(Hydrozoa Owen, 1843) Класс Гидрозои, Гидроидные, Гидроидные полипы, Гидроидные медузы, Гидромедузы, Class Hydrozoa Owen, 1843 (Hydralike animals, Hydroids, Hydrozoans, Hydromedusae) 8 отрядов
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
Гидроидные (Hydrozoa).
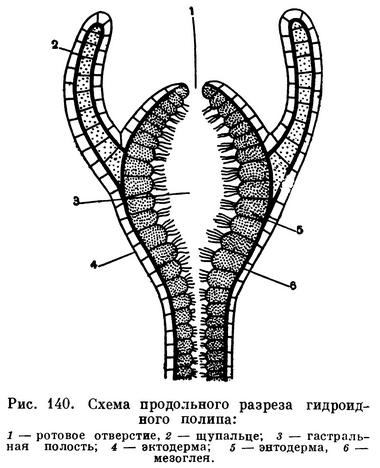
Тело гидроидного полипа цилиндрической или яйцевидной формы, в своей нижней части снабжено обычно ножкой. При помощи ножки одиночные полипы прикрепляются к грунту, водным растениям, раковинам моллюсков, т. е. вообще к каким-либо подводным предметам. У колониальных форм ножка соединяет полипа с колонией. На верхнем конце тела полипа помещается ротовое отверстие, окруженное щупальцами. Щупальца могут располагаться правильными венчиками или без строгого порядка. Число щупалец различно у разных видов. У гигантского одиночного гидроидного полипа бранхиоцериантуса (Branchiocerianthus imperator), достигающего метра в высоту, насчитывается до 380 щупалец, а у маленького монобрахиума (Monobrachium parasiticum) — всего одно щупальце. Чаще же каждый полип снабжен 10—30 щупальцами, которые могут быть или простыми, или снабженными на конце булавовидным утолщением. У одного вида (Cladocoryne) щупальца даже ветвятся. Существуют также полипы вовсе лишенные щупалец. К ним относится пресноводная микрогидра (Microhydra).
Щупальцами полипы захватывают пищу, а такие одиночные полипы, как пресноводная гидра, пользуются ими при передвижении.
Внутреннее строение гидроидного полипа очень простое. Он имеет вид двуслойного мешка. Снаружи полип покрыт слоем клеток эктодермы, а его кишечная полость выстлана энтодермой.
Эктодерма образована особыми эпителиально-мускульными клетками. Тело такой клетки имеет вид многогранной призмочки, клетки расположены тесно, как пчелиные соты. Призматическая часть выполняет покровную функцию. На своем внутреннем конце эпителиальные клетки имеют длинный вырост, тянущийся вверх и вниз вдоль тела полипа. Внутри отростка проходит тонкое мускульное волокно. Такие же сократительные отростки отходят и от клеток энтодермы, но они расположены перпендикулярно к отросткам эктодермальных клеток. Совокупность волокон многих эктодермальных и энтодермальных клеток дает возможность полипу и его щупальцам вытягиваться и сокращаться.
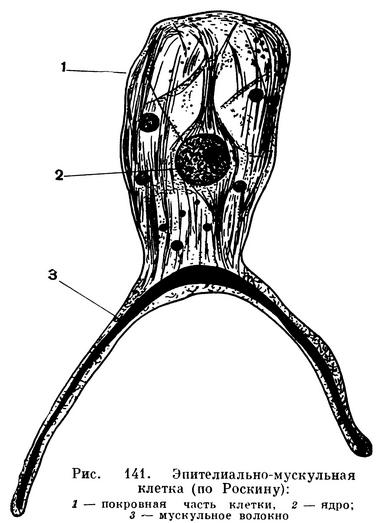
Между эпителиально-мускульными клетками располагаются особые стрекательные клетки, они сложно устроены и имеют для всех кишечнополостных очень важное значение как орудие нападения и защиты. Описание строения и действия этих клеток будет дано ниже, в специальном разделе о стрекательных клетках и действии их яда. В эктодерме есть и особые нервные клетки, снабженные длинными тонкими отростками, образующими в своей совокупности сетевидное нервное сплетение, несколько более плотное на концах щупалец и вокруг рта. У полипов, способных размножаться половым путем (например, у гидры), в эктодерме развиваются также половые клетки. Обычно они скапливаются в нижней или средней части тела и получают название половых желез или гонад. Наконец, здесь же в эпителии имеются резервные клетки (они называются промежуточными или интерстициальными), из которых развиваются эпителиально-мускульные, стрекательные, нервные и половые клетки гидроидных.
Эктодерма многих видов полипов выделяет снаружи тонкую оболочку, состоящую из хитиноподобного вещества. Эта оболочка выполняет у гидроидных полипов роль наружного скелета, служащего опорой и защитой.
У некоторых видов оболочка вокруг полипа образует род чашечки, в которую полипы могут втягиваться в случае опасности.
Энтодерма состоит из железистых и эпителиально-мускульных клеток. Железистые клетки вырабатывают пищеварительный сок (протеолитический фермент), способствующий усвоению белков. Мелкие животные, попавшие в гастральную полость полипа, под влиянием пищеварительных соков перевариваются, распадаются на части. Клетки энтодермы имеют по 2—5 тонких жгутиков, которые все время извиваются и перемешивают содержимое гастральной полости. Частицы пищи, оказавшиеся при этом вблизи стенок гастральной полости, захватываются псевдоподиями, образующимися на поверхности эпителиально-мускульных клеток. Дальнейшее пищеварение происходит уже внутри клетки, подобно тому как это происходит у одноклеточных животных. Гастральные полости отдельных полипов колонии сообщаются друг с другом, образуя единую пищеварительную полость колонии.
В энтодерме имеются и нервные клетки, но их меньше, чем в эктодерме; стрекательные же клетки в энтодерме вообще отсутствуют. Наличие нервных клеток в энтодерме свойственно кишечнополостным, отчасти — иглокожим, но несвойственно всем другим животным.
Между слоями эктодермы и энтодермы у гидроидных полипов расположен тонкий слой неклеточного вещества — мезоглеи.
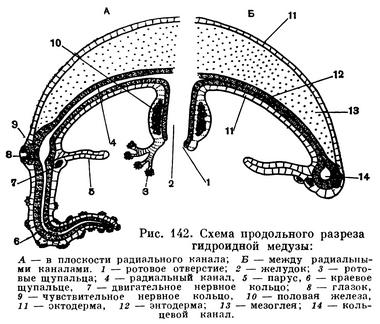
Гораздо сложнее устроены гидроидные медузы. Внешне гидромедуза имеет вид прозрачного диска, зонтика или колокола. Встречаются и причудливые формы медуз с кольцевыми перетяжками посередине тела или медузы почти шарообразной формы.
От внутреннего центра зонтика свешивается ротовой хоботок со ртом на конце. Края рта могут быть гладкими или снабженными четырьмя более или менее бахромчатыми ротовыми лопастями. У некоторых гидромедуз по краям рта сидят маленькие булавовидные ротовые щупальца.
Рот ведет в желудок, занимающий всю полость ротового хоботка, от желудка к периферии зонтика отходят 4 (изредка больше) радиальных канала. На краю зонтика они впадают в кольцевой канал. Совокупность желудка и каналов носит название гастроваскулярной системы. По краю зонтика гидромедуз располагаются щупальца и органы чувств.
