(Secernentea) Класс Сецерненты, Class Secernentea 9 отрядов
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
Систематика нематод. Это, пожалуй, один из наиболее сложных вопросов.
Нематоды — это непрерывно расширяющийся поток вновь открываемых видов и родов. Приведем пример этого. В 1949 г. числилось 200 видов нематод из отряда тиленхид, среди которых много паразитов растений. Американский исследователь этих нематод Торн (Thorne, 1949) указывал, что это число, вероятно, составляет 5% числа видов названного отряда, в действительности живущих в почве и в растениях. К 1962 г. число их достигло 800 видов, т. е. с 1949 по 1961 г. было вновь открыто 600 видов тиленхид.
Большинством ученых класс нематод делится на два подкласса — подкласс Аденофореи (Adenophorea) и подкласс Сецерненты (Secernentea).
Главная масса аденофореи — жители бентоса морей и океанов; многие (хотя и меньшинство из них) обитают в почве и в пресных водах. Известны среди них сравнительно немногие группы, приспособленные к паразитизму в органах растений и животных. Сецерненты, напротив, в большинстве приспособлены к паразитическому существованию в тканях и в органах растений и животных.
Остановимся на некоторых важнейших группах этих подклассов.
ПОДКЛАСС СЕЦЕРНЕНТЫ (SECERNENTEA)
Не случайно знакомство с подклассом сецернентов мы начинаем во вторую очередь. Аденофореи — свободные формы моря — типичные нематоды. Подкласс сецернентов включает множество групп нематод, на организации которых лежит печать специализации.
В пределах отряда хромадорид, проникших в пресные воды и в почву, развивается семейство плектид (Plectidae), представители которого проникли в сапробиотические очаги, где плектиды нашли готовый «полуфабрикат» — доступный источник их питания. Когда возникло семейство плектид, неизвестно, так как фактически нематоды «не оставили» каких-либо палеонтологических документов. Но во всяком случае, они должны были возникнуть тогда, когда уже существовали растительные организмы и когда в почве началось накопление органического материала. Фактически это очень древний процесс. Как указывает выдающийся советский ученый акад. В. И. Вернадский (1926), «чем более мы изучаем химические явления биосферы, тем более мы убеждаемся, что в ней нет случаев, где они были бы независимы от жизни... И так, — продолжает акад. Вернадский, — длилось в течение всей геологической истории». Нам приблизительно известно, когда на Земле появились грибы, и в частности низшие, гифомицеты. Вероятно, они появились еще в каменноугольном периоде. Бактерии появились много раньше. Но сапробиотические очаги, возникающие в результате загнивания трупов растений, появились в почве уже тогда, когда развилась пышная наземная флора каменноугольного периода. Весьма вероятно, что именно к этому времени, ввиду сказанного выше, начинается становление семейства плектид в его современном облике, т. е. в виде форм, приспособленных к питанию за счет продуктов распада белков и растворимых сахаров, содержащихся в сапробиотических очагах. В водоемах процесс мог возникнуть и раньше.
Мы здесь напомнили о семействе Plectidae потому, что современная наука установила наличие черт сходства между Plectidae, с одной стороны, и формами семейства Rhabditidae — с другой. Интерес этого вопроса связан с тем, что семейство Rhabditidae принадлежит не к подклассу Adenophorea, с которым выше мы ознакомили читателя, а к другому подклассу нематод — Secernentea. Таким образом, оказывается, что эти два семейства — Plectidae и Rhabditidae — связывают узами родства оба названных подкласса нематод, а это восполняет наши представления об эволюции класса нематод.
Но семейство Rhabditidae глубоко интересно и независимо от вопросов эволюции развития класса нематод.
