(Malacostraca Latreille, 1802) Класс Малакостраки, Высшие ракообразные, Class Malacostraca Latreille, 1802 (Malacostracans) 14 отрядов
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ПОДКЛАСС ВЫСШИЕ РАКООБРАЗНЫЕ (MALACOSTRACA)
Основные признаки видов подкласса — постоянное количество грудных и брюшных сегментов, а также присутствие брюшных конечностей. Грудь состоит из 8, а брюшко из 6 или 7 сегментов и заканчивается тельсоном. Нередко передний или несколько передних грудных сегментов срастаются с головой и их конечности превращаются в ногочелюсти. Иногда срастаются между собой или с тельсоном и брюшные сегменты. Мужские половые отверстия всегда открываются на последнем, а женские на шестом грудном сегменте. Желудок подразделен на жевательную и фильтрующую камеры. Всегда есть сильно развитая пищеварительная железа, сердце и кровеносные сосуды.
Название «высшие» не совсем точно. Если постоянство числа грудных и брюшных сегментов можно считать признаком более высокой организации, чем их непостоянство, присущее другим подклассам ракообразных, то присутствие брюшных конечностей, наоборот, говорит о меньшей специализации, чем их исчезновение, наблюдающееся у других подклассов. По всей вероятности, высшие ракообразные развивались независимо от других подклассов, каждый из которых сохранил те или иные примитивные признаки, унаследованные от общих предков.
Представители высших ракообразных, как правило, крупнее, чем рачки, относящиеся к другим подклассам. Они распространены чрезвычайно широко: населяют самые разнообразные водоемы и частично приспособились к обитанию на суше. Систематика подкласса очень сложна: он делится на надотряды, которые в свою очередь подразделяются на отряды. Таких надотрядов насчитывается пять.
НАДОТРЯД ФИЛЛОКАРИДЫ (РНYLLOCARIDА)
Этот надотряд включает единственный очень бедный видами отряд, сохраняющий признаки примитивной организации.
ОТРЯД ТОНКОПАНЦИРНЫЕ РАКООБРАЗНЫЕ (LEPTOSTRACA)
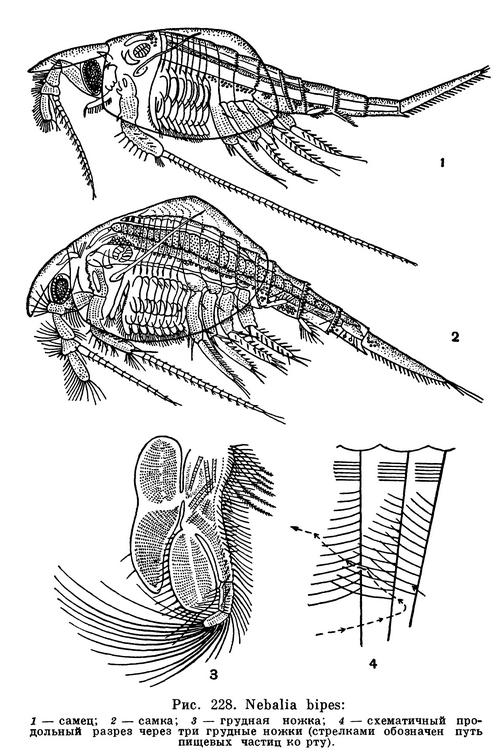
Тонкопанцирные ракообразные обитают исключительно в море. Большая их часть встречается в прибрежной области и ведет придонный образ жизни и только один вид — Nebaliopsis typica — планктонное глубоководное животное. Известно всего 8 видов этих своеобразных рачков, но распространены они очень широко.
Карапакс тонкопанцирных имеет вид тонкой двустворчатой раковинки, покрывающей весь грудной и переднюю часть брюшного отдела рачка. С ним подвижно сочленен направленный вперед овальный пластинчатый рострум, прикрывающий сверху большие стебельчатые глаза и основания антенн. Наклоняя или приподнимая рострум, рачок регулирует силу тока воды, возбуждаемого грудными ножками и направляемого спереди назад между створками раковины.
Передняя и задняя антенны довольно длинные и богаты чувствительными щетинками. Ротовые придатки обычного строения и служат для фильтрации и размельчения пищи. Кроме того, удлиненный щупик передней челюсти очищает полость раковины от попадающих туда посторонних частиц. Грудной отдел состоит из 8 сегментов, каждый из которых снабжен парой одинаково устроенных ножек, приспособленных одновременно и для передвижения, и для дыхания, и для подачи пищи ко рту. Эти ножки пластинчатые, листовидные. Их наружные лопасти — эпи- и экзоподит — служат жабрами, а снабженная длинными щетинками более узкая и сильнее хитинизированная внутренняя лопасть — эндоподит — функционирует преимущественно как орган движения. На внутреннем крае основной части каждой ноги сидят щетинки двух типов: одни более короткие и тонкие, но более многочисленные отфильтровывают пищевые частицы, которые передаются сзади наперед другими более длинными и малочисленными щетинками.
Грудные ножки сгибаются и разгибаются последовательно. Их движения напоминают движения ног жаброногов. При встречном изгибании ножек соседние пары образуют фильтрующие камеры. Вода проходит спереди назад по продольному желобку вдоль брюшной стороны тела, оставляя между фильтрующими щетинками мелкие взвешенные частицы, а затем двумя потоками расходится в стороны. Эпиподиты и частично экзоподиты ножек используют при этом растворенный в воде кислород, но, кроме того, он усваивается и тонкими покровами внутренней стенки раковины. Даже когда рачки неподвижно сидят на грунте, их ножки продолжают совершать свои ритмичные взмахи. Только при зарывании в грунт, когда рострум загибается вниз и закрывает спереди вход в раковину, работа грудных ножек прекращается.
В состав длинного цилиндрического брюшка входит 7 сегментов. Из них 4 передних несут длинные двуветвистые ножки, богатые щетинками и служащие для плавания. Ножки следующих 2 сегментов короткие, редуцированные, а последний брюшной сегмент совсем лишен конечностей. За ним следует удлиненный тельсон с двумя длинными ветвями фурки.
Самцы по внешнему виду и по своему поведению несколько отличаются от самок. Их раковинка уже, брюшной отдел с тельсоном относительно головы и груди длиннее, глаза крупнее, а число чувствительных щетинок на передних антеннах гораздо больше. У донных видов самцы быстрее и более продолжительное время, чем самки, плавают, сильно взмахивая своими передними брюшными ножками, и активно отыскивают менее подвижных самок.
Самки носят откладываемые яйца на себе, удерживая их щетинками внутренних краев задних грудных ножек. Из яиц выходят молодые, еще не вполне сформировавшиеся рачки, первое время остающиеся на ножках матери. После двух линек они переходят к самостоятельному существованию.
Тонкопанцирные ракообразные, сохраняющие многие примитивные признаки, только сравнительно недавно были найдены в ископаемом состоянии. Из пермских отложений Германии описали Nebalia bentzi, очень близкую к обыкновенной Nebalia bipes. Это подтверждает глубокую древность отряда.
В палеозое существовал близкий к тонкопанцирным отряд Archaeostraca, принадлежащий к тому же надотряду Phyllocarida. Среди Archaeostraca известны виды, которые считаются самыми примитивными из всех высших ракообразных, как в частности Nahecaris из девонских отложений Германии. Однако систематическое положение многих Archaeostraca не вполне ясно.
НАДОТРЯД СИНКАРИДЫ (SYNCARIDA)
Этот надотряд также характеризуется примитивностью организации и глубокой древностью. Замечательно, что сначала были открыты его ископаемые представители (в каменноугольных отложениях) и только через 40 лет были найдены ныне живущие.
Все современные Syncarida обитают в пресной воде. Они населяют высокогорные озера и ручьи Тасмании и подземные воды юго-восточной части Австралии, а также подземные воды других материков, за исключением Северной Америки.
В отличие от тонкопанцирных они полностью лишены карапакса. Все их грудные и брюшные сегменты имеют сходное строение и приблизительно одинаковые размеры. Грудные ножки двуветвисты. Конечности последнего брюшного сегмента преобразованы в рулевые ноги — так называемые уроподы. Левая и правая жвалы симметричны. Самки откладывают яйца в воду и не носят их на себе.
Надотряд включает 3 отряда — Anaspidacea, Bathynellacea и Stygocaridacea.
ОТРЯД АНАСПИДОВЫЕ РАКООБРАЗНЫЕ (ANASPIDACEA)
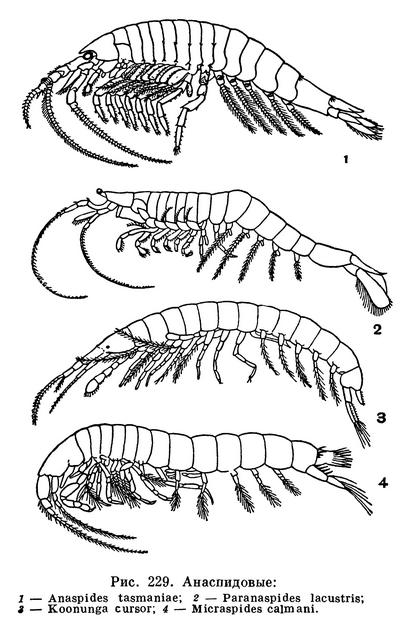
Ныне живущие представители анаспидовых принадлежат всего к 5 видам. Они обитают только в высокогорных озерах и ручьях Тасмании, и лишь один из них живет в грунтовых водах близ г. Мельбурна в Австралии. В палеозойское время анаспидовые были распространены гораздо шире. Их ископаемые остатки найдены во многих местах Европы и Северной Америки, причем древнейшие из этих ракообразных были морскими животными. Современные виды можно считать последними остатками древней, некогда процветавшей группы, дожившими до наших дней в очень ограниченной области, в водоемах, где нет или очень мало их конкурентов и врагов.
На голове этих рачков находятся стебельчатые (у Anaspides и Paranaspides) или сидячие (у Koonunga) глаза, совсем отсутствующие у подземного Micraspides. Антенны довольно длинные. В основном членике передней антенны расположен орган равновесия — статоцист, весьма своеобразно устроенный. В нем нет статолита, как это обычно бывает, но на его стенках сидят булавовидные щетинки, утолщенные концы которых сгибаются под действием земного притяжения.
Передний грудной сегмент срастается с головой, однако его конечности сохраняют характер ходильных. Только у Koonunga они приспособлены к рытью грунта, а у остальных родов имеют такое же строение, как последующие, если не считать присутствия внутренних выростов, участвующих в подаче пищи ко рту. Грудные ножки при основании снабжены тонкостенными наружными выростами — эпиподитами, выполняющими функцию жабр. На передних 6—7 парах грудных ножек есть и наружные ветви — экзоподиты. Они совершают беспрерывные взмахи (у Anaspides 100, а у Paranaspides 250—300 раз в минуту), благодаря чему жабры все время омываются свежей водой. Две передние пары брюшных ножек самцов преобразованы в копулятивный аппарат. Они имеют вид трубок и направлены вперед. Следующие три пары снабжены сильно развитой многочлениковой, богатой щетинками наружной и короткой, иногда совсем отсутствующей внутренней ветвью. При помощи наружных ветвей этих ножек Anaspides и Paranaspides могут некоторое время плавать, держась спинной стороной вверх. Последние брюшные ножки — уроподы — пластинчатые, направлены назад и вместе с тельсоном составляют хвостовой веер. Резко изгибая тело и отталкиваясь хвостовым веером от воды, рачки способны передвигаться скачками задним концом вперед.
Самка Anaspides откладывает крупные (диаметром около 1 мм) яйца на поверхность водных растений или на другие подводные предметы. Из яиц выходят молодые рачки длиной 2,7 мм, отличающиеся от взрослых недоразвитыми стебельчатыми глазами, вместо которых имеется непарный глаз, отсутствием внутренних ветвей передних брюшных конечностей и расщепленным на конце тельсоном, задне-боковые отростки которого напоминают ветви фурки других ракообразных. После нескольких линек молодые рачки становятся вполне подобными взрослым.
Anaspides tasmaniae — самый крупный представитель отряда, достигающий в длину 5 см, живет преимущественно в ручьях и мелких речках нагорного плато Тасмании. Значительно более мелкие Anaspides spinulae и Paranaspides lacustris обитают там же, но в высокогорных озерах. Все они питаются главным образом донными водорослями и бактериями и органическими остатками грунта, который они взмучивают грудными ножками и отфильтровывают задними челюстями. Anaspides может захватывать и более крупную пищу, например червей и других водных животных.
Сравнение физико-химических условий водоемов, населенных этими тремя видами, показало, что они выносят довольно значительные колебания температуры, различное содержание в воде кислорода и различный химический состав ее. Очевидно, ограниченность области их распространения объясняется не какими-нибудь особыми, присущими ей физико-химическими условиями, а геологической историей нагорной части Тасмании, позволившей сохраниться только там «живым ископаемым».
Четвертый тасманийский вид — Micraspides calmani длиной около 8 мм — обитает в полостях торфяника. Он лишен глаз, что указывает на его подземный образ жизни. То же можно сказать о зарывающемся в грунт единственном австралийском представителе анаспидовых — Koonunga cursor.
ОТРЯД БАТИНЕЛЛИЕВЫЕ РАКООБРАЗНЫЕ (BATHYNELLACEA)
Не только капиллярные, пропитанные соленой водой ходы морских пляжей населены специально приспособленными к жизни в таких условиях ракообразными — гарпактицидами и мистакокаридами. На значительном удалении от моря на обширных пространствах суши, особенно там, где имеются отложения известняка, под сравнительно тонким слоем почвы циркулируют подземные воды. Они пропитывают горные породы, протекают через узкие пространства между частицами грунта, заполняют колодцы и образуют водоемы в глубине известковых пещер. В этих водах также обитает своеобразное население, в том числе некоторые ракообразные. Наиболее характерными из них следует считать представителей отряда Bathynellacea.
Микроскопические размеры (обычно до 2 мм, Baicalobathynella magna до 5,4 мм), тонкое червеобразное тело и короткие конечности позволяют этим рачкам пробираться по узким подземным ходам и проползать между частицами грунта. Вместе с грунтовыми водами они проникают в колодцы, пещерные водоемы, а иногда и в придонные слои воды крупных озер и живут и в таких условиях, в том случае если температура воды достаточно низка. Первый представитель Bathynellacea — Bathynella natans — был обнаружен чешским зоологом В. Вейдовским в колодце в Праге в 1886 году, а затем в течение 28 лет никому не удавалось находить этих рачков. Впоследствии, после того как методы исследования подземных вод были усовершенствованы и на их фауну стали обращать серьезное внимание, Bathynellacea были открыты во многих странах и оказались очень широко распространенными. Теперь они известны из ряда мест Европы, экваториальной Африки, Мадагаскара, полуострова Малакка, Японии и Южной Америки. В СССР их нашли в пещерах Кавказа, в колодцах близ Дона, в озере Байкал, в пещерах Средней Азии и Дальнего Востока. Всего известно 64 вида и подвида Bathynellacea.
Тело Bathynellacea удлиненное, цилиндрическое. В связи с подземным образом жизни глаза отсутствуют. Антенны довольно короткие, с многочисленными чувствительными щетинками. Все 8 грудных сегментов свободные. Их конечности двуветвистые и снабжены эпиподитами, участвующими в процессе дыхания. Однако газообмен происходит, по-видимому, и через стенки тела. У самцов конечности последнего грудного сегмента сильно видоизменены и превращены в совокупительный аппарат, а у самок они могут отсутствовать. Брюшные сегменты по длине и ширине не отличаются от грудных. Последний брюшной сегмент срастается с тельсоном, на конце которого расположены фуркальные ветви. Короткие редуцированные ножки есть только на одном или двух передних брюшных сегментах, или их нет совсем, но на последнем сросшемся с тельсоном брюшном сегменте всегда имеются сильно развитые двуветвистые уроподы.
При помощи грудных ножек Bathynellacea медленно ползают по узким ходам между частицами грунта, изгибая при этом свое тело. Попав в более обширное пространство, они могут некоторое время плавать, последовательно взмахивая своими грудными ножками и оставляя тело выпрямленным. Однако их движения в толще воды настолько слабы, что рачки совсем не могут противостоять течению любой силы, которое легко их сносит. Обыкновенно они находятся на грунте или в самом придонном слое воды, и их удается поймать планктонной сеткой только в том случае, если она зачерпнет поверхностный слой грунта.
Размножение изучено только у Bathynella natans. Самка откладывает на грунт поодиночке довольно крупные (диаметром 0,11 мм) по отношению к ее телу яйца. При температуре 11°С из них через 2 месяца выходят малоподвижные личинки длиной около 0,45 мм. У личинок уже имеются антенны, все ротовые придатки и передняя пара грудных ножек, а также 10 туловищных сегментов. Они еще не питаются, используя находящиеся в их теле запасы желтка. После первой линьки добавляется вторая пара грудных ножек, еще один туловищный сегмент, а также возникают зачатки недостающих конечностей. Личинки становятся более активными и питаются самостоятельно. С каждой последующей линькой (а их до окончания превращения личинки насчитывается пять) личинки постепенно приближаются по своему строению к взрослым. Через 9 месяцев после рождения заканчивается формирование взрослого животного, но для достижения половозрелости самцы должны перелинять еще 4 раза, а самки 5 раз. В общем жизнь рачков при температуре 12—14°С длится не менее 2 лет.
Большинство Bathynellacea обнаружено в различных выходах грунтовых вод при температуре не выше 12—14°С. Несколько видов обитает в воде, пропитывающей песок близ берегов озер или лагун, как например Parabathynella caperti, найденная в песке на расстоянии 1 м от уреза воды озера Танганьика в Африке, или виды того же рода с острова Мадагаскар. Bathynella baicalensis и Baicalobathynella magna живут на значительной глубине (до 1440 м) в озере Байкал. К совершенно необычным для Bathynellacea условиям приспособились Thermobathynella adami и Thermobathynella amyxi. Первый вид населяет теплые (55°С) подземные воды близ озера Упемба в экваториальной Африке, а второй — пропитывающие песок солоноватые воды в дельте Амазонки.
Широкое и разбросанное географическое распространение Bathynellacea при полном отсутствии у них покоящихся стадий и, следовательно, способности к пассивному расселению до сих пор остается нерешенной загадкой. Некоторые исследователи допускают давнее морское происхождение этих рачков, предполагая, что они сохранились в участках суши, некогда покрывавшихся морем, однако прямых доказательств правильности этой гипотезы пока не приведено. Во всяком случае, несомненна глубокая древность этих мельчайших высших ракообразных.
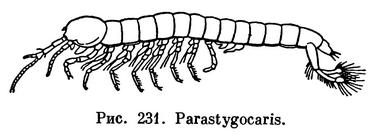
ОТРЯД СТИГОКАРИДЫ (STYGOCARIDACEA)
Представители отряда Stygocaridacea по своему внешнему виду, размерам и по образу жизни напоминают Bathynellacea. Они были открыты только в 1963 году при обследовании фауны грунтовых вод Южной Америки. Известно всего 2 рода и 4 вида Stygocaridacea.
По характеру сегментации тела этот отряд довольно резко отличается от Bathynellacea. Передний грудной сегмент Stygocaridacea полностью срастается с головой, но задний брюшной сегмент свободный, отчлененный от тельсона. Тельсон снабжен рудиментарной фуркой. В основании передних антенн имеется орган равновесия — статоцист. Передние брюшные ножки самца преобразованы в копулятивный аппарат. У обоих видов рода Stygocaris отсутствуют наружные ветви грудных конечностей.
Все Stygocaridacea живут в интерстициальных пресных водах Аргентины и Чили, причем Parastygocaris andina известен в Андах на высоте 2000 м над уровнем моря. Биология этих рачков не изучена. Сходство их с Bathynellacea объясняется одинаковой средой обитания (интерстициальные воды), к которой представители обоих отрядов приспособились независимо друг от друга.

НАДОТРЯД ГОПЛОКАРИДЫ (HOPLOCARIDA)
Гоплокариды — крупные морские ракообразные, пять передних грудных сегментов которых срастаются с головой. Глаза стебельчатые. Короткий карапакс не покрывает трех задних грудных сегментов. Брюшко очень длинное, передние его конечности несут жаберные придатки. Самки вынашивают яйца под грудью. К этому надотряду относится всего один отряд.
ОТРЯД РОТОНОГИЕ РАКООБРАЗНЫЕ, ИЛИ РАКИ-БОГОМОЛЫ (STOMATOPODA)
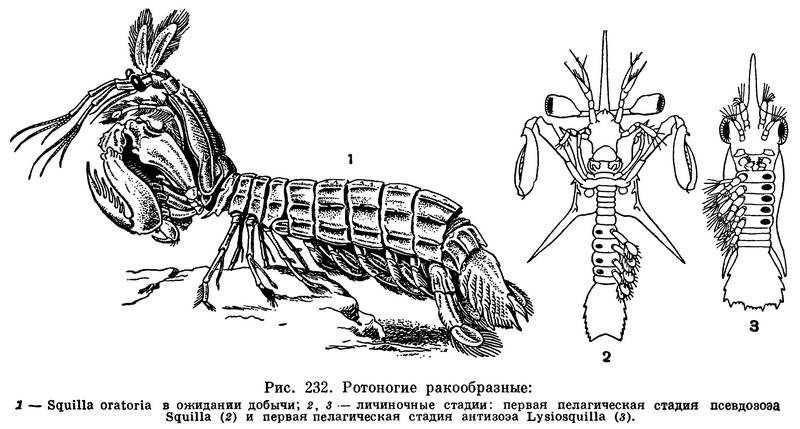
На мелководьях тропических и субтропических морей, а также среди коралловых рифов, в норах или в расселинах коралловых массивов укрываются крупные (от 1—2 до 34 см длиной) хищные раки. Наружу они выставляют только самый передний конец тела со стебельчатыми глазами, чувствительными антеннами и мощными ловчими ногами. Если мимо них проплывет креветка или мелкая рыба, они молниеносно выскакивают из норы и хватают жертву своими ловчими ногами, последний членик которых вооружен многочисленными острыми шипами и прижимается к предпоследнему, как лезвие перочинного ножа к его ручке. Пронзенная шипами жертва оказывается зажатой между последним и предпоследним члениками. По способу охоты и строению ловчих ног эти ракообразные несколько напоминают широко известных хищных насекомых — богомолов, почему они и получили название раков-богомолов. Нередко они сами оказываются добычей рыб или человека. В Средиземном и Адриатическом морях, а также в Индийском и Тихом океанах существует регулярный промысел ротоногих ракообразных, считающихся очень вкусными.
Строение их весьма своеобразно. Голова и передняя часть их грудного отдела сверху покрыты карапаксом, имеющим вид щита, лишь в малой степени защищающего бока животного. Передний край карапакса сочленен с подвижным рострумом, по бокам которого располагаются сидящие на длинных стебельках фасеточные глаза. Между ними находится науплиальный глаз. Фасеточные глаза раков-богомолов отличаются необыкновенной подвижностью. Они могут поворачиваться вслед за проплывающими мимо животными и в процессе ловли добычи играют очень важную роль. Каждая из передних антенн состоит из длинной 3-члениковой основной части и трех прикрепленных к ней жгутов. На жгутах расположены многочисленные органы химического чувства и вкуса — цилиндры и щетинки. Перед тем как схватить замеченную глазами добычу, рак пускает в ход передние антенны и только после того, как убедится в съедобности объекта, приступает к его ловле. В аквариуме рак-богомол пристально следил за привязанным к нитке шариком из пластилина, которым экспериментатор двигал перед ним, но после исследования шарика передними антеннами не делал никаких попыток его поймать. Совсем иначе рак вел себя, когда к нитке вместо пластилинового шарика привязывали кусок рыбы. Задние антенны короче передних. Их наружная ветвь преобразована в чешуйку, как у многих десятиногих ракообразных и у мизид.
