(Enteropneusta) Класс Энтеропнейсты, Кишечнодышащие, Class Enteropneusta 1 отряд
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
В настоящее время точка зрения на положение кишечнодышащих в системе животных, видимо, установилась окончательно. Академик И. И. Шмальгаузен и В. Н. Беклемишев помещают класс Enteropneusta среди раздела вторичноротых животных в составе особого типа полухордовых — Hemichorda, в который вместе с кишечнодышащими входят и почти нацело вымершие в настоящее время граптолиты. Кишечнодышащие стоят посредине между двумя основными ветвями вторичноротых животных — типом иглокожих, в древние времена перешедших к сидячему образу жизни и в связи с этим подвергшихся кардинальной перестройке всего плана строения тела своих предков, и типом хордовых, активно двигающихся животных, с активным питанием и прогрессивным развитием различных систем органов, в том числе нервной системы. Кишечнодышащие не могут быть отнесены ни к тому ни к другому типу. Однако современных кишечнодышащих нельзя рассматривать и как прямых предков позвоночных животных. Таким образом, нам предстоит познакомиться еще с одним типом животных — Hemichorda, в который включаются два класса животных: 1 — палеозойские вымершие Graptolithoidea с современными их представителями — Pterobranchia и 2 — Enteropneusta.
Кишечнодышащие (Enteropneusta) представлены в настоящее время 64 видами, относящимися к 12 родам и 4 семействам. Группа крыложаберных (Pterobranchia) включает 21 вид, 3 рода и 2 семейства.
Представители этих двух классов животных сильно отличаются друг от друга и по внешнему виду, и по образу жизни. Но и те и другие исключительно морские животные, не выносящие даже небольшого опреснения. Живут на дне моря.
Кишечнодышащие — свободноподвижные червеобразные животные, с очень сильно вытянутым в длину мягким, обильно покрытым слизью телом. Большинство из них живет в мягком иле либо илистом песке в прибрежной приливо-отливной зоне океана, заселяя тихие куты морских губ и заливов. Грунт в таких заливах в некоторых случаях бывает даже сильно заражен сероводородом и издает неприятный сильный запах. И все-таки животные обитают в нем. Другие виды предпочитают песок с галькой, которые не обнажаются во время отлива, а некоторые, в частности уже известная нам Ptychodera, встречаются на коралловых рифах.
Роясь в грунте, кишечнодышащие заглатывают огромное его количество и извлекают из него мелких животных, растительные остатки и другие органические вещества, которыми они питаются. Они являются, таким образом, в основной своей массе грунтоядными животными, безвыборочно заглатывающими грунт. Однако некоторые крупные виды (Ptychodera) могут быть и хищниками. Есть среди них, по-видимому, и такие, которые питаются мелкими планктонными водорослями, взвешенными в воде (рис. 165).
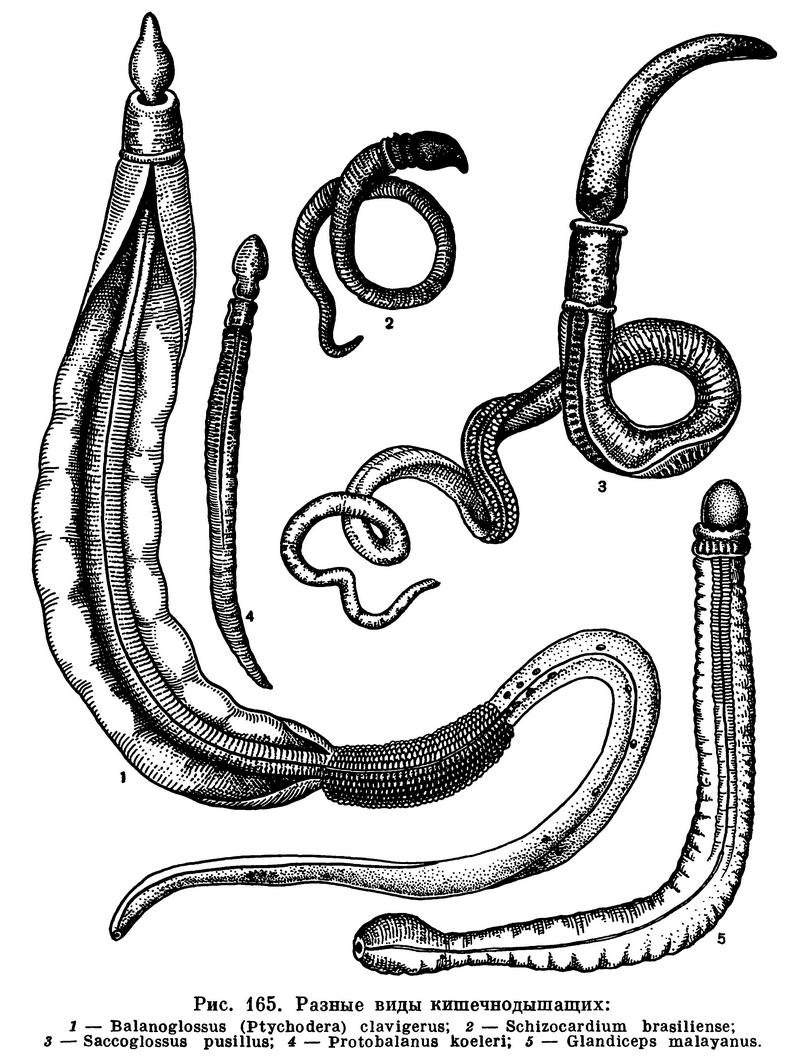
