(Ascomycota) Тип Аскомикоты, Аскомицеты, Сумчатые грибы, Phylum Ascomycota 16 классов
Информация из Интернета
Аскомикоты, или Аскомицеты, или Сумчатые грибы (Ascomycota) — отдел (тип) в царстве грибов, объединяющий организмы с септированным (разделённым на части) мицелием и специфическими органами полового спороношения — сумками (асками), содержащими чаще всего по 8 аскоспор. Имеют и бесполое спороношение, причём во многих случаях половой процесс утрачивается (такие виды грибов традиционно относили к несовершенным грибам).
Название отдела происходит от греческого слова ασκος (askos) — «сумка», «мешок» и латинского слова mycota — «грибы», указывает на специфические органы полового спороношения — сумки.
К аскомицетам относят примерно 6400 родов, включающих более 64 000 видов. Среди них — многие дрожжи (в том числе широко используемые в производстве хлебопекарной и алкогольной продукции пекарские дрожжи, Saccharomyces cerevisiae) — вторично одноклеточные организмы, не образующие мицелия. Нужно упомянуть также ряд макромицетов — съедобные (сморчки, трюфели, алеврия) и ядовитые (строчки). Многие аскомицеты являются паразитами высших растений, вызывающими у последних ряд заболеваний. Это представители родов тафрина (Taphrina), вентурия (Venturia), подосфера (Podosphaera), эризифе (Erysiphe), ботритис (Botrytis), гибберелла (Gibberella), спорынья (Claviceps) и др. Представители рода кордицепс (Cordyceps) паразитируют на членистоногих. Среди родов аскомицетов имеются и патогенные для человека: пневмоцистис (Pneumocystis; вызывает пневмоцистную пневмонию), трихофитон (Trichophyton; вызывает трихофитию), некоторые виды родов аспергилл (Aspergillus) и пеницилл (Penicillium), способные вызывать различные дерматомикозы и глубокие микозы (а также приводить к порче пищевых продуктов; в то же время пенициллы и аспергиллы находят существенное применение в микробиологической промышленности). Ряд видов аскомицетов, и прежде всего Neurospora crassa, широко используют в генетических исследованиях.
В палеозое (девон — карбон) в ряде оформившихся к тому времени эволюционных линий аскомицетов имела место лихенизация — возникновение лишайников как симбиотических ассоциаций грибов с микроскопическими водорослями. Характерные представители лишайников — аспицилия (Aspicilia), ксантория (Xanthoria), кладония (Cladonia), пармелия (Parmelia), цетрария (Cetraria), эверния (Evernia), уснея (Usnea), нередко существенно различные по внешнему виду. Некоторые виды кладонии образуют ягель — основной корм северных оленей; цетрария исландская («исландский мох») используется в традиционной исландской кулинарии, а аспицилию («лишайниковую манну») в прошлом употребляли в пищу жители пустынных районов Ближнего Востока. Экстракты из эвернии и пармелии нашли применение в парфюмерной промышленности, а ряд лишайников используют в производстве антибиотиков и других биологически активных веществ.
Вегетативное тело аскомицетов обычно представляет собой септированный (разделённый поперечными перегородками — септами — на отдельные клетки) гаплоидный мицелий (встречаются и дрожжевые формы, у которых вегетативное тело образовано одиночными почкующимися клетками). Наличие септированного мицелия свойственно и базидиомицетам, но у последних он на протяжении большей части жизненного цикла не гаплоидный, а дикариотичный. Разделение мицелия септами позволяет гифе в случае повреждения терять меньше клеточного содержимого и обусловливает большую выживаемость аскомицетов по сравнению с ценоцитными зигомицетами. Так, наименьший жизнеспособный фрагмент мицелия, способный дать начало новому организму, для зигомицета составляет около 100 мкм, а для аскомицета — 15—20 мкм.
В середине септы имеется простая пора, через которую возможен обмен цитоплазмой, органеллами и даже ядрами. У представителей подотдела Pezizomycotina рядом с септой встречается тельце Воронина — белковое микротельце, способное при повреждении гифы закупоривать пору и предотвращать потери цитоплазмы. Клетки аскомицетов обычно многоядерные, иногда (например, в порядке Erysiphales) — одноядерные. Стенки клеток, как правило, являются двуслойными (с тонким наружным и более или менее толстым внутренним слоями), в то время как у базидиомицетов они многослойные. В состав клеточной стенки входят полисахариды: β-глюканы и хитин (доля последнего не превышает 20—25 %, а у некоторых сахаромицетов его заменяет маннан).