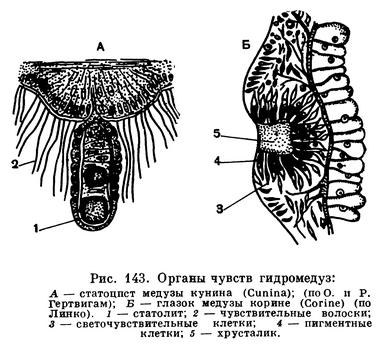
Щупальца служат для осязания и ловли добычи, они густо усажены стрекательными клетками.Часть щупалец может быть видоизменена в особые чувствительные органы, называемые в зависимости от их формы колбочками или усиками. У одной группы гидромедуз (трахилид) щупальца видоизменяются в органы равновесия. Такое щупальце сильно укорочено и сидит как бы на тонком стебельке. На его конце помещается известковое зернышко — статолит. Снаружи щупальце окружено длинными чувствительными волосками (рис. 143, А). Когда тело медузы наклоняется, щупальце под действием силы тяжести остается висеть отвесно и при этом касается чувствительных волосков, которые передают раздражение через нервную систему к эпителиально-мускульным клеткам, что вызывает сокращение их мускульных волокон. Существует предположение, что органы равновесия (они называются статоцистами) служат главным образом для того, чтобы мускулатура работала ритмично. Движение медузы осуществляется благодаря сокращению мускульных волокон на краю зонтика. Выталкивая воду из полости зонтика, медуза получает реактивный толчок и движется верхней стороной зонтика вперед. Усиление реактивной способности достигается благодаря наличию на внутренней стороне зонтика кольцевидного выроста, называемого парусом, который сужает выход из полости зонтика. Каждое сокращение кольцевых мускульных волокон вызывает колебания статоцистов, которые раздражают при этом клетки нервной системы и вызывают новое сокращение. У медуз с вырезанными статоцистами регулярность сокращений зонтика резко нарушается, а их частота уменьшается. У гидромедуз из группы лептолид статоцисты отсутствуют или устроены в виде пузырька, внутри которого находится несколько статолитов, а стенки покрыты чувствительными клетками. Статоцисты лептолид не имеют ничего общего с щупальцами, но несут ту же функцию, что и статоцисты трахилид.
Некоторые гидромедузы обладают светочувствительными органами — глазками, которые всегда расположены на основании щупалец и хорошо заметны благодаря темной окраске. Глазок состоит из двух родов клеток — светочувствительных и пигментных, т. е. несущих в себе красящее вещество. Благодаря наличию пигментных клеток свет падает на чувствительные клетки только с одной стороны. Световое раздражение чувствительные клетки передают нервной системе. Наиболее простые глазки имеют вид пятен, более сложные устроены в виде ямок. В наиболее сложно устроенных глазках полость ямки заполнена прозрачным веществом, выполняющим роль хрусталика (рис. 143, Б).
В связи со свободноподвижным образом жизни гидромедуз их нервная система развита несравненно выше, чем у гидрополипов. Хотя сплетение имеет также вид сети, но на краю зонтика нервные клетки скапливаются очень густо и образуют два кольца. Одно из них (наружное) чувствительное, другое (внутреннее) двигательное.
Чувствительное кольцо проходит вблизи статоцистов, глазков и оснований щупалец и воспринимает полученные от них раздражения. Двигательное кольцо лежит в основании паруса, где сосредоточено большое количество кольцевых мускульных волоконец, которые иннервируются от двигательного нервного кольца.
Медузы раздельнополы, их половые железы располагаются либо в эктодерме ротового хоботка, либо в эктодерме зонтика под радиальными каналами. Здесь они ближе всего к питательным веществам, необходимым для развития половых продуктов. Строение клеток эктодермы и энтодермы медуз такое же, как и у полипов, и потому не требует дополнительного описания, зато мезоглея у медуз развита несравненно сильнее. Она богата водой и имеет студенистый характер, благодаря чему гидромедузы очень прозрачны, многих, даже довольно крупных, медуз трудно увидеть в воде. Особенно сильно развита мезоглея в зонтике.
Общая характеристика, данная выше, относится только к одному подклассу гидроидов (Hydroidea), но в класс гидроидных (Hydrozoa) входит также весьма своеобразный подкласс сифонофор (Siphonophora). Сифонофоры живут только в море. Они похожи на колонии гидроидных полипов, целиком, всей колонией перешедших к пелагическому существованию.
Колониям сифонофор свойственно в наибольшей степени явление полиморфизма, равное которому можно найти только среди муравьев и термитов. В состав колонии сифонофор входят особи, имеющие особое строение и выполняющие различные функции. Одни из них выполняют функции движения, другие — питания, третьи — выделения, четвертые — размножения, пятые — защиты. Сифонофоры — это одни из самых удивительных и самых красивых созданий морской стихии.
ЖИЗНЕННЫЙ ЦИКЛ КИШЕЧНОПОЛОСТНЫХ, ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
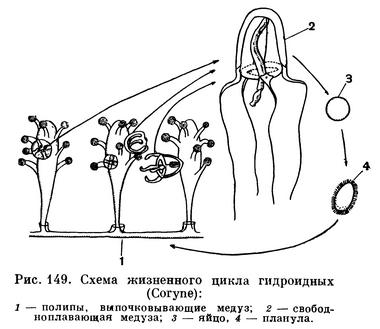
Иначе протекает жизненный цикл у гидроидных и сцифоидных. Дробление яйца и развитие планулы происходит у них так же, как у коралловых полипов, аналогичным способом возникает и первый полип — основатель колонии, а потом и вся колония (у сцифоидных колонии обычно не образуется). Однако полипы гидроидных и сцифоидных совершенно лишены способности к половому размножению, зато они выпочковывают или отделяют не только подобных себе полипов, но совершенно не похожих на них медуз. Медузы и являются половыми особями. Образование медуз у гидроидных и сцифоидных происходит по-разному. У гидрополипов медузы возникают из почек. Вначале такая почка внешне похожа на обычную почку, из которой развивается полип, но затем появляются отличия. Медузоидная почка сильно увеличивается в размере, становится полупрозрачной, внутри ее появляются 4 радиально расположенных канала и отходящий от места их скрещивания ротовой хоботок. Теперь молодая медузка внешне напоминает маленький колокольчик или зонтик. Вскоре она отрывается от колонии полипов и уплывает. Медузы самостоятельно питаются и увеличиваются в размерах, затем у них возникают половые железы. Яйца и сперма выбрасываются прямо в морскую воду, где происходит оплодотворение яиц. Из яиц возникает личинка — планула. На рисунке 149 показан типичный жизненный цикл на примере гидроида из рода Корине (Coryne sarsii).
КЛАСС ГИДРОИДНЫЕ (HYDROZOA) (Растения или животные?)
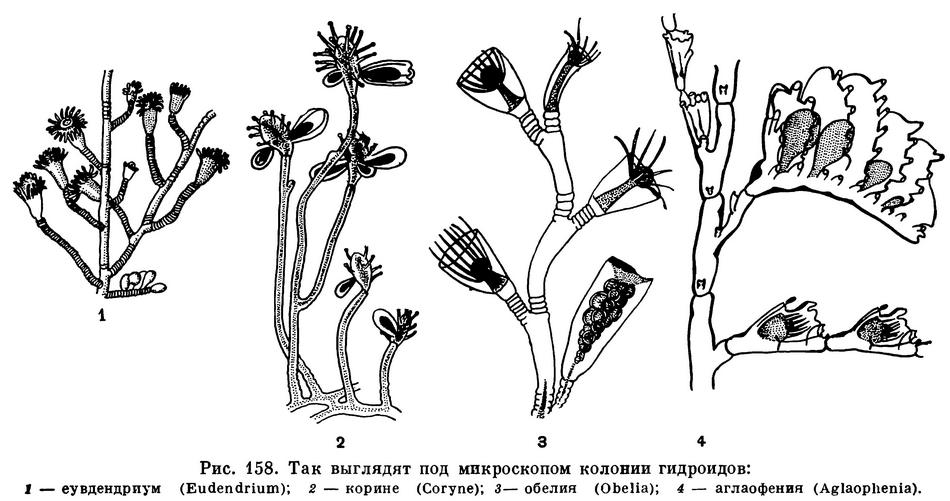
Бродя по берегу моря, мы часто видим гряды выброшенных волнами зеленоватых, бурых или коричневых спутанных комков жестких нитей. Очень мало кто знает, что значительная часть этой «морской травы» имеет не растительное, а животное происхождение. Всякий, кто бывал на море, конечно, видел, что все камни, сваи и другие подводные предметы обрастают какими-то нежными кустиками, извивающимися в волнах. Если собрать такие кустики и посмотреть их под микроскопом, то наряду с настоящими водорослями можно увидеть и нечто совсем особенное. Вот перед нами коричневая членистая веточка с розовыми комочками на концах. Вначале розовые комочки неподвижны, но стоит им несколько минут постоять спокойно, и они начинают шевелиться, вытягиваться в длину, приобретая форму маленького кувшинчика с венчиком щупалец на верхнем конце тела. Это полипы гидроида эудендриум (Eudendrium), живущего в наших северных морях, в Черном море и в морях на Дальнем Востоке. Рядом другая, также членистая, но более светлая веточка. Полипы на ней также розовые, но по форме напоминают веретено. Щупальца сидят на теле полипа без всякого порядка, и каждое снабжено на конце маленькой головкой — скоплением стрекательных клеток. Движения полипов медлительны, они то сгибают свое тело, то медленно покачиваются из стороны в сторону, но чаще сидят неподвижно, широко расставив щупальца — подстерегают добычу. На некоторых полипах можно заметить почки или молодых развивающихся медуз. Подросшие медузки энергично сжимают и разжимают свой зонтик, тонкая нить, связывающая медузу с полипом, при этом обрывается, и медузка толчками уплывает прочь. Это полипы корине (Coryne) и их медузы. Они также обитают и в арктических и в умеренных морях.
