Домен Прокариоты, Доядерные (Prokaryota Chatton, 1925), Дробянки, Монеры (Monera Haeckel, 1866), Надцарство Прокариоты, Доядерные, Domain Prokaryota Chatton, 1925 (Prokaryotes) 2 царства
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Прокариоты, Доядерные (Prokaryota Chatton, 1925), Дробянки, Монеры (Monera Haeckel, 1866) — одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный и автотрофный (фотосинтез и хемосинтез). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды.
Прокариоты разделяют на два таксона в ранге домена (надцарства): Бактерии (Bacteria) и Археи (Archaea).
Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 г. Этот процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация), а также с помощью вирусов — бактериофагов (трансдукция).
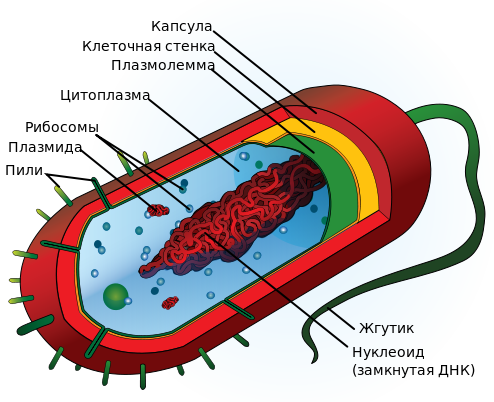
Строение типичной клетки прокариот: капсула, клеточная стенка, плазмалемма, цитоплазма, рибосомы, плазмида, пили, жгутик, нуклеоид.
Характерные особенности.
• Отсутствие четко оформленного ядра
• Наличие жгутиков, плазмид и газовых вакуолей
• Структуры, в которых происходит фотосинтез
• Формы размножения — бесполый способ, имеется псевдосексуальный процесс, в результате которого происходит лишь обмен генетической информацией, без увеличения числа клеток.
• Размер рибосомы — 70s (по коэф. седиментации различают и рибосомы др. типов, а также субчастицы и биополимеры, входящие в состав рибосом)
История понятия.
Монеры — этим именем Геккель назвал простейшие одноклеточные организмы без ядра. Так как присутствие ядра во многих случаях трудно констатируется, то первоначально, пока методы микроскопического исследования были сравнительно несовершенны, безъядерными считались очень многие формы. Вопрос о монерах представляет некоторый интерес ввиду того, что первоначальное возникновение организмов на земле, вероятно, произошло в форме тел, не дифференцированных ещё на ядро и протоплазму.
В настоящее время термин «монеры» не применяется.
Отличия эукариот от прокариот.
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит «хорошее ядро»). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд лет назад, от них около 1,2 млрд лет назад произошли эукариотические организмы.
Систематика микроорганизмов.
Естественная (филогенетическая) систематика микроорганизмов имеет конечной целью объединение родственных форм, связанных общностью происхождения, и установление иерархического соподчинения отдельных групп. До настоящего времени отсутствуют единые принципы и подходы к объединению (или разделению) их в различные таксономические единицы, хотя для них пытаются использовать сходство геномов как общепринятый критерий. Очень многие микроорганизмы имеют одинаковые морфологические признаки, но различаются по строению геномов, родственные связи между ними часто бывают неясными, а эволюция многих просто неизвестна.
Более того, краеугольное для каждой классификации понятие вид для бактерий до сих пор не имеет чёткого определения, а в ряде случаев истинное родство между бактериями может оказаться спорным, поскольку оно лишь отражает общность происхождения от одного далекого предка. Такой упрощённый критерий, как размер, применявшийся на заре микробиологии, в настоящее время абсолютно неприемлем. Кроме того, микроорганизмы значительно различаются по своей архитектуре, системам биосинтезов, организации генетического аппарата. Их разделяют на группы для демонстрации степени сходства и предполагаемой эволюционной взаимосвязи. Базовый признак, используемый для классификации микроорганизмов — тип клеточной организации.
Искусственная (ключевая) систематика микроорганизмов.
Более скромные задачи у искусственной систематики, объединяющей организмы в группы на основе сходства их важнейших свойств. Эту последнюю характеристику применяют для определения и идентификации микроорганизмов. С позиций медицинской микробиологии микроорганизмы обычно подразделяют в соответствии с влиянием, которое они оказывают на организм человека на патогенные, условно-патогенные и непатогенные.
