(Nematoda Diesing, 1861) Тип Нематоды, Круглые черви, Phylum Nematoda Diesing, 1861 = Nemata Cobb, 1919 (Nematodes, Roundworms) 2 класса
Синонимы:
Nemata Cobb, 1919
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
КЛАСС НЕМАТОДЫ, ИЛИ КРУГЛЫЕ ЧЕРВИ (NEMATODA)
Если бы тип немательминтов был ограничен только рассмотренными выше классами животных, то едва ли была бы построена та общая характеристика этого типа, которая возникла в современной науке. Кроме того, его значение и в экономике природы, и в хозяйственной деятельности человека было бы настолько ограниченным, что изучение немательминтов не достигло бы того прогресса, который наблюдается в нашей современности. Глубокий интерес к типу немательминтов не столько определяется характеристиками гастротрих, киноринх и даже коловраток, сколько становится следствием изучения центральной группы, главного и наиболее крупного класса немательминтов — класса нематод (Nematoda).
Естественно, что этот класс немательминтов будет в центре и нашего внимания.
Биологический прогресс. Известный советский ученый академик А. Н. Северцов (1866—1936), исследуя эволюцию позвоночных, подверг глубокому изучению проблему основных направлений филогенеза животных. Он установил, в частности, что для многих естественных групп животных типично состояние эволюционного развития, характеризующееся рядом ведущих признаков, определяющих жизненный успех этих групп. А. Н. Северцов обозначил это как специфическое проявление биологического прогресса.
Биологический прогресс, указывает А. Н. Северцов, характеризуется следующими признаками: 1) численным увеличением особей данной систематической (таксономической) группы; 2) прогрессирующим расселением, т. е. захватом новых сред обитания; 3) возрастающим многообразием форм (подвидов, видов, родов и т. п.).
Класс нематод характеризуется именно этими общими биологическими признаками — признаками биологического прогресса. Нематоды овладели всеми известными науке местообитаниями.
Мы не знаем таких сред обитания, таких биотопов, в которых не было бы нематод. Дно морей и океанов от северного до южного полюса (об этом можно говорить с полной уверенностью) заселено огромным количеством видов и особей нематод. Свободноживущие нематоды известны решительно всюду, во всех точках морского дна, которые подверглись специальным исследованиям. Завоевав дно всех морей и океанов, нематоды проникли, видимо позднее, в солоноватые водоемы. Поэтому очень большое число видов нематод обитает на дне солоноватоводных бассейнов, в том числе в лиманах — этих преддвериях рек, впадающих в моря. Известен ряд фактов, свидетельствующих о том, что в древней истории класса нематод наступил такой важный этап развития, когда они стали проникать в пресные водоемы, и в конце концов многие группы свободноживущих нематод заселили реки. В дальнейшем нематоды совершили еще один важный шаг исторического развития — они проникли в почвенные воды и стали компонентами почвенной фауны — комплекса почвенных биоценозов. Отметим, что на этом экологическое развитие части групп и остановилось. Для некоторых групп нематод оказался открытым путь в сапробиотические очаги. Органические останки растений и животных подвергаются гниению, обусловленному жизнедеятельностью сменяющих друг друга групп гнилостных бактерий, под влиянием которых в почве образуются очаги гнилостного распада. В этих очагах соответствующие группы сапрофитных бактерий последовательно разлагают органический материал на более простые составные части. В частности, белки расщепляются на более простые, растворимые в воде компоненты; полисахариды трансформируются в растворимые ди- и моносахариды; распадаются клетчатка, жиры, пектиновые оболочки клеток растительной ткани и т. д. Поэтому сапробиотические очаги оказались наиболее доступными источниками питания нематод. Так сформировалась богатая видами группа сапробиотических нематод. Именно эта группа стала источником развития других групп нематод, перешедших к паразитическому существованию за счет организмов животных и растений. В конечном счете формируются два больших потока родов и видов нематод, приспособившихся, с одной стороны, к паразитированию в органах человека и животных, а с другой — в органах растений.