А вот еще кустик, полипы на нем сидят внутри прозрачных колокольчиков. Внешне они очень похожи на полипов эудендриум, но ведут себя совершенно иначе. Стоит слегка дотронуться до полипа концом иголки, как он стремительно втягивается в глубь своей защитной оболочки — колокольчика. На этом же кустике можно найти и медузок: они так же, как полипы, скрыты внутри прозрачной защитной оболочки. Медузы плотно сидят на тонком бесщупальцевом полипе. Это колония гидроида обелии (Obelia).
Теперь, когда мы можем отличать гидроидов от водорослей, следует обратить внимание на перовидную колонию аглаофении (Aglaophenia). У этого вида, очень обычного у нас на Черном море, кормящие полипы сидят на веточке в один ряд. Каждый заключен в чашечку — гидротеку и окружен тремя защитными полипами.
Свободноплавающих медуз у аглаофении не образуется, а недоразвитые особи медузоидного поколения спрятаны внутрь очень сложного образования — корзиночки (видоизмененной веточки колонии).
Колонии гидроидов поселяются чаще всего на небольших глубинах — от литорали до 200—250 м и предпочитают каменистый грунт или же прикрепляются к различным деревянным и металлическим предметам. Нередко они очень густо разрастаются на подводных частях судов, покрывая их мохнатой «шубкой». В этих случаях гидроиды приносят значительный вред судоходству, так как такая «шуба» резко снижает скорость судна. Известно немало случаев, когда гидроиды, поселяясь внутри труб морского водопровода, почти совсем закрывали их просвет и препятствовали подаче воды. Бороться с гидроидами довольно трудно, так как эти животные неприхотливы и вполне хорошо развиваются, казалось бы, в неблагоприятных условиях. Кроме того, они отличаются быстрым ростом — за месяц вырастают кустики 5—7 см высотой. Чтобы очистить от них днище корабля, приходится ставить его в сухой док. Здесь корабль очищают от наросших гидроидов, полихет, мшанок, морских желудей и других животных-обрастателей.
В последнее время стали применять специальные ядовитые краски — покрытые ими подводные части корабля подвержены обрастаниям в значительно меньшей степени.
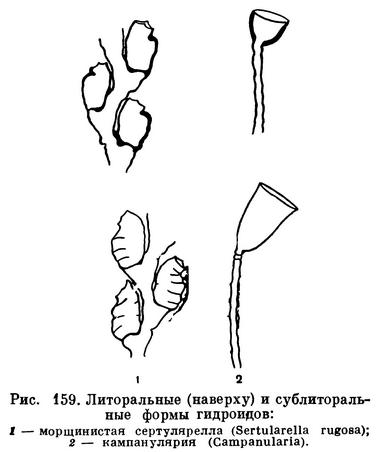
Гидроиды, поселяющиеся в литоральной зоне, совершенно не боятся прибоя. У многих из них полипчики защищены от ударов скелетной чашечкой — текой; на колониях, растущих в самой прибойной зоне, теки всегда значительно толще, чем у тех же видов, живущих поглубже, где прибойные волны не ощущаются (рис. 159).
У других гидроидов из прибойной зоны колонии имеют длинные, очень гибкие ствол и ветви, или же они поделены на членики. Такие колонии извиваются вместе с волнами и потому не ломаются и не рвутся.
На больших глубинах живут особые гидроиды, не похожие на литоральные виды. Здесь преобладают колонии в форме елочки или пера, многие похожи на деревца, а есть виды, напоминающие ершик. Они достигают высоты 15—20 см и покрывают морское дно густым лесом. В зарослях гидроидов живут черви, моллюски, ракообразные, иглокожие. Многие из них, например рачки морские козочки, находят среди гидроидов убежище, другие, как, например, морские «пауки» (многоколенчатые), не только прячутся в их зарослях, но и питаются гидрополипами.
Если поводить вокруг поселений гидроидов мелкоячеистым сачком или, что еще лучше, использовать для этого специальную, так называемую планктонную, сеть, то среди массы маленьких рачков и личинок различных других беспозвоночных животных попадутся гидроидные медузы. Большинство видов гидромедуз — не очень крупные животные, редко они достигают более 10 см в диаметре зонтика, обычно же размеры гидромедузы 2—3 см, а часто всего 1—2 мм. Гидроидные медузы очень прозрачны. Даже пойманных и помещенных в стеклянную посуду медузок сразу и не заметишь: видны лишь беловатые ниточки каналов и ротовой хоботок. Только внимательно приглядевшись, можно заметить контуры зонтика.
Рассматривая колонию гидроида Корине (Coryne), мы уже видели только что отпочковавшихся маленьких медузок этого вида. У вполне сформированной медузы колокольчатый зонтик 1—8 см высотой, четыре щупальца и длинный, червеобразный ротовой хоботок. Резкими сокращениями зонтика медуза быстро передвигается в горизонтальной плоскости или поднимается вверх. Вниз она медленно опускается под влиянием тяжести, застыв в воде с распущенными щупальцами. Морские планктонные рачки, составляющие главную пищу медузы, постоянно совершают вертикальные перемещения: днем погружаются в глубины, а к ночи поднимаются к поверхности. Они опускаются в более глубокие, спокойные слои воды также и во время волнения. Медузы постоянно двигаются вслед за ними, преследовать свою добычу им помогают два чувства — осязание и зрение. В спокойной воде зонтик медузы все время ритмично сокращается, поднимая животное к поверхности. Как только медуза начинает ощущать вызванное волнами движение воды, ее зонтик перестает сокращаться и она медленно погружается в глубину. Свет она различает при помощи глазков, находящихся в основании щупалец. Слишком яркий свет действует на нее подобно волнению — зонтик перестает сокращаться и животное погружается в более темную глубину. Эти простые рефлексы помогают медузе преследовать добычу и спасаться от гибельного для нее волнения.
Как уже было сказано выше, медуза Корине питается планктонными организмами, преимущественно веслоногими рачками. Глаза медузы не настолько совершенны, чтобы она могла видеть свою добычу, ловит она ее вслепую. Ее щупальца могут очень значительно растягиваться, превосходя высоту зонтика в десятки раз. Вся поверхность щупальца усеяна многочисленными стрекательными клетками. Как только к щупальцу прикоснется рачок или какое-нибудь другое маленькое планктонное животное, оно сразу же поражается стрекательными клетками.
Щупальце при этом быстро сокращается и подтягивает добычу ко рту. Длинный хоботок вытягивается в направлении добычи. Если попался более крупный рачок, медуза оплетает его не одним, а двумя, тремя или всеми четырьмя щупальцами.

Совсем иначе ловят свою добычу медузы с плоским зонтиком и многочисленными щупальцами, например тиаропсис (Tiaropsis) — гидромедуза размером с двухкопеечную монету, очень обычная в наших северных морях. По краям ее зонтика находится до 300 тонких щупалец. У покоящейся медузы щупальца широко расставлены и охватывают значительное пространство. При сокращении зонтика медуза как бы сметает ими рачков, подгоняя их к середине нижней стороны зонтика (см. рис. 160). Рот у тиаропсис широкий, снабженный четырьмя большими бахромчатыми лопастями, которыми медуза захватывает подогнанных рачков.
