(Arthropoda von Siebold, 1848) Тип Артроподы, Членистоногие, Phylum Arthropoda von Siebold, 1848 (Arthropods) 17 классов
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
Среди всех групп беспозвоночных животных тип членистоногих выделяется наибольшим разнообразием приспособлений к самым различным условиям существования, изумительным богатством форм и огромным числом видов. Число видов членистоногих приближается к 3 миллионам и намного превышает число видов всех остальных типов животных и растений, вместе взятых, включая микроорганизмы. Особенно обильны видами насекомые, на долю которых падает более 90% известных видов членистоногих.
Именно тонкими приспособлениями к использованию всевозможных местообитаний, всяких источников пищи, часто недоступных для других животных, объясняется многообразие и обилие видов членистоногих.
Трудно найти такие места, где не было бы членистоногих. Они живут во всех морях и океанах — и в толще воды, и на дне, и в грунте на разных глубинах; они обитают и во всевозможных пресных водоемах — не только в реках и озерах, но и в мелких пересыхающих лужах, в подземных водах и пещерах, в скоплениях воды в дуплах деревьев. Некоторые виды приспособились к жизни в очень соленых водоемах, другие — в горячих источниках. Кажется, все разнообразие водной среды на поверхности нашей планеты освоено членистоногими.
Большая армия представителей этого типа приспособилась и к жизни вне воды — на суше, где членистоногие особенно многообразны. Весь слой почвы, пронизанный корнями растений, обильно заселен и членистоногими — во многих лесных и луговых почвах миллион особей клещей и ногохвосток на 1 м2 не редкость. Огромное количество членистоногих обитает на поверхности различных частей растений и внутри растительных тканей.
Многие членистоногие — паразиты, постоянные или временные, наружные или внутренние, как других видов членистоногих, так и животных, относящихся к другим типам, причем, например, среди насекомых число видов паразитов не многим, вероятно, уступает числу остальных видов. Иногда членистоногие столь изменяются под влиянием паразитического образа жизни, что даже перестают быть похожими на членистоногих.
Какими только способами, какими только веществами членистоногие не питаются. Одни фильтруют воду, потребляя мелкие планктонные организмы и мелкие взвешенные частицы разлагающихся остатков организмов; другие роются в грунте или почве, пропуская через кишечник большое количество минеральных частиц, чтобы усвоить рассеянные между ними органические вещества; есть членистоногие, которые питаются такими трудноперевариваемыми веществами, как рог и воск; есть существующие за счет клетчатки. Некоторые способны усваивать с помощью симбиотических микроорганизмов атмосферный азот. Огромное число представителей разных классов членистоногих — хищники и паразиты; для наземных членистоногих, особенно насекомых, один из основных источников пищи — ткани живых высших растений.
Во всех климатических зонах можно встретить членистоногих. Огромно их количество и велико многообразие в тропиках, но есть они и в высоких широтах Арктики и на побережье Антарктиды, где недавно были обнаружены новые виды клещей и ногохвосток. Членистоногих можно встретить и в самых сухих пустынях. Образно можно сказать, что членистоногие — это тип животных, который по полноте овладения поверхностью земного шара не имеет равных.
По многим признакам строения членистоногие близки к кольчатым червям, особенно к полихетам. Замечательный зоолог начала прошлого века Жорж Кювье, предложивший деление животных на типы, объединил кольчатых червей и членистоногих в единый тип членистых животных (Articulata). Знаменательно, что и в наши дни некоторые авторитетные зоологи, например В. Н. Беклемишев, ратуют за восстановление типа членистых. Это показывает, что членистоногие во всяком случае очень близки к многощетинковым кольчатым червям. От близких к ним форм когда-то давно, еще в те геологические эпохи, в отложениях которых не сохранилось ископаемых остатков, отделились предки современных членистоногих.
Членистоногие, как и кольчатые черви, — это двустороннесимметричные сегментированные животные. Сходство членистоногих с кольчецами проявляется не только в наружной сегментации. Центральная нервная система у более примитивных членистоногих очень сходна с лестничной нервной системой полихет.
Специфические признаки членистоногих связаны с тем, что у них тело покрыто очень плотной и прочной кутикулой, которая нередко образует очень твердый панцирь. Кутикула у членистоногих выделяется наружным слоем клеток, так называемой гиподермой, имеет очень сложное строение и сложный химический состав. Раньше считали, что у членистоногих кутикула состоит только из хитина. Теперь выяснилось, что, кроме хитина, в состав ее входят многие другие соединения, в основном задубленные белки, и что у большинства членистоногих на долю хитина падает не больше трети веществ, из которых состоит кутикула. Хитин — это сложный полисахарид, в состав молекулы которого входят атомы азота. Вероятно, это не определенное соединение, а группа близких соединений, формулы которых точно не установлены. Хитин эластичен, прочен и устойчив к химическим воздействиям. Нередко в кутикуле содержится много углекислого кальция.
Прочность и твердость кутикулы членистоногих настолько велики, что в тех местах, где она достигает большой толщины, покровы становятся негибкими. Подвижность сегментов тела друг относительно друга, обеспечивающая движение, у членистоногих достигается тем, что между твердыми участками (склеритами) сохраняются перепончатые участки с очень тонкой кутикулой; такие перепонки на сгибах образуют складки, расправляющиеся при растягивании тела. Далеко не все сегменты у членистоногих подвижны друг относительно друга, очень часто наблюдается полное слияние соседних сегментов, особенно в передних отделах тела.
В теле членистоногих обычно можно различить 3 отдела — голову, на которой находятся основные органы чувств, служащие для ориентировки в пространстве (глаза, усики и др.), и органы захватывания пищи. Сегменты головы у членистоногих слитные. Далее следует грудь — тот отдел тела, на котором располагаются основные двигательные придатки — ноги, а у насекомых и крылья. Задний отдел тела называется брюшком. У некоторых членистоногих (например, у многоножек, а из ископаемых — у трилобитов) все туловищные сегменты более или менее однородны, и тогда в теле животного различают только 2 отдела — голову и туловище. У многих членистоногих (у паукообразных, десятиногих раков) сливаются головной и грудной отделы, образуя головогрудь. А у клещей вообще утрачиваются границы между отделами тела, тело слитное.
Передвигаются членистоногие с помощью конечностей.
Поскольку конечности у них, как и все тело, покрыты кутикулой, их движение возможно тоже только при разделении ног на членики. По строению конечностей, на которые опираются членистоногие при движении по твердой поверхности или с помощью которых они плавают, весь тип и получил свое название.
Некоторое количество пар конечностей на голове у всех членистоногих превращается в ротовые органы — челюсти, использующиеся хотя бы частично для захватывания или размельчения пищи. Это тоже признак, характерный для типа членистоногих. Ротовые части бывают настолько видоизменены, что их трудно даже признать конечностями, например, хоботок комара или клопа. Число пар ног, которые служат для передвижения, неодинаково у разных членистоногих. Для насекомых характерно развитие 3 пар двигательных конечностей, для паукообразных — 4 пар, а у некоторых многоножек число ног превышает сотню.
Нередко туловищные конечности служат не только как ноги, но и как жабры, присоски и другие органы.
У членистоногих двигательная мускулатура состоит из отдельных мышц, концы которых присоединяются к разным утолщенным участкам покровов, отделенным друг от друга мягкой перепонкой. Движение членистоногих обеспечивается не изгибами тела при сокращении всей мускулатуры туловища, как у червей, а сокращениями обычно немногих мышц, связанных с конечностями или другими двигательными придатками (крылья насекомых, вилочка ногохвосток и др.). Мышцы членистоногих обладают исключительно высокой способностью к сокращению. Все они поперечнополосатые, причем обнаруживаемая под микроскопом поперечная исчерченность мышечных волоконец у членистоногих выражена больше, чем у позвоночных,— мышцы членистоногих сильнее наших, если, конечно, учитывать размеры. Так как панцирь членистоногих служит и местом прикрепления двигательных мышц, он является и скелетом, называемым «наружным» в отличие от привычного нам внутреннего скелета позвоночных.
Панцирь членистоногих лишь в результате долгой эволюции стал скелетом. А сначала утолщение и упрочнение покровов у обитавших в морях далеких червеобразных предков членистоногих служило, надо полагать, приспособлением к защите внутренних органов от повреждений. Утолщение и уплотнение кутикулы делает ее менее проницаемой. Поэтому понятно, что только очень мелкие членистоногие, имеющие тонкие покровы, живущие в воде или в очень влажных местах, могут дышать всей поверхностью тела.
У всех более крупных членистоногих, имеющих и более плотные покровы, обязательно развиваются специальные дыхательные органы, поверхность которых покрыта такой тонкой кутикулой, что через нее легко может поступать кислород.
У живущих в воде членистоногих это жабры. Для дыхания в воде важно, чтобы поверхность жабр была возможно большей. Поэтому жабры всегда представляют собой тонкие листовидные выросты или пучки пальцевидных отростков. Обычно жабрами у членистоногих становятся конечности или части конечностей. В воде благодаря большой ее плотности даже тоненькие жаберные листочки не слипаются друг с другом. Поэтому на небольшом участке тела (в остальных местах защищенного плотными покровами) может разместиться много листовидных жабр.
Для доставки кислорода от жабр ко всем удаленным от них органам служит кровеносная система и полостная жидкость. У членистоногих кровеносная система представлена расположенным в спинной части тела пульсирующим, обычно продолговатым сосудом — сердцем. Кровь поступает в сердце через закрывающиеся клапанами боковые отверстия, а выталкивается из него в полость тела или непосредственно, или через немногие крупные сосуды — система кровообращения незамкнутая. У крупных морских членистоногих, имеющих толстый панцирь, — у омаров, у мечехвостов — в крови есть дыхательный пигмент гемоцианин, сходный по химическому составу с нашим гемоглобином, но придающий крови не красный, а синеватый цвет: в состав гемоцианина входит не железо, как в гемоглобине, а медь. Гемоцианин легко присоединяет кислород при прохождении крови через жабры и отдает его внутренним органам. Так как у членистоногих кровеносная система незамкнутая, правильнее называть их «кровь» «гемолимфой», но и термином «кровь» зоологи широко пользуются.