Несмотря на очевидную важность этого утилитарного подхода, их систематика всё же основана на принципах, общих для всех форм жизни. Для облегчения диагностики и принятия решений, касающихся лечения и прогноза заболевания, предложены идентификационные ключи. Сгруппированные в таком ключе микроорганизмы не всегда находятся в филогенетическом родстве, но перечисляются вместе, поскольку обладают несколькими, легко выявляемыми сходными свойствами.
Разработаны разнообразные доступные и быстрые тесты, позволяющие, как минимум в общих чертах, идентифицировать выделенные от пациента микроорганизмы. В отношении бактерий наибольшее распространение нашли предложенные американским бактериологом Дэвидом Бёрджи подходы к систематизации, учитывающие один или несколько наиболее характерных признаков. "Определитель бактерий Бёрджи" — характерный пример искусственной систематики. Согласно его принципам, легко выявляемые свойства являются основой для объединения бактерий в большие группы.
Названия таксонов у микроорганизмов.
Образование и применение научных названий микроорганизмов регламентируют "Международный кодекс номенклатуры бактерий", "Международный кодекс ботанической номенклатры" (грибы), "Международный кодекс зоологической номенклатуры" (простейшие) и решений Международного комитета по таксономии вирусов. Все изменения научных названий микроорганизмов возможны лишь решениями соответствующих международных конгрессов и постоянных комитетов по номенклатуре.
Категории таксономической иерархии.
Для микроорганизмов приняты следующие категории (таксоны) таксономической иерархи (по восходящей): Вид (Species)—> Род (Genus) —> Триба, или колено (Tribus) —> Семейство (Familia) —> Порядок (Ordo) —> Класс (Classis) —> Отдел(Divisio) —> Царство (Regnum). При необходимости вводят категории (по восходящей): Подтриба, или подколено (Subtribus) —> Подсемейство (Subfamilia) —> Подпорядок (Subordo) —> Подкласс (Subclassis) —> Подотдел (Subdivisio).
Названия таксонов у микроорганизмов.
Род и выше. Названия таксонов, имеющих ранг рода и выше, униноминальны (унитарны), то есть обозначаются одним словом, например Herpesviridae (семейство герпесвирусов).
Вид. Названия видов биноминальны (бинарны), то есть обозначаются двумя словами — название рода и вида. Например, Escherichia coli (кишечная палочка). Второе слово бинарного названия вида, взятое отдельно, не имеет статуса в номенклатуре и не может быть использован для научного обозначения микроорганизма. Исключением выступают вирусы, видовые названия которых не бинарны, то есть включают только видовое название (например, вирус бешенства).
Инфравидовые таксоны. Систематика бактерий включает также внутривидовые таксоны названия которых не подчиняются правилам «Международного кодекса номенклатуры бактерий».
Подвид. Названия подвидов триноминальны (тринарны); для их обозначения применяют слово подвид (subspecies) после видового названия, например Klebsiella pneumoniae subsp ozena (палочка озены, где ozenae — название подвида).
Вариант. Разнообразные механизмы изменчивости бактерий приводят к определённой нестабильности признаков, совокупность которых определяет тот или иной вид. Поэтому в систематике бактерий широко применяют понятие «вариант». Различают морфологические, биологические, биохимические, серологические и многие другие варианты.
В медицинской бактериологии обычно выделяют серологические варианты (серовары), варианты, устойчивые антибиотикам (резистенсвары), бактериофагам (фаговары), а также варианты, различающиеся по биохимическим (хемовары), биологическим или культуральным признакам (биовары). Например, Vibrio cholerae биовар Eltor (холерный вибрион Эль-Тор) или Escherichia со серовар 0157:Н7 (представитель группы энтерогеморрагических кишечных палочек).
Штамм и клон. В микробиологии также применяют специализированные термины — «штамм» и «клон».
Штаммом [от нем. stammen, происходить] называют культуру микроорганизмов, выделенную из определённого конкретного источника (какого-либо организма или объект окружающей среды).
Клоном [от греч. klon, отводок] называют культуру микроорганизмов, полученную из одной материнской клетки. Например, вирус гриппа A (H3N2) штамм Ленинград 34/72.