Сапробиос. Что такое сапробиотическая среда? Прежде всего это сложное биологическое явление, сложный биологический процесс, связанный с биологическими циклами азота, углерода и минеральных элементов. Циклы названных элементов обусловлены деятельностью организмов, с одной стороны, синтезирующих сложные органические соединения, характерные для живых форм (белки, углеводы, жиры, пектин, клетчатку и т. д.), и, с другой стороны, возвращающих продукты распада этих веществ в почву в результате гнилостного распада тканей растений и животных. Синтетические процессы всегда связаны с процессами разрушения, распада, и обе эти, диалектически связанные, стороны жизненных процессов присущи прежде всего почве как биологическому явлению. Распад органического материала, возникающий при разложении трупов животных и растений, и составляет основу характеристики сапробиотической среды. В почве сапробиотическая среда возникает в форме отдельных сапробиотических очагов. В конце вегетации культурных растений нередко можно наблюдать, такие очаги после сбора урожая сельскохозяйственных культур. Так, на свекольном поле, особенно после дождей, можно найти разлагающиеся листья свеклы, причем особенно ясно следы сапробиотического распада видны на нижней стороне листьев, обращенной к почве. Если взять каплю распадающегося материала (от листа) и рассмотреть ее под микроскопом, можно легко убедиться в том, что в этой капле видны различные одноклеточные организмы, в том числе бактерии, низшие грибы, простейшие и многочисленные энергично двигающиеся нематоды. Все эти организмы всегда весьма специфичны, т. е. образуют группы, встречающиеся только в данной гнилостной среде. Поэтому их обычно называют сапрофитными бактериями, сапрофитными грибами, сапрозойными нематодами и т. д. Все эти организмы играют определенную роль в сложных процессах распада органического материала и составляют действующие факторы сапробиотической среды. Их деятельность обеспечивает возвращение в почву элементов, за счет которых синтезируются живые ткани животных и растений. На языке биохимика этот сложный процесс созидания и разрушения выражается в циклах главных элементов, участвующих в синтезе и распаде органических соединений, присущих живым организмам и определяющих само понятие о жизни.
Таким образом, сапробиотическая среда есть процесс распада органического материала, причем в этом распаде принимают участие и даже неизбежно обеспечивают его специфические, приспособленные к жизни в сапробиотической среде организмы, в первую очередь бактерии и грибы. Эти организмы выделяют специфические ферменты, обеспечивающие распад органических соединений — белков, углеводов, клетчатки и др. — на более элементарные соединения.
Характерный признак сапробиотической среды — ее динамичность. Она всегда существует в ее изменениях и превращениях, конечным звеном которых является минерализация органического материала — возвращение бывших живых тел в почву. Сапробиотические процессы всегда происходят в почве, и в ней живут организмы, в том числе и нематоды, которые так или иначе участвуют и в этих сложных процессах распада органического материала. Эти сапробиотические нематоды — корень всех сецернентов, всего этого подкласса.
Все основные группы сапробиотических нематод принадлежат к отряду Rhabditida. В составе этого отряда очень много различных групп — от сапробиотических и до специализированных паразитов. Среди представителей подкласса сецернентов очень много паразитов животных, человека и растений — гораздо больше, чем в пределах подкласса аденофорей. Причина этого заключается в том, что в пределах подкласса аденофорей нет ни одного семейства, которое было бы так тесно связано с сапробиотической средой, как семейство рабдитид. Правда, читатель познакомился с семейством плектид, принадлежащим к аденофореям. Но плектиды — источник развития рабдитид, а не каких-либо аденофорей.
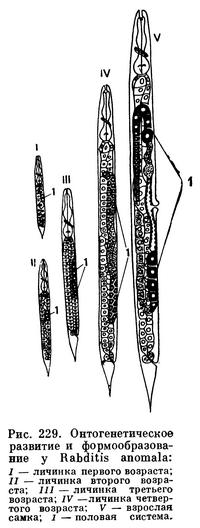
Первый, наиболее характерный признак представителей семейства рабдитид (Rhabditidae) — небольшие размеры форм. Длина их тела около 1 мм, а часто и меньше. Вторым внешним признаком этих сапробиотических нематод назовем свойственную им веретеновидную форму тела: к голове и к хвосту тело рабдитид обычно сужено, в среднем отделе заметно расширено. Голова, как правило, несет осязательные сосочки (папиллы), а не щетинки, как у плектид. Амфиды, или боковые органы, всегда расположены на передней плоскости головы, т. е. на губах. Ротовая полость имеет форму удлиненного цилиндра (рис. 229). В глубинном отделе этого цилиндра видны «глоточные бугры». В суженной части ротовой полости, лежащей между вдающимися в нее упомянутыми выше «глоточными буграми», у большинства представителей семейства рабдитид расположены мелкие зубчики, названные онхами.
