(Isopoda) Отряд Изоподы, Равноногие ракообразные, Order Isopoda 116 семейств
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
НАДОТРЯД ПЕРАКАРИДЫ (PERACARIDA)
Этот обширный надотряд объединяет очень различных как по внешнему виду, так и по образу жизни ракообразных, обитающих в море, в пресных водах и на суше, ползающих по дну и плавающих в толще воды, большей частью свободноживущих, но частично и паразитирующих на других животных. Однако все они обладают некоторыми общими признаками, указывающими на их родство между собой.
Голова всех перакарид срастается с одним или изредка с двумя передними грудными сегментами, конечности которых превращаются в ногочелюсти. Правая и левая жвалы устроены различно: на левой обычно есть подвижная пластинка, отсутствующая на правой. Самки вынашивают яйца под грудью в выводковой сумке, образованной направленными внутрь листовидными отростками грудных ножек (только у отряда Thermosbaenacea выводковая сумка помещается на спинной стороне тела, под карапаксом). Из яиц выходят почти вполне сформировавшиеся молодые рачки, так что настоящих личинок у перакарид нет.
К надотряду Peracarida относятся отряды мизид, кумовых, клешненосных осликов, спелеогрифовых, термосбеновых, равноногих и разноногих ракообразных.
ОТРЯД РАВНОНОГИЕ РАКООБРАЗНЫЕ (ISOPODA)
Равноногие ракообразные (табл. 33) — один из немногих отрядов этого класса, многочисленные (4500 видов) представители которого приспособились к самым различным условиям существования. Большинство относящихся к нему видов обитает в море, причем населяет все глубины, начиная от приливо-отливной зоны и кончая предельными океаническими глубинами порядка 11 км. Среди морских равноногих есть обитатели дна, но есть и немногочисленные планктонные виды; есть также и паразиты других ракообразных и рыб. Чрезвычайно широко распространены равноногие и в пресных водах, как поверхностных, так и подземных; пресноводные водяные ослики могут считаться очень обычными и многочисленными представителями фауны самых различных озер и прудов северного полушария. Наконец, богатый видами подотряд равноногих — мокрицы — выработал весьма совершенные приспособления к жизни на суше. Они встречаются почти во всех климатических зонах земного шара — от холодных приполярных районов до влажных тропических лесов и раскаленных пустынь. Очевидно, в процессе эволюции равноногие приобрели какие-то важные особенности, позволившие им впоследствии специализироваться в разных направлениях и освоить различные среды обитания. Одной из таких особенностей можно считать строение брюшного отдела и его конечностей, выполняющих различные функции — половую, двигательную и дыхательную — и хорошо защищенных от неблагоприятных внешних воздействий, в частности у сухопутных рачков от высыхания.
Тело равноногих обычно уплощено сверху вниз, реже цилиндрическое или даже сжатое с боков (у подотряда Phreatoicidea, табл. 33, 4). Его покровы очень разнообразны, часто гладкие, но нередко снабжены килями, шипами или различными бугорками. У некоторых мокриц бугорки поверхности тела образуют сложную водопроводную систему. Если рачок прикоснется к капельке росы, влага под действием капиллярных сил начинает распространяться в промежутках между бугорками, достигая заднего конца тела и орошая находящиеся там жабры. Также разнообразна окраска равноногих. Обычно она бурая или желтоватая, но у видов, живущих на морских растениях, она зеленая, под цвет субстрата, а подземные и глубоководные виды лишены пигмента, белые или иногда розоватые.
Карапакс полностью отсутствует. Голова срастается с одним или изредка с двумя передними грудными сегментами. На ней с боков находятся сидячие глаза. Число составляющих их фасеток колеблется в очень широких пределах: у самого крупного вида равноногих — Bathynomus giganteus (табл. 33, 10), достигающего в длину 27 см, в каждом глазе насчитывается 3000 фасеток, у более мелких хищных морских Aega (табл. 33, 6) — около 400, у мокриц — обычно 20—30, а у водяных осликов — 4. У глубоководных и подземных видов глаза не развиты совсем. Обе пары антенн имеют разную длину у различных видов. У многих глубоководных равноногих задние антенны значительно длиннее тела. Строение ротовых придатков зависит от способа питания. У большинства равноногих они жующего типа, но у некоторых хищников и паразитов жвалы и челюсти превращены в шиловидные острия и сложены вместе в виде пирамиды, так что ротовой аппарат приобретает характер колюще-сосущего.У взрослых особей из подотряда Gnathiidea ротовые придатки, за исключением жвал самцов, редуцированы, и рачки вовсе не питаются (табл. 33, 1,2).
Семь свободных грудных сегментов несут по паре ходильных ног, всегда лишенных наружных придатков. У многих равноногих эти ножки имеют приблизительно одинаковое строение и одинаковую длину, почему отряд и получил свое название. Однако у других они специализированы в разных направлениях. Нередко на одной передней или даже на трех передних ножках развивается хватательное приспособление — подклешня. У многих представителей подотряда Asellota, в особенности часто у глубоководных, конечные членики трех задних пар грудных ножек расширены и снабжены по краям многочисленными щетинками. Такие ножки служат для плавания (табл. 33, 11,12). У половозрелых самок на передних пяти или четырех (у некоторых Arcturidae на одной) парах грудных ножек с внутренней стороны возникают пластинки, образующие выводковую сумку.
Особенно характерно строение брюшного отдела и его конечностей. Он всегда гораздо короче грудного, и количество составляющих его сегментов сокращено за счет того, что один, несколько или даже все сегменты срастаются с полукруглым или треугольным тельсоном, образуя крупный плеотельсон. Только у подотряда Anthuridea этого не происходит и последний брюшной сегмент и тельсон сохраняют самостоятельность. Конечности пяти передних брюшных сегментов состоят из короткого основания и двух широких листовидных, часто тонкостенных ветвей. Эти ветви накладываются друг на друга, как страницы книги, и направлены назад таким образом, что они находятся под плеотельсоном (рис. 244, 2). Основная функция передних брюшных ножек дыхательная. Они служат жабрами. Через их тонкие стенки растворенный в воде кислород проникает в проходящие в ножках кровеносные лакуны и окисляет кровь. В связи с этим, как уже указывалось, сердце у равноногих ракообразных в значительной степени смещено в брюшной отдел и расположено близко от жабр. Наружные ветви одной из пар брюшных ножек обычно сильно хитинизированы и служат крышкой, покрывающей остальные нижние жаберные листочки, которые, таким образом, оказываются сверху защищенными плеотельсоном, а снизу этой крышкой (рис. 244, 1).