Поселения энтеропнейст на мелководьях бывают иногда очень значительны, но, видимо, непостоянны. Бывают годы, когда эти животные появляются на некоторых участках литорали в огромном количестве, а в другое время в том же районе моря можно встретить лишь единичные экземпляры. Например, живущий у нас на литорали Белого и Баренцева морей Saccoglossus mereschkowskii иногда образует такие массовые скопления, при которых в 1 дм3 грунта встречается 15—20 экземпляров животных размером до 20 см и более. Видимо, их массовое развитие зависит от каких-то особо благоприятных комбинаций условий — температуры, солености, состава грунта и других факторов.
Кишечнодышащие обитают в основном в поверхностной — литоральной зоне, и к ней приурочено в своем распространении большинство их видов. Но они тем не менее встречаются в океане на всех глубинах, хотя количество глубоководных форм очень невелико. До недавнего времени наиболее глубоководным представителем Enteropneusta считался Glandiceps abyssicola, пойманный на глубине 4500 м. Однако среди коллекций глубоководных животных, собранных на советском экспедиционном судне «Витязь» в Тихом океане, есть кишечнодышащие, которые были обнаружены в Курило-Камчатской и Алеутской впадинах на глубине 700—800 м.
Большинство видов кишечнодышащих (60%) обитает в теплых морях, 30% — в морях умеренной зоны. И только некоторые представители этого класса встречаются в арктических областях. Saccoglossus mereschkowskii, например, имеет циркумполярное распространение. Он найден у нас в Белом, Баренцевом, Беринговом, Охотском и Японском морях. Наиболее богата видами кишечнодышащих тропическая зона, причем виды, обитающие в тропиках, распространены, как правило, очень широко. Но среди них есть и такие, которые имеют ограниченный ареал. Некоторые обитают только в Индийском океане и в морях Малайского архипелага.
Известны также свои виды для Ла-Манша, Канады, Южной Африки и других мест. Ни один вид Enteropneusta неизвестен в Антарктике. Только Saccoglossus otagoensis был найден на северо-восточном побережье Новой Зеландии.
Представители класса Enteropneusta имеют четкий план строения тела, не измененный под влиянием сидячего образа жизни, как у Pterobranchia. Размеры тела Enteropneusta очень варьируют и в среднем составляют 10—15 см в длину. Самые мелкие из них имеют 3—5 см. Но известны и очень длинные кишечнодышащие. Balanoglossus cornosus, Balanoglossus clavigerus и др. могут достигать в длину 1,5 м. А самый крупный экземпляр Balanoglossus gigas имел в длину 2,5 м. Диаметр его тела составлял при этом 2 см. Этот гигантский представитель Enteropneusta был найден впервые на Атлантическом побережье Южной Америки, а затем в значительном количестве был отмечен у побережья Бразилии. В 1962 г. в тех же районах была найдена другая гигантская энтеропнейста. Размеры наиболее крупных экземпляров имели в длину более 2 м. Животные относились к новому виду редкого рода Willeya.