Цилиндрическая ротовая полость рабдитид гладкостенная и достаточно широкая, чтобы легко пропустить комочек сапробиотического субстрата, который обычно глотают рабдитиды. Пройдя сквозь ротовую полость, комочек пищи попадает в просвет пищевода. Стенки его мускулисты и несут обычно два утолщения, известные под названием бульбусов. Один из них средний, а другой — задний. Задний бульбус обладает усиленной мускулатурой и особым внутренним дробильным аппаратом, который раздробляет проглоченный комок пищи (рис. 229). За пищеводом следует средняя кишка, переходящая в заднюю кишку. Последняя открывается наружу заднепроходным отверстием, лежащим, как у всех нематод, на брюшной стороне тела, в основании хвоста.
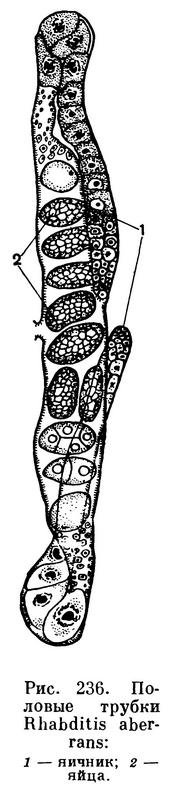
Половые органы самок хорошо развиты. Оба яичника (бывает и один) содержат большое число зародышевых половых клеток — овогониев, за счет которых формируются яйца (рис. 236). Самцы имеют один семенник, удлиненный семяпровод, за которым следует семяизвергательный канал, впадающий в заднюю кишку. В ней же лежат выдвижные совокупительные органы самца — спикулы. Самым замечательным органом самца, несомненно, являются хвостовые бурсальные крылья. Одно из них лежит справа, а другое слева, начинаясь несколько впереди от хвоста и часто достигая его кончика.
Таковы основные особенности организации рабдитид, которые повторяются в различных не слишком резких вариантах у более чем 240 видов этого семейства. Мы не случайно остановились на описании организации рабдитид. Задача, которую мы должны теперь решить, состоит в том, чтобы попытаться объяснить, почему сформировались описанные выше признаки.
Оказывается, что все перечисленные признаки легко и естественно объясняются тем, что они полностью отвечают основным особенностям сапробиотической среды. К тем признакам этой среды, которые выше были описаны, надо присоединить еще один: сапробиотическая среда, как правило, представлена не крупными площадями почвы, но значительно чаще отдельными сапробиотическими очагами. Каждый кусочек органического материала, попавший на землю, каждый лист, обрывок стебля и т. п. становится причиной развития маленького сапробиотического очага. В почве могут быть, и фактически имеются, большие количества таких отдельных, взаимно изолированных сапробиотических очагов, и в каждом из них под влиянием бактерий и грибов в первую очередь развивается сапробиотический процесс. Здесь же, в этом крохотном подчас очажке, в почве покоится, быть может, несколько крохотных личинок рабдитид, покрытых плотной шкуркой. Они ждут возникновения сапробиотического очага. Когда гнилостные, сапрофитные бактерии начинают свою разрушительную работу и когда начинается формирование и развитие сапробиотического очага в почве, этот процесс становится стимулом к развитию личинок рабдитид. Быстро развиваясь, рабдитиды превращаются в самок и самцов, и начинается быстротечная, стремительная жизнь.
С биологической точки зрения, если воспользоваться спортивной терминологией, можно сказать, что рабдитиды настоящие спринтеры, но не в смысле быстроты движения, а в смысле необычайной быстроты развития.
