Царство Археи (Archaea Woese, Kandler & Wheelis, 1990), Regnum Archaea Woese, Kandler & Wheelis, 1990 2 типа
Синонимы:
• Archaebacteria
Типы:
• Кренархеоты (Crenarchaeota)
• Эвриархеоты (Euryarchaeota)
• Корархеоты (Korarchaeota)
• Наноархеоты (Nanoarchaeota)
• Thaumarchaeota
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Археи (лат. Archaea от др.-греч. «извечный, древний, первозданный, старый») — домен живых организмов (по трёхдоменной системе Карла Вёзе наряду с бактериями и эукариотами). Археи представляют собой одноклеточные микроорганизмы, не имеющие ядра, а также каких-либо мембранных органелл.
Ранее археи объединяли с бактериями в общую группу, называемую прокариоты (или царство Дробянки (лат. Monera)), и они назывались архебактерии, однако сейчас такая классификация считается устаревшей: установлено, что археи имеют свою независимую эволюционную историю и характеризуются многими биохимическими особенностями, отличающими их от других форм жизни.
В настоящее время археи подразделяют на 5 типов. Из этих групп наиболее изученными являются кренархеоты (лат. Crenarchaeota) и эвриархеоты (лат. Euryarchaeota). Классифицировать археи по-прежнему сложно, так как подавляющее большинство из них никогда не выращивались в лабораторных условиях и были идентифицированы только по анализу нуклеиновых кислот из проб, полученных из мест их обитания.
Археи и бактерии очень похожи по размеру и форме клеток, хотя некоторые археи имеют довольно необычную форму, например, клетки Haloquadratum walsbyi плоские и квадратные. Несмотря на внешнее сходство с бактериями, некоторые гены и метаболические пути архей сближают их с эукариотами (в частности ферменты, катализирующие процессы транскрипции и трансляции). Другие аспекты биохимии архей являются уникальными, к примеру, присутствие в клеточных мембранах липидов, содержащих простую эфирную связь. Большая часть архей — хемоавтотрофы. Они используют значительно больше источников энергии, чем эукариоты: начиная от обыкновенных органических соединений, таких как сахара, и заканчивая аммиаком, ионами металлов и даже водородом. Солеустойчивые археи — галоархеи (лат. Haloarchaea) — используют в качестве источника энергии солнечный свет, другие виды архей фиксируют углерод, однако, в отличие от растений и цианобактерий (синезелёных водорослей), ни один вид архей не делает и то, и другое одновременно. Размножение у архей бесполое: бинарное деление, фрагментация и почкование. В отличие от бактерий и эукариот, ни один известный вид архей не формирует спор.
Изначально археи считали экстремофилами, живущими в суровых условиях, таких как горячие источники и солёные озёра, однако потом они были обнаружены в самых различных местах, включая почву, океаны, болота и толстую кишку человека. Архей особенно много в океанах, и, возможно, планктонные археи являются самой многочисленной группой ныне живущих организмов. В наше время археи признаны важной составляющей жизни на Земле и играют роль в круговоротах углерода и азота. Ни один из известных представителей архей не является паразитом или патогенным организмом, однако часто они бывают мутуалистами и комменсалами. Некоторые представители являются метаногенами и обитают в пищеварительном тракте человека и жвачных, где очень многочисленны и помогают осуществлять пищеварение. Метаногены используются в производстве биогаза и очистке канализационных сточных вод, а ферменты экстремофильных микроорганизмов, сохраняющие активность при высоких температурах и в контакте с органическими растворителями, находят своё применение в биотехнологии.
История открытия.
Первые представители группы были обнаружены в различных экстремальных средах обитания, например геотермальных источниках.
На протяжении большей части XX века прокариоты считались единой группой и классифицировались по биохимическим, морфологическим и метаболическим особенностям. К примеру, микробиологи пытались классифицировать микроорганизмы в зависимости от формы клеток, деталей строения клеточной стенки и потребляемых микроорганизмами веществ. В 1965 году было предложено устанавливать степень родства разных прокариот на основании сходства строения их генов. Этот подход, филогенетика, в наши дни является основным.
Впервые археи были выделены в качестве отдельной группы прокариот на филогенетическом древе в 1977 году Карлом Вёзе и Джорджем Эдвардом Фоксом при сравнительном анализе 16S рРНК. Изначально эти две группы были обозначены как архебактерии (лат. Archaebacteria) и эубактерии (лат. Eubacteria) и рассматривались как царства или подцарства, которые Вёзе и Фокс называли термином Urkingdoms. Вёзе утверждал, что эта группа прокариот есть фундаментально отличный тип жизни. Чтобы подчеркнуть это отличие, впоследствии две группы прокариот были названы археями и бактериями. В трёхдоменной системе Карла Вёзе обе эти группы и эукариоты были возведены в ранг домена. Этот термин был предложен Вёзе в 1990 году для обозначения самого верхнего ранга в классификации организмов, включающей одно или несколько царств.
В первое время к новому домену причисляли только метаногенные микроорганизмы. Считалось, что археи населяют только места с экстремальными условиями: горячие источники, солёные озёра. Тем не менее, к концу XX века микробиологи пришли в выводу, что археи являются большой и разнообразной группой организмов, широко распространённой в природе, вполне обычной и для менее экстремальных сред обитания, например, для почвы или вод океана. Причиной такой переоценки стало применение метода полимеразной цепной реакции для идентификации прокариот в образцах воды и почвы по их нуклеиновым кислотам. Данный метод позволяет выявлять и идентифицировать организмы, которые по тем или иным причинам не культивируются в лабораторных условиях.
Происхождение и эволюция.
