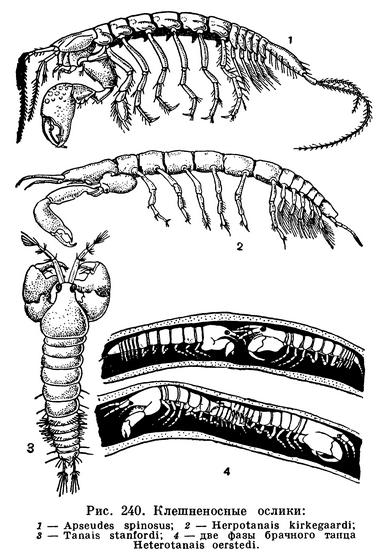
(Tanaidacea) Отряд Танаидацеи, Танаидовые ракообразные, Клешненосные ослики, Order Tanaidacea 24 семейства
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
НАДОТРЯД ПЕРАКАРИДЫ (PERACARIDA)
Этот обширный надотряд объединяет очень различных как по внешнему виду, так и по образу жизни ракообразных, обитающих в море, в пресных водах и на суше, ползающих по дну и плавающих в толще воды, большей частью свободноживущих, но частично и паразитирующих на других животных. Однако все они обладают некоторыми общими признаками, указывающими на их родство между собой.
Голова всех перакарид срастается с одним или изредка с двумя передними грудными сегментами, конечности которых превращаются в ногочелюсти. Правая и левая жвалы устроены различно: на левой обычно есть подвижная пластинка, отсутствующая на правой. Самки вынашивают яйца под грудью в выводковой сумке, образованной направленными внутрь листовидными отростками грудных ножек (только у отряда Thermosbaenacea выводковая сумка помещается на спинной стороне тела, под карапаксом). Из яиц выходят почти вполне сформировавшиеся молодые рачки, так что настоящих личинок у перакарид нет.
К надотряду Peracarida относятся отряды мизид, кумовых, клешненосных осликов, спелеогрифовых, термосбеновых, равноногих и разноногих ракообразных.
ОТРЯД КЛЕШНЕНОСНЫЕ ОСЛИКИ (TANAIDACEA)
При сборах донных морских животных наряду с кумовым, бокоплавами, равноногими изредка попадаются мелкие высшие ракообразные, принадлежащие к отряду клешненосных осликов, или танаидовых. Они ведут скрытый образ жизни, укрываясь в построенных ими трубках, зарываясь в грунт или прячась среди водорослей, камней и т. д. Клешненосные ослики встречаются как на мелководье, так и на больших океанических глубинах, но нигде не образуют массовых скоплений. На Курильских островах и в Южной Америке один или два вида обитают и в пресных водах, но, как правило, клешненосные ослики не переносят опреснения и могут считаться настоящими морскими животными. Длина их тела составляет обычно от 2 до 5 мм, и только некоторые глубоководные виды достигают значительно более крупных размеров (Herpotanais kirkegaardi до 26 мм). Из-за сравнительной редкости и немногочисленности этих рачков их биология мало изучена. Описано около 250 видов клешненосных осликов.
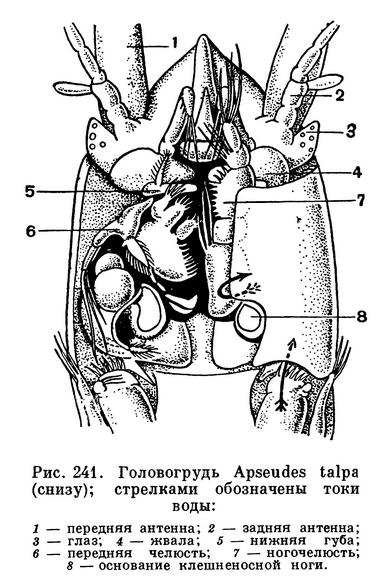
Тело этих ракообразных удлиненное, цилиндрическое или уплощенное в спинно-брюшном направлении. Короткий щитовидный карапакс покрывает голову и слившиеся с нею два передних грудных сегмента. Между его боковыми краями и стенкой тела, так же как у кумовых, с каждой стороны находится дыхательная полость. Передний край карапакса часто образует плоский треугольный или полукруглый рострум. У многих мелководных видов по бокам от основания рострума на специальных выростах головы располагаются маленькие глаза с очень небольшим количеством фасеток (у Heterotanais — 12, у Apseudes — 8). Глазные выросты Tanaidaсеа существенно отличаются от глазных стебельков тонкопанцирных, мизид и десятиногих, поскольку они не сочленены с головой и не могут поворачиваться.