Конечности пяти передних слившихся с головой и прикрытых сверху карапаксом грудных сегментов превращены в ногочелюсти. Первая из них короткая, тонкая, густо покрыта щетинками и служит органом осязания. При помощи этих конечностей рак также удаляет со своего тела посторонние частицы. Остальные четыре пары замечательны тем, что их последний и предпоследний членики образуют так называемую подклешню за счет способности последнего членика пригибаться к предпоследнему. Так возникает приспособление к захватыванию добычи или, в случае необходимости, комочков грунта. Из этих конечностей особенно сильно развита передняя (т. е. вторая ногочелюсть), превосходящая остальные по размерам не менее чем вдвое, с вооруженным острыми шипами последним члеником. Именно эта пара ног и осуществляет схватывание добычи. Остальные ногочелюсти передают пойманную добычу вперед, ко рту, и, кроме того, выполняют основную работу по рытью грунта при постройке нор. Все пять пар ногочелюстей лишены наружных ветвей, но несут маленькие эпиподиты, в незначительной степени участвующие в процессе дыхания.
Три задних грудных сегмента подвижно сочленены между собой и не покрыты карапаксом. Их конечности функционируют как ходильные ноги. Они имеют длинные внутренние и короткие наружные ветви и лишены эпиподитов.
Брюшной отдел длиннее головы и груди вместе взятых. Все его 6 сегментов сильно развиты, они толще и длиннее, чем предшествующие им задние свободные грудные сегменты. Пять передних пар брюшных конечностей довольно крупные, двуветвистые, листовидные, причем обе ветви по краям снабжены многочисленными перистыми щетинками. При помощи зубцов, имеющихся на внутреннем крае основания каждой ножки, левая и правая конечности каждой пары могут соединяться между собой и производить взмах как единое целое.
Функции передних брюшных ножек весьма разнообразны. Благодаря их взмахам ротоногие плавают. Кроме того, на всех: передних брюшных ножках располагаются жабры, имеющие вид тонкостенных, многократно ветвящихся придатков. Так, например, у обыкновенного рака-богомола Средиземного моря — Squilla mantis длиной 15 см — на брюшных ножках насчитывается в общей сложности около 10 000 жаберных придатков (основной придаток каждой из 10 ножек разветвляется на 30 вторичных ветвей, каждая из которых дает 25—30 третичных ответвлений) длиной около 1 см при диаметре 2 мм. Дыхательная поверхность всех жабр составляет огромную величину порядка 600 см2. Брюшные ножки непрерывно совершают последовательные взмахи со скоростью 25—40 раз в минуту, возбуждая таким образом ток воды, достаточный для дыхания. Когда рак сидит в норе, эти взмахи приводят к постоянной смене воды в его жилище.
Наконец, у самцов внутренние ветви первой и второй пар брюшных ножек преобразованы в копулятивный аппарат. Последняя пара брюшных ножек — уроподы — плоские, сильно хитинизированные и направлены назад, располагаясь по бокам крупного лопатообразного тельсона. Его задний конец снабжен зубцами, а спинная поверхность — различными килями и буграми. Продольные кили есть и на поверхности брюшных сегментов. Они устроены так, что вода поступает в нору при движении брюшных ножек определенным образом направленными струями.
Большинство ротоногих роет норы в морском грунте. Мелкие виды родов Gonodactylus и Coronida прячутся в расселинах и щелях между ветвями кораллов, а мелкий (2,7 см длины) Lysiosquilla vicina использует для жилья норы Balanoglossus, причем оба животных обитают в одной норе, по-видимому не мешая друг другу.
Раки-богомолы зарываются в грунт передним концом тела, орудуя рострумом и ногочелюстями и выбрасывая грунт из норы брюхом, тельсоном и уроподами.
Готовая нора обычно имеет два выхода, и вода, направляемая взмахами передних брюшных ножек, свободно протекает через нее. Норы Lysiosquilla excavathrix достигают глубины 1 м, Squilla mantis обычно зарывается на глубину около 15 см.
Большую часть времени раки-богомолы проводят в норах. Вылезая наружу, они ползают по поверхности грунта при помощи задниг грудных ног, а также ловчих ногочелюстей, которые при этом сгибаются и на которые рак опирается, как на костыли. Раки могут и довольно быстро плавать. При этом они гребут передними брюшными ножками, ногочелюсти сложены и прижаты к телу, а чешуйки задних антенн, тельсон и уроподы изгибаются под разными углами и служат рулями глубины. Кроме того, животные могут передвигаться скачками вперед и назад, резко изгибая и выпрямляя тело.
Squilla mantis и, вероятно, другие виды обладают очень хорошим зрением. Лучше всего рак видит объекты, находящиеся от него на расстоянии 10 см. Доказано, что число зрительных элементов, участвующих при этом в зрении, у него такое же, как у человека, смотрящего на тот же объект с дистанции 1,2—1,5 м. Рак свободно различает на таком расстоянии креветок длиною 3—5 см, но обращает внимание преимущественно на двигающиеся объекты. Лучше всего Squilla видит при сравнительно слабом освещении (10—20, до 100 люксов), соответствующем тому, которое господствует на глубине его обитания (60—100 м).
Как уже указывалось, зрение имеет важное значение при добывании пищи раками-богомолами.
Большинство этих животных охотится на других ракообразных, прежде всего на креветок, но Lysiosquilla охотно нападает также на рыб и моллюсков, a Gonodactylus поедает актиний.
Икрометание продолжается около 4 часов. При этом самки сгибают брюшко и вместе с яйцами выделяют липкий секрет, который склеивает яйца в крупные яйцевые мешки. Формирование яйцевого мешка осуществляется главным образом тремя задними ногочелюстями. Они придают ему форму эллипса, длиной у Squilla mantis до 14 и шириной до 10 см. В таком мешке у этого вида содержится до 50 000 яиц, расположенных в три слоя. У мелких Gonodactylus длина мешка всего 8—14 мм. Яйцевой мешок помещается между ногочелюстями самки, и она носит его в течение 10—11 недель. В это время она не питается.
Из яиц выходят личинки, разные у разных родов. У большинства родов, в том числе у Squilla, из яйца появляется так называемая псевдозоэа. Она обладает плоским карапаксом с длинным рострумом и с двумя парами длинных шипов на переднем и заднем краях, стебельчатыми глазами, сформированными головными придатками и двумя передними парами ногочелюстей, из которых вторая уже имеет характерное строение ловчих ног. Брюшные сегменты также хорошо развиты, за исключением последнего, еще слитого с тельсоном, и несут плавательные ножки (рис. 232,2). Псевдозоэа сначала живет на дне, но после двух линек, происходящих у Squilla в течение 2—3 дней, при длине 2—2,5 мм переходит к планктонному существованию. В толще воды она линяет еще 4 раза и превращается в стадию синзоэа длиною 8 мм, у которой развиты все грудные конечности и которая и по другим признакам более сходна с взрослыми раками-богомолами. Однако синзоэа также плавает в толще воды и, только перелиняв еще 4 раза, опускается на дно и дает начало маленькому ротоногому рачку.
У родов Lysiosquilla и Coronida личиночное развитие начинается с планктонной стадии антизоэа. В отличие от псевдозоэа ее глаза сидячие, брюшной отдел короткий и нерасчлененный, зато уже обособлены все грудные сегменты (рис. 232,3). Пять пар передних грудных ножек двуветвисты и служат для плавания. После пяти линек, в результате которых у личинки развиваются брюшные сегменты и недостающие конечности, антизоэа переходит в только что описанную стадию синзоэа. У Lysiosquilla в Неаполитанском заливе личиночное развитие занимает 2—3 месяца.
Личинки ротоногих вскоре после рождения начинают вести хищный образ жизни. Псевдозоэа ловит мелких животных ловчими ножками, а антизоэа сначала шипами передних челюстей; после третьей линьки у нее появляются ловчие ноги и функции добывания пищи переходят к ним. Достаточно продолжительный период планктонного существования обеспечивает разнос личинок течениями и способствует расселению раков-богомолов.
Из 170 известных видов этого отряда подавляющее большинство обитает в тропических и субтропических морях на небольшой глубине. В отдельных случаях раки-богомолы опускаются до глубины 760 м. Как правило, они избегают сколько-нибудь значительного опреснения, но некоторые тропические виды могут обитать и в солоноватых водах, а один из них даже проникает в низовья рек на острове Тайвань. За пределами тропической и субтропической зон раки-богомолы встречаются в Средиземном и Адриатическом морях (Squilla mantis, Lysiosquilla maculata и др.).
В морях СССР зарегистрирован всего один вид — Squilla oratoria, попадающийся в Японском море и на мелководьях залива Петра Великого.
НАДОТРЯД ПЕРАКАРИДЫ (PERACARIDA)
Этот обширный надотряд объединяет очень различных как по внешнему виду, так и по образу жизни ракообразных, обитающих в море, в пресных водах и на суше, ползающих по дну и плавающих в толще воды, большей частью свободноживущих, но частично и паразитирующих на других животных. Однако все они обладают некоторыми общими признаками, указывающими на их родство между собой.
Голова всех перакарид срастается с одним или изредка с двумя передними грудными сегментами, конечности которых превращаются в ногочелюсти. Правая и левая жвалы устроены различно: на левой обычно есть подвижная пластинка, отсутствующая на правой. Самки вынашивают яйца под грудью в выводковой сумке, образованной направленными внутрь листовидными отростками грудных ножек (только у отряда Thermosbaenacea выводковая сумка помещается на спинной стороне тела, под карапаксом). Из яиц выходят почти вполне сформировавшиеся молодые рачки, так что настоящих личинок у перакарид нет.
К надотряду Peracarida относятся отряды мизид, кумовых, клешненосных осликов, спелеогрифовых, термосбеновых, равноногих и разноногих ракообразных.
ОТРЯД МИЗИДЫ (MYSIDACEA)
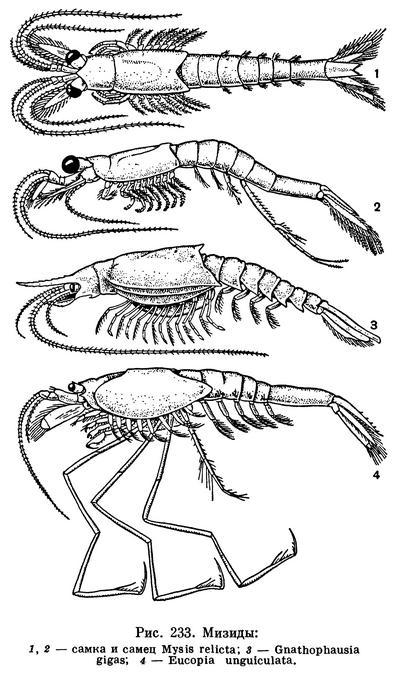
По внешнему виду мизиды несколько напоминают маленьких креветок. Тело их вытянуто в длину, глаза стебельчатые, голова и грудь покрыты цилиндрическим карапаксом, брюшко тонкое и заканчивается хвостовым веером, состоящим из пластинчатого тельсона, по краям которого расположены плоские двуветвистые уроподы. Подобно креветкам они плавают у дна или в толще воды.
Однако при более внимательном рассмотрении мизид нетрудно заметить их важные отличия от креветок. Карапакс мизид срастается не больше чем с тремя передними, а не со всеми грудными сегментами. Над задними грудными сегментами он просто нависает. У мизид всего одна пара ногочелюстей, да и то очень слабо специализированных, а не три, как у креветок, клешней, как правило, нет. Самки мизид вынашивают яйца под грудью в выводковой сумке, а не на брюшных ножках, как креветки. По-разному устроены и жвалы обеих групп ракообразных. Трудно сомневаться в том, что мизиды и креветки приобрели общий облик независимо друг от друга. Это объясняется их сходным способом передвижения.
Мизиды распространены чрезвычайно широко. Всего их известно около 500 видов. Они обитают во всех морях и океанах, как у дна, так и в толще воды. Некоторые виды проникают из морей в реки, причем поднимаются по ним очень высоко. Каспийские и азовско-черноморские мизиды заселили не только Волгу, Днепр, Дон, Донец и их притоки, но и водохранилища, построенные на этих реках. Есть и чисто пресноводные виды, как например обычная в крупных озерах северной части Европы и Америки Mysis relicta. Несколько своеобразных видов открыто в пресных подземных водах.
Многие мизиды служат излюбленной пищей рыб, в том числе промысловых. Для улучшения условий питания рыб как во вновь созданных водохранилищах, так и в ранее существовавших водоемах у нас в стране были в широких масштабах произведены искусственные переселения мизид. Их перевезли в Аральское море, в озеро Балхаш и во многие водохранилища Украины. В последнее время и в США осуществляется заселение мизидами водоемов, в которых раньше этих рачков не было.
Длина тела большинства мизид 10—20 мм, но среди глубоководных видов многие гораздо крупнее (до 18 см). Тело мелководных видов полупрозрачное. У многих мелководных видов через тонкие покровы просвечивают находящиеся в соединительной ткани крупные звездчатые клетки — хроматофоры, заполненные темным пигментом. Они расположены по одной на верхней стороне брюшных сегментов и тельсона, а более мелкие разбросаны по другим частям тела. На свету или на светлом грунте зерна пигмента собираются в центре хроматофора и рачок светлеет, в темноте или на темном грунте пигмент рассеивается по всей клетке, что приводит к потемнению рачка. Эти передвижения пигмента в хроматофорах, так же как у десятиногих ракообразных, регулируются деятельностью находящихся в глазных стебельках органов внутренней секреции. Глубоководные мизиды, как правило, лишены хроматофоров и окрашены в красный цвет.
Карапакс мизид обычно гладкий, без выростов, с коротким рострумом или без него, и только у видов глубоководного рода Gnathophausia снабжен длинным рострумом, продольными килями и оттянутыми в шипы задне-боковыми углами (рис. 233, 3). Стебельчатые глаза с многочисленными фасетками на конце, но у многих глубоководных и пещерных видов фасетки и пигмент отсутствуют, а у глубоководного рода Pseudomma оба слепых глаза срастаются между собой, образуя единую пластинку. Передняя и задняя антенны очень длинные. Передняя антенна с двумя жгутами, богатыми чувствительными щетинками и цилиндрами, а у самцов между обоими жгутами находится специальный чувствительный придаток, помогающий отыскивать самок. Наружная ветвь задней антенны превращена в чешуйку, а внутренняя имеет вид длинного жгута. Из ротовых придатков наибольший интерес представляют передние челюсти: их наружная ветвь преобразована в широкую пластинку, движения которой обеспечивают смену воды под краем карапакса, что необходимо для процесса дыхания. Внутренние лопасти обеих челюстей и щупик задней челюсти снабжены многочисленными щетинками и, пригибаясь друг к другу, образуют фильтрующую камеру, ограниченную спереди большой верхней губой. Большинство мизид питается отфильтровывая мелкие взвешенные в воде частицы, но, кроме того, они могут захватывать и крупные куски отмерших растений и животных и даже трупы себе подобных, которые они разрывают и размельчают жвалами.
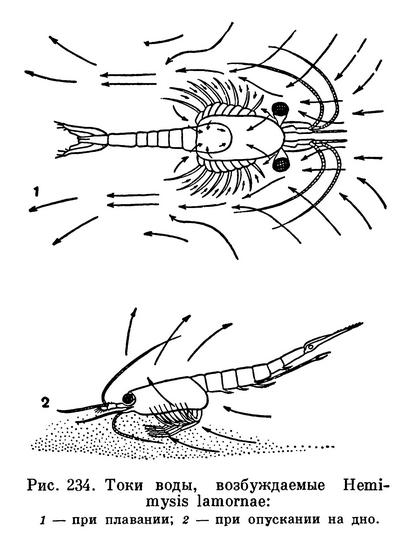
Все 8 пар грудных ножек снабжены наружными ветвями — экзоподитами. Передняя из них — ногочелюсть — отличается от остальных меньшими размерами и присутствием внутренних лопастей при основании. Она принимает участие в процессе фильтрации и захватывания пищевых частиц. Остальные служат для передвижения. Их экзоподиты совершают взмахи, благодаря которым по бокам тела рачка возникают два круговорота воды, проталкивающие животное вперед (рис. 234). Перебирая внутренними ветвями грудных ножек, мизиды ползают по дну. У некоторых видов часть этих внутренних ветвей расширена и приспособлена для рытья грунта. Такие виды могут очень быстро зарываться в поверхностный слой грунта. У подотряда Lophogastrida, который объединяет почти исключительно глубоководные виды, на основных частях грудных ножек находятся разветвленные жабры (эпиподиты), остальные многочисленные мизиды относящиеся к другому подотряду — Mysida, лишены жабр; газообмен у них совершается через тонкие стенки карапакса, в полостях которых циркулирует кровь. Некоторые хищные глубоководные мизиды имеют длинные ловчие ноги, приспособленные для захватывания других животных. У половозрелых самок всех мизид к основаниям грудных ножек прикреплены направленные внутрь пластинки (оостегиты), образующие выводковую сумку.
Длинный и тонкий брюшной отдел мизид состоит из 6 цилиндрических, очень подвижных друг по отношению к другу сегментов, из которых последний самый крупный. У рода Gnathophausia на нем ясно заметен поперечный шов, свидетельствующий о том, что этот сегмент двойной, представляющий результат слияния шестого и седьмого сегментов. Действительно, у эмбрионов мизид шестой и седьмой брюшные сегменты самостоятельны и срастаются только на поздних стадиях развития. Очевидно, некогда мизиды имели 7 брюшных сегментов подобно тонкопанцирным.
У всех мизид хорошо развиты конечности шестого брюшного сегмента — уроподы. Они двуветвистые, плоские и расположены по бокам крупного пластинчатого тельсона, образуя вместе с ним хвостовой веер. В основании внутренней ветви уроподов у всех мизид, за исключением Lophogastrida, находится орган равновесия — статоцист. На стенках полости статоциста расположены чувствительные щетинки (у Praunus около 60), на которые опирается статолит, состоящий из органического вещества и известковых наслоений. Он давит на те или иные чувствительные щетинки, и рачок, ощущая это давление, повертывает свое тело так, чтобы спинная его сторона была обращена вверх. Конечности передних пяти брюшных сегментов хорошо развиты только у Lophogastrida и рода Boreomysis, у которых они служат основным органом плавания. У остальных мизид они в той или иной степени редуцированы, особенно у самок. У самцов одна из них (часто четвертая пара) имеет удлиненную ветвь, помогающую при оплодотворении яиц.
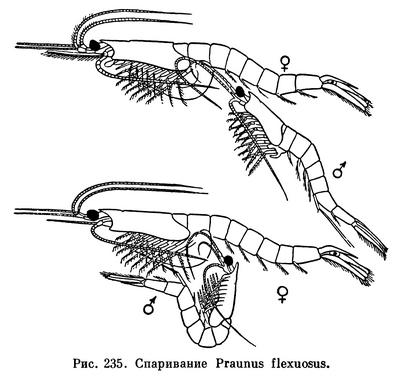
Спаривание мизид происходит ночью. Оно наблюдалось лишь у немногих видов. Самцы при помощи химического чувства отыскивают недавно слинявших самок и плавают под ними брюшной стороной вверх. Приблизившись к самке, самец Mesopodopsis просто выпускает сперматозоиды в воду, а затем направляет их удлиненными внутренними ветвями четвертых брюшных ножек в выводковую камеру самки. Самец Hemimysis и Praunus схватывает самку на 1—2 секунды внутренними ветвями грудных ножек и вводит сперму в ее выводковую сумку при помощи трубкообразных отростков последнего грудного сегмента, в которые открываются его семепроводы. Такой способ спаривания, не связанный с необходимостью длительно удерживать самку, объясняет одну своеобразную особенность мизид: в отличие от большинства других высших ракообразных их самцы мельче и слабее самок.
Через 20—30 минут после проникновения спермы в выводковую сумку самка откладывает туда же яйца. Число яиц различно у разных видов и, кроме того, зависит от размера, т. е. от возраста самки. Так, например, самка пресноводной Mysis relicta длиной 13—15 мм откладывает 10—20, а длиною 17—21 мм — 25—40 яиц; крупные, пережившие зиму самки Neomysis vulgaris в районе г. Киля носят до 55 яиц в выводковой сумке, а мелкие (10 мм) самки, родившиеся летом, — всего 9 яиц. В Северном Каспии перезимовавшие самки Paramysis baeri откладывают весной в среднем 89, максимально 156 яиц.
Яйца развиваются в выводковой сумке, причем составляющие ее пластинки двигаются так, что вода в сумке все время сменяется. Вышедшие из яиц молодые рачки также первое время остаются в выводковой камере матери, а окрепнув, переходят к самостоятельному существованию. Вынашивание яиц и эмбрионов у разных видов длится разное время, при высокой температуре оно заканчивается гораздо раньше, чем при низкой, В прибрежных водах Индии, близ Мадраса, при температуре воды 25—29°С вынашивание яиц и эмбрионов Mesopodopsis indica продолжается всего 4 дня, а обитатель холодных (до 14°С) озер Mysis relicta носит яйца и эмбрионов в течение 2—3 месяцев. В холодном Белом море у Mysis oculata, Mysis mixta и Mysis litoralis вынашивание яиц и эмбрионов растягивается на 6—7 месяцев.
Рост молоди, вышедшей из выводковой камеры матери, также зависит от температуры. В хорошо прогреваемом мелководном Северном Каспии родившиеся в мае рачки уже в июне достигают половозрелости и приступают к размножению. До наступления зимы северокаспийские мизиды успевают дать 4 поколения. За весь период размножения одна пара Paramysis baeri может произвести на свет 430 000 потомков.
Многие виды совершают регулярные горизонтальные и вертикальные миграции. Neomysis integer, Mesopodopsis slabberi и Praunus flexuosus осенью откочевывают от берегов, зиму проводят на сравнительно больших глубинах, а весною возвращаются на мелководье. В Рижском заливе зимой мизиды также концентрируются на глубине, а весной переходят на мелководье. При этом более теплолюбивая Neomysis vulgaris занимает глубины до 15 м, на глубине от 15 до 25 м преобладает Mysis mixta, а на большей глубине обитает холодолюбивая Mysis relicta.
Мигрирующие виды нередко образуют стаи или косяки. Мигрирующие стаи Neomysis integer достигают 1 км в длину и от одного до многих метров в ширину.
Не менее характерны для мизид и вертикальные миграции. Многие мелководные виды днем живут на дне, а ночью поднимаются в поверхностные слои воды. Gastrosaccus sanctus зимой появляется в поверхностных слоях воды в 4 часа, а летом в 9 часов вечера, т. е. в обоих случаях в сумерки. Суточные вертикальные миграции широкого диапазона свойственны каспийским мизидам Mysis microphthalma, Mysis amblyops и Paramysis loxolepis. Ночью они поднимаются в поверхностный 50-метровый слой, а днем опускаются до 600—750 м. Четвертая пелагическая мизида Каспия — Mysis caspia — совершает суточные вертикальные миграции между 150 и 400 м.
Среди мизид, обитающих в наших южных морях, многие способны существовать при различной солености, причем несколько видов выдерживает даже полное опреснение. Такие виды, как Paramysis ullskyi, Paramysis lacustris, Paramysis intermedia, Paramysis baeri, Limnomysis benedeni и некоторые другие, распространены не только во всем Каспийском море и в опресненных участках Черного и Азовского морей, но заходят и во впадающие в них реки — Волгу, Урал, Терек, Дон, Днепр, Донец, Прут, Дунай и др. Особенно широко расселилась по речным системам Paramysis ullskyi, поднявшаяся по Волге выше Рыбинска. Некоторые из речных мизид сильно размножились и в построенных на наших реках водохранилищах, благодаря чему улучшились условия питания обитающих там рыб. Как уже указывалось, мизид переселили в такие водохранилища и другие водоемы, куда они не могли проникнуть самостоятельно. Так, например, понто-каспийские мизиды Paramysis lacustris, Paramysis intermedia, Limnomysis benedeni успешно размножились во многих водохранилищах Украины и Литвы, а также в озере Балхаш и в Аральском море и используются там в пищу промысловыми рыбами.
