Clupea harengus Linnaeus, 1758
Атлантическая сельдь, Атлантическо-скандинавская сельдь, Океаническая сельдь, Морская сельдь, Многопозвонковая сельдь, Обыкновенная сельдь, Балтийская сельдь, Салака, Мурманская сельдь, Норвежская сельдь, Датская сельдь, Шотландская сельдь, Исландская сельдь, Гренландская сельдь, Банковская сельдь, Полярный залом, Стремлинг, Гигантский стремлинг, Клупея харенгус (Rus),
Atlantic herring, Herring, Sea herring, Sea Atlantic herring, Baltic herring, Salaka, Murman herring, Norwegian herring, Labrador herring, Bank herring, Shore herring, Summer herring, Spring herring, Fall herring, Mesh herring, Sea stick, Yawling, Split
(Eng).
Синонимы:
Clupea alba Yarrell, 1829
Clupea borealis Schnakenbeck, 1931
Clupea elongata Lesueur, 1818
Clupea halec Mitchill, 1814
Clupea harengus atlanticus Schnakenbeck, 1931
Clupea harengus borealis Schnakenbeck, 1931
Clupea harengus britannicus Schnakenbeck, 1931
Clupea harengus caledonicus Schnakenbeck, 1931
Clupea harengus cimbricus Schnakenbeck, 1931
Clupea harengus frisius Schnakenbeck, 1931
Clupea harengus harengus Linnaeus 1758 Атлантическая сельдь (Rus),
Clupea harengus harengus membras Linnaeus, 1761
Clupea harengus harengus murmanica Tikhonov, 1941 Мурманская сельдь (Rus),
Clupea harengus islandicus Schnakenbeck, 1931
Clupea harengus ivernicus Schnakenbeck, 1931
Clupea harengus Linnaeus, 1758
Clupea harengus membras Linnaeus, 1761 Балтийская сельдь, салака (Rus),
Clupea harengus norvegivus Schnakenbeck, 1931
Clupea harengus scandicus Schnakenbeck, 1931
Clupea harengus schelderensis Nilsson, 1832
Clupea harengus scoticus Schnakenbeck, 1931
Clupea harengus septemtrionalis Schnakenbeck, 1931
Clupea islandicus Schnakenbeck, 1931
Clupea leachii Yarrell, 1832
Clupea minima Storer, 1839
Clupea palassi probatowi Makushok, 1935
Clupea septemtrionalis Schnakenbeck, 1931
Clupea vittata Mitchill, 1815
Cyprinus esca Walbaum, 1792
Rogenia alba Valenciennes, 1847
Рыбы открытого океана. Н.В.Парин 1998 г.
Атлантическо-скандинавская (так называемая океаническая) сельдь (Clupea harengus) размножается в прибрежных водах Норвегии и Исландии, откладывая клейкую икру на водоросли, а в период нагула ведет пелагический образ жизни вдали от берега. В годы хорошего состояния запасов она совершает довольно протяженные миграции в открытую часть Северной Атлантики и достигает Баренцева моря, Шпицбергена, Гренландии (сейчас численность этого вида находится в состоянии депрессии).
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Сельдь атлантическая — Clupea harengus harengus Linnaeus, 1758.
Как и у других представителей рода, рот верхний, большой, на верхней челюсти нет центральной выемки. За брюшными плавниками, а иногда и перед ними есть слабовыраженный киль. Последние лучи анального плавника не удлинены. Для вида характерны короткие грудные плавники, длина которых составляет не более 14% абсолютной длины тела. Длина головы составляет менее 23% длины тела. Морской вид. Зоопланктофаг. Достигает длины 40 см. Обитает в северо-восточной части Атлантики, Баренцевом море и в горле Белого моря. Ценнейший объект промысла.
Салака — Clupea harengus membras Linnaeus, 1761
От атлантической сельди хорошо отличается большей длиной грудных плавников (боле 14% длины тела) и большей головой, длина которой составляет не менее 23% длины тела. Имеет меньшие размеры, длина тела редко достигает 20 см. Ареал подвида охватывает Балтийское море.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
При содержании рыб воду лучше подсаливать (5—18 ‰). В аквариуме содержат следующие виды:
Мурманская сельдь - Clupea harengus Linnaeus, 1758, 33 см; есть четыре подвида.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Наиболее известна из семейства сельдевых (Clupeidae) океаническая сельдь (Clupea harengus) распространенная в Атлантическом и Тихом океанах, имеющая много родственных видов. Мировой улов сельдей еще недавно превышавший 6 млн. тонн, что составляло 37% добычи всех рыб. Сельдь является пелагической рыбой. Сельдям родственны сардины рода Sardinа, а также анчоусы рода Engraulis семейства Engraulidae.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.2.1 (1). Clupea harengus Linnaeus, 1758 — Атлантйческая сельдь (Св., 1952 : 123).
[1] русск. Атлантическая сельдь — Св., 1952 : 123. Бельчуг — Гр. : 215. Галлёя кольск. — Подв. Многопозвонковая сельдь — Пром. р. : 120. Морская сельдь — Св., 1952 : 117. Мурманская сельдь, Норвежская сельдь — Б., 1932 : 94. Обыкновенная сельдь — Кессл., 1864 : 185. Океаническая сельдь — Б. : 148. Селёдка — А. Ник. : 792. Сельдина пек. — Опыт. Сельдь — Пром. р. : 120. ~ Seld, Seloetka — Pallas : 209. [12] латыш. Renge — ZK. Silke — Hoek : 47. [13] лит. Silka, Silke, Silke paprastoji, Silkie, Silkis — Majew. [14] дат. Sild — Hoek : 47: Names of fishes : 36. [15] шведск. Sill — Hoek : 47; Б. : 148; Names of fishes : 37. Sillen — Smitt : 954. Stromming — Names of fishes : 37. [16] норв. Sild — Hoek : 47; Б. : 148; Names of fishes : 37. - [17] исл. Hafsild, Sild — Saemundsson : 105. [18] англ. Atlantic herring США, Канада — Bailey : 7; Ricker : 111. Herring — Smitt : 954; Poll : 129; Б. : 148; Saemundsson : 105; Names of fishes : 36; Канада — Ricker : 111; Европа и Сев. Африка — Мартинсеп, 1961 : 87. Sea herring Канада — Ricker : 111. [19] нем. Hering — Hoek : 47; Б. : 148; Names of fishes : 37. [20] голланд. Bliek, Haring — Poll : 129. [21] флам. Haring — Names of fishes : 36. Hering — Hoek : 47. [22] франц. Ering валл. — Poll : 129. Hareng — Hoek : 47; Poll : 120; Б. : 148. Haring валл., Hering валл., Hinring валл. — Poll : 129. [24] исп. Ijitu sardina — Names of fishes : 37. [25] португ. Arenque — Names of fishes : 37.
