тело хозяина, а затем выделяют из придаточных желез легко застывающую жидкость, стекающую по яйцекладу и превращающуюся в трубочку. Затем самка наездника осторожно вынимает яйцеклад и через трубочку начинает питаться соками хозяина. Так поступает, например, паразит зерновой моли (Nemapogon) хаброцитус (Habrocytus cerealellae, рис. 363), паразиты листового люцернового долгоносика (Hypera postica) спинтерус (Spintherus) и дибрахоидес (Dibrachoides dynastes).

ПОДОТРЯД ЖАЛЯЩИЕ ПЕРЕПОНЧАТОКРЫЛЫЕ (ACULEATA)
К жалящим перепончатокрылым относятся наиболее высокоорганизованные представители отряда. У них встречаются удивительнейшие строительные инстинкты, поразительные примеры заботы о потомстве и сложные формы общественной жизни. К этому подотряду относятся хорошо знакомые всем пчелы, осы и муравьи.
Как и у наездников, части груди жалящих перепончатокрылых сильно слились и включают в себя первый сегмент брюшка. Брюшко этих насекомых стебельчатое, как и у наездников. Самым надежным признаком, отличающим жалящих перепончатокрылых от паразитических, является то, что вертлуги их ног одночлениковые, тогда как у всех паразитических перепончатокрылых — 2-члениковые.
Яйцеклад жалящих перепончатокрылых превратился в жало, служащее им как орудие защиты и нападения. Придаточные железы яйцеклада превращаются в ядовитые железы, кислую и щелочную (дюфурову). В яде разных видов преобладают выделения разных желез. У рабочих медоносных пчел яйцеклад служит уже только для защиты: жало зазубрено на конце, и если пчела ужалит крупное животное или человека, жало останется в теле жертвы. Пчела при этом погибнет, а мышцы, вырванные вместе с жалом, будут продолжать автоматически сокращаться и впрыскивать все новые порции яда. Еще удивительнее изменения, происшедшие с яйцекладом рабочих муравьев из подсемейства формицин. Яйцеклад их превратился в крошечные хитиновые пластинки, заметные лишь на препарате под большим увеличением. Зато половину брюшка муравьев занимает огромная кислая ядовитая железа, окруженная мощным мускульным мешком. При сокращении мышц яд, состоящий в основном из муравьиной кислоты, очень ядовитой для насекомых, выбрасывается на расстояние до полуметра, т. е. на расстояние, в 500 раз превышающее длину тела муравья.
Биология жалящих перепончатокрылых отличается большой сложностью и разнообразием. На примере этого подотряда можно видеть, как постепенно усложнялись у насекомых инстинкты заботы о потомстве, что привело в конце концов к созданию одного из удивительнейших явлений природы — «общества» насекомых.
Сложные инстинкты заботы о потомстве можно наблюдать у представителей семейств дорожных ос (Pompilidae), роющих ос (Sphecidae) и ампулицид (Ampulicidae). Хотя каждое из этих семейств возникло, очевидно, самостоятельно, эволюция инстинктов шла у них параллельно, и поэтому образ жизни их довольно сходен. Все эти семейства часто объединяют под общим названием одиночных ос. Знанию биологии этих интересных насекомых мы обязаны великому французскому энтомологу Ж. А. Фабру. В последние годы было получено много новых сведений благодаря исследованиям советского энтомолога С. И. Малышева и энтомолога из ГДР Г. Ольберга.
Семейство складчатокрылых ос (Vespidae) распадается на несколько подсемейств, из которых мы будем рассматривать четыре: стенных ос (Eumeninae), бумажных ос (Vespinae), мазарид (Masaridinae) и полибий (Polybiinae). Для всех складчатокрылых ос характерна одна особенность — они откладывают в ячейку яйцо еще до того, как начнут носить добычу. Благодаря этой особенности в этом семействе по крайней мере дважды (в линиях Eumeninae —> Vespinae и Маsaridinae —> Polybiinae) возник «общественный» образ жизни.
Стенные осы (Eumeninae), как и многие роющие осы, приносят своим личинкам несколько парализованных насекомых. Как правило, в данном случае это личинки мелких насекомых — бабочек (листоверток, огневок, толстоголовок) и жуков (слоников, листоедов, точильщиков). Яйцо самки откладывают не на пол, а подвешивают на потолке ячейки. В СССР наиболее обычны представители двух родов — настоящих стенных ос (Odynerus) и пилюльных ос (Eumenes).

Пчелы (Apidae) — это одно из самых многочисленных семейств перепончатокрылых, насчитывающее около 20000 видов, распространенное всюду, где есть цветковые растения, и переживающее сейчас период своего расцвета. Все представители этого семейства выкармливают своих личинок тестом из пыльцы и нектара цветков, содержащим не меньшее количество белков, чем мясная пища личинок большинства других перепончатокрылых.
Эволюция пчел неразрывно связана с эволюцией цветковых растений, и, наоборот, существование большинства цветковых растений (примерно 90% видов) невозможно без насекомых-опылителей, основными среди которых являются пчелы. Эти насекомые в поисках нектара и пыльцы залезают в цветы и переносят налипшую на них пыльцу на пестики других растений, осуществляя таким образом перекрестное опыление растений.
Строение тела пчел и особенно строение их ног и ротового аппарата (рис. 386) приспособлено для сбора нектара и пыльцы. Лишь наиболее примитивные пчелы, например прозописы, трудноотличимые от роющих ос, переносят, подобно мазаридам, пыльцу в зобике. Все остальные пчелы собирают и переносят ее на поверхности тела. Наиболее совершенно строение собирательного аппарата медоносных пчел, расположенного на задних ногах.