Не только понто-каспийские мизиды вселяются в пресные воды. В северной части Тихого океана некоторые виды рода Neomysis заходят в низовья рек и связанные с ними озера, есть такие виды и в Юго-Восточной Азии, на Зондских островах, в Австралии и т. д.
Особый интерес представляет распространение северной пресноводной мизиды Mysis relicta. Она живет в крупных холодных чистых озерах Карелии, в Ладожском и Онежском озерах, в нескольких озерах Белоруссии и Смоленской области, в устьях северных рек, в озерах северной Германии и скандинавских стран, в некоторых озерах Канады и США. Расселение Mysis relicta по всем этим озерам связано с четвертичным оледенением, когда перед краем наступающего или отступающего ледника возникали огромные водоемы, фауна которых частично содержала и виды морского происхождения. Вместе с Mysis relicta в таких остаточных озерах обычно обитают веслоногий рачок Limnocalanus grimaldii и некоторые виды бокоплавов. Mysis relicta, как правило, живет при температуре не выше 14°С, при высоком содержании кислорода в воде. Рачок совершает регулярные суточные миграции, поднимаясь ночью в поверхностные слои воды.
Практическое значение мизид довольно велико, поскольку они служат излюбленной пищей многих рыб, в том числе промысловых. Снеток и некоторые сиги питаются преимущественно Mysis relicta, молодь судака в большом количестве поедает мизид, прежде чем перейти к потреблению рыб. Мизиды усиленно поедаются салакой в Балтийском море, кильками и некоторыми сельдями в Каспийском море, многими рыбами в наших северных и дальневосточных морях.
В некоторых странах Юго-Восточной Азии мизид охотно едят и люди. Из рачков приготовляют пасту или соус, которыми сдабривают рис. На острове Ява это кушанье носит специальное название «джембрет».
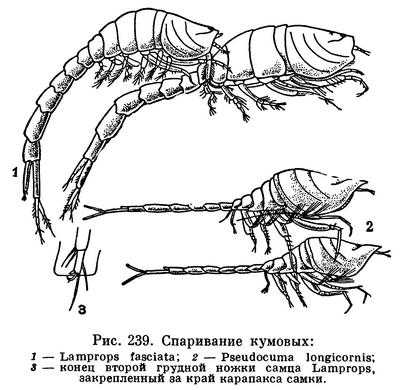
ОТРЯД КУМОВЫЕ РАКООБРАЗНЫЕ (CUMACEA)
Кумовые ракообразные по некоторым особенностям своей организации близки к мизидам, но по образу жизни довольно резко от них отличаются. Все кумовые приспособлены к зарыванию в поверхностный слой морского грунта и лишь на короткие промежутки времени покидают грунт и плавают или ползают по его поверхности. Несмотря на сравнительно большое количество относящихся к этому отряду видов (около 600), все они имеют сходное строение, соответствующее их однообразной биологии. Они обитают почти исключительно в море, и лишь несколько понто-каспийских видов и один дальневосточный вид встречаются в пресных водоемах. Длина тела этих рачков обычно не достигает 1 см, и лишь немногие виды крупнее (до 35 мм).
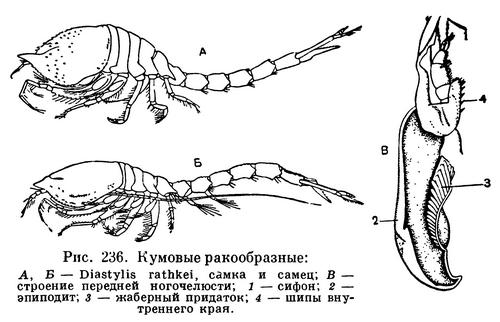
Тело кумовых резко разделяется на более широкую переднюю часть, включающую голову и грудь, и тонкую заднюю часть, состоящую из брюшка и тельсона. Передняя часть тела покрыта коротким карапаксом, оставляющим свободными 5, реже меньше задних грудных сегментов. Строение карапакса весьма своеобразно. Передне-боковые его углы вытянуты вперед и сближены между собой так, что образуют часто довольно длинный так называемый «ложный рострум». За основанием ложного рострума на поверхности карапакса находится непарный глаз, состоящий из двух соприкасающихся друг с другом фасеточных глаз. Число фасеток в каждом из них очень невелико по сравнению с тем, которое свойственно другим высшим ракообразным, обычно 3—4, самое большее 11, и зрение не играет существенной роли в жизни кумовых ракообразных. Боковые края карапакса сзади и с боков плотно прилегают к стенкам тела, и только по бокам от ложного рострума остаются отверстия, ведущие слева и справа в пространства между краем карапакса и стенкой тела; эти пространства представляют собой жаберные полости. Такое их строение позволяет рачку дышать, когда он сидит, зарывшись в грунт и выставив над его поверхностью только самый передний конец тела. Грунт не попадает в защищенные краями карапакса жаберные полости, а вода проникает в них через отверстия по бокам рострума. Передние антенны очень короткие, каждая с двумя жгутами, как у мизид. Задние антенны у самок также очень короткие, но у половозрелых самцов они удлинены, нередко превосходят по длине тело и богаты чувствительными щетинками. С их помощью самцы отыскивают самок. Жвалы лишены щупиков. На обеих парах челюстей с внутренней стороны находятся выросты с фильтрующими щетинками.
Три передних, покрытых карапаксом сегмента грудного отдела срастаются с головой, и их конечности превращены в ногочелюсти. Первая из них короткая, но устроена особенно сложно и выполняет несколько ответственных функций. Левая и правая ногочелюсти сцеплены воедино шипами на внутреннем крае и направлены вперед. Крупный эпиподит каждой ножки разделен на две части. Передняя из них также направлена вперед, и конец ее свернут так, что образует нечто вроде трубки; это сифон, по которому вода проходит в жаберную полость. Задняя часть направлена назад, она тонкостенная, листовидная и снабжена жаберным придатком, на котором сидят в ряд 4—12, иногда до 40 пальцевидных жаберных листочков. Задняя часть эпиподита первой ногочелюсти служит основным органом дыхания, но в дыхании участвует и внутренняя поверхность стенки карапакса. Ток воды в жаберную полость вызывается движениями пластинчатой задней части эпиподита, совершающей около 40 взмахов в минуту. Остальные две пары ногочелюстей гораздо длиннее и служат главным образом для захватывания пищевых частиц и передачи их вперед, к ротовому отверстию. Задняя из них снабжена многочисленными перистыми щетинками, при помощи которых она очищает жаберные полости от попадающих туда посторонних частиц.
На пяти свободных задних грудных сегментах расположено по паре ходильных ножек. Все они, обычно за исключением последней пары, помимо внутренних, имеют многочлениковые, несущие многочисленные щетинки наружные ветви, при помощи которых рачки плавают. Внутренние ветви служат для разгребания грунта, а передние, кроме того, загребают пищевые частицы. У половозрелых самок на последней паре ногочелюстеи и на трех передних ходильных ножках возникают пластинки, образующие выводковую сумку.
Брюшко тонкое, длинное, цилиндрическое, и составляющие его 6 сегментов подвижно сочленены между собой. Поэтому брюшко может легко изгибаться как на спинную, так и на брюшную сторону. Передние брюшные конечности у самок отсутствуют, а у половозрелых самцов частично или полностью развиваются. Они короткие, обычно двуветвистые, с длинными щетинками и служат для плавания. К последнему брюшному сегменту у обоих полов прикреплены направленные назад длинные тонкие двуветвистые уроподы. Загибая брюшко на спинную сторону, рачок может очищать ими спинную поверхность карапакса и свободных грудных сегментов. Расположенный между основаниями уроподов тельсон имеет различную форму у разных видов, а у некоторых срастается с последним брюшным сегментом.
Большую часть жизни, во всяком случае светлое время суток, кумовые проводят зарывшись в поверхностный слой грунта. Опускаясь на дно, рачок загибает вверх брюшко и передний конец тела и начинает разгребать грунт внутренними ветвями трех пар задних грудных ног. Им помогает изгибающееся в разные стороны брюшко. В конце концов все тело рачка погружается в грунт и только его самый передний конец оказывается на уровне поверхности грунта. У некоторых видов над грунтом торчат и концы уроподов. Изогнув брюшко на спинную сторону, рачок спокойно сидит в грунте. При помощи задних ногочелюстей и передних ходильных ног он вырывает перед своим ротовым отверстием воронку и приступает к питанию (рис. 237).
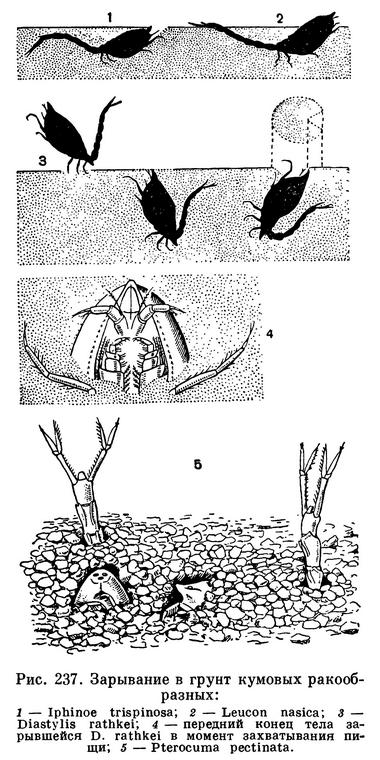
Виды, зарывающиеся в песчаный грунт (Lamprops, Pseudocumidae, Cumopsis и др.), удлиненными внутренними ветвями передней пары ходильных ног подгребают к ротовому отверстию, находящемуся на дне воронки, отдельные песчинки. Ногочелюсти передают их челюстям и жвалам, которые счищают с песчинок налет из водорослей и бактерий, служащих пищей рачкам. Некоторые кумовые могут использовать таким образом частицы грунта, не уступающие по размерам их карапаксу. «Обработанные» частицы отбрасываются назад через голову. Другие виды, живущие на мягком илистом грунте (Diastylis, Cumella и др.), разводят в стороны внутренние ветви своих последних ногочелюстей и начинают пропускать ил через щетинки передних ногочелюстей и челюстей. Бактерии и находящиеся в грунте разные органические частицы поедаются, а минеральные частицы направляются вперед между основаниями передних антенн.
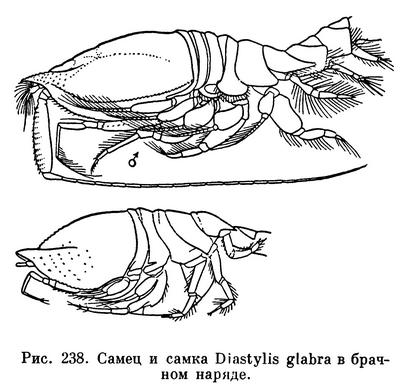
Спаривание происходит в ночное или сумеречное время в толще воды, вне грунта, после линьки самки. Самец Lamprops прикрепляется к спинной стороне тела самки, держась за края ее свободных грудных сегментов своими грудными ножками, снабженными специально приспособленными для этого коготками и щетинками, а также жгутами задних антенн. Остается пока неизвестным, оплодотворяются ли яйца в выводковой сумке или в половых путях самки. Самка откладывает в выводковую сумку яйца, число которых различно у разных видов (от 3 до 200) и, кроме того, зависит от размера самки. Так, например, в выводковой сумке самок Diastylis rathkei длиной 10 мм насчитывается от 12 до 49 яиц, а у самок того же вида длиной 15 мм — от 85 до 102 яиц.
Кумовые ракообразные широко распространены во всех морях и океанах. Некоторые семейства, как например Diastylidae, Leuconidae, Lampropidae, приурочены к холодным водам и встречаются только в высоких и умеренных широтах северного и южного полушарий, отсутствуя в тропиках, другие, как Bodotriidae и Nannastacidae, наоборот, приурочены к тропической и субтропической областям.
Многие виды способны жить как на малых, так и на больших глубинах. Так, например, Leucon fulvus и Leucon pallidus, населяют глубины от 9—10 до 2560 м, Eudorella emarginata — от 9 до 2000 м и т. д. Однако большинство кумовых (в наших дальневосточных морях 70% известных видов) не опускается глубже материковой отмели. С другой стороны, известен один род (Makrokylindrus), объединяющий только глубоководные виды.
Весьма своеобразна фауна кумовых Каспийского моря. Она состоит из 17 видов, принадлежащих к семейству Pseudocumidae. Из них 6 ограничены в своем распространении Каспием, а остальные встречаются, кроме того, в опресненных частях Черного и Азовского морей. Некоторые из этих видов заходят в русла рек и поднимаются по ним вверх на 100—150 и иногда даже на 350 км от устья. Единственным чисто пресноводным видом кумовых можно считать Lamprops korroensis, обитающего в озерах и низовьях рек Охотского моря, Сахалинского залива, залива Де-Кастри, Сахалина и Курильских островов.
Кумовые ракообразные используются в пищу многими рыбами, в частности промысловыми. В Каспийском море их поедают молодь осетровых, вобла, некоторые бычки и особенно охотно лещ, в Азовском и Черном морях — тарань, камбалы, бычки. В Баренцевом море и в дальневосточных морях они входят в состав пищи пикши, трески, камбал, морского окуня и других рыб.
Учитывая важное кормовое для рыб значение кумовых, их акклиматизируют в некоторых водохранилищах. Так, например, Pseudocuma cercaroides была перевезена в Днепровское водохранилище, где прижилась и размножилась.
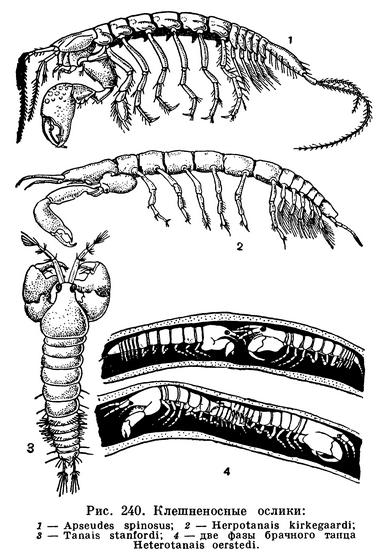
ОТРЯД КЛЕШНЕНОСНЫЕ ОСЛИКИ (TANAIDACEA)
При сборах донных морских животных наряду с кумовым, бокоплавами, равноногими изредка попадаются мелкие высшие ракообразные, принадлежащие к отряду клешненосных осликов, или танаидовых. Они ведут скрытый образ жизни, укрываясь в построенных ими трубках, зарываясь в грунт или прячась среди водорослей, камней и т. д. Клешненосные ослики встречаются как на мелководье, так и на больших океанических глубинах, но нигде не образуют массовых скоплений. На Курильских островах и в Южной Америке один или два вида обитают и в пресных водах, но, как правило, клешненосные ослики не переносят опреснения и могут считаться настоящими морскими животными. Длина их тела составляет обычно от 2 до 5 мм, и только некоторые глубоководные виды достигают значительно более крупных размеров (Herpotanais kirkegaardi до 26 мм). Из-за сравнительной редкости и немногочисленности этих рачков их биология мало изучена. Описано около 250 видов клешненосных осликов.
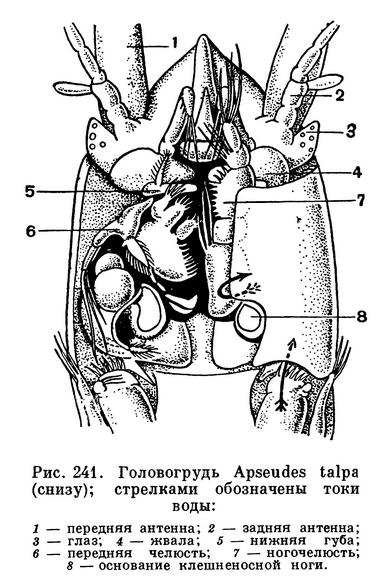
Тело этих ракообразных удлиненное, цилиндрическое или уплощенное в спинно-брюшном направлении. Короткий щитовидный карапакс покрывает голову и слившиеся с нею два передних грудных сегмента. Между его боковыми краями и стенкой тела, так же как у кумовых, с каждой стороны находится дыхательная полость. Передний край карапакса часто образует плоский треугольный или полукруглый рострум. У многих мелководных видов по бокам от основания рострума на специальных выростах головы располагаются маленькие глаза с очень небольшим количеством фасеток (у Heterotanais — 12, у Apseudes — 8). Глазные выросты Tanaidaсеа существенно отличаются от глазных стебельков тонкопанцирных, мизид и десятиногих, поскольку они не сочленены с головой и не могут поворачиваться.
Обе пары антенн обычно короткие. У представителей подотряда Monokonophora передние антенны с двумя жгутами, а задние с чешуевидной наружной ветвью, у представителей другого подотряда — Dikonophora — на передних антеннах всего один жгут, а задние лишены наружной ветви. Ротовые придатки хорошо развиты только у самок, а у самцов многих видов они частично или даже полностью редуцируются. Жвалы у Monokonophora несут щупик, отсутствующий на жвалах Dikonophora. Челюсти снабжены внутренними лопастями, многочисленные щетинки которых образуют фильтрующий аппарат. На передней ногочелюсти, так же как у кумовых, имеется крупный эпиподит, приспособленный для возбуждения тока воды в дыхательной полости, под боковым краем карапакса. Вода заходит под край карапакса сзади, омывает дыхательную полость и выходит позади места прикрепления передней ногочелюсти. Кислород усваивается внутренней поверхностью боковых краев карапакса, а также стенкой тела, прилегающей к дыхательной полости. Здесь располагается густая сеть капиллярных кровеносных лакун, впадающих близ заднего края карапакса в околосердечную сумку.
Конечности второго, слившегося с головой грудного сегмента крупнее остальных и снабжены на конце настоящей клешней или у некоторых видов подклешней. Благодаря характерному строению этих ножек клешненосные ослики и получили свое название. У самцов клешненосные ноги обычно бывают крупнее, чем у самок и нередко имеют несколько иное строение, но биологическое значение этих различий пока остается неизвестным. Остальные 6 пар грудных ножек устроены одинаково и представляют собой ходильные конечности. Все они одноветвисты и только у некоторых Apseudidae и Kalliapseudidae на передней из них, а также на клешненосных ногах есть короткие наружные ветви, напоминающие соответствующие придатки мизид и кумовых. Кроме того, у рода Apseudes ноги переднего свободного грудного сегмента отличаются от остальных тем, что их последние членики расширены и приспособлены для рытья грунта. У самок перед размножением на четырех, а у некоторых видов всего на одной паре грудных ножек развиваются пластинки, образующие выводковую сумку. Так же как у кумовых, они исчезают после линьки, следующей за окончанием вынашивания яиц и молоди, а затем, перед новым периодом размножения, возникают вновь.
Брюшной отдел всегда значительно короче и обычно несколько уже грудного. Пять передних составляющих его сегментов свободные, а последний, шестой сегмент срастается с тельсоном, образуя так называемый плеотельсон. Конечности передних пяти брюшных сегментов короткие, но двуветвистые и снабжены по краям многочисленными щетинками. Они служат плавательными ножками и обычно у самцов развиты сильнее, чем у самок. Самки некоторых видов совсем лишены этих ножек. К плеотельсону прикрепляется последняя пара брюшных ножек — уроподы. Они обычно двуветвисты, причем и ствол и обе ветви тонкие, палочковидные. У некоторых клешненосных осликов, например у Apseudes, ветви уроподов удлиненные и расчлененные.
Большинство клешненосных осликов живет в выкапываемых ими норах или в трубках. Внутреннюю поверхность норы рачки обкладывают песчинками, остатками растительности и другими частицами, которые они собирают и перетаскивают клешненосными ногами. Весь этот материал скрепляется специальным секретом. На боках трех передних свободных грудных сегментов располагаются железы, вырабатывающие этот секрет. Протоки желез очень длинные; они тянутся через каждую из трех пар передних ходильных ножек и открываются в их когтевых члениках. Этими ходильными ножками рачки и скрепляют обкладку своих нор и стенки своих трубок. Сидя в норе или в трубке, рачки энергично машут брюшными ножками и таким образом обновляют воду в своем убежище.
Время от времени они вылезают из нор и трубок и передвигаются по дну, частично ползая при помощи грудных, частично плавая над дном при помощи брюшных ножек. Кроме того, они могут двигаться скачками, сгибая и разгибая тело.
Пищей клешненосным осликам служат находящиеся в грунте остатки отмерших растений и другие органические частицы. Некоторые виды отфильтровывают их щетинками челюстей, используя ток воды, направляемый в дыхательные полости.
Самцы клешненосных осликов отличаются от самок большим количеством чувствительных щетинок на жгутах антенн и сильнее развитыми плавательными ножками, что позволяет приписывать им активную роль в поисках самок. Наряду с этим самцы многих видов частично или полностью лишены ротовых придатков, следовательно, не могут питаться и, очевидно, недолговечны.
Механизм определения пола у клешненосных осликов очень интересен. Еще недавно предполагалось, что некоторым из них свойственна протогиния — явление, довольно редкое в животном мире. При протогинии созревающий организм превращается в самку, которая затем переходит в самца.
Новые наблюдения Рамиреца над Heterotanais oerstedi позволяют считать по крайней мере этот вид потенциальным гермафродитом. У него самцы могут возникать либо непосредственно из молодых особей, либо из половозрелых самок, закончивших вынашивание молоди или даже еще не отметавших яиц. Это зависит от условий существования рачков. При совместном содержании самца и самки в одном сосуде перемены пола не происходит. При содержании молодой особи вместе с самцом из нее получается самка и, наоборот, если ее подсадить к самке, то — самец. Содержание молодой особи в одиночестве приводит к превращению ее в самку. Таким образом, пол у Heterotanais oerstedi не связан с присутствием определенного гена.
Гермафродитные особи известны не только для Heterotanais oerstedi, но также для крупного (15 мм) антарктического Apseudes spectabilis, Tanais stanfordi и глубоководного Neotanais serratispinosus. При этом у широко распространенного Tanais stanfordi (рис. 240, 3) гермафродитизм наблюдается только в отдельных частях его обширного ареала, в частности на Курильских островах, а в других местах этот вид представлен нормальными, не переходящими друг в друга самцами и самками. Замечательно, что все известные до сих пор гермафродитные танаидовые живут при низкой температуре.
Размножение клешненосных осликов удалось изучить лишь недавно, сконструировав для наблюдений специальные микроаквариумы. Самец Heterotanais oerstedi забирается в трубку самки, и в течение 11 часов рачки исполняют там брачный танец. Сначала самец преследует самку, затем она переворачивается через голову, и оба партнера оказываются «лицом к лицу», соприкасаясь антеннами (рис. 240, 4). После этого они поворачиваются друг к другу задними концами тела и контактируются уроподами, для чего самцу приходится предварительно переворачиваться через голову. Наконец, самец оказывается лежащим на спине, а самка располагается над ним. В таком положении (в течение 3 секунд) происходит копуляция, при которой сперматозоиды попадают в выводковую сумку самки. Вслед за этим самец покидает трубку самки, а она закрывает вход в трубку. Через некоторое время самка откладывает в выводковую сумку яйца.
Распространение клешненосных осликов изучено далеко не достаточно. Наибольшее количество их видов приурочено к небольшим глубинам полносоленых морей умеренной зоны и высоких широт. Можно отметить четкие различия в составе фауны этих ракообразных на разных по характеру грунтах — песчаных, илистых и каменистых. На больших и предельных океанических глубинах обитают преимущественно представители семейства Neotanaidae, характеризующегося многими примитивными признаками.

ОТРЯД СПЕЛЕОГРИФОВЫЕ РАКООБРАЗНЫЕ (SPELAEOGRIPHACEA)
Столовая гора близ Кейптауна в Южной Африке богата различными подземными ходами и пещерами. В некоторых из них есть водоемы. При обследовании этих пещер в одном пещерном ручье, температура воды которого равнялась 10°С, были обнаружены небольшие (7,5 мм) слепые и лишенные пигмента рачки. Они были названы Spelaeogriphus lepidops. Их строение оказалось настолько своеобразным, что их пришлось выделить в особый отряд, имеющий некоторые общие признаки с клешненосными осликами и кумовыми. Больше нигде Spelaeogriphacea найти пока не удалось.