3.2.1 (2). Ciupea harengus membras Linne, 1761 — Салака (Св., 1952 : 140).
[1] русск. Салака — Б., 1932 : 95. Салакушка Балт. м.: Финский зал. — Кессл., 1864 : 185. Сельдь — Пром. р. : 124. Стремишка — Гр. : 214. Стремлинг Балт. м. — Гейнем. : 429. ~ Обыкновенная сельдь — Озерецк., 1791 : 77; Блуменбах : 408; Теряев : 53; Ловецкий : 488. Салакуша — Нордстет; САР; Сл. 1847 г. Салакушка — Начерт. : 397; Нордстет; САР; Сл. 1847 г. Стремлинг — Торговый устав 1724 г. [4] польск. Krocz, Perk, Rap — Majew. Sledz — Majew.; Names of fishes : 37. Szliedz — Majew. [15] шведск. Sill — Б., 1940б : 6. Stromming — Б. : 150. [18] англ. Baltic herring Канада — Ricker : 111. [19] нем. Stromling — Б. : 150. [36] финск. Haili — Hoek : 47; Names of fishes : 36. Silakka — Hoek : 47; Names of fishes : 36; Brofeldt : 157. Silli — Hoek : 47. [37] карел. Салатти Коткозеро, Салкой Крошнозеро — А. [38] эст. Raim — Б., 19406 : 6. Silk — Hoek : 47. [40] ижор. Aili — Кессл., 1864 : 185.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Атлантическая, или многопозвонковая,
сельдь (Clupea harengus) внешне очень
мало отличается от восточной. Для нее
характерно большее число позвонков,
54—59 (60), чаще всего 55—58, большее
число продольных рядов чешуи, наличие
сравнительно сильных зубов на сошнике,
иной характер кариотипа (набора
хромосом). Как показано ниже, она
существенно отличается от восточной сельди по
биологии, особенно по биологии
размножения. Различают две формы (подвида)
атлантической сельди — собственно
атлантическую сельдь (основная, или
номинальная, форма), распространенную в
водах северной части Атлантического океана
и сопредельных морей Ледовитого океана,
и балтийскую сельдь, или салаку.
Атлантическая сельдь (Clupea harengus
harengus) достигает длины до 36 см,
у Исландии — до 42 см. Она
распространена от мыса Гаттераса на западе и
Бискайского залива на востоке до
Гренландии, северо-западных берегов
Шпицбергена и Новой Земли. Область
распространения ограничивается водами
атлантического происхождения, и очень редко
восточные или северные границы нахождения
этого вида заходят за границу плавучих
льдов. Размножение атлантической
сельди происходит только в южной половине
ареала, самые северные нерестилища
расположены у Лофотенских островов и
островов округа Тромсе (до 70—71° с. ш. на
север). Дальше к северу и востоку
встречается либо молодь, занесенная в
Баренцево море Нордкапским течением,
либо взрослые особи, проникающие в
окраины Гренландского моря по
Шпицбергенскому течению. Размножение всех рас
атлантической сельди происходит при
температуре не ниже 4—5° С.
Различают несколько рас
атлантической сельди.
Наиболее многочисленной расой
являются весенненерестующие атлантическо-скандинавские сельди. Они приближаются
к берегам только в период размножения,
в марте — апреле. Размножение атлантическо-скандинавских сельдей
происходит у берегов Норвегии, у Оркнейских
и Шетландских островов со стороны
океана, нерестилища известны на свалах
цоколя Фарерского архипелага и вдоль
южных берегов Исландии. Особенно крупные
нерестилища находятся у юго-западных
берегов Норвегии. Все районы
размножения сельдей находятся под сильным
влиянием атлантических течений.
Подхваченные течением личинки уносятся далеко
на север. От берегов Южной Норвегии
молодь приносится к Вест-фьорду; от
Лофотенских островов — к берегам Мурмана,
в центральные и восточные районы
Баренцева моря, а также к острову
Медвежьему; часть молоди выносится в
открытые районы восточной половины Норвержского моря; из южных районов
Исландии молодь приносится течением Ирмингера к ее северным берегам.
Условия откорма молодых сельдей
предопределяются районом распространения
молоди в итоге ее дрейфа. Чем севернее
и восточнее заносится молодь, тем в
худшие условия существования она попадает.
Сельди, выросшие в западных районах
Баренцева моря, в пять лет достигают длины
24—25 см и в этом возрасте становятся
половозрелыми. В восточных районах за
5 лет сельди вырастают только до 18—19 см и созревают только в 7—8 лет.
Во время возвратной миграции к
нерестилищам стаи сельди группируются
по размерному признаку, который отражает в известной мере их
физиологическое состояние. К преобладающей в стае
возрастной группе присоединяются особи
старших возрастов, отставшие в росте,
и молодые экземпляры с высоким темпом
роста.
После размножения атлантическо-скандинавские сельди вступают в новую фазу
своего жизненного цикла. Вначале,
ослабевшие после нереста, они сносятся
течением, а дальше уже предпринимают
активные миграции в районы откорма —
в область полярного фронта, к северным
берегам Исландии, в район порога Мона
и далеко на север по Шпицбергенскому
течению.