Хотя возможные окаменелости прокариотических клеток датированы возрастом в 3,5 млрд лет, большинство прокариот не имеет характерных морфологических особенностей, и поэтому окаменелые формы нельзя идентифицировать как останки архей. В то же время химические остатки уникальных для архей липидов более информативны, так как эти соединения у других организмов не встречаются. В некоторых публикациях указывается, что останки липидов архей или эукариот присутствуют в породах возрастом 2,7 млрд лет, однако достоверность этих данных остаётся под сомнением. Эти липиды были также обнаружены в докембрийских формациях. Древнейшие из подобных остатков были найдены в Исуанском зеленокаменном поясе на западе Гренландии, где находятся самые старые на Земле осадочные породы, сформировавшиеся 3,8 млрд лет назад. Археи могут быть древнейшими живыми существами, населяющими Землю.
Вёзе утверждал, что археи, бактерии и эукариоты представляют собой три раздельные линии, рано отделившиеся от общей предковой группы организмов. Возможно, это произошло ещё до клеточной эволюции, когда отсутствие типичной клеточной мембраны давало возможности к неограниченному горизонтальному переносу генов, и предки трёх доменов различались между собой по фиксируемым комплектам генов. Не исключено, что последний общий предок архей и бактерий был термофилом, это даёт основания предположить, что низкие температуры были «экстремальной средой» для архей, и организмы, приспособившиеся к ним, появились только позже. Сейчас археи и бактерии связаны между собой не больше, чем с эукариотами, и термин «прокариоты» обозначает лишь «не эукариоты», что ограничивает его применимость.
Сравнительная характеристика архей и других доменов.
В приведённой таблице показаны некоторые главные черты архей, свойственные или не свойственные другим доменам, чтобы показать их родственные связи. Многие из этих свойств также обсуждаются ниже.
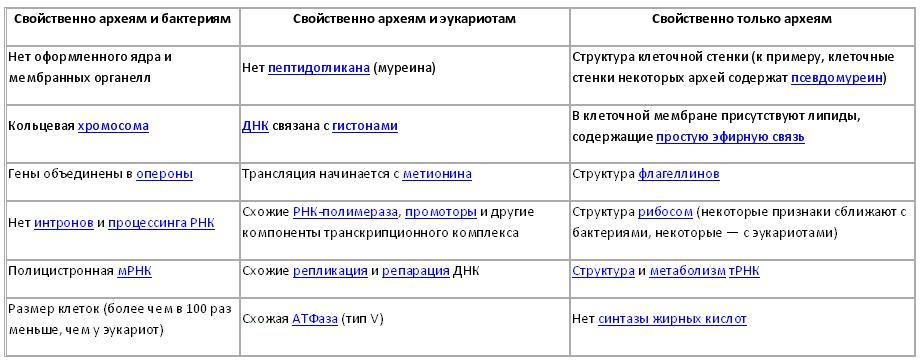
Родство с другими прокариотами.
Установление степени родства между тремя доменами имеет ключевое значение для понимания возникновения жизни. Большинство метаболических путей, в которых задействована большая часть генов организма, схожи у бактерий и архей, в то время как гены, отвечающие за экспрессию других генов, очень похожи у архей и эукариот. По строению клеток археи наиболее близки к грамположительным бактериям: клетка покрыта единственной плазматической мембраной, дополнительная внешняя мембрана, характерная для грамотрицательных бактерий отсутствует, клеточные стенки различного химического состава, как правило, толстые. В филогенетическом древе, основанном на сравнительном анализе структур гомологичных генов/белков прокариот, гомологи архей наиболее близки к таковым грамположительных бактерий. В некоторых важнейших белках архей и грамположительных бактерий, таких как Hsp70 и глутаминовая синтетаза I, обнаруживаются одинаковые эволюционно консервативные вставки и делеции.
Гупта (англ. Gupta) предположил, что археи отделились от грамположительных бактерий в результате отбора по признаку устойчивости к действию антибиотиков. Это основывается на наблюдении, что археи устойчивы к огромному числу антибиотиков, в основном производимых грамположительными бактериями, и что эти антибиотики действуют главным образом на гены, которые отличают бактерий от архей. Согласно гипотезе Гупты, давление отбора в направлении формирования устойчивости к антибиотикам грамположительных бактерий в конце концов привело к существенным изменениям в структуре генов-мишеней антибиотиков у некоторых микроорганизмов, которые стали общими предками современных архей. Предполагаемая эволюция архей под действием антибиотиков и других неблагоприятных факторов также может объяснить их адаптацию к экстремальным условиям, таким как повышенные температура и кислотность, как результат поиска ниш, свободных от продуцирующих антибиотики организмов. Кавалир-Смит (англ. Cavalier-Smith) выдвинул схожее предположение. Версия Гупты также подтверждается другими работами, изучающими родственные черты в структурах белков, и исследованиями, показавшими, что грамположительные бактерии могли быть первой ветвью, отделившейся от общего древа прокариот.
Родство с эукариотами.
Эволюционное родство между археями и эукариотами остаётся неясным. Помимо сходства в структуре и функциях клеток, между ними существуют сходство на генетическом уровне. Установлено, что группа архей кренархеоты стоят ближе к эукариотам, чем к другому типу архей — эвриархеотам. Кроме того, у некоторых бактерий, как Thermotoga maritima обнаружены архееподобные гены, переданные путём горизонтального переноса. Наиболее распространена гипотеза, согласно которой предок эукариот рано отделился от архей, а эукариоты возникли в результате слияния археи и эубактерии, ставших цитоплазмой и ядром новой клетки. Эта гипотеза объясняет различные генетические сходства, но сталкивается с трудностями в объяснении клеточной структуры.
Строение.
Форма клеток и колоний.
