(Sporozoa) Класс Спорозои, Споровики, Class Sporozoa 1 отряд
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
КЛАСС СПОРОВИКИ (SPOROZOA)
Класс споровиков представляет собой обширную группу простейших организмов, ведущих исключительно паразитический образ жизни. Приспособление к паразитизму у них очень глубокое и совершенное. У всех споровиков отсутствуют специальные органоиды захвата пищи, и они питаются осмотическим путем. Хозяевами их являются самые различные беспозвоночные и позвоночные животные. Несколько видов споровиков паразитирует в человеке.
В процессе эволюции они приспособились к паразитированию в самых различных органах и тканях. Многие споровики — паразиты кишечника и различных органов, связанных с пищеварительной системой (в том числе печени). Имеются виды, паразитирующие в органах выделительной системы — почках. Органы кровеносной системы и кровь также служат средой обитания некоторых споровиков.
В этой группе простейших есть немало видов, приспособившихся к внутриклеточному паразитизму. Паразиты проникают внутрь клеток различных тканей хозяина, питаются, растут и развиваются за их счет.
Одной из форм глубокого приспособления споровиков к паразитизму явилась выработка сложных и разнообразных жизненных циклов, обеспечивающих заражение хозяина. По ходу циклов происходит смена разных форм размножения, ведущих, с одной стороны, к увеличению числа паразитов в данной особи хозяина, а с другой — к образованию таких стадий, которые служат для заражения новых особей хозяина.
У некоторых споровиков выработалось не только чередование различных форм размножения, но и смена хозяев, относящихся к разным видам и группам животного мира.
Многие виды споровиков приносят большой вред как возбудители заболеваний человека, домашних и промысловых животных.
ОТРЯД ГРЕГАРИНЫ (GREGARINIDA)
Грегарины — большая группа споровиков, включающая около 500 видов. Все без исключения грегарины — паразиты беспозвоночных. Наибольшее число видов их обитает в кишечнике членистоногих, в особенности часто у насекомых. Некоторые грегарины паразитируют и в других группах беспозвоночных — в кольчатых червях, иглокожих, оболочниках. Кроме кишечных грегарин, каковых большинство, имеются виды, живущие в полости тела, в органах размножения и др.
Наиболее сложно устроены грегарины, живущие в кишечнике членистоногих. На их примере мы и ознакомимся с организацией представителей этого отряда.
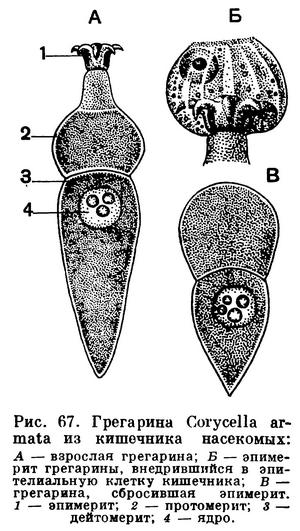
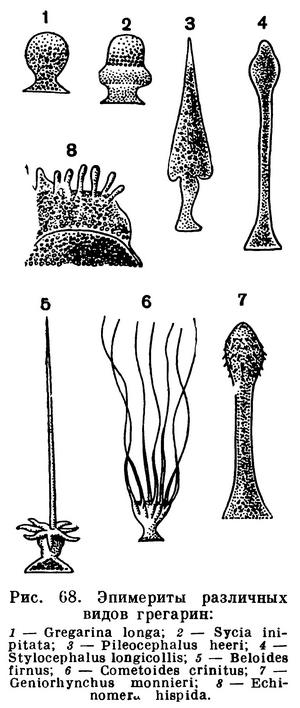
На рисунке 67 показано строение взрослой формы Corycella armata — грегарины из кишечника личинки жука-вертячки Gyrinus natator. Тело ее, достигающее довольно значительной длины, разделено на 3 отдела. В наиболее крупном заднем отделе помещается ядро. Этот отдел называется дейтомеритом. Впереди от него расположен протомерит. Он отделен от дейтомерита тонкой прослойкой светлой цитоплазмы, представляющей собой продолжение слоя эктоплазмы. На самом переднем конце грегарины расположен эпимерит. Этот участок тела служит для прикрепления грегарины к стенке кишечника. Часто он бывает снабжен крючьями, иногда нитевидными выростами и может принимать весьма причудливую форму. На рисунке 68 изображены некоторые характерные формы эпимеритов разных видов грегарин. По достижении предельных размеров, когда у грегарины наступают процессы полового размножения, эпимерит обычно отбрасывается. Он целиком состоит из эктоплазмы.