Голень задней ноги медоносной пчелы расширена и ее наружная сторона имеет гладкую вогнутую поверхность, окаймленную по бокам бахромой волосков (корзиночка). Корзиночка расширяется к вершине голени.Несколько отступя от бахромы волосков, торчит один большой волос, изогнутый к входу. И наконец, по самому краю голени расположен ряд острых длинных зубцов, образующих гребень. Первый членик лапки также расширен и имеет форму прямоугольника. С наружной стороны он покрыт 10—12 рядами тонких волосков, носящих название щеточки. Первый членик лапки причленен к голени не всем основанием, а только его передням углом, так что членик может совершать поперечные маятникообразные движения.
Перелетая с цветка на цветок, пчела собирает пыльцу, которая прилипает к многочисленным густым, иногда разветвленным волоскам головы и груди. Затем, главным образом передними ногами, пчела счищает пыльцу с тела, и в конце концов она скапливается в щеточках средних ног. После этого щеточки средних ног зажимаются и протаскиваются между щеточками задних ног, так что пыльца со средней правой ноги переходит на заднюю левую. Таким же путем с левой средней ноги пыльца попадает на заднюю правую. Когда в щеточках набирается достаточное количество пыльцы, пчела гребнем левой ноги вычесывает щеточку правой, и наоборот. В результате на наружной поверхности каждого гребня скапливается по комочку пыльцы. И, наконец, последний акт работы заключается в том, что лапка начинает совершать маятникообразные движения и сдвигает комочек пыльцы внутрь корзиночки, где он удерживается при помощи боковых волосков и изогнутого волоса. Операция эта повторяется многократно, и в конце концов в корзиночке образуется большой комок пыльцы, носящий название обножки. Прилетев в улей, пчела упирается средней ногой в верхний конец комка и выталкивает его из корзиночки в ячейку, предназначенную для хранения пыльцы.
Сходным образом устроены собирательные аппараты шмелей, мелипон и других высших пчел. У стенных пчел, панургусов и др. голени задних ног полностью покрыты длинными волосками, между которыми и набивается пыльца. У короткохоботных пчел для этой цели служат также и волоски на бедрах. У пчел-листорезов, осмий и других мегахилин собирательный аппарат размещен на брюшке. На нижней его стороне имеется брюшная щетка из густых золотистых волосков, куда пыльца набивается при помощи щеточек задних ног. У самцов всех видов и у паразитических пчел собирательный аппарат отсутствует. Редуцирован он и у самок медоносной пчелы, которым никогда не приходится собирать пыльцу.
Ротовой аппарат пчел приспособлен для добывания нектара из венчиков цветков. У высших пчел нижние челюсти и нижняя губа образуют длинный вытянутый хоботок, оканчивающийся заостренным язычком. У разных пород медоносных пчел длина хоботка варьирует от 5,7 до 6,8 мм. У шмелей он несколько длиннее; именно поэтому шмели лучше опыляют цветки красного клевера, имеющие узкие и длинные венчики. А вот у тропических южноамериканских пчел-эвглоссин (Euglossinae) длина хоботка даже превосходит длину тела.
Хоботок примитивных пчел (Colletes и др.) лишь немногим длиннее, чем у роющих ос, и оканчивается тупым широким раздвоенным язычком. У остальных одиночных пчел он имеет строение, промежуточное между хоботком такого примитивного типа и хоботком высших пчел.
Чтобы закончить обзор строения пчел, остановимся на строении их восковых желез. Высшие пчелы для строительства гнезда используют воск. У медоносной пчелы восковые железы расположены на тергитах четырех последних сегментов брюшка. Местоположение их можно обнаружить по светлым пятнышкам, носящим название зеркалец, на внутренней стороне которых расположен слой железистых клеток. Через поры воск выступает наружу и свободно лежит в кармашках зеркалец в виде тонкой треугольной пластинки. Пчела накалывает пластинку на волоски щеточки задней лапки и переносит ее к челюстям. Каждая пластинка весит около 0,25 мг. Для строительства одной ячейки требуется 13 мг воска, или 50 пластинок, а для трутневой ячейки — 30 мг, или 120 пластинок. Восковые железы имеются у рабочих пчел с момента их выхода из куколки, но начинают действовать они только на 3—5-й день и наибольшего развития достигают на 12—18-й день, после чего начинают уменьшаться и перерождаться. У самок и самцов (трутней) этих желез нет.
У шмелей и мелипон воск выделяется тергитами брюшка, у одиночной пчелы-мелитты (Melitta), покрывающей стенки ячеек воскоподобным веществом, оно выделяется придаточной половой железой («щелочной железой» жала).

К подсемейству шмелей (Bombinae) относятся настоящие шмели (Bombus, табл. 51, 4) и паразитирующие в их гнездах шмели-кукушки (Psithyrus).
Шмели — это общественные насекомые. У них имеется разделение самок на более крупных плодущих маток и мелких бесплодных рабочих, выполняющих все основные работы в гнезде. Правда, процесс дифференциации зашел у них еще не слишком далеко и при отсутствии матки рабочие сами могут откладывать яйца.
Ранней весной можно видеть крупных шмелей, летающих низко над землей. Время от времени они садятся на землю и заползают под листву или во всякие норки в земле. Это перезимовавшие самки ищут место для того, чтобы основать новую семью.