Несмотря на незначительную величину, гидроидные медузы очень прожорливы. Они поедают массу рачков и потому считаются вредными животными — конкурентами планктоноядных рыб. Обильная пища необходима медузам для развития половых продуктов. Плавая, они разбрасывают в море огромное количество яиц, которые впоследствии дают начало полипоидному поколению гидроидов.
Способностью светиться обладают также некоторые медузы, живущие у поверхности моря, среди них маленькая гидромедуза раткеа (Rathkea), довольно крупная (до 10—15 см в диаметре зонтика) экворея (Aequorea) и сцифоидная медуза пелагия ночесветка (Pelagia noctiluca). Часто эти медузы появляются в очень больших количествах, и тогда ночью волны кажутся пламенеющими, а на лопастях весел возникают огненные шары — так ярко светятся прилипшие к ним медузы. Из донных кишечнополостных светятся некоторые гидроиды и многие морские перья. Однако свечение этих животных, по-видимому, не связано с их питанием, так как они вспыхивают ярким светом только при механическом раздражении. Возможно, способность неожиданно излучать сильный свет является защитной реакцией и служит для отпугивания животных, случайно наткнувшихся в темноте на морское перо.
Выше мы назвали кишечнополостных типичными обитателями моря. Это действительно так для 9000 видов, относящихся к этому типу, но около полутора-двух десятков видов кишечнополостных живет в пресных водах и в морях уже не встречается. Видимо, их предки очень давно переселились в пресные воды.
Очень характерно, что все эти формы как пресноводных, так и солоноватоводных бассейнов относятся только к классу гидроидных и даже только к одному его подклассу — гидроидей (Hydroidea).
Среди всех других кишечнополостных никакой склонности к воде пониженной солености не наблюдается.
К самым типичным обитателям пресных вод всего земного шара, часто образующим очень плотные популяции, относится несколько видов гидр, составляющих отряд гидровых (Hydrida).
ПРЕСНОВОДНАЯ ГИДРА
В каждой группе животного царства имеются излюбленные зоологами представители, которых они используют в качестве основных объектов при описании развития и строения животных и над которыми ставят многочисленные опыты по физиологии. В типе кишечнополостных таким классическим объектом служит гидра. Это и понятно. Гидр легко найти в природе и сравнительно просто содержать в лабораторных условиях. Они быстро размножаются, и потому в короткий срок можно получить массовый материал. Гидра — типичный представитель кишечнополостных животных, стоящих у основания эволюционного древа многоклеточных. Поэтому ее используют при выяснении всех вопросов, касающихся изучения анатомии, рефлексов и поведения низших многоклеточных. Это в свою очередь помогает понять происхождение более высокоорганизованных животных и эволюцию их физиологических процессов. Кроме того, гидра служит прекрасным объектом при разработке таких общебиологических проблем, как регенерация, бесполое размножение, пищеварение, осевой физиологический градиент и многое другое. Все это делает ее незаменимым животным как для учебного процесса — от средней школы до старших курсов университета, так и в научной лаборатории, где решаются проблемы современной биологии и медицины в разных их отраслях.
Первым человеком, который увидел гидру, был изобретатель микроскопа и крупнейший натуралист XVII—XVIII вв. Антон Левенгук.
Разглядывая водные растения, Левенгук увидел среди других мелких организмов странное животное с многочисленными «рогами». Он наблюдал также рост почек на его теле, образование у них щупалец и отделение молодого животного от материнского организма. Левенгук изобразил гидру с двумя почками, а также нарисовал кончик ее щупальца со стрекательными капсулами, каким он видел его под своим микроскопом.
Однако находка Левенгука почти не привлекла внимания его современников. Лишь через 40 лет гидрой заинтересовались в связи с необычайным открытием молодого учителя Трамбле (Trambley). Занимаясь в свободное время изучением малоизвестных тогда водных животных, Трамбле обнаружил существо, похожее и на животное и на растение. Чтобы установить его природу, Трамбле разрезал это существо пополам. Регенеративные способности низших животных тогда были еще почти неизвестны и считалось, что восстанавливать утраченные части могут только растения. К удивлению Трамбле, из каждой половинки выросла целая гидра, обе они шевелились, хватали добычу, значит, это было не растение. Возможность превращения куска тела гидры в целое животное была воспринята как значительное открытие в науке о жизни, и Трамбле занялся глубоким и серьезным изучением гидры. В 1744 г. он опубликовал книгу «Мемуары к истории одного рода пресноводных полипов с руками в виде рогов». В книге было очень подробно изложено строение гидры, ее поведение (движения, ловля добычи), размножение почкованием, некоторые моменты физиологии. Для проверки своих предположений Трамбле проделал с гидрой ряд опытов, положив начало новой науке — экспериментальной зоологии.
Если собрать в прибрежной части озера или реки водные растения и поместить их в аквариум с чистой водой, то вскоре на них можно увидеть гидр. Вначале они почти незаметны. Потревоженные животные сильно сжимаются, их щупальца сокращаются. Но по истечении некоторого времени тело гидры начинает вытягиваться, ее щупальца удлиняются. Теперь гидру можно как следует разглядеть. Форма ее тела трубковидная, на переднем конце находится ротовое отверстие, окруженное венчиком из 5—12 щупалец. Сразу под щупальцами у гидр большинства видов имеется небольшое сужение — шейка, отделяющая «голову» от туловища. Задний конец гидры сужен в более или менее длинную ножку, или стебелек, с подошвой на конце (у некоторых видов ножка не выражена). Посередине подошвы находится отверстие, так называемая аборальная пора. Гастральная полость гидры сплошная, перегородок в ней нет, щупальца полые, похожие на пальцы перчатки.
Стенка тела гидры, как и у всех кишечнополостных, состоит из двух слоев клеток, их тонкое строение уже было описано выше, и потому здесь мы остановимся только на одной особенности клеток тела гидры, которая полностью изучена пока лишь на этом объекте и не обнаружена у других кишечнополостных.
Гидра — типичное пресноводное животное, лишь в очень редких случаях гидр находили в слабо осолоненных водоемах, например в Финском заливе Балтийского моря, и в некоторых солоноватоводных озерах, если содержание солей в них не превышало 0,5%. Гидры живут в озерах, реках, ручьях, прудах и даже в канавах, если вода в них достаточно чистая и содержит большое количество растворенного кислорода. Держатся гидры обычно вблизи берегов, в неглубоких местах, так как они светолюбивы. При содержании гидр в аквариуме они всегда перебираются на его освещенную сторону.
Гидры — малоподвижные животные, большую часть времени они сидят на одном месте, прикрепившись подошвой к веточке водного растения, камню и т. д. Излюбленная поза гидры в спокойном состоянии — висеть вниз «головой», спустив несколько расставленные щупальца.
Гидра — прожорливый хищник, она питается инфузориями, планктонными рачками, малощетинковыми червями, нападает также на мальков рыб. Гидры подстерегают свою добычу, подвесившись на какой-нибудь сучок или стебель водного растения, и, широко расставив щупальца, постоянно делают круговые поисковые движения. Как только одно из щупалец гидры коснется жертвы, к ней устремляются остальные щупальца и парализуют животное стрекательными клетками. Теперь от медлительности гидры не остается и следа, она действует быстро и «решительно». Добыча подтягивается щупальцами ко рту и быстро заглатывается. Мелких животных гидра глотает целиком. Если жертва несколько крупнее самой гидры, она также может ее заглотить. При этом рот хищницы широко раскрывается, а стенки тела сильно растягиваются. Если добыча не помещается в гастральную полость целиком, гидра заглатывает лишь один ее конец, по мере переваривания проталкивая жертву все глубже и глубже. Сытая гидра несколько съеживается, и ее щупальца сокращаются.
В крупнейшем центре физиологии — в созданном академиком Павловым институте в Колтушах стоит памятник собаке. Большая часть законов, изложенных в учении Павлова, была открыта при постановке опытов на собаках. Возможно, такой же памятник заслуживает и маленький пресноводный полип.