Отдельные клетки архей достигают от 0,1 до 15 мкм в диаметре и могут иметь различную форму: шара, палочки, спирали или диска. Некоторые кренархеоты имеют другую форму, например, Sulfolobus — неправильной дольчатой формы; Thermofilum — тонкой нитевидной формы и меньше 1 мкм в диаметре, а Thermoproteus и Pyrobaculum почти идеально прямоугольные. Haloquadratum walsbyi — плоские квадратные археи, живущие в сверхсолёных водоёмах. Такие необычные формы клеток, вероятно, обеспечиваются клеточной стенкой и прокариотическим цитоскелетом. У архей обнаружены белки, родственные компонентам цитоскелета других организмов, а также показано присутствие филаментов в их клетках, однако у архей, в отличие от других организмов, эти структуры плохо изучены. У Thermoplasma и Ferroplasma клеточная стенка отсутствует, поэтому их клетки имеют неправильную форму и похожи на амёб.
Клетки некоторых видов архей могут объединяться в агрегаты и филаменты длиной до 200 мкм. Эти организмы могут формировать биоплёнки. В культурах Thermococcus coalescens клетки сливаются друг с другом, формируя одну крупную клетку. Археи рода Pyrodictium образуют сложные многоклеточные колонии, в которых клетки объединены с помощью длинных, тонких, полых трубок, называемых cannulae, которые выступают над поверхностями клеток и собирают их в густое кустовидное скопление. Функции этих трубок не ясны, но, возможно, они осуществляют коммуникацию и обмен питательными веществами между соседними клетками. Существуют и многовидовые колонии, как, например, «нить жемчуга», обнаруженная в 2001 году в болоте в Германии. Круглые беловатые колонии некоторых необычных эвриархеот перемежаются тонкими нитями, которые могут достигать до 15 см в длину и состоят из особых видов бактерий.
Археи и бактерии имеют очень похожую структуру клеток, однако их состав и организация отделяют архей от бактерий. Как у бактерий, у них отсутствуют внутренние мембраны и органеллы, клеточные мембраны, как правило, ограничены клеточной стенкой, а плавание осуществляется за счёт одного или более жгутиков. Структурно археи наиболее схожи с грамположительными бактериями. Большинство имеет одну плазматическую мембрану и клеточную стенку, периплазматическое пространство отсутствует. Исключением из этого главного правила является Ignicoccus, у которого крупное периплазматическое пространство, ограниченное наружной мембраной, содержит окружённые мембраной везикулы.
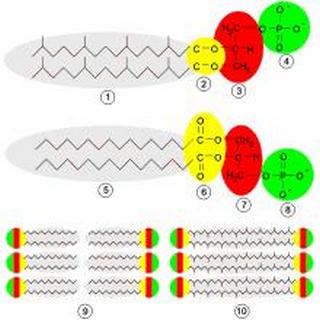
Структура мембран. Вверху — фосфолипиды архей: 1 — изопреновые цепочки; 2 — простые эфирные связи; 3 — остаток L-глицерина; 4 — фосфатная группа. Посередине — бактериальные или эукариотические фосфолипиды: 5 — цепочки жирных кислот; 6 — сложноэфирные связи; 7 — остаток D-глицерина; 8 — фосфатная группа. Снизу: 9 — липидный бислой бактерий и эукариот; 10 — липидный монослой некоторых архей.
Мембраны.
Молекулы, из которых построены мембраны архей, сильно отличаются от тех, которые используются в мембранах других организмов. Это указывает на то, что археи состоят лишь в отдалённом родстве с бактериями и эукариотами. У всех живых организмов клеточные мембраны построены из фосфолипидов. Молекулы фосфолипидов состоят из двух частей: гидрофильной полярной, состоящей из фосфатов, и гидрофобной неполярной, состоящей из липидов. Эти компоненты объединены через остаток глицерина. В воде молекулы фосфолипидов кластеризуются, при этом фосфатные «головки» оказываются обращёнными к воде, а липидные «хвосты» — обращёнными от неё и спрятанными внутрь кластера. Главной составляющей мембраны являются два слоя таких фосфолипидов, именуемые липидным бислоем.
Эти фосфолипиды у архей обладают четырьмя необычными чертами:
• У бактерий и эукариот мембраны состоят главным образом из глицерин-сложноэфирных липидов, тогда как у архей они сложены из глицерин-эфирных липидов. Различным является тип связи между остатками липидов и глицерина. Связи двух типов обозначены жёлтым на схеме справа. В сложноэфирных липидах связь сложноэфирная, а в эфирных — эфирная. Эфирные связи химически более стойкие, чем сложноэфирные (эстерные). Эта стабильность помогает археям выживать при высоких температурах, а также в сильнокислых и сильнощелочных средах. Бактерии и эукариоты содержат некоторое количество эфирных липидов, но по сравнению с археями они не являются главной составляющей мембран.
• Имеется отличие в стереохимии — у архей асимметрический центр глицериновой составляющей имеет L-конфигурацию, а не D-, как у других организмов. Поэтому для синтеза фосфолипидов археи используют совершенно другие ферменты, чем бактерии и эукариоты. Такие ферменты появились очень рано в истории жизни, что указывает на то, что археи рано отделились от двух других доменов.
• Липидные «хвосты» архей химически отличны от таковых у других организмов. Основу липидов архей составляет изопреноидная боковая цепь, и их липиды представляют собой длинные цепи с множеством побочных ветвей, иногда даже с циклопропановыми и циклогексановыми кольцами. Хотя изопреноиды играют важную роль в биохимии многих организмов, только археи используют их для создания фосфолипидов. Предполагают, что эти разветвлённые цепи, как и эфирные связи, служат для приспособления к обитанию при высоких температурах. Установлено, что изопреноидные мембраны сохраняют в широком диапазоне температур (0—100 °C) жидкокристаллическое состояние, что необходимо для их нормального биологического функционирования. Проницаемость таких мембран для ионов и низкомолекулярных органических веществ также мало изменяется с повышением температуры, в отличие от мембран из «обычных» липидов, у которых она резко возрастает.