Окраска кишечнодышащих также варьирует, чаще всего от серовато-белых до светло-желтых тонов. Часто встречаются и желтовато-коричневые и коричневые особи. Но среди них есть и ярко окрашенные виды — ярко-красные Saccoglossus pusillus, оранжевые Ptychodera flava, темно-коричневые Glandiceps и черно-фиолетовые Ptychodera erythraea. Обычны разные вариации окраски, и разные части тела животных могут быть окрашены различно. Наш северный Saccoglossus mereschkowskii имеет хобот ярко-кирпичного цвета. Воротник и передняя часть тела темнее хобота и другого оттенка — кроваво-алого цвета. К заднему концу окраска постепенно слабеет и тело становится более прозрачным с желтоватыми или зеленоватыми оттенками. Животные, добытые из глубоководных океанических впадин, имели грязную розовато-бежевую окраску и достигали в длину 10—15 см.
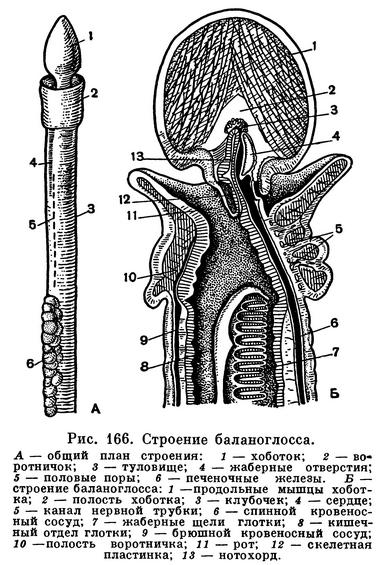
Длинное червеобразное тело кишечнодышащих четко дифференцировано на три отдела — хоботок, воротничок и собственно туловище, где располагается большинство органов животного (рис. 166, А). Хоботок представляет собой мускулистое вытянутое или желудевидное образование. Основная функция хоботка — роющая. Основание его сужено в тонкую шейку, которая охватывается следующим отделом тела — коротким, также мускулистым воротничком, несколько приподнимающимся над поверхностью тела в виде валика. Внутри воротничка, как и внутри хобота, имеется полость, соединяющаяся двумя канальцами через особые воротничковые поры с внешней средой. Следом за воротничком идет длинное туловище животного, составляющее 9/10 всей длины тела. В передней части туловище с боков прорезано двумя рядами многочисленных узких жаберных щелей, расположенных поперечно. Все тело животного одето нежным однослойным эпителием, несущим движущиеся реснички и называемым поэтому мерцательным. Кроме того, сверху животное обильно покрыто слизью, вырабатываемой клетками воротничка. Внутри следуют два слоя мышечных волокон: наружных — кольцевых и внутренних — продольных.
На брюшной стороне тела у основания хобота, ближе к границе последнего с воротничком, лежит рот (рис. 166, Б). Рот ведет в короткую глотку, лежащую на протяжении воротничка. От спинной стороны глотки отходит внутрь хоботка слепое выпячивание. Этот вырост называется нотохордом и считается зачатком спинной струны, или хорды, позвоночных животных. Нотохорд доходит до середины длины хоботка, прилегая к его брюшной стороне. Между ним и брюшной стенкой хоботка развивается плотная, хрящеобразная пластинка, которая называется скелетом хоботка. Вместе с нотохордом эта пластинка придает хоботку известную плотность.
Далее глотка переходит в пищевод, или дыхательный отдел кишечника. В то время как спинная и брюшная стенки пищевода сплошные, боковые стенки пронизаны двумя рядами подковообразных жаберных щелей. Через эти щели полость пищевода сообщается с внешней средой.
Вода, проходящая через рот, поступает в жаберный отдел и через жаберные щели выходит наружу. В перегородках между щелями проходят многочисленные кровеносные сосуды, и кислород, растворенный в воде, передается прямо в кровь животного. Такое строение жаберного аппарата — связь его с кишечником — неизвестно для представителей беспозвоночных животных. Только среди хордовых этот тип строения дыхательного аппарата получает широкое распространение. Благодаря строению дыхательного аппарата эти животные и стали называться кишечнодышащими.