Эта миграция происходит с большой
скоростью, особенно у стай,
мигрирующих на север,— до 8—10 км в сутки.
Течения ускоряют нагульные миграции.
В конце июля стаи сельдей достигают уже
наиболее удаленных районов и, накопив
запасы жира, начинают обратную
миграцию. Осенний миграционный путь
проходит значительно западнее. После нереста
течения способствуют движению рыб,
идущих нагуливаться. Во время нерестовой
миграции эти же течения замедляют
движение и стаи сельди, экономя силы,
обходят основные потоки Норвежского течения
с запада.
Откармливаясь в поверхностной толще,
где развитие жизненных процессов,
связанных с откормом сельди, протекает
очень быстро, стаи сельди уже в начале
августа достигают наивысшей
упитанности, после чего у них быстро развиваются
половые продукты. Двигаясь с прежней
скоростью, около 7 км в сутки, стаи сельди
уже в декабре могли бы прибыть на
нерестилища, но в это время в водах Норвегии
еще зима, планктон не начал развиваться,
корма для личинок нет, и сельди
задерживаются в пути, в районе Восточноисландского течения, где низкие
температуры способствуют снижению их обмена.
Стаи держатся на глубине нескольким
сотен метров при температуре 1—2° С.
С наступлением весны они быстро
переходят к нерестилищам и первые приступают
к размножению. Вслед за сельдью идут
хищные рыбы — сайда, треска, пикша.
Сотни норвежских ботов, вооруженных
кошельковыми неводами и дрифтерными
сетями, встречают стаи сельди,
приближающиеся к берегам. В практике
норвежского промысла известны случаи, когда
за сутки добывалось более 100 тыс. т
сельди, а за сезон, продолжающийся
менее месяца, улов составлял 1,0—1,2 и
даже 1,5 млн. т.
У норвежских берегов издавна
различаются четыре промысловые категории
сельди: 1) мелкая сельдь, 7—19 см длины,
в возрасте 1—2,5 года; 2) жирная,
подрастающая сельдь от 19 до 26 см длины,
в возрасте 2,5—4 года; 3) крупная,
преднерестовая сельдь и 4) весенняя
нерестовая сельдь, имеющие длину от 27 до 32 см
и возраст от 4 до 8 лет и выше. Промысел
производится в периоды подхода их к
берегам: мелкой сельди — на севере,
жирной — у Средней Норвегии, крупной и
нерестовой — у Южной Норвегии.
Долгое время лов атлантическо-скандинавской сельди производился только
вблизи берегов. Исследованиями
советских ученых были прослежены миграции
сельди и обнаружены скопления ее в
открытом море, вдали от берегов. На этих
скоплениях был организован эффективный
промысел.
В Баренцевом море живут только
молодые возрастные группы атлантическо-скандинавских сельдей (до 5—7 лет).
С наступлением половой зрелости они
переходят в Норвежское море и вливаются
в общее стадо атлантическо-скандинавских сельдей. У Мурманского побережья,
как и в Норвегии, молодая сельдь
нередко заходит в заливы (губы). Существовал
специальный «запорный» промысел такой
сельди. Выход зашедшей в губу стае
запирался огромной сетью, и запертая
сельдь успешно вылавливалась. Особенно
большие уловы сельди в губах Мурмана
были получены в 1933—1935 гг. (от 68
до 103 тыс. т в год).
Значительные колебания численности
отдельных поколений сельди при небольшом числе их в Баренцевом море (всего
2—3 группы) не позволили создать здесь
большого и устойчивого промысла.
Хорошие подходы сельди в Баренцево море
наблюдаются периодически. В соседнем,
Норвежском, море взрослое стадо
состоит из 8—10 взрослых групп, и
возможности сельдяного промысла здесь более
стабильны.
Атлантическо-скандинавские сельди
обладают огромным высокопродуктивным
нагульным ареалом и достигают больших
размеров, чем другие расы; имея высокий
темп роста, они живут до 15—18 лет и,
как следствие этого, обладают
многовозрастной структурой нерестового стада.
В последние годы в связи с
интенсификацией промысла много внимания
уделялось изучению численности и
величины стада сельди.
Многие десятки тысяч рыб были
помечены небольшими стальными пластинками,
которые с помощью пружинных
пистолетов загонялись в полость тела. Такой
способ пометки сельди переносят вполне
удовлетворительно. Обнаружение меток
среди пойманных рыб делалось с
помощью специальных электромагнитов,
которые извлекали метки из рыбной муки,
приготовленной из сельди. Запас
определялся по формуле, согласно которой
общее количество помеченных рыб относится
ко вторично пойманным как запас к
улову. Этим методом оценивали запасы
норвежские ихтиологи. Советскими учеными
был применен метод определения объема
скоплений сельди на местах зимовки с
помощью гидроакустических приборов и
определения плотности скоплений путем
подводного фотографирования.
Оба метода дали сходные результаты.
Запас атлантическо-скандинавской
сельди в период наилучшего пополнения
достигал 12,0—12,5 млн. т.
Вторая раса — летненерестующие
сельди — объединяет несколько стад,
населяющих воды Исландии и Фарерских
островов, южные фиорды Гренландии, и в
особенности (наиболее многочисленное
стадо) воды шельфа Новой Англии и
Новой Шотландии, на банке Джорджес.
Нерест летненерестующих сельдей
происходит в течение второй половины лета,
и протяженность их миграций гораздо
меньше, чем у весенненерестующих
сельдей. Откорм их разделяется на два
периода: весенний, до размножения, и осенний,
после нереста. От южных берегов
Исландии они удаляются на север всего на 200—300 миль. Миграции сельдей, живущих
на шельфе Северной Америки,
ограничены банкой Джорджес и заливом Фэндибей.
Все летненерестующие сельди
отличаются небольшим ростом в первый год
жизни, но на втором-третьем году они
почти достигают размеров сельдей,
размножающихся весной.