Об образе жизни спелеогрифовых ничего не известно.
Тело Spelaeogriphus длинное, немного уплощенное сверху вниз, с параллельными боковыми краями.
Голова срастается только с одним передним грудным сегментом и вместе с ним покрыта коротким карапаксом с направленным вперед коротким треугольным рострумом. Под боковыми краями карапакса расположены, как у кумовых и клешненосных осликов, дыхательные полости. Большой ложковидный эпиподит ногочелюсти функционирует, очевидно, так же как у кумовых и клешненосных осликов, возбуждая ток воды, проходящий мимо сочлененных с головой подвижных глазных стебельков, полностью лишенных фасеток и пигмента.
Конечности семи свободных грудных сегментов устроены приблизительно одинаково и представляют собой ходильные ножки.
Все они, за исключением ножек последней пары, снабжены короткими наружными ветвями, а у самок на передних пяти парах есть направленные внутрь пластинки, образующие выводковую камеру.
Все шесть брюшных сегментов свободные. На передних четырех брюшных сегментах находится по паре двуветвистых плавательных ножек. Их одночлениковые ветви несут по краям многочисленные щетинки, и, судя по их строению, рачок может не только ползать, но и плавать. Пятый брюшной сегмент лишен конечностей, а на шестом помещаются направленные назад двуветвистые уроподы. Тельсон короткий, овальный.
Судя по содержимому кишечника, рачки заглатывают грунт.
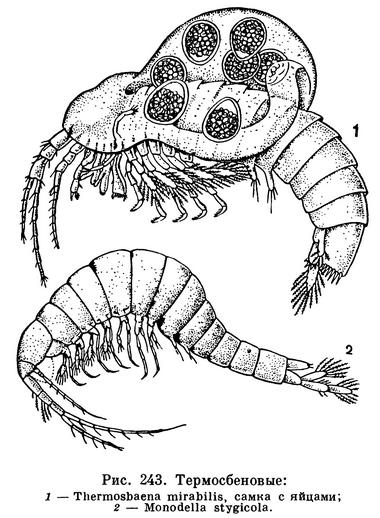
ОТРЯД ТЕРМОСБЕНОВЫЕ РАКООБРАЗНЫЕ (THERMOSBAENACEA)
Среди перакарид есть еще один отряд, представители которого обитают только в подземных водах. Сначала они были открыты в горячем солоноватом ключе в Тунисе, но потом близкие виды удалось обнаружить в пещерах южной Италии, в солоноватых интерстициальных водах морского побережья Югославии, а в последнее время в пещерах США. По своему строению термосбеновые во многих отношениях сходны с кумовыми и клешненосными осликами, но по способу размножения они резко отличаются от всех перакарид: выводковая сумка, в которой самки вынашивают яйца, образуется у них на спинной стороне под краем карапакса, как у ветвистоусых ракообразных, а не под грудью за счет отростков грудных ножек, как у всех остальных перакарид. На этом основании некоторые исследователи считают возможным выделять термосбеновых в особый надотряд Pancarida, но другие, учитывая строение их жвал и многие детали анатомии, включают их в состав надотряда Peracarida. Всего известно 6 видов этих ракообразных, из которых один принадлежит к роду Thermosbaena, а остальные к роду Monodella.
Длина их тела не превышает 3,5 мм. Туловище цилиндрическое, грудной и брюшной отделы одинаковой ширины. Будучи исключительно подземными животными, они совершенно слепы и полностью лишены пигмента. Передний грудной сегмент объединен с головой. Цилиндрический карапакс срастается с головой и передним грудным сегментом, но нависает и над несколькими следующими грудными сегментами. Особенно сильно он разрастается назад у половозрелых самок, у которых он может покрывать все грудные сегменты. У них, как уже указывалось, между карапаксом и спинной стороной тела образуется выводковая сумка, куда откладываются яйца. Внутренняя поверхность карапакса служит органом дыхания.
На всех свободных грудных сегментах у Monodella или только на пяти передних у Thermosbaena находится по паре двуветвистых ходильных ножек. Брюшной отдел, как обычно, состоит из 6 сегментов, но у Thermosbaena последний брюшной сегмент срастается с крупным широким тельсоном, образуя плеотельсон. На двух передних брюшных сегментах имеются короткие нерасчлененные ножки. Последний брюшной сегмент снабжен направленными назад двуветвистыми уроподами.
Thermosbaena может только ползать по грунту, перебирая своими грудными ножками, Monodella argentaria из итальянских пещерных озер, кроме того, плавает, часто поднимаясь к поверхности воды, Monodella halophila, как и другие обитатели интерстициальных вод, пробирается, изгибая свое узкое тело, по капиллярным ходам между песчинками. Все эти рачки заглатывают грунт и усваивают находящееся в нем органическое вещество.
Перед откладкой яиц карапакс самки удлиняется. У самок Monodella от заднего конца каждого яичника вверх отходит канал, по которому яйца, вероятно, проходят в выводковую сумку. Кроме него, есть и другой канал, открывающийся в основании ноги пятого грудного сегмента половым отверстием. Можно предполагать, что именно по последнему каналу проникает сперма после спаривания и яйца откладываются в выводковую сумку уже оплодотворенными, однако ни спаривания, ни откладки яиц у термосбеновых еще никто не наблюдал. У Monodella в выводковой сумке насчитывается от 4 до 16 яиц. Из них там же выходят почти полностью сформированные рачки, отличающиеся от взрослых отсутствием ног на двух последних грудных сегментах. Они покидают выводковую сумку матери и начинают жить самостоятельно. Через неделю они достигают в длину 2 мм, и к этому времени у них появляется сначала шестая, а потом и седьмая пара ходильных ног.
Условия существования немногочисленных известных видов термосбеновых весьма разнообразны. По-видимому, исходной средой их обитания был морской грунт. В наиболее близких к исходным условиях существует Monodella halophila, населяющая заполненные солоноватыми водами интерстициальные ходы адриатического побережья Югославии. В солоноватых водах живет и Monodella relicta из ключей близ Мертвого моря в Израиле; она считается реликтом морей, некогда покрывавших этот район. Два вида Monodella (Monodella argentaria и Monodella stygicola) из пещерных озер южной Италии, по всей вероятности, проникли в эти сравнительно крупные подземные водоемы из системы грунтовых вод, поскольку их местонахождения расположены недалеко от моря и им приписывалось морское происхождение. Однако недавнее открытие Monodella texana в пещерах США, находящихся на расстоянии 200 км от ближайших морских берегов, опровергает сложившиеся ранее представления о связи распространения Monodella со Средиземным морем и сравнительно недавними этапами его геологического прошлого. Вероятно, род древнее, а история его расселения сложнее, чем это ранее предполагалось.
В совершенно необычайных условиях существует первый по времени открытия представитель термосбеновых, по имени которого назван весь отряд — Thermosbaena mirabilis. Теплый ключ в Тунисе — единственное место обитания этого вида — круглый год имеет температуру 45°С. Рачок узко приспособлен к жизни при такой высокой температуре и при понижении температуры до 30°С погибает. Monodella texana и Monodella argentaria живут в пресной, а остальные виды отряда в солоноватой воде.
Обширное и разорванное географическое распространение, а также условия обитания термосбеновых позволяют видеть в их современных немногочисленных представителях остатки древней, в значительной степени исчезнувшей фауны. В этом отношении они подобны цефалокаридам, анаспидовым и батинеллиевым.
ОТРЯД РАВНОНОГИЕ РАКООБРАЗНЫЕ (ISOPODA)
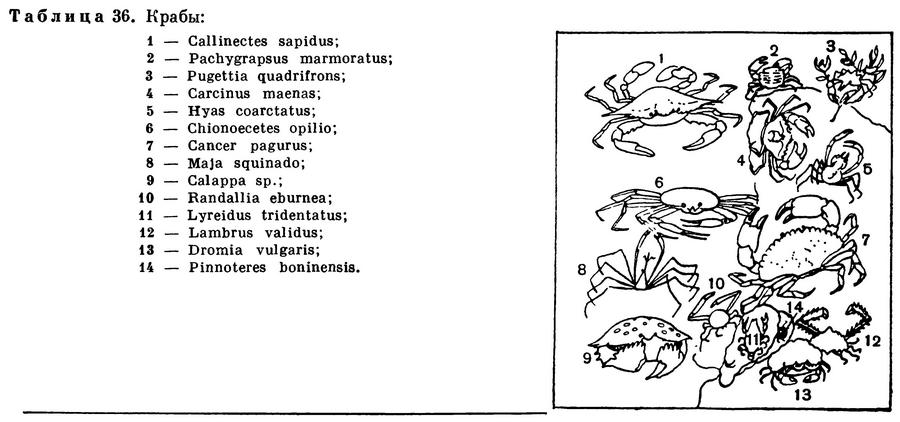
Равноногие ракообразные (табл. 33) — один из немногих отрядов этого класса, многочисленные (4500 видов) представители которого приспособились к самым различным условиям существования. Большинство относящихся к нему видов обитает в море, причем населяет все глубины, начиная от приливо-отливной зоны и кончая предельными океаническими глубинами порядка 11 км. Среди морских равноногих есть обитатели дна, но есть и немногочисленные планктонные виды; есть также и паразиты других ракообразных и рыб. Чрезвычайно широко распространены равноногие и в пресных водах, как поверхностных, так и подземных; пресноводные водяные ослики могут считаться очень обычными и многочисленными представителями фауны самых различных озер и прудов северного полушария. Наконец, богатый видами подотряд равноногих — мокрицы — выработал весьма совершенные приспособления к жизни на суше. Они встречаются почти во всех климатических зонах земного шара — от холодных приполярных районов до влажных тропических лесов и раскаленных пустынь. Очевидно, в процессе эволюции равноногие приобрели какие-то важные особенности, позволившие им впоследствии специализироваться в разных направлениях и освоить различные среды обитания. Одной из таких особенностей можно считать строение брюшного отдела и его конечностей, выполняющих различные функции — половую, двигательную и дыхательную — и хорошо защищенных от неблагоприятных внешних воздействий, в частности у сухопутных рачков от высыхания.
Тело равноногих обычно уплощено сверху вниз, реже цилиндрическое или даже сжатое с боков (у подотряда Phreatoicidea, табл. 33, 4). Его покровы очень разнообразны, часто гладкие, но нередко снабжены килями, шипами или различными бугорками. У некоторых мокриц бугорки поверхности тела образуют сложную водопроводную систему. Если рачок прикоснется к капельке росы, влага под действием капиллярных сил начинает распространяться в промежутках между бугорками, достигая заднего конца тела и орошая находящиеся там жабры. Также разнообразна окраска равноногих. Обычно она бурая или желтоватая, но у видов, живущих на морских растениях, она зеленая, под цвет субстрата, а подземные и глубоководные виды лишены пигмента, белые или иногда розоватые.
Карапакс полностью отсутствует. Голова срастается с одним или изредка с двумя передними грудными сегментами. На ней с боков находятся сидячие глаза. Число составляющих их фасеток колеблется в очень широких пределах: у самого крупного вида равноногих — Bathynomus giganteus (табл. 33, 10), достигающего в длину 27 см, в каждом глазе насчитывается 3000 фасеток, у более мелких хищных морских Aega (табл. 33, 6) — около 400, у мокриц — обычно 20—30, а у водяных осликов — 4. У глубоководных и подземных видов глаза не развиты совсем. Обе пары антенн имеют разную длину у различных видов. У многих глубоководных равноногих задние антенны значительно длиннее тела. Строение ротовых придатков зависит от способа питания. У большинства равноногих они жующего типа, но у некоторых хищников и паразитов жвалы и челюсти превращены в шиловидные острия и сложены вместе в виде пирамиды, так что ротовой аппарат приобретает характер колюще-сосущего.У взрослых особей из подотряда Gnathiidea ротовые придатки, за исключением жвал самцов, редуцированы, и рачки вовсе не питаются (табл. 33, 1,2).
Семь свободных грудных сегментов несут по паре ходильных ног, всегда лишенных наружных придатков. У многих равноногих эти ножки имеют приблизительно одинаковое строение и одинаковую длину, почему отряд и получил свое название. Однако у других они специализированы в разных направлениях. Нередко на одной передней или даже на трех передних ножках развивается хватательное приспособление — подклешня. У многих представителей подотряда Asellota, в особенности часто у глубоководных, конечные членики трех задних пар грудных ножек расширены и снабжены по краям многочисленными щетинками. Такие ножки служат для плавания (табл. 33, 11,12). У половозрелых самок на передних пяти или четырех (у некоторых Arcturidae на одной) парах грудных ножек с внутренней стороны возникают пластинки, образующие выводковую сумку.
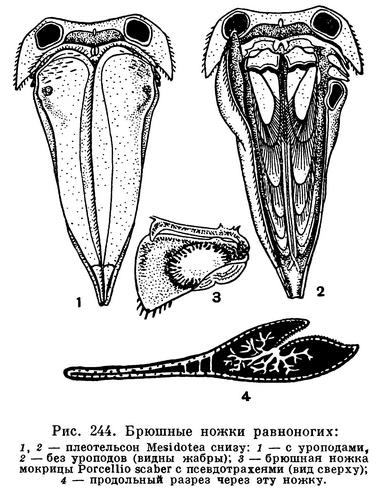
Особенно характерно строение брюшного отдела и его конечностей. Он всегда гораздо короче грудного, и количество составляющих его сегментов сокращено за счет того, что один, несколько или даже все сегменты срастаются с полукруглым или треугольным тельсоном, образуя крупный плеотельсон. Только у подотряда Anthuridea этого не происходит и последний брюшной сегмент и тельсон сохраняют самостоятельность. Конечности пяти передних брюшных сегментов состоят из короткого основания и двух широких листовидных, часто тонкостенных ветвей. Эти ветви накладываются друг на друга, как страницы книги, и направлены назад таким образом, что они находятся под плеотельсоном (рис. 244, 2). Основная функция передних брюшных ножек дыхательная. Они служат жабрами. Через их тонкие стенки растворенный в воде кислород проникает в проходящие в ножках кровеносные лакуны и окисляет кровь. В связи с этим, как уже указывалось, сердце у равноногих ракообразных в значительной степени смещено в брюшной отдел и расположено близко от жабр. Наружные ветви одной из пар брюшных ножек обычно сильно хитинизированы и служат крышкой, покрывающей остальные нижние жаберные листочки, которые, таким образом, оказываются сверху защищенными плеотельсоном, а снизу этой крышкой (рис. 244, 1).
Такое строение дыхательного аппарата послужило, несомненно, предпосылкой, позволившей части равноногих перейти к сухопутному существованию. Их жабры оказались хорошо защищенными от высыхания и не потребовалось дополнительных усовершенствований для того, чтобы полностью их обезопасить в новых условиях существования. Большинство мокриц, несмотря на то что они живут на суше, дышат при помощи своих брюшных ножек, как настоящие водные животные: они используют кислород, растворенный в тонком слое покрывающей их жабры влаги. Следовательно, их жабры должны все время увлажняться. Это достигается у разных мокриц разными путями. У одних, как уже упоминалось, приток воды к жабрам обеспечивается благодаря своеобразной водопроводной системе, образованной скульптурой покровов. У других задние брюшные ножки — уроподы — устроены таким образом, что при соприкосновении с капелькой воды она, также под влиянием капиллярных сил, поступает в жабры по уроподам. Наконец, третьи орошают свои жабры жидкостью, выделяемой из анального отверстия и добываемой из поедаемой рачками сочной растительности. Однако есть мокрицы, способные при дыхании использовать не только растворенный в воде, но и атмосферный кислород. У таких мокриц в толще наружных ветвей передних брюшных ножек находится полость, соединяющаяся щелью с наружной средой, так что в эту полость свободно проникает воздух. От нее отходят ветвящиеся, заканчивающиеся слепо каналы, напоминающие трахеи насекомых и других наземных членистоногих и поэтому называемые псевдотрахеями (рис. 244, 3,4). Через них кислород воздуха проникает в кровь, циркулирующую в лакунах в толще наружных ветвей брюшных ножек, а кислород, растворенный в смачивающей жабры воде, используется внутренними ветвями.
Последняя пара брюшных ножек — уроподы — имеет разное строение у разных подотрядов равноногих. Так, например, у Asellota и мокриц они прикреплены к заднему краю плеотельсона и направлены назад, у Valvifera они с боков подвернуты под брюшную сторону плеотельсона и, как крышки, прикрывают передние нежные дыхательные брюшные ножки (рис. 244, 1,2), у Anthuridea и Flabellifera они сочленены с основанием плеотельсона и вместе с ним образуют хвостовой веер.
У паразитических Epicaridea, в особенности у их самок, организация всего тела обыкновенно очень упрощена. Они нередко теряют сегментацию и конечности и превращаются в бесформенный мешок, набитый яйцами. Однако самцы и личинки полностью сохраняют признаки равноногих ракообразных.
Основным способом передвижения всех равноногих ракообразных, за исключением паразитических, следует считать ползание по дну, почве или растениям при помощи грудных конечностей. Большинство видов двигается только таким способом. Среди морских видов многие, кроме того, могут плавать. Обыкновенно они проплывают небольшие расстояния над дном, с тем чтобы потом снова перейти к ползанию. При этом представители некоторых семейств Asellota гребут специально приспособленными к плаванию задними грудными ножками, a Anthuridea, Cymothoidae и Serolidae — брюшными ножками. Idoteidae орудуют при плавании и задними грудными и брюшными ножками.
Некоторые пресноводные равноногие также могут глубоко зарываться в грунт и таким образом спасаться от пересыхания водоемов. Так поступает, например, обычный американский водяной ослик Asellus communis, а также представители подотряда Phreatoicidea, обитающие в болотах Южной Африки. В засушливыз периоды Phreatoicidea вырывают во влажном мхе ямки глубиной 2,5—5 см и массами сидят в них, прикрывая задними грудными ножками жабры и ожидая появления воды.
Питание растениями присуще очень многим равноногим, как морским, так и пресноводным и сухопутным. Виды рода Idotea питаются водорослями, на которых они живут, а также обрастающими их диатомовыми. Преимущественно водорослевыми и бактериальными обрастаниями питаются литоральные Jaera, Sphaeroma, Eurydice, Ligia и некоторые другие. Интересно, что изменчивая окраска широко распространенной Idotea viridis, обычной, в частности, у нас в Балтийском и Черном морях, зависит от ее пищи. Живущие на зостере и зеленых водорослях рачки — зеленые, на бурых — коричневые, на красных — красные. Пигмент растений окрашивает не только содержимое их кишечника, но также кровь и полостную жидкость, просвечивающие через покровы тела. Если рачков пересадить с одних растений на другие, их окраска через 10—15 дней соответственно изменится. Поедая растительную пищу, Idotea не брезгает и попадающимися ей среди обрастаний животными — гидроидами, мшанками, личинками хирономид и т. д.
Обычные обитатели пресных вод — водяные ослики — также типичные растительноядные организмы. Помимо водных растений, живых и отмирающих, они в большом количестве поедают листья деревьев, падающие в водоемы. За всю свою жизнь каждый водяной ослик потребляет 168 мг листьев (в сыром весе). В водоемах, в которые попадает много древесных листьев, водяные ослики обыкновенно живут в большом количестве. В пруду на территории парка Тимирязевской академии насчитывается 5000 рачков на 1 м2.
Африканские и австралийские Phreatoicidea едят растительные остатки и водяные мхи.
К питанию зелеными частями растений и их остатками близко использование в пищу древесины. Среди равноногих известны древоточцы, вредящие деревянным сооружениям в море и питающиеся древесиной. Это, во-первых, многочисленные виды рода Limnoria и, во-вторых, некоторые виды рода Sphaeroma. Виды Limnoria встречаются почти во всех морях. Только в наших дальневосточных и северных морях известно 4 вида, в Черном море — 2 вида и т. д. Некоторые из них живут на водорослях и питаются ими, но большинство поселяется на сваях пристаней, на стоящих у причалов деревянных судах и т. д. Острыми жвалами они жуют древесину, причем разрушают преимущественно более мягкие летние слои, не трогая зимних. Под поверхностью сваи на глубине 1—2 см они выгрызают ходы, превосходящие по диаметру ширину их тела, следуя по летним слоям нарастания дерева. Каждый такой ход одним или двумя отверстиями соединяется с наружной средой. В нем обычно обитает самка (длиной до 5 мм) и более мелкий самец, но точит дерево главным образом самка. При сильном повреждении на 1 см2 поверхности древесины насчитывается 12—15 отверстий, ведущих в ходы рачков. Источенная рачками поверхность сваи становится рыхлой, губчатой, иногда сваи истончаются и теряют первоначальную прочность. Их приходится заменять новыми.
Рачки заглатывают опилки, образующиеся при бурении древесины, и пропускают их через свой кишечник. При этом клетчатка переваривается, а лигнин выделяется с испражнениями. При прохождении через кишечник рачка древесные опилки теряют 50—95% своего исходного сухого веса.
В проточенных в дереве ходах происходит и размножение Limnoria. Летом каждая самка приносит 8—10, изредка до 30, осенью 1—4 молодых рачка. Вылезая из ходов, рачки при помощи брюшных ножек довольно быстро плавают спинной стороной вниз, но не удаляются дальше чем на 1 м от места своего рождения. Расселение лимнорий на далекие расстояния происходит пассивно, главным образом путем переноса течениями зараженных рачками бревен, стволов, кусков дерева и т. д.

Заглатывание грунта и использование содержащегося в нем органического вещества распространено среди морских равноногих не менее широко, чем питание растениями.
Немало среди морских равноногих и хищников. Mesidotea при помощи чувствительных щетинок и цилиндров своих задних антенн разыскивает зарывшихся в грунт бокоплавов, полихет, личинок хирономид и других мелких животных, хватает их подклешнями передних грудных ножек и поедает. Нередко она нападает на рыб, в особенности попавших в рыболовные сети, и местами, например в низовьях наших северных рек, приносит заметный ущерб промыслу, так как портит улов. По-видимому, она может иногда распространяться вверх по рекам, прицепившись к мигрирующим рыбам. Astacilla охотится на различных мелких животных следующим образом: она держится задними грудными ножками за водоросли, ветви гидроидов или мшанок и приподнимает вытянутую переднюю часть тела так, чтобы длинные, богатые крепкими щетинками передние грудные ножки и чувствительные задние антенны были направлены вперед; в таком положении она легко хватает передними ножками свои жертвы, прежде всего разных мелких рачков (рис. 246).
Многочисленные представители семейства Cirolanidae образуют ряд постепенных переходов от хищничества к паразитизму. Виды родов Cirolana и Conilera нападают на рыб, правда преимущественно больных или попавших в сети, и на недавно слинявших десятиногих раков. Особенно охотно они поедают трупы рыб, оставляя от них буквально только кожу да кости.
У этих родов ротовые придатки жующего типа. Виды рода Aega — временные паразиты рыб. Они быстро плавают, прочно прикрепляются тремя парами крючковидных передних грудных ног к коже рыб, там, где она тонкая (жабры, плавники, жаберные крышки), прокалывают ее колюще-сосущим ротовым аппаратом и сосут кровь рыб. Однако, наевшись, они оставляют своего хозяина. Наконец, виды подсемейства Cymothoinae (Cymothoa, Meinertia, Livoneca, Anilocra и др.) постоянно живут на рыбах, причем не только на жабрах, но и во рту, а также на поверхности тела. Они плотно прикрепляются к рыбе, благодаря тому что все (иногда за исключением последней пары) их грудные ножки крючковидны и своими концами внедряются в кожу хозяина. Иногда прикрепившиеся рачки обрастают кожей рыбы, образуя на ее поверхности опухоли. Самки Cymothoinae теряют двустороннюю симметрию, и часто тело их искривляется. Они паразитируют как на морских, так и на пресноводных рыбах. В бассейне Амура на теле многих рыб можно найти вздутия, в которых помещаются попарно крупная асимметричная самка и более мелкий симметричный самец Livoneca amurensis.