По брюшной стенке пищевода проходит продольный желобок, образованный частично клетками, выделяющими секрет, а частично клетками, несущими реснички. Это эндостиль — образование, которое считается гомологом щитовидной железы позвоночных животных. Его более подробное описание приводится при разборе анатомии оболочников.
Мелкие частицы пищи, попадающие с водой в пищевод, склеиваются слизью эндостиля и гонятся его ресничками вдоль желобка, в пищеварительный отдел кишечника. За жаберным отделом кишечника следует печеночный отдел, несущий на спинной стороне многочисленные печеночные придатки. Затем в виде трубки идет средняя кишка, которая переходит в заднюю. На заднем конце тела она открывается наружу анальным отверстием.
Кровеносная система состоит из продольных кровеносных сосудов — брюшного и спинного. Брюшной сосуд залегает лишь в туловище, тогда как спинной продолжается в хоботке. В хоботке образуется большая центральная кровеносная лакуна — полость, куда поступает кровь. По спинному сосуду кровь течет вперед. Она частично поступает в отходящие от спинного сосуда парные приносящие жаберные сосуды, проходящие через жаберные стенки, и окисляется. Затем окисленная кровь через уносящие жаберные сосуды возвращается назад в спинной кровеносный сосуд. Из центральной кровеносной лакуны кровь но двум окологлоточным сосудам, окружающим глотку в области воротничка, возвращается в брюшной сосуд и направляется к заднему концу тела. Через сеть лакун в области кишечника она вновь поступает в спинной сосуд. Циркуляцию крови обеспечивает сердечный пузырь. Но кровь не проходит через сердце. Этот полый замкнутый мускулистый мешок, лежащий у спинной стенки хобота над нотохордом, своим сокращением проталкивает кровь из центральной кровеносной лакуны по сосудам. Чтобы кровь двигалась в нужном направлении, имеется система специальных клапанов.
Выделительная система состоит из двух пар особых трубочек, посредством которых полости хобота и воротничка, где накапливаются продукты выделения, сообщаются с внешней средой. Эти трубочки называются нефридиями. Как правило, в хоботе сохраняется только один левый нефридий, а в воротничке их имеется два. Кроме того, в хоботке, в центральной кровеносной лакуне, в перегородке между нею и полостью хоботка, имеется особый складчатый орган, называемый клубочком. Этот клубочек является как бы своеобразным фильтром, через который продукты обмена поступают из крови в полость хобота и оттуда выводятся наружу.
Нервная система состоит из двух нервных стволов — брюшного и спинного. Брюшной нервный ствол развит значительно слабее спинного. Он залегает по средней брюшной линии туловища. Спинной нервный ствол продолжается в хоботок. В области воротничка оба ствола соединяются нервным воротничковым кольцом. В области воротничка нервный ствол залегает в виде толстостенной нервной трубки, или цилиндра. Эта часть нервной системы наиболее развита и называется иногда спинным мозгом. Наличие хорошо развитой нервной системы на спинной стороне тела является очень важным признаком хордовых животных. Кишечнодышащие и по этому признаку также принципиально отличаются от остальных беспозвоночных. Но, с другой стороны, они отличаются от хордовых наличием брюшного нервного ствола.
Специальных органов чувств у кишечнодышащих нет, но под наружным покровом их тела расположено множество светочувствительных клеток.
Кишечнодышащие — раздельнополые животные. Самцы и самки у них по внешнему виду неразличимы, хотя иногда их окраска не вполне одинакова. Половая система устроена очень просто. Половые железы образуют от 16 до 100 пар овальных мешочков, лежащих по бокам кишечника в средней части туловища вслед за жаберным его отделом. Каждая гонада открывается наружу коротким протоком, через который половые продукты выводятся в окружающую воду. Кишечнодышащие, за единственным исключением, размножаются половым путем. Почти все они яйцекладущие животные. Представители обоих полов у большинства их видов встречаются в норках, где и происходит спаривание. Оплодотворение наружное. Яйца через половые отверстия или выбрасываются в воду, или прикрепляются к стенкам подземных ходов. Только у тропического рода Glandiceps самцы и самки во время периода размножения покидают дно и плавают некоторое время у поверхности воды, образуя плотные скопления размером от 1 :до 5 м в поперечнике и толщиной до 2 м. Животные беспорядочно двигаются в таких скоплениях, видимо отыскивая партнеров для размножения. Однако среди них присутствуют и неполовозрелые особи. Явление это довольно загадочно, но оно безусловно способствует широкому расселению вида в океане.