Летненерестующие сельди обладают
значительно большей плодовитостью. Атлантическо-скандинавские сельди длиной
около 32—33 см редко имеют количество
икринок больше 70—75 тыс., обычно
50—60 тыс. У летненерестующих сельдей
тех же размеров плодовитость достигает
150—200 тыс. икринок. Однако запасы
этих сельдей гораздо меньше запасов весенненерестующих.
Шельф Северного моря и прилегающие
районы населяют так называемые
банковые сельди, размножающиеся летом
и осенью вдали от берегов, на
мелководных банках, и сельди солоноватых вод
(Датских проливов, Зюдерзее),
нерестующие в весеннее время вблизи берегов,
в зонах значительного опреснения.
Все стадии жизненного цикла сельдей
Северного моря протекают в его
пределах. Несмотря на длительные
исследования расового состава сельдей Северного
моря, вопрос этот не решен в полной
мере и до сегодняшнего дня. По ряду
признаков может быть выделено три
стада: северное, размножающееся на банках,
прилегающих к Северной Шотландии;
второе стадо, имеющее местом своего
размножения Доггер-банку; и третье, ламаншское стадо, нерестующее в Ламанше,
Наибольшее количество молоди сельди
в Северном море находится в его
юго-восточной части, куда она, несомненно,
заносится из более северных районов моря.
В последние годы наряду с более
интенсивным использованием взрослой части
стада стал развиваться и промысел
молоди для получения кормовой муки и
жира.
Темп роста сельдей Северного моря
значительно ниже, чем у атлантическо-скандинавских. Они редко достигают длины
30 см, обычные их размеры 26—28 см.
Они становятся половозрелыми в 3—4 года
и никогда не бывают старше 8—10 лет.
В стаде при современном интенсивном
промысле преобладают рыбы 3—6 лет.
Подвид балтийская сельдь, или салака
(Clupea harengus membras), населяет
Балтийское море к востоку от Датского
пролива. Она отличается своей малой
величиной, обычно менее 20 см длины, причем
становится половозрелой начиная от
13—14 см длины, в возрасте 2—3 лет.
Живет салака до 6—7 лет. Однако среди
обычной салаки попадаются и так называемые
гигантские салаки, растущие гораздо
быстрее и достигающие 33 и даже 37,5 см
длины. Тогда как обычная салака
питается планктоном, гигантская салака —
хищная рыба, нередко питающаяся трехиглой
колюшкой.
Кроме малой величины, салака
отличается от собственно атлантической
сельди меньшим числом позвонков, которых
у нее 54—57, и биологией. Населяя всю
восточную часть Балтийского моря и его заливы, постоянно обитая в воде
пониженной солености, салака встречается
иногда и в совершенно пресной воде
некоторых озер Швеции. Нерестится салака на
твердом, каменисто-гравиевом грунте, на
глубине от 2—3 до 20 м. Нерест
происходит весной, отчасти летом и осенью, в
связи с чем различаются две группы форм —
весенние и осенние сельди.
Салака — главная промысловая рыба
Балтийского моря, дающая около
половины всего улова, добываемого в этом
водоеме. Ловят ее главным образом у
берегов, ставными сетями и неводами.
В 1954—1956 гг. была сделана
интересная попытка акклиматизации салаки
в Аральском море. Через три года
после завоза первой партии икры выросшая
из нее салака начала нереститься, а в
1960 г. происходил массовый нерест
салаки аральского происхождения. В первые
годы салака в Аральском море очень
хорошо росла и нагуливалась, гораздо лучше,
чем в исходных Рижском и Вислинском
заливах. Но затем запасы кормовых для
салаки рачков диаптомуса и дикерогаммаруса сильно сократились, темп роста
и упитанность салаки понизились.
Промыслового стада салаки в Аральском
море пока не образовалось вследствие
недостатка корма и обилия врагов и
конкурентов.
Первое упоминание о лове
атлантической сельди встречается уже в 702 г.
в монастырских хрониках Англии. Уже
тогда сельдь служила источником
благосостояния.
С XI и вплоть до XV столетия соленая
(сухим, стоповым посолом) сельдь была
важным объектом торговли ганзейских
купцов, и на базе этой торговли возросло
и базировалось в течение не менее 350 лет
морское могущество Ганзейского союза
городов. Ганзейские рыбаки промышляли
сельдь главным образом у немецких и
датских берегов Балтийского моря. Однако
в XV в. подходы сельди к этим берегам
стали гораздо меньше; были годы, когда
она вовсе не подходила, и уловы здесь
стали катастрофически падать. В то же
время были обнаружены громадные
подходы сельди к берегам Голландии и
Шотландии.
Голландцами был открыт способ
мокрого посола сельди в бочках на кораблях,
появились и специальные суда — логгеры — для лова сельди в море. Промысел
сельди сыграл огромную роль в развитии
экономики Голландии в XV—XVI вв.
Тогда уже сельдь промышляли вдали от
берегов с помощью дрифтерных сетей со
специальных парусных логгеров, на
которых сельдь засаливалась в бочках и уже
в готовом виде доставлялась на берег.
С XVII столетия морской промысел
сельди начинает развиваться в Англии,
которая очень скоро заняла первое место
в сельдяном рыболовстве европейских
стран, сохранявшееся за ней до начала
первой мировой войны.
Затем больше всего сельди стали
добывать в Норвегии. Наконец, в 1950—1967 гг. наибольшие уловы сельди брали
Норвегия, Советский Союз, Исландия,
Канада и Дания.
Обитатели бездны. Уильям Кроми 1971 г.
Для всех океанских рыб характерны темная спина и светлое брюхо. Серебристое или белое брюхо снизу трудно разглядеть на фоне освещенной солнцем поверхности воды и врагам, и жертвам этих рыб. Если же глядеть сверху, синие и зеленые цвета на спине сливаются с морем и темным дном. Когда свет сверкает на спине, на белое брюхо падает тень, благодаря чему рыба сбоку кажется как бы плоской и равномерно окрашенной под цвет воды. Отблески солнечного света делают неясным пигмент на боках и спине, а нижняя сторона тела, несмотря на тень, остается довольно бледной, поскольку пигмента на ней нет.