Из сказанного выше можно видеть, что все биотопы нашей планеты, все среды жизни, вероятно, почти все многоклеточные животные и все растения оказались средой обитания нематод. Это явление демонстрирует нам один из наиболее ярких примеров биологического прогресса. Общее число видов нематод в настоящее время еще не выяснено наукой. Известный американский специалист по нематодам Н. А. Кобб (N. A. Cobb) полагал, что общее число видов нематод — свободных и паразитических — приближается к одному миллиону.
Сказанное выше оправдывает интерес, который проявляют многие зоологи к этой группе животных. Ввиду этого знакомство с их организацией и биологией полезно и читателю книги.
Строение. Как мы видели, другие классы немательминтов представлены очень мелкими формами: большая часть из них меньше 1 мм, реже встречаются более крупные формы. Среди нематод имеются очень мелкие формы. Одна из них — Trichoderma minutum — достигает всего 80 мк в длину. Такие мелкие формы известны в морях и в почве, где встречаются нематоды длиной 200—300 мк. Однако наряду с этими микроскопическими формами известны настоящие гиганты из мира нематод. Самки лошадиной аскариды достигают 37 см в длину. Свайник-великан (Dioctophyme renale) длиной в 1 м, a Placentonema gigantissima — паразит кашалота, этого огромного китообразного, по величине тела, без преувеличения, напоминает удава, самки его до 8 с лишним метров в длину! Таким образом, у нематод мы наблюдаем очень значительные колебания длины тела — от 80 мк до 8 м. Эти колебания длины тела — одно из свидетельств многообразия тех сред жизни, которыми овладели нематоды.
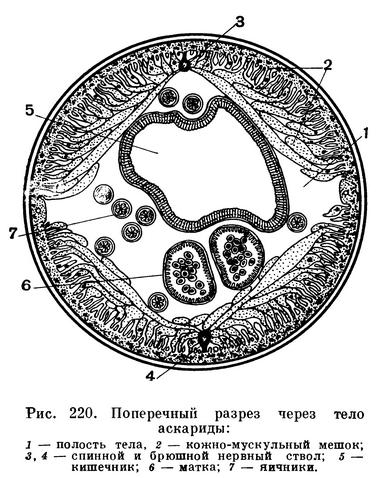
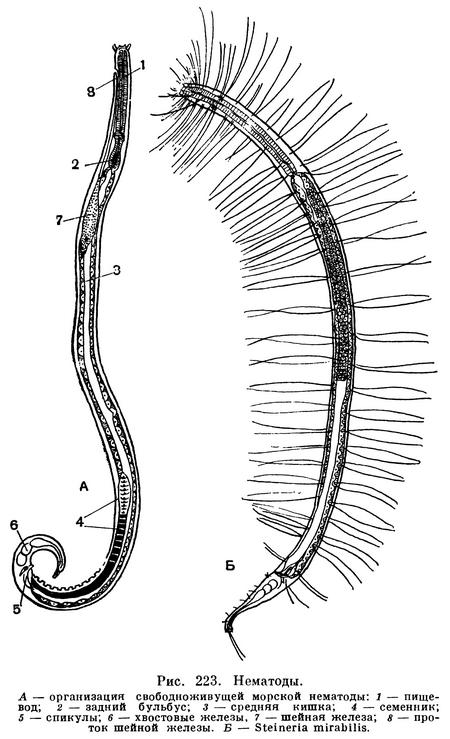
Каков же диаметр тела этих животных? Известный русский зоолог, автор многочисленных специальных работ и учебников и вместе с тем талантливый переводчик проф. Н. А. Холодковский называл нематод «струнцами». Теперь этот термин получил более узкое значение, но он все же хорош тем, что дает образное представление об общей форме тела нематод. Действительно, они длиннотелые и тонкие, как струна (рис. 223). При этом в поперечном сечении их вытянутое тонкое тело, как правило, образует правильный круг (рис. 220).