У аскомицетов выражены две стадии развития: анаморфа, в которой организм размножается бесполым путём, и телеоморфа, в которой он формирует половые структуры. Вместе обе стадии носят название голоморфа. Анаморфа и телеоморфа не схожи, в зависимости от условий организм может переходить в телеоморфу, а может и нет, в результате чего часто один и тот же аскомицет описывался как два разных вида. Например, название Aspergillus fisheri дано анаморфе, телеоморфа которой описана как Neosartoria fisheri, аналогичная ситуация с Amorphotheca resinae и Hormoconis resinae и многими другими. В наши дни молекулярные методы позволяют обнаружить описанные дважды виды.
Бесполое размножение аскомицетов осуществляется практически всегда конидиями. Морфология конидий очень разнообразна. Они могут быть одноклеточными, с разным числом поперечных перегородок, с продольными и поперечными перегородками (муральными). Чаще всего конидии эллипсоидные, могут быть округлыми. Особенно сложное строение имеют конидии некоторых водных грибов, обитающих, в частности, на погружённых в воду разлагающихся листьях листопадных пород деревьев и кустарников.
Половое размножение аскомицетов проходит в форме гаметангиогамии, без образования дифференцированных гамет. Для этого на разных гифах образуются половые органы: мужские — антеридии и женские — архикарпы, представляющие собой видоизменённые оогонии.
У низших форм половой процесс напоминает зигогамию у зигомицетов (Zygomycota), поскольку образующиеся гаметангии внешне сходны и после их объединения сразу происходит слияние ядер. Однако у низших асковых грибов сливаются только два ядра многоядерных гаметангиев, а не все, то есть отсутствует множественная кариогамия. Образовавшееся диплоидное ядро без периода покоя делится мейотически, образуя гаплоидные ядра, а зигота трансформируется в сумку — аск (от греческого слова ασκος (askos) — «сумка», «мешок»).
У высших форм половой процесс проходит более сложно. Архикарп у них дифференцирован на расширенную нижнюю часть — аскогон и верхнюю в виде изогнутой трубочки — трихогину. Антеридий представляет собой одноклеточную структуру цилиндрической формы. При соприкосновении гаметангиев трихогина врастает своим концом в антеридий, после чего содержимое антеридия перетекает по ней в аскогон. Слияние цитоплазмы гаметангиев (плазмогамия) не сопровождается слиянием их гаплоидных ядер, хотя они сближаются и располагаются попарно, образуя дикарионы. Затем из аскогона вырастают аскогенные гифы, одновременно ядра дикарионов делятся, что ведёт к увеличению числа дикарионов. В каждую из аскогенных гиф проникают дикарионы. Завершается процесс образованием сумок (асков), которыми становятся концевые клетки, расположенные на концах аскогенных гиф, содержащие дикарион. Вначале гаплоидные ядра дикариона сливаются (происходит кариогамия), образуя диплоидное ядро. Без периода покоя это ядро делится мейотически, образуя четыре гаплоидных ядра, а те, в свою очередь, делятся митотически. В итоге появляются восемь гаплоидных клеток, которые становятся аскоспорами, а клетка, в которой они находятся, становится аском. У некоторых видов количество спор может быть меньшим (четыре) за счёт отсутствия митотического деления после мейотического или за счёт дегенерации части гаплоидных ядер или большим.
Таким образом, в жизненном цикле асковых грибов имеется три стадии: гаплоидная стадия, когда мицелий размножается бесполым путём; стадия дикариона и самая короткая — диплоидная стадия, когда молодая сумка непродолжительное время содержит диплоидное ядро.
У многих сумчатых грибов половой процесс упрощается. У них не образуются антеридии, а вместо них функционируют вегетативные гифы, конидии или мелкие клетки спермации, которые могут образовываться даже не на соседних гифах, а на достаточном расстоянии от архикарпа. Потоками воздуха, воды или через насекомых спермации переносятся на трихогину, после чего происходит слияние цитоплазмы клеток. Случается, что могут отсутствовать оба гаметангия, тогда половой процесс протекает в форме соматогамии, то есть сливаются вегетативные клетки. При этом следует отметить, что у сумчатых грибов есть формы с гомоталличным и гетероталличным мицелиями, причём последние всегда отличаются набором аллелей.
Аскогенные гифы с асками могут образовываться не только беспорядочно (в любом месте мицелия), как это происходит у низших форм, но и на плодовых телах, состоящих из плотно переплетённых гиф. У сумчатых грибов имеются четыре типа плодовых тел:
• Клейстотеции (клейстокарпии) представляют собой полностью замкнутое плодовое тело с находящимися внутри асками, освобождающимися после разрушения его стенок. Характерны для группы порядков плектомицеты.
• Перитеции (от древнегреческих слов περι- (peri-) — «возле», «около» и θηκη (teki) — «хранилище») — почти замкнуты («полузамкнуты»), то есть сумки окружены перидием, обычно имеют кувшинообразную форму с выводным отверстием в верхней части. Характерны для группы порядков пиреномицеты.