Гнездо шмелей представляет собой неправильный шар из травы, мха, прутиков и т. п. Часто оно делается в каком-либо укрытии — в брошенных норах грызунов, между стенами домов и обшивкой, в скворечниках. Известен даже случай,
когда шмели устроили гнездо в стоявшем на террасе чучеле лисицы.
Муравьи (Formicidae) — это самое многочисленное по количеству особей семейство насекомых. Да и по количеству видов немногие семейства могут поспорить с ним. Сейчас насчитывается примерно 6000 видов муравьев, но, учитывая, что каждый год описываются все новые и новые виды и роды и то, что муравьи многих районов земного шара почти не изучены, число это будет значительно увеличено. Все муравьи имеют характерный признак, по которому их можно отличить от любых других перепончатокрылых: между грудью и брюшком у них имеется тонкий стебелек из одного или двух члеников, тогда как у всех остальных перепончатокрылых брюшко непосредственно причленяется к груди.
В этой главе мы уже не раз встречались с примерами общественной жизни у насекомых. Но если у пчел и у ос в пределах одного семейства мы можем наблюдать все этапы перехода от одиночного образа жизни к общественному, то муравьи все без исключения являются общественными насекомыми.
Размножение и расселение муравьев происходит следующим образом: раз в году (у некоторых видов дважды), у каждого вида в свое время, в гнезде появляется множество крылатых муравьев. Это молодые самки и самцы. Некоторое время они еще живут в гнезде, а потом начинают покидать его. В теплые осенние дни, особенно после дождей, над гнездами некоторых наших видов поднимаются целые облака крылатых муравьев. Издали кажется, будто с земли поднимается легкий дым. Массы ласточек, стрижей и других птиц с криками летают в этих роях, хватая муравьев. В воздухе или на земле происходит оплодотворение, после чего самцы вскоре погибают, а самки отгрызают крылья и начинают отыскивать место, подходящее для основания гнезда. Здесь самка роет небольшую норку и откладывает первую порцию яичек, обычно не больше десятка. Когда из яичек вылупятся личинки, она начинает кормить их. У муравьев-бульдогов и некоторых других самки выходят из гнезд и охотятся на насекомых. Но у большинства муравьев самка так и не покидает гнезда до конца жизни, а личинок она кормит выделениями слюнных желез. На образование питательных веществ идут запасы жира и ненужная уже теперь мускулатура крыльев. В пищу идет и большая часть яиц, так что в конце концов до взрослого состояния доживают всего 2—3 рабочих. Обычно они бывают гораздо мельче нормальных. Теперь самка уже перестает кормить личинок и занимается только откладкой яиц. Всю работу берут на себя молодые рабочие.
Просто не верится, что самки некоторых видов могут около года жить не питаясь, и не только жить, но и ухаживать за молодью, копать землю и даже кормить личинок. В том, что это действительно так, можно легко убедиться. В начале осени всюду происходит массовый лёт черного садового муравья (Lasius niger), самого многочисленного муравья в средней полосе Европейской части СССР. В это время бескрылые оплодотворенные молодые самки во множестве бегают всюду в поисках места для устройства гнезда. Их можно встретить даже в больших городах на тротуарах людных улиц. Если наловить таких самок, каждую из них посадить в пробирку с землей, заткнутую ватой, и периодически увлажнять почву, то к концу зимы в некоторых пробирках можно обнаружить первых крошечных рабочих. В природе они появляются к середине лета, так как зимой развитие приостанавливается.
В жизни муравьиной семьи период самостоятельного существования самки наиболее опасен. Подавляющее большинство семей гибнет, так и не успев возникнуть, именно в это время. Поэтому у целого ряда видов появились приспособления, позволяющие избежать гибели. Самый простой способ — это деление старых семей, что аналогично роению медоносных пчел. Так размножаются бродячие муравьи и иногда рыжие лесные муравьи. Самка малайского муравья каребары (Carebara), около 2 см длиной, вылетая из гнезда, прихватывает несколько рабочих, которые имеют длину всего 1—2 мм. Эти рабочие держатся челюстями за ножки самки.
Среди муравьев широко распространено явление временного «социального» паразитизма. Самки таких видов обычно гораздо мельче, чем у видов, самостоятельно основывающих гнезда. После брачного лёта они проникают в уже сформировавшиеся гнезда других видов. Самки рыжих лесных муравьев (Formica rufa) основывают новые семьи в гнездах бурого лесного муравья (Formica fusca) и близких к нему видов, в которых почему-либо нет своей самки. Самки волосистого желтого муравья (Lasius umbratus), проникнув в гнездо черного садового муравья (Lasius niger), убивают его самку. Так или иначе в конце концов получается гнездо, где самка одного вида, а рабочие — другого. Самка кладет яички, а рабочие ухаживают за ней и за молодью. Постепенно рождаются рабочие вида паразита и сменяют естественным путем рабочих хозяина.
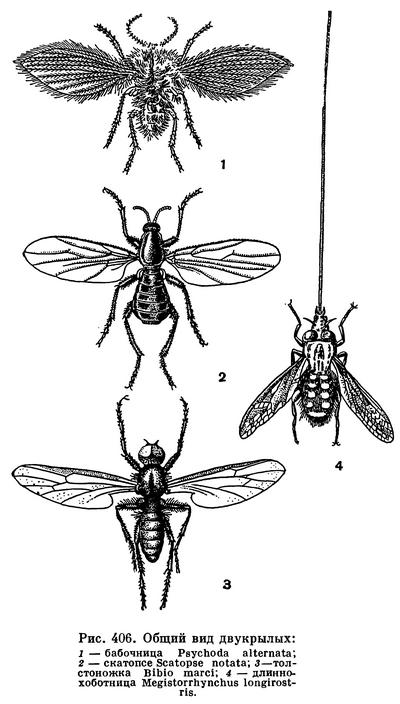
ОТРЯД ДВУКРЫЛЫЕ, ИЛИ МУХИ И КОМАРЫ (DIPTERA)
Среди 33 современных отрядов насекомых отряд двукрылых занимает по численности и разнообразию представителей одно из первых мест, уступая в этом отношении лишь жукам, бабочкам и перепончатокрылым. К настоящему времени в этом отряде известно 80000 видов. Несомненно, в ближайшем будущем эта цифра значительно возрастет, так как изучение двукрылых еще очень далеко от своего завершения.