Разделение цитоплазмы грегарин на два слоя — эктоплазму и эндоплазму — выражено всегда очень отчетливо. Наружный слой эктоплазмы образует прочную пелликулу, определяющую постоянство формы тела грегарин. У некоторых грегарин в эктоплазме найдены тонкие скелетно-опорные волоконца, еще более увеличивающие прочность наружных слоев тела. Немногие виды грегарин способны несколько менять форму своего тела, сокращаться вдоль продольной оси. У таких видов в эктоплазме, на границе с эндоплазмой, найдены сократимые волоконца — миофибриллы.
Эндоплазма грегарины, резко отграниченная от эктоплазмы, при рассматривании под микроскопом живых грегарин выглядит темно-серой. Это обусловлено наличием в эндоплазме большого количества гликогена, нередко сконцентрированного в виде мелких гранул. В этом проявляется одна из характерных особенностей многих внутренностных паразитов, тело которых богато гликогеном. Кишечные паразиты живут в среде, в которой свободный кислород присутствует в ничтожных количествах. Это затрудняет или делает невозможными дыхательные процессы, сопровождающиеся поглощением кислорода. В этих условиях паразитические организмы вынуждены искать взамен дыхания другие источники энергии. Такими источниками оказываются процессы анаэробного (без участия свободного кислорода) расщепления гликогена, процессы гликолиза. He вдаваясь в химическую характеристику этих процессов, отметим лишь, что при этом сложная молекула гликогена распадается на более простые органические соединения (в частности, некоторые органические кислоты). Этот процесс сопровождается выделением энергии. Однако в отношении количества освобождаемой энергии на единицу веса (на грамм-молекулу) гликогена при окислении выделяется во много раз больше энергии, чем при гликолизе. Окисление — гораздо более «экономичный» процесс, чем гликолиз. Поэтому обычно у организмов, основным источником энергии которых являются анаэробные гликолитические процессы, в теле имеются значительные скопления гликогена. Грегарины — один из ярких примеров, иллюстрирующих это правило.
Ядра, расположенные всегда в дейтомерите, имеют обычно вид пузырька, богатого ядерным соком и снабженного одним или несколькими крупными ядрышками.
Рассмотренное выше строение грегарины свойственно преимущественно кишечным паразитам. Оно характерно для группы видов, объединяемых в семейство Polycystidae. У представителей другого семейства — Monocystidae, которые паразитируют главным образом в полости тела, половых железах и других органах, строение более простое. У Monocystidae нет разделения на эпи-, прото- и дейтомерит, и тело их часто имеет червеобразную форму.
Наблюдая под микроскопом за живыми грегаринами, легко обнаружить, что они способны двигаться, медленно перемещаясь в содержимом кишечника. Это движение носит характер плавного скольжения, не сопровождающегося изменением формы тела.
Механизм этого процесса до настоящего времени не вполне ясен, поскольку у грегарин нет никаких специальных органоидов движения.
Исследования Шевякова и других авторов показали, что на заднем конце движущейся грегарины выделяется слизь. По-видимому, она выбрасывается наружу с известной силой, благодаря чему и происходит — по принципу ракетного двигателя — поступательное движение грегарины.
У настоящих грегарин (Кроме настоящих грегарин (отряд Eugregarinida), имеется небольшой отряд шизогрегарин (Schizogregannida), размножающихся бесполым путем.) отсутствует бесполое размножение, они размножаются только половым путем. Рассмотрим типичный цикл развития грегарин (рис. 69).