Помимо Cirolanidae, на рыбах паразитируют личинки (праницы) Gnathiidea (рис. 249, 2). Они подплывают к рыбам, прикрепляются к их плавникам крючковидными передними ногами и начинают разрывать ими ткани хозяина, погружая все глубже свой колюще-сосущий ротовой аппарат, пока не доберутся до кровеносных сосудов. При этом они удерживаются в ране жвалами, как гарпунами, а сосущий хоботок составляют челюсти. Они сосут кровь чрезвычайно жадно. Средний отдел их кишечника переполняется кровью, и задняя часть груди раздувается настолько, что сегментация ее исчезает. Впоследствии кровь переходит в пищеварительные железы. Ее запасов хватает на всю жизнь рачка. После линьки личинок возникшие из них взрослые рачки больше не питаются, их ротовые придатки, за исключением жвал самца, редуцируются, а кишечник у некоторых родов частично даже зарастает.
Наиболее глубокие приспособления к паразитическому существованию свойственны подотряду Epicaridea, все представители которого паразитируют на различных ракообразных. Их организация, в особенности самок, в различной степени упрощена вплоть до полной потери сегментации и конечностей и превращения тела в бесформенный мешок. Самки представителей семейств Bopyridae, Dajidae и Entoniscidae, поселяющиеся в жаберной полости креветок и крабов, а иногда (Dajidae) и мизид, в той или иной степени сегментированы, но обычно асимметричны, ротовые придатки их превращены в сосущий хоботок, которым они сосут кровь своих хозяев. Выводковая сумка очень крупная и содержит большое количество яиц. Самцы двустороннесимметричны, гораздо мельче самок и живут на них (рис. 247, А). Виды семейства Cryptoniscidae паразитируют на самых различных ракообразных — на ракушковых, усоногих, мизидах, кумовых, других равноногих, бокоплавах, клешненосных осликах. Половозрелые самки Cryptoniscidae имеют мешковидное тело, без всяких следов сегментации и конечностей, с хоботком на переднем конце, которым они протыкают покровы хозяина и сосут его кровь. Такие самки образуются из маленьких самцов, сохраняющих основные признаки равноногих ракообразных. Таким образом, Cryptoniscidae, в отличие от других раздельнополых Epicaridea, представляют собой протерандрических гермафродитов: при созревании они становятся самцами, а затем преобразуются в самок.
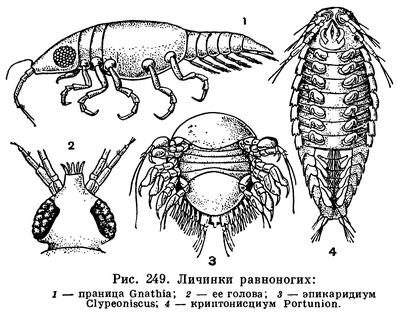
Все Epicaridea имеют свободноплавающих личинок, при помощи которых они расселяются и заражают своих хозяев.
Размножение подавляющего большинства равноногих происходит так же, как у других перакарид. Однако, помимо раздельнополых видов, среди паразитических видов, как уже упоминалось, есть протерандрические гермафродиты (Cymothoinae, Cryptoniscidae), а для Cyathura carinata описана протогиния. Наконец, среди мокриц известно несколько видов, которым свойственно партеногенетическое размножение — явление исключительно редкое у высших ракообразных.
Половой диморфизм у раздельнополых видов, если исключить Epicaridea, проявляется главным образом в строении передних брюшных ножек, приспособленных у самцов для копуляции, и нередко в различиях размеров самцов и самок: у водяных осликов, например, самцы крупнее самок, но у некоторых других видов наблюдаются обратные соотношения. У Gnathiidea самцы имеют жвалы, отсутствующие у самок (табл. 33, 1,2). У ряда Sphaeromidae у самцов две передние пары грудных ножек снабжены подклешнями, а у самок только одна.
О стадии манка Gnathiidea, носящей специальное название праница, уже говорилось выше. У Epicaridea эта стадия называется эпикаридиум. Эпикаридиум обычно достигает в длину 0,2—0,35 мм и активно плавает при помощи крупных задних антенн и брюшных ножек. Грудные ножки его крючковидные (рис. 249, 5). Он питается только за счет желтка, унаследованного от яйца. Судя по наблюдениям над Portunion (для других видов жизненный цикл не установлен), эпикаридиум прикрепляется к планктонным веслоногим и после линьки превращается в паразитическую стадию микронноциум, которая теряет членистые конечности и плавательные щетинки, но, слиняв несколько раз, дает начало новой свободноживущей стадии — криптонисциуму. Криптонисциум имеет нормальную сегментацию, 7 пар крючковидных грудных и 6 пар плавательных брюшных ножек, хорошо развитые антенны (рис. 249, 4).
Партеногенетическое размножение известно у некоторых видов мокриц из рода Trichoniscus (Trichoniscus elisabethae, Trichoniscus provisorius) и также у обычных широко распространенных мокриц Armadillidium vulgare и Cylisticus convexus. Все эти виды распадаются на отдельные расы, различающиеся по числу хромосом. Расы с обычным диплоидным числом хромосом включают и самцов и самок, и у них размножению предшествует оплодотворение яиц. Наряду с ними существуют расы с триплоидным числом хромосом (у Trichoniscus диплоидное число хромосом 16, триплоидное 24), представленные исключительно самками, к которым иногда добавляются в ничтожном количестве (менее 1,6%) самцы. Яйца триплоидных самок при созревании не испытывают второго редукционного деления и оказываются способными к развитию без оплодотворения. Такие размножающиеся партеногенетически расы приурочены к сравнительно неблагоприятным условиям существования, в частности к засушливым местам, и обычно обитают близ границ области распространения каждого данного вида.
У свободноживущих равноногих, как водных, так и сухопутных, вышедшие из яиц и находящиеся на стадии манка молодые рачки некоторое время остаются в выводковой сумке матери и линяют там два или три раза. После этого, будучи еще на той же стадии, рачки покидают выводковую сумку матери. У морских видов из семейства Arcturidae молодь перебирается из выводковой сумки матери на ее длинные задние антенны и некоторое время живет на них (рис. 250). Позднее молодые рачки время от времени линяют и в конце концов достигают половозрелости. Промежутки между линьками и число их до достижения половозрелости различны у разных видов. Так, например, у мокрицы Porcellio эмбриональное развитие длится 35 дней, первая стадия манка продолжается 21 день, из которых 16 приходится на пребывание в выводковой сумке, вторая и третья — по 23 дня, а всего до возникновения вполне сформированного, но еще неполовозрелого рачка требуется 102 дня. У морского рачка Sphaeroma rugicauda при температуре 15°С яйца в выводковой сумке развиваются 45 дней, у обычного водяного ослика под Москвой — от 14 дней в мае до 22 дней в августе. Самки водяного ослика достигают половозрелости через 50—55 дней после рождения, перелиняв за это время 13—15 раз.
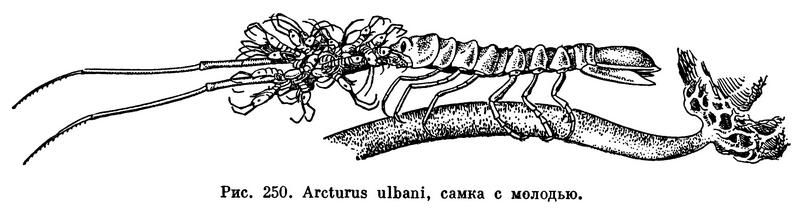
Общая продолжительность жизни установлена для немногих видов. Мокрица Oniscus asellus живет 3—4 года, Aega — не менее 2,25 лет, водяной ослик — около 2 лет, Cryptoniscidae — менее полугода.
Для многих глубоководных Asellota характерны удлиненные грудные ножки, препятствующие погружению их тела в мягкий грунт (табл. 33, 8,11,12), а также антенны, богатые чувствительными щетинками и цилиндрами. Однако особенно интересна четко выраженная у них тенденция к увеличению размеров тела по мере увеличения глубины. Чем глубже обитает какой-нибудь вид определенного рода, тем крупнее его размеры по сравнению с близким видом с меньшей глубины. Так, например, длина тела видов рода Haplomesus с глубины менее 3 км не достигает 5 мм, а Haplomesus gigas с глубины 8330—8430 м имеет в длину 15 мм.
Специальные исследования показали, что увеличение тела глубоководных равноногих происходит главным образом за счет увеличения размеров составляющих их организм клеток, а не за счет возрастания их количества. Это явление не связано с понижением температуры среды обитания. Предполагают, что в качестве основного фактора, вызывающего «глубоководный гигантизм», выступает повышенное на больших глубинах гидростатическое давление, действующее на обмен веществ.
Среди морских мелководных равноногих многие способны переносить значительные колебания солености. Jaera albifrons и Cyathura carinata (табл. 33, 7) часто встречаются в опресненных участках моря и даже заходят в пресные воды. Mesidotea entomon (табл. 33, 5) обитает преимущественно в солоноватых или даже в пресных водах, причем при большом опреснении она достигает более крупных размеров. В западной части Балтийского моря наибольшая длина самцов 50 мм, а в опресненной восточной — более 80 мм. Этот вид населяет Средний и Южный Каспий и некоторые озера, покрывавшиеся во время великого оледенения морем.
Фауна пресноводных равноногих значительно беднее морской. Наиболее широко распространено и обильно видами чисто пресноводное семейство водяных осликов (Asellidae). Обыкновенный водяной ослик (Asellus aquaticus, табл. 33, 3) обитает во множестве стоячих водоемов всей Европы, исключая Пиренейский полуостров. На Кавказе, кроме него, известен близкий вид Asellus monticola. На значительном протяжении Азии водяные ослики отсутствуют. Только в Байкале живет пять присущих лишь этому озеру видов. Кроме того, по одному эндемичному виду есть в Ципоципиканских озерах (бассейн Витима), в бассейне Иртыша и в бассейне Уссури. Местонахождения этих видов могут считаться остатками некогда более широкого распространения рода Asellus.
Морское происхождение свойственно также чрезвычайно мелким равноногим, обитающим в интерстициальных пресных и морских водах. Как и другие представители интерстициальной фауны, они имеют длинное червеобразное тело и укороченные конечности, что позволяет им активно передвигаться по узким пространствам между песчинками и другими частицами грунта. Род Microcharon (табл. 33, 13) объединяет 16 видов и 5 подвидов, большинство которых обитает в пресных грунтовых водах бассейна Средиземного моря, но один вид известен из Закавказья, а два из Средней Азии, причем Microcharon halophilus, известный из пещеры Каптархана в восточной Туркмении, живет в соленом подземном озере, соленость воды которого равняется 12‰. Кроме этих обитателей внутренних водоемов, к роду Microcharon принадлежат еще два вида из интерстициальных морских вод Ламанша. Нет особых сомнений в том, что виды этого рода переселились из системы морских в систему пресных интерстициальных вод. Сходная история расселения приписывается и интерстициальному семейству Microcerberidae, принадлежащему к подотряду Anthuridea. Только к пресным водам приурочен ныне подотряд Phreatoicidea. Относящиеся к нему 47 видов распространены в Австралии, Тасмании, Новой Зеландии, Южной Африке и Индии как в наземных, так и в подземных водах. Однако в ископаемом состоянии они известны из морских отложений пермского периода в Архангельской области, Англии и Германии. Очевидно, в конце палеозоя они жили в море и населяли северное полушарие, а до наших дней сохранились только в пресных водах некоторых стран южного полушария (рис. 252).
К жизни на суше приспособился только один подотряд равноногих — Oniscoidea, или мокрицы. Они обитают во всех ландшафтных зонах земли, но особенно многочисленны и разнообразны в степях. Наиболее примитивные из них — представители рода Ligia — еще связаны с морем. Ligia живет на морских берегах и может длительное время существовать под водой. В условиях эксперимента эти мокрицы оставались под водой более 80 дней, причем нормально питались в таких условиях. Так же тесно связаны, но с пресными водами виды рода Ligidium, часто находимые в ручьях и ключах. Для всего семейства Ligiidae, к которому принадлежат оба названных рода, характерно отсутствие приспособлений к дыханию атмосферным воздухом. Все остальные мокрицы существуют совершенно независимо от водоемов и представляют собой настоящих сухопутных животных. Однако они могут жить только в условиях повышенной влажности, обеспечивающей их нормальное дыхание и размножение. Поэтому они выработали ряд приспособлений, благодаря которым они заселили даже такие, казалось бы, не подходящие для них местообитания, как пустыни.
Практическое значение равноногих, несмотря на их широкое распространение и видовое разнообразие, довольно ограниченно. С одной стороны, они приносят некоторый вред человеку, поскольку Limnoria u Sphaeroma разрушают деревянные сооружения в море, а мокрица Hemilepistus zachvatkini объедает луговую растительность в Узбекистане. Но, с другой стороны, мокрицы, перерабатывая значительное количество растений, способствуют увеличению плодородия почвы. В особенности важную роль в этом деле играют виды рода Hemilepistus в пустынях, где они выполняют такую же огромную работу, как земляные черви в других климатических зонах.

ОТРЯД РАЗНОНОГИЕ РАКООБРАЗНЫЕ, ИЛИ БОКОПЛАВЫ (AMPHIPODA)
Бокоплавов хорошо знают не только зоологи, но и все рыбаки. В разных частях СССР местные жители называют их по-разному: «стонога» на Каспии, «мормыш» или «мормышка» на Урале и в Западной Сибири, «бармаш» на Байкале и в Восточной Сибири. На Байкале существует подледный зимний промысел омуля — «бармашенье»: бокоплавов из окрестных озер живьем привозят в бочках на Байкал, пробивают во льду проруби и горстями швыряют туда рачков, привлекая таким образом омуля, которого ловят на поддев. Широко известно кормовое значение бокоплавов для рыб. Они служат удильщикам прекрасной наживкой. Рыбоводы перевозят их вместе с мизидами во вновь созданные водохранилища для улучшения условий откорма рыб. Делались попытки искусственного разведения этих рачков на рыбоводных заводах. В естественных условиях многие рыбы используют в пищу бокоплавов, а некоторые, как например форели, питаются исключительно ими. Наряду с этим также хорошо известно, что бокоплавы в отдельных случаях портят рыболовные сети и объедают попавших в них рыб.
Бокоплавы подобно равноногим распространены очень широко. Наибольшее количество их видов обитает в море, где они населяют все глубины и живут как на дне, так и в толще воды. Только в море встречается и небогатая видами группа бокоплавов, приспособившаяся к паразитированию на китах — так называемые китовые вши (семейство Cyamidae). В пресных водах видовое разнообразие бокоплавов гораздо меньше, чем в море; по необычайному богатству видами из всех пресных водоемов выделяется замечательное озеро Байкал, где живет около 240 видов бокоплавов, причем почти все они за пределы Байкала не выходят. Довольно богата и разнообразна фауна бокоплавов подземных пресных вод. Однако, в отличие от равноногих, бокоплавы не сумели приспособиться к сухопутному существованию. Правда, есть виды бокоплавов, большую часть жизни проводящие на суше, но они живут только на морских пляжах или в других местах в непосредственной близости от моря и остаются с ним очень тесно связанными. Всего в настоящее время известно около 4500 видов бокоплавов.
По своему строению они во многом сходны с равноногими ракообразными, однако тело их часто бывает сжато с боков, а не сверху вниз, как у равноногих. Впрочем, и среди бокоплавов есть виды с уплощенным в спинно-брюшном направлении, а также с цилиндрическим телом. Голова, так же как у равноногих, срастается с первым, изредка с двумя первыми грудными сегментами, а карапакс отсутствует. Глаза сидячие и расположены по бокам головы. У пелагических Phronima каждый глаз разделен на две, а у семейства Ampeliscidae даже на 3 части (табл. 34, 12). С другой стороны, у Oedicerotidae оба глаза на спинной стороне соединяются так, что образуется один громадный непарный глаз. Глубоководные и подземные виды, как это бывает обычно, слепы, но у некоторых из них на месте глаз находятся темные, лишенные фасеток «глазные пятна», назначение которых остается неизвестным. Под покровами головы, близ ее спинной стороны, у многих бокоплавов имеется пара статоцистов, с 1—3 статолитами в каждом. Обе пары антенн обычно длинные и снабжены чувствительными цилиндрами и щетинками. Ротовые придатки жующего типа.
Конечности всех семи свободных грудных сегментов устроены по-разному, что и отражено в названии отряда «разноногие». Первые две пари ножек обычно снабжены подклешнями, в редких случаях имеются настоящие клешни или эти ножки не специализированы как хватательные. У следующих двух пар ножек когти обращены назад, а у последних трех пар — вперед. У некоторых планктонных глубоководных видов подклешни есть не только на передних, но и на следующих за ними или даже на всех парах грудных ножек. При помощи этого приспособления рачки временно прикрепляются к медузам и гребневикам, которыми питаются. У видов, ползающих по водорослям, гидроидам и т. д., когти задних грудных ножек заметно удлинены; особенно сильно они развиты на задних ногах паразитических Cyamidae, которые вонзают их: в кожу китов и таким образом прочно прикрепляются к хозяину. У этого семейства, а также у семейства «морских козочек» (Caprellidae) третья и четвертая пары грудных ног редуцированы.
У всех бокоплавов эпиподиты грудных ножек, за исключением ног первой пары, а иногда и некоторых других, превращены в листовидные тонкостенные жабры. Во многих случаях жабры складчатые, благодаря чему увеличивается их дыхательная поверхность, а иногда они снабжены пальцевидными выростами. Поскольку дыхание осуществляется придатками грудных ножек, сердце целиком помещается в грудном отделе. У половозрелых самок к некоторым из грудных ножек с внутренней стороны от жабр прикреплены пластинки выводковой сумки. В отличие от равноногих, кумовых и других выводковая сумка бокоплавов не исчезает после конца каждого периода размножения.
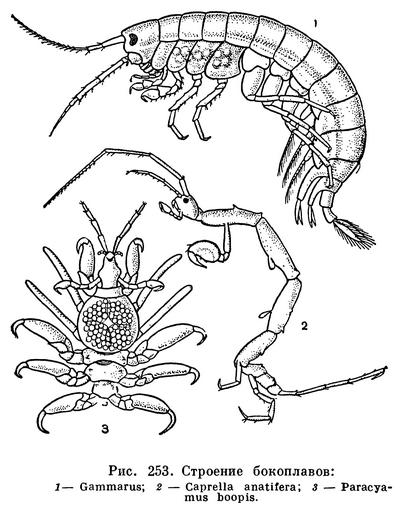
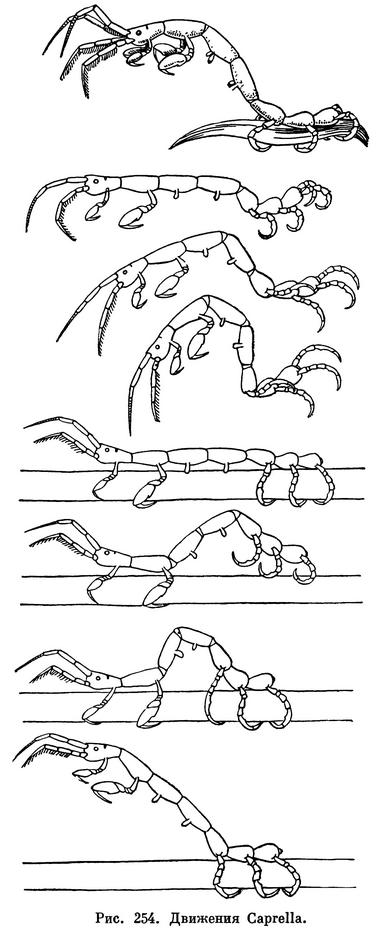
Брюшной отдел состоит из 6 сегментов. Обычно он несколько короче грудного, но имеет такую же ширину. Однако у многих планктонных бокоплавов он сужен, благодаря чему все тело приобретает каплевидную форму (рис. 255, 2). Конечности трех передних брюшных сегментов приспособлены для плавания. Их ветви многочлениковые и снабжены многочисленными плавательными щетинками. Конечности трех задних брюшных сегментов направлены назад, и ветви их не расчленены (за исключением наружных ветвей последней пары, которая часто состоит из 2 члеников). Это прыгательные ножки, или уроподы. У представителей подотряда Laemodipodea, в который объединяются морские козочки и китовые вши, брюшной отдел очень сильно укорочен и лишен сегментации, а брюшные ножки редуцированы и нередко отсутствуют совсем. У бедного видами подотряда Ingolfiellidea плавательные ножки превращены в маленькие нерасчлененные пластинки. За брюшным отделом следует короткий тельсон, имеющий форму треугольника, овала или расщепленный выемкой на две лопасти.
Покровы тела бокоплавов часто гладкие, но во многих случаях вооружены различными килями, зубцами и шипами. Такая скульптура покровов иногда имеет защитное значение. Среди многочисленных бокоплавов Байкала часть гладкие, а часть «вооруженные». В кишечниках байкальских бычков, питающихся преимущественно бокоплавами, резко преобладают гладкие виды. Очевидно, «вооруженные» в какой-то мере защищены от нападения бычков.
Бокоплавы, как правило, окрашены довольно однообразно в буроватый, зеленоватый и желтоватый тона. Исключение составляют байкальские виды, среди которых есть пестрые, синие, красные, зеленые (табл. 34, 1,4,5,7). Глубоководные и подземные виды бесцветны, но среди планктонных глубоководных видов есть и красные, как например Cyphocaris (табл. 34, 13), Paracyphocaris и ряд близких родов.
Зеленоватая окраска пресноводного Gammarus вызывается каротиноидами, вырабатываемыми из содержащихся в поедамых рачком растениях каротинов. В аквариуме представителей лишенной пигмента подземной расы Gammarus содержали в течение длительного времени в полной темноте и в условиях постоянного освещения. Независимо от этого они приобретали нормальную зеленоватую окраску, если получали в пищу растения. Однако есть и наследственные факторы, определяющие окраску. Изредка наряду с зеленоватыми встречаются красные особи бокоплавов. Эксперименты по скрещиванию их между собой и с нормальными особями показали, что окраска зависит от трех пар генов, причем ген зеленоватой окраски резко доминирует.
В соответствии с различным строением ног движения разноногих весьма разнообразны. Большинство этих рачков может ползать по дну и растениям, перебирая грудными ногами, плавать при помощи передних брюшных ножек и прыгать, отталкиваясь от субстрата задними брюшными ножками. Следует иметь в виду, что название всего отряда «бокоплавы» неточно. Только в очень мелких ручьях или у самого берега водоема рачки действительно плавают на боку, а там, где глубина это позволяет, они плавают спиной вверх, но нередко лежат на грунте на боку. Однако поскольку их легче всего заметить на самой ничтожной глубине, за ними и утвердилось заведомо неверное наименование.
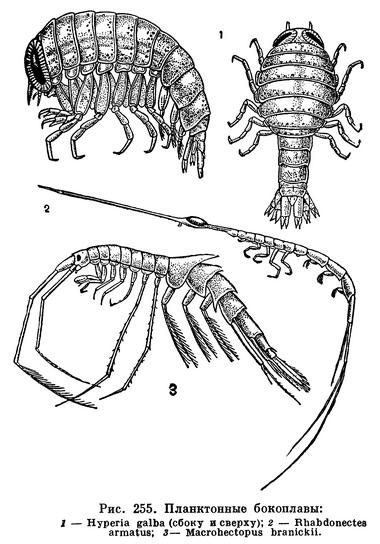
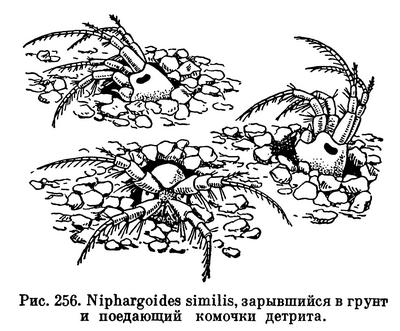
Все Corophiidae живут в построенных ими трубках. Corophium volutator цементирует секретом стенки вырываемого им туннеля длиною 4—8 см, а перед наступлением зимы углубляет туннель до 20 см. Corophium curvispinum прикрепляет свои трубки к поверхности грунта, камней, раковин моллюсков, а также к днищам судов. Благодаря своим прикрепленным к судам домикам этот каспийский вид распространился очень широко: суда развезли его по всей Волге и другим русским рекам, он проник в бассейн Балтийского моря и даже в Англию. При строительстве убежищ корофииды пользуются удлиненными задними антеннами, которыми они захватывают подходящий строительный материал (рис. 257).