Обе пары антенн обычно короткие. У представителей подотряда Monokonophora передние антенны с двумя жгутами, а задние с чешуевидной наружной ветвью, у представителей другого подотряда — Dikonophora — на передних антеннах всего один жгут, а задние лишены наружной ветви. Ротовые придатки хорошо развиты только у самок, а у самцов многих видов они частично или даже полностью редуцируются. Жвалы у Monokonophora несут щупик, отсутствующий на жвалах Dikonophora. Челюсти снабжены внутренними лопастями, многочисленные щетинки которых образуют фильтрующий аппарат. На передней ногочелюсти, так же как у кумовых, имеется крупный эпиподит, приспособленный для возбуждения тока воды в дыхательной полости, под боковым краем карапакса. Вода заходит под край карапакса сзади, омывает дыхательную полость и выходит позади места прикрепления передней ногочелюсти. Кислород усваивается внутренней поверхностью боковых краев карапакса, а также стенкой тела, прилегающей к дыхательной полости. Здесь располагается густая сеть капиллярных кровеносных лакун, впадающих близ заднего края карапакса в околосердечную сумку.
Конечности второго, слившегося с головой грудного сегмента крупнее остальных и снабжены на конце настоящей клешней или у некоторых видов подклешней. Благодаря характерному строению этих ножек клешненосные ослики и получили свое название. У самцов клешненосные ноги обычно бывают крупнее, чем у самок и нередко имеют несколько иное строение, но биологическое значение этих различий пока остается неизвестным. Остальные 6 пар грудных ножек устроены одинаково и представляют собой ходильные конечности. Все они одноветвисты и только у некоторых Apseudidae и Kalliapseudidae на передней из них, а также на клешненосных ногах есть короткие наружные ветви, напоминающие соответствующие придатки мизид и кумовых. Кроме того, у рода Apseudes ноги переднего свободного грудного сегмента отличаются от остальных тем, что их последние членики расширены и приспособлены для рытья грунта. У самок перед размножением на четырех, а у некоторых видов всего на одной паре грудных ножек развиваются пластинки, образующие выводковую сумку. Так же как у кумовых, они исчезают после линьки, следующей за окончанием вынашивания яиц и молоди, а затем, перед новым периодом размножения, возникают вновь.
Брюшной отдел всегда значительно короче и обычно несколько уже грудного. Пять передних составляющих его сегментов свободные, а последний, шестой сегмент срастается с тельсоном, образуя так называемый плеотельсон. Конечности передних пяти брюшных сегментов короткие, но двуветвистые и снабжены по краям многочисленными щетинками. Они служат плавательными ножками и обычно у самцов развиты сильнее, чем у самок. Самки некоторых видов совсем лишены этих ножек. К плеотельсону прикрепляется последняя пара брюшных ножек — уроподы. Они обычно двуветвисты, причем и ствол и обе ветви тонкие, палочковидные. У некоторых клешненосных осликов, например у Apseudes, ветви уроподов удлиненные и расчлененные.
Большинство клешненосных осликов живет в выкапываемых ими норах или в трубках. Внутреннюю поверхность норы рачки обкладывают песчинками, остатками растительности и другими частицами, которые они собирают и перетаскивают клешненосными ногами. Весь этот материал скрепляется специальным секретом. На боках трех передних свободных грудных сегментов располагаются железы, вырабатывающие этот секрет. Протоки желез очень длинные; они тянутся через каждую из трех пар передних ходильных ножек и открываются в их когтевых члениках. Этими ходильными ножками рачки и скрепляют обкладку своих нор и стенки своих трубок. Сидя в норе или в трубке, рачки энергично машут брюшными ножками и таким образом обновляют воду в своем убежище.
Время от времени они вылезают из нор и трубок и передвигаются по дну, частично ползая при помощи грудных, частично плавая над дном при помощи брюшных ножек. Кроме того, они могут двигаться скачками, сгибая и разгибая тело.
Пищей клешненосным осликам служат находящиеся в грунте остатки отмерших растений и другие органические частицы. Некоторые виды отфильтровывают их щетинками челюстей, используя ток воды, направляемый в дыхательные полости.