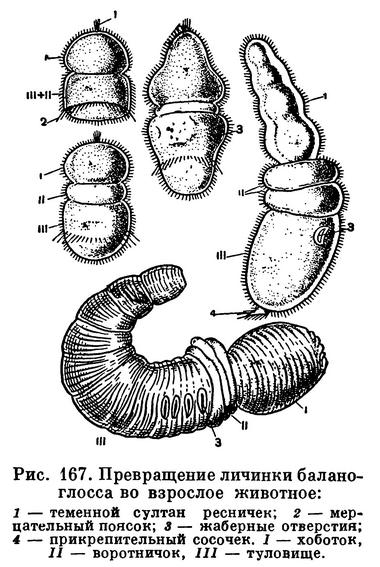
В зависимости от большего или меньшего количества желтка, содержащегося в яйце, развитие у разных видов кишечнодышащих может происходить двумя путями — без превращения и с превращением, или метаморфозом. И у тех и у других видов кишечнодышащих имеется в принципе сходная свободноживущая личинка, но у первых она мало отличается от взрослого организма (Saccoglossus kowalevskii, Dolichoglossus). Развитие у этих видов сильно сокращено. Из яйцевой оболочки выходит уже готовая личинка, состоящая из трех сегментов, каждый из которых соответствует хоботку, воротничку или туловищу взрослых форм (рис. 167). Такая личинка очень тяжела для того, чтобы свободно плавать в воде. Она держится на дне, где ползает или роется в иле. При помощи своих ресничек она может только немного плавать в придонных слоях воды. И хотя эта личинка очень похожа на молодое животное, она все-таки остается настоящей личинкой. У нее имеются временные личиночные органы — чувствительная теменная пластинка с султаном ресничек, движущим органом ей служит прианальный венчик ресничек — мерцательный поясок. После утери этих органов превращение ее во взрослое животное происходит путем вытягивания в длину главным образом третьего сегмента, формирования воротничка и внутренних органов. Появляется нотохорд, закладываются жабры, оформляется нервная система.
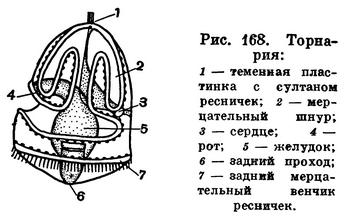
У тех же Enteropneusta, яйца которых бедны желтком, — у Ptychodera, Spengelia и др. — развитие происходит через стадию свободноплавающей пелагической личинки. Эта личинка очень характерна и, как уже указывалось выше, чрезвычайно похожа на пелагических личинок морских звезд (рис. 168). Развитие здесь сопровождается довольно сложным метаморфозом. Вследствие своего постоянного вращения вокруг оси личинка называется торнарией. Зрелая торнария по существу устроена по тому же плану, что и рассмотренная выше личинка. Она имеет кишечник с ротовым отверстием на брюшной стороне и анальным на заднем полюсе тела. На переднем полюсе имеется теменная пластинка с венчиком длинных чувствующих ресничек. Внешний вид этой прозрачной личинки усложнен сильно извитым мерцательным шнуром, проходящим по наружной ее поверхности. Имеется и прианальный венчик ресничек. На первых стадиях своего развития личинка активно плавает в воде султаном вверх и вращается вокруг своей оси. Постепенно объем ее тела несколько уменьшается, утолщаются стенки и личинка теряет свою прозрачность. Она начинает расти в длину, и у нее возникает 5—6 пар жаберных отверстий. А затем наступает резкое превращение — тело ее делится на верхнюю и нижнюю половину. Верхняя часть в дальнейшем растягивается в хобот и обособляется перетяжкой от остального тела, которое формируется из нижней части. Позже около хобота образуется валик — зачаток воротничка, происходит упорядочивание органов, за счет роста заднего конца тело удлиняется, животное приобретает облик взрослой формы и садится на дно.