• У некоторых архей липидный бислой заменяется монослоем. Фактически при этом липидные «хвосты» двух разных фосфолипидных молекул сливаются с образованием одной молекулы с двумя полярными головками. Эти слияния делают мембрану более стойкой и лучше приспособленной для суровых условий. К примеру, ферроплазма имеет липиды этого типа, и они помогают ей выживать в сильнокислых условиях.
Клеточная стенка.
Большинство архей (но не Thermoplasma и Ferroplasma) обладают клеточной стенкой. У большей части из них она сформирована молекулами поверхностных белков, образующих наружный S-слой. S-слой представляет собой жёсткую сетку из белковых молекул, покрывающих клетку снаружи, подобно кольчуге. Этот слой защищает клетку от физических и химических воздействий, а также предотвращает контакт макромолекул с клеточной мембраной. В отличие от бактерий, клеточная стенка архей не содержит пептидогликан. Метанобактерии (лат. Methanobacteriales) имеют клеточные стенки, содержащие псевдопептидогликан, который напоминает пептидогликан эубактерий по морфологии, функции и физической структуре, но отличен по химической: в нём нет остатков D-аминокислот и N-ацетилмурамовой кислоты.
Жгутики.
Жгутики архей работают так же, как и у бактерий: их длинные нити приводятся в движение вращательным механизмом в основании жгутика. Этот механизм работает за счёт трансмембранного протонного градиента. Тем не менее жгутики архей значительно отличаются от бактериальных по строению и способу сборки. Два типа жгутиков развились из разных предковых структур. Бактериальный жгутик и система секреции III типа имели общую предковую структуру, а архейный жгутик произошёл от бактериальных пилей IV типа. Жгутик бактерий полый внутри и собирается из субъединиц, которые проходят вверх по центральной поре к концу жгутика. Жгутики же архей строятся путём добавления субъединиц в их основание. Кроме того, в отличие от бактериальных жгутиков, в жгутики архей входит несколько видов флагеллинов.
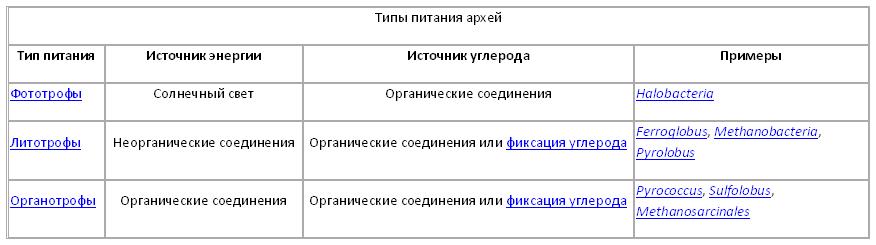
Метаболизм.
Археи демонстрируют огромное разнообразие химических реакций, протекающих в их клетках в процессе метаболизма, а также источников энергии. Эти реакции классифицируются по группам питания в зависимости от источников энергии и углерода. Некоторые археи получают энергию из неорганических соединений, таких как сера или аммиак (они являются литотрофами). К ним относятся нитрифицирующие археи, метаногены и анаэробные метаноокислители. В этих реакциях одно соединение отдаёт электроны другому (окислительно-восстановительные реакции), а выделяющаяся при этом энергия служит топливом для осуществления различных клеточных процессов. Соединение, отдающее электроны, называется донором, а принимающее — акцептором. Выделяющаяся энергия идёт на образование АТФ путём хемиосмоса. В сущности, это основной процесс, протекающий в митохондриях эукариотических клеток.
Другие группы архей используют в качестве источника энергии солнечный свет (их называют фототрофами). Однако ни один из этих организмов не образует кислород в процессе фотосинтеза. Многие базовые метаболические процессы являются общими для всех форм жизни, например, археи используют модифицированный вариант гликолиза (путь Энтнера-Дудорова), а также полный или частичный цикл Кребса (трикарбоновых кислот). Это, вероятно, отражает раннее возникновение этих путей в истории жизни и их высокую эффективность.
Некоторые эвриархеоты являются метаногенами и обитают в анаэробных средах, таких как болота. Такой тип метаболизма появился рано, и возможно даже, что первый свободноживущий организм был метаногеном. Обычная для этих организмов биохимическая реакция представляет собой окисление водорода с использованием углекислого газа в качестве акцептора электронов. Для осуществления метаногенеза необходимо множество различных коферментов, уникальных для этих архей, таких как кофермент М и метанофуран. Некоторые органические соединения, такие как спирты, уксусная и муравьиная кислоты, могут использоваться метаногенами в качестве альтернативных акцепторов электронов. Подобные реакции протекают у архей, живущих в пищеварительном тракте. У ацидотрофных архей уксусная кислота распадается непосредственно на метан и углекислый газ. Такие ацидотрофные археи относятся к отряду Methanosarcinales. Они являются важной составляющей сообществ микроорганизмов, продуцирующих биогаз.
Другие археи используют атмосферный углекислый газ как источник углерода благодаря процессу фиксации углерода (то есть они являются автотрофами). Этот процесс включает в себя либо сильно изменённый цикл Кальвина, либо недавно открытый метаболический путь, известный как 3-гидроксилпропионат/4-гидроксибутиратный цикл. Кренархеоты также используют обратный цикл Кребса, а эвриархеоты — восстановительный ацетил-СоА процесс. Фиксация углерода осуществляется за счёт энергии, получаемой из неорганических соединений. Ни один известный вид архей не фотосинтезирует. Источники энергии, которые используют археи, чрезвычайно разнообразны, начиная от окисления аммиака Nitrosopumilales до окисления сероводорода или элементарной серы, проводимого Sulfolobus, при этом в качестве акцепторов электронов могут использоваться кислород или ионы металлов.