Это двухцветное одеяние как нельзя подходит для жизни в открытом море. Вдали от берегов и дна нет нужды в кричащих, пестрых красках и быстрых сменах маскировки. Среда тут остается неизменной, поэтому неизменен и покров. Обыкновенная сельдь (Clupea harengus), которая огромными стаями скитается по Северной Атлантике, всегда одета в синевато-зеленый «пиджак» и серебристо-серые «брюки».
Типичный житель открытого моря, где обитают многие виды рыб, маленькая сельдь, пожалуй, самая многочисленная рыба в приповерхностных слоях воды. Атлантическая сельдь и ее тихоокеанская сестра Clupea pallasi передвигаются огромными многомиллиардными косяками, которые подчас достигают длины в несколько миль и такой же ширины. Иногда кажется, что море битком набито их серебристыми, длиной сантиметров в 30 телами.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
la. Clupea harengus harengus Linne — Атлантическая сельдь, мурманская сельдь, многопозвонковая сельдь.
Clupea harengus Linne, 1758 : 317 (Атлантич. океан у берегов Европы). — Неinсkе, Naturgesch. d. Herings, I, 1898 : LIX—LXIII и др., многочисл. табл. намерений, иллюстр. (частью: Атлантич. океан у берегов Европы; моногр., расовый анализ). — Sсhnakеnbесk, Zeitschr. Morph. Okol. Tiere, XXI, 1931 : 482—566 (подвиды: harengus, britanicus, atlanticus, ivernicus, caledonicus, scoticus, frisicus, cimbricus, scandicus, norvegicus, septemtrionalis, borealis, islandicus). — Le Gall, Ann. Inst. Oceanogr., Paris, XV, 1, 1935 : 99—165 (расовый анализ восточноатлантических сельдей; цит. по Световидову). — Карантонис, Тр. Полярн. научно- иссл. инст. морск. рыбн. хоз. океаногр., 1, 1938 : 59—73, фиг. 1—6 (морфология трех типов сельдей Баренц. м.).
Clupea harengus harengus Берг, Рыбы пресн. вод России, 1923 : 500. — Берг, Рыбы пресн. вод СССР, I, 1948 : 148. — Световидов, 1952 : 123, табл. II (диагноз сельди из Алезунда; биология, библиография).
Clupea harengus harengus n. murmanica Тихонов, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 7, 1941 : 3—37 (Баренц. м.).
Распространение, биология. (По мурманской сельди, как и по другим формам сельдей, указываются лишь основные работы последних лет; подробнее см. у Есипова (1938, 1939) и Световидова (1952).) Есипов, Промысл. рыбы Баренц. м., 1937 : 22. — Глебов, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 1, 1938 : 5—58 (условия захода в губы). — Мурашкинцева, там же : 74—87 (типы зимних колец). — Мантейфель и Болдовский, там же : 88—101 (причины зимних заходов в губы). — Болдовский, там же : 102—119 (питание в губах). — Алеев, там же: 120—133 (распределение в южн. части Баренц. м.). — Есипов, там же : 160—169 (библиография до 1937 г. вкл.). — Соловьева, там же, 2, 1938 : 109—129 (время закладки зимних колец). — Тихонов, там же, 4, 1939 : 5—40 (зимнее распределение в южн. части Баренц. м.). — Мантейфель и Марти, там же : 41—94 (нерест). — Юданова, там же : 95—108 (сезонные изменения жирности). — Пчелкина, там же : 109—128 (распределение в связи с планктоном). — Есипов, там же : 149—222 (размножение; литературн. сводка). — Расс, там же, 6, 1939 : 93—164 (нерест и жизненный цикл). — Рыженко, Сб., посвящ. Н. М. Книповичу, 1939 : 394—410 (личинки и мальки на Мурмане). — Есипов, Тихонов, Марти, Болдовский, Мантейфель и др., Биолог. и пром. мурм. сельди, изд. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 1939 : 1—100. — Марти, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 7, 1941 : 38—124 (жизненный цикл). — Мантейфель, там же : 125—218 (распределение в связи с планктоном). — Болдовский, там же : 219—286 (питание в море по сезонам).— Andersson, Fiskar Fiske Norden, I, 1942 : 227—246, pl. 64 (биология, цветн. фотогр.). — Марти, Рыбн. промышл. СССР, 3, 1945 : 3 (современный промысел). — Дмитриев, Биолог. и пром. сельди в Белом м., 1946 : 41—52 (заходы в Белое м.). — Расс в изд.: Промысл. рыбы СССР, 1949 : 120, атлас табл. 30 (сводка, цветн. рис.). — Sаеmundsson, Zool. Iceland, IV, 72, 1949 : 104 (Исландия; литература). — Красикова, Световидов, Природа, 8, 1951 : 46—47 (Енисейский зал.). — Марти в изд.: Промысл. рыбы Баренц. Белого м., 1952 : 47—63 (биологич. очерк).
D (всего лучей) (16) 17—19 (20), среднее 18.2, А (14, 15) 16—18 (19), ср. 17.0, жаберных тычинок (61—63) 64—71 (72), ср. 67.1. (По данным Рабинерсона (Изв. Отд. прикл. ихт., III, 1, 1925), для 200 сельдей из Тюва-губы.) Позвонков, по данным Тихонова (1941) на основании анализа более 50 тыс. мурманских сельдей (55) 56—58 (59), средние цифры за 9 лет (1930—1938 гг.) варьируют в пределах 57.15—57.35. По Шнакенбеку, большинство рас атлантических сельдей по числу позвонков укладываются в формулу (54) 55—58 (59). (По данным М. П. Сомова, число позвонков в урожайных поколениях в среднем выше, чем в неурожайных.) Световидов для норвежской сельди (38 экз. из Алезунда) дает следующую формулу: D 17—21, ср. 18.9, А 16—20, ср. 18.0, жаберных тычинок 66—73, ср. 70.1, килевых чешуек позади основания брюшных плавников 13—16, ср. 14.3, позвонков 56—59, ср. 57.06.