Среди кишечнодышащих есть один живородящий вид — Xenopleura vivipara, а один вид — Balanoglossus proliferans — способен размножаться как половым, так и бесполым путем. В результате простого поперечного деления тела взрослого животного, у которого совершенно отсутствуют половые железы, получаются новые особи, способные к половому размножению. Таким образом, здесь происходит настоящее чередование поколений — полового и бесполого. Причем оба эти поколения внешне настолько резко отличаются друг от друга, что первое давно было известно науке под названием Balanoglossus capensis, а второе — Balanoglossus proliferans.
Как уже говорилось, большинство видов кишечнодышащих роет в грунте длинные ходы или постоянные норы. Другие же живут на поверхности дна. Все эти виды образуют за счет своей обильной слизи трубки, в которых они и живут. На поверхности грунта обитает большинство Ptychodera, Spengelia, Glandiceps и Glossobalanus. Все они прячутся под камнями, обломками кораллов, иногда глубоко внедряясь в блоки мадрепоров, в зарослях водорослей или под корнями высших водных растений в прибрежной зоне. Некоторых из них находили в мертвых раковинах Муа, наполненных песком. Все эти виды образуют за счет своей обильно выделяемой слизи трубки, в которых они живут. Смешиваясь с песком, слизь быстро застывает в плотную коркообразную массу. Животные могут свободно двигаться в своих трубках. Постройка трубки происходит следующим образом. Хобот животного внедряется в песок, и песчинки, окружающие его, прилипают и вкрапливаются в слизь, обильно покрывающую все тело животного. На конце хобота получается маленький поясок толщиной в 1—2 песчинки. С помощью бьющих в одном направлении назад ресничек эпидермиса (наружного покрова тела) целым потоком один за другим такие поясочки песчинок, смешанных со слизью, передвигаются к концу тела животного. Образуется плотный кожух, охватывающий, как рубашка, все тело животного. Очень быстро он засыхает в прочную трубку.
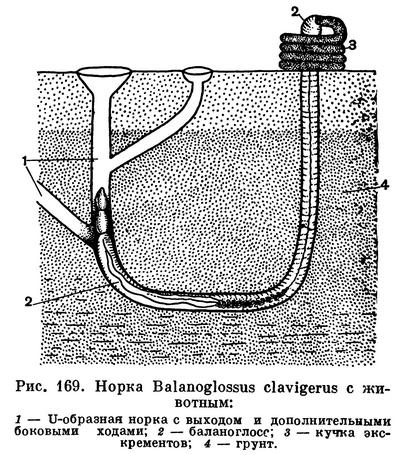
Большинство видов Enteropneusta роет в грунте длинные U-образные норы (рис. 169). Как правило, такая норка вначале уходит вертикальной шахтой вниз на довольно большую глубину. У Balanoglossus и нового гигантского вида Willeya эти ходы достигают 60—80 см. Затем следует изгиб норы, и она снова выходит на поверхность. Постоянно сам ход выстилается быстро засыхающей слизью. Эта слизь склеивает материал, из которого состоят стенки хода, и образует гладкую внутреннюю поверхность. Животные почти никогда не покидают норок. Во время прилива они поднимаются до поверхности грунта и выставляют наружу или передний конец тела, или задний, для того чтобы выбросить фекалии, которые лежат у выхода из норки горкой спирально закрученных колбасок высотой в 3—6 см так, как это наблюдается у червя пескожила (Arenicola marina). По этим кучкам закрученных экскрементов и по небольшим дырочкам входов и можно обнаружить домики Enteropneusta на литорали морей.
При малейшем сотрясении грунта животные молниеносно прячутся в свои норки (или в трубки) и уходят на спасительную глубину. Таким образом они спасаются от врагов, и прежде всего от хищных рыб, питающихся донной фауной. В качестве защиты у многих видов, возможно, служат содержащиеся в слизи иодистые выделения кожных желез. Некоторые Enteropneusta, например Balanoglossus clavigerus, издают столь резкий запах йодоформа, что можно, идя на этот запах, найти их в песке.