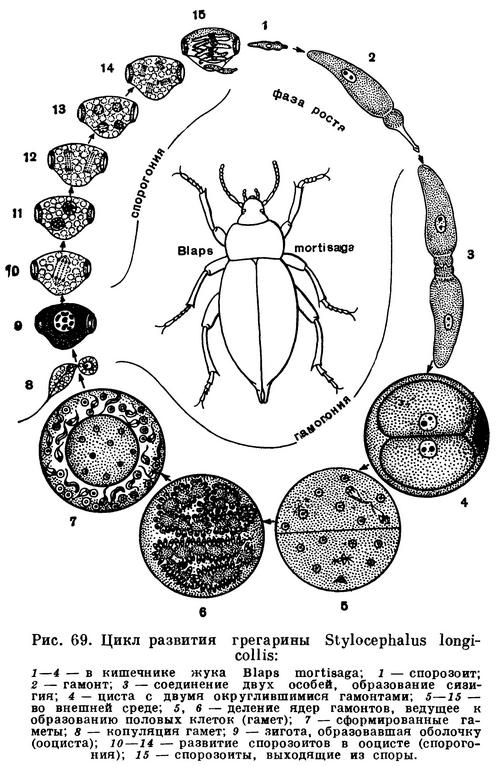
Живущие в кишечнике насекомого грегарины, достигнув предельного роста, теряют эпимерит и соединяются попарно. Вокруг такой парочки (сизигия) выделяется общая оболочка (циста). Вскоре после инцистирования в каждой из соединившихся в сизигий особи начинается многократное последовательное деление ядра. В каждой особи образуется множество ядер. К концу этого процесса все ядра располагаются по периферии грегарины и вокруг каждого из них обособляется небольшой участок цитоплазмы. Так формируется множество мелких одноядерных клеток. Большая часть цитоплазмы при этом остается неиспользованной. Это так называемое остаточное тело, которое в дальнейшем процессе размножения участия не принимает. Мелкие одноядерные клетки представляют собой половые клетки — гаметы. Эти клетки сближаются попарно и сливаются.
У некоторых видов грегарин гаметы, образуемые обоими соединившимися в сизигий особями, по размерам и строению неотличимы друг от друга. У других видов они различны. Одна особь дает начало округлым и неподвижным гаметам. Это женские гаметы. Другая особь образует удлиненные гаметы, снабженные жгутиковидным выростом. Это мужские подвижные гаметы.
Образовавшиеся в результате оплодотворения зиготы сразу же покрываются оболочкой, принимая при этом веретенообразную, а нередко бочонковидную форму. Покрывшаяся оболочкой зигота носит название ооцисты. Оболочка цисты в этот период цикла развития еще не разрушается. Таким образом, многочисленные образовавшиеся после копуляции гамет ооцисты лежат внутри оболочки цисты. На этой стадии, а иногда и несколько позже, цисты с ооцистами выводятся из кишечника наружу. Последним этапом цикла развития грегарины является процесс формирования ооцисты, получивший название спорогонии. Он сводится к следующему. Внутри ооцисты ядро (образовавшееся в результате слияния ядер гамет) трижды делится. Вокруг каждого из возникших таким путем ядер обособляется участок цитоплазмы. В каждой ооцисте получается восемь очень маленьких червеобразной формы клеток, называемых спорозоитами. На этой стадии ооцисты способны к заражению новых особей хозяина. Если циста с ооцистами или даже отдельная ооциста будет проглочена насекомым — хозяином данного вида грегарин, то под воздействием кишечного сока спорозоиты выходят из ооцисты. Они подвижны. В дальнейшем из каждого спорозоита развивается грегарина. Достигнув типичного для данного вида размера, грегарины вновь соединяются попарно, инцистируются, и цикл начинается снова.
Таким образом, рассмотренный выше цикл развития грегарин можно отчетливо разделить на три этапа. Первый — это рост и развитие взрослой грегарины. Второй — образование половых клеток (гамет) и оплодотворение. Третьим этапом цикла является спорогония — развитие зиготы (ооцисты) и формирование спорозоитов.