Килевые чешуйки развиты как позади, так и впереди основания брюшных плавников. Зубы на сошнике сравнительно хорошо развиты,
расположены в 2—3 ряда; число зубов у разных форм атлантических сельдей варьирует от 7 до 22, чаще более 10, средние цифры 10.7—15.8. (См. у Гейнке, 1898 : 42, 47, 71, 111, 141, 146, 152; об изменчивости этого признака см. у Световидова, 1952.) Длина взрослых в Баренцевом море 25—33 см, у берегов Шпицбергена — до 37 см («полярный залом»); крупные исландские сельди достигают 40—42 см.
Из многочисленных форм атлантических сельдей в пределах Баренцева моря встречается лишь две расы, обе относящиеся к группе атлантическо-скандинавских весенних сельдей: 1) норвежско-мурманская (или мурманская) сельдь, нерестящаяся главным образом у Лофотенских островов и проводящая большую часть жизни в Баренцовом море, и 2) норвежская сельдь (Повидимому, соответствует сельдям III морфологического типа Ф. 3. Карантониса (см. у Марти, 1941).), которая нерестится у юго-западных берегов Норвегии и ловится в небольшом числе в юго-западной части Баренцова моря (см. ниже).
Распространение. Атлантическая сельдь распространена в северной части Атлантического океана и частично в прилежащих арктических водах. В западной части океана от северных берегов Лабрадора на юг до о. Блок (около 41° с. ш.), а в зимнее время спускается до м. Гаттераса; юго-зап. Гренландия (в теплые годы до 72.5° с. ш.); юго-вост. Гренландия; вокруг Исландии. У берегов Европы от Бискайского залива до Британских, Шетландских и Фарерских островов; проливы Скагеррак, Каттегат и западная часть Балтийского моря до о. Эланд и Гданьской бухты (восточнее n. membras L.); берега Норвегии и открытая часть Норвежского моря. В Баренцовом море норвежская сельдь заходит лишь в юго-западную часть, не восточнее 35° в. д. Ареал норвежско-мурманской сельди по сравнению с таковым норвежской сдвинут к северу — от Лофотенских островов до Медвежинской банки, западных берегов Шпицбергена (78° с. ш.) и южных склонов возвышенности Персея (76°48' с. ш., 33°19' в. д.); на восток до Новой Земли (до широты Маточкина Шара), а в южной части Баренцова моря в летнее время заходит до Чёшской губы, о. Колгуева и западной части Печерской губы; заходит во все заливы Белого моря, где часто ловится с беломорской (малопозвонковой) сельдью. В августе 1948 г. в бухте Омулевой Енисейского залива был пойман 1 экз. сельди, который А. II. Световидов относит к типичному (многопозвонковому) подвиду (позвонков 58, килевых чешуек позади V 14).
Биология. [Биология мурманской сельди прекрасно изучена и данные опубликованы в «Трудах» Полярного института рыбного хозяйства (см. синонимию); основные сведения обобщены в монографии А. Н. Световидова и в атласе «Промысловые рыбы СССР». Важные новые сведения о биологии мурманской сельди изложены в книге «Промысловые рыбы Баренцова и Белого морей» (Марти).] Морская стайная рыба, обитающая преимущественно в поверхностных слоях воды; в зимнее время и для нереста опускается в придонные горизонты, но не глубже 200—300 м. Обычна при положительных температурах и высоких соленостях, но в некоторых районах переносит и значительное опреснение; изредка заходит и в устья рек; некоторые южные популяции (Каттегат, Зунд, ранее — Зюйдерзее) нерестятся при солености 8—22%. Вообще во времени и условиях нереста глубине до 40—50 м, в то время как банковские сельди Северного моря, как и мурманская форма, нерестятся на глубине до 200 м. По времени нереста различают весенних, летних, осенних и зимних сельдей; из них наиболее многочисленны весенние и осенние сельди. Нерестилища мурманской сельди расположены у Лофотенских островов, в основном от Сере до Рёста, в значительно меньшей степени и в более северных районах (Финмаркен). Нерест в марте—апреле. Икра выметывается на песчаный или гравиевый грунт на глубине (50) 130—225 м при температуре около 4—7° и солености 34—35‰. Плодовитость атлантической сельди колеблется в пределах от 10 до 100 тыс. икринок, у мурманской составляет 15—23 тыс. икринок диаметром 1.6—2.1 мм. Через 2—3 недели после нереста выклевываются личинки Длиной 5—8 мм, которые через неделю начинают вести пассивно пелагический образ жизни и далеко разносятся течениями, достигая о. Медвежьего и берегов Мурмана. К августу—сентябрю, по достижению длины 4—6 см, они массами подходят к берегам, часто встречаясь близ устьев рек. Сформировавшиеся сеголетки размером более 6—7 см (так называемая «килька») начинают вести активно пелагический образ жизни, широко расселяясь почти по всему Баренцову морю. Молодая сельдь (до 2—3 лет — «мелкая сельдь», в 3—4-летнем возрасте — «жирная сельдь») периодически подходит к берегам — «килька» и мелкая сельдь преимущественно летом, жирная сельдь зимой. Созревающая сельдь постепенно отходит в юго-западные районы Баренцова моря, приближаясь к местам нереста у Лофотенских островов. Повторно нерестующие крупные сельди (в возрасте 9—10 лет и более) отходят после икрометания далеко на север, достигая западных берегов Шпицбергена («полярный залом»). Рост сельди в Баренцовом море происходит следующим образом («Промысл. рыбы СССР»; длина до конца средних лучей С в см, вес в г):

Половозрелости мурманская сельдь достигает в массе в 5—6-годовалом возрасте. О дальнейшем ее росте можно судить по анализам крупных (уже нерестовавших) сельдей из шпицбергенского района (Марти, 1941) : 7+ 28.8 см, 9+ 30.2 см, 11+ 31.4 см, 13+ 32.2 см. Продолжительность жизни 15—20 лет.