Мелкие формы часто уподобляются тонкой нити. Да и самое слово «нематода» произошло от греческого слова nemas, что означает нить.
Форме тела нематод соответствует и главный, типичный, способ перемещения нематод в пространстве: они движутся наподобие микроскопических или видимых и невооруженным глазом змей. Лежа всегда на боку, нематоды изгибаются в спинно-брюшной плоскости и перемещаются по дну водоемов, в тесных водных пленках почвы, в кишечнике и других органах человека и животных, меж клетками корней, стеблей, листьев и других частей растений.
Все тело нематод покрыто гибкой, эластичной и прочной кутикулой. Эта кутикула — производное лежащего под ней: тонкого слоя кожного эпителия, называемого у нематод гиподермой. Гиподерма — живая эпителиальная ткань, которая выделяет на своей поверхности кутикулу. Кутикула у нематод может быть гладкой или кольчатой, причем кольца построены совершенно правильно, все они у каждого вида определенного размера и часто несут различные уплотнения — склероции, имеющие форму правильно расположенных точек, линий (палочек), пластин и т. д. Гиподерма очень тонкая. Но по бокам тела, а также вдоль спины и брюха она утолщена, особенно по бокам, где образованы правый и левый гиподермальные валики, известные под названием «хорд» или полей (не имеющих, конечно, никакого отношения к хорде хордовых животных). Внутри боковых «хорд» у части нематод лежат правый и левый выделительные каналы. Кутикула и гиподерма составляют периферию кожно-мускульного мешка тела нематод. Под гиподермой расположена продольная мускулатура. Однако мышечный слой не сплошной. Он тянется вдоль тела в виде четырех мышечных тяжей — двух спинно-боковых и двух брюшно-боковых, отделенных друг от друга четырьмя упомянутыми «хордами» (рис. 223). Мышечные клетки удлинены и всегда расположены в одном направлении, что очень характерно для так называемых поляризованных клеточных компонентов ткани. В этих случаях длинные и перпендикулярные им оси клеток одинаково ориентированы в пределах всего тела. Поэтому все клетки мышц работают согласованно, синхронно, что естественно повышает их кинетическую энергию. Недаром тонкие нематоды легко проникают в узкие пространства между нитями водорослей, между гифами мицелия грибов, между частицами почвы, облеченными капиллярной пленкой воды, в поры тела животных, в устьица листьев, межклетники корневых, стеблевых и других тканей растений и т. п.
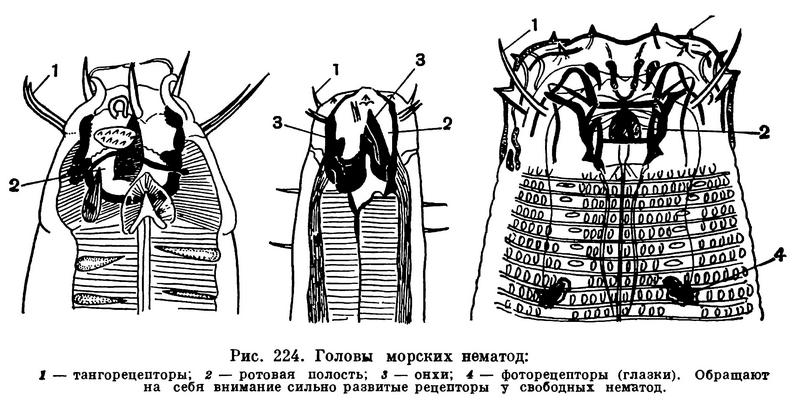