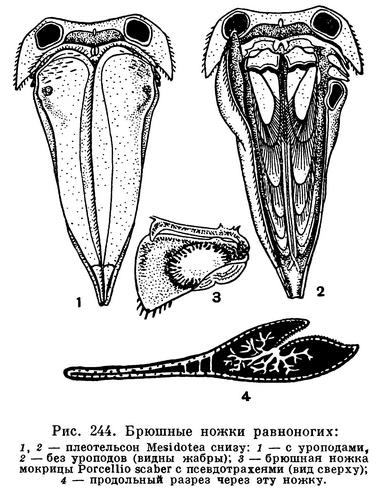
Такое строение дыхательного аппарата послужило, несомненно, предпосылкой, позволившей части равноногих перейти к сухопутному существованию. Их жабры оказались хорошо защищенными от высыхания и не потребовалось дополнительных усовершенствований для того, чтобы полностью их обезопасить в новых условиях существования. Большинство мокриц, несмотря на то что они живут на суше, дышат при помощи своих брюшных ножек, как настоящие водные животные: они используют кислород, растворенный в тонком слое покрывающей их жабры влаги. Следовательно, их жабры должны все время увлажняться. Это достигается у разных мокриц разными путями. У одних, как уже упоминалось, приток воды к жабрам обеспечивается благодаря своеобразной водопроводной системе, образованной скульптурой покровов. У других задние брюшные ножки — уроподы — устроены таким образом, что при соприкосновении с капелькой воды она, также под влиянием капиллярных сил, поступает в жабры по уроподам. Наконец, третьи орошают свои жабры жидкостью, выделяемой из анального отверстия и добываемой из поедаемой рачками сочной растительности. Однако есть мокрицы, способные при дыхании использовать не только растворенный в воде, но и атмосферный кислород. У таких мокриц в толще наружных ветвей передних брюшных ножек находится полость, соединяющаяся щелью с наружной средой, так что в эту полость свободно проникает воздух. От нее отходят ветвящиеся, заканчивающиеся слепо каналы, напоминающие трахеи насекомых и других наземных членистоногих и поэтому называемые псевдотрахеями (рис. 244, 3,4). Через них кислород воздуха проникает в кровь, циркулирующую в лакунах в толще наружных ветвей брюшных ножек, а кислород, растворенный в смачивающей жабры воде, используется внутренними ветвями.
Судя по наблюдениям над туркменскими мокрицами рода Protracheoniscus, размеры трахейного поля наружных ветвей брюшных ножек зависят от условий существования. Виды, обитающие в пустыне и на других открытых сухих местах, имеют относительно крупное трахейное поле, а виды, существующие при повышенной влажности, наоборот, относительно небольшое. Совсем маленькое трахейное поле свойственно виду Protracheoniscus birsteini, который вторично перешел от жизни на суше к жизни в воде и был обнаружен среди зарослей водных растений в обширных мелководных озерах Делили в бассейне реки Атрек.
У некоторых морских равноногих дыхательная поверхность брюшных ножек увеличивается за счет образования складок, как у некоторых Sphaeromidae, пальцевидных выростов, как у Bathynomus, или продольного расщепления ветвей, как у некоторых представителей паразитического подотряда Epicaridea.
Однако передние брюшные ножки равноногих служат не только для дыхания. У самцов некоторые из них частично преобразованы в совокупительные органы. Эту функцию у многих выполняют внутренние ветви второй пары. У водяных осликов, например, они представляют собой нечто вроде шприцев весьма сложного строения. При спаривании самец сначала набирает в этот шприц свою сперму, а затем впрыскивает ее в половые отверстия самок. Первая пара брюшных ножек также принимает участие в этом процессе. Сходным образом, при помощи двух пар передних брюшных ножек, производят перенос спермы в половые пути самок мокрицы. Понятно, что по строению передних брюшных ножек самцы равноногих всегда отличаются от самок, у которых первая пара брюшных ножек нередко совсем отсутствует. Некоторые группы равноногих используют брюшные ножки не только как органы дыхания и совокупления, но и как органы движения. Anthuridea при плавании над дном прижимают грудные ножки к бокам тела и гребут брюшными. Также при помощи трех пар передних брюшных ножек плавают плоские антарктические Serolidae (табл. 33, 9), некоторые Cymothoidae и др.
Последняя пара брюшных ножек — уроподы — имеет разное строение у разных подотрядов равноногих. Так, например, у Asellota и мокриц они прикреплены к заднему краю плеотельсона и направлены назад, у Valvifera они с боков подвернуты под брюшную сторону плеотельсона и, как крышки, прикрывают передние нежные дыхательные брюшные ножки (рис. 244, 1,2), у Anthuridea и Flabellifera они сочленены с основанием плеотельсона и вместе с ним образуют хвостовой веер.
У паразитических Epicaridea, в особенности у их самок, организация всего тела обыкновенно очень упрощена. Они нередко теряют сегментацию и конечности и превращаются в бесформенный мешок, набитый яйцами. Однако самцы и личинки полностью сохраняют признаки равноногих ракообразных.
Основным способом передвижения всех равноногих ракообразных, за исключением паразитических, следует считать ползание по дну, почве или растениям при помощи грудных конечностей. Большинство видов двигается только таким способом. Среди морских видов многие, кроме того, могут плавать. Обыкновенно они проплывают небольшие расстояния над дном, с тем чтобы потом снова перейти к ползанию. При этом представители некоторых семейств Asellota гребут специально приспособленными к плаванию задними грудными ножками, a Anthuridea, Cymothoidae и Serolidae — брюшными ножками. Idoteidae орудуют при плавании и задними грудными и брюшными ножками.