Основными чертами, обособившими двукрылых от других отрядов насекомых, являются, во-первых, сохранение у взрослой стадии лишь первой пары крыльев — органов быстрого и совершенного полета и, во-вторых, коренное преобразование личиночной стадии, выразившееся в утрате ног, а у высших двукрылых также в редукции головной капсулы и, в конечном итоге, в развитии внекишечного пищеварения.
Форма тела взрослых двукрылых весьма разнообразна. Всем известны стройные длинноногие комары и коренастые короткотелые мухи, но только специалисты отнесут к этому отряду микроскопическую бескрылую «пчелиную вошь» или встречающуюся в муравейниках самку одного из видов горбаток, которая больше похожа на очень мелкого таракана.
Органы зрения — крупные фасеточные глаза — у двукрылых занимают нередко большую часть поверхности их округлой головы. Дополнительно на темени имеются, хотя и не у всех, 2—3 точечных глазка.
Усики, или антенны, расположены на лобной поверхности головы, между глазами. У комаров они длинные, многочлениковые, что является одним из наиболее отчетливых признаков, отличающих подотряд длинноусых двукрылых (Nematocera). У мух, входящих в два других подотряда, антенны сильно укорочены и обычно состоят всего из трех коротких члеников, последний из которых несет простую или перистую щетинку. Антенны — главным образом органы восприятия запахов. На поверхности каждого из члеников имеются специально приспособленные для этого обонятельные бугорки. Нередко антенны самцов двукрылых бывают много сложнее, чем антенны самок. Эти вторичные половые отличия наблюдаются обычно у комаров; у мух же они проявляются чаще в размерах глаз.
Ротовые части двукрылых (рис. 407) сильно изменены и пригодны для приема главным образом жидкой пищи. Наиболее совершенным приспособлением для этого является хоботок высших мух, образованный нижней губой и заканчивающийся сосательными лопастями.
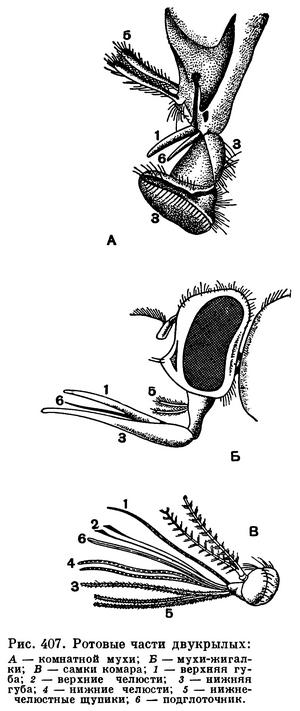

У кровососущих комаров ротовые части сильно вытянуты, нижняя губа образует желобок, в котором располагаются колющие стилеты: игловидные верхние челюсти (мандибулы) и нижние челюсти (максиллы). Между ними находится подглоточник, по которому проходит проток слюнных желез. Сверху желоб нижней губы прикрывается верхней губой.
У некоторых кровососущих мух мандибулы не развиваются и хоботок устроен иначе, чем у комаров. Их нижняя губа образует стилетообразный твердый желоб, вырез которого прикрывает такая же по форме верхняя губа, сцепленная с нижней специальными выростами. Зубчики, которые в хоботке высших мух расположены на сосательных лопастях и служат большинству видов для соскребания твердых пищевых частиц, у кровососов сильно увеличены и используются для вскрытия покровов животных. При этом муха приставляет хоботок вертикально к коже животного и приводит в движение валики, на которых расположены предротовые зубцы. Надрезав верхний защитный слой кожи, эти зубцы довольно быстро высверливают рану. Такими хоботками обладают жигалки, муха це-це и другие близкие виды двукрылых. При проколе покровов насекомых хищными мухами — ктырями и зеленушками — основную роль выполняет нижняя губа вместе с подглоточником. У таких кровососов, как слепни, ранка наносится в основном мандибулами.
Три грудных сегмента двукрылых плотно спаяны между собой, образуя прочный грудной отдел — вместилище мощной мускулатуры. Он служит надежной опорой для крыльев во время быстрого полета. Здесь же расположены жужжальца — короткие булавовидные придатки, представляющие собой видоизмененную вторую пару крыльев. Их считают органами равновесия. Среднегрудь — наиболее мощный грудной сегмент — на заднем крае сверху снабжена полукруглым выростом — щитком.
Передняя пара крыльев у представителей отряда, как правило, хорошо развита. Однако известны также случаи полной утраты крыльев и не только у паразитических форм, но и у свободноживущих видов. Удивительный облик приобретают, например, бескрылые хионеи (Chionea araneoides) из семейства лимониид (Limoniidae), напоминающие скорее пауков, чем мух. Встретить их можно ранней весной, когда они ползают прямо по снегу. Большинство же комаров из этого очень крупного семейства, включающего более 5500 видов, имеют хорошо развитые крылья и внешне очень похожи на комаров-долгоножек.