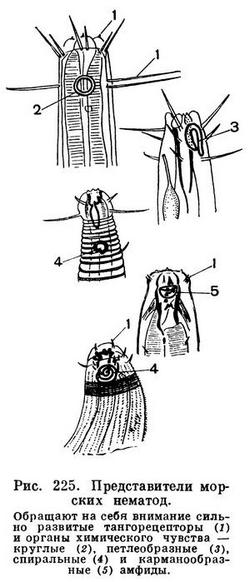
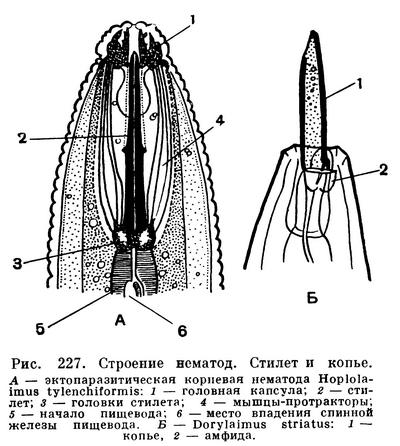
Головной конец тела нематод снабжен головной капсулой, опирающейся на внутренний опорный скелет из плотной кутикулы. Головная капсула состоит из двух основных частей — головных бугров и подвижных губ. Но у многих форм губы и головные бугры сливаются в общую головную капсулу. На ней расположены органы осязания — тангорецепторы, имеющие форму либо щетинок (рис. 224), либо сосочков, т. е. папилл. На переднем конце головной капсулы, строго посередине и лишь изредка сместившись несколько на брюшную сторону, лежит ротовое отверстие, окруженное губами. На головной капсуле, или сзади от нее, или на боковых губах лежат боковые обонятельные ямки, известные у нематод под названием боковых органов или амфид. От амфид отходят обонятельные нервы (рис. 225). У некоторых свободноживущих нематод развиты также глаза, снабженные у ряда форм линзой и глазным пигментированным бокалом — зеленым, оранжевым, фиолетовым, красным, черным (рис. 224). Иногда вдоль всего тела торчат щетинки (рис. 223).
Тело нематод очень ясно дифференцировано на три участка. Передний участок несет органы чувств, о которых сказано выше, и ему соответствует передний отрезок кишечника — передняя кишка. Второй отрезок тела соответствует средней кишке и включает, кроме нее, половые трубки. Третий — образует хвост, ограниченный на брюшной стороне тела заднепроходной щелью (анусом). Конец хвоста имеет у разных видов различную форму (рис. 223).
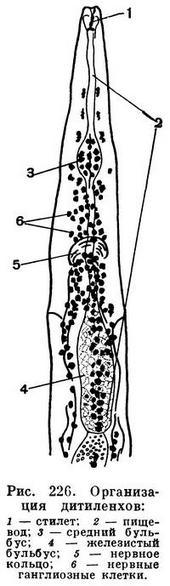
Центральная нервная система состоит из нервных стволов, расположенных вдоль тела, и кольцевых комиссур, связывающих продольные стволы в единую систему. У нематод наиболее типична кольцевая нервная комиссура, охватывающая пищевод. Она образует «нервное кольцо» нематод, которое не несет нервных ядер и состоит из неврофибрилл. Однако впереди от него и позади него, к пищеводу, расположена сложная система ганглиозных клеток (рис. 226). Именно эта система ганглиозных клеток в своей совокупности образует нечто подобное «мозгу» турбеллярий и гастротрих. В этом «мозгу» можно различить несколько ганглиев (рис. 226). От этих ганглиев вперед, к тангорецепторам и амфидам головы, отходят нервные тяжи. Кроме того, нервные ядра заложены в самой ткани пищевода; нервные ядра регулируют движение мускулатуры пищевода, работу зубов, копий и стилетов, которыми вооружены многие нематоды, и выделение экскретов пищеводных желез.