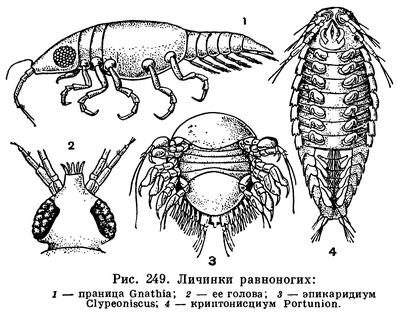
Для некоторых равноногих плавание играет важную роль в их жизни. Полупаразитические полухищные виды Aega и некоторых других Cymothoidae вплавь нападают на рыб, прикрепляются к ним и начинают сосать их кровь. Личинки паразитических Epicaridea плавают при помощи задних антенн и брюшных ножек до тех пор, пока не найдут своих хозяев — различных ракообразных, после чего начинается превращение личинок во взрослые формы. Так же ведут себя личинки равноногих подотряда Gnathiidea, так называемые праницы (рис. 249, 1), которые сначала плавают, а потом паразитируют на рыбах, с тем отличием от Epicaridea, что взрослые ведут свободный, а не паразитический образ жизни. Наконец, известно несколько планктонных видов равноногих, которые плавают всю жизнь. Это преимущественно глубоководные животные, как например широко распространенная Munneurycope murrayi и Anuropus bathypelagicus, свойственный только северной половине Тихого океана. Idotea algirica живет на плавающих по поверхности моря саргассовых и других водорослях, кусках дерева и вместе с этими предметами пассивно переносится с места на место. Однако, как и другие виды этого рода, она хорошо плавает и сама, перебираясь с одного скопления «плавника» на другое.
Среди донных морских видов многие способны зарываться в грунт. Очень быстро зарываются, например, крупная Mesidotea (табл. 33, 5) и мелкие представители подотряда Asellota — Desmosoma, Macrostylisn др., имеющие специальные копательные грудные ножки. Cyathura carinata и, вероятно, другие виды подотряда Anthuridea делают в илистом грунте направленные вертикально глубокие ходы, которые они покидают только ночью. Если экспериментальный аквариум с начавшим строительство хода рачком повернуть на 90°, придав ходу горизонтальное направление, рачок продолжает рыть ход, также изгибая новый его отрезок под прямым углом к прежнему. Способность ориентироваться по отношению к силе земного притяжения объясняется присутствием у Cyathura в основании тельсона парного статоциста. Оперативное удаление статоциста нарушает способность рачка рыть ходы в определенном направлении. Самцы Gnathiidea своими жвалами вырывают ходы длиной по 15—20 см в илистых крутых склонах дна. В конце ход расширяется в камеру, в которой живет около десятка самок.
Некоторые пресноводные равноногие также могут глубоко зарываться в грунт и таким образом спасаться от пересыхания водоемов. Так поступает, например, обычный американский водяной ослик Asellus communis, а также представители подотряда Phreatoicidea, обитающие в болотах Южной Африки. В засушливыз периоды Phreatoicidea вырывают во влажном мхе ямки глубиной 2,5—5 см и массами сидят в них, прикрывая задними грудными ножками жабры и ожидая появления воды.
Однако наиболее совершенные норы строят приспособившиеся к жизни в пустынях мокрицы из рода Hemilepistus (табл. 33, 16). В пустыне Каракум норы Hemilepistus cristatus достигают глубины 60—100 см. Температура в них круглый год не падает ниже 10° и не поднимается выше 26°С, относительная влажность воздуха близка к 100%. На поверхности почвы днем температура достигает 60°С, а влажность уменьшается до 15%. Летом мокрицы покидают норы только утром и вечером, а зимой впадают в спячку и совсем не выходят из нор. Там же в конце мая самки рождают по 60—80 молодых рачков. Родители не выпускают их из норы и защищают их от врагов, закрывая вход в нору своими передними грудными сегментами. Для питания детей родители затаскивают в нору растительные остатки. Через 2,5 месяца молодь вырастает, но продолжает жить в норе до следующей весны, и только в марте рачки выходят на поверхность и приступают к строительству собственных жилищ. В старой норе остаются самец и самка, обзаводящиеся снова детьми. Таким образом, в каждой норе всегда живет одна семья и эти семьи не смешиваются между собой.
Питание растениями присуще очень многим равноногим, как морским, так и пресноводным и сухопутным. Виды рода Idotea питаются водорослями, на которых они живут, а также обрастающими их диатомовыми. Преимущественно водорослевыми и бактериальными обрастаниями питаются литоральные Jaera, Sphaeroma, Eurydice, Ligia и некоторые другие. Интересно, что изменчивая окраска широко распространенной Idotea viridis, обычной, в частности, у нас в Балтийском и Черном морях, зависит от ее пищи. Живущие на зостере и зеленых водорослях рачки — зеленые, на бурых — коричневые, на красных — красные. Пигмент растений окрашивает не только содержимое их кишечника, но также кровь и полостную жидкость, просвечивающие через покровы тела. Если рачков пересадить с одних растений на другие, их окраска через 10—15 дней соответственно изменится. Поедая растительную пищу, Idotea не брезгает и попадающимися ей среди обрастаний животными — гидроидами, мшанками, личинками хирономид и т. д.
Обычные обитатели пресных вод — водяные ослики — также типичные растительноядные организмы. Помимо водных растений, живых и отмирающих, они в большом количестве поедают листья деревьев, падающие в водоемы. За всю свою жизнь каждый водяной ослик потребляет 168 мг листьев (в сыром весе). В водоемах, в которые попадает много древесных листьев, водяные ослики обыкновенно живут в большом количестве. В пруду на территории парка Тимирязевской академии насчитывается 5000 рачков на 1 м2.
Африканские и австралийские Phreatoicidea едят растительные остатки и водяные мхи.
Почти все мокрицы питаются растениями, которые служат для них источником не только пищи, но и влаги, необходимой для увлажнения жабр. Поглощая растительный материал, массовые виды мокриц, например некоторые виды родов Oniscus и Porcellio, выступают в качестве важных факторов почвообразовательного процесса. Их экскременты частично непосредственно входят в состав почвенного слоя, частично подвергаются дальнейшей переработке ногохвостками, которые их поедают. Особенно важное значение имеет почвообразовательная деятельность мокриц рода Hemilepistus в некоторых пустынях, в частности в пустыне Каракум в Туркмении. При рытье нор они выносят на поверхность частички глубже лежащего грунта. Кроме того, они откладывают вокруг нор свои экскременты, которые вместе с частичками грунта образуют вокруг отверстия норы своеобразные кольцевые валики. На 1 га площади обычно приходится более 1 миллиона мокриц. В течение 3 месяцев своей ежегодной активной деятельности они дают около 0,5 т богатых водорастворимыми солями частичек почвы глубинных слоев и до 1 т экскрементов. На образованных ими плодородных выбросах хорошо растут растения. Кроме того, благодаря многочисленным (20 000 на 1 га) глубоким норам мокриц повышается скважность почвы и улучшаются условия существования почвенных бактерий. Все это приводит к значительному повышению плодородия почвы.