В покое крылья складываются над брюшком кровлеобразно, горизонтально одно над другим, или же попросту отводятся назад и в стороны. Многие семейства двукрылых лучше всего различаются по жилкованию крыла — рисунку, который образуется на прозрачных крыльях их каркасом — жилками. У хороших летунов особенно прочно укреплен жилками передний край крыла. Поверхность крыльев нередко бывает покрыта крупными и мелкими волосками или чешуйками, а иногда несет дополнительно чувствующие поры. В основании крыла у многих мух обособляются грудная и крыловая чешуйки, а также крылышко.
Строение ног двукрылых тесно связано с их образом жизни. Подвижные, быстро бегающие мухи имеют короткие крепкие ноги. Комары же, днем обычно скрывающиеся среди растительности, обладают длинными конечностями, приспособленными для лазания среди сплетения стеблей травы или в листве деревьев и кустарников. Лапки ног заканчиваются коготками, у основания которых прикрепляются 2—3 особые подушечки-присоски. С их помощью двукрылые могут свободно передвигаться по совершенно гладкой поверхности.
Остроумными опытами доказано, что у мух эти подушечки служат не только для передвижения, но являются дополнительными вкусовыми органами, сигнализирующими о съедобности того субстрата, на который села муха. Если голодную муху поднести к раствору сахара так, чтобы она коснулась его лапками, то муха выдвигает хоботок для сосания. Когда же раствор сахара заменяют водой, муха в этом случае никак не реагирует.
Как грудь, так и брюшко, состоящее у двукрылых из 5—9 видимых сегментов, часто имеют характерную расцветку и усажены волосками и щетинками. Расположение этих щетинок часто используется как признак для различения отдельных семейств, родов и видов отряда.
Представление о личинках двукрылых как о беловатых, безногих и безголовых «червях», копошащихся в навозе и помойках, совершенно не отражает истинного многообразия их форм и основано на самом поверхностном знакомстве с отрядом.
Прежде всего следует подчеркнуть, что у личинок всех длинноусых двукрылых голова хорошо развита и часто снабжена сильными челюстями, с помощью которых личинки питаются корешками растений или гниющими органическими веществами. Единственным исключением является редкое семейство длинноусых двукрылых — гиперосцелидиды (Hyperoscelididae). У личинок гиперосцелидид полностью отсутствует головная капсула, их головной сегмент несет лишь пару усиков и ротовое отверстие. Живут эти личинки в разлагающейся древесине и питаются исключительно жидкой пищей.
Головная капсула никогда не развивается у личинок высших мух, весь ротовой аппарат которых представлен обычно только двумя склеротизованными крючьями.
Утрата головной капсулы, столь характерная для личинок высших двукрылых, связана с развитием у них своеобразного способа пищеварения, которое получило название внекишечного. При этом типе пищеварения пища предварительно переваривается вне тела личинки под воздействием выделяемых ею пищеварительных соков и лишь потом заглатывается и усваивается.
Разнообразна форма тела личинок. Обычно она червеобразная, но иногда бывает настолько необычной, что может поставить в тупик неопытного систематика.
Пожалуй, единственным постоянным признаком личинок двукрылых является отсутствие грудных (истинных) ног. Отсутствие ног у личинок мух в ряде случаев компенсируется развитием у них различных выростов тела, напоминающих «ложные ноги» гусениц бабочек. С помощью этих выростов личинки могут сравнительно быстро передвигаться по поверхности субстрата.
В обильном пищевом субстрате личинки двукрылых встречаются большими скоплениями. Обычными местами массового развития личинок высших мух являются разлагающиеся трупы животных, помойки, уборные и т. п.
Процесс превращения взрослой личинки в куколку у двукрылых имеет свои особенности. Обычно у насекомых с полным превращением, после того как под покровами личиночной шкурки сформируется куколка, эти покровы сбрасываются и куколка полностью освобождается.
Длинноусые двукрылые не представляют исключения из этого правила. Зато у целой группы высших мух имеется специальное дополнительное защитное приспособление, оберегающее куколку от повреждений и получившее название пупария. В этом случае шкурка взрослой личинки не только не сбрасывается, как ненужная оболочка, но, наоборот, затвердевает, приобретает бочонковидную форму и укрепляется различными отложениями. Куколка же формируется внутри этой шкурки, а взрослая муха, чтобы оказаться на свободе, выламывает в ней круглое выходное отверстие.
Эта биологическая особенность положена в основу при выделении в отряде двукрылых, кроме подотряда длинноусых, или комаров (Nematocera), еще двух подотрядов: короткоусых прямошовных двукрылых (Brachycera-Orthorrhapha), не имеющих пупария, и короткоусых круглошовных двукрылых (Brachycera-Cyclorrhapha), развивающихся с пупарием. Интересно, что личинки некоторых групп двукрылых хотя и не формируют типичного пупария, но окукливаются все же внутри личиночной шкурки. Среди длинноусых двукрылых такой способ окукливания характерен для небольшого семейства скатопсид (Scatopsidae), насчитывающего около 130 видов, и для немногих видов семейства галлиц (Cecidomyiidae), таких, как гессенская мушка и некоторые другие. Из короткоусых прямошовных двукрылых внутри слабо измененной личиночной шкурки окукливаются личинки львинок.