Пищеварительная система нематод сложнее, чем у форм предшествующих классов. Передняя кишка делится на ротовую полость, или стому, и пищевод. Хотя и стома и пищевод, строго говоря, представляют собой не что иное, как глотку, тем не менее в системе знаний о нематодах, или нематологии, утвердилась указанная номенклатура: стома, или ротовая полость, и пищевод. Для этого имеются солидные основания. Стома — это часть глотки, которая функционирует как ротовая полость и часто вооружена различными особо дифференцированными придатками, заслуживающими наименования органов. Пищевод — это часть глотки, способная к перистальтическим движениям, проталкивающим комок пищи в среднюю кишку (рис. 226). Поэтому нематологи (специалисты по нематодам) не называют стому и пищевод глоткой (фаринксом). Пусть правы сравнительные морфологи, что и стома и пищевод нематод — это фаринкс, глотка. Однако функционально это стома и пищевод. В стоме различают неподвижные придатки, или онхи, и подвижные зубы; у некоторых нематод имеются особо дифференцированные «челюсти», у других — острый сосущий стилет и, наконец, копье (рис. 227).
Средняя кишка такая же, как у гастротрих. Стенка ее состоит из одного слоя клеток. Задняя часть кишки переходит в прямую кишку, открывающуюся наружу уже упоминавшимся заднепроходным отверстием. Пищеварение у нематод своеобразное. В пищеводе лежат специальные железы, которые выделяют экскреты, содержащие ферменты. Эти ферменты либо поступают с пищей в среднюю кишку, где пища переваривается, либо выделяются наружу, и тогда возникает своеобразный процесс переваривания пищи в наружной среде, в капле ферментов нематоды, после чего быстро переваренная пища попадает в просвет стомы и пищевода и усваивается в кишке.
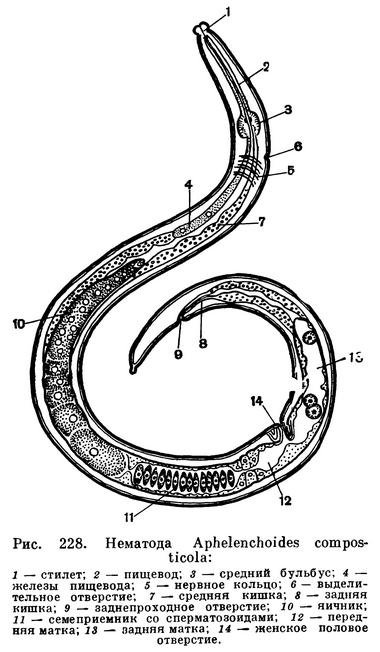
Выделительная система нематод бывает двух типов. У одних форм она состоит всего лишь из одной шейной железистой клетки, проток которой открывается наружу брюшной порой. У других, кроме этой шейной железы, имеются боковые выделительные каналы. Содержимое их выделяется наружу через брюшную выделительную пору (рис. 228). Мы не будем вдаваться в подробности процессов выделения. Отметим, что продукты обмена проникают в полостную жидкость. Здесь они с помощью особых клеточных систем обезвреживаются, диффундируют в шейную железу и выделяются наружу.
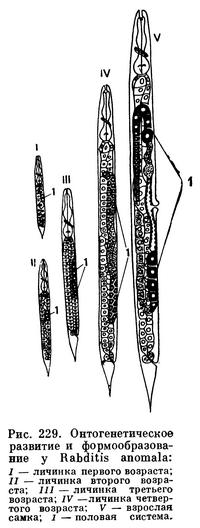
Все нематоды, как правило, раздельнополые животные. У самцов развиты семенники, семяпроводы и семяизвергательный канал. Семенников может быть два или один. Кроме того, у самцов имеются специальные совокупительные органы — спикулы и рулек, направляющий их движения. Женские половые органы состоят из яичников, яйцепроводов и матки. Женское половое отверстие расположено на брюшной стороне тела. Самцы вводят спикулы в женское половое отверстие и оплодотворяют самок. Сперма нематод не имеет подвижных жгутиков. Ни в одном органе нематод вообще нет подвижных клеточных органелл, и в частности ресничек. Нет, как выше сказано, и хвостов у спермы. Она перемещается амебоидными движениями. В половых путях самок формируются яйца. Они оплодотворяются спермой самцов в женских половых путях, и в частности в особых семеприемниках (рис. 228). Затем оплодотворенные яйца выделяются наружу через женское половое отверстие или же развиваются внутри половых трубок. В этом случае из женского полового отверстия выходят наружу личинки (живорождение). Яйца нематод заключены в яйцевые оболочки, предохраняющие их от физических повреждений и химических воздействий среды. Личинки четыре раза линяют, последовательно, после каждой линьки переходят в следующую стадию развития, превращаясь в личинок второго, третьего и четвертого возрастов. Из личинки четвертого возраста развиваются молодые формы — самец или самка (рис. 229). Очень часто личинки не похожи на взрослые формы. В таких случаях нематологи говорят о развитии с превращением.