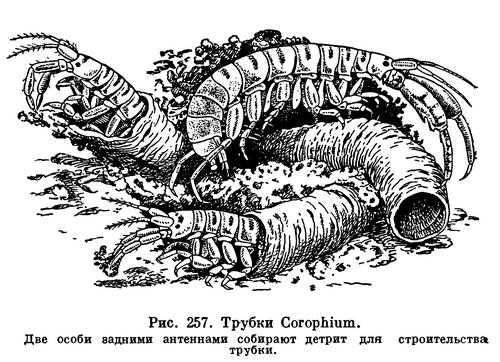
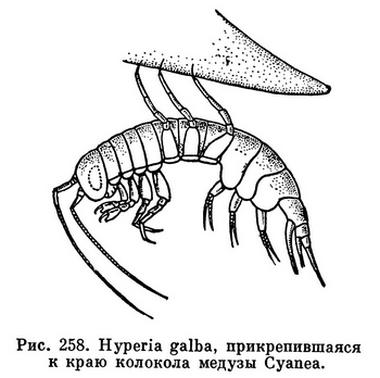
Донные бокоплавы, не зарывающиеся в грунт и не делающие нор и домиков, обыкновенно прячутся среди водорослей, зарослей гидроидов и губок или под камнями, в расселинах скал и т. д. Многие представители семейства Leucothoidae живут в каналах губок и в мантийной полости асцидий, пользуясь этими животными только как местом поселения. Морские козочки ползают по водорослям и ветвям гидроидов и, держась за них тремя парами задних грудных ножек, приподнимают остальное тело так, чтобы иметь возможность схватывать проплывающих мимо животных хватательными передними ножками. Их охотничья поза напоминает соответствующее положение тела равноногих рачков Astacilla (рис. 246).
Большинство бокоплавов можно считать всеядными животными, это значит, что они способны использовать органическое вещество в разных формах. Пресноводные и многие морские бокоплавы едят растения, как живые, так и отмершие, грунт, трупы и остатки животных, а при случае и мелких живых животных. Они откусывают жвалами куски пищи и измельчают их, причем челюсти задерживают мелкие частицы, не давая им вывалиться из сферы действия ротовых придатков. Некоторые виды могут наряду с этим добывать пищу путем фильтрации. Массовый бокоплав побережий Каспийского и Азовского морей — Niphargoides maeoticus — пассивно профильтровывает взвесь, приносимую волнами. Когда волна начинает отходить от берега, рачки сидят в грунте, высунув из него передний конец тела, при обнажении грунта они зарываются в него целиком.
Хищников среди донных бокоплавов сравнительно мало. К ним относятся морские козочки. Молодые рачки едят гидроидных полипов, а подросшие ловят передними хватательными ножками веслоногих рачков, мелких бокоплавов, червей и т. д. Среди планктонных бокоплавов хищничество преобладает. Судя по постоянному присутствию в кишечниках Hyperiidea стрекательных клеток кишечнополостных, гиперииды нападают на медуз и питаются ими. Под колоколом ушастой медузы часто живет по нескольку экземпляров Hyperia galba, которая, вероятно, находит там и стол и дом. Гиперииды прицепляются не только к медузам, но и к гребневикам. Phronima выедает тело сальп и пирозом, а затем забирается в пустую мантию отдельных особей этих животных и в ней плавает. Планктонные гаммариды также, по-видимому, питаются преимущественно кишечнополостными. В их кишечниках, так же как у гипериид, были обнаружены стрекательные клетки медуз, а строение ротовых придатков и грудных ножек с подклешнями многих планктонных Lysianassidae позволяет предполагать их полупаразитическое существование на медузах и, возможно, на гребневиках.
Настоящими паразитами среди бокоплавов следует считать только китовых вшей (Cyamidae). Огромными массами они сидят на китах, обычно близ полового и анального отверстий, и грызут их кожу, нередко вызывая глубокие язвы, доходящие до жирового слоя. В отличие от других паразитических ракообразных у китовых вшей нет никаких свободноплавающих стадий. Вероятно, они переходят с одной особи хозяина на другую во время спаривания китов, а китята заражаются при рождении или при кормлении молоком.
Все бокоплавы раздельнополы. Половой диморфизм часто бывает хорошо выражен, но по-разному у различных семейств и родов. У представителей семейства Gammaridae самцы, как правило, крупнее самок, но у представителей семейства Lysianassidae наблюдаются обратные соотношения размеров. У некоторых бокоплавов Байкала, принадлежащих к семейству Gammaridae, самцы настолько мельче самок,что их называют карликовыми. Они значительно раньше самок достигают половозрелости, после чего их рост прекращается. Так, например, длина половозрелых самцов планктонного Macrohectopus branickii не превосходит 5,5 мм, в то время как длина половозрелых самок колеблется между 14 и 30 мм. У самцов многих Gammaridae и всех Talitridae подклешни передних грудных ножек сильнее развиты, чем у самок. Нередко самцы обладают более длинными антеннами с более многочисленными чувствительными органами на них. У многих видов подземного рода Niphargus самцы резко отличаются от самок удлиненным конечным члеником наружной ветви задних уроподов, а иногда, кроме того, удлиненными ветвями одной или двух пар передних уроподов. Половозрелые самки всегда имеют выводковую сумку.
Спаривание продолжается обычно несколько дней. Самец располагается на спинной стороне самки, удерживаясь своими подклешнями за передний край ее первого и за задний край ее пятого свободного грудного сегмента и ожидая ее линьки. После линьки самки самец передвигается под ее брюшную сторону, складывает вместе свои передние брюшные ножки, несколько раз просовывает их между задними пластинками ее выводковой сумки и в это же время выделяет из своих половых отверстий сперму. При помощи передних брюшных ножек сперма переносится в выводковую сумку, куда через 1,5—4 часа (у Gammarus) откладываются яйца, которые здесь же оплодотворяются.
Нормальная откладка яиц может происходить только тогда, когда в выводковой сумке есть сперма. В опытах с Gammarus duebeni удавалось закупоривать половые отверстия самцов. После спаривания с такими самцами, происходившего нормально, если не считать того, что они не выделяли сперму, половина самок вообще не отложила яиц, а остальные отложили их не полностью, в малом количестве.
Наиболее богата и разнообразна фауна бокоплавов материкового склона. В Баренцевом море здесь обитает около 260 видов, в Японском — 250 видов. Некоторые из видов бокоплавов материкового склона встречаются в огромном количестве. В Чукотском море на 1 м2 дна приходится до 24 тысяч экземпляров Pontoporeia и до 14 тысяч экземпляров Lembos. Трал в этом море приносил такую массу бокоплавов, что, высыпанные на палубу, они образовывали копошащиеся кучи высотой до полуметра.
С глубиной видовое разнообразие и количество бокоплавов уменьшаются, однако даже с огромных океанических глубин, более 6000 м, в настоящее время известно около 300 видов. Большая часть их относится к широко распространенным родам, встречающимся и на меньших глубинах, но среди них есть и очень своеобразные представители. Так, например, в Курило-Камчатской впадине живет пелагический бокоплав Vitjaziana gurjanovae, выделенный в особое семейство и не поднимающийся на глубины менее 6000 м.
Пресные воды населяет сравнительно небольшое количество видов бокоплавов. В северном полушарии чрезвычайно широко распространен озерный бокоплав Gammarus lacustris, обитающий в самых различных озерах нередко в громадном количестве. Он может существовать как в пресных, так и в сильно минерализованных водоемах и переносить различные неблагоприятные условия, в том числе зимнее уменьшение содержания кислорода в воде. При наступлении зимнего замора массы рачков скапливаются под нижней поверхностью льда. В Сибири бокоплавов добывают, пробивая во льду проруби и разными способами облавливая нижнюю его поверхность. В текучих водах обитают другие виды того же рода — Gammarus pulex, Gammarus balcanicus и т. д.
Практическое значение бокоплавов, как уже указывалось, очень велико и определяется использованием их в пищу многими рыбами, в том числе и промысловыми. Так, например, в Каспии и Азовском море они составляют значительную часть пищи леща, молоди осетровых, на Дальнем Востоке — многих камбал, в устьях северных рек — муксуна, омуля, ряпушки, в пресных озерах — различных сигов, форелей и т. д. Для улучшения условий откорма ценных рыб бокоплавов перевезли во многие вновь созданные водохранилища и в озера, где их раньше не было.


НАДОТРЯД ЭВКАРИДЫ (EUCARIDA)
У представителей надотряда эвкарид всегда имеется карапакс, он покрывает все головные и грудные сегменты и срастается со спинной стороной последних. Глаза стебельчатые, жвалы симметричные, без подвижной пластинки. Самки носят яйца прикрепленными к брюшным ножкам или реже откладывают их прямо в воду. Из яиц обычно выходят личинки, непохожие на взрослых, у немногих видов развитие прямое. Надотряд объединяет два отряда — эвфаузиевых и десятиногих ракообразных.
ОТРЯД ЭВФАУЗИЕВЫЕ РАКООБРАЗНЫЕ (EUPHAUSIACEA)
Каждый год наши китобойные флотилии промышляют китов в Антарктике. Туда же направляются китобои из многих других стран. Хорошо известно, что наибольшее количество усатых китов сосредоточено в антарктических морях. Киты находят там огромные запасы своей основной пищи — скопления «криля», т. е. эвфаузиевых рачков Euphausia superba. Эти скопления настолько велики, что в настоящее время делаются попытки непосредственно использовать рачков для изготовления из них пищевых продуктов для людей, кормовой муки, жира, витаминов и т. д.
Эвфаузиевые рачки живут не только в Антарктике. Они широко распространены во всем Мировом океане, избегая лишь его опресненных районов. Повсюду, в частности в наших северных и дальневосточных морях, они служат важным объектом питания рыб и китообразных. Всего известно 85 видов эвфаузиид.
Все эвфаузиевые — планктонные животные. Тело их прозрачное, у некоторых глубоководных видов красное. Его длина колеблется от 7 до 96 мм. По внешнему виду эвфаузииды напоминают небольших креветок, однако отличаются от них многими признаками. Боковые края карапакса эвфаузиид короткие и оставляют непокрытыми кустистые жабры, прикрепленные к основаниям грудных ног. Ни одна из 8 пар грудных ног не превращена в ногочелюсти. Задние грудные ножки укорочены. На заднем крае тельсона, по бокам от его заостренного конца, располагается пара пластинок, отсутствующая у десятиногих ракообразных. Чрезвычайно характерны для эвфаузиид органы свечения — фотофоры. Обычно их бывает 10: одна пара на глазных стебельках, по одному на основных члениках второй и седьмой пар грудных ножек и по одному на нижней поверхности каждого из 4 передних брюшных сегментов. Они испускают желтый или желто-зеленый свет, причем светятся как при раздражении, так и сами по себе, однако не беспрерывно, то вспыхивая, то затухая. Каждая вспышка продолжается 3—6 секунд, после чего фотофоры медленно гаснут. Иногда вспыхивают только наиболее яркие фотофоры глазных стебельков, иногда все вместе. Когда фотофоры не действуют, рачки в темноте становятся невидимыми благодаря прозрачности своего тела. Фотофоры помогают рачкам собираться в стаи, а самцам находить самок. Только у одного вида — глубоководной Bentheuphausia amblyops — фотофоры отсутствуют, но у нее, в отличие от других эвфаузиид, редуцированы и глаза. У остальных эвфаузиид, наоборот, глаза хорошо развиты. Они стебельчатые и очень крупные: нередко их длина равняется 1/6—1/8 длины тела рачка. У многих хищных видов каждый глаз подразделен на две части: одна, более крупная, направлена вперед, а другая — вверх. Такое строение глаз, по всей вероятности, увеличивает кругозор рачка и помогает ему охотиться на движущихся животных. Особенно эффективными оказываются двураздельные глаза при слабом освещении. Способность видеть при слабом освещении достигается еще высокой концентрацией в теле эвфаузиид витамина А, который на 90—95% сосредоточен в их глазах, а также астаксантина. Эти вещества участвуют в образовании светочувствительных соединений, в частности определяющих возможность сумеречного зрения. По содержанию в теле витамина А и астаксантина эвфаузииды превосходят остальных ракообразных.
У видов с двойными глазами есть еще одно приспособление, свидетельствующее об их хищных повадках: грудные ноги второй или третьей пары, а иногда и те и другие у них очень сильно удлинены и нередко на одной из этих пар развиты клешни. Это так называемые ловчие или разбойничьи ноги, которыми рачки хватают свои жертвы (рис. 260, 4).
Эвфаузииды — прекрасные пловцы. На пяти передних брюшных сегментах у них расположено по паре сильных двуветвистых плавательных ножек — плеоподов. Эти ножки уплощены по направлению спереди назад, а их ветви снабжены по краям перистыми щетинками. На основаниях ножек имеются направленные внутрь крючковидные шипы, при помощи которых левая и правая ножки каждой пары могут сцепляться и действовать как единое целое. Своими плавательными ножками эвфаузииды гребут. При этом Meganyctiphanes norvegica передвигается со средней скоростью 271 м в час. Но, кроме того, резко сгибая и распрямляя брюшко и отталкиваясь тельсоном и пластинчатыми уроподами от воды, они совершают молниеносные прыжки. Судя по наблюдениям в аквариуме, разные виды в толще воды ведут себя различно. Растительноядная Euphausia беспрерывно двигается во всех направлениях, а хищные Stylocheiron и Nematoscelis в течение длительного времени неподвижно парят в воде, держа тело в вертикальном положении и расправив свои длинные ловчие ноги в ожидании добычи.
Подвижность эвфаузиид позволяет многим из них совершать регулярные вертикальные миграции. Мелководные виды ночью поднимаются к поверхности и там питаются, а днем опускаются в более глубокие слои. Однако, по-видимому, не все особи каждые сутки проделывают такое путешествие; значительная часть остается на обычной глубине их обитания, так как отдельные особи поднимаются в поверхностные слои через сутки или даже через двое суток, а самые крупные мигрируют только для размножения. В Баренцевом море суточные миграции наблюдаются только весной и осенью.
Кроме того, в некоторых морях происходят сезонные миграции эвфаузиид. Так, например, в Баренцевом море рачки зимой опускаются и образуют на мелководьях и банках мощные придонные скопления. Весной половозрелые рачки для размножения поднимаются в поверхностные слои воды. Это происходит начиная с середины апреля и совпадает с весенним развитием водорослей, которые служат пищей личинкам эвфаузиид. Подъем начинается под воздействием усиливающегося освещения.
Поднимающиеся для размножения эвфаузииды образуют стаи, хорошо видимые с борта корабля. Э. А. Зеликман следующим образом описывает «роение» Thysanoessa raschii в Баренцевом море: «Рачки концентрировались на площади, приблизительно равной 9 кв. милям. Эвфаузииды плавали на этом участке как рассеянно, поодиночке, так и стайками. Стайки были двух типов — шаровидные, от 0,5 до 2 м диаметром, и лентовидные, длиною около 5 м и шириной около 1 м. Основная масса рачков придерживалась верхнего двухметрового слоя. С борта судна стайки ярко-розовых рачков в воде выглядели красновато-рыжими пятнами. Кое-где под стайками эвфаузиид шла мойва, которая питалась рачками и преследовала их; рачки уходили от рыбы в поверхностный слой, и многие выпрыгивали из воды. Эвфаузиид пожирали и многочисленные чайки, главным образом моевки. За два часа наблюдений с корабля каждая из стаек рачков сохранялась как целое».
Массовый подъем антарктической Euphausia superba также связан с размножением. Огромные скопления половозрелых рачков приурочены к зоне так называемой антарктической дивергенции, где происходит поднятие к поверхности глубинных вод, богатых солями фосфора и азота. Эти соли дают возможность развиться огромному количеству водорослей, используемых в пищу рачками. Самки Euphausia superba выметывают яйца на прибрежных мелководьях. Затем яйца погружаются на глубины 1800—3000 м, где из них выходят личинки. По мере развития личинки постепенно поднимаются в поверхностные слои. Все эти вертикальные перемещения приводят к тому, что разные личиночные стадии переносятся то поверхностными, то глубинными течениями в различных направлениях, однако в результате все рачки остаются в пределах обычной и наиболее благоприятной для них области распространения, окружающей довольно узким кольцом берега Антарктиды.
Поднимающиеся к поверхности воды самки всех эвфаузиид уже оплодотворены. Процесс оплодотворения у них происходит еще во время пребывания у дна. Непосредственно наблюдать его не удавалось, но несомненно, что при этом важную роль играет сложно устроенный придаток на передних брюшных ножках самца, так называемая петазма. У самок на брюшной поверхности шестого грудного сегмента имеется карманообразный орган — теликум, в который открываются половые отверстия. При помощи петазмы самец прикрепляет два образующихся в его половых путях сперматофора к теликуму самки. Иногда наблюдались самки не с двумя, а с четырьмя и даже с шестью сперматофорами, оплодотворенные, очевидно, дважды и трижды.
Одни эвфаузииды (Euphausia, Thysanoessa, Meganyctiphanes и др.) выметывают оплодотворенные яйца прямо в воду, а другие носят их на себе в виде яйцевых мешков, прикрепленных к задним грудным ногам. У Nyctiphanes образуется два удлиненных грушевидных мешка с 60 яйцами в каждом, у Nematoscelis и Stylocheiron — один мешок, содержащий у первого рода 36—60, а у второго — всего 18 яиц.
Плодовитость эвфаузиид, откладывающих яйца прямо в воду, значительно больше. Каждая самка антарктической Euphausia superba длиною 33—45 мм выметывает в зависимости от размера от 200 до 3600, а по некоторым данным до 11 000 яиц, Thysanoessa inermis, Thysanoessa raschii, Thysanoessa longicaudata и Euphausia pacifica в наших северных и дальневосточных морях — от 1000 до 2000 яиц.
Из яиц эвфаузиид выходит типичный науплиус. Он линяет, оставаясь на той же стадии, а затем, после второй линьки, переходит в стадию метанауплиуса. Через 2—3 дня (у Thysanoessa Баренцева моря) метанауплиус превращается в присущую только эвфаузиидам стадию — калиптопис, у которой брюшко удлинено и четко отграничено от грудного отдела, а глаза находятся под карапаксом. Калиптопис имеет сквозной кишечник и начинает питаться водорослями. Калиптопис трижды линяет; после третьей линьки возникает последняя стадия — фурцилия, характеризующаяся постепенным развитием всех грудных и брюшных ножек и освобождением из-под карапакса стебельчатых глаз. Фурцилия линяет несколько раз и постепенно переходит во вполне сформированного рачка. Число ее линек у разных видов, по-видимому, различно. Фурцилии старших возрастов, задние антенны которых уже не служат для плавания, иногда выделяются в особую стадию — циртопию. У некоторых эвфаузиид, в особенности у носящих яйца на себе и населяющих прибрежные воды, число личиночных стадий уменьшается и метаморфоз оказывается сокращенным.
Яйца, науплиусы и метанауплиусы мелководных эвфаузиид наших северных и дальневосточных морей находятся в поверхностных слоях воды. Калиптопис и фурцилии опускаются несколько глубже, а взрослые особи начинают совершать вертикальные миграции.
Через год после рождения эвфаузииды наших морей, а также некоторые другие, в частности Euphausia superba, достигают половозрелости и приступают к размножению.
На следующий год эвфаузииды размножаются вторично, после чего погибают, в значительной своей части становясь добычей рыб. Небольшая часть рачков достигает трехлетнего возраста и размножается еще раз. Крупные глубоководные
виды, несомненно, долговечнее, но их жизненный цикл еще не изучен.
Пища эвфаузиид разнообразна. Некоторые виды питаются главным образом планктонными водорослями, отфильтровывая их из воды. Euphausia superba поедает преимущественно диатомовые водоросли, диаметр которых не превосходит 0,04 мм. Грудные ножки рачка с внутренней стороны снабжены длинными, густыми перистыми щетинками, образующими фильтрующий аппарат. Если в эту ловчую сеть попадают более крупные частицы, рачок удаляет их щетинками последних члеников грудных ног. Большинство других эвфаузиид питается сходным образом, но наряду с водорослями потребляет и планктонных животных — веслоногих и ракушковых ракообразных, глобигерин, тинтинноидей и т. д. Молодые Meganyctiphanes norvegica длиной до 19 мм едят детрит и диатомовые водоросли, рачки длиной 21—29 мм — преимущественно детрит, а крупные особи, длиной 31—39 мм, — планктонных ракообразных. Глубоководные виды рода Thysanopoda — хищники, нападающие исключительно на животных, в том числе и на довольно крупных (щетинкочелюстные, медузы). Среди мелководных видов хищниками могут считаться виды Nematoscelis, Tessarobrachion, Thysanoessa longipes и др. Своими ловчими ногами они хватают жертву. Эти ноги обычно вооружены крупными шипами, которые прокалывают пойманное животное. Глубоководная Bentheuphausia amblyops, помимо животных, в большом количестве потребляет водоросли, для чего время от времени поднимается в поверхностные слои воды.
Эвфаузииды служат основной пищей многим морским млекопитающим и рыбам. Усатые киты питаются главным образом этими рачками. Как уже указывалось, скопления китов в Антарктике связаны с зонами массового развития Euphausia superba. Этот вид распространен на глубинах до 40—50 м вокруг берегов Антарктиды; зимой и весной он держится среди льдов, летом и осенью — в открытой воде, образуя стаи самых разных размеров — площадью от нескольких до 2000 м2. Каждая стая состоит из одноразмерных рачков. Euphausia superba составляет основную, а часто и единственную пищу финвала, синего кита, горбача и сейвала. В течение года они выедают 38 миллионов тонн рачков. Однако количество Euphausia superba остается значительным. На одну квадратную морскую милю приходится около 14 миллионов рачков, что достаточно для наполнения желудков более чем 50 китов.
В северной части Тихого океана желудки китов также бывают обычно набиты эвфаузиидами. Вес содержимого желудка финвала, наполненного одними эвфаузиидами, достигает 360 кг. Для Северной Атлантики доказана зависимость присутствия китов от распространения скоплений эвфаузиид. Столь же охотно поедают этих рачков некоторые ластоногие, например акиба, миграции которой связаны с районами концентрации эвфаузиид, а также многие птицы — пингвины, чайки и др.
В огромном количестве поедают эвфаузиид в Баренцевом море сельдь (до 60% по весу от всей пищи), морской окунь (50%), треска, молодь многих других рыб; в дальневосточных морях — скумбрия, сардина, сельдь, лососевые, минтай, молодь трески и т. д. Места обитания многих из этих рыб в период их откорма приурочены к районам концентрации эвфаузиид. Интенсивное выедание рыбами рачков может уменьшить их численность в море. После резкого сокращения в 1957 и 1958 гг. запасов одного из основных потребителей эвфаузиид — трески — численность этих рачков в южной части Баренцева моря увеличилась в 4 раза.

ОТРЯД ДЕСЯТИНОГИЕ РАКООБРАЗНЫЕ (DECAPODA)
Из всех ракообразных десятиногие издавна пользуются наиболее широкой известностью. Популярный представитель этого отряда — речной рак — неизменный герой бесчисленных сказок и басен. Он фигурирует в числе знаков Зодиака, а с крабами связаны многие мифы инков, ацтеков и древних египтян. К десятиногим принадлежат крупнейшие из ракообразных: длина омаров иногда превосходит 80 см, расстояние между когтями вытянутых в стороны средних ног японского краба Macrocheira составляет 3 м.