Необычайно широка приспособляемость двукрылых к различным жизненным условиям. Их личинки освоили самые разнообразные среды обитания: быстрые потоки и стоячие воды, чистые, прозрачные водоемы, в том числе и моря с соленой водой, и зловонные сточные канавы, толщу почвы, различные попадающие в почву гниющие растительные вещества, ткани живых растений и, наконец, полость тела насекомых и других беспозвоночных животных, а также кишечный тракт, подкожную клетчатку и дыхательные пути позвоночных животных, а в ряде случаев и человека.
Личинки двукрылых ведут скрытый образ жизни и неспособны к длительным передвижениям. Пристроить свое потомство в подходящие условия — задача взрослых мух, которые поэтому являются хорошими летунами. Многие из них имеют интересные приспособления, повышающие выживаемость личинок. Достаточно вспомнить о распространенном среди высших двукрылых рождении живых личинок, а в некоторых случаях о вскармливании личинок выделениями специальных желез, когда личинка покидает тело матери, будучи уже совсем взрослой.
Однако обычно не взрослые мухи кормят своих личинок, а, наоборот, личинки запасают питательные вещества, необходимые для жизни взрослой фазы.
Двукрылые являются одной из самых многочисленных групп насекомых и поэтому представляют собой большую силу природы. И сила эта, если оценивать значение двукрылых в целом, наносит огромный ущерб не только хозяйству, но и здоровью человека.
В природе существуют многочисленные очаги различных болезней, которыми болеют дикие животные. Во многих случаях эти болезни не опасны для человека, но некоторые из них представляют для людей чрезвычайно серьезную угрозу. Существуют также болезни, которые не передаются от человека к человеку, но тем не менее очень широко распространены. Кровососущие двукрылые, нападающие на животных и человека, наряду с другими кровососущими членистоногими, широко разносят эти болезни, передавая возбудителя во время сосания крови.
Главная опасность от малярийного комара не в том, что он нанес болезненный укус, а в том, что одновременно он может занести в кровь возбудителей малярии, а только одна эта болезнь унесла человеческих жизней много больше, чем все войны в истории человечества, вместе взятые.
Столь же опасными переносчиками инфекций являются синантропные двукрылые, т. е. виды, обитающие в жилищах человека. Посещая отбросы и фекалии, они переносят болезнетворные микроорганизмы и яйца глистов на своем теле и в кишечнике, оставляя их на посуде, пище, мебели и т. д. Недаром многие коллективы ученых трудятся над изучением биологии одного из таких насекомых — комнатной мухи — с целью ее истребления.
Среди двукрылых немало видов, которые приносят человеку вред, уничтожая плоды его трудов. Печальной известностью пользуются злаковые мухи, личинки которых губят посевы зерновых культур. Трудно назвать такое культурное растение, которое бы в той или иной степени не повреждалось представителями этого отряда. То же в полной мере относится и к домашним животным. Паразитические двукрылые, развивающиеся за счет домашних животных, не только истощают и частично губят их, но и делают непригодными их мясо и шкуры, снижают удои молока и т. д. Достаточно упомянуть, что только различные виды оводов наносят ежегодно огромный урон животноводству.
Для здоровья человека личинки сырной мухи представляют опасность тогда, когда употребляются в пищу зараженные ими продукты. В кишечнике человека личинки способны продолжительное время сохранять жизнеспособность, вызывая изъязвления кишечной стенки, с симптомами, напоминающими заболевание тифом.
Нельзя недооценивать и отрицательного значения тех двукрылых, которые нападают на человека во время его работы в полевых условиях, значительно снижая производительность труда, а в ряде случаев делая этот труд в определенные периоды невозможным.
Положительная роль двукрылых в природе и в хозяйстве человека невелика по сравнению с приносимым ими вредом. Они являются неутомимыми санитарами, очищающими поверхность суши от накапливающихся здесь отбросов. Некоторые группы двукрылых известны как почвообразователи и как враги вредных насекомых, сдерживающие их размножение.
Распространены двукрылые очень широко: от тропиков до границ льдов на севере и в горах. Но даже среди тропических представителей отряда почти нет особенно крупных и ярко окрашенных видов. Любители насекомых уделяют им мало внимания, отдавая предпочтение жукам и бабочкам, хотя биологически двукрылые не менее интересны и своеобразны.
ПОДОТРЯД ДЛИННОУСЫЕ ДВУКРЫЛЫЕ (NEMATOCERA)
Комары имеют стройное удлиненное тело и тонкие, обычно длинные ноги, реже они плотные, приземистые, с короткими ногами. Их антенны состоят более чем из трех члеников. У личинок головная капсула хорошо развита. Куколки покрытого типа.
Велико значение комаров как переносчиков возбудителей таких тяжелых заболеваний, как малярия в различных ее формах, желтая лихорадка, вызываемая вирусом, японский энцефалит, энцефаломиелит и др. Лишь хорошо разработанная научная система профилактики этих болезней в СССР и некоторых других странах позволила резко снизить заболеваемость людей. Для борьбы с комарами успешно были применены не только химические, но и биологические меры борьбы. Завезенная из Америки небольшая живородящая рыбка гамбузия акклиматизировалась в Средней Азии, где стала одним из основных врагов комариных личинок.
Комары являются первым из пяти основных семейств кровососущих двукрылых, комплекс которых получил меткое название «гнус». Вместе со слепнями, мокрецами, мошками, а на юге также и москитами комары образуют полчища двукрылых, которые, особенно в болотистых таежных местах, в летние месяцы не дают ни минуты покоя, нападая на животных и человека.