Таковы самые общие представления об организации нематод. Во многом их организация сходна с организацией гастротрих, киноринх и коловраток. Однако нематоды существенно отличаются от любых представителей этих групп следующими важными признаками: формой тела; способом движения; отсутствием протонефридий; отсутствием в любой системе органов ресничного эпителия и подвижных жгутов в каких бы то ни было клетках, в том числе половых; четкой половой дифференцировкой (самцы и самки), несвойственной по крайней мере гастротрихам; большим числом видов и их групп и ярко выраженными признаками биологического прогресса. Как и у других групп немательминтов, у нематод нет ни органов дыхания, ни кровообращения.
Систематика нематод. Это, пожалуй, один из наиболее сложных вопросов.
Нематоды — это непрерывно расширяющийся поток вновь открываемых видов и родов. Приведем пример этого. В 1949 г. числилось 200 видов нематод из отряда тиленхид, среди которых много паразитов растений. Американский исследователь этих нематод Торн (Thorne, 1949) указывал, что это число, вероятно, составляет 5% числа видов названного отряда, в действительности живущих в почве и в растениях. К 1962 г. число их достигло 800 видов, т. е. с 1949 по 1961 г. было вновь открыто 600 видов тиленхид.
Большинством ученых класс нематод делится на два подкласса — подкласс Аденофореи (Adenophorea) и подкласс Сецерненты (Secernentea).
Главная масса аденофореи — жители бентоса морей и океанов; многие (хотя и меньшинство из них) обитают в почве и в пресных водах. Известны среди них сравнительно немногие группы, приспособленные к паразитизму в органах растений и животных. Сецерненты, напротив, в большинстве приспособлены к паразитическому существованию в тканях и в органах растений и животных.
Остановимся на некоторых важнейших группах этих подклассов.
ПОДКЛАСС АДЕНОФОРЕИ (ADENOPHOREA)
Большинство аденофореи, как сказано выше, живет в морях и океанах. Часть из них обитает в пресных водах и в почве. Некоторые группы представлены паразитами растений и животных.
Большое число видов различных родов и семейств аденофорей ведет свободное существование на дне водоемов, т. е. в бентосе. Это типичные нематоды, с наиболее полной и примитивной организацией. Среди них множество очень мелких форм, длина тела которых равна долям миллиметра. Упомянутая выше Trichoderma minutum принадлежит к этому подклассу и обитает в море. Имеются среди аденофорей и относительно крупные формы. Их длина измеряется не миллиметрами, а сантиметрами, нередко десятками сантиметров. Но таких все же меньше. Основная масса аденофорей представлена более мелкими формами, достигающими длины 1—5 мм.