ПРЕСНОВОДНАЯ МЕДУЗА
В 1880 г. в бассейне с тропическими растениями лондонского ботанического общества вдруг появились медузы. Сразу два зоолога Ланкестер (Lankester) и крупный знаток кишечнополостных Олмен (Аllman) сообщили об этой находке на страницах журнала «Нечур» («Природа»). Медузки были очень маленькие, самые крупные из них едва достигали 2 см в диаметре зонтика, однако их появление взволновало тогдашних зоологов: до этого и не предполагали, что могут существовать пресноводные медузы. Медуз считали типичными обитателями моря. Незадолго перед этим в бассейн было посажено великолепное южноамериканское водное растение виктория-регия, поэтому высказывались предположения, что медузы были завезены в Лондон вместе с посадочным материалом из Амазонки. Через некоторое время медузы исчезли из бассейна так же таинственно, как и появились. Их обнаружили снова лишь через пять лет тоже в Лондоне, но уже в другом бассейне с тем же тропическим растением. В 1901 г. эти медузы появились в Лионе (Франция), также в оранжерейном бассейне с викторией-регией. Затем их стали находить в Мюнхене, Вашингтоне, Петербурге, Москве. Медуз обнаруживали то в бассейнах ботанических садов, то в аквариумах с тропическими рыбками. К удивлению любителей-аквариумистов, у них неожиданно появились новые питомцы. Крохотные медузки (часто всего 1—2 мм в диаметре зонтика) вдруг оказывались в большом количестве в аквариуме, в котором накануне не было ни одной. Несколько дней можно было наблюдать, как медузки толчками передвигаются в воде и охотно поедают маленьких рачков. Но в один прекрасный день, заглянув в свой аквариум, хозяин находил в нем только рыб, никаких медуз там не было.
К этому времени пресноводная медуза была подробно описана в специальной зоологической литературе. Оказалось, что она принадлежит к классу гидроидных. Назвали ее краспедакустой (Craspedacusta). Самые маленькие медузки имеют полусферический зонтик, 4 радиальных канала и 8 щупалец. По мере роста медузы форма ее зонтика становится все более плоской, а число щупалец увеличивается.
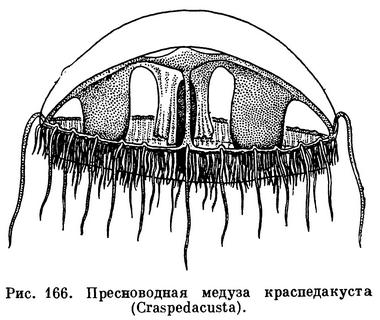
Половозрелые медузы достигают 2 см в диаметре и несут по краю зонтика широкий парус и около 400 тонких щупалец, усаженных стрекательными клетками. Ротовой хоботок четырехгранный, с крестообразным ротовым отверстием, края рта слабоскладчатые. В месте отхождения от ротового хоботка радиальных каналов развиваются 4 гонады. Медузы очень прозрачны, их мезоглея бесцветна, а щупальца, радиальные каналы, ротовой хоботок и гонады имеют беловатую или кремовую окраску.
Эта медузка загадала зоологам сложную загадку. Если согласиться с мнением о том, что она попадает в оранжереи вместе с растениями из тропиков, то как она выживает при перевозке? Виктория-регия транспортировалась с берегов Амазонки в виде семян или корневищ. Нежные медузы, случайно захваченные вместе с корневищами, несомненно, должны погибнуть во время длительного пути через океан. Но если даже предположить, что медуза, несмотря на подсыхание, может выжить, то как она попадает в маленькие аквариумы любителей экзотических рыбок?
Вскоре медуз стали находить и в природных водоемах. Первый раз ее поймали в реке Янцзы в Китае, потом в Германии, затем в США. Однако и в естественных и в искусственных водоемах находки были очень редкими и всегда неожиданными: так, однажды медуз обнаружили в хранилищах вашингтонского водопровода.
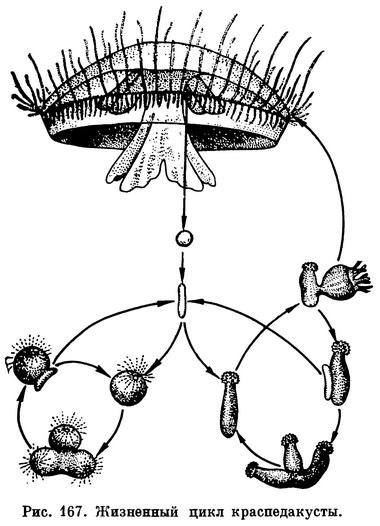
Наблюдениями над медузой удалось установить, что она отпочковывается от крошечных бесщупальцевых полипчиков, названных микрогидрами (Microhydra). Эти полипы были найдены еще в 1884 г. в тех же бассейнах в Лондоне, где ловили и медуз, но тогда никто не предполагал о связи между этими двумя столь непохожими существами. Полипы микрогидры видны простым глазом как белые точки на фоне зеленых листьев водных растений, на которых они обычно поселяются. Их высота обычно не превышает 0,5—1 мм, форма тела напоминает кеглю: туловище в виде бутылки, а на короткой шейке сидит шаровидная «головка» с ртом посередине. Головка густо усажена стрекательными клетками, щупалец нет. Полипы иногда образуют примитивные колонии из 2—7 особей. Микрогидра размножается почкованием и образует подобных себе бесщупальцевых полипов. Время от времени от одной из сторон тела полипа отделяется группа клеток, имеющая форму маленького червячка. Такие группы клеток называются фрустулами. Фрустула способна, извиваясь, ползать по дну и забираться на водные растения, здесь она превращается в молодую микрогидру.
Однажды удалось наблюдать, как на теле микрогидры из почки стала развиваться медуза; когда она отделилась от полипа и начала плавать, то в ней легко было узнать молодую краспедакусту. Удалось также проследить за развитием яиц краспедакусты. Вначале из яйца образуется червеобразная личинка, лишенная ресничек и очень похожая на фрустулу микрогидры. После некоторого периода ползания по субстрату личинка прикрепляется к нему и превращается в бесщупальцевого полипа. Так было установлено, что медуза краспедакуста и полип микрогидра принадлежат к одному виду кишечнополостных, но к разным его поколениям.
Проведенные опыты показали, что на смену поколений у этого вида гидроидных чрезвычайно большое влияние оказывают условия среды. Выпочковывание медуз на полипах происходит только при температуре воды не ниже 26—33°С, а выпочковывание полипов и отделение фрустул — при температуре 12—20°С. После этого стало ясно, что существование вида может длительное время поддерживаться за счет размножения полипов. На маленьких неподвижных микрогидр ни аквариумисты, ни ботаники в оранжереях не обращают внимания, так как они почти и не видны простым глазом, очень трудно найти их и в природе. Полипы могут долго жить в аквариуме, а при повышении температуры у всех полипов появляются медузоидные почки и они отделяют медуз. Медузы краспедакусты подвижны и могут быть замечены в воде невооруженным глазом. Теперь становится понятным, почему их почти всегда находили в бассейнах с тропическими растениями и рыбками: эти бассейны искусственно подогревались. Неясно только одно: всегда ли медузы жили в Европе или были привезены туда? (Полипы, возможно, способны перенести некоторое высыхание и длительный путь в неблагоприятных условиях.) И где же родина микрогидры — краспедакусты?
Ответить на этот вопрос довольно трудно. С момента первого обнаружения медуз в Лондоне было описано свыше 100 случаев их нахождения в самых различных частях мира. Вот краткое описание распространения вида. В СССР местом их обитания является Любовское водохранилище вблизи Тулы, река Дон, озеро Караязы около Тбилиси (на высоте почти 2000 м над уровнем моря), река Кура, искусственные водоемы в Старой Бухаре. Кроме того, медузы и полипы неоднократно появлялись в аквариумах у любителей-рыбоводов и в университетах Москвы и Ленинграда. За пределами нашей страны этот вид был обнаружен почти во всех странах Европы, в Индии, Китае и Японии, в Австралии, Северной и Южной Америке. Указать теперь, где его родина и куда он был завезен, невозможно.