На открытой поверхности суши ни кожное дыхание, ни дыхание с помощью жабр невозможно — легкопроницаемые участки покровов пропускают воду и быстро подсыхают, теряя проницаемость. Поэтому водные организмы, извлеченные на сушу, погибают от потери воды или от недостатка поступающего кислорода. На суше только скрытоживущие мелкие членистоногие, обитающие в таких местах, где воздух всегда насыщен водяным паром, например в почве, могут дышать всей поверхностью тела (мелкие клещи, ногохвостки, симфилы). Жизнь на суше вне влажных укрытий требует развития непроницаемых для испарения покровов, а такие покровы не пропускают и кислород.
У наземных членистоногих развиваются особые дыхательные органы — либо «легкие», представляющие глубокие мешковидные впячивания со складчатыми стенками, открывающиеся наружу лишь небольшими отверстиями (у пауков, у скорпионов), либо (у большинства наземных членистоногих) трахеи. Трахеи представляют собой тонкие, обычно обильно ветвящиеся трубочки, открывающиеся наружу маленьким входным отверстием («дыхальцем») на поверхности тела. Ветви этих трубочек омываются полостной жидкостью, а самые мелкие ответвления доходят до отдельных клеток. Из тонких трубочек трахей вода через дыхальца испаряется очень медленно, и потери ее незначительны, а кислород по ним проникает быстро и легко.
Уже далекие предки разных групп членистоногих приспособились к жизни в разных средах. Можно выделить группу водных членистоногих, к которым относятся ракообразные, выделяемую в подтип жабродышащих. Правда, среди современных ракообразных есть и такие, которые живут на суше (мокрицы), но по всем признакам они очень близки к своим водным родичам. Другой подтип — трахейнодышащие — представлен типично наземными членистоногими — различными группами многоножек и насекомых. Они прекрасно приспособлены к жизни на суше. В их покровах содержатся воско- и жироподобные вещества, защищающие организм от высыхания, а все системы органов, связанных с расходованием воды, — выделительная и дыхательная — действуют так, что влага расходуется минимально. Среди насекомых есть отдельные виды, проводящие всю жизнь или часть ее в воде, но строение таких водных насекомых явно показывает их происхождение от обитавших на суше предков.
За длившуюся сотни миллионов лет историю эти группы обособились в связи с приспособлениями к жизни в двух разных средах — в воде и на суше.
Третий подтип членистоногих — хелицеровые — очень существенно отличается и от ракообразных, и от трахейнодышащих. Среди хелицеровых есть и водные представители — мечехвосты, обитающие в морях, и наземные — паукообразные.
В ископаемом состоянии сохранилось много остатков хелицеровых. Их изучение, дополняющее изучение современных представителей, показывает, как шло разделение классов в результате приспособления к жизни в разных средах — в воде и на суше. Изучение всех групп членистоногих убеждает в том, что переход их к жизни на суше был связан со способностью к выработке предохраняющих от испарения влаги покровов, к сохранению в теле воды, без которой невозможна жизнь.
У членистоногих, хорошо приспособившихся к жизни на суше, панцирь бывает нередко тоньше, чем у обитающих в воде, но зато на его поверхности отлагается очень тонкий слой воскоподобных и жироподобных веществ, не пропускающих пары воды. Этот слой защищает организм от высыхания и препятствует испарению. Поэтому у всех членистоногих, живущих открыто на поверхности суши, есть специальные органы дыхания. Те членистоногие, у которых специальных органов дыхания нет, которые дышат всей поверхностью кожи, вынуждены обитать во влажных укрытиях — в почве, в лесной подстилке и т. д., только иногда по ночам и после сильных дождей показываясь на поверхности.
У наилучшим образом приспособившихся к жизни на суше членистоногих чаще всего развивается трахейная система. Не только дыхательная, но и другие системы органов меняются у членистоногих с переходом к жизни на суше.
Выведение растворенных вредных продуктов разрушения белков у водных членистоногих происходит непосредственно во внешнюю среду через отверстия выделительных органов, открывающиеся у основания конечностей головного или грудного отдела. А у всех членистоногих, наилучшим образом приспособившихся к жизни на суше, — у пауков, многоножек, насекомых — выделительные трубочки («мальпигиевы сосуды») открываются в начале задней кишки. По мере прохождения по ней непереваренных остатков пищи и продуктов выделения и из тех и из других стенки задней кишки всасывают воду, столь труднодоступную для жителей суши, а экскременты и продукты обмена выводятся из кишечника в обезвоженном состоянии.
Приспособившиеся к жизни на суше членистоногие выработали способность запасать много жира в особых образованиях — в жировом теле. Главная роль жирового тела состоит в том, чтобы в критические периоды, когда организм не может получить воду извне, снабжать его... водой!
При окислении жиров образуется много воды (так называемая «метаболическая вода»), которая и идет на покрытие расхода влаги животным. В этом отношении насекомые сходны с верблюдами, у которых при длительных переходах через пустыню используется вода, образующаяся при окислении запасов жира в горбу. У водных групп членистоногих (ракообразных, мечехвостов) жирового тела не бывает.
Исключительно многообразны у членистоногих типы питания и характер пищи, от которых зависит строение ротовых органов и кишечника. При переходе к жизни в воздушной среде большую роль в завоевании членистоногими суши сыграло приспособление к питанию высшими растениями. Цветковые растения черпают влагу из почвы и сами становятся источником воды для наземных животных. Представители многих групп насекомых, некоторые клещи и совсем немногие многоножки и мокрицы способны к питанию живыми тканями высших растений. Многие наземные членистоногие питаются разлагающимися тканями растений, а многие — хищники или паразиты. Но все они, завоевывая сушу, следовали за высшими растениями — не только создателями поедаемых питательных веществ, но и живыми резервуарами влаги.
Наружный панцирь членистоногих настолько плотен, что в затвердевшем состоянии не растягивается. Поэтому рост членистоногих сопровождается линьками. У растущего животного старый панцирь становится тесным, лопается, обычно сперва на спине, и сбрасывается. И только в короткий период, пока новый панцирь, образовавшийся под ним, не затвердеет, животное может увеличиваться в размерах. Во время сбрасывания старых покровов, которые сходят не только со всей наружной поверхности тела, но и со всех конечностей, в том числе и с ротовых органов, а также с передней и задней кишки и со всех дыхательных органов, членистоногие бывают вялыми, малоподвижными, не питаются и стараются спрятаться в каких-либо укрытиях.
Рост у членистоногих — не непрерывный процесс, а ступенчатый, размеры тела увеличиваются скачкообразно — сразу после линьки. У многих членистоногих во время линьки происходит и большее или меньшее изменение строения тела. Часто взрослое животное приспособлено к совсем иному образу жизни, чем молодое, и потому особи разных возрастов (возрастом у членистоногих называется период между линьками) бывают даже совсем непохожи друг на друга. В таком случае развитие сопровождается метаморфозом. Превращение гусеницы в куколку, а куколки в бабочку может служить общеизвестным примером.
Образ жизни разных членистоногих неодинаков. Многие из них отличаются очень сложным поведением, что всегда связано с высоким развитием органов чувств и центральной нервной системы.
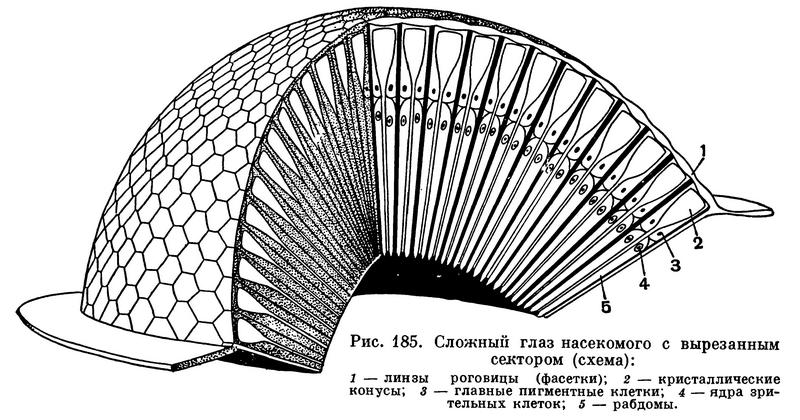
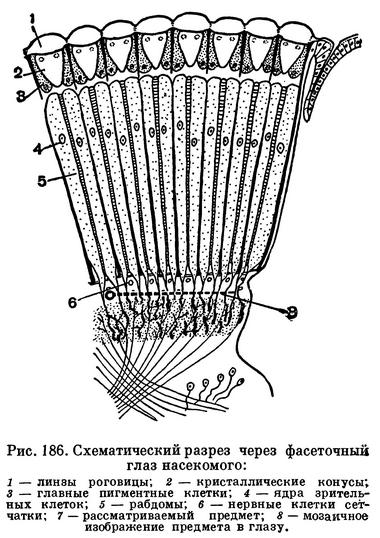
У большинства членистоногих хорошо развиты глаза. Глаза у членистоногих бывают простые, имеющие одну линзу, и сложные, или фасеточные. У высших представителей всех подтипов глаза сложные, фасеточные, состоящие из множества расположенных плотно рядом друг с другом маленьких глазков. Иногда число глазков (так называемых омматидиев) в фасеточном глазу измеряется несколькими тысячами (рис. 185). Фасеточные глаза характерны для высших ракообразных, мечехвостов и насекомых. Такие глаза были и у живших в палеозойскую эру трилобитов — членистоногих, полностью вымерших в конце этой эры. Каждый омматидий воспринимает только одну точку находящегося перед ним предмета (рис. 186). В результате сложный глаз дает изображение, состоящее из большого числа отдельных точек, т. е. мозаичное (как тоновое клише в книге) и прямое (рис. 187). Вспомним, что изображение, получаемое в нашем глазу, перевернутое и цельное!