Фототрофные археи используют солнечный свет для получения химической энергии в виде АТФ. У Halobacteria активируемые светом ионные насосы как бактериородопсин и галородопсин создают ионный градиент путём выкачивания ионов из клетки через плазматическую мембрану. Запасённая в этом электрохимическом градиенте энергия преобразуется в АТФ с помощью АТФ-синтазы. Этот процесс представляет собой форму фотофосфорилирования. Способность этих насосов переносить ионы через мембраны при освещении обусловлена изменениями, которые происходят в структуре ретинолового кофактора, скрытого с центре белка, под действием света.
Генетика.
Как правило, археи имеют одиночную кольцевую хромосому, размер которой может достигать 5 751 492 пар нуклеотидов у Methanosarcina acetivorans, обладающей самым большим известным геномом среди архей. Одну десятую размера этого генома составляет геном с 490 885 парами нуклеотидов у Nanoarchaeum equitans, имеющего самый маленький известный геном среди архей; он содержит лишь 537 генов, кодирующих белки. Также у архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды. Возможно, плазмиды могут передаваться между клетками при физическом контакте, в ходе процесса сходного с конъюгацией бактерий.
Археи могут поражаться вирусами, содержащими двухцепочечную ДНК. Эти виды неродственны другим группам вирусов и имеют различные необычные формы, включая бутылки, крючки и капли. Эти вирусы были тщательно изучены на термофилах, в основном отрядов Sulfolobales и Thermoproteales. В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражающий галофильные археи. Защитные реакции архей против вирусов могут включать механизм, близкий к РНК-интерференции эукариот.
Археи генетически отличны от эукариот и бактерий, причём до 15 % белков, кодируемых одним геномом археи, уникальны для этого домена, хотя функции большинства этих белков неизвестны. Большая часть уникальных белков, функция которых известна, принадлежит эвриархеотам и задействована в метаногенезе. Белки, общие для архей, бактерий и эукариот, участвуют в основных клеточных функциях и касаются в основном транскрипции, трансляции и метаболизма нуклеотидов. К другим особенностям архей можно отнести организацию генов, выполняющих связанные функции (к примеру, гены, ответственные за разные этапы одного и того же метаболического процесса), в опероны и большие отличия в строении генов тРНК и их аминоацил-тРНК синтетаз.
Транскрипция и трансляция архей больше напоминают эти процессы в клетках эукариот, чем бактерий, причём РНК-полимераза и рибосомы архей очень близки к аналогичным структурам у эукариот. Хотя у архей есть лишь один тип РНК-полимеразы, по строению и функции в транскрипции она близка к РНК-полимеразе II эукариот, при этом схожие группы белков (главные факторы транскрипции) обеспечивают связывание РНК-полимеразы с промотором гена. В то же время другие факторы транскрипции архей более близки к таковым у бактерий. Процессинг РНК у архей проще, чем у эукариот, так как большинство генов архей не содержит интронов, хотя в генах их тРНК и рРНК их достаточно много, также они присутствуют в небольшом количестве генов, кодирующих белки.
Размножение.
Археи размножаются бесполым путём: бинарным или множественным делением, фрагментацией или почкованием. Мейоза не происходит, поэтому даже если представители конкретного вида архей существуют более чем в одной форме, все они имеют одинаковый генетический материал. Клеточное деление определяется клеточным циклом: после того, как хромосома реплицировалась и две дочерние хромосомы разошлись, клетка делится. Детали были изучены лишь у рода Sulfolobus, но особенности его цикла очень схожи с таковыми и у эукариот, и у бактерий. Репликация хромосом начинается с множественных точек начала репликации с помощью ДНК-полимеразы, похожей на аналогичные ферменты эукариот. Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки, и компоненты септы, проходящей через центр клетки, схожи с их бактериальными эквивалентами.
Археи не образуют споры. Некоторые виды Haloarchaea могут претерпевать смену фенотипа и существовать как клетки нескольких различных типов, включая толстостенные клетки, устойчивые к осмотическому шоку и позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания.
Среды обитания.
Археи живут в широком диапазоне сред обитания и являются важной частью глобальной экосистемы, могут составлять до 20 % общей биомассы. Первые открытые археи были экстремофилами. Действительно, многие археи выживают при высоких температурах, часто свыше 100 °C, и обнаружены в гейзерах, чёрных курильщиках и маслосборниках. Другие приспособились к жизни в очень холодных условиях, в сильносолёных, сильнокислых и сильнощелочных средах, а также при высоком давлении — до 700 атмосфер (барофилы). Однако среди архей есть и мезофилы, обитающие в мягких условиях, в болотистых местностях, сточных водах, океанах и почве.
Экстремофильные археи относятся к четырём главным физиологическим группам: галофилам, термофилам, ацидофилам (кислотоустойчивые) и алкалифилам (щелочеустойчивые). Эти группы нельзя рассматривать в ранге типа или как другие самостоятельные таксоны. Они не взаимоисключают друг друга, и некоторые археи относят одновременно к нескольким группам. Тем не менее, они являются удобной стартовой точкой для классификации.
Галофилы, включая род Halobacterium, живут в экстремально солёных средах, таких как солёные озёра, и при минерализации больше 20—25 % превосходят по численности своих соседей-бактерий. Термофилы лучше всего растут на температурах свыше 45 °C в таких местах, как горячие источники; для гипертермофилов оптимальная температура — 80 °C и выше. Methanopyrus kandleri (штамм 116) растёт при 122 °C, рекордно высокой температуре для всех организмов.
Другие археи обитают в очень кислых или щелочных средах. Например, наиболее устойчивый ацидофил Picrophilus torridus растёт при pH = 0, что эквивалентно 1,2 молярной серной кислоте.
