(Harpacticoida) Отряд Гарпактициды, Order Harpacticoida 45 семейств
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ОТРЯД ВЕСЛОНОГИЕ РАКООБРАЗНЫЕ (COPEPODA)
Дыхание веслоногих осуществляется всей поверхностью тела, жабры отсутствуют. С этим, возможно, связано и слабое развитие или даже отсутствие кровеносной системы. Сердце есть только у представителей подотряда Calanoida, да и у них оно невелико, хотя и бьется очень часто: например, у морского рачка Labidocera оно совершает более 150 ударов в минуту. У остальных веслоногих полостная жидкость приводится в движение сокращениями кишечника.
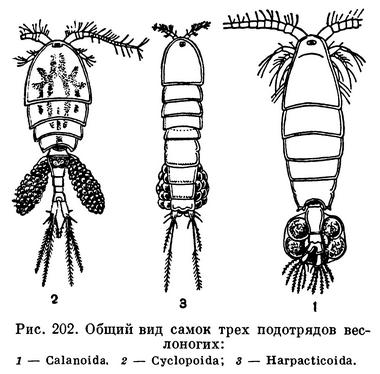
В пределах отряда веслоногих принято различать 5 подотрядов, из которых 2 объединяют исключительно паразитические виды. (Некоторые исследователи разделяют паразитических веслоногих на 5 или 6 подотрядов.) Свободноживущие веслоногие относятся к подотрядам Calanoida, Cyclopoida и Harpacticoida (рис. 202).
Calanoida — исключительно планктонные животные. Их голова и грудь значительно длиннее узкого брюшка, передние антенны очень длинные, превосходят голову и грудь, а нередко и все тело рачка, если есть яйцевой мешок, то один.
Cyclopoida обитают преимущественно на дне или в придонных слоях воды. Среди них есть и паразиты. Их голова и грудь относительно всего тела гораздо короче, чем у Calanoida, передние антенны короткие, не достигают и середины головогрудного отдела, брюшко уже груди. Самки всегда носят два яйцевых мешка.
Harpacticoida, за единичными исключениями, живут на дне и больше ползают, чем плавают. Тело их червеобразно благодаря тому, что брюшной отдел почти не отличается по ширине от грудного. Передние антенны очень короткие, самки большинства видов образуют один яйцевой мешок.
Представители всех трех подотрядов населяют как моря, так и пресные воды.
ПОДОТРЯД ГАРПАКТИЦИДЫ (HARPACTICOIDA)
Об образе жизни представителей третьего подотряда — Harpacticoida — известно гораздо меньше. Эти червеобразные, большей частью очень мелкие рачки, чрезвычайно разнообразны как в морских, так и в пресных водах, но никогда не встречаются в массовом количестве. Описано более 30 семейств и несколько сотен видов Harpacticoida.
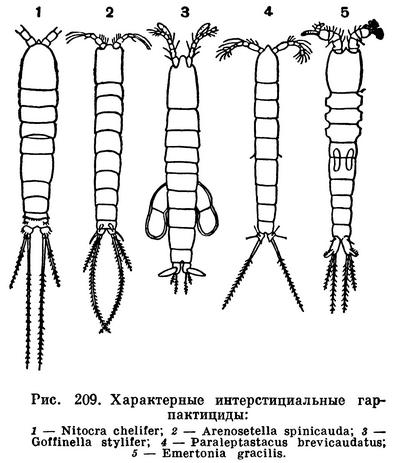
Большинство гарпактицид ползает по дну и донным растениям. Лишь единичные виды способны продолжительное время плавать и входят в состав морского планктона (Microsetella). Гораздо более характерны целые группы родов и видов гарпактицид, приспособившиеся к обитанию в особых, необычных условиях, в частности в капиллярных ходах между песчинками морских пляжей и в подземных пресных водах.
Всего несколько лет тому назад зоологи применили очень простой прием, позволяющий изучать население капиллярных ходов морского песка. На пляже, выше уровня моря, выкапывается яма. В ней постепенно накапливается вода, заполнявшая капилляры песка. Эту воду фильтруют через планктонную сеть и таким образом добывают представителей своеобразной фауны, получившей название интерстициальной.