ПОДОТРЯД КОРОТКОУСЫЕ ПРЯМОШОВНЫЕ ДВУКРЫЛЫЕ (BRACHYCERA-ORTHORRHАРНА)
Это типичные мухи с компактным коротким телом и широкими сильными крыльями. Их антенны состоят из 3 члеников, но последний из них может сохранять следы дополнительного расчленения. Головная капсула личинок сильно редуцирована. Личиночная шкурка при окукливании, как правило, сбрасывается. Куколка покрытая, при выходе мухи ее головогрудь растрескивается по Т-образной линии.
Слепни (семейство Tabanidae) — крупные кровососущие двукрылые. Самка слепня способна за одно кровососание принять до 200 мг крови, т. е. столько, сколько выпивают 70 комаров или 4000 мокрецов. Если к этому добавить, что в болотистых местностях в жаркие летние месяцы стада домашних животных атакуют десятки тысяч слепней, становится понятным огромное отрицательное значение слепней в природе и хозяйстве человека. Их вредность усугубляется еще и тем, что в момент кровососания слепни переносят возбудителей сибирской язвы, туляремии, полиомиелита и других тяжелых заболеваний, а также передают некоторые болезни, вызываемые нематодами.
Слепней иногда неправильно называют оводами. Однако достаточно убедиться, что пойманная на животном муха имеет короткий колющий хоботок, чтобы с уверенностью отнести ее к слепням. Красивы крупные глаза слепней — золотистые, переливающиеся всеми цветами радуги. Крылья их иногда прозрачны, иногда с дымчатыми пятнами, брюшко всегда уплощено.
ПОДОТРЯД КОРОТКОУСЫЕ КРУГЛОШОВНЫЕ ДВУКРЫЛЫЕ (BRACHYCERA-CYCLORRHAPHA)
Типичные мухи с коротким компактным телом и широкими сильными крыльями. Их антенны укороченные, 3-члениковые, со щетинкой на третьем членике. Головная капсула личинок полностью редуцирована, сохраняются лишь ротовые крючки. Личиночная шкурка при окукливании не сбрасывается, приобретает бочонковидную форму и затвердевает, пропитываясь специальными выделениями, образуя ложный кокон — пупарий. Куколка свободная. При выходе взрослой мухи пупарий открывается по округлой линии под давлением головы или лобного пузыря, который в большинстве случаев хорошо развит.
На цветках зонтичных и сложноцветных часто рядом с осами и шмелями сидят очень похожие на них мухи из семейства журчалок (Syrphidae, табл. 59). Хотя эти мухи совершенно безобидны, птицы не решаются их трогать, принимая за вооруженных жалом перепончатокрылых. В семействе журчалок около 4500 видов.
Навозные мухи (семейство Scatophagidae) получили свое название потому, что их наиболее массовые виды обычны на экскрементах животных, т. е. являются копробионтами. Такова рыжая навозница (Scatophaga stercoraria) — крупная муха, длиной до 10 мм, желто-бурого цвета с густыми ржаво-желтыми волосками и несколько более светлыми крыльями того же оттенка (табл. 60, 8). Ее личинки хищничают в навозе и фекалиях.
Свыше 3000 видов насчитывается в семействе настоящих мух (Muscidae). Их облик легко представить, вспомнив всем известную комнатную муху.
Биология настоящих мух очень разнообразна, и ее трудно охарактеризовать в целом. Большая часть входящих в семейство видов развивается в гниющих органических остатках растительного и животного происхождения, где их личинки либо перерабатывают сами остатки, либо хищничают. Ряд видов настоящих мух является вредителями растений. Нет в составе семейства только специализированных паразитов, хотя паразитизм в наиболее примитивных, несовершенных формах у этих мух также известен.
Многие виды настоящих мух являются синантропными, т. е. более или менее тесно связанными с человеком. Некоторые из них, например комнатная муха (Musca domestica, рис. 423), уже не встречаются в дикой природе, вне поселков и городов. Навоз, фекалии, различный мусор — вот те отбросы, где развиваются личинки комнатной мухи — постоянного спутника людских поселений. Поразительна скорость размножения этого вида. За один раз самка в среднем откладывает около 100—150 яиц, но при достаточном питании яйцекладки повторяются с интервалом в 2—4 дня, так что ее общая плодовитость составляет в итоге 600, а в странах с жарким климатом 2000 и более яиц. Если бы личинки, куколки и сами мухи не погибали, то потомство только одной самки к концу лета могло бы превысить 5 триллионов (5 000 000 000 000) экземпляров.
Личинки комнатной мухи, как и других высших мух, не имеют головы. Они разжижают пищу, выпуская на нее пищеварительные соки, — такой способ пищеварения носит название внекишечного. В результате вся колония личинок мух оказывается плавающей в разжиженной полупереваренной среде, которая ими постоянно заглатывается. В результате пища используется с удивительной экономией. В одном литре конского или коровьего навоза или в таком же количестве кухонных отбросов может одновременно развиться от 1000 до 1500 личинок мух, а в свином навозе — до 4000.