Самцы клешненосных осликов отличаются от самок большим количеством чувствительных щетинок на жгутах антенн и сильнее развитыми плавательными ножками, что позволяет приписывать им активную роль в поисках самок. Наряду с этим самцы многих видов частично или полностью лишены ротовых придатков, следовательно, не могут питаться и, очевидно, недолговечны.
Механизм определения пола у клешненосных осликов очень интересен. Еще недавно предполагалось, что некоторым из них свойственна протогиния — явление, довольно редкое в животном мире. При протогинии созревающий организм превращается в самку, которая затем переходит в самца.
Новые наблюдения Рамиреца над Heterotanais oerstedi позволяют считать по крайней мере этот вид потенциальным гермафродитом. У него самцы могут возникать либо непосредственно из молодых особей, либо из половозрелых самок, закончивших вынашивание молоди или даже еще не отметавших яиц. Это зависит от условий существования рачков. При совместном содержании самца и самки в одном сосуде перемены пола не происходит. При содержании молодой особи вместе с самцом из нее получается самка и, наоборот, если ее подсадить к самке, то — самец. Содержание молодой особи в одиночестве приводит к превращению ее в самку. Таким образом, пол у Heterotanais oerstedi не связан с присутствием определенного гена.
Гермафродитные особи известны не только для Heterotanais oerstedi, но также для крупного (15 мм) антарктического Apseudes spectabilis, Tanais stanfordi и глубоководного Neotanais serratispinosus. При этом у широко распространенного Tanais stanfordi (рис. 240, 3) гермафродитизм наблюдается только в отдельных частях его обширного ареала, в частности на Курильских островах, а в других местах этот вид представлен нормальными, не переходящими друг в друга самцами и самками. Замечательно, что все известные до сих пор гермафродитные танаидовые живут при низкой температуре.
Размножение клешненосных осликов удалось изучить лишь недавно, сконструировав для наблюдений специальные микроаквариумы. Самец Heterotanais oerstedi забирается в трубку самки, и в течение 11 часов рачки исполняют там брачный танец. Сначала самец преследует самку, затем она переворачивается через голову, и оба партнера оказываются «лицом к лицу», соприкасаясь антеннами (рис. 240, 4). После этого они поворачиваются друг к другу задними концами тела и контактируются уроподами, для чего самцу приходится предварительно переворачиваться через голову. Наконец, самец оказывается лежащим на спине, а самка располагается над ним. В таком положении (в течение 3 секунд) происходит копуляция, при которой сперматозоиды попадают в выводковую сумку самки. Вслед за этим самец покидает трубку самки, а она закрывает вход в трубку. Через некоторое время самка откладывает в выводковую сумку яйца.
Число откладываемых самкой в выводковую сумку яиц незначительно: у мелких видов обычно от 3 до 8, у крупного Apseudes antarcticus — 16. Яйца и молодь все время омываются током воды, вызываемым движениями составляющих сумку пластинок. Молодые рачки, оставаясь в сумке, линяют 2 раза и выходят из нее на стадии манка. У Heterotanais oerstedi при температуре 17°С это происходит через 18—19 дней, а при температуре 20°С — через 14 дней после откладки яиц. Первая свободноживущая стадия манка достигает в длину 0,6 мм, в то время как длина ее матери около 2 мм. Через 10—11 дней наступает первая на свободе линька молодого рачка. Вскоре после освобождения от потомства самка также линяет и теряет выводковую сумку, с тем чтобы приобрести ее вновь после 3 линек, последняя из которых предшествует следующему периоду размножения.
Дальнейшее развитие молоди прослежено у антарктического вида Heterotanais antarcticus. После выхода из выводковой сумки и до возникновения у самок выводковой сумки рачки линяют 7 раз, причем первые 4 стадии развития должны считаться стадией манка, поскольку у них отсутствует или недоразвита последняя пара грудных ножек. Как уже указывалось, эта стадия, помимо клешненосных осликов, свойственна только кумовым и равноногим ракообразным.
Распространение клешненосных осликов изучено далеко не достаточно. Наибольшее количество их видов приурочено к небольшим глубинам полносоленых морей умеренной зоны и высоких широт. Можно отметить четкие различия в составе фауны этих ракообразных на разных по характеру грунтах — песчаных, илистых и каменистых. На больших и предельных океанических глубинах обитают преимущественно представители семейства Neotanaidae, характеризующегося многими примитивными признаками.