Устойчивость к экстремальным условиям внешней среды сделала архей центральной темой в обсуждениях возможных свойств жизни на других планетах. Некоторые среды, в которых обитают экстремофилы, не сильно отличаются от таковых на Марсе, что наводит на мысль о возможном переносе таких устойчивых микроорганизмов между планетами на метеоритах.
Недавно несколько работ показали, что археи обитают не только в термофильных и мезофильных условиях, но также встречаются, иногда в большом количестве, и в местах с низкими температурами. Например, археи встречаются в холодных водах, таких как полярные моря. Ещё более важно, что огромное количество архей обнаружено повсеместно в океанах в неэкстремальных условиях в составе планктона (как часть пикопланктона). Хотя эти археи могут присутствовать в поистине колоссальном количестве (до 40 % от общей биомассы микробов), почти ни один из этих видов не был изолирован, выращен и изучен в чистой культуре. Поэтому наше понимание роли архей в экологии океана, их влияния на глобальный биогеохимический круговорот остаётся в значительной мере неполным. Некоторые морские кренархеоты способны к нитрификации, поэтому вероятно, что они оказывают влияние на океанический круговорот азота, хотя эти океанические кренархеоты могут использовать и другие источники энергии. Большое число архей также обнаружено в осадке, покрывающем океаническое дно, причём они составляют большинство живых клеток на глубине больше 1 м от уровня океанического дна.

Пятно цветных термофильных архей (Национальный парк Йеллоустоун, США)
Роль в круговороте веществ.
Археи вторично используют такие элементы, как углерод, азот и серу в своих различных средах обитания. Хотя такие превращения необходимы для нормального функционирования экосистемы, археи могут также содействовать вредным изменениям, вызванным деятельностью человека, и даже вызвать загрязнение.
Археи осуществляют многие этапы круговорота азота. Это включает в себя как реакции, удаляющие азот из экосистемы, к примеру, азотное дыхание и денитрификация, так и процессы, в ходе которых поглощается азот, такие как усвоение нитратов и фиксация азота. Недавно была открыта причастность архей к окислению аммиака. Эти реакции особенно важны в океанах. Археи также играют важную роль в почвенном окислении аммиака. Они образуют нитриты, которые затем окисляются другими микробами в нитраты. Последние потребляются растениями и другими организмами.
В круговороте серы археи, живущие за счёт окисления соединений серы, получают их из каменистых пород и делают их доступными для других организмов. Однако виды, осуществляющие это, такие как Sulfolobus, образуют серную кислоту как побочный продукт, и существование таких организмов в заброшенных шахтах может, совместно с кислотными шахтными водами, причинить вред окружающей среде.
В круговороте углерода метаногены удаляют водород и играют важную роль в разложении органической материи популяциями микроорганизмов, выступающих как разлагатели в анаэробных экосистемах, таких как илы, болота и водоочистные сооружения. Однако метан является одним из самых распространённых газов в земной атмосфере, вызывающих парниковый эффект, достигая 18 % от общего объёма парниковых газов. Он в 25 раз более эффективен по способности вызывать парниковый эффект, чем углекислый газ. Метаногены являются главным источником атмосферного метана и выделяют большую часть ежегодного выброса метана. Поэтому эти археи причастны к созданию парникового эффекта на Земле и глобальному потеплению.
Взаимодействие с другими организмами.
Хорошо изученные отношения между археями и другими организмами — мутуализм и комменсализм. Пока не существует чётких доказательств существования патогенных или паразитических видов архей. Однако была предположена связь между некоторыми видами метаногенов и инфекциями полости рта. Кроме того, вид Nanoarchaeum equitans, возможно, является паразитом другого вида архей, поскольку он выживает и размножается только в клетках кренархеона Ignicoccus hospitalis и не приносит никакой очевидной выгоды своему хозяину. С другой стороны, архееподобные ацидофильные наноорганизмы Ричмондских рудников (англ. Archaeal Richmond Mine acidophilic nanoorganisms) (ARMAN) иногда прикрепляются к клеткам других архей в биоплёнках кислых сточных вод рудников. Природа этого взаимодействия не ясна, но, в отличие от случая Nanorchaeaum—Ignicoccus, сверхмелкие клетки ARMAN всегда остаются независимыми от клеток Thermoplasmatales.

ARMAN — новая группа архей, обитающая в дренажных кислых шахтных водах.
Мутуализм.
Одним хорошо понятным примером мутуализма является взаимодействие простейших и метанообразующих архей, обитающих в пищеварительном тракте животных, способных переваривать целлюлозу, таких как жвачные и термиты. В этих анаэробных условиях простейшие разлагают целлюлозу для получения энергии. В этом процессе в качестве побочного продукта освобождается водород, однако высокий его уровень сокращает получение энергии. Метаногены превращают водород в метан, и простейшие могут дальше нормально получать энергию.
В случае анаэробных простейших вроде Plagiopyla frontata археи живут внутри клетки простейшего и потребляют водород, образуемый в его гидрогеносомах. Археи также взаимодействуют и с более крупными организмами. Например, морская архея Cenarchaeum symbiosum живёт внутри (как эндосимбионт) губки Axinella mexicana.
Комменсализм.
Археи могут быть комменсалами, то есть существовать совместно с другим организмом, не принося ему ни пользы, ни вреда, но с выгодой для себя. К примеру, метаноген Methanobrevibacter smithii является наиболее типичным представителем архей в микрофлоре человека. Каждый десятый прокариот в человеческом пищеварительном тракте принадлежит к этому виду. В пищеварительном тракте термитов и человека эти метаногены в действительности могут быть мутуалистами, взаимодействущими с другими микробами пищеварительного тракта и способствующими пищеварению. Археи также взаимодействуют с другими организмами, к примеру, живут на внешней поверхности кораллов и в части почвы, прилегающей к корням растений (ризосфере).