Благодаря своим крупным размерам и отличным вкусовым качествам некоторые десятиногие ракообразные служат важным объектом промысла. В 1962 году во всем мире было выловлено около 1 миллиона тонн ракообразных — креветок, крабов, омаров, лангустов и т. д., т. е. приблизительно в 2 раза больше, чем всех лососевых. В СССР промысел камчатского краба производится огромными судами — плавучими консервными заводами, на которых работает несколько тысяч человек, и существует специальная крабоконсервная промышленность.
Десятиногие ракообразные распространены чрезвычайно широко. Они населяют все океаны и моря, от уреза воды до глубины около 5 км. Особенно богата видами десятиногих тропическая мелководная фауна. В пресных водах обитают общеизвестные речные раки, а в тропических и субтропических странах — крабы и креветки. Наконец, в тропиках обычны сухопутные десятиногие — крабы и сильно видоизмененные раки-отшельники.

По образу жизни и по внешнему виду десятиногие весьма разнообразны. К этому отряду принадлежат плавающие креветки с длинным мускулистым брюшком, ползающие по дну крабы, брюшко которых подвернуто под грудь, раки-отшельники, прячущие свое спирально извитое брюшко в пустые раковины брюхоногих моллюсков, также ползающие по дну речные раки, омары и лангусты, обладающие подобно креветкам длинным симметричным брюшком, и некоторые другие. Общее число видов десятиногих достигает 8500; их приблизительно столько же, сколько видов птиц.
Что же заставляет объединять столь различных животных в один отряд? Всем им свойственны некоторые общие признаки, свидетельствующие об их тесном родстве и едином происхождении.
У всех десятиногих ракообразных три передних грудных сегмента срастаются с головой, а их конечности превращены в ногочелюсти. Пять пар задних грудных ног, принадлежащих к свободным грудным сегментам, служат для передвижения, почему весь отряд и получил название десятиногих. Часто на некоторых из этих ног есть клешни или иногда подклешни. Жабры целиком покрыты боковыми краями карапакса и снаружи не видны. Между боковыми краями карапакса и стенкой тела расположено с каждой стороны по жаберной полости. Самки вынашивают яйца, прикрепляя их к брюшным конечностям, и только самые примитивные креветки просто откладывают яйца в воду. Из яиц выходят планктонные личинки, совсем непохожие на взрослых, у многих пресноводных и глубоководных видов развитие прямое.
Голова и грудь десятиногих покрыты карапаксом, плоским и коротким у крабов и цилиндрическим, удлиненным у остальных представителей отряда. Передний конец карапакса часто вытянут в шиповидный, острый на конце рострум. Поверхность карапакса иногда снабжена килями, буграми или шипами, имеющими защитное значение. По бокам от основания рострума прикрепляются стебельчатые глаза. Они могут поворачиваться в разные стороны и обеспечивают широкий кругозор.
Каждый глаз состоит обычно из большого (у креветки Palaemon serratus — 3020) количества фасеток, число которых увеличивается с возрастом. Фасетки отделены друг от друга пигментными клетками, и каждая фасетка воспринимает лишь лучи, падающие перпендикулярно к ее роговице. Она видит только небольшую часть объекта, на который смотрит рак, а другие фасетки видят иные части этого объекта. Таким образом осуществляется «мозаичное зрение». Ночью пигмент расходится к концу и к основанию глаза, и косые лучи могут достигать сетчатки — рак видит объект целиком, но довольно туманно. В воде десятиногие видят только близко расположенные предметы. Рак Galathea на расстоянии 1,5 м не замечает даже крупные движущиеся объекты, но на расстоянии 40 см хорошо различает белые квадраты площадью 2 см2. Поэтому водные десятиногие в поисках пищи, самок, убежища пользуются не столько зрением, сколько обонянием, осязанием и химическим чувством. Зато в жизни сухопутных видов, обладающих особенно длинными глазными стебельками, зрение играет очень важную роль.
В глазном стебельке находится несколько органов внутренней секреции. Их выделяющиеся в кровь гормоны регулируют расположение пигмента в пигментных клетках, процесс линьки, обмен веществ, содержание в крови сахара и кальция и взаимодействуют с другими органами внутренней секреции(рис. 263).
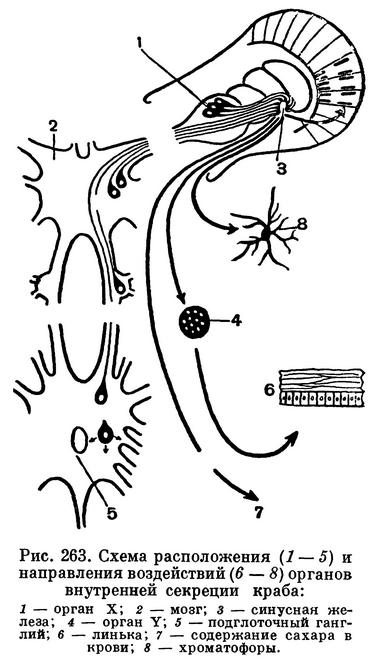
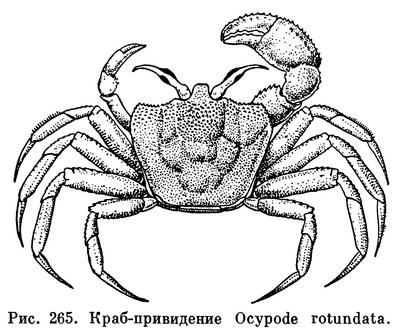
Передние и задние антенны у крабов очень короткие, а у остальных десятиногих длинные, нередко, особенно у глубоководных креветок, превосходящие по длине тело. У многих «длиннохвостых», т. е. обладающих длинным брюшком, десятиногих на каждой из передних антенн находится по два или даже по три жгута. Эти жгуты богаты чувствительными щетинками и цилиндрами, выполняющими функции органов обоняния, осязания и химического чувства. В основном членике каждой из передних антенн расположен орган равновесия — статоцист.
У большинства десятиногих полость статоциста соединяется с наружной средой узкой щелью. На стенках полости находятся чувствительные щетинки двух типов — крючковидные и нитевидные. На крючковидные щетинки опираются один или несколько статолитов. Большинство десятиногих использует в качестве статолитов песчинки, которые после линьки рак запихивает в полость статоциста, у некоторых статоцист образуется за счет затвердевания выделяемого специальными железами вещества; у многих крабов статолитов нет. Ощущая давление статолита на те или иные чувствительные щетинки, животное ориентируется по отношению к силе земного притяжения.
Жвалы десятиногих всегда мощные и служат для перетирания и разрывания пищи. Передние челюсти маленькие, а на задних имеется крупная наружная лопасть — скафогнатид, благодаря движениям которой осуществляется смена воды в жаберной полости. Она совершает от 120 до 200 взмахов в минуту, причем в жаберной полости в течение 10 минут сменяется от 2 до 25 объемов воды, что зависит от температуры воды, содержания в ней кислорода и видовой принадлежности рака.
Из 8 пар грудных конечностей 3 передние превращены в ногочелюсти. Они принимают участие в захватывании пищевых частиц и в передаче их к ротовому отверстию. Остальные 5 пар грудных ног служат главным образом для ползания и, кроме того, передние из них — для хватания пищи. На таких хватательных ногах развиваются клешни. У крабов, раков-отшельников и других близких к последним форм клешни есть только на передней паре ходильных ног, у большинства креветок — на двух передних парах, а у речных раков, омаров и креветок из семейств Penaeidae (табл. 35, 7) и Sergestidae — на трех передних парах, причем клешни первой пары значительно крупнее остальных. У креветок из семейства Crangonidae на передних ходильных ногах настоящие клешни заменены подклешнями (табл. 35, 4). У лангустов, раков-медведей и некоторых других клешней нет совсем.
Нередко левая и правая клешни первой пары имеют разное строение: одна из них крупнее или они различны по форме. Так, например, у омаров одна (то левая, то правая) более массивная клешня приспособлена для раздавливания панцирей морских ежей, раковин моллюсков, домиков морских желудей и носит название дробящей, а другая, более тонкая, режущая клешня помогает делить на куски тело рыб, креветок и т. д. Пресноводные родственники омаров — речные раки унаследовали от своих предков асимметрию клешней. Особенно резко выражена асимметрия клешней у самцов тропических полусухопутных так называемых манящих крабов, обладающих маленькой левой и колоссальной правой клешней, сгибая которую они производят своеобразные манящие движения (рис. 284). У раков-щелкунов (сем. Alpheidae) одна из двух передних клешней также значительно крупнее другой и устроена таким образом, что подвижный палец может с силой прижиматься к неподвижному, причем раздается резкий звук (рис. 278).
Задние грудные ноги в некоторых случаях также имеют какие-нибудь специальные приспособления. У крабов-плавунцов (сем. Portunidae) последние их членики уплощены, ими крабы отталкиваются от воды, когда плавают (табл. 36, 1). У раков-отшельников задняя пара грудных ног укорочена и служит для удерживания раковин брюхоногих, в которых они живут. Родственные им Lithodidae в процессе эволюции утеряли способность жить в раковине, но сохранили укороченные задние грудные ноги. Эти конечности у Lithodidae подвернуты под край карапакса и используются для очистки жабр.
С грудными конечностями связаны жабры. Они прикрепляются или к основному членику ногочелюстей и ходильных ног, или к сочленению между этим члеником и телом, или, наконец, к стенке тела над местом прикрепления той или иной грудной конечности. Часто на каждом грудном сегменте расположены жабры всех трех или двух типов. Как уже указывалось, жабры прикрыты боковым краем карапакса, ограничивающим снаружи жаберную полость.
У плавающих креветок вода свободно проникает в жаберные полости и, направляемая движениями скафогнатида задней челюсти, омывает жабры. У ползающих десятиногих нижние края карапакса плотнее прижимаются к стенке тела, основные членики ног развиты сильнее и благодаря этому жаберные полости почти полностью изолированы от внешней среды, что препятствует загрязнению жабр грунтом. Для проникновения воды в жаберные полости имеются специальные отверстия, расположенные у крабов по одному перед основанием клешней, а у раков-отшельников, речных раков, омаров и других между основаниями ходильных ног. Из жаберной полости вода выходит из-под нижнего края карапакса в его передней части.
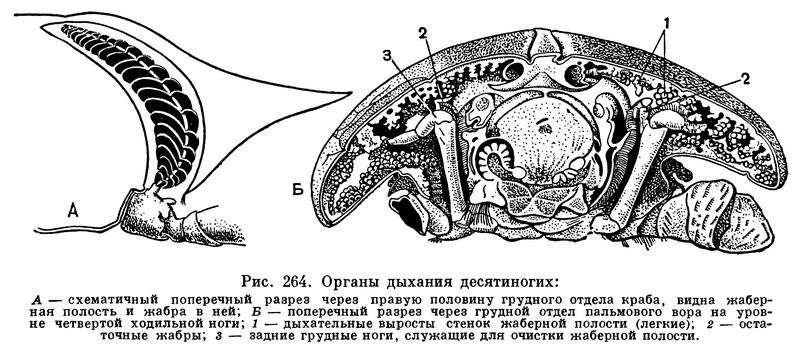
За грудным отделом тела следует брюшной, строение которого различно у разных десятиногих и тесно связано с их образом жизни. У плавающих десятиногих — креветок — брюшко сильно развито, длиннее головы и груди, часто сжато с боков и снабжено относительно крупными плавательными ножками — плеоподами. По паре плеоподов прикреплено к каждому из пяти передних брюшных сегментов, а конечности последнего брюшного сегмента — уроподы — широкие, пластинчатые и образуют вместе с тельсоном хвостовой веер. Ползающие по дну длиннохвостые ракообразные (омары, речные раки, лангусты и др.) также имеют длинное брюшко, но оно более или менее сжато в спинно-брюшном направлении, а плеоподы относительно короткие и слабые и для плавания не служат. У большинства раков-отшельников покровы брюшка мягкие, кожистые и оно спирально завернуто, так как помещается внутри пустой раковины брюхоногих моллюсков и следует за ее оборотами. Асимметрия брюшка раков-отшельников проявляется и в строении его конечностей: плеоподы развиваются только на стороне брюшка, прилегающей к наружной стенке раковины, а уропод с этой стороны гораздо крупнее, чем с противоположной. У некоторых родственных ракам-отшельникам десятиногих, например у представителей семейства Lithodidae, к которому, между прочим, принадлежит камчатский краб, а также у сухопутных пальмового вора и Coenobita брюшко укорочено и подвернуто под грудь, но и у них в расположении хитиновых щитков, покрывающих брюшко, и плеоподов заметны признаки асимметрии, свидетельствующие об их происхождении от форм со спирально закрученным брюшком. Наконец, у настоящих крабов брюшко сильно уменьшено в размерах и всегда подвернуто под грудь, число составляющих его сегментов, особенно у самцов, сокращено, плеоподы недоразвиты, а уроподы отсутствуют.
Первичная функция плеоподов плавательная, как это свойственно креветкам. Однако даже у креветок плеоподы, кроме того, участвуют в процессе размножения. У самцов креветок первая или вторая пара плеоподов частично преобразована в совокупительный орган, а самки, за исключением представителей семейств Penaeidae и Sergestidae, прикрепляют к плеоподам яйца. У ползающих десятиногих половая функция плеоподов стала основной, самки используют их только для вынашивания яиц, самцам передние плеоподы служат копулятивным органом. У самцов крабов сохраняются только первая или две первые пары плеоподов, превращенные в копулятивный аппарат.
Десятиногие — весьма подвижные животные. По способу передвижения их можно разделить на плавающих и ползающих, причем к первым принадлежат почти все креветки, а ко вторым остальные. Резкой границы между этими группами нет. Большинство креветок может не только плавать, но и ползать по дну или растениям, а среди таких типичных ползающих десятиногих, как крабы, известно обширное семейство крабов-плавунцов (Portunidae), представители которого приспособились к плаванию.
Креветки плавают при помощи плеоподов, отталкиваясь ими от воды, причем прижимают антенны и грудные ноги к телу и выпрямляют брюшко. Планктонные виды держатся в воде вертикально, работая не только плеоподами, но и наружными ветвями грудных ног. При этом они вытягивают в стороны свои длинные антенны и внутренние ветви грудных ног и таким образом препятствуют погружению. При приближении врага креветки резкими скачками уплывают задом наперед, порывисто сгибая брюшко и отталкиваясь тельсоном и уроподами от воды.
Ползание по дну осуществляется при помощи четырех пар задних грудных ног. Десятиногие сгибают и разгибают их последовательно, причем никогда левая и правая ноги одной пары не действуют одновременно. Омары, лангусты, раки-отшельники ползают передним концом вперед, но скорость их передвижения невелика. Гораздо быстрее бегают десятиногие с укороченным грудным отделом и подогнутым под него брюшком, т. е. крабы и крабовидные родичи раков-отшельников — камчатский краб (табл. 37), пальмовый вор (табл. 40), Coenobita и др. Все они передвигаются боком, достигая иногда очень большой скорости. Обычный «травяной краб» Carcinus maenas бегает со скоростью 1 м в секунду, а сухопутный краб-привидение Ocypode мчится на своих вытянутых ногах так быстро, что умудряется ловить даже мелких птиц. Крабы-плавунцы также плавают боком. При этом вторая — четвертая пары грудных ног делают 630—780 взмахов в минуту, а последняя пара работает еще более интенсивно.
Способы и объекты питания десятиногих довольно разнообразны. Большинство этих ракообразных можно считать хищниками. Морские ползающие виды находят различных животных — полихет, моллюсков, иглокожих, других ракообразных, разрывают или раздробляют добычу клешнями, перетирают ее жвалами и поедают. Многие способны расправляться с крупными и сильными жертвами. Краб Loxorhynchus grandis (с карапаксом длиной 26 см и шириной 21 см) разрывал клешнями осьминогов и больших морских звезд. Некоторые десятиногие наряду с животными поедают и богатый органическим веществом грунт, а также растения. Раки-отшельники охотно едят разных беспозвоночных, но наряду с ними и грунт, который они захватывают своей меньшей (обычно левой) клешней. Креветки Pandalus, Palaemon, Crangon и др. также едят главным образом мелких животных, но заглатывают и грунт и водоросли. Наши речные раки питаются преимущественно водными растениями — урутью, роголистником, рдестом, харовыми водорослями, но при случае охотно поедают моллюсков, личинок насекомых, а также трупы любых животных. Чрезвычайно широк ассортимент объектов питания некоторых сухопутных десятиногих. Пальмовый вор ест падающие на землю плоды различных пальм, особенно охотно маслянистые плоды панданового дерева, грунт, а также нападает на других сухопутных ракообразных, в том числе на больных особей своего вида. Мнение о питании его кокосовыми орехами неверно. Несмотря на огромную силу своих клешней, он не может разгрызть скорлупу зрелых кокосовых орехов и поедает только разбитые. Помещенный в садок с кокосовыми орехами, пальмовый вор через несколько недель погибал от голода. Неверно также мнение, что пальмовый вор забирается на кокосовые пальмы, чтобы сбросить орехи вниз. Он не может влезать высоко на деревья и совсем не способен слезать с них. Вообще никакой связи между ним и кокосовыми пальмами нет, поскольку он обитает на многих островах, на которых отсутствуют кокосовые пальмы.
Будучи в большинстве хищниками, десятиногие сами подвергаются нападению многочисленных врагов — рыб, головоногих моллюсков и других десятиногих, а сухопутные виды — птиц и грызунов. Для защиты от врагов десятиногие выработали разнообразные и совершенные приспособления. К таким приспособлениям принадлежат разобранные ранее примеры покровительственной окраски, свойственной некоторым креветкам и крабам, и угрожающие позы многих крабов. Той же цели служат и другие особенности строения и поведения десятиногих.
Многие ползающие десятиногие, как например омары, лангусты (табл. 38), крабы, в том числе обычные в Черном море Pachygrapsus и Carcinus (табл. 36, 2,4), укрываются между камнями и раковинами моллюсков, в расщелинах скал и т. д. Существуют крабы и креветки, которые, стремясь к безопасности, закапываются в грунт. Креветки Crangon, Nectocrangon и, вероятно, другие представители семейства Crangonidae, а также донные креветки из семейства Penaeidae днем зарываются в грунт и сидят в нем горизонтально, выставив наружу глаза и антенны, а ночью выходят на охоту. Так же поступают крабы-плавунцы, Galatheidae, некоторые раки-отшельники. Многие крабы, зарывшись в грунт, располагают свое тело вертикально.
Различные десятиногие строят более или менее совершенные норы, в которых проводят значительную часть жизни. Представителей семейства Callianassidae называют «раками-кротами» за то, что они вырывают глубокие ходы, часто имеющие форму латинской буквы U, а иногда и более сложную. Рачок роет грунт клешнями, выбрасывая его из норы задними грудными ногами. Стенки хода обмазываются илом или выделениями кожных желез рачка (рис. 268).
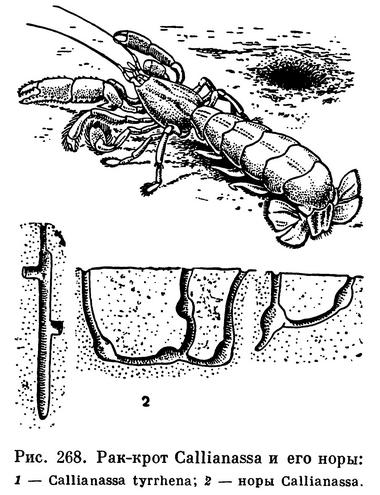
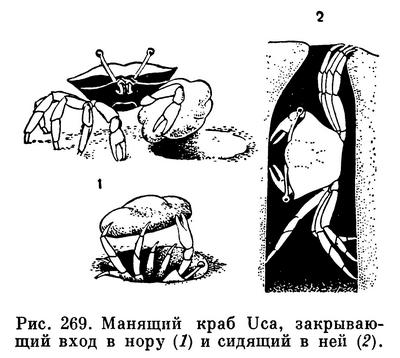
Почти все десятиногие, приспособившиеся к жизни на суше, сооружают норы, имеющие в их жизни большое значение. Обычно норы сухопутных крабов настолько глубоки, что достигают уровня грунтовых вод и их обитатели могут пить находящуюся в их домах воду. В других случаях норы расположены так, что они заливаются во время прилива. Часто норы имеют два выхода. Некоторые манящие крабы, влезая в нору, закрывают вход в нее крышкой из грунта. В каждой норе обитает один краб. Рытье нор осуществляется ходильными ножками одной стороны тела и меньшей клешненосной ногой, причем краб располагается в строящейся норе боком (рис. 269, 2).
Некоторые крабы сами нор не роют, а используют в качестве жилища норы или трубки других животных, например трубки сидячих полихет. В трубках червя Chaetopterus поселяются разные представители семейств крабов Pinnoteridae (табл. 36, 14) и крабовидных Porcellanidae.
Последние живут также в ходах, выточенных в известняке моллюском-камнеточцем Pholas.
Раки-отшельники используют пустые раковины брюхоногих моллюсков, пряча в них свое длинное, лишенное твердых покровов брюшко. Они подбирают себе раковины по размерам и плотно в них удерживаются уроподами и укороченными задними грудными ногами. На врагов рак-отшельник реагирует, забираясь целиком в раковину и закрывая ее устье своими клешнями. Раки-отшельники одного вида могут заселять раковины разных видов моллюсков.
От поселения в чужих ходах и пустых раковинах один шаг к жизни на поверхности тела и в полости тела живых животных, которые могут быть также использованы как убежища. Такой шаг многие десятиногие ракообразные сделали.
Существуют креветки, проводящие всю жизнь внутри губок. В двух губках Speciospongia vespara общим объемом 50 000 см3 было обнаружено 18 000 экземпляров двух видов раков-щелкунов Synalpheus. Креветки из рода Spongicola сидят попарно (самец и самка) в полостях стеклянных губок, куда они попадают еще будучи личинками и откуда затем, увеличившись в размерах, выбраться уже не могут.
Десятиногие ракообразные используют других животных не только в качестве убежища, но и для маскировки собственного тела, а также в качестве оружия, направляемого против своих врагов. Мелководные крабы из семейства Dorippidae при помощи загнутых на спинную сторону грудных ножек последней пары таскают над собой створку или обломок раковины какого-нибудь двустворчатого моллюска, полностью закрывая свое тело, за что и получили меткое название «стыдливых крабов». Крабы из семейства Majidae срезают своими клешнями различных обрастающих камни животных — губок, гидроидов, мшанок, колониальных асцидий — и помещают их на спинную сторону карапакса, где все эти животные продолжают расти. Этому способствуют специальные крючковидные щетинки поверхности карапакса, к которым хорошо прикрепляется живой наряд краба, а также секрет желез задних ногочелюстей, склеивающий в кучки оторванных клешнями животных.
Крабы из семейства Dromiidae покрывают свое тело губкой, выкраивая из нее кусок, совершенно подходящий для них по размеру. Обычные обитатели коралловых рифов — крабики Lybia и Polydectus всегда держат своими специально для этого приспособленными клешнями мелких актиний Bunedeopsis, которых выставляют вперед при нападении врага. Их клешни служат только для удерживания актиний, а пищу они захватывают задними ногочелюстями. На поверхности левой клешни рака-отшельника Diogenes edwardsi обычно прикреплена актиния; когда он закрывает клешней отверстие своей раковины, актиния остается снаружи и как бы охраняет вход в жилище. Другой рак-отшельник — Paguropsis typica — держит над собой защищающую его актинию Mammilifera.
Сожительство раков-отшельников с актиниями — обычное явление. Для раков-отшельников актинии служат средством защиты от врагов, актинии питаются остатками пищи своих хозяев. Поскольку как раки-отшельники, так и актинии получают от совместной жизни обоюдную пользу, это сожительство считается одним из наиболее ярких примеров симбиоза.