К питанию зелеными частями растений и их остатками близко использование в пищу древесины. Среди равноногих известны древоточцы, вредящие деревянным сооружениям в море и питающиеся древесиной. Это, во-первых, многочисленные виды рода Limnoria и, во-вторых, некоторые виды рода Sphaeroma. Виды Limnoria встречаются почти во всех морях. Только в наших дальневосточных и северных морях известно 4 вида, в Черном море — 2 вида и т. д. Некоторые из них живут на водорослях и питаются ими, но большинство поселяется на сваях пристаней, на стоящих у причалов деревянных судах и т. д. Острыми жвалами они жуют древесину, причем разрушают преимущественно более мягкие летние слои, не трогая зимних. Под поверхностью сваи на глубине 1—2 см они выгрызают ходы, превосходящие по диаметру ширину их тела, следуя по летним слоям нарастания дерева. Каждый такой ход одним или двумя отверстиями соединяется с наружной средой. В нем обычно обитает самка (длиной до 5 мм) и более мелкий самец, но точит дерево главным образом самка. При сильном повреждении на 1 см2 поверхности древесины насчитывается 12—15 отверстий, ведущих в ходы рачков. Источенная рачками поверхность сваи становится рыхлой, губчатой, иногда сваи истончаются и теряют первоначальную прочность. Их приходится заменять новыми.
Рачки заглатывают опилки, образующиеся при бурении древесины, и пропускают их через свой кишечник. При этом клетчатка переваривается, а лигнин выделяется с испражнениями. При прохождении через кишечник рачка древесные опилки теряют 50—95% своего исходного сухого веса.
В проточенных в дереве ходах происходит и размножение Limnoria. Летом каждая самка приносит 8—10, изредка до 30, осенью 1—4 молодых рачка. Вылезая из ходов, рачки при помощи брюшных ножек довольно быстро плавают спинной стороной вниз, но не удаляются дальше чем на 1 м от места своего рождения. Расселение лимнорий на далекие расстояния происходит пассивно, главным образом путем переноса течениями зараженных рачками бревен, стволов, кусков дерева и т. д.

Заглатывание грунта и использование содержащегося в нем органического вещества распространено среди морских равноногих не менее широко, чем питание растениями. Многие представители Asellota, как мелкие, так и крупные, а также многие Valvifera, в частности из семейства Arcturidae, питаются именно таким образом. В особенности характерно питание грунтом для глубоководных равноногих. По мере увеличения глубины количество растительноядных и хищных видов равноногих постепенно уменьшается, и, начиная с 3—4 км и кончая предельными океаническими глубинами порядка 11 км, все равноногие оказываются грунтоедами. При этом некоторые из них способны сортировать грунт, заглатывая преимущественно наиболее мелкие его частицы с несколько повышенным содержанием органического вещества. Приблизительно таким же образом питаются обитатели подземных пресных вод, в частности подземные водяные ослики и Cirolanidae.
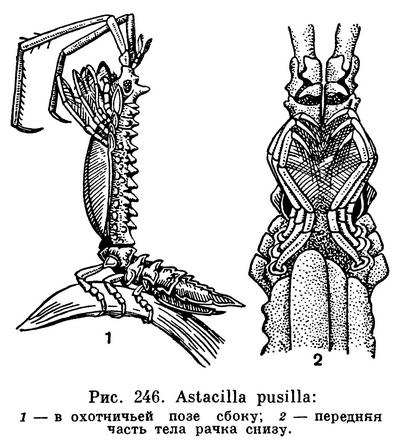
Немало среди морских равноногих и хищников. Mesidotea при помощи чувствительных щетинок и цилиндров своих задних антенн разыскивает зарывшихся в грунт бокоплавов, полихет, личинок хирономид и других мелких животных, хватает их подклешнями передних грудных ножек и поедает. Нередко она нападает на рыб, в особенности попавших в рыболовные сети, и местами, например в низовьях наших северных рек, приносит заметный ущерб промыслу, так как портит улов. По-видимому, она может иногда распространяться вверх по рекам, прицепившись к мигрирующим рыбам. Astacilla охотится на различных мелких животных следующим образом: она держится задними грудными ножками за водоросли, ветви гидроидов или мшанок и приподнимает вытянутую переднюю часть тела так, чтобы длинные, богатые крепкими щетинками передние грудные ножки и чувствительные задние антенны были направлены вперед; в таком положении она легко хватает передними ножками свои жертвы, прежде всего разных мелких рачков (рис. 246).
Многочисленные представители семейства Cirolanidae образуют ряд постепенных переходов от хищничества к паразитизму. Виды родов Cirolana и Conilera нападают на рыб, правда преимущественно больных или попавших в сети, и на недавно слинявших десятиногих раков. Особенно охотно они поедают трупы рыб, оставляя от них буквально только кожу да кости.
У этих родов ротовые придатки жующего типа. Виды рода Aega — временные паразиты рыб. Они быстро плавают, прочно прикрепляются тремя парами крючковидных передних грудных ног к коже рыб, там, где она тонкая (жабры, плавники, жаберные крышки), прокалывают ее колюще-сосущим ротовым аппаратом и сосут кровь рыб. Однако, наевшись, они оставляют своего хозяина. Наконец, виды подсемейства Cymothoinae (Cymothoa, Meinertia, Livoneca, Anilocra и др.) постоянно живут на рыбах, причем не только на жабрах, но и во рту, а также на поверхности тела. Они плотно прикрепляются к рыбе, благодаря тому что все (иногда за исключением последней пары) их грудные ножки крючковидны и своими концами внедряются в кожу хозяина. Иногда прикрепившиеся рачки обрастают кожей рыбы, образуя на ее поверхности опухоли. Самки Cymothoinae теряют двустороннюю симметрию, и часто тело их искривляется. Они паразитируют как на морских, так и на пресноводных рыбах. В бассейне Амура на теле многих рыб можно найти вздутия, в которых помещаются попарно крупная асимметричная самка и более мелкий симметричный самец Livoneca amurensis.