Глаза членистоногих, оказывается, могут воспринимать не только те лучи, которые видимы для нашего глаза, но и ультрафиолетовую часть спектра. Например, многие цветы, которые нам кажутся равномерно белыми, для насекомых оказываются пестрыми; у таких цветов есть не видимый нами ультрафиолетовый рисунок.
И наземные и водные членистоногие способны различать обычный свет и так называемый поляризованный (те световые лучи, электромагнитные колебания которых совершаются в одной плоскости). Поскольку для рассеянного света неба характерна определенная степень поляризации лучей в меридиональной плоскости, членистоногие по небу, как по компасу, могут ориентироваться и в ясные и в пасмурные дни. Таким образом, членистоногие различают не только частоту, но и плоскость колебаний световых лучей.
Мы не ощущаем магнитные волны. А например, самки термитов в гнезде лежат так, что ось их тела направлена по магнитному меридиану, а если поместить рядом сильный магнит, меняют свое положение. Как воспринимают членистоногие магнитные волны, пока не выяснено.
Отмечено, что членистоногие могут реагировать и на короткие радиоволны.
Хорошо развит и слух у членистоногих. Звуковые волны определенной длины (чаще короткие) они воспринимают с помощью тонких слуховых волосков на поверхности тела, особенно на усиках, с помощью
специальных слуховых органов, напоминающих по строению струну, натянутую над резонаторной ямкой (как струна на балалайке), и других приспособлений. Часто наземные членистоногие издают ультразвуки, которых мы не слышим, с помощью различных «стридуляционных» органов, по принципу действия напоминающих струну и смычок.
Особенно замечательна способность членистоногих воспринимать запахи — ничтожные концентрации многих веществ, растворенных в воде или взвешенных в воздухе. Никакие наши химические приборы не дают возможности уловить такие слабые концентрации вещества, а насекомые, например, могут по ним ориентироваться. Бабочки совки могут прилетать за километр на тарелку с бродящей патокой, а самцы шелкопрядов летят за километры на не ощущаемый нами и вблизи запах коробки, в которой сидела самка. Не менее тонко восприятие и запахов, и содержания солей водными членистоногими. Многие наземные членистоногие (мокрицы, личинки насекомых) способны улавливать разницу в относительной влажности воздуха порядка 1—2%.
У водных членистоногих прекрасно развиты органы равновесия.
Наиболее универсальное чувство, присущее всем членистоногим, — осязание. Осязательную функцию выполняют многочисленные волоски на поверхности тела членистоногих. Для многих членистоногих, особенно для донных, почвенных и живущих в тканях растений и животных, характерно стремление к наибольшей поверхности контакта с твердыми предметами — «тигмотаксис».
Многообразная «информация», получаемая из внешнего мира органами чувств членистоногих, требует «обработки информации» центральной нервной системой. Головной мозг — парные скопления нервных клеток над пищеводом — у высших активных членистоногих очень хорошо развит, в нем различается несколько отделов, связанных с основными органами ориентировки в пространстве, расположенными в головном отделе.
Чем сложнее поведение членистоногого, чем многообразнее те условия, с которыми животное сталкивается, и чем сложнее ответы организма на воздействия среды, тем более сложно строение головного мозга. Головной мозг у членистоногих связан окологлоточным нервным кольцом с брюшной нервной цепочкой. Исходным у членистоногих, по-видимому, является соответствие каждому сегменту тела пары ганглиев брюшной нервной цепочки. Но при слиянии отдельных сегментов и даже отделов тела друг с другом происходит уменьшение числа нервных узлов, частично сливающихся друг с другом, частично исчезающих. Степень расчленения или слитности брюшной цепочки связана со степенью расчленения или слитности тела, и концентрация узлов брюшной нервной цепочки совсем не может служить показателем сложности поведения.
Поведение членистоногих, особенно в период размножения (спаривание, забота о потомстве), бывает очень сложным, в чем можно убедиться из знакомства с приводимыми ниже описаниями образа жизни разных представителей этого типа. Очень часто поведение определяется прямой реакцией на раздражение, например приближение к источнику света («положительный фототаксис») или уход от света («отрицательный фототаксис»). Еще более распространены реакции на раздражение путем усиления или ослабления подвижности («кинезисы»). Например, многие насекомые активнее двигаются при более сильном или слабом освещении и малоподвижны при среднем. Таким образом, после ненаправленного быстрого движения они, наконец, попадают в благоприятные для себя условия освещения и перестают двигаться.
Кроме непосредственных стандартных ответов на раздражения (безусловных рефлексов), для членистоногих очень характерны и сложные цепи наследственно закрепленных рефлекторных действий (инстинкты). Инстинкты представляют такое сложное поведение, какое в нормальных для данного вида условиях является целесообразным, но оно остается неизменным и при изменении условий и поэтому может становиться нецелесообразным.
Например, роющие осы вырывают норку, приносят в нее парализованное насекомое, которым должна питаться личинка, откладывают на добычу яйцо и заделывают норку. Все действия осы «целесообразны». Но если в процессе деятельности осы, когда она приступает к заделыванию норки, вынуть и добычу и яйцо, оса завершает свою уже ненужную работу по замуровыванию норки.
Наряду со сложными формами инстинктивного поведения у членистоногих (высшие ракообразные, пауки, высшие насекомые) прослеживаются и индивидуальные навыки; у них можно выработать и условные рефлексы. Мало того, например, пчелы могут передавать друг другу приобретенные навыки, обмениваться информацией. Определенными телодвижениями рабочая пчела сообщает другим пчелам в улье, в каком направлении надо лететь за взятком.
Членистоногим, несомненно, свойственны многие проявления высшей нервной деятельности, которую еще недавно считали характерной только для позвоночных.
Все членистоногие размножаются половым путем — обычно с оплодотворением, но иногда и без оплодотворения (партеногенетически). Большинство членистоногих раздельнополые, но некоторые, ведущие неподвижный образ жизни (например, усоногие ракообразные) или паразитические формы (например, некоторые равноногие ракообразные), — гермафродиты.
Типы оплодотворения у членистоногих разнообразны.
У водных членистоногих, например мечехвостов, бывает настоящее наружное оплодотворение, когда самка откладывает яйца в воду, а самец туда же выводит семенную жидкость. Проникновение сперматозоида в яйцо происходит при наружном оплодотворении во внешней среде — в воде.
Часто при наружном оплодотворении у водных членистоногих самец прикрепляет «пакеты» с семенной жидкостью (сперматофоры) к выводным половым отверстиям самок — откладываемые яйца оплодотворяются во внешней среде, но вероятность оплодотворения каждого яйца в таком случае увеличивается.
С переходом к жизни на суше низшие членистоногие, обитающие во влажной среде, — в почве, в гнилой древесине и т. п. (многие клещи, скорпионы, ложноскорпионы, многоножки, ногохвостки и др.), выработали новый способ оплодотворения. Самцы откладывают сперматофоры во внешнюю среду, как при наружном оплодотворении (на землю, на остатки растений), а самки подбирают их своими половыми отверстиями, и, таким образом, проникновение сперматозоидов в яйцо осуществляется внутри организма самки, как при внутреннем оплодотворении. Такое оплодотворение называется наружно-внутренним.
У обитающих на поверхности суши в открытой атмосфере членистоногих оплодотворение внутреннее: самцы вводят семенную жидкость непосредственно в половые отверстия самок. Проникновение сперматозоида в яйцо происходит в защищенных от высыхания условиях внутри материнского организма.
В отдельных группах водных членистоногих, например у усоногих раков, также выработалась способность к внутреннему оплодотворению, повышающая плодовитость.
У большинства членистоногих наблюдается откладка яиц, живорождение наблюдается редко, например у некоторых двукрылых насекомых. Чаще наблюдается яйцеживорождение: самка рождает живых детенышей, однако развитие детенышей происходит за счет питательных запасов яйца, но не во внешней среде, а в яйцеводах самок (например, у тлей).
Как упоминалось, развитие многих членистоногих происходит со сложным превращением. В тех случаях, когда взрослая особь и форма, вышедшая из яйца, резко различаются по строению, молодые особи называются личинками.
У малоподвижных или неподвижных морских ракообразных расселяются подвижные личинки. У очень подвижных во взрослом состоянии крылатых насекомых личинки — в основном питающаяся стадия.
В тех случаях, когда яйца богаты желтком, из них выходят молодые членистоногие, уже похожие на взрослых. Такое развитие характерно для многих пресноводных ракообразных, паукообразных и некоторых насекомых.
Членистоногие — древние животные.
Самые достоверные данные, на основании которых можно судить о происхождении тех или иных групп животных, дают исследования ископаемых окаменевших остатков организмов или их отпечатков в слежавшихся и окаменевших наслоениях ила, песка или глины, под наносами которых трупы животных были погребены. Но не от всех прошедших эпох, когда уже существовала на Земле жизнь, сохранились ископаемые остатки организмов. Отложения самых древних периодов развития жизни на Земле подвергались в течение последующих эпох таким сильным изменениям, что точно установить, какие организмы обитали на Земле около миллиарда лет тому назад, невозможно, сколько-нибудь уцелевших остатков животных из отложений более древних, чем палеозойские, не сохранилось. А уже в самых глубоких («нижних») осадочных породах кембрийского периода палеозойской эры (отлагавшихся более полумиллиарда лет назад) встречаются трилобиты — животные с явно выраженными признаками членистоногих, имевшие развитой наружный скелет, расчлененные конечности, сложные глаза и другие свойственные представителям типа Arthropoda признаки организации. Трилобиты — класс членистоногих, полностью вымерший в конце палеозойской эры (длившейся 300—350 миллионов лет).