Главным пищевым объектом сельди Баренцова моря служат Euphausiidae и Calanus finmarchicus. Болдовский (1941) установил следующую схему сезонного изменения интенсивности питания мурманской сельди. После очень слабого зимнего периода питания, с марта—апреля, интенсивность питания сельди возрастает (в основном за счет Euphausiidae), но возобновление роста и увеличение жирности начинается позднее, с конца мая. Наиболее интенсивный период откорма приходится на июнь—июль (Calanus), после чего осенью (август—октябрь) постепенно ослабевает (период докорма); в этот период в желудках сельди преобладают Euphausiidae, а временами также молодь рыб (трески, песчанки, собственные мальки), Limacina и др. Наибольший рост происходит в период летнего откорма, а также осенью, наибольшая жирность достигается в августе—сентябре.
Нерест норвежской сельди происходит у юго-западных берегов Норвегии в феврале—апреле, личинки вдоль норвежских берегов относятся
течением к Финмаркену (smaasild норвежцев), а по достижении 2—3-летнего возраста активно мигрируют на юг обратно (fetsild). С приближением половозрелости, по предположениям норвежских авторов, часть сельди отходит в открытое море, проходя, так называемую, океаническую стадию (обнаруживается по строению чешуи). Подходящая к берегам Норвегии для нереста крупная сельдь (storsild, vaarsild) имеет возраст 4—7 лет.
В Баренцовом море норвежская сельдь имеет ограниченное распространение, повидимому, заходя лишь в его юго-западную часть в молодом возрасте.
Хозяйственное значение. Важнейшая промысловая рыба северной части Атлантического океана, занимающая второе место в мировом промысле (15—16 млн ц). Наибольшее количество сельди вылавливается в Северном море (до 9—10 млн ц), у берегов Норвегии и Исландии. В Баренцовом море наши уловы в урожайные годы достигают 1 млн ц.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*1. Clupea harengus L. — Морская, или океаническая сельдь.
Литературу см. при подвидах.
Зубы на сошнике есть. Sp. br. 58—73, на нижней части дуги 40—51.
Морской вид, но некоторые формы размножаются и отчасти живут в солоноватой воде.
Разделяется на два подвида, которые в свою очередь распадаются на ряд форм, причем подвиды и их формы по количеству позвонков, степени развития зубов и ряду биологических особенностей разделяются на две группы — многопозвонковых и малопозвонковых сельдей. К многопозвонковым сельдям относятся атлантическая (Clupea harengus harengus) и балтийская (Clupea harengus harengus n. membras) сельди, к малопозвонковым — тихоокеанская (Clupea harengus pallasi), чёшско-печорская (Clupea harengus pallasi n. suworowi) и беломорская (Clupea harengus pallasi n. maris-albi) сельди. Биологические различия обеих групп сводятся к следующему. 1) У многопозвонковых сельдей икрометание происходит на протяжении всего года, в связи с чем среди них, в частности у атлантической сельди, по времени нереста различают весенних, осенних, летних и зимних сельдей. Малопозвонковые сельди размножаются весной, и поэтому у них по времени икрометания есть только весенние сельди. 2) Икрометание у многопозвонковых сельдей происходит в общем при несколько более высокой температуре, чем у малопозвонковых. 3) Нерестилища многопозвонковых сельдей располагаются наичаще на значительном отдалении от берегов, иногда на банках среди моря, и на больших глубинах, чем у малопозвонковых, у некоторых форм атлантической сельди на глубине до 200 м. Нерест малопозвонковых сельдей происходит в узко прибрежной зоне на небольших глубинах, максимально до 10—15 м. 4) В связи с этим икра малопозвонковых сельдей, как правило, откладывается на подводную растительность, между тем как у многопозвонковых сельдей икра откладывается на грунт, так как на таких глубинах, где они размножаются, подводной растительности обычно нет. 5) Малопозвонковым сельдям свойственна большая по сравнению с многопозвонковыми приуроченность в период икрометания к опреснениям. 6) Формам малопозвонковых сельдей свойственна большая локальность и привязанность к узким районам распространения и в общем меньшие по размерам миграции, чем многопозвонковым, причем у последних миграции происходят и в личиночной стадии в виде пассивного дрейфа их с течениями (см. об этом «Биология»). Эти биологические различия обеих групп сельдей обусловлены различным происхождением их. Биологические особенности малопозвонковых сельдей выработались в условиях арктических морей, где большинство их (Clupea harengus pallasi n. maris-albi и Clupea harengus pallasi n. suworowi) обитает и в настоящее время. В арктических морях температурные условия, при которых происходит икрометание и развитие икры и личинок Clupea harengus, кратковременны, и, чтобы иметь, соответствующую температуру для развития икры и личинок, размножаться эта сельдь здесь может лишь весной. Поэтому в арктических морях и имеются лишь весенние сельди. По этой же причине и у Clupea harengus harengus в северных частях ареала (у Норвегии, Исландии, склонов континентального плато Северного моря и др.) имеются лишь весенние сельди, размножающиеся весной и отчасти в конце зимы и летом, а осенние свойственны южным частям ареала (Северное море, Ламанш и др.). Температурными условиями арктических морей обусловлено и икрометание малопозвонковых сельдей на небольшой глубине у самых берегов, так как вода только здесь и прогревается до той температуры, при которой возможны икрометание и развитие икры и личинок. Склонность малопозвонковых сельдей к опреснениям в период икрометания обусловлена тем, что более теплая вода рек и поверхностного стока оказывает отепляющее влияние на прибрежные воды, вследствие чего в арктических морях икрометание частично и происходит вблизиустьев рек. Биологические особенности Clupea harengus pallasi, аналогичные биологическим особенностям других малопозвонковых сельдей, выработались несомненно также в арктических морях. Попав из арктических морей в умеренные части Тихого океана, малопозвонковая сельдь образовала здесь особый подвид, широко расселившийся здесь, но сохранивший все биологические особенности этих сельдей арктических морей. В силу консерватизма своей наследственности этот подвид оказался неспособным дать формы, размножающиеся на больших глубинах в отдалении от берегов и в другое время кроме весны, вследствие чего у него нет и осенних форм (подробнее об этом см.: Световодов, 1949б).