• Апотеции — открытые вместилища аск. Образуют чаши (блюдца). По верхней стороне плодового тела расположен слой сумок и парафиз; парафизы, пока сумки незрелые, могут смыкаться над ними своими вершинами и выполнять защитную функцию. Также образуют сморщенную поверхность сморчков (причём рёбра складок стерильны), могут, однако, не раскрываться (у трюфелей). Аскомицеты с апотециями считаются наиболее высокоорганизованными и объединяются в группу порядков дискомицеты.
• Псевдотеции характерны для класса Loculoascomycetes. При их формировании вначале образуются мицелиальные стромы, в полостях которых формируются гаметангии, происходит половой процесс и образование сумок.
Клейстотеции и перитеции часто развиваются на особых образованиях — стромах — плотных сплетениях гиф, которые часто ошибочно принимают за плодовые тела. Они могут находиться на поверхности стромы или быть погружёнными в её тело.
Во второй половине XX века аскомицеты рассматривались как класс Ascomycetes, включавшийся в состав отделов Mycota (Грибы) или Eumycota (Настоящие грибы). По месту формирования сумок класс делили на три подкласса: Hemiascomycetidae (гемиаскомицеты; сумки располагаются непосредственно на мицелии, плодовых тел нет), Loculoascomycetidae (локулоаскомицеты; сумки образуются в аскостромах и содержатся в полостях — локулах, настоящих плодовых тел нет) и Euascomycetidae (эуаскомицеты; сумки образуются в плодовых телах). В составе последнего подкласса, в свою очередь, выделяли, сообразуясь со строением плодовых тел, три группы порядков: дискомицеты (плодовые тела — апотеции), пиреномицеты (плодовые тела — обычно перитеции) и плектомицеты (плодовые тела — обычно клейстотеции).
В 1980—1990-е годы ряд микологов предложил повысить таксономический ранг аскомицетов до ранга отдела с названием Ascomycota. Название было валидизировано в 1998 году Т. Кавалье-Смитом; отдел стал крупнейшим по числу входящих в него видов в царстве Fungi. Основные его подразделения получили, соответственно, статус классов.
Развитие методов молекулярной филогенетики позволило в 1990-е годы приступить к построению новой системы аскомицетов, отражающей филогенетические связи между отдельными их порядками. Первый её вариант был предложен в 1997 году У. Э. Эрикссоном и К. Винкой (в статье «Supraordinal taxa of Ascomycota»). Названия основных таксонов новой системы были теперь новыми (типифицированными), однако преемственность с предшествующими классификациями во многом сохранялась: локулоаскомицеты в основном вошли в класс Dothideomycetes (часть была отнесена к классу Chaetothyriomycetes, который в дальнейшем был включён в состав Eurotiomycetes как подкласс Chaetothyriomycetidae), пиреномицеты — в класс Sordariomycetes, плектомицеты — в класс Eurotiomycetes. Дискомицеты оказались сборной группой, и их представители были включены в различные классы новой системы (при этом большинство лихенизированных видов дискомицетов вошло в класс Lecanoromycetes, нелихенизированных — в класс Leotiomycetes). Гемиаскомицеты в их старом понимании были разделены на два подотдела (Taphrinomycotina с четырьмя классами и Saccharomycotina с одним), а эуаскомицеты — объединены вместе с локулоаскомицетами в подотдел Pezizomycotina.