Из кембрийских отложений известны и ракообразные, некоторые из которых относятся к современным отрядам, с силурийского периода известны родичи мечехвостов и даже скорпионов, в девонских отложениях были найдены остатки ногохвосток, а недавно московский палеонтолог Б. Б. Родендорф установил, что в девоне жили и крылатые насекомые. Все эти данные свидетельствуют о том, что тип членистоногих отделился от каких-то животных, похожих на кольчецов, очень давно. Поэтому точно установить родственные связи различных членистоногих очень трудно. Несомненно, что одной из первых оформившихся групп членистоногих были трилобиты. В палеозое трилобиты достигли расцвета, они буквально кишели в прибрежной зоне морей и океанов 500 миллионов лет назад. Трилобиты сохранились в ископаемом состоянии настолько хорошо, что по некоторым остаткам удалось не только исследовать их внутреннее строение, но на шлифах изучить даже микроскопическую структуру тканей этих животных, а также проследить их развитие. А применяя некоторые методы современной химии, удалось даже выяснить, что в их крови был гемоцианин, как у современных высших ракообразных.
Трилобиты, обычно выделяемые в отдельный класс членистоногих, заслуживают выделения в самостоятельный подтип Trilobita.
Из современных групп членистоногих наиболее близки к трилобитам, по-видимому, мечехвосты, личинки которых несколько сходны с трилобитами, а зародыши — с личинками трилобитов («протаспис»).
Мечехвосты и паукообразные имеют ряд общих черт строения, позволяющих их объединить в единый подтип хелицеровых (Chelicerata).
По характеру органов дыхания и некоторым другим признакам некоторые общие с трилобитами черты имеют и ракообразные (класс Crustacea). Однако непосредственных родственных связей между трилобитами и ракообразными установить нельзя. Ракообразные представляют особый подтип жабродышащих (Branchiata) в типе членистоногих. А так как среди современных ракообразных многие имеют более примитивное строение, чем хелицеровые, принято рассмотрение типа членистоногих начинать с них.
Последний подтип — трахейнодышащие (Tracheata) — объединяет различных многоножек и насекомых, т. е. типично наземных членистоногих.
В настоящее время не вызывает сомнений происхождение насекомых от каких-то древних членистоногих, близких к многоножкам, а многоножек — от примитивных червеобразных членистоногих.
Вероятно, водные хелицеровые произошли от общих предков с трилобитами — это была другая ветвь эволюционного развития членистоногих. А приспособившиеся к жизни в водной среде ракообразные, сохранившие много черт древних членистоногих, представляют третью основную ветвь развития членистоногих (рис. 188).
Знакомство с типом и с его многообразными представителями удобнее всего начинать с подтипа жабродышащих, с класса ракообразных.
ПОДТИП ЖАБРОДЫШАЩИЕ (BRANCHIATA)
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
Бесконечно разнообразны водоемы, существующие на нашей Земле, — от необозримого океана, глубина которого достигает почти 11 км, до маленьких пересыхающих луж и капиллярных вод, пропитывающих морской и речной песок. Различны их температура, соленость, кислотность, газовый режим и все остальные условия обитания в них живых организмов.
Немногие группы многоклеточных животных приспособились к жизни во всех этих столь разнообразных условиях. Большинство обитает или в морской, или в пресной воде, или на дне, или в толще воды, ведет или свободный, или паразитический образ жизни. Этого никак нельзя сказать о ракообразных.
Ракообразные населяют любые существующие на Земле водоемы, пресноводные и морские, маленькие и большие, живут на дне и в толще воды, проникают в подземные воды, опускаются на предельные глубины Мирового океана. Большинство ракообразных свободно передвигается по дну или в толще воды, но среди них есть и сидяче-прикрепленные виды, например морские желуди и морские уточки, есть и паразиты, прикрепляющиеся к рыбам и другим водным животным. Водная среда — царство ракообразных, однако некоторые из них приспособились и к жизни на суше. Таковы общеизвестные мокрицы, некоторые тропические крабы и раки-отшельники.
Многие ракообразные чрезвычайно многочисленны. Морской планктон под всеми широтами и на всех глубинах в основном (не менее чем на 90% по весу) состоит из ракообразных. Очень велико значение ракообразных и в планктоне пресных вод. Донные ракообразные также нередко встречаются в массовом количестве.
Ракообразные играют очень важную роль в экономике природы. Органическое вещество в водоемах создается главным образом за счет жизнедеятельности микроскопических водорослей. Ракообразные поедают эти водоросли и сами, в свою очередь, поедаются рыбами. Они выступают в качестве посредников, делающих создаваемое в водоемах органическое вещество доступным для рыб. С другой стороны, они используют в пищу огромные массы погибших водных животных, обеспечивая таким образом очищение водоемов.
Существование почти всех рыб, как морских, так и пресноводных, в значительной степени зависит от ракообразных. Некоторые рыбы, как например сельдь, всю свою жизнь питаются планктонными ракообразными, другие используют их только вскоре после выхода из икринки, а затем переходят на какую-нибудь другую пищу. Для гигантов моря — беззубых китов — ракообразные служат основной пищей.
Многие ракообразные непосредственно используются человеком в качестве высокоценного пищевого продукта. Во многих странах развит промысел креветок, крабов, омаров, лангустов и некоторых других съедобных видов. В последнее время проводятся успешные опыты использования морских планктонных ракообразных для добывания витаминов, жиров и других важных веществ. На рыбоводных заводах разводятся некоторые виды ракообразных, необходимые для кормления молодых рыб.
Размеры тела ракообразных колеблются в очень широких пределах — от долей миллиметра до 80 см. Столь же разнообразны форма их тела, строение его частей, окраска и другие признаки.
В настоящее время известно более 20 000 видов ракообразных.
Тело ракообразных, так же как и других членистоногих, обычно покрыто хитиновым панцирем. Он отсутствует или слабо выражен только у паразитических и сидяче-прикрепленных видов. Панцирь прекрасно защищает животных от всевозможных вредных для них физических и химических воздействий, но, с другой стороны, препятствует их беспрерывному росту. Поэтому время от времени ракообразные линяют — сбрасывают старый панцирь и, пока не затвердел новый, быстро растут. Многие виды после линьки оказываются совершенно беспомощными и вынуждены укрываться от врагов в каких-нибудь убежищах.
Тело ракообразных состоит из отдельных сегментов, частично срастающихся друг с другом. Каждый сегмент снабжен парой членистых конечностей, но нередко в процессе эволюции некоторые конечности исчезают. Сегменты тела группируются в три отдела, имеющиеся и у других членистоногих, — голову, грудь и брюшко.
Голова представляет собой результат слияния головной лопасти и 5 следующих за ней сегментов и снабжена соответственно 5 парами придатков — передними и задними антеннами, находящимися у взрослых особей перед ротовым отверстием (только у Cephalocarida задние антенны расположены позади рта), жвалами, или мандибулами, и двумя парами челюстей, или максилл. Обе пары антенн служат в первую очередь органами чувств. Они богаты различными чувствительными щетинками, осуществляющими функции обоняния, осязания и чувства химического состава воды. Жвалы размельчают пищу, челюсти фильтруют пищевые частицы и участвуют в процессе подачи пищи ко рту.
На голове помещаются также глаза. Обычно у ракообразных есть пара сложных фасеточных глаз, устроенных так же, как у других членистоногих. Иногда эти глаза сидят на стебельках, иногда непосредственно на поверхности головы. У некоторых ракообразных, например у ветвистоусых, сложные глаза сливаются в один непарный глаз. У личинки ракообразных — науплиуса — сложные глаза еще не развиты, но имеется один непарный простой, так называемый науплиальный глаз. У многих ракообразных этот глаз сохраняется и у взрослых, а у веслоногих он служит единственным светочувствительным органом в течение всей их жизни. У глубоководных и подземных видов глаза исчезают.
Задний край головы многих ракообразных снабжен покрытой хитином складкой, которая охватывает сверху и с боков весь грудной отдел тела или его переднюю часть. Это — карапакс, имеющий форму щита, двустворчатой раковины или полуцилиндра. В тех многочисленных случаях, когда карапакс отсутствует (жаброногие, веслоногие, бокоплавы, равноногие и др.), он исчез вторично. Карапакс не только служит защитным образованием, но и определяет направление токов воды, необходимых для питания и дыхания.
Грудной и брюшной отделы тела у разных ракообразных состоят из различного количества сегментов. Нередко один или несколько передних грудных сегментов срастаются с головой и их конечности преобразуются в так называемые ногочелюсти, принимающие участие в захватывании пищи.
Брюшко заканчивается тельсоном, соответствующим пигидию многощетинковых червей.
У наиболее примитивных ракообразных грудные ножки выполняют три основные функции — двигательную, дыхательную и функцию подачи пищи ко рту. Каждая ножка снабжена несколькими лопастями, расположенными на ее внутреннем и наружном краях. Некоторые из наружных лопастей превращены в жабры, другие лопасти вызывают ток воды ко рту, третьи служат веслами. У более высокоорганизованных ракообразных наблюдается разделение функций между разными конечностями: одни используются только для передвижения, другие — только для дыхания, третьи — только для захватывания пищи.
Чрезвычайно характерным для ракообразных следует считать способ их дыхания. Как все настоящие водные животные, они дышат жабрами, через тонкие стенки которых растворенный в воде кислород проникает в кровь. Даже сухопутные ракообразные — мокрицы имеют жабры и используют кислород смачивающего их тонкого слоя влаги. Обыкновенно жабрами служат выросты грудных ног, но у равноногих и ротоногих ракообразных в жабры превращены брюшные конечности. У части ракообразных (веслоногие, мистакокариды, усоногие, ракушковые) жабр нет и дыхание совершается всей поверхностью их тела.
Кровь приводится в движение биением сердца, которое помещается недалеко от жабр. У ракообразных, жабры которых находятся на брюшном отделе, сердце помещается сзади, у остальных оно расположено в грудном отделе. Кровеносные сосуды есть только у представителей подкласса высших ракообразных, у остальных кровь течет по полостям между внутренними органами (рис. 189).