Помимо Cirolanidae, на рыбах паразитируют личинки (праницы) Gnathiidea (рис. 249, 2). Они подплывают к рыбам, прикрепляются к их плавникам крючковидными передними ногами и начинают разрывать ими ткани хозяина, погружая все глубже свой колюще-сосущий ротовой аппарат, пока не доберутся до кровеносных сосудов. При этом они удерживаются в ране жвалами, как гарпунами, а сосущий хоботок составляют челюсти. Они сосут кровь чрезвычайно жадно. Средний отдел их кишечника переполняется кровью, и задняя часть груди раздувается настолько, что сегментация ее исчезает. Впоследствии кровь переходит в пищеварительные железы. Ее запасов хватает на всю жизнь рачка. После линьки личинок возникшие из них взрослые рачки больше не питаются, их ротовые придатки, за исключением жвал самца, редуцируются, а кишечник у некоторых родов частично даже зарастает.
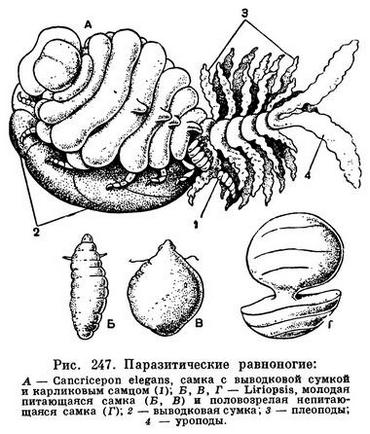
Наиболее глубокие приспособления к паразитическому существованию свойственны подотряду Epicaridea, все представители которого паразитируют на различных ракообразных. Их организация, в особенности самок, в различной степени упрощена вплоть до полной потери сегментации и конечностей и превращения тела в бесформенный мешок. Самки представителей семейств Bopyridae, Dajidae и Entoniscidae, поселяющиеся в жаберной полости креветок и крабов, а иногда (Dajidae) и мизид, в той или иной степени сегментированы, но обычно асимметричны, ротовые придатки их превращены в сосущий хоботок, которым они сосут кровь своих хозяев. Выводковая сумка очень крупная и содержит большое количество яиц. Самцы двустороннесимметричны, гораздо мельче самок и живут на них (рис. 247, А). Виды семейства Cryptoniscidae паразитируют на самых различных ракообразных — на ракушковых, усоногих, мизидах, кумовых, других равноногих, бокоплавах, клешненосных осликах. Половозрелые самки Cryptoniscidae имеют мешковидное тело, без всяких следов сегментации и конечностей, с хоботком на переднем конце, которым они протыкают покровы хозяина и сосут его кровь. Такие самки образуются из маленьких самцов, сохраняющих основные признаки равноногих ракообразных. Таким образом, Cryptoniscidae, в отличие от других раздельнополых Epicaridea, представляют собой протерандрических гермафродитов: при созревании они становятся самцами, а затем преобразуются в самок.
Среди Cryptoniscidae особенно интересны роды Dunalia и Liriopsis, паразитирующие на корнеголовых ракообразных Sacculina и Peltogaster, которые в свою очередь паразитируют на крабах и раках-отшельниках. Самка Dunalia помещается в жаберной полости пораженного саккулиной краба, но своим длинным хоботком через покровы краба достигает ответвлений «корней» саккулины и питается за ее счет. Срок продолжительности жизни Dunalia (2—3 месяца) значительно меньше, чем саккулины, так что наступает момент, когда паразит освобождается от своего сверхпаразита. Молодые самки Liriopsis овальные, прикрепляются к мешковидному телу Peltogaster и сосут его соки. Затем животное проникает в мантийную полость Peltogaster, округляется, тело его перешнуровывается на две части, оно прекращает питаться и достигает половозрелости (рис. 247, Б—Г).
Все Epicaridea имеют свободноплавающих личинок, при помощи которых они расселяются и заражают своих хозяев.
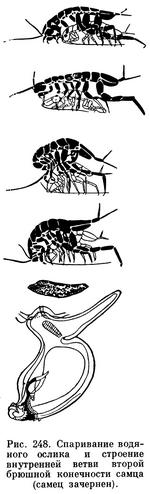
Размножение подавляющего большинства равноногих происходит так же, как у других перакарид. Однако, помимо раздельнополых видов, среди паразитических видов, как уже упоминалось, есть протерандрические гермафродиты (Cymothoinae, Cryptoniscidae), а для Cyathura carinata описана протогиния. Наконец, среди мокриц известно несколько видов, которым свойственно партеногенетическое размножение — явление исключительно редкое у высших ракообразных.
Половой диморфизм у раздельнополых видов, если исключить Epicaridea, проявляется главным образом в строении передних брюшных ножек, приспособленных у самцов для копуляции, и нередко в различиях размеров самцов и самок: у водяных осликов, например, самцы крупнее самок, но у некоторых других видов наблюдаются обратные соотношения. У Gnathiidea самцы имеют жвалы, отсутствующие у самок (табл. 33, 1,2). У ряда Sphaeromidae у самцов две передние пары грудных ножек снабжены подклешнями, а у самок только одна.