В XXI веке система аскомицетов была дополнена новыми классами, и их общее число достигло 20. Филогенетические связи между классами аскомицетов можно представить в виде такой кладограммы:
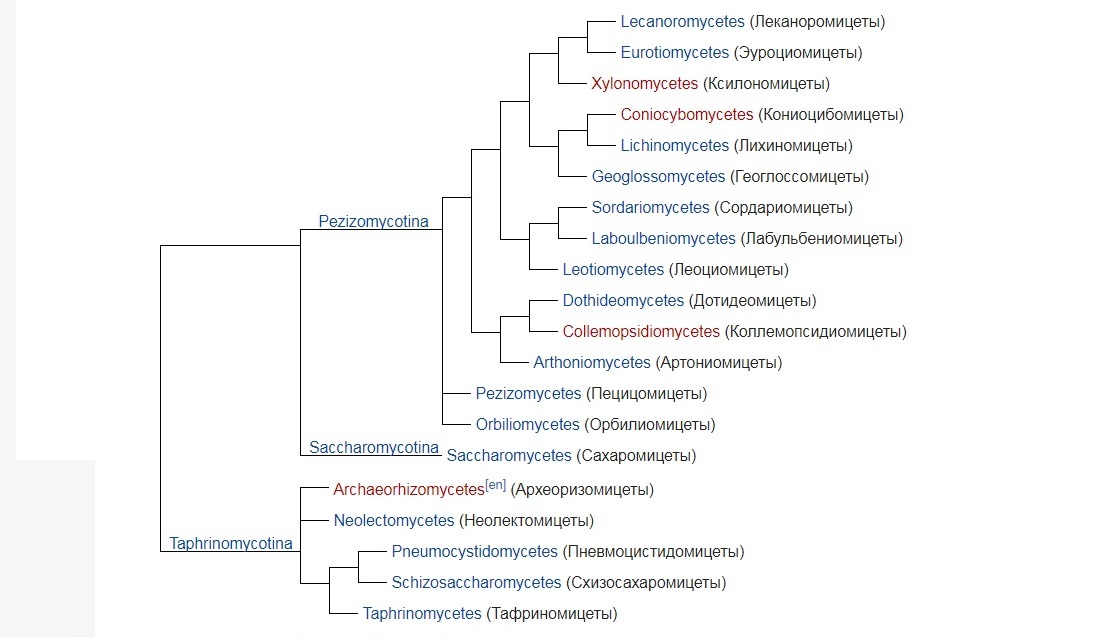
Представленная здесь кладограмма носит компромиссный характер: результаты различных исследований не вполне согласуются друг с другом. Однако считаются более или менее установленными: сестринское родство подотделов Saccharomycotina и Pezizomycotina и базальное положение подотдела Taphrinomycotina, а также их монофилия (хотя с включением в систему класса Archaeorhizomycetes монофилия подотдела Taphrinomycotina подтверждается не всегда); положение Orbiliomycetes и Pezizomycetes в основании Pezizomycotina и монофилия клады Leotiomyceta(Pezizomycotina за вычетом двух перечисленных классов); взаимная близость Arthoniomycetes и Dothideomycetes, Leotiomycetes и Sordariomycetes (сестринское родство Eurotiomycetes и Lecanoromycetes в некоторых исследованиях ставится под сомнение). В то же время нельзя считать вполне установленными отображённые на кладограмме филогенетические связи четырёх основных групп Leotiomyceta; особенно нестабильно положение классов Geoglossomycetes и Xylonomycetes.
Три порядка аскомицетов (Lahmiales, Medeolariales, Triblidiales) не включены в какие-либо классы; их рассматривают как Pezizomycotina incertae sedis.
Аскомицеты играют важную роль в наземных экосистемах. Разрушая такие субстраты, как опад, мёртвые ветки и стволы деревьев (в том числе входящие в их состав целлюлозу и лигнин, недоступные большинству организмов), они вносят существенный вклад в биологические циклы углерода и азота. Сами по себе аскомицеты являются пищей для ряда бактерий, простейших, микотрофных и микопаразитических грибов, беспозвоночных и позвоночных животных (птицы, кроты, грызуны и более крупные: олени, кабаны). Много аскомицетов и в водной среде — в пресноводных и морских местообитаниях, иногда крайне экстремальных.
Аскомицеты являются микобионтами подавляющего большинства лишайников (до 98 %). Помимо этого, более 1700 описанных видов аскомицетов, принадлежащих к 19 порядкам из 7 классов, относятся к группе лихенофильных грибов: они обитают исключительно в лишайниках, выступая как паразиты или комменсалы. С корневыми системами высших растений ряд видов аскомицетов образуют микоризу.
Значительная часть аскомицетов — сапротрофы. В то же время среди них немало паразитов растений, включая возбудителей весьма тяжёлых болезней растений. Среди этих болезней — парша яблони, плодовая гниль, мучнистая роса, голландская болезнь вязов, серая гниль, фузариозы, спорынья злаков и др.
Среди грибов, паразитирующих на животных и человеке, большинство — аскомицеты. Ряд аскомицетов — это плесени, повреждающие пищевые продукты и выступающие как биодеструкторы, которые разрушают материалы и конструкции.
Некоторые сумчатые грибы используются в хозяйственной деятельности человека — например, ряд представителей пецицомицетов (особенно трюфели и сморчки) употребляют в пищу. Дрожжи незаменимы в хлебопечении и на бродильных производствах, другие виды аскомицетов широко используются для получения биологически активных веществ (антибиотиков, витаминов, ферментов, алкалоидов, гиббереллинов, фитогормонов и пр.), органических кислот, кормового белка.
Ряд видов аскомицетов — нейроспору густую (Neurospora crassa), называемую иногда «дрозофилой мира грибов», некоторые виды сордарии (Sordaria), эмерицеллу лежачую (Emericella nidulans) и др. — широко используют в генетических исследованиях. В частности, к виду Neurospora crassa был успешно применён тетрадный анализ, позволивший уточнить представления о ходе кроссинговера при мейозе.