Классификация.
Классификация архей, как и прокариот в целом, быстро меняется и во многом остаётся спорной. Современные системы классификации стремятся объединить археи в группы организмов со схожими структурными свойствами и общими предками. Эти классификации основаны на анализе структуры генов рРНК для установления родственных отношений между организмами (молекулярная филогенетика). Большую часть архей, выращиваемых в лабораториях и хорошо изученных, относят к двум главным типам, кренархеоты (лат. Crenarchaeota) и эвриархеоты (лат. Euryarchaeota). Другие группы были выделены в порядке рабочей гипотезы. Например, довольно необычный вид Nanoarchaeum equitans, открытый в 2003 году, был выделен в самостоятельный тип Nanoarchaeota. Был также предложен новый тип Korarchaeota. Он объединяет небольшую группу термофильных видов, обладающих особенностями обоих основных типов, но более родственно близких к кренархеотам. Другие недавно открытые виды имеют лишь дальнее родство с вышеперечисленными группами, например, Archaeal Richmond Mine acidophilic nanoorganisms (ARMAN), открытые в 2006 году и являющиеся одними из самых мелких известных на сегодняшний момент организмов.
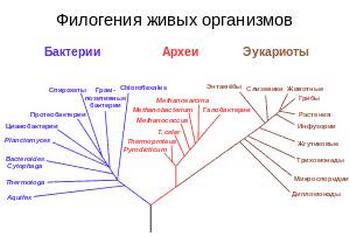
Филогенетическое древо, построенное на основании анализа рРНК, показывает разделение бактерий, архей и эукариот.
Разделение архей на виды также является спорным. В биологии вид определяется как группа близкородственных организмов. Обычный критерий, которым пользуются в подобных ситуациях — организмы одного вида могут скрещиваться друг с другом, но не с особями других видов, — в данном случае не работает, поскольку археи размножаются только бесполым путём.
Археи демонстрируют высокий уровень горизонтального переноса генов между линиями. Некоторые исследователи предполагают, что особи можно объединять в популяции, похожие на виды, при условии высокой степени схожести их геномов и редко случающегося переноса генов между организмами с менее схожими геномами, как в случае рода ферроплазма (лат. Ferroplasma). С другой стороны, изучение рода Halorubrum показало существование значимой передачи генов между дальнеродственными популяциями, что ограничивает применимость этого критерия. Вторая проблема состоит в том, какое практическое значение может иметь подобное разделение на виды.
Современные данные о генетическом разнообразии архей фрагментарны, и общее число их видов не может быть оценено с какой-либо точностью. Сравнительный анализ структур 16S рРНК архей позволил предположить существование 18—23 филогенетических групп уровня типов, причём представители лишь 8 групп были непосредственно выращены в лаборатории и изучены (с учётом вероятной полифилии некоторых выделяемых в данный момент типов). Многие из этих гипотетических групп известны лишь по одной последовательности рРНК, что говорит о том, что пределы разнообразия этих организмов остаются неясными. Многие бактерии также никогда не были культивированы в лаборатории, что приводит к схожим проблемам при их характеристике.
По устоявшейся классификации в настоящее время выделяют 5 типов архей:
• Crenarchaeota — термофилы, термоацидофилы, серные анаэробные бактерии;
• Euryarchaeota — метаногенные и галофильные археи;
• Nanoarchaeota — единственный известный представитель Nanoarchaeum equitans;
• Korarchaeota — ДНК обнаружена в геотермальных источниках США, Исландии, на рисовых полях Японии, культивируемые виды пока неизвестны;
• Thaumarchaeota — в основном окислители аммония, как например, морской аммоний-окислитель Nitrosopumilus maritimus и аммоний-окислитель преимущественно почвенного происхождения Nitrososphaera gargensis. Недавние филогенетические исследования, основанные на сравнительном анализе структур рибосомальных белков и других важнейших генов, подтвердили существование типа Таумархеот.
Иногда выделяют также шестой тип — Aigarchaeota.
Значение в технологии и промышленности.
Экстремофильные археи, особенно устойчивые к высоким температурам или повышенной кислотности/щёлочности среды, являются источником ферментов, работающих в этих суровых условиях. Эти ферменты находят множество применений. Например, термостабильные ДНК-полимеразы, такие как Pfu ДНК-полимераза вида Pyrococcus furiosus, полностью изменили молекулярную биологию, дав возможность использовать полимеразную цепную реакцию для простого и быстрого клонирования ДНК. В промышленности амилазы, галактозидазы и пуллуланазы других видов Pyrococcus, функционирующие при температуре свыше 100 °C, применяются при производстве продуктов питания при высоких температурах, к примеру, при производстве молока и сыворотки с низким содержанием лактозы. Ферменты этих термофильных архей остаются очень стабильными в органических растворителях, что позволяет использовать их в безопасных для окружающей среды процессах в зелёной химии для синтеза органических соединений. Их стабильность делает эти ферменты удобными для использования в структурной биологии, поэтому аналоги ферментов бактерий и эукариот, получаемые из экстремофильных архей, часто применяются в структурных исследованиях.
По сравнению с применением ферментов архей, использование самих организмов в биотехнологии развито слабо. Метанообразующие археи являются важной частью водоочистных сооружений, так как они входят в сообщество микроорганизмов, осуществляющих анаэробное разложение и образование биогаза. В обогащении полезных ископаемых ацидофильные археи могут использоваться для получения металлов из руд, в том числе золота, кобальта и меди.
Археи могут дать потенциально полезные антибиотики. Пока описано мало археоцинов, но предполагается, что их существует сотни, особенно много их может быть получено из родов Haloarchaea и Sulfolobus. Эти антибиотики по структуре отличны от бактериальных, поэтому они могут иметь другие механизмы действия. Кроме того, они могут позволить создать селектируемые маркеры для использования в молекулярной биологии архей.