Половому процессу предшествует длительное спаривание. Самец удерживает самку грудными ножками, располагаясь на ее спинной стороне и ожидая ее линьки. У водяных осликов и, вероятно, у других видов перенос спермы возможен только в течение 10 часов после линьки самки, когда ее половые отверстия расширены и не образовались пластинки выводковой сумки. Линька всегда происходит в два приема: сначала старый панцирь сбрасывается с задней, а потом с передней части тела. В промежуток между этими двумя линьками самец своими копулятивными брюшными ножками переносит сперму в половые пути самки. Через 2—10 часов у самки образуется выводковая сумка и туда откладываются оплодотворенные в половых путях яйца. Число их различно у разных видов, причем по мере развития оно уменьшается, так как часть яиц вываливается из выводковой сумки и погибает. Так, например, у водяного ослика из 150 откладываемых сначала яиц к концу развития остается 80, у Idotea из 30—40 — только 4—6, у Jaera albifrons из 15—16 не больше 12, а обычно 3—4 яйца. Наибольшее количество яиц, как и у других животных, образуется у паразитических видов. В выводковой сумке Cymothoa oestrum насчитывается 2450 яиц. Наоборот, очень низкой плодовитостью отличаются подземные и глубоководные виды. Мелкие интерстициальные виды Microcharon носят в сумке всего 1—2 яйца. Из всех равноногих только некоторым Gnathiidea, в частности роду Paragnathia, свойственно живорождение. Из половых отверстий самок сразу выходят подвижные личинки на стадии манка, и выводковой сумки не образуется совсем. У всех остальных яйца развиваются в выводковой сумке. Движения ее пластинок приводят к тому, что вода все время омывает яйца и молодь. У мокриц полость выводковой сумки увлажняется за счет водопроводной системы покровов, но у некоторых видов на брюшной стороне тела яйценосных самок образуются пальцевидные выросты, выделяющие влагу в выводковую сумку. Из яиц в выводковой сумке выходят молодые рачки, находящиеся, так же как у кумовых и некоторых других отрядов, на стадии манка. Обычно эти молодые рачки отличаются от взрослых главным образом недоразвитием последней пары грудных ножек, но у Gnathiidea и Epicaridea стадия манка имеет ряд специальных приспособлений к активному существованию и поискам хозяина, на котором ей или последующим стадиям предстоит паразитировать.
О стадии манка Gnathiidea, носящей специальное название праница, уже говорилось выше. У Epicaridea эта стадия называется эпикаридиум. Эпикаридиум обычно достигает в длину 0,2—0,35 мм и активно плавает при помощи крупных задних антенн и брюшных ножек. Грудные ножки его крючковидные (рис. 249, 5). Он питается только за счет желтка, унаследованного от яйца. Судя по наблюдениям над Portunion (для других видов жизненный цикл не установлен), эпикаридиум прикрепляется к планктонным веслоногим и после линьки превращается в паразитическую стадию микронноциум, которая теряет членистые конечности и плавательные щетинки, но, слиняв несколько раз, дает начало новой свободноживущей стадии — криптонисциуму. Криптонисциум имеет нормальную сегментацию, 7 пар крючковидных грудных и 6 пар плавательных брюшных ножек, хорошо развитые антенны (рис. 249, 4). Он отыскивает раков, которые служат окончательными хозяевами данному виду паразита, прикрепляется к ним, линяет и начинает преобразовываться во взрослую стадию. У раздельнополых Epicaridea криптонисциум, прикрепившийся к еще не зараженному хозяину, превращается в самку. Если же личинка садится поблизости от уже имеющейся самки, она становится самцом, причем ее облик меняется незначительно: редуцируются только глаза, антенны, плавательные ножки, но характер сегментации и строение грудных ножек остаются прежними. У протерандрических гермафродитов Cryptoniscidae личинка криптонисциум, садясь на хозяина, без всякой линьки превращается в самца, так как в ней развиваются семенники. Затем в половых железах, помимо сперматозоидов, начинают созревать яйца, паразит несколько раз линяет, постепенно теряя при этом сегментацию, конечности и другие признаки ракообразных, и в конце концов приобретает мешковидную форму, характерную для половозрелой самки.
Партеногенетическое размножение известно у некоторых видов мокриц из рода Trichoniscus (Trichoniscus elisabethae, Trichoniscus provisorius) и также у обычных широко распространенных мокриц Armadillidium vulgare и Cylisticus convexus. Все эти виды распадаются на отдельные расы, различающиеся по числу хромосом. Расы с обычным диплоидным числом хромосом включают и самцов и самок, и у них размножению предшествует оплодотворение яиц. Наряду с ними существуют расы с триплоидным числом хромосом (у Trichoniscus диплоидное число хромосом 16, триплоидное 24), представленные исключительно самками, к которым иногда добавляются в ничтожном количестве (менее 1,6%) самцы. Яйца триплоидных самок при созревании не испытывают второго редукционного деления и оказываются способными к развитию без оплодотворения. Такие размножающиеся партеногенетически расы приурочены к сравнительно неблагоприятным условиям существования, в частности к засушливым местам, и обычно обитают близ границ области распространения каждого данного вида.
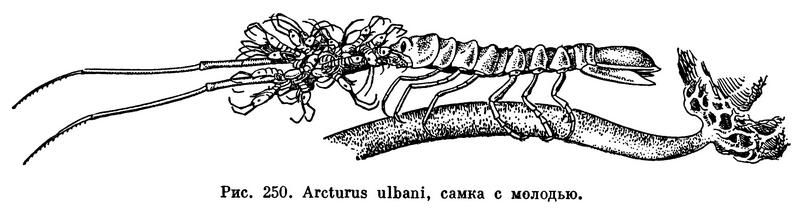
У свободноживущих равноногих, как водных, так и сухопутных, вышедшие из яиц и находящиеся на стадии манка молодые рачки некоторое время остаются в выводковой сумке матери и линяют там два или три раза. После этого, будучи еще на той же стадии, рачки покидают выводковую сумку матери. У морских видов из семейства Arcturidae молодь перебирается из выводковой сумки матери на ее длинные задние антенны и некоторое время живет на них (рис. 250). Позднее молодые рачки время от времени линяют и в конце концов достигают половозрелости. Промежутки между линьками и число их до достижения половозрелости различны у разных видов. Так, например, у мокрицы Porcellio эмбриональное развитие длится 35 дней, первая стадия манка продолжается 21 день, из которых 16 приходится на пребывание в выводковой сумке, вторая и третья — по 23 дня, а всего до возникновения вполне сформированного, но еще неполовозрелого рачка требуется 102 дня. У морского рачка Sphaeroma rugicauda при температуре 15°С яйца в выводковой сумке развиваются 45 дней, у обычного водяного ослика под Москвой — от 14 дней в мае до 22 дней в августе. Самки водяного ослика достигают половозрелости через 50—55 дней после рождения, перелиняв за это время 13—15 раз.