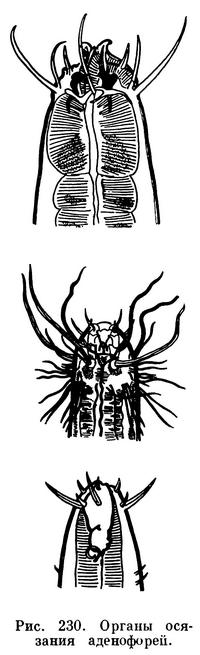
Какими же признаками характеризуется подкласс аденофорей, его многочисленные свободноживущие представители? Первое — прогрессивным развитием органов чувств. На рисунке 230 показаны головные концы нескольких форм, у которых хорошо заметно прежде всего мощное развитие органов осязания, которые у морских аденофорей обычно имеют форму длинных щетинок. Каждая щетинка состоит из кутикулярного чехла, по оси которого располагается нерв (тангорецепторы). Таких тангорецепторов может быть четыре, шесть, десять, двенадцать. Очень часто по одному тангорецептору расположено по бокам головной капсулы — один справа, другой слева; кроме того, по паре рецепторов — по субвентральным и субдорсальным секторам головы (рис. 230). Всего, следовательно, шесть рецепторов. Однако это число у других форм либо возрастает до десяти (если и по бокам головы развито по два рецептора), либо падает до четырех. Развитие тангорецепторов в виде длинных щетинок очень характерно именно для свободных аденофорей. Это прямое следствие и приспособления к свободному существованию, в условиях которого подвижная морская нематода сталкивается со многими другими донными беспозвоночными, в том числе и с хищными нематодами.
ПОДКЛАСС СЕЦЕРНЕНТЫ (SECERNENTEA)
Не случайно знакомство с подклассом сецернентов мы начинаем во вторую очередь. Аденофореи — свободные формы моря — типичные нематоды. Подкласс сецернентов включает множество групп нематод, на организации которых лежит печать специализации.
В пределах отряда хромадорид, проникших в пресные воды и в почву, развивается семейство плектид (Plectidae), представители которого проникли в сапробиотические очаги, где плектиды нашли готовый «полуфабрикат» — доступный источник их питания. Когда возникло семейство плектид, неизвестно, так как фактически нематоды «не оставили» каких-либо палеонтологических документов. Но во всяком случае, они должны были возникнуть тогда, когда уже существовали растительные организмы и когда в почве началось накопление органического материала.
Мы здесь напомнили о семействе Plectidae потому, что современная наука установила наличие черт сходства между Plectidae, с одной стороны, и формами семейства Rhabditidae — с другой. Интерес этого вопроса связан с тем, что семейство Rhabditidae принадлежит не к подклассу Adenophorea, с которым выше мы ознакомили читателя, а к другому подклассу нематод — Secernentea. Таким образом, оказывается, что эти два семейства — Plectidae и Rhabditidae — связывают узами родства оба названных подкласса нематод, а это восполняет наши представления об эволюции класса нематод.
Первый, наиболее характерный признак представителей семейства рабдитид (Rhabditidae) — небольшие размеры форм. Длина их тела около 1 мм, а часто и меньше. Вторым внешним признаком этих сапробиотических нематод назовем свойственную им веретеновидную форму тела: к голове и к хвосту тело рабдитид обычно сужено, в среднем отделе заметно расширено. Голова, как правило, несет осязательные сосочки (папиллы), а не щетинки, как у плектид. Амфиды, или боковые органы, всегда расположены на передней плоскости головы, т. е. на губах. Ротовая полость имеет форму удлиненного цилиндра (рис. 229). В глубинном отделе этого цилиндра видны «глоточные бугры». В суженной части ротовой полости, лежащей между вдающимися в нее упомянутыми выше «глоточными буграми», у большинства представителей семейства рабдитид расположены мелкие зубчики, названные онхами.
Цилиндрическая ротовая полость рабдитид гладкостенная и достаточно широкая, чтобы легко пропустить комочек сапробиотического субстрата, который обычно глотают рабдитиды. Пройдя сквозь ротовую полость, комочек пищи попадает в просвет пищевода. Стенки его мускулисты и несут обычно два утолщения, известные под названием бульбусов. Один из них средний, а другой — задний. Задний бульбус обладает усиленной мускулатурой и особым внутренним дробильным аппаратом, который раздробляет проглоченный комок пищи (рис. 229). За пищеводом следует средняя кишка, переходящая в заднюю кишку. Последняя открывается наружу заднепроходным отверстием, лежащим, как у всех нематод, на брюшной стороне тела, в основании хвоста.
Таковы основные особенности организации рабдитид, которые повторяются в различных не слишком резких вариантах у более чем 240 видов этого семейства.