Слизь кишечнодышащих может светиться. Свечение прежде всего появляется при сотрясении грунта. Оно менее интенсивно в дневное время. Glossobalanus minutus светится в течение всего дня, а у Ptychodera имеется четкий суточный ритм свечения, который сохраняется неизменным до 8 суток, если содержать животное в темноте. Химизм и биологический смысл их свечения неизвестны.
Видимо, в связи с роющим образом жизни, при котором нежное тело животных может быть легко поранено, а также в качестве защиты от врагов у кишечнодышащих развилась необычайная способность к регенерации, т. е. к восстановлению тела (при его повреждении или разрыве) из отдельных, даже небольших частей. Например, маленький хвостовой кусочек тела Ptychodera способен восстановить целый организм. Интересно, что передняя часть тела не может достроить заднюю, а задние части легко восстанавливают передние.
Мы уже знаем, что специальных органов чувств у кишечнодышащих не имеется. Но все части тела этих животных отчетливо реагируют на механическое раздражение и на свет. Хобот всегда избегает света, и его поведение позволяет установить, что животные могут воспринимать не только сам свет, но и его направление. Свет и даже небольшое механическое раздражение всегда вызывают бегство, уход, закапывание и сжатие тела. Сильнее всего на эти раздражения реагирует хобот.
Как же происходит закапывание животных? За счет прилива и отлива крови к хоботу, а также за счет напряжения его мышц хоботок может становиться то более упругим и сжиматься в жесткий кулак, то расслабляться. Ptychodera, например, косо внедряет в грунт его раздутый передний конец. Потом она сжимает продольные мускулы и перистальтическая волна вздутия движется к заднему концу хобота. Она доходит до его конца за 5 секунд. Когда вздутие доходит до конца хобота, животное прочно зацепляется в грунте, а утонченный передний конец хобота за счет сокращения кольцевых и продольных мышц внедряется глубже в песок. Ставшая в это время тонкой задняя часть хобота подтягивается вперед, к передней его части. При рытье хобот опирается на воротничок, служащий для него в данном случае твердой опорой.
Внутреннее напряжение воротничка при рытье также сильно увеличивается за счет сокращения его толстых мышечных стенок. Кроме того, в полость воротничка через воротничковые поры нагнетается вода. Видимо, при рытье эти поры могут герметически закрываться и тургор воротничка за счет несжимаемой воды увеличивается еще сильнее. Остальное тело совершенно не принимает участия в передвижении. Оно просто подтягивается вслед за передними двумя отделами, при необходимости сильно утончаясь (до 1 мм у некоторых видов) и вытягиваясь. Зарывание происходит довольно быстро. За одну минуту происходит до 12 перистальтических сжатий хоботка. В это же время реснички покровов тела синхронно и постоянно уводят песчинки от кончика хоботка ко рту и к концу тела. Эти песчинки расширяют проход и облегчают внедрение в него туловища. Одновременно стенки хода обрабатываются выделившейся слизью.
Двигаясь вперед, кишечнодышащие заглатывают грунт. Причем заглатывание грунта происходит не просто механически, как у других грунтоедов. Реснички хобота бьют по направлению назад и переносят прилипшие к слизи песчинки и другие частицы длинными параллельными потоками назад. У основания хобота удар ресничек направлен ко рту, так что нити слизи и песка у рта сливаются и создается один мощный поток. Край воротничка при этом отодвигается далеко в сторону от основания хобота и толстый поток пищи проникает в рот. В пищеварительном канале частицы грунта гонятся сначала реснитчатыми клетками эндостиля по жаберному отделу и далее по кишечнику. Пищевой поток все время переворачивается и вращается. Переваривание происходит в печеночном отделе. Вместе с поступившим грунтом в кишечник попадают микроскопические животные и водоросли, которыми, собственно, и питаются Enteropneusta. У некоторых видов кишечник столь сильно набит грунтом, что их нежное тело рвется от тяжести при поднимании животного. Фекалии выбрасываются через задний конец тела, который, высовываясь из норки, совершает коловращательное движение у поверхности грунта. При этом и получаются те спирально закрученные колбаски экскрементов, о которых мы говорили выше.