Известны виды рабдитид, весь цикл индивидуального развития которых — от яйца до взрослой плодущей самки — протекает всего лишь за несколько часов. Сроки, равные примерно 12 или 24 часам, — обычное явление. Если маленький сапробиотический очажок просуществовал, скажем, 20 дней, за этот срок в нем перебывает до 10—15 видов рабдитид, которые примерно через каждые 2—3 дня сменяют друг друга. Эта быстрая смена видов просто поразительна. Немецкая исследовательница Райтер (1928) наблюдала картину смены видов в одном из своих опытов. На третий день в экспериментальном сапробиотическом субстрате содержались тысячи особей Rhabditis inermis, от нескольких дюжин до сотен Rhabditis elongata и еще три других вида. Через два дня первый из названных видов угас, три других размножились в тысячах экземпляров. Еще через три дня картина снова изменилась и в массовых количествах появился один из видов, ранее малочисленный. Таким образом, восьми дней оказалось достаточно для развития в небольшом экспериментально организованном «микрокосмосе» огромных количеств рабдитид, исчисляемых многими тысячами, возникших из ограниченного числа личинок, имевшихся в комочке почвы, использованной для опыта. Наблюдая за этими тысячами рабдитид под покровным стеклом, можно заметить, что большинство этих маленьких животных находится в состоянии почти неутомимого движения. При более сильном увеличении микроскопа видно, что все они очень часто работают средним бульбусом своего пищевода и «дробильным» аппаратом второго, кардиального, бульбуса и заглатывают массу пищи, состоящей из мелких органических частиц сапробиоса — сапробиотического детрита, бактерий и спор грибов. При сильном увеличении мы обнаружим, что на хвосте рабдитид оседают целые колонии бактерий сапробиотической среды. Кишечник часто заполнен, и нередко можно видеть в нем темные массы пищи. Однако эта пища содержит много воды; концентрация растворенных в ней продуктов расщепления белков, сахаров и других веществ невелика, и рабдитиды должны энергично глотать пищу, чтобы обеспечить не только свое питание, но и «спринтерское» развитие яиц. А ведь каждая самка, едва достигающая 1 мм в длину, дает за свою жизнь, измеряемую днями, до 250—300 яиц, из которых за 1—2 дня развиваются новые самки и самцы. Пища малопитательна, ее нужно много, и поэтому рабдитиды должны непрерывно двигаться, чтобы всегда иметь эту пищу около себя. Вот почему рабдитиды почти непрерывно поглощают пищу и столь же непрерывно двигаются. При малом увеличении микроскопа поле наблюдения буквально мерцает — все оно в динамике, в движении от множества движущихся маленьких змеевидных животных! Здесь возникает стремительный оборот питания, усвоения пищи, выделения отбросов, стремительный рост зародышевых половых клеток, «спринтерские» скорости развития, смерти и новых рождений. Самка рабдитид находится в непрерывном движении. Даже когда самец осеменяет ее, она все-таки хватает пищу и, извиваясь и суетясь, движется вперед или в стороны под покровным стеклом в свете лучей микроскопа (или в темноте). Вот почему самцу и нужны бурсальные крылья. Он прилаживает их плотно к телу самки и выполняет свою жизненную функцию все в той же симфонии движения!
Как видим, вся организация рабдитид отвечает условиям сапробиотической среды. Эта среда быстротечна, так как в ней под влиянием различных групп сапробиотических бактерий идет распад органического материала и каждый день меняется химическая характеристика сапробиоса. Этим объясняется столь быстрое развитие рабдитид. И именно поэтому они столь малых размеров — малые размеры упрощают проблему быстрого развития. Рабдитидам нужно все время питаться, так как пища непитательна; надо накопить огромное поколение, обеспечивающее будущее, т. е. жизнь личинок, которые останутся в почве в ожидании возникновения нового сапробиотического очага, когда вновь возобновится эта феерическая смена поколений. Вся организация рабдитид подчинена этому требованию сапробиотической среды.
Кинооператор мог бы сделать изумительную картину, рисующую эту неисчерпаемую энергию жизни рабдитид и их организации, во всех деталях приспособленную к стремительной феерии реальной жизни!
Таковы рабдитиды. Подчиняясь ритму распада органического материала, они участвуют в великом процессе возвращения почве азота, углерода, серы и других химических элементов. Под «руководством» великих химиков природы — бактерий сапробиотической среды и с помощью низших грибов — обладателей мощных ферментов сапробиотические рабдитиды участвуют в процессах конечной минерализации органического материала, возвращая его почве и становясь тем самым одним из источников новых бесчисленных поколений живых форм. Так погибшие организмы, через распад, становятся ступенью к новой жизни.
Какая красота и какой оптимизм жизни в этом творческом круговороте веществ!
В настоящее время можно с известной уверенностью предполагать, что рабдитиды произошли от древних плектид. Основным фактором развития рабдитид была сапробиотическая среда, в которой в результате влияния естественного отбора, открытого Чарлзом Дарвином (см. Ч.Дарвин, Происхождение видов или К. А. Тимирязев, Чарлз Дарвин и его учение), рабдитиды и приобрели свои особенности организации, столь полно отвечающие условиям сапробиоса.
Рабдитиды сыграли важную роль в эволюции группы нематод, с которой человек вынужден был серьезно считаться.