Гарпактициды составляют заметную часть интерстициальной фауны. Их обнаружили всюду, где проводились соответствующие исследования, — на пляжах Англии, вдоль европейского и американского берегов Атлантического океана, на Средиземном и Черном морях, у берегов Африки и Индии, на островах Мадагаскар, Реюньон и Багамских. Большинство интерстициальных гарпактицид принадлежит к особым, обитающим только в таких условиях родам, отличающимся необыкновенно тонким и длинным телом (рис. 209), позволяющим рачкам двигаться в узких капиллярных ходах. Замечательно, что некоторые из этих специализированных видов были найдены в очень отдаленных друг от друга местах. Так, например, на Багамских островах оказались Arenosetella palpilabra, ранее известная только из Шотландии, и Horsiella trisaetosa, ранее известная только из Кильской бухты. Трудно объяснить подобное распространение, поскольку покоящихся яиц у интерстициальных гарпактицид нет.
Гарпактициды пресных подземных вод также представлены рядом специализированных родов — Parastenocaris, Elaphoidella, Ceuthonectes и других, частично очень широко распространенных, частично имеющих узкие и разорванные ареалы. Так, например, два вида рода Ceuthonectes обитают только в пещерах Закавказья, Югославии, Румынии, Италии и Южной Франции. Эти удаленные друг от друга местонахождения считаются остатками некогда гораздо более обширной области распространения древнего рода. В некоторых случаях можно допускать тропическое происхождение подземных гарпактицид Европы. Среди многочисленных видов рода Elaphoidella есть как тропические, так и европейские виды. Первые обитают в наземных, вторые (за единичными исключениями) в подземных водах. По всей вероятности, под землей сохранились остатки древней тропической фауны, погибшей на поверхности земли под влиянием климатических изменений. В тропических наземных водоемах некоторые гарпактициды приспособлены к условиям существования, напоминающим условия жизни в подземных водах. Известны тропические виды Elaphoidella, живущие в своеобразных микроводоемах, образующихся в пазухах листьев водных растений из семейства Bromeliaceae. В таких же условиях живет тропическая Viguierella coeca, обнаруженная на этих растениях в ботанических садах почти всех стран.
Чрезвычайно богата видами гарпактицид своеобразная фауна Байкала. Она состоит из 43 видов, из которых 38 эндемичны для этого озера. Особенно много этих рачков в прибрежной части Байкала, на камнях и водных растениях, а также на растущих здесь губках. По-видимому, они питаются за счет губок и в свою очередь становятся жертвами также ползающего по губкам бокоплава Brandtia parasitica.
Некоторые виды гарпактицид приурочены только к водоемам, очень бедным солями, характеризующимся повышенной кислотностью, т. е. связанным с верховыми, сфагновыми, болотами. Таков, например, Arcticocamptus arcticus, биология которого подробно изучена Е. В. Боруцким.
Arcticocamptus arcticus широко распространен на севере Европы, от Большеземельской тундры до Скандинавии, на западном побережье Гренландии, на Новой Земле. Кроме того, он найден в Альпах и в нескольких пунктах средней полосы Европейской части СССР, в том числе в Косине под Москвой, в Звенигороде, под Ярославлем и т. д. Повсюду он обитает в водоемах, связанных со сфагновыми болотами.
Из многочисленных водоемов, расположенных близ Косина, рачок живет только в двух болотах и в Святом озере, лежащем среди сфагнового торфяника. Очевидно, условия существования в других разнообразных соседних водоемах неблагоприятны для Arcticocamptus arcticus. При этом даже в немногих населенных им водоемах рачок в активном состоянии существует только в течение 1,5—2 месяцев весной, остальную часть года, т. е. 10—10,5 месяцев, он проводит в стадии покоящегося яйца.