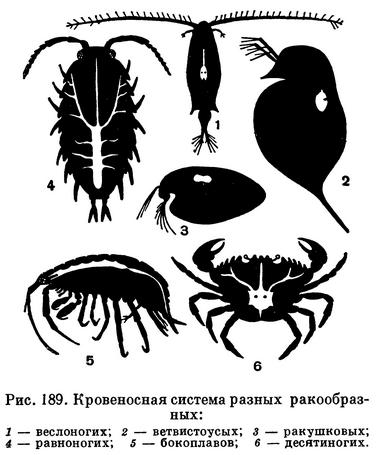
В крови часто растворены дыхательные пигменты — красный гемоглобин и синий гемоцианин, в котором железо заменено медью. Дыхательные пигменты могут связывать большое количество кислорода.
Питаются ракообразные различной пищей. Одни фильтруют имеющуюся в воде взвесь, включающую и мелкие организмы — бактерии, одноклеточные водоросли и т. д., другие своими жвалами откусывают куски мертвых и живых животных или растений. Пища перетирается жвалами и попадает в рот, а оттуда в пищевод. Задняя часть пищевода у большинства ракообразных преобразована в желудок, в котором иногда имеются хитиновые зубы, завершающие измельчение пищи. Экскременты выбрасываются через анальное отверстие, расположенное на заднем конце тела.
Органами выделения служат железы, открывающиеся наружу или при основании задних максилл (максиллярные железы), или при основании задних антенн (антеннальные железы). Эти железы соответствуют метанефридиям кольчатых червей. Выделяемая ими моча у многих пресноводных ракообразных очень бедна солями. Животные удаляют из полости тела воду, и концентрация солей в их полостной жидкости оказывается выше, чем в окружающей среде.
Почти все ракообразные раздельнополы, только усоногие и некоторые десятиногие — гермафродиты. Обыкновенно самцы резко отличаются от самок своим внешним видом или размерами. У одних ракообразных самцы крупнее, у других мельче самок. Часто у самцов есть какие-нибудь приспособления, помогающие им удерживать самку во время спаривания. Спаривание заключается в том, что самец прикрепляет близ половых отверстий самки сперматофор, т.е. кучку сперматозоидов, одетую оболочкой. Когда самка начинает откладывать яйца, вместе с ними выделяется вещество, растворяющее оболочку сперматофора; сперматозоиды оказываются на свободе и оплодотворяют яйца. В других случаях сперма вводится при помощи специальных копулятивных органов непосредственно в половые пути самки. Яйца представителей подкласса жаброногих ракообразных способны развиваться партеногенетически, т. е. без оплодотворения. Имеются виды, состоящие из одних самок, дающих начало новому поколению самок.
Сравнительно редко яйца откладываются прямо в воду. Обычно самка носит их на себе в специальной выводковой камере или прикрепляет их к поверхности тела или к конечностям.
Из яйца развивается характерная для всех ракообразных личинка науплиус. Тело науплиуса внешне не сегментировано. Он имеет три пары конечностей — передние и задние антенны и мандибулы. У некоторых ракообразных науплиус выходит из яйца и приступает к самостоятельному существованию, у других эта стадия формируется внутри яйцевой оболочки и личинка покидает яйцо, достигнув более поздней стадии развития. Наконец, есть ракообразные с прямым развитием, у которых все личиночные стадии проходят в яйце, а из него рождается уже сформировавшийся маленький рачок.
Личинки ракообразных часто непохожи на взрослых и нередко ведут совсем иной образ жизни. Так, например, личинки многих донных ракообразных плавают в толще воды.
Сравнение между собой по многим признакам различных ракообразных позволяет разделить этот огромный класс на 5 подклассов, к рассмотрению которых мы и переходим.
ПОДКЛАСС ЖАБРОНОГИЕ РАКООБРАЗНЫЕ (BRANCHIOPODA)
ПОДКЛАСС ЦЕФАЛОКАРИДЫ (CEPHALOCARIDA)
ПОДКЛАСС МАКСИЛЛОПОДЫ (MAXILLOPODA)
ПОДКЛАСС РАКУШКОВЫЕ РАКООБРАЗНЫЕ (OSTRACODA)
ПОДКЛАСС ВЫСШИЕ РАКООБРАЗНЫЕ (MALACOSTRACA)
Жизнь животных. Том 3. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1969 г.
ПОДТИП ХЕЛИЦЕРОВЫЕ (CHELICERATA)
Хелицеровые — обширный и разнообразный подтип членистоногих. В современной фауне они представлены почти исключительно наземными формами, обычно объединяемыми в класс паукообразных (Arachnida). Однако по происхождению хелицеровые — водные членистоногие, первоначально населявшие море. Водные формы, объединяемые в класс меростомовых (Merostomata), вымерли в отдаленные геологические эпохи. Из них до нашего времени дожило только несколько видов мечехвостов (отряд Xiphosura) — удивительным образом сохранившиеся «живые ископаемые». По происхождению хелицеровые связаны с трилобитами.
Наиболее характерные признаки хелицеровых — исчезновение усиков (антеннул) и своеобразный способ объединения сегментов тела в отделы, что связано с функциональными особенностями конечностей (рис. 4). Усики у всех хелицеровых отсутствуют, и их место занимает первая пара послеротовых конечностей, которые обычно имеют на конце клешни и получили особое название — хелицеры. Отсюда и название подтипа, что буквально значит «клешнеусые». Сегментальный состав отделов тела здесь иной, чем в других подтипах членистоногих, почему и отделы тела имеют особые названия. Тело делится на головогрудь — просому — и брюшко — опистосому. Конечности головогруди служат для захвата и пережевывания пищи и для передвижения, а конечности брюшка выполняют дыхательную, половую и другие специальные функции. В состав головогруди входят элементы головной лопасти — акрона — и последующих 6 сегментов. Акрону принадлежат глаза, расположенные на щите, покрывающем головогрудь, и срединный надротовой вырост — эпистом с верхней губой. Первый сегмент несет клешневидные хелицеры, которые, как говорилось, заняли место усиков перед ртом. Однако функционально они не заменяют усики, а служат для захвата пищи. За ними следует вторая пара ротовых конечностей, также имеющих особое название. Это педипальпы, или ногощупальца. Они состоят из основного членика — коксы, снабженного жевательным отростком — коксэндитом, и членистого щупальца. Среди конечностей головогруди функции педипальп наиболее разнообразны. Их основные членики с эндитами почти всегда так или иначе участвуют в приеме пищи, а щупальца в основном служат органами осязания, но могут участвовать в передвижении или захвате добычи. Конечности остальных четырех сегментов головогруди — ноги — служат для передвижения. Но у ряда форм на тазиках первой — третьей пар ног сохраняются жевательные отростки, и они также участвуют в пережевывании пищи.
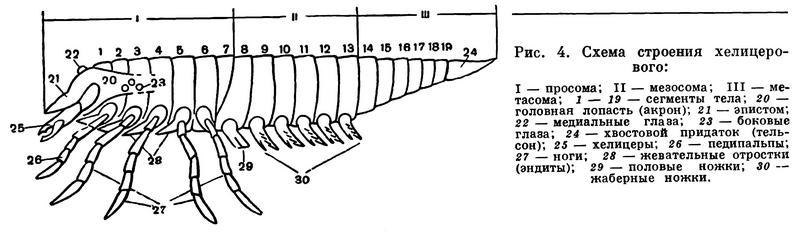
Головогрудь и брюшко у всех хелицеровых соединяются посредством седьмого, так называемого предполового, сегмента, который обычно лишен конечностей и в той или иной степени недоразвит. В составе брюшка объединяются две группы сегментов. Шесть передних, т. е. с восьмого по тринадцатый, имеют в той или иной степени недоразвитые видоизмененные конечности и образуют передний отдел брюшка — мезосому. Следующие за ними шесть сегментов всегда лишены конечностей и образуют задний отдел брюшка — метасому, которая оканчивается хвостовым придатком (тельсон). В пределах подтипа наблюдается постепенная атрофия сегментов метасомы и укорачивание брюшка. Первый брюшной сегмент, по общему счету восьмой, — половой, на нем всегда расположено половое отверстие, а его конечности у ряда форм образуют половые крышечки. Конечности следующих пяти сегментов у первичноводных форм представлены жаберными ножками, а у наземных превращаются в парные легкие, число которых различно и которые могут частично или полностью исчезать, заменяясь дыхательными трубками — трахеями.
Во внутреннем строении хелицеровых характерна сложная система выростов кишечника — так называемая печень, клетки которой способны к внутриклеточному перевариванию пищи (фагоцитозу). Сохраняются примитивные выделительные органы — коксальные железы, соответствующие у членистоногих выделительным органам кольчатых червей — целомодуктам. Характерна также атрофия отдела мозга, иннервирующего усики, — так называемого дейтоцеребрума, что связано с их исчезновением.
В подавляющем большинстве хелицеровые откладывают яйца, но есть и живородящие формы. Яйца богаты желтком, благодаря чему вылупляющаяся особь по своей организации сходна со взрослой, и послезародышевое развитие, как правило, протекает без превращений — метаморфоза, сопровождаясь ростом и линьками.