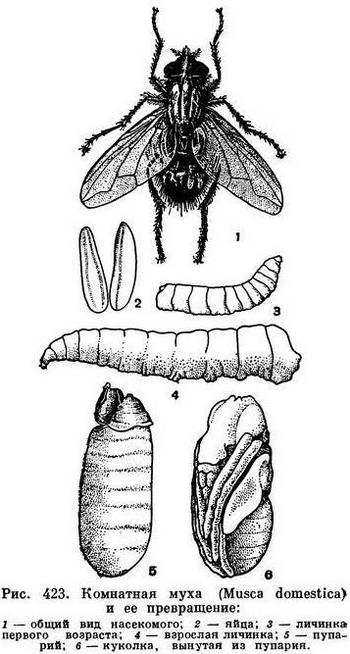
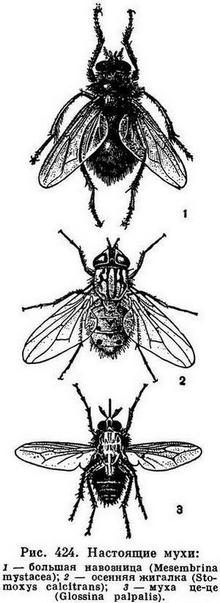
ОТРЯД БЛОХИ (SIPHONАРТЕRА)
Среди насекомых с полным превращением блохи — единственный отряд, все представители которого совершенно бескрылы. Обычно бескрылость блох объясняют паразитическим образом жизни. Однако существует много групп паразитических насекомых, у которых крылья развиты.
Блохи — наружные паразиты млекопитающих и птиц, поверхность тела которых по сравнению с поверхностью тела этих мелких паразитов огромна, а переход с одного хозяина на другого легко осуществляется активным переползанием при встрече животных-хозяев или прыжками в местах их постоянного обитания (норы и т. д.). Отсюда их бескрылость.
Строение тела блох приспособлено к движению в шерстном покрове животного-хозяина примерно так же, как строение тела саранчового к движению в травянистом покрове — оно сильно сплющено с боков. Задние ноги у блох прыгательные, лапки всех ног прекрасно развитые, 5-члениковые, заканчивающиеся 2 коготками. Голова маленькая, на голове короткие усики, перед ними по простому глазку.
Ротовые части блох приспособлены для прокалывания кожи и высасывания крови, прокол кожи осуществляют зазубренные жвалы. Питаясь, блохи наполняют кровью способный сильно раздуваться желудок.
Самцы у блох мельче самок. Оплодотворенные самки с силой выбрасывают яйца, обычно порциями по нескольку штук так, что яйца не остаются на шерсти животного, а падают на землю, обычно в норе животного-хозяина или в других местах, им постоянно посещаемых.
Из яйца выходит безногая, но очень подвижная червеобразная личинка (рис. 430, 2) с хорошо развитой головой. Для дальнейшего развития личинка нуждается в достаточной влажности, поэтому она зарывается в землю или мусор в гнезде или норе хозяина. Личинка питается различными разлагающимися остатками, а у многих видов нуждается, кроме того, в питании остатками непереваренной крови, содержащейся в испражнениях взрослых блох.
Выросшая личинка прядет себе паутинный кокон, сверху защищенный пылью и песчинками, и окукливается в нем. Куколка у блох типичная свободная. Вышедшая из куколки взрослая блоха подкарауливает животное-хозяина.
Некоторые более многоядные блохи могут нападать на любое животное; известно, что блоха однопроходных и сумчатых в Австралии (Echidnophaga ambulans) временно питается на змеях, а некоторые наши блохи — даже на гусеницах! Но нормально существовать и размножаться блохи могут только на подходящих им животных.
На человеке паразитирует человечья блоха (Pulex irritans), достигающая длины 3—4 мм (самка). Эта блоха может жить также на собаках, кошках и даже лошадях, т. е. на животных, имеющих постоянный контакт с человеком.
Много блох бывает там, где есть места для развития их личинок — грязные трещины в полу, грязные ковры и т. д., где в пыли и мусоре личинки могут питаться разными разлагающимися остатками и испражнениями взрослых блох.
В XVII—XVIII веках во дворцах французской знати было так много блох, что одним из предметов обихода придворных дам были «блохоловки» — небольшие золотые коробочки вроде медальона с маленьким отверстием, носившиеся на цепочке на шее, в которые блохи могли попасть чисто случайно. Сколько же там было блох!
На кошках живет кошачья блоха (Ctenocephalus felis), редко и неохотно переходящая на людей, а на собаках — собачья (Ctenocephalus canis).
В предгорьях Тянь-Шаня на лошадях, верблюдах и рогатом скоте паразитирует блоха алакурт (Vermipsylla alakurt). Оплодотворенная самка прикрепляется к телу животного и высасывает очень много крови, ее тело при этом сильно раздувается. Эта блоха, в массе размножаясь, истощает животных (рис. 431, 1).
Много блох разных родов и видов связано с грызунами. На сусликах живет сусличья блоха (Ceratophyllus tesquorum) и Neopsylla setosa, на тарбаганах — тарбаганья (Oropsylla silantievi), на крысах — Xenopsylla cheopis, Ceratophyllus fasciatus и др.
Удивительная блоха обитает в тропиках. Эта мелкая (1 мм), известная еще К. Линнею блоха Dermatophilus penetrans, родина которой — тропическая Америка, расселилась, будучи завезенной человеком, и по всей Африке, и на Мадагаскаре. Оплодотворенная самка внедряется в кожу человека, грызуна или другого животного-хозяина, сосет кровь, и под кожей ее брюхо пузыревидно расширяется (рис. 431, 3). На коже около тела блохи образуется язва, часто гноящаяся, через которую блоха выстреливает свои яйца. Если такую блоху в ранке раздавить, начинаются опасные воспаления. В экономически неразвитых странах живущие в тяжелых условиях неподвижные больные сильно страдают от этой блохи. У грызунов нередко происходит обламывание пальцев конечностей, на которых поселяются эти блохи, вызывающие сильные воспаления.
Блохи — отряд насекомых, который трудно сблизить с другими группами. По строению личинки они напоминают двукрылых, куколка и некоторые детали строения взрослых блох позволяют сближать их с жуками.