Впервые археи были обнаружены в экстремальных местах обитания — горячих вулканических источниках
Определитель Берджи.
Определитель Берджи систематизирует все известные бактерии по нашедшим в практической бактериологии наибольшее распространение принципам идентификации бактерий, основанным на различиях в строении клеточной стенки и отношении к окраске по Граму. Определитель выделяет четыре основных категории бактерий — Gracillicutes [от лат. gracilis, изящный, тонкий, + cutis, кожа] — виды с тонкой клеточной стенкой, окрашивающиеся грамотрицательно; firmicutes [от лат. flrmus, крепкий, + cutis, кожа] — бактерии с толстой клеточной стенкой, окрашивающиеся грамположительно; Tenericutes [от лат. tener, нежный, + cutis, кожа] — бактерии, лишённые клеточной стенки (микоплазмы и прочие представители класса Mollicutes) и Mendosicutes [от лат. mendosus, неправильный, + cutis, кожа] — архебактерии (метан- и сульфатредуцирующие, галофильные, термофильные и архебактерии, лишённые клеточной стенки). Описание бактерий даётся по группам (секциям), в состав которых включены семейства, роды и виды; в некоторых случаях в состав групп входят классы и порядки. Патогенные для человека бактерии входят в небольшое число групп.
Группа 1 определителя Берджи. Спирохеты. Включает свободноживущие и паразитические виды; для человека патогенны представители родов Treponema, Borrelia и Leptospira.
Группа 2 определителя Берджи. Аэробные и микроаэрофильные подвижные извитые и изогнутые грамотрицательные бактерии. Патогенные для человека виды входят в роды Campylobacter, Helicobacters, Spirillum.
Группа 3 определителя Берджи. Неподвижные (редко подвижные) грамотрицательные бактерии. Не содержит патогенные виды.
Группа 4 определителя Берджи. Грамотрицательные аэробные и микроаэрофильные палочки и кокки. Патогенные для человека виды включены в состав семейств Legionellaceae, Neisseriaceae и Pseudomonadaсеае, в группу входят также патогенные и условно-патогенные бактерии родов Acinetobacter, Afipia, Alcaligenes, Bordetella, Brucella, Flavobacterium, Francisella, Kingella и Moraxella.
Группа 5 определителя Берджи. Факультативно анаэробные грамотрицательные палочки. Группа образована тремя семействами — Enterobacteriaceae, Vibrionaceae и Pasteurellaceae, каждое из которых включает патогенные виды, а также патогенные и условно-патогенные бактерии родов Calymmobaterium, Cardiobacterium, Eikenetta, Gardnerella и Streptobacillus.
Группа 6 определителя Берджи. Грамотрицательные анаэробные прямые, изогнутые и спиральные бактерии. Патогенные и условно-патогенные виды входят в состав родов Bacteroides, Fusobacterium, Porphoromonas и Prevotelta.
Группа 7 определителя Берджи. Бактерии, осуществляющие диссимиляционное восстановление сульфата или серы Не включает патогенные виды.
Группа 8 определителя Берджи. Анаэробные грамотрицательные кокки. Включает условно-патогенные бактерии poда Veillonella.
Группа 9 определителя Берджи. Риккетсии и хламидии. Три семейства — Rickettsiaceae, Bartonellaceae и Chlamydiaсеае, каждое из которых содержит патогенные для человека виды.
Группы 10 и 11 определителя Берджи включают анокси- и оксигенные фототрофные бактерии, не патогенные для человека.
Группа 12 определителя Берджи. Аэробные хемолитотрофные бактерии и родственные организмы. Объединяет серо- железо- и марганецокисляющие и нитрифицирующие бактерии, не вызывающие поражения у человека.
Группы 13 и 14 определителя Берджи включают почкующиеся и/или обладающие выростами бактерии и бактерии образующие футляры. Представлены свободноживущими видами, не патогенными для человек;
Группы 15 и 16 определителя Берджи объединяют скользящие бактерии, не образующие плодовые тела и образующие их. Группы не включают виды, патогенные для человека.
Группа 17 определителя Берджи. Грамположительные кокки. Включает условно-патогенные виды родов Enterococcus, Leuconostoc, Peptococcus, Peptostreptococcus, Sarcina, Staphylococcus, Stomatococcus, Streptococcus.
Группа 18 определителя Берджи. Спорообразующие грамположительные палочки и кокки. Включает патогенные, условно-патогенные палочки родов Clostridium и Bacillus.
Группа 19 определителя Берджи. Споронеобразующие грамположительные палочки правильной формы. Включая условно-патогенные виды родов Erysipelothrix и Listeria.
Группа 20 определителя Берджи. Споронеобразующие грамположительные палочки неправильной формы. В состав группы входят патогенные и условно-патогенные виды родов Actinomyces, Corynebacterium, Gardnerella, Mobiluncus и др.
Группа 21 определителя Берджи. Микобактерии. Включает единственный род Mycobacterium, объединяющий патогенные и условно-патогенные виды.
Группы 22-29. Актиномицеты. Среди многочисленных видов лишь нокардиоформные актиномицеты (группа 22) родов Gordona, Nocardia, Rhodococcus, Tsukamurella, Jonesia, Oerskovi и Terrabacter способны вызывать поражения у человека.
Группа 30 определителя Берджи. Микоплазмы. Патогенны для человека виды, включённые в состав рода Acholeplasma, Mycoplasma и Ureaplasma.
Остальные группы определителя Берджи — метаногенные бактерии (31), сульфатредуцируюшие бактерии (32 экстремально галофильные аэробные архебактерии (33), архебактерии, лишённые клеточно стенки (34), экстремальные термофилы и гипертермофилы, метаболизируюшие серу (35) — не содержат патогенные для человека виды.