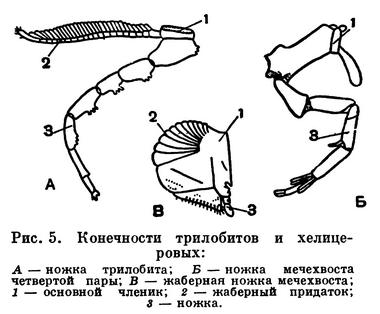
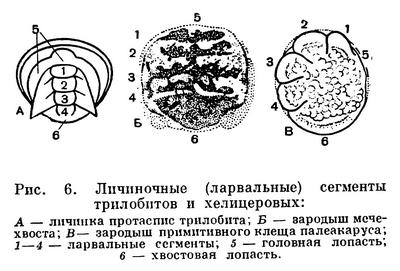
Хелицеровые очень древняя группа. Первые морские представители подтипа известны уже из кембрийского периода. На родство хелицеровых с трилобитами с определенностью указал в 1924 году Д. М. Федотов, затем Стермер, и эта точка зрения принята теперь большинством зоологов. Однако хелицеровых можно сближать лишь с наиболее примитивными трилобитами, у которых задние сегменты тела не слиты в пигидий. Родство с трилобитами проявляется во многих признаках. Существенно соответствие (гомология) частей двуветвистых конечностей трилобитов и жаберных ножек водных хелицеровых. У тех и других конечность состоит из основания и двух ветвей: внутренней — собственно членистой ножки наружной — членистого придатка с жаберными листочками (рис. 5). У водных хелицеровых, как у трилобитов, рот расположен вентрально, в виде продольной щели между коксальными эндитами ног. Четыре передних сегмента головогруди хелицеровых соответствуют сегментам головы трилобитов и представляют ларвальные сегменты (рис. 6). Как говорилось, они характерны для личинки трилобитов — протасписа — и, в отличие от последующих, образуются все одновременно. Одновременное появление этих сегментов и их анатомические отличия от остальных установлены впервые у зародыша мечехвостов в 1933 году П. П. Ивановым, а недавно обнаружены автором этого раздела у примитивных клещей. У некоторых хелицеровых (сольпуг и др.) и во взрослом состоянии четыре передних сегмента объединены в отдел, соответствующий голове трилобитов, а сегменты третьей и четвертой пар ног расчленены. Постларвальные (туловищные) сегменты у мечехвостов формируются, как у трилобитов, один за другим за счет деятельности околоанальной зоны роста, с тем только различием, что у трилобитов этот процесс происходил в послезародышевом развитии, а у мечехвостов наблюдается в эмбриогенезе.
Прототип хелицеровых наиболее полно олицетворяют древние морские меростомовые — эвриптериды (отряд Eurypterida), организация которых может быть выведена из трилобитообразной. При этом имели место две основные группы процессов: во-первых, атрофия органов чувств головы — в первую очередь усиков, а затем и сложных глаз, во-вторых, подразделение однородных трилобитных конечностей на двигательно-жевательные и дыхательные и соответственное объединение сегментов тела в отделы. От усиков, по крайней мере внешне, ни у кого из хелицеровых не осталось и следов. Сложные глаза атрофировались более постепенно, у большинства водных форм они имеются, но становятся меньше, чем у трилобитов, и в конце концов на их месте остается несколько простых глазков. Первая пара конечностей превратилась в хватательные хелицеры, следующие пять пар стали двигательно-жевательными, окружив укоротившуюся ротовую щель своими коксэндитами. В результате всех этих изменений голова как обособленный отдел исчезла, а все 6 передних сегментов и элементы головной лопасти объединились в передний отдел тела — просому. Остальные сегменты образовали брюшко — опистосому, причем конечности его переднего отдела — мезосомы — в большей своей части сохранили дыхательную функцию жабр, а конечности заднего отдела — метасомы — атрофировались. Общее число сегментов сократилось до 19; роль соединения головогруди и брюшка взял на себя седьмой сегмент, за которым всегда следует сегмент с половым отверстием.
Как мы видим, характернейшая черта организации хелицеровых — это отсутствие головы как обособленного отдела, и дело тут не только в том, что сегменты головы объединились с двумя туловищными в головогрудь. Ведь голова членистоногих, например насекомых, — это подвижный передний отдел тела, на котором расположены основные органы чувств — усики и сложные глаза — и набор ротовых конечностей, специально приспособленных для приема пищи. Ни один из этих компонентов головы у хелицеровых не выражен. Усики у них исчезли, зрение ослабло, а ротовые конечности в собственном смысле (челюсти) так и не образовались — большинство конечностей просомы одновременно жевательные и двигательные. Природу этого своеобразия организации в какой-то мере раскрывает образ жизни первичноводных форм, о чем мы можем судить по ныне живущим мечехвостам и по косвенным данным в отношении вымерших эвриптерид. Это крупные формы с щитовидной головогрудью, тяжелым панцирем и ограниченными возможностями движения, в большинстве донные животные, обитатели мелководных участков моря и опресненных водоемов. По способу добывания пищи большинство водных хелицеровых, по-видимому, было сходно с современными мечехвостами. Это роющиеся бентоядные хищники — потребители малоподвижных беспозвоночных: моллюсков, кольчатых червей и т. п. (эвриптериды и древние мечехвосты, вероятно, питались также трилобитами). Они не настигают добычу в погоне, а отыскивают «на ощупь» у себя под ногами, схватывают хелицерами и измельчают жевательными отростками ног. Ил роют передним острым краем головогрудного щита, а упором при рытье служит крепкая хвостовая игла, характерная для большинства меростомовых. Естественно, что при таком образе жизни необходимость совершенствования глаз как органов предметного зрения отпадает, а длинные нитевидные усики у переднего края тела становятся помехой при описанном способе рытья. Зато развитие клешневидных хелицер, служащих для схватывания добычи, и сохранение набора жевательных эндитов близ рта на брюшной стороне весьма существенны.
Значительные отличия от трилобитов возникли в индивидуальном развитии. У хелицеровых произошло обогащение яиц желтком. В результате те стадии, которые у трилобитов были свободными, послезародышевыми и развивались путем длительного первичного метаморфоза, у хелицеровых стали проходить в яйце в сокращенном виде, в порядке эмбриогенеза, и из яйца стала вылупляться особь, сразу похожая на взрослую, т. е. имело место явление, называемое эмбрионизацией развития (понятие, введенное в общей форме И. И. Шмальгаузеном и разработанное А. А. 3ахваткиным).
Историческая судьба хелицеровых сложилась иначе, чем судьбы двух других подтипов членистоногих — жабродышащих (ракообразных) и трахейнодышащих. Родина ракообразных — море, здесь они возникли, и вся их длительная эволюция, за немногими исключениями, связана с водной средой. Родина трахейнодышащих, в первую очередь насекомых, — суша, здесь они сформировались, достигли невиданного расцвета и господствуют по сей день. Родиной хелицеровых была водная среда — мелководье морей и, быть может, пресные воды. Здесь сложились основные черты организации этих животных. Однако в дальнейшем, будучи уже водными членистоногими с целым комплексом приспособлений к водной среде обитания и, по-видимому, даже к довольно узкой форме жизни бентоядных хищников, хелицеровые начали выходить на сушу, что, судя по имеющимся данным, происходило не раз и в различные эпохи. В море они не удержались и в большинстве вымерли. Живым свидетельством их прошлой водной жизни осталось несколько видов мечехвостов.
Переход к жизни на суше требует многих специальных приспособлений. По типу индивидуального развития хелицеровые были подготовлены к этому. Путем эмбрионизации они избавились от архаического водного метаморфоза — наследия трилобитов, при наличии которого выход на сушу вряд ли возможен. Однако по своей организации хелицеровые были типично водными членистоногими, для которых переход к сухопутной жизни был нелегкой задачей. И хотя она была разрешена наземными хелицеровыми — паукообразными, печать водного происхождения лежит на всей их эволюционной судьбе. В дальнейшем мы увидим, что по тем формам, которые приняла жизнь хелицеровых на суше, у них наблюдается много общего. В то же время каждый отряд паукообразных имеет свои особенности приспособления к сухопутной жизни и дальнейшей эволюции на суше, что в значительной мере обусловливает разнообразие наземных форм и помогает осветить их природу. Но если общие направления эволюции хелицеровых вырисовываются достаточно отчетливо, то этого нельзя сказать о филогенетических соотношениях группировок внутри подтипа. Накопившиеся результаты сравнительного изучения современных форм и палеонтологические материалы все менее укладываются в традиционную классификацию хелицеровых, и в последнее время не раз предпринимались попытки ее пересмотра. Однако зоологи не пришли к единому мнению, и это вынуждает нас в настоящем издании придерживаться традиционного подразделения хелицеровых на два класса — меростомовых (Merostomata) и паукообразных (Arachnida).
Вообще надо заметить, что отрасль знания о хелицеровых — арахнология далеко не так разработана, как, например, наука о насекомых — энтомология. Отечественных руководств по общей арахнологии не существует; вопросы происхождения и эволюции хелицеровых затронуты в немногих специальных работах. Иностранные руководства в большинстве также специальны, а в трактовке некоторых общих вопросов не всегда совпадают с нашими представлениями.
Желание придать по возможности эволюционное освещение материалу привело к необходимости научной постановки ряда проблем, при изложении, однако, в достаточно доступной форме, требуемой настоящим изданием. Такими проблемами явились происхождение хелицеровых, выход хелицеровых на сушу и различные пути эволюции паукообразных, сравнительное рассмотрение клещей как трех независимых отрядов и ряд других. При этом особенно существенной для нас была трактовка хелицеровых в трудах наших зоологов В. Н. Беклемишева и А. А. Захваткина.
ПОДТИП TPАХЕЙНОДЫШАЩИЕ (TRACHEATA)
Этот подтип включает только наземных членистоногих, таких, как многоножки и насекомые. Правда, среди последних есть и обитатели водоемов, например личинки стрекоз, водные жуки и др., но в этом случае ясно выявляется их вторичная водность, их происхождение от наземных предков, примерно так же, как легко устанавливается происхождение китов и ластоногих от наземных млекопитающих.
Хотя все трахейнодышащие — наземные животные, степень их приспособленности к жизни на суше разная, представители разных групп неодинаково легко выносят недостаток влаги в окружающем воздухе.
Многие низшие трахейнодышащие вообще не могут жить, если воздух постоянно не насыщен водяными парами. Низшие многоножки, низшие насекомые (протуры, вилохвостки) быстро погибают, если их насильно держать на открытом воздухе. Они могут жить только в таких укрытиях, где воздух очень влажен. Легче всего многоножки, низшие насекомые и другие наземные членистоногие находят необходимую для себя влагу в почве или в сходных средах (в гнилой древесине, в трещинах скал и т. п.). Поэтому все низшие трахейнодышащие, как правило, обитатели почвы, да и ранние стадии развития многих групп высших насекомых проходят в почве.