Жизненный цикл Arcticocamptus arcticus тесно связан с изменениями растительного покрова болота. Е. В. Боруцкий пишет: «Как только с первыми весенними теплыми лучами рыхлый снег начинает таять и на поверхности болота образуются лужи — все животные, проведшие суровую зиму в тех или других стадиях в ледяном заключении, начинают реагировать на первые весенние лучи. Arcticocamptus arcticus одним из первых выходит из состояния анабиоза и появляется в водоеме. Уже в небольших лужах, еще среди снега, там, где поверхностные слои сфагнума оттаяли, можно найти его личинок, медленно и неуклюже передвигающихся среди листочков мха в поисках пищи. Личинки — на первой науплиальной стадии и, очевидно, только что вылупившиеся из яйца. С каждым днем науплиус крепнет, движения его становятся более уверенными и быстрыми, наконец, наступает момент первой линьки — он меняет старую узкую оболочку на новую, более просторную. За первой линькой следует вторая, третья и т. д., и вот спустя две-три недели мы уже встречаем взрослые экземпляры или личинок на последней копеподитной стадии. Но они уже не пользуются тем простором, который имели в своем раннем личиночном состоянии: вместо обширных луж, полных воды, в которых они свободно переплывали с одного сфагнового кустика на другой, теперь только влажный мох и незначительное количество воды. Вместо жалких голых ветвей теперь нежные розовые цветы кассандры и клюквы, белые чашечки андромеды и шапки цветущего богульника. Болото изменилось — яркий зеленый сфагновый ковер пестрит розовыми и белыми пятнами цветов. И это изменение картины болота как нельзя лучше совпадает с определенным моментом в биологии Arcticocamptus arcticus, именно с периодом копуляции. В продолжение нескольких дней мы встречаем почти исключительно копулирующие пары. Но эти цветы — роковые для Arcticocamptus arcticus: с постепенным увяданием их наблюдается постепенное уменьшение численности рачков, копуляция встречается все реже и реже, чаще попадаются самки с яйцевыми мешками и, наконец, к середине или к концу июня Arcticocamptus arcticus совсем исчезает из водоема, и лишь только запоздалые экземпляры встречаются в незначительном количестве в июле или в начале августа».
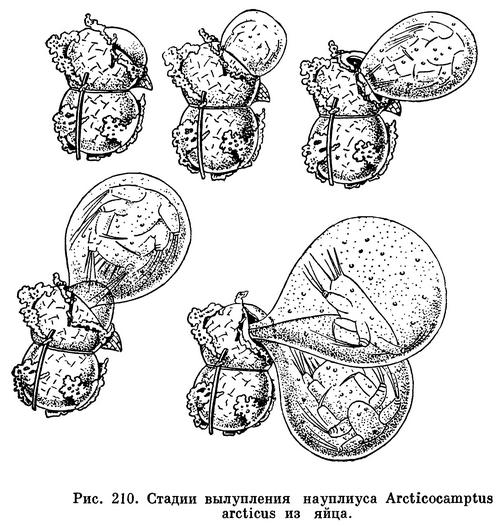
Рачок оставляет в водоеме свои яйцевые мешки, имеющие форму двух соединенных вместе шаров, покрытых общей «мешковой» оболочкой, осуществляющей механическую защиту яиц, а также защищающей их от высыхания. Кроме того, каждое яйцо имеет свою более тонкую прозрачную оболочку. Она непроницаема как для солей, так и для воды. К осени в каждом яйце формируется науплиус, причем науплиусы двух соединенных яиц всегда направлены своими передними концами в протлвоположные стороны. Науплиусы одеты еще одной очень тонкой и эластичной внутренней оболочкой, снабженной различными тяжами и нитями. Эта оболочка проницаема для воды, но не для солей.