Раки-отшельники сожительствуют не только с актиниями, но и с представителями другой близкой группы кишечнополостных — зоантариями, играющими в этом симбиозе такую же роль, как актинии. Некоторые виды актиний и зоантарий живут только на определенных видах раков-отшельников, другие, как например Sagartia parasitica, сожительствуют с многими видами.
Помимо кишечнополостных, раковины раков-отшельников часто обрастают губки. При этом очень часто губка почти полностью растворяет раковину, и рак-отшельник оказывается окруженным губкой.
Защитное значение имеет, по-видимому, и способность некоторых десятиногих издавать звуки. В этом отношении особенно интересны раки-щелкуны (сем. Alpheidae), широко распространенные на тропических и субтропических мелководьях и встречающиеся также у нас в Черном и Японском морях. Они обычно прячутся между стволами кораллов, среди колоний мшанок, водорослей и камней, а также в губках.
Лангусты также издают громкие звуки, отпугивая ими своих врагов. На первом членике задних антенн у лангустов есть лопасть, которая скребет о килевидный выступ передней части карапакса, причем возникает трескучий звук. Калифорнийский лангуст перед стрекотаньем угрожающе машет задними антеннами, усаженными острыми шипами. Рыбы отскакивают от лангуста и, по наблюдениям Моултона, иногда теряют при этом свою добычу, которую подбирает лангуст. Известны и некоторые другие стрекочущие и скрипящие десятиногие, но биологическое значение издаваемых ими звуков остается пока неясным.
К защитным приспособлениям десятиногих принадлежит также присущая многим из них способность к автотомии — произвольному отрыванию своих конечностей. Когда враг хватает за ногу какого-нибудь рака, тот оставляет свою конечность врагу, а сам на оставшихся ногах спасается бегством. Привязанный за ногу краб отрывает ее при виде осьминога. Таким образом можно лишить краба шести ног, но, если краба ослепить, автотомии не происходит. Отрывание конечностей осуществляется всегда в строго определенном месте, недалеко от их основания, при помощи специальных мышц — автотомизаторов, которые загибают ногу вверх до тех пор, пока она не обломится. В месте отрыва внутри конечности находятся два расположенных поперечно соединительнотканных листка, помогающих быстро приостановить кровотечение. Некоторые десятиногие могут в случае необходимости отрывать себе конечности своими клешнями или задними ногочелюстями.
Способность к автотомии всегда сопровождается способностью к регенерации. Утраченная конечность постепенно отрастает вновь, тем скорее, чем чаще животное линяет, поскольку рост регенерирующей конечности происходит только после линьки. У крупных омаров полная регенерация клешней происходит только через 2 года после их потери, у самок речных раков — через 3—4 года, а у чаще линяющих самцов — через 1,5—2 года. Если при утере конечности или глазного стебелька были повреждены связанные с ними нервные центры, то на их месте может вырасти придаток ненормального строения. Так, у многих креветок взамен глазного стебелька появляется усик, напоминающий переднюю антенну. Такое явление называют гетероморфозом.
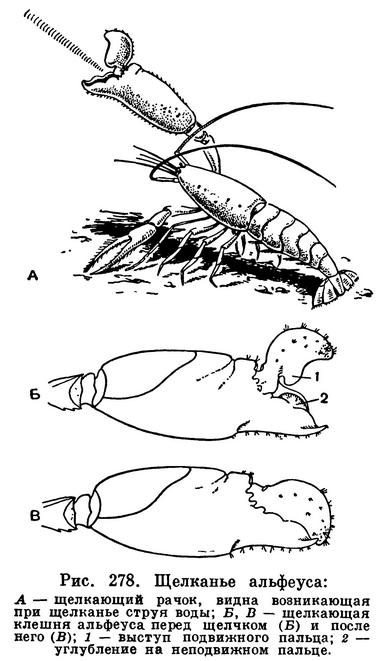
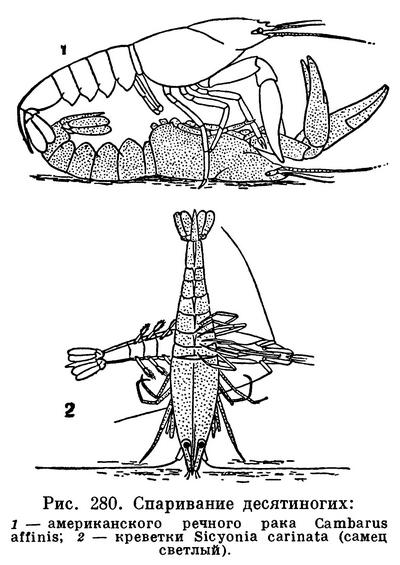
Почти все десятиногие ракообразные раздельнополы, причем самцы внешне хорошо отличаются от самок. У креветок, как правило, самцы мельче самок, а у ползающих десятиногих наоборот, и только у крабов, живущих в других животных (Hapalocarcinidae, Pinnoteridae), самцы, так же как у креветок, уступают по своим размерам самкам. Часто самцы имеют более сильно развитые клешни, помогающие удерживать самку при спаривании. Иногда, как например у краба-плавунца Neptunus, самец и самка различаются по цвету, а у травяного краба Carcinus по форме карапакса.
Особенно четко проявляется половой диморфизм в строении брюшка и его придатков. Как уже указывалось, передние брюшные ножки самцов целиком или частично преобразованы в органы, при помощи которых самец переносит сперматофоры, прикрепляя их к половым отверстиям самки, а у крабов вводя их в ее половые пути. Нередко брюшко самок бывает шире, чем у самцов (самки носят под ним икру), а у крабов оно, кроме того, часто состоит из большего числа сегментов, так как у самцов брюшные сегменты частично сливаются между собой.
У некоторых крабов наблюдается полиморфизм самцов. Так, например, среди самцов Inachus различаются три категории: мелкие крабики с толстыми клешнями, несколько более крупные, но с плоскими, как у самок, клешнями и самые крупные, опять с толстыми клешнями, причем семенники хорошо развиты только у крабов первой и третьей категории.
Гермафродитизм свойствен лишь немногим креветкам. Таковы все представители семейства Pandalidae, в том числе и промысловые виды Pandalus borealis и Pandalus latirostris. Это типичные протерандрические гермафродиты. При наступлении половозрелости они становятся самцами, а на третий год жизни превращаются в самок. Сходным образом происходит смена пола у Lysmata seticauda и Calocaris macandrae.
Самцы водных десятиногих находят самок, руководствуясь чувством обоняния и осязания. На жгутах передних антенн самцов креветок особенно много чувствительных щетинок, помогающих, вероятно, в поисках самок. Сухопутные крабы пользуются для этого зрением. Самцы издали видят самок и быстро бегут к ним.
У разных видов спаривание осуществляется по-разному. Креветки Sicyonia carinata и Athanas nitescens располагаются перпендикулярно друг к другу, причем самец оказывается лежащим на спине снизу, а самка спиной кверху над ним. У речных раков, омаров, лангустов, Galatheidea и некоторых других самка лежит на спине, а самец находится над ней спиной вверх. У многих крабов и у крабовидных отшельников самец сидит сверху, на спинной стороне самки. Молодые самцы (длиной 10 мм) Athanas nitescens в течение нескольких минут спариваются с разными самками 4—5 раз, более крупные (17—20 мм) — один раз. Самцы других креветок, китайского и камчатского крабов, речного рака Cambarus также часто спариваются с несколькими самками. Самкам некоторых крабов для оплодотворения яиц достаточно одного спаривания в несколько лет, так как сперматозоиды в семеприемниках остаются живыми. Самка американского голубого краба (Callinectes) спаривается один раз в жизни, в возрасте 3 лет, а живет 7 лет и откладывает яйца ежегодно.
Промежуток между спариванием и откладкой яиц у разных видов различен. Самки Athanas и других креветок начинают откладку яиц через несколько минут после спаривания, речные раки — через 10—45 дней, краб Carcinus — через несколько месяцев. Для откладки яиц омары, речные раки и раки-отшельники опрокидываются на спину, а креветки ложатся на бок и подгибают брюшко так, чтобы между ним и нижней поверхностью груди образовалось пространство, куда и попадают яйца. Вместе с яйцами выделяется секрет, растворяющий оболочки сперматофоров, сперматозоиды освобождаются и оплодотворяют яйца. Самки лангустов разрывают оболочку сперматофора когтем задней ходильной ноги. Секрет цементных желез, находящихся у крабов в семеприемниках, а у остальных десятиногих на нижней поверхности брюшка и на брюшных ножках, образует наружную оболочку каждого яйца, а также тяж, который прикрепляет его к брюшным ножкам матери. Крабы Carcinus при откладывании яиц зарываются в грунт, без чего не происходит прикрепления яиц. В дальнейшем яички остаются на плеоподах вплоть до выхода личинок или молодых рачков. Только у самых примитивных креветок, объединяемых в семейство Penaeidae, процесс откладывания яиц происходит иначе: они откладывают яйца прямо в воду.
Плодовитость десятиногих колеблется в широких пределах. При этом чем больше яиц, тем меньше их размеры. Очень плодовиты некоторые крабы: число яиц, выметываемых Cancer pagurus и Carcinus maenas, достигает 3 миллионов (но в Черном море у последнего не более 180 000 яиц), у Callinectes sapidus — 2 миллионов, у Eriocheir sinensis оно колеблется от 270 000 до 920 000 и т. д. Гораздо ниже плодовитость наших северных крабов (Hyas coarctatus — 1000—16000 яиц, Hyas araneus — до 110 000 яиц). Очень высока плодовитость лангустов — от 500 000 до 1,5 миллионов яиц. У раков-отшельников число яиц обычно не доходит до 10 000, но у близкого к ним камчатского краба оно значительно больше — от 70 000 до 270 000. Среди креветок наиболее плодовиты Penaeidae, откладывающие яйца в воду: Penaeus trisulcatus мечет около 800 000, a Parapenaeopsis longirostris около 400 000 яиц. Креветки, вынашивающие яйца на себе, откладывают их в меньшем количестве: Pandalus latirostris — 150—450, Pandalus borealis — 400—3000, Palaemon adspersus — 160—3600, Palaemon elegans — 280—1600, Crangon crangon — 1500—15000. Сравнительно невелика и плодовитость европейского и норвежского омаров (8000—32000 и 1300—4000 яиц).
У всех пресноводных и глубоководных десятиногих плодовитость значительно меньше, чем у морских мелководных. Наши речные раки откладывают от 60 до 500—600 яиц, тропические пресноводные крабы — 20—150, глубоководные планктонные креветки — 15—60, глубоководный (2000—2600 м) Munidopsis rostrata — 230. Из их крупных яиц выходит не личинка, а уже почти сформировавшийся рачок.
Яйца остаются прикрепленными к брюшным ножкам матери в течение продолжительного времени. Камчатский краб и европейский омар вынашивают их почти целый год, норвежский омар — 9 месяцев, краб Cancer pagurus — 7—8 месяцев, речные раки — около полугода, Pandalus borealis — 5 месяцев, Carcinus maenas — летом 2 месяца, зимой 5—6 месяцев, китайский мохнаторукий краб и креветки рода Palaemon — 1—1,5 месяца, пальмовый вор — 3—4 недели, краб-плавунец Portunus trituberculatus — 2—3 недели. Эти различия зависят в первую очередь от температуры, которая влияет на скорость эмбрионального развития.
У большинства десятиногих из яйца выходит личинка, резко отличающаяся от взрослых как по своему строению, так и по образу жизни. Только у многих пресноводных, глубоководных и обитающих при низкой температуре видов развитие прямое, т. е. все личиночные стадии проходят под покровом яйцевой оболочки и из яйца появляется маленький почти сформировавшийся рачок.
Различают несколько стадий плавающих личинок десятиногих ракообразных (табл. 39). Первая из них — науплиус — свойственна только примитивному семейству креветок Penaeidae (табл. 39, 12).
Продолжительность существования личинок в планктоне различна у разных видов: у креветок Palaemon adspersus и Crangon crangon и краба Carcinus maenas — 4—5 недель, у пресноводной креветки Atyaephyra desmaresti и у европейского омара — 2—3 недели, у креветки Pandalina brevirostris — 2 месяца, у обыкновенного лангуста Palinurus — 3 месяца, у креветки Sergestes arcticus — 4—5 месяцев, а у лангуста Panulirus — до 6 месяцев. За время своего пребывания в планктоне филлозомы лангустов разносятся течениями на далекие расстояния. С этим связывают циркумтропическое распространение некоторых лангустов.
Многие планктонные личинки десятиногих — хорошие пловцы. Когда они плавают при помощи ногочелюстей, т. е. на стадии зоэа, они двигаются задним концом вперед или, как многие зоэа крабов, спинной стороной вперед. При этом зоэа Galathea проплывает 1 м за 45—56 секунд, Porcellana — за 65—92 секунды. После превращения в последующие стадии, когда органами движения становятся плеоподы, личинки двигаются передним концом вперед. Пищей личинкам служат разные планктонные водоросли и животные.
Огромное количество личинок погибает, становясь жертвами различных хищников или попадая в неподходящие условия. По расчетам японского зоолога Марукава, из 200 000 зоэа первой стадии, которых дает одна самка камчатского краба, до второй стадии зоэа доживает 13, а до стадии глаукотоэ 7 личинок.
У большинства пресноводных, глубоководных и холодноводных десятиногих яйца крупные, богатые желтком, и всё превращение личинки проходят под покровом яйцевой оболочки. Из яйца выходит маленький рачок, который отличается от взрослых главным образом отсутствием последней пары брюшных конечностей. Новорожденные рачки удерживаются на плеоподах матери при помощи своих конечностей. У речных раков северного полушария (сем. Astacidae) для этого служат клешни, а у речных раков южного полушария (сем. Parastacidae) — задние грудные ножки. Сходным образом ведут себя вылупившиеся из яиц пресноводные крабы (сем. Potamonidae), полуглубоководная креветка Sclerocrangon ferox и глубоководная Sclerocrangon zenkevitchi.
С размножением связаны миграции многих десятиногих. Они стремятся к моменту выхода из яиц личинок попасть в условия, наиболее благоприятные для существования своих новорожденных потомков.
Некоторые десятиногие живут очень долго: американский омар до 50 лет, широкопалый речной рак до 20 лет, камчатский краб до 23 лет, лангуст более 15 лет, мохнаторукий краб до 6, иногда до 10 лет. Пальмовый вор длиной более 10 см имеет от роду не менее 5 лет.
Продолжительность жизни креветок обычно не превышает 4 лет (Crangon crangon, Pandalus), а мелкие виды не доживают и до года.
Подавляющее большинство десятиногих обитает в области морского мелководья. Особенно богата ими фауна тропиков. Коралловые рифы населены бесчисленным множеством самых разнообразных крабов, креветок и раков-отшельников, часто очень пестро окрашенных и имеющих причудливую форму. По направлению на север и на юг от тропической зоны фауна десятиногих беднеет. В пределах умеренных широт по богатству десятиногими выделяется северная часть Тихого океана, где обитает наибольшее количество родов и видов таких семейств креветок, как Crangonidae, Hippolytidae, Pandalidae, а также крабовидных отшельников Lithodidae. В Беринговом море известно 117, в Охотском 113, в Японском 82 вида десятиногих. Из северной части Тихого океана шло расселение некоторых групп десятиногих в Полярный бассейн и даже в северную часть Атлантического океана. Очень бедна десятиногими ракообразными Антарктика, откуда указано всего 8 видов.
Морские десятиногие обитают от приливо-отливной полосы до глубины 5300 м. В этом отношении они уступают многим другим отрядам ракообразных (бокоплавам, равноногим и др.), которые приспособились к жизни и на больших глубинах, вплоть до максимальных (11 км). Среди глубоководных донных десятиногих много древних родов, отсутствующих на мелководьях, но известных в ископаемом состоянии (Eryonidae, крабы Prosoponidae и Homolidae и др.). Очень характерны глубоководные планктонные креветки, принадлежащие к семействам Oplophoridae (табл. 35, 8), Penaeidae, Sergestidae и др. Обычно они темно-красного цвета, их конечности удлинены, и многие из них обладают способностью к свечению.
В пресных водах обитает сравнительно немного видов десятиногих. Среди них широко известны речные раки, принадлежащие к трем семействам — Astacidae, Parastacidae и Austrastacidae. Astacidae живут в умеренной и отчасти южной полосе северного полушария, Parastacidae — в умеренной полосе южного полушария — в Южной Америке, Австралии, а также на острове Мадагаскар, Austrastacidae — только в южной части Австралии. Почти на всем протяжении тропиков речных раков нет. В пределах СССР речные раки представлены двумя родами — Astacus и Cambaroides. Род Astacus занимает почти всю Европу и западную часть Азии, род Cambaroides — бассейн Амура, северную Японию, Корею и Сахалин.
Наиболее широко распространены и имеют довольно важное промысловое значение два наших вида речных раков — широкопалый (Astacus astacus) и узкопалый (Astacus leptodactylus). Широкопалый рак живет главным образом в водоемах, относящихся к бассейну Балтийского моря. Он обычен в Латвии, Литве, Эстонии, в Ленинградской области, в некоторых районах Украины и Белоруссии. Область распространения узкопалого рака гораздо шире и охватывает водоемы бассейнов Каспийского, Черного и Азовского морей, Каспийское и Азовское моря, а также реки и озера Западной Сибири.
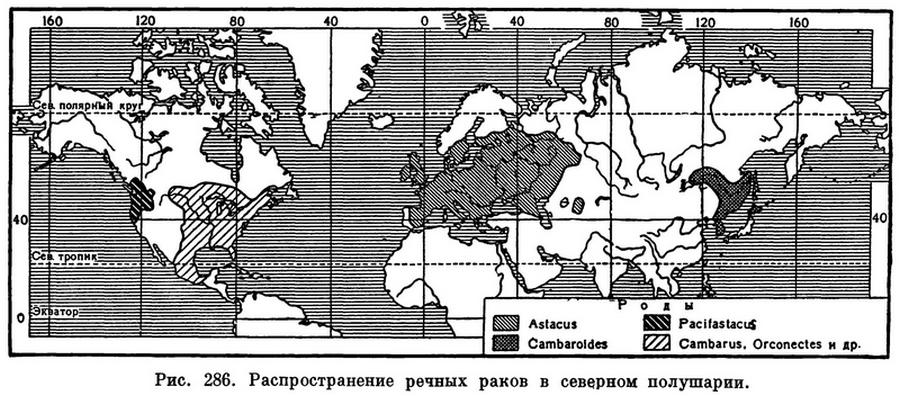
В пресных водах тропиков взамен отсутствующих там речных раков водится много различных крабов и креветок. К пресным водам целиком приурочено богатое видами семейство крабов Potamonidae, включающее около 20 родов и более 200 видов. Некоторые виды этого семейства встречаются даже в высокогорных водоемах (до 2100 м над уровнем моря в Индии). Один из видов Potamonidae — Роtamon potamios — широко распространен в бассейне Средиземного моря и обычен у нас на Кавказе, в Крыму и в Средней Азии. В сырых лесах Закавказья он встречается и во влажной земле, в которой роет свои норы. Среди креветок есть также одно целиком пресноводное семейство — Atyidae, характеризующееся присутствием пучков щетинок на пальцах клешней, при помощи которых рачки захватывают грунт. Разбросанное географическое распространение этого семейства, а также его морфологические особенности указывают на его глубокую геологическую древность. В бассейне Амура водится два вида атиид, представляющих вместе со многими рыбами и моллюсками тропический элемент в амурской фауне. Еще одна креветка из этого семейства — Troglocaris anophthalmus (табл. 35, 12) — живет в пещерных водоемах Закавказья. Кроме этих двух семейств, в тропических пресных водах обитают виды крабов и креветок, относящиеся к преимущественно морским родам.
Особый интерес представляют пещерные десятиногие. Известно около 20 слепых пещерных видов Atyidae и 10 видов Palaemonidae, разбросанных по различным континентам и островам. В пещерах США живет 18 видов слепых речных раков, а в Мексике — слепой пещерный краб Typhlopseudothelphusa mocinoi, относящийся к семейству Potamonidae. Наиболее удивительным следует считать существование представителя рода Munidopsis (сем. Galatheidae) в солоноватом подземном озере на одном из Канарских островов; все остальные многочисленные виды этого рода обитают на больших океанических глубинах. Очевидно, и они и подземный Munidopsis polymorpha некогда произошли от вымерших мелководных морских предков и дожили до наших дней в подземных и глубинных водах, как в убежищах, сохранивших много различных остатков древних фаун.
Десятиногие используются в пищу людьми и интенсивно добываются почти во всех странах. В СССР особенно большое экономическое значение имеет промысел камчатского краба (Paralithodes camtschatica), из мускулов ног которого делают консервы. Он принадлежит к семейству Lithodidae, т. е. крабовидных раков-отшельников, и населяет Японское, Охотское и Берингово моря.
Вследствие позднего наступления половозрелости и медленного роста камчатского краба его запасы, подорванные интенсивным промыслом, восстанавливаются с большим трудом. Из огромного количества ежегодно рождающихся личинок до взрослого состояния доживают очень немногие. Остальные становятся жертвами различных хищников или уносятся течениями. Заросли, подходящие для оседания личинок, есть только в Хайрюзовском районе охотоморского побережья Камчатки. Отсюда молодь краба заселяет более южные участки. Однако в годы с пониженной температурой воды на мелководьях такая миграция невозможна, и южные стада камчатского краба совсем не получают пополнения.
Уже более 30 лет тому назад было сделано предложение об акклиматизации камчатского краба в Баренцевом море. В настоящее время есть сторонники и противники этой идеи. Методика перевозки крабов и их личинок с Дальнего Востока разработана, перевозка осуществлена, и сейчас ведутся наблюдения в аквариальных и естественных условиях над поведением камчатского краба в новом для него водоеме.
Краба ловят ставными сетями, которые стремятся поставить так, чтобы перегородить основные пути миграции. Небольшие суда привозят сети с пойманными крабами на краболов, где производят выпутку крабов, отрывают у них ноги и клешни, варят их, рубят, вытряхивают мясо и изготовляют из него консервы. Помимо камчатского краба, в наших дальневосточных морях в меньшем количестве добывают креветок из рода Pandalus. Этот промысел может быть расширен, так как запасы различных креветок в наших морях очень велики. В пресных водах, преимущественно на Украине, ведется промысловый лов речных раков.
За рубежом добывают много различных десятиногих, из которых на первом месте стоят креветки. В 1962 году их было поймано 700 тысяч тонн, причем особенно развит этот промысел в Китае, США, Индии, Японии. Наибольшее значение в уловах имеют семейства Penaeidae, Pandalidae и Crangonidae. В некоторых странах Южной и Юго-Восточной Азии ведется искусственное разведение креветок, преимущественно пенеид. Прибрежные низменные пространства при помощи дамб превращают в цепь прудов, соединяющихся каналом с морем. В прилив вода заливает пруды и остается там благодаря тому что шлюзы во время отлива закрывают. В такие пруды креветки заходят сами, или туда выпускают пойманных в море личинок. Креветок подкармливают рисовыми отрубями. При таком ведении хозяйства удается получать до 500 кг креветок с гектара.
Меньшее значение имеет промысел настоящих крабов, особенно развитый в США. Их продают свежезамороженными, вареными и в виде консервов, причем высоко ценятся недавно слинявшие особи. Из других десятиногих считаются деликатесом лангусты и омары. Только на Кубе ловом лангуста занято более 1000 судов, вылавливающих ежегодно свыше 7500 тонн, что составляет около 1/4 всего улова республики. Запасы европейского омара подорваны промыслом, и Канада экспортирует в Европу довольно значительное количество американских омаров. Для восстановления запасов омара производится его разведение в специальных садках.
Наша страна использует десятиногих еще совершенно недостаточно. Нет сомнения в том, что в будущем их промысел у нас значительно возрастет.