Общая продолжительность жизни установлена для немногих видов. Мокрица Oniscus asellus живет 3—4 года, Aega — не менее 2,25 лет, водяной ослик — около 2 лет, Cryptoniscidae — менее полугода.
Как уже указывалось, большинство известных видов равноногих ракообразных обитает в море, где они встречаются от приливо-отливной зоны до предельных океанических глубин. Литораль населена небольшим количеством видов, из которых наиболее характерны представители родов Jaera и Sphaeroma. Jaera albifrons и Jaera marina широко распространены в северной части Атлантического океана, а первая из них обычна также в Баренцевом и Белом морях. Окраска этих рачков очень изменчива: некоторые особи целиком темные, у других вдоль тела проходит темная полоса, у третьих темная голова или плеотельсон и т. д. (рис. 251). Соотношение особей с разным типом окраски в различных поселениях рачков различно. Специальные исследования по скрещиванию между собой различно окрашенных особей показали, что каждый тип окраски зависит от определенного гена. Одни из них доминантны, другие рецессивны. Есть гены некоторых типов окраски, сцепленные с генами пола. Это значит, что определенный тип окраски встречается только у самцов или, наоборот, только у самок. Некоторые поселения рачков обеднены генами по сравнению с другими, и соответственно разнообразие окраски составляющих их особей меньше. Таким образом, окраска Jaera определяется совершенно иначе, чем у Idotea.
На морских мелководьях, глубже приливо-отливной зоны, богатство фауны равноногих возрастает. Среди водорослей, в зарослях гидроидов и мшанок преобладают виды с узким телом, позволяющим рачкам пролезать между ветвями, например различные виды Idotea, Arcturidae и др. На мягких илистых грунтах обитают уплощенные сверху вниз равноногие вроде Mesidotea, Synidotea, Serolidae и др. Зарывающиеся в мягкий грунт виды часто имеют удлиненное цилиндрическое тело (Anthuridea, Gnathiidea, Ischnomesidae и др.). На больших океанических глубинах обитают почти исключительно представители подотряда Asellota. Этот подотряд явно тяготеет к холодным водам. Его виды многочисленны на мелководьях в высоких и умеренных широтах северного и южного полушарий, а также на больших глубинах всего Мирового океана, но почти не встречаются на тропических мелководьях.
Для многих глубоководных Asellota характерны удлиненные грудные ножки, препятствующие погружению их тела в мягкий грунт (табл. 33, 8,11,12), а также антенны, богатые чувствительными щетинками и цилиндрами. Однако особенно интересна четко выраженная у них тенденция к увеличению размеров тела по мере увеличения глубины. Чем глубже обитает какой-нибудь вид определенного рода, тем крупнее его размеры по сравнению с близким видом с меньшей глубины. Так, например, длина тела видов рода Haplomesus с глубины менее 3 км не достигает 5 мм, а Haplomesus gigas с глубины 8330—8430 м имеет в длину 15 мм.
Специальные исследования показали, что увеличение тела глубоководных равноногих происходит главным образом за счет увеличения размеров составляющих их организм клеток, а не за счет возрастания их количества. Это явление не связано с понижением температуры среды обитания. Предполагают, что в качестве основного фактора, вызывающего «глубоководный гигантизм», выступает повышенное на больших глубинах гидростатическое давление, действующее на обмен веществ.
Среди морских мелководных равноногих многие способны переносить значительные колебания солености. Jaera albifrons и Cyathura carinata (табл. 33, 7) часто встречаются в опресненных участках моря и даже заходят в пресные воды. Mesidotea entomon (табл. 33, 5) обитает преимущественно в солоноватых или даже в пресных водах, причем при большом опреснении она достигает более крупных размеров. В западной части Балтийского моря наибольшая длина самцов 50 мм, а в опресненной восточной — более 80 мм. Этот вид населяет Средний и Южный Каспий и некоторые озера, покрывавшиеся во время великого оледенения морем.
Фауна пресноводных равноногих значительно беднее морской. Наиболее широко распространено и обильно видами чисто пресноводное семейство водяных осликов (Asellidae). Обыкновенный водяной ослик (Asellus aquaticus, табл. 33, 3) обитает во множестве стоячих водоемов всей Европы, исключая Пиренейский полуостров. На Кавказе, кроме него, известен близкий вид Asellus monticola. На значительном протяжении Азии водяные ослики отсутствуют. Только в Байкале живет пять присущих лишь этому озеру видов. Кроме того, по одному эндемичному виду есть в Ципоципиканских озерах (бассейн Витима), в бассейне Иртыша и в бассейне Уссури. Местонахождения этих видов могут считаться остатками некогда более широкого распространения рода Asellus. Далее, вдоль восточного берега Азии, включая северную Японию и северный Китай, широко распространен близкий к Asellus aquaticus вид водяного ослика — Asellus hilgendorfi. В Западной Европе наряду с Asellus aquaticus встречаются и некоторые другие виды водяных осликов, в частности Asellus meridianus, Asellus banyulensis и т. д. Как правило, Asellus aquaticus и Asellus meridianus, обладающие чрезвычайно сходной биологией, тем не менее не живут вместе в одном водоеме. Между ними существуют антагонистические отношения. Asellus aquaticus постепенно вытесняет Asellus meridianus, но механизм этого вытеснения, так же как и в других подобных случаях, разгадать пока не удалось. Есть особые виды водяных осликов в наземных водоемах Северной Америки и Северной Африки.
Много видов водяных осликов приспособилось к жизни в подземных пресных водах. Такие виды известны из Закавказья, Европы (особенно много их на Балканском полуострове), бассейна Амура и Японии. Только в подземных водах обитают особые роды этого семейства — Stenasellus, Stygasellus и Johanella. Несколько видов Stygasellus населяет только Португалию, Johanella найдена лишь в одном ключе в Северной Африке, a Stenasellus имеет очень широкое, но разорванное распространение. Наибольшее количество видов этого рода приурочено к пещерам стран, лежащих к северу от Средиземного моря (южная Франция, Италия, Пиренейский полуостров, Балканы), 4 вида открыто в подземных водах экваториальной Африки, а один вид обитает в теплом источнике Ходжа-Койнар, температура воды которого круглый год равняется 20—22°С и который находится в горной системе Кугитангтау, в восточной Туркмении. По всей вероятности, Stenasellus — очень древний род тропического происхождения, сохранившийся только в подземных водах. Ему свойственны многие чрезвычайно примитивные морфологические признаки (чешуевидная наружная ветвь на задних антеннах, сравнительно крупные передние брюшные сегменты и т. д.).