Когда наступает время вылупления науплиусов, т. е. в период весеннего таяния льда, в мешковой оболочке с одной стороны образуется трещина, через которую начинает выпячиваться эластичная внутренняя оболочка. Сначала этот процесс идет очень медленно, но после того, как около половины окруженной оболочкой личинки окажется снаружи, происходит резкий толчок и личинка, заключенная в полый шар, как бы «выстреливает» из яйцевого мешка и отскакивает в сторону или задерживается за края щели придатками эластичной оболочки. Замечательно, что сам науплиус при этом остается почти все время совершенно пассивным. Только в самом начале процесса вылупления науплиус производит несколько довольно слабых движений, приводящих, по-видимому, к разрыву яйцевой оболочки. Основную роль здесь играет полупроницаемая эластичная оболочка, через которую диффундирует вода, вызывающая ее разбухание, что сначала заставляет лопнуть мешковую оболочку, а затем выпятиться из нее окруженному эластичной оболочкой науплиусу. Тяжи и нити этой оболочки действуют как пружины, причем расположены они таким образом, что науплиус внутри образованного оболочкой полого шара всегда выходит своим головным концом вперед. За первым науплиусом через ту же щель в мешковой оболочке выпячивается таким же способом второй, или оба «выстреливают» одновременно. Первым толчком к разбуханию эластичной оболочки служит, по-видимому, разрыв науплиусом яйцевой оболочки (рис. 210).
Только через некоторое время новорожденный науплиус, находящийся внутри полностью раздутой, принявшей шаровидную форму эластичной оболочки, начинает двигаться и пытается ее разорвать. Это удается ему не сразу, после чего оболочка спадается и личинка оказывается на свободе. Утомленная тяжелой работой, она сначала почти не способна быстро передвигаться, но это ей и не нужно, так как она находит достаточное количество пищи на поверхности только что покинутой мешковой оболочки, которая обычно за многомесячное пребывание в водоеме обрастает водорослями и покрывается частицами детрита.
При помещении яиц Arcticocamptus arcticus в воду неблагоприятных для его существования водоемов науплиусы внутри яиц развивались нормально, но их вылупления не происходило. Специально поставленными опытами Е. В. Боруцкий доказал, что при сравнительно высоком содержании солей в воде не происходит диффузии воды через эластичную внутреннюю оболочку и она не разбухает. Если вода не имеет кислой реакции, яйцевая оболочка частично не растворяется и не размягчается, что также исключает возможность вылупления науплиуса. Таким образом, обе эти оболочки препятствуют вылуплению науплиуса при попадании яйца в неблагоприятные для рачка условия, обрекающие его на гибель. Действительно, личиночные стадии и взрослые рачки погибали в воде низовых (несфагновых) болот, а также других водоемов, содержащей обычное количество солей и имеющей нейтральную или щелочную реакцию. Все это делает понятным строгую приуроченность Arcticocamptus arcticus к верховым, сфагновым, болотам с их специфическим гидрохимическим режимом.
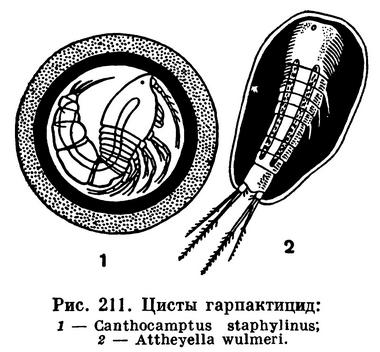
Если Arcticocamptus arcticus в активном состоянии существует весной, то некоторые другие виды пресноводных гарпактицид встречаются только зимой или только летом. При этом известны виды, проводящие период покоя не в стадии покоящегося яйца, как Arcticocamptus arcticus, а в стадии цист, несколько напоминающих описанные выше цисты циклопов. У Canthocamptus staphylinus такие цисты округлые, у Attheyella wulmeri и Attheyella northumbrica они овальные, причем фуркальные щетинки рачка торчат из оболочки (рис. 211).
Среди пресноводных гарпактицид есть виды, способные к партеногенетическому размножению, не свойственному всем остальным веслоногим ракообразным. У широко распространенной в Европе Еlаphoidella bidens самцы вообще неизвестны, а в условиях эксперимента у этого вида удалось получить 5 поколений партеногенетических самок. К партеногенетическому размножению оказался способным и Epactophanes richardi, хотя в естественных условиях он представлен как самками, так и самцами. По-видимому, и некоторые другие виды гарпактицид могут размножаться партеногенетически.
Практическое значение гарпактицид несравненно меньше, чем каланид и циклопов. В некоторых водоемах они составляют заметную часть пищи рыб, в особенности их молоди.