Поэтому же к жизни в почве, особенно в ее верхних слоях, легче всего приспосабливаются представители тех водных групп животных, у которых нет специализированных органов дыхания. У типичных обитателей почвы, таких, например, как нематоды, дождевые черви и др., дыхание осуществляется всей поверхностью кожи; во влажной почве животное может иметь обеспечивающие кожное дыхание проницаемые покровы, через которые легко поступает кислород и легко выходит углекислота. То, что через проницаемые для газов покровы легко может испаряться вода, при жизни в почве неважно — в почве воздух насыщен водяным паром и потому потеря влаги животным ничтожна. По-видимому, предки трахейнодышащих и были такими обитавшими в почве примитивными червеобразными мелкими членистоногими, которые дышали всей поверхностью тела. И среди современных низших многоножек, ногохвосток, протур и личинок насекомых есть немало таких, у которых дыхание кожное, трахей нет. Конечно, применительно к таким бестрахейным представителям членистоногих название «трахейнодышащие» довольно условно.
Переход от жизни в почве и в других укрытиях к жизни на открытой поверхности суши возможен для таких мелких животных только в том случае, если у них покровы становятся непроницаемыми для воды.
Переход от жизни в почве к жизни на суше требовал качественного изменения кутикулы у трахейнодышащих. При жизни в почве и дыхании всей поверхностью тела, как это бывает у самых примитивных наземных животных и, вероятно, было свойственно и предкам членистоногих, покровы должны были быть механически прочными и проницаемыми для газов. Именно такие покровы бывают у бестрахейных низших многоножек, у коллембол и др. В кутикуле таких членистоногих различается внутренний более эластичный богатый хитином слой — эндокутикула и наружный более плотный — экзокутикула. Если даже такая кутикула бывает толстой, она полностью не предохраняет от потери воды. Чем мельче животное, тем больше поверхность тела, которая приходится на единицу его объема и массы.
Трахейнодышащие — довольно мелкие организмы, обычно не крупнее нескольких сантиметров; поэтому относительно объема поверхность испарения у них довольно велика. Снижение проницаемости покровов шло у трахейных членистоногих двумя путями. Первый путь — уплотнение и утолщение покровов. Это тот путь, по которому шло приспособление к наземному образу жизни двупарноногих многоножек, которые, кстати, так полностью и не расстались с влажными местами и почвой. Другое, более совершенное приспособление выработалось у губоногих многоножек и у высших насекомых. У них предохранительным приспособлением к защите от потери влаги стал самый наружный очень тонкий, часто тоньше 0,001 мм, слой кутикулы, содержащий водонепроницаемые воскоподобные и жироподобные вещества — эпикутикула. Но эпикутикула препятствует проникновению кислорода. Как приспособление к воздушному дыханию в более или менее сухом воздухе у трахейнодышащих и вырабатывается трахейная система. У обитающих на суше ракообразных и паукообразных бывают и другие системы органов дыхания, а у трахейнодышащих только трахеи. Это самая экономная в отношении расхода влаги дыхательная система. Интересно, что у тех полихет, которые переходят к жизни на суше (хотя и во влажных местах), вырабатывается и уплотненная кутикула, и зачаточная трахейная система в виде углублений кожи, и даже брюшные ветви их параподий становятся похожими на расчлененные ноги многоножек. Особенно хорошо эти признаки выражены у обитающих на Зондских островах наземных полихет (Lycastis vivax и Lycastopsis amboinensis), которые могут служить «моделью» того, как от червеобразных предков произошли формы, напоминающие многоножек (рис. 85).
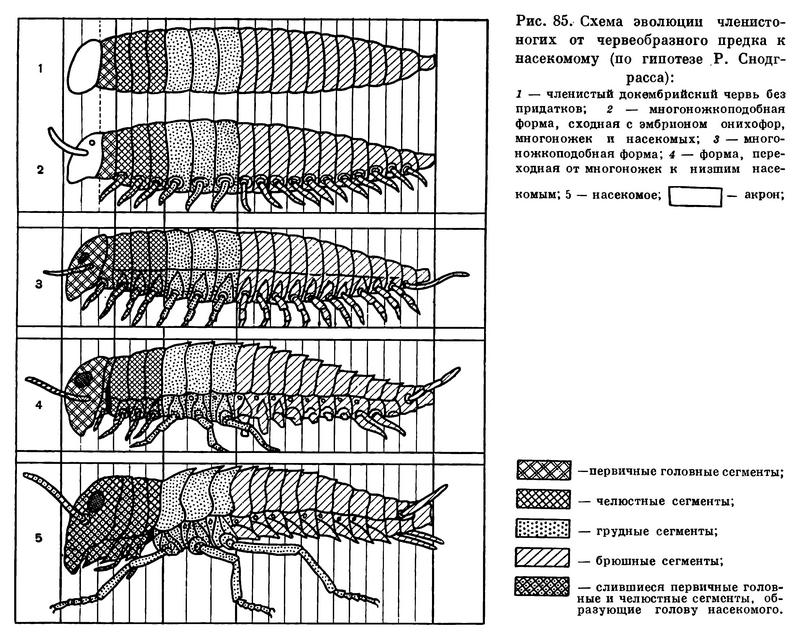
Приспособления к жизни на суше охватывают не только покровы и дыхательную систему у трахейнодышащих, но и другие системы органов, связанные с расходованием влаги. Выделение продуктов белкового обмена происходит у них не непосредственно во внешнюю среду, что всегда сопровождается потерей воды (мочевой жидкости), а в задний отдел кишечника, где вода всасывается его стенками. Выделительными органами у трахейных становятся или сами стенки кишечника (у низших многоножек, двухвосток, ногохвосток), или трубчатые выросты кишечника, лежащие в полости тела, — мальпигиевы трубочки. Даже биохимические реакции в организме трахейнодышащих перестраиваются в направлении экономии расхода воды — у них в результате распада белков получается мочевая кислота, которая легко выпадает в виде кристаллов и не требует для выведения столько воды, сколько нужно для выделения мочевины или аммиака, образующихся у водных членистоногих. И именно у трахейнодышащих вырабатывается особый орган для получения необходимой воды в те периоды, когда ее не хватает. У них полость тела заполнена особой жировой тканью — «жировым телом». Когда жир окисляется, выделяется много воды, которую и потребляют трахейнодышащие. Таким образом, у трахейнодышащих все системы органов приспособлены к экономии воды, что и позволяет многим из них оставить жизнь в почве и выйти на открытую поверхность. Низшие трахейнодышащие еще тесно связаны с жизнью в земле (многоножки, протуры, ногохвостки и др.), а высшие насекомые перешли к открытой жизни в воздушной среде, что полностью характерно только для взрослых; ранние же стадии (яйца, личинки или молодь) часто обитают в почве и других укрытиях.
Переход трахейнодышащих к жизни на суше потребовал и обязательного внутреннего оплодотворения.
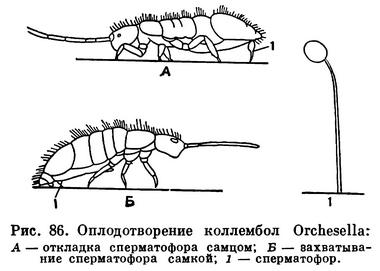
Гермафродитизм наблюдается редко (муха Termitoxenia, червец Icerya purchasi). Подавляющее большинство трахейнодышащих раздельнополы; многоножки, коллемболы, двухвостки, щетинохвостки и другие примитивные трахейнодышащие имеют особый свойственный почвенным членистоногим тип оплодотворения — наружно-внутреннее оплодотворение. При таком оплодотворении самец откладывает каплю семенной жидкости, обычно заключенную в тонкий футляр, — сперматофор — во внешнюю среду, а самка его подбирает половым отверстием, иногда через много часов после откладки (рис. 86). Этот способ оплодотворения возможен только во влажной среде. А на открытом воздухе такой способ оплодотворения исключен — во внешней среде капля семенной жидкости быстро высыхает и сперматозоиды погибают. Поэтому у «настоящих» наземных членистоногих — у насекомых вырабатывается внутреннее оплодотворение, достигаемое в разных группах очень разными путями.
Трахейнодышащие представляют единую группу членистоногих, в пределах которой намечается несколько путей исторического развития, совершавшегося уже несколько сотен миллионов лет, бурно продолжающегося и в наши дни. Часть сохранила облик, близкий к каким-то кольчатым червям. Есть основания думать, что трахейнодышащие произошли от далеких предков членистоногих, еще близких к кольчатым червям, перешедших к жизни в почве. Таковы различные многоножки. А у насекомых сократилось число сегментов и наметилось разделение туловища на 2 отдела — грудь, несущую двигательные придатки, и брюшко. Голова у всех трахейнодышащих довольно однотипна. Общим является наличие одной пары усиков в отличие от ракообразных. А вот ротовой аппарат выявляет довольно много общего у трахейнодышащих и ракообразных. Но нет оснований считать трахейнодышащих потомками ракообразных, как не оправдался и взгляд на них как на потомков трилобитов.
Вероятно, эти три группы происходят от разошедшихся задолго до того времени, от которого сохранились ископаемые остатки, трех ветвей тех групп кольчатых червей, которые дали начало членистоногим.
Среди трахейнодышащих явно более простыми являются многоножки.
ПОДТИП ТРИЛОБИТЫ (TRILOBITA)
Трилобиты — вымершие морские членистоногие, особенно обильные в первой половине палеозойской эры и полностью исчезнувшие к ее концу. Панцири трилобитов находят в ископаемом состоянии в морских отложениях, их описано около 4000 видов. Трилобиты — одни из древнейших наиболее примитивных членистоногих, отличавшиеся однородностью (гомономностью) конечностей и первичным метаморфозом. Их изучение позволяет полнее понять организацию и развитие вышестоящих групп, в особенности родственных им хелицеровых.