Беломорская и чёшско-печорская сельди приняты Л. С. Бергом за nationes Clupea harengus pallasi, что безусловно правильно отражает их родственное и подчиненное отношение к последней. Балтийская сельдь, имеющая от Clupea harengus harengus такие же отличия, как формы малопозвонковых сельдей от Clupea harengus pallasi, и являющаяся в сущности одной из местных форм атлантической сельди, принята им в качестве подвида; следует отметить, что некоторые из подвидов других видов семейства (например Sprattus sprattus, Sardinops sagax, отчасти Sardina pilchardus и др.) имеют такую же степень различия, что и формы многопозвонковых и малопозвонковых сельдей и поэтому следовало бы считать и формы обеих групп также подвидами, подобно тому как это принято для Gadus morhua, который также разделяется на две группы подвидов, аналогичных группам и формам Clupea harengus и имеющих сходные и биологические особенности, обусловленные их происхождением (Световидов, 1944, 1948). Однако при таком наименовании разделения Clupea harengus на две группы подчиненного значения форм каждой из этих групп не видно. Кроме того, и степень различия форм обоих подвидов Clupea harengus значительно меньше, чем подвидов обоих групп Gadus morhua. Поэтому в данном случае удобнее, несмотря на громоздкость наименования, принять разделение Clupea harengus на два подвида — многопозвонковых (Clupea harengus harengus) и малопозвонковых (Clupea harengus pallasi) сельдей, а формы их за nationes. При этом балтийскую сельдь, принимая во внимание то, что она является такой же формой Clupea harengus harengus, как, например, атлантическо-скандинавские сельди или близкие к ней сельди датских проливов, следует считать за natio атлантической сельди. Резких различий между этими подвидами в основных систематических признаках, так же как и в других, приводимых ниже в диагнозах, нет. Развитие зубов, на сошнике, более сильных и многочисленных у атлантической сельди, имеет также переходный характер, так как одинаковая степень развития встречается у той и другой. Наблюдаемые между многопозвонковыми и малопозвонковыми группами сельдей резкие различия личинок в положении анального отверстия (на 47—49-м сегменте и под 82—83-й клеткой хорды у Clupea harengus harengus и на 42—45-м сегменте и под 62—67-й клеткой хорды у Clupea harengus pallasi n. maris-albi) и в числе клеток хорды (более 100 у Clupea harengus harengus и приблизительно 90 у Clupea harengus pallasi n. maris-albi) обусловлено малым числом исследованных личинок. При исследовании большего числа их и при этом у всех форм как Clupea harengus harengus, так и Clupea harengus pallasi, эти, несомненно варьирующие в значительно более широких пределах признаки, в особенности число клеток хорды, будут иметь заходящий характер. Наоборот, в нижеследующем описании подвидов hiatus имеется в некоторых признаках между атлантической и балтийской сельдями, с одной стороны, и тихоокеанской и чёшско-печорской и беломорской, — с другой. Различия эти обусловлены тем, что характеристика атлантической сельди дана лишь по одной весенней форме (атлантическо-скандинавской), так же как и балтийской (весенняя сельдь). При дальнейшем исследовании обеих этих сельдей, основанном на изучении и других форм их, в частности и осенних той и другой и более мелких по размерам форм атлантической, имеющиеся резкие различия в некоторых признаках (длина головы, размеры глаз) несомненно сгладятся. Равным образом и описание подвидов и малопозвонковых сельдей дано лишь по одной из форм каждой из них: тихоокеанской— по южносахалинско-хоккайдской, беломорской — по мелкой сельди Онежского зал., а чёшско-печорской — вообще по недостаточному числу экземпляров.Резкие различия и между этими подвидами наблюдаются в тех же признаках, что и между атлантической и балтийской сельдями, и в обоих случаях обусловлены, помимо указанных причин, в значительной степени и разными размерами их (более крупными атлантической и тихоокеанской сельдей и более мелкими остальных).
Когда работа была закончена и сдана в Издательство, автор познакомился со статьей Пономаревой (1951), цитируемой при описании Clupea harengus harengus и Clupea harengus pallasi. В этой статье Л. А. Пономарева рассматривает многопозвонковых и малопозвонковых сельдей за самостоятельные виды — Clupea harengus и Clupea pallasi, а все формы — за подвиды: Clupea harengus membras, Clupea pallasi maris-albi и Clupea pallasi suworowi. Однако все приводимые ею в пользу этого доводы основаны на малой компетенции в рассматриваемом вопросе и на недостаточном знакомстве с литературой. Л. А. Пономарева, считая число позвонков важнейшим признаком, положенным в основу систематики рода Clupea, утверждает, что «по этому признаку многопозвонковые или океанические и малопозвонковые или восточные сельди всегда резко различны» и в качестве «наглядной иллюстрации» приводит кривые числа позвонков (рис. 3). Однако даже на этом рисунке кривые числа позвонков атлантической сельди и малопозвонковых — тихоокеанской и ее форм явно трансгрессивны. Нельзя не отметить несоответствия между количеством позвонков на этих кривых и в описаниях. Так, у атлантической сельди по кривой позвонков 55—59, а в описании 53—55, у чёшской сельди по кривой позвонков 52—56, в описании 54, в среднем: 53.5 и т. д. Если же принять во внимание пределы колебаний количества позвонков, известное по литературным данным, то в действительности мы имеем дело не с трансгрессией, а почти с полным совпадением, так как у тихоокеанской сельди количество позвонков колеблется от 47 до 57, а у атлантической сельди, если принять во внимание все ее формы, а не одну лишь мурманскую, — от 51 до 60. У атлантической сельди даже средние числа позвонков у разных рас колеблются от 54.7 до 57.5, между тем как на кривой пределы колебаний указаны от 55 до 59, что