Помимо водяных осликов, в пещерах Европы обитают своеобразные роды Typhlocirolana, Caecosphaeroma, Cirolanides, Typhlosphaeroma, принадлежащие к морским семействам Sphaeromidae и Cirolanidae. Вместе с некоторыми другими животными они образуют группу видов морского происхождения в составе пресноводной подземной фауны.
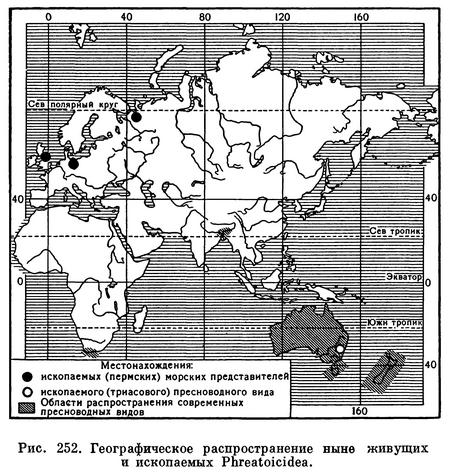
Морское происхождение свойственно также чрезвычайно мелким равноногим, обитающим в интерстициальных пресных и морских водах. Как и другие представители интерстициальной фауны, они имеют длинное червеобразное тело и укороченные конечности, что позволяет им активно передвигаться по узким пространствам между песчинками и другими частицами грунта. Род Microcharon (табл. 33, 13) объединяет 16 видов и 5 подвидов, большинство которых обитает в пресных грунтовых водах бассейна Средиземного моря, но один вид известен из Закавказья, а два из Средней Азии, причем Microcharon halophilus, известный из пещеры Каптархана в восточной Туркмении, живет в соленом подземном озере, соленость воды которого равняется 12‰. Кроме этих обитателей внутренних водоемов, к роду Microcharon принадлежат еще два вида из интерстициальных морских вод Ламанша. Нет особых сомнений в том, что виды этого рода переселились из системы морских в систему пресных интерстициальных вод. Сходная история расселения приписывается и интерстициальному семейству Microcerberidae, принадлежащему к подотряду Anthuridea. Только к пресным водам приурочен ныне подотряд Phreatoicidea. Относящиеся к нему 47 видов распространены в Австралии, Тасмании, Новой Зеландии, Южной Африке и Индии как в наземных, так и в подземных водах. Однако в ископаемом состоянии они известны из морских отложений пермского периода в Архангельской области, Англии и Германии. Очевидно, в конце палеозоя они жили в море и населяли северное полушарие, а до наших дней сохранились только в пресных водах некоторых стран южного полушария (рис. 252).
К жизни на суше приспособился только один подотряд равноногих — Oniscoidea, или мокрицы. Они обитают во всех ландшафтных зонах земли, но особенно многочисленны и разнообразны в степях. Наиболее примитивные из них — представители рода Ligia — еще связаны с морем. Ligia живет на морских берегах и может длительное время существовать под водой. В условиях эксперимента эти мокрицы оставались под водой более 80 дней, причем нормально питались в таких условиях. Так же тесно связаны, но с пресными водами виды рода Ligidium, часто находимые в ручьях и ключах. Для всего семейства Ligiidae, к которому принадлежат оба названных рода, характерно отсутствие приспособлений к дыханию атмосферным воздухом. Все остальные мокрицы существуют совершенно независимо от водоемов и представляют собой настоящих сухопутных животных. Однако они могут жить только в условиях повышенной влажности, обеспечивающей их нормальное дыхание и размножение. Поэтому они выработали ряд приспособлений, благодаря которым они заселили даже такие, казалось бы, не подходящие для них местообитания, как пустыни.
Эти приспособления направлены главным образом на то, чтобы свести к минимуму потерю влаги организмом, что достигается разными способами. Все мокрицы активны только ночью, что защищает их от действия солнечных лучей. Светлое время суток они проводят зарывшись в поверхностный слой почвы, спрятавшись под камнями, в различных других укрытиях или в норах, которые выкапывают и в которых поддерживается повышенная влажность. Есть мокрицы, живущие в норах грызунов и муравейниках. Ночью они поедают растения, получая не только пищу, но и влагу. В условиях эксперимента, да и в природе они отрицательно фототропичны: активно стремятся уйти от света, чтобы избежать высыхания. Для большинства мокриц смертельно пребывание в условиях влажности воздуха менее 86% насыщения, причем Oniscus с менее совершенными ложными трахеями погибает раньше, чем Porcellio, ложные трахеи которого более сложны.
При помещении мокриц в условия пониженной влажности они сначала очень быстро теряют воду, но затем этот процесс замедляется, благодаря тому что из крови выделяются некоторые вещества, увеличивающие непроницаемость покровов. С другой стороны, испарение влаги позволяет мокрицам в известных пределах регулировать температуру своего тела, избегать перегрева. Разница между температурой нагретого солнцем до 36°С пляжа и температурой тела Ligia равняется 8°, для Oniscus и Armadillidium эта разница составляет 4—5°, для Porcellio — 2—3°.
Способность регулировать температуру своего тела свойственна только живым мокрицам. Температура тела мертвых животных совпадает с температурой субстрата. Все эти морфологические и физиологические особенности позволяют утверждать, что из всех ракообразных мокрицы наиболее полно приспособились к жизни на суше.
Практическое значение равноногих, несмотря на их широкое распространение и видовое разнообразие, довольно ограниченно. С одной стороны, они приносят некоторый вред человеку, поскольку Limnoria u Sphaeroma разрушают деревянные сооружения в море, а мокрица Hemilepistus zachvatkini объедает луговую растительность в Узбекистане. Но, с другой стороны, мокрицы, перерабатывая значительное количество растений, способствуют увеличению плодородия почвы. В особенности важную роль в этом деле играют виды рода Hemilepistus в пустынях, где они выполняют такую же огромную работу, как земляные черви в других климатических зонах.