Совсем недавно этот вид кишечнополостных снова заставил зоологов задуматься. Теперь, когда распространение, образ жизни, строение полипов и медуз, казалось, были хорошо изучены, вдруг открылось, что из яиц краспедакусты могут развиваться полипы двух родов — описанные выше бесщупальцевые и снабженные щупальцами. Оба рода полипов образуют фрустулы. Щупальценосные полипы при помощи почкования образуют и подобных себе и бесщупальцевых полипов, медуз они выпочковывать не могут. Бесщупальцевые полипы образуют подобных себе полипов и медуз, но не способны выпочковывать полипов, снабженных щупальцами. Из фрустул образуются обе формы полипов. Щупальценосные полипы пока были обнаружены только два раза: в 1960 г. в Венгрии и в 1964 г. в аквариуме Ленинградского университета. Условия, вызывающие их появление, пока неясны. В реках Индии и великих озерах Африки обитает еще два вида пресноводных медуз, близких родственников краспедакусты. Хорошо известна медуза из африканского озера Танганьика, называемая лимнокнидой (Limnocnida tanganjice).
ПРОИСХОЖДЕНИЕ ПРЕСНОВОДНЫХ КИШЕЧНОПОЛОСТНЫХ
Как уже говорилось выше, число чисто пресноводных кишечнополостных ограничивается тремя видами медуз (и их полипов), десятком видов гидр и паразитом икры осетровых рыб полиподиумом, свободная стадия которого живет в реках. Кроме этого, имеются также некоторые виды гидроидных, способные выдерживать значительное опреснение и даже проникающие в пресные воды, но здесь они не могут размножаться, и потому их нельзя считать полноценными пресноводными животными.
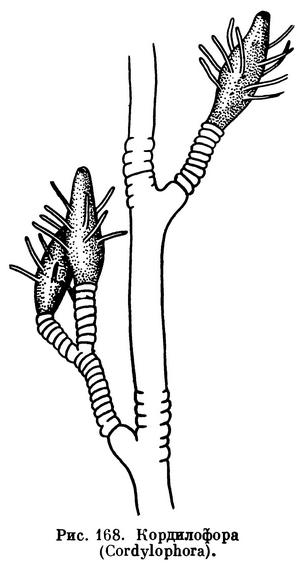
Из числа таких гидроидов в первую очередь нужно сказать о кордилофоре.
Кордилофора образует небольшие нежные колонии в виде кустиков высотой до 10 см. Полипы сидят на концах ветвей и имеют веретеновидную форму. У каждого полипа 12—15 щупалец, сидящих без строгого порядка в срединной части тела. Свободноплавающих медуз у кордилофоры нет, особи медузоидного поколения прикреплены к колонии.
Этот вид был впервые обнаружен академиком Российской академии П. С. Палласом в 1771 г. в северной части Каспийского моря, потому-то кордилофора и называется каспийской (Cordylophora caspia). Однако ее распространение вовсе не ограничивается этим бассейном, она обитает в Балтийском, Черном и Азовском морях, а также встречается вдоль всего атлантического побережья Европы и в устьях всех крупных рек Азии, Америки и Австралии. Этот вид поселяется только в сильно опресненных участках моря и живет на небольшой глубине, обычно не глубже 20 м.
В названии, данном Палласом кордилофоре, — каспийская — имеется и свой смысл. Дело в том, что родина кордилофоры — Каспийское море. Только в середине прошлого столетия кордилофора по Волге и Мариинской системе проникла в Балтийское море, где в силу его малой солености (0,8%) нашла свою вторую родину. Кордилофора — организм-обрастатель; она селится на всех твердых подводных предметах, как неподвижных, так и подвижных. Дальнейшую помощь в расселении ей оказали бесчисленные корабли, стекающиеся со всех сторон в Балтийское море. Возвращаясь домой, они увозили из Балтийского моря на своем днище незваного гостя, «нарушителя границ».
По руслам рек кордилофора проникает иногда довольно высоко, но здесь она не получает полного развития, ее колонии по мере удаления от моря становятся маленькими, на них уже не возникают особи медузоидного поколения.
Попробуем представить себе, какие же у кишечнополостных есть возможности попасть в пресноводный водоем. Таких возможностей три. Одна из них — это переход к паразитизму. Возможно, что так попал в наши реки полиподиум. Предки современных осетровых рыб были настоящими морскими животными, в море они и получили этого паразита. Затем, когда осетровые стали проходными рыбами (такими, которые часть жизни проводят в море, а часть в пресноводных водоемах), полиподиум вместе с хозяином попал в реку. Известно, что паразитический образ жизни в значительной мере изменяет физиологию паразита, вынуждая его приспосабливаться к новым условиям жизни. Вместе с осетровыми рыбами к жизни в пресной воде приспособились и их паразиты.
А как же попадали в пресные водоемы свободноживущие кишечнополостные? Не могут ли они использовать для этого устья рек, впадающих в море? Конечно, могут, но при этом им придется преодолеть два препятствия. Одно из них — это понижение солености. В реки могут попасть только виды, способные выдерживать очень значительное опреснение.
Среди типичных морских обитателей есть такие, на которых даже самое незначительное понижение процентного содержания соли в морской воде действует губительно. К ним относятся почти все коралловые полипы, сцифоидные медузы и большинство гидроидных. Но часть гидроидных все же может существовать и при некотором опреснении. Из кишечнополостных, упомянутых в этой книге, к эвригалинным относится Корине. Этот вид может жить как в воде с нормальной океанической соленостью, так и в опресненных морях, например в Белом и Черном.
Из числа эвригалинных видов и вышли те, потомки которых активно пробирались в пресноводные водоемы. Процесс завоевания рек и озер шел постепенно. Вначале выделилась группа солоноватоводных гидроидов, которые уже не могли вернуться в океан, так как не выносили высокой солености его вод. Затем уже солоноватоводные вплотную приблизились к устьям рек. Далеко не все из них смогли преодолеть этот «барьер», большинство так и осталось в речном устье. В настоящее время по этому пути идет кордилофора.
Попав в реку, морские животные встретили на своем пути и другой «барьер» — течение. При активном проникновении морских или солоноватоводных кишечнополостных в пресные воды им неизбежно приходилось преодолевать встречный поток воды, сносивший обратно в море планктонных медуз и неспособных к самостоятельному движению прикрепленных полипов или их колонии. Продвижение таких прикрепительных полипов навстречу течению было затруднено.
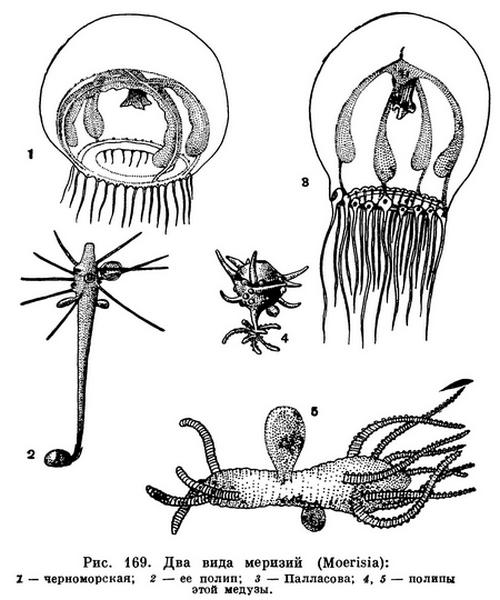
В отдаленные геологические эпохи карта Земли была иной, чем мы видим ее сейчас. Во многих местах современную сушу покрывало море. Когда море ушло, остались замкнутые соленые бассейны, а в них сохранились морские животные. Некоторые из этих бассейнов постепенно опреснялись, причем животные либо погибали, либо приспосабливались к новым условиям. Замкнутое теперь Каспийское море, которое по сути дела является огромным солоноватоводным озером, раньше было соединено с океаном, и в нем сохранилось много животных морского происхождения. Среди них интересное кишечнополостное — Палласова меризия (Moerisia pallasi). У этого вида гидроидных две формы полипов: одни живут колонией на дне, другие ведут планктонный образ жизни. Плавающие полипы образуют колонии из двух особей, соединенных друг с другом своими ножками. Время от времени колония разрывается пополам, и на месте разрыва у каждого полипа образуются новый венчик, щупальце и рот. Кроме того, полипы размножаются и почкованием, отделяя от себя маленьких свободноплавающих медуз. Один близкий вид меризии обитает в Черном и Азовском морях, другой — в соленых озерах Северо-Восточной Африки.