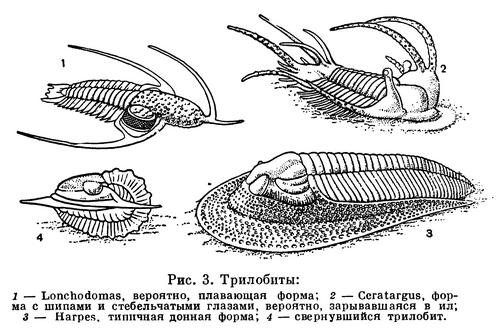
Длина тела трилобитов — 2—10 см, редко до 75 см. У примитивных форм тело состоит из двух отделов: слитной головы и расчлененного многосегментного туловища (рис. 1). Тело заканчивается небольшой анальной лопастью (тельсон). У большинства трилобитов анальная лопасть и несколько задних сегментов сливаются, образуя третий, хвостовой отдел — пигидий. Тело делится продольными спинными бороздами на выпуклую срединную часть (ось, или рахис) и уплощенные боковые (плевры). С тройным продольным и поперечным делением тела связано название «трилобиты», что значит трехдольчатые. Покров спинной стороны очень плотный, а брюшной — тонкий перепончатый. Голова и пигидий были покрыты щитами, сегменты тела между ними соединялись подвижно, что позволяло трилобиту свертываться на брюшную сторону.
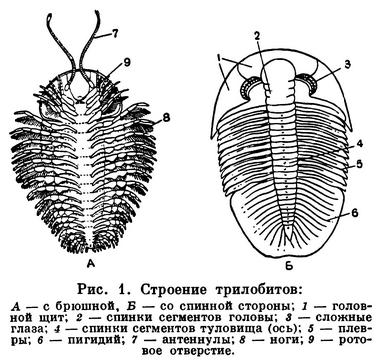
В состав головы входит головная лопасть — акрон и 4 послеротовых сегмента. Акрон образует переднюю и боковые части полулунного головного щита, ему принадлежат на спинной стороне пара сложных фасеточных глаз и пара простых глаз, а на брюшной стороне — пара нитевидных членистых усиков — антеннул, прикрепленных перед ртом. Позади рта расположены 4 пары конечностей сегментов головы, которые по строению подобны друг другу и следующим за ними многочисленным ножкам туловища. Все они двуветвистые, состоят из основания и двух членистых ветвей.
Долгое время считалось, что двуветвистые ножки трилобитов и ракообразных соответствуют друг другу по строению. Однако, как установил Стермер, это не так. У трилобитов от основного членика отходит неветвистая расчлененная ножка, а вторая ветвь представлена жаброй — членистым придатком основного членика с жаберными листочками. У раков двуветвиста сама ножка. В то же время конечностям трилобитов подобны брюшные жаберные ножки водных хелицеровых, в частности мечехвостов, что служит одним из доказательств родства этих членистоногих. Ножки трилобитов, судя по их строению, выполняли различные функции: служили органами дыхания, плавания и ползания по грунту, по-видимому, гнали воду по брюшной стороне тела вперед, к ротовому отверстию, доставляя органические частицы и мелкие донные организмы, вероятно, служившие пищей трилобитам. Пища дополнительно измельчалась с помощью жевательных отростков — эндитов, имевшихся на члениках ножек у ряда форм.
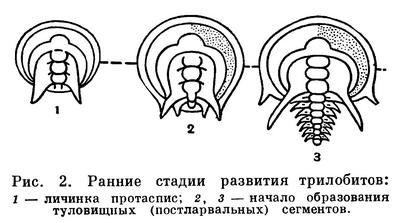
Судя по ископаемым остаткам неполовозрелых стадий, развитие трилобитов представляло примитивный метаморфоз, который по некоторым признакам напоминает развитие кольчатых червей. Из яйца вылуплялась очень мелкая (до 1 мм) личинка — протаспис (рис. 2). Самые ранние известные стадии личинки имеют цельное тело. Далее оно сразу делится на головную и анальную лопасти и 4 сегмента между ними, которые являются сегментами головы трилобита и, в отличие от остальных сегментов, образуются все одновременно и называются личиночными или ларвальными. Дальнейшее развитие сопровождалось появлением туловищных сегментов, длительным ростом и линьками. Новые, послеличиночные, или постларвальные, сегменты, как и у аннелид, один за другим обособлялись вперед от зоны роста, расположенной у переднего края анальной лопасти, т. е. путем анаморфоза, и в развитии следовал длинный ряд стадий с неполным числом сегментов, вплоть до полносегментного состояния. После этого трилобит продолжал расти и линять, но новых сегментов обычно уже не появлялось, т. е. начинался период развития, называемый эпиморфозом.
Среди современных членистоногих первичный метаморфоз сохранился у некоторых ракообразных, у которых из яйца вылупляется личинка — науплиус, имеющая только ларвальные сегменты, а далее следует процесс наращивания постларвальных сегментов. Однако число ларвальных сегментов у протасписа и науплиуса различно: у первого 4, у второго 2. Это, а также различия в строении конечностей препятствуют сближению ракообразных с трилобитами. В то же время в эмбриональном развитии водных хелицеровых — мечехвостов — обнаружено 4 ларвальных сегмента, что в сочетании с другими признаками служит основанием для сближения хелицеровых с трилобитами.
Трилобиты были исключительно морскими формами, на что указывает характер отложений, содержащих их панцири и сопутствующие остатки других морских беспозвоночных. В большинстве трилобиты обитали в мелководных участках моря и были донными животными, медленно ползающими и плавающими у дна. На это указывает плоское снизу тело, тяжелый панцирь, относительно слабые конечности, вентральное положение рта. Многие могли зарываться в ил, особенно формы с острым краем головного щита. Обитателями толщи ила считаются некоторые формы с торчащими шипами и стебельчатыми глазами, которые они, видимо, выставляли над поверхностью дна наподобие перископов. Некоторые формы с длинными шипами на голове считают плавающими. Шипы увеличивали поверхность тела, повышая плавучесть, и такие виды, вероятно, могли парить в толще воды. Утрата глаз некоторыми группами трилобитов в одних случаях может быть связана с жизнью в иле или в сильно взмученных течениями водах в зонах илонакопления, в других — с жизнью в темноте на больших глубинах. Так, некоторые мелкие слепые трилобиты считаются обитателями больших глубин, измельчавшими в связи с недостатком пищи. Расселение трилобитов, вероятно, происходило на ранних личиночных стадиях. Мелкие личинки — протасписы, почти равные по размерам науплиусам современных веслоногих раков, очевидно, были планктонными организмами и могли далеко переноситься течениями, способствуя широкому распространению, которое характерно для многих видов трилобитов.
Трилобиты были очень многочисленны в кембрийский и ордовикский периоды, в силуре, девоне и карбоне число видов постепенно, но неуклонно сокращается, и в перми они полностью исчезают. Отчетливое изменение морфологических признаков во времени, обилие видов и хорошая сохранность панцирей сделали трилобитов важнейшей группой «руководящих ископаемых» при изучении раннепалеозойских геологических систем. Наличие остатков трилобитов позволяет определять морские отложения, их возраст и условия образования, что важно при разведке полезных ископаемых.
При изучении трилобитов невольно возникает вопрос: что привело к полному вымиранию такой огромной группы организмов, некогда господствовавшей в морях? Обычно указывают на возможные изменения абиотических условий и на хищников. Врагами трилобитов считают ракоскорпионов, головоногих моллюсков и древних рыб, численность которых возрастала как раз с силура, а средствами защиты от хищников — наличие у многих трилобитов шипов и способность свертываться на брюшную сторону. Однако внешние факторы сами по себе вряд ли могут объяснить исчезновение трилобитов. Ведь рядом с ними жили другие группы беспозвоночных, в том числе не менее древние ракообразные, которые также подвергались действию абиотических перемен и уничтожались хищниками, но тем не менее благополучно дожили до нашего времени, а многие достигли расцвета. Видимо, дело не только в уничтожающих факторах, но и в самой природе трилобитов. Если сравнить их с ракообразными, то при всем обилии видов трилобитов бросается в глаза крайняя однотипность их примитивной организации и особенно развития. Как мы видели, формы индивидуального развития ракообразных эволюционировали с давних времен. Одни из них пошли по пути обогащения яиц желтком, благодаря чему из яйца стал вылупляться организм, сразу похожий на взрослого, у других возник сложный метаморфоз со сменой типов личинок и образа жизни, в частности у паразитов, третьи приобрели способность очень быстро размножаться, особенно мелкие планктонные формы, и т. д. Обогащение яиц желтком произошло и у водных хелицеровых, о чем речь пойдет ниже Ничего подобного у трилобитов мы не находим по-видимому, все они обладали первичным метаморфозом в его архаическом, так сказать, наиболее длинном варианте Чтобы стать взрослым и способным размножаться, трилобит был вынужден пройти длиннейший ряд стадий со множеством линек, начиная от крошечного протасписа, через периоды наращивания сегментов тела и последующего роста в полносегментном состоянии. При этом, кроме самых ранних стадий, вероятно планктонных, на всех последующих образ жизни существенно не менялся, т. е. жизненный цикл был монотонным и в экологическом отношении. Естественно, что при таком способе развития противостоять уничтожающим факторам среды весьма трудно. Быстрое восстановление численности вида невозможно, а сохранение хотя бы части особей в каких-то иных экологических условиях маловероятно благодаря однотипному образу жизни в течение большей части жизненного цикла. Иными словами, наиболее правдоподобно предположение, что трилобиты не смогли устоять против уничтожающих факторов изменявшейся среды, вероятно в основном против появившихся хищников, благодаря несовершенству метаморфоза, которое они так и не сумели преодолеть в своем историческом развитии.
Отметим еще, что в кембрийских отложениях открыты остатки странных членистоногих, по некоторым признакам похожих на трилобитов, а по другим на хелицеровых и на ракообразных. Эти недостаточно изученные формы рассматривают как представителей нескольких классов, которые при таком подразделении объединяются с трилобитами в подтип трилобитообразных (Trilobitomorpha). Возможно, что дальнейшее изучение этих животных позволит полнее осветить соотношения трилобитов и других членистоногих.