Совершенно ясно, что все три вида меризий произошли от одного общего предка, который когда-то обитал в древнем Сарматском море. Когда Сарматское море ушло, на его месте остался ряд водоемов, в том числе замкнутые Каспийское море и озера Египта. В них развились самостоятельные виды меризий.
Если представить себе, что опреснение водоема идет еще дальше, то можно понять, как могут возникнуть пресноводные медузы. Их способ завоевания пресноводных бассейнов — длительное приспособление к увеличивающемуся опреснению. Двигаться при этом им никуда не нужно, они проделывают путь из моря в пресную воду не в пространстве, а во времени.
В 1910 г. на Атлантическом побережье Северной Америки было поймано несколько маленьких гидромедуз. Оказалось, что они принадлежат к ранее неизвестному виду. Сам по себе этот факт особого значения не имеет. И теперь ежегодно описывают несколько новых видов кишечнополостных — в море еще много неизученного. Интересно другое. Эта медуза — ее назвали блакфордией (Blackfordia) — через 15 лет была поймана в Черном море. Ни в Средиземном море, фауна которого известна очень хорошо, ни на европейском побережье Атлантического океана этот вид не живет. Как же оказалась американская блакфордия в Черном море? Второй случай произошел совсем недавно. Один из видов гидроидов, обитающих в Кильском канале, — бугенвиллия — неожиданно был обнаружен опять-таки в Черном море. И блакфордия и упомянутый балтийский гидроид (Bougainvillia megas) — солоноватоводные виды; чтобы попасть из одного бассейна с пониженной соленостью в другой, они должны, подобно кордилофоре, преодолеть препятствие — море с его высокой соленостью.
До постройки канала между Волгой и Доном в Каспийском море было всего два вида кишечнополостных — каспийская меризия и кордилофора. Когда канал был готов и по нему началось судоходство, из Азово-Черноморского бассейна в Каспийское море перебрались еще три вида. Уже через год после ввода канала в строй переселилась в Каспийское море блакфордия, еще через год черноморская меризия, а вслед за ней и тот балтийский гидроид (Bougainvillia megas), который незадолго перед тем попал в Черное море из Кильской бухты. Конечно, так путешествуют не одни только кишечнополостные, но и моллюски, и ракообразные, и черви, и другие солоноватоводные организмы.
«ПАРУСНЫЙ ФЛОТ» КИШЕЧНОПОЛОСТНЫХ
Класс гидроидных делится на два подкласса — гидроидов и сифонофор. К описанию этих удивительных пелагических колониальных кишечнополостных мы и переходим.
Целый мир живых существ обитает на грани двух стихий — воды и воздуха. На плавающих водорослях, обломках древесины, кусках пемзы и других предметах можно обнаружить разнообразных приросших или крепко уцепившихся животных. Не следует думать, что они попали сюда случайно — «терпят бедствие». Напротив, многие из них теснейшим образом связаны и с водной и с воздушной средой и в иных условиях они существовать не могут. Кроме таких «пассивных пассажиров», здесь же можно видеть животных, активно плавающих у самой поверхности, снабженных различно устроенными органами — поплавками, или животных, которые удерживаются, используя пленку поверхностного натяжения воды. Весь этот комплекс организмов (плейстон) особенно богат в субтропиках и тропиках, где не ощущается губительного действия низких температур.
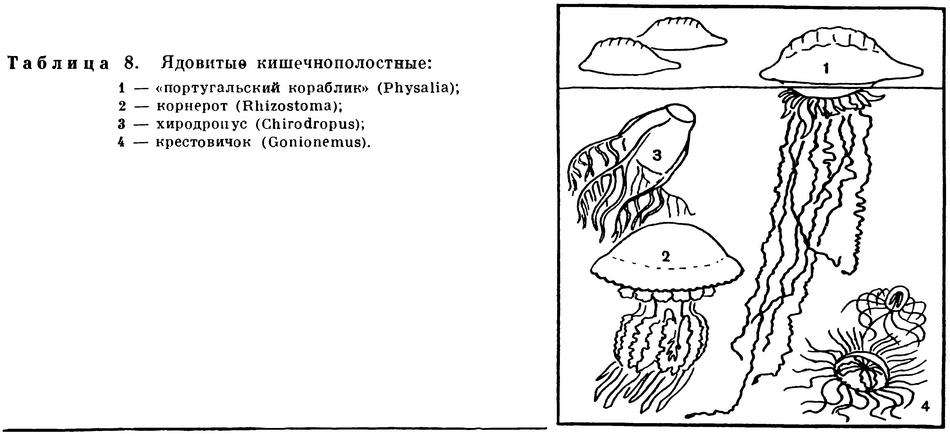
Выше, когда речь шла о действии стрекательных клеток, уже был упомянут «португальский военный кораблик» — крупная сифонофора физалия (Physalia, см. цветную таблицу 8).
Как и все сифонофоры, физалия представляет собой колонию, в состав которой входят как полипоидные, так и медузоидные особи. Над поверхностью воды возвышается воздушный пузырь, или пневматофор, — видоизмененная медузоидная особь колонии. У крупных экземпляров пневматофор достигает 30 см, он обычно имеет ярко-голубой или красноватый цвет. Воздушный пузырь плавает по поверхности моря как туго надутый резиновый шар. Наполняющий его газ близок по составу к воздуху, но отличается повышенным содержанием азота и углекислого газа и уменьшенным количеством кислорода. Этот газ вырабатывается специальными газовыми железами, находящимися внутри пузыря. Стенки пневматофора выдерживают довольно сильное давление газа, так как образованы двумя слоями эктодермы, двумя слоями энтодермы и двумя слоями мезоглеи. Кроме того, эктодерма выделяет тонкую хитиноидную оболочку, благодаря которой прочность пневматофора также значительно увеличивается, хотя его стенки остаются очень тонкими. Верхняя часть пневматофора имеет вырост в виде гребня. Гребень расположен на пневматофоре несколько по диагонали и имеет слегка выгнутую S-образную форму. Все остальные особи колонии расположены на нижней стороне пневматофора и погружены в воду.
Питающие полипы, или гастрозоиды, сидят в один ряд. Они имеют более или менее бутылковидную форму и обращены ротовым отверстием вниз. Каждый питающий полип снабжен одним длинным щупальцем — арканчиком. По всей длине арканчик густо покрыт стрекательными клетками. Рядом с каждым питающим полипом на нижней стороне пузыря прикрепляется основание гонодендра — особи полипоидного поколения. На гонодендре и его боковых выростах расположены грозди редуцированных медузоидных особей — гонофоров, в которых развиваются половые продукты. Здесь же сидят и защитные бесщупальцевые полипы — пальпоны. На каждом гонодендре имеется одна медузоидная особь, называемая нектофором или плавательным колоколом. Половые клетки у нектофора не образуются, а его зонтик достигает значительной величины и способен сокращаться, как у свободноплавающих медуз. Перед наступлением половой зрелости гонофоров гонодендры отрываются от колонии и плавают у поверхности моря, причем нектофор выполняет локомоторные функции.
Благодаря косому расположению гребня на плавательном пузыре физалия асимметрична, причем известны две формы физалий — «правая» и «левая», которые являются как бы зеркальным отображением друг друга. Было замечено, что все физалии, обитающие в одном участке моря, имеют одинаковое строение, т. е. все они либо «правые», либо «левые». В связи с этим высказывалось предположение, что существует два вида или две географические расы физалий.
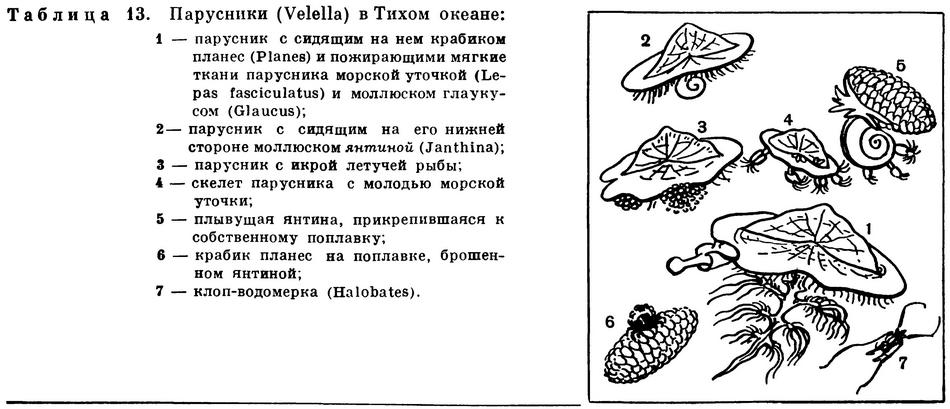
К плейстонным организмам относятся также весьма своеобразные кишечнополостные — порпита (Porpita) и велелла (Velella), которую также называют парусником.
Долгое время этих животных относили к сифонофорам, причем отдельные придатки их считали специализированными особями колонии. Теперь все больше зоологов склоняется к мнению о том, что порпита и парусник представляют собой не колонию, а крупного одиночного плавающего полипа, и относит их к отряду хондрофор (Chondrophora) из класса гидроидных. Тело у них уплощенное; у порпиты оно имеет форму круга, у парусника — овала. Верхняя сторона диска покрыта хитиноидной оболочкой, под которой помещается сложно устроенный воздушный колокол — пневматофор. Он состоит из центральной камеры, большого количества окружающих его кольцевых камер и отходящих от них ко всем частям тела тонких трубочек — трахей, служащих для дыхания. На нижней стороне диска расположены органы полипа. В центре находится ротовой конус, а по периферии располагаются многочисленные щупальца. Между ротовым конусом и щупальцами имеются особые выросты тела — гонодендры, на которых выпочковываются особи медузоидного поколения. Верхняя сторона диска прибрежной порпиты гладкая; у велеллы, живущей в открытом океане, на ней находится высокий вырост треугольной формы — парус. Парус велеллы имеет такое же значение, что и гребень на воздушном пузыре физалии. Он расположен на овальном теле парусника асимметрично и слегка S-образно изогнут. Парус позволяет животному двигаться не по прямой, а маневрировать, хотя, конечно, не произвольно, а более или менее случайно.
В субтропических частях океана, где температура не падает ниже 15°С, парусники встречаются в очень больших количествах. Местами эти крупные кишечнополостные (они достигают 12 см по длинной оси диска) собираются в огромные стаи протяженностью в несколько десятков миль, причем на каждый квадратный метр поверхности океана приходится по паруснику. Вместе с крупными парусниками плавают и молодые, величина которых измеряется миллиметрами.
Ветер, ударяя в парус, гонит стаю велелл по морю, и они могут проходить многие сотни миль.
Обитая в открытом океане, парусники не боятся воды: они не могут утонуть, так как имеют очень совершенный пневматофор, состоящий из большого числа независимых камер. Если волна все же опрокинет велеллу, то при помощи движений краев диска она принимает нормальное положение и снова подставляет парус ветру. Кроме парусников, здесь же можно встретить и много других животных, которые, однако, вначале почти незаметны.
Общеизвестно, что открытое море тропиков имеет интенсивный синий цвет. В связи с этим парусники и большинство животных, которые обитают вместе с ними, также окрашены в голубые или синие тона — это служит им хорошей защитой.
Парусники и обитающие среди них другие животные создают в открытом море особый тесно связанный мирок — плейстонный биоценоз, который по воле течения и ветра все время плывет по поверхности океана.
Велелла, как и все кишечнополостные, — хищник; она питается планктоном, в состав ее пищи входят рачки, личинки различных беспозвоночных, мальки рыб. Все же остальные животные, которые входят в состав плавающего биоценоза, либо питаются парусниками, либо используют их в качестве постоянного или временного субстрата для прикрепления. Таким образом, весь биоценоз существует за счет планктона, но непосредственно используют планктон только одни парусники.
На верхней стороне диска велеллы, как на палубе корабля, путешествуют маленькие голубые крабики планес (Planes). Здесь они находят защиту от врагов, а также получают пищу. Голодный крабик быстро перебирается на нижнюю сторону диска парусника и отнимает у него захваченных планктонных рачков. Наевшись, крабик опять взбирается на верхнюю сторону диска и устраивается под парусом, тесно прижавшись к нему. Крабики никогда не пожирают свой корабль, чего нельзя сказать о многих других плейстонных животных.
На нижней стороне парусника часто можно найти хищного брюхоногого моллюска янтину (Janthina). Янтины объедают мягкие ткани до тех пор, пока от парусника не останется один хитиноидный скелет. Потеряв опору, янтина не тонет, так как хорошо приспособлена к жизни у поверхности воды. Как только поедаемый парусник начинает тонуть, янтина выпускает обильную слизь, образующую пузырьки, наполненные воздухом. Слизь эта очень быстро затвердевает, и получается хороший поплавок, на котором моллюск может самостоятельно плавать, передвигаясь от одного парусника к другому. Подплыв к новой жертве, янтина покидает ненужный ей теперь поплавок и быстро переползает на велеллу. Брошенный поплавок янтины вскоре заселяется гидроидами, мшанками, морскими уточками и другими прикрепленными животными, а также маленькими крабиками; иногда они поселяются и на раковине самого моллюска.
Вместе с янтиной на парусниках поселяется и другой хищный моллюск — голожаберник эолис (Aeolis).
Иногда рядом с парусником можно заметить сопровождающих его голожаберных моллюсков глаукусов (Glaucus). Тело этого безраковинного моллюска вытянутое, рыбообразное, по бокам имеется три пары разветвленных щупальцевидных выростов, при помощи которых моллюск прикрепляется к поверхностной пленке воды. Плавает он темно-синей брюшной стороной кверху, его спинная сторона серебристо-белого цвета. Это делает плывущего глаукуса незаметным и с воздуха и из воды. Проголодавшийся глаукус, подгребая щупальцевидными выростами, подплывает к паруснику и, придерживаясь за него, вырывает и поедает большие куски края диска.
Объеденные моллюсками, парусники гибнут, но от них остается хитиноидный скелет, в котором еще сохраняется система воздушных камер. Такие мертвые парусники плавают некоторое время на поверхности, и на них поселяются личинки усоногих рачков — морских уточек (Lepas fasciculatus). По мере роста новых поселенцев скелет парусника погружается все глубже, а на ножке, при помощи которой морская уточка прикрепляется к субстрату, развивается дополнительный шаровидный поплавок, увеличивающий плавучесть рачка.
Все свободноживущие усоногие раки — прикрепленные животные, исключение составляет только указанный выше вид морской уточки. Когда ее шаровидный поплавок достигает значительной величины, он отделяется от парусника, и после этого морская уточка может самостоятельно держаться на поверхности воды и даже плавать, размахивая ножками. У остальных усоногих рачков взмахи ножек подгоняют к рачку пищу — мелкие планктонные организмы, но этот вид морской уточки, в отличие от всех своих сородичей, ведет хищнический образ жизни. Подплыв к паруснику, морская уточка захватывает край его диска ножками и, передвигаясь вдоль края, быстро выедает значительную часть велеллы.
Кроме описанных здесь животных, в биоценоз велеллы входят также некоторые креветки, ресничные черви, клопы-водомерки и ряд других животных, в том числе один вид летучей рыбы прогнихтис (Prognichthys agae), которая откладывает на парусниках икру. Клопы-водомерки галобатесы живут в тесном контакте с велеллой и порпитой, используя их и как «пирог» и как «плот».
Носящийся в открытом океане мирок велеллы очень ограничен, но все его обитатели тесно связаны друг с другом. Интересно отметить, что большинство видов, составляющих этот биоценоз, относится к таким группам животных, которые обычно ведут донный образ жизни. Исходя из этого, можно с уверенностью сказать, что плейстонные животные происходят от донных (а не от планктонных) организмов, которые потеряли связь с дном и стали прикрепляться к различным плавающим предметам или использовать в качестве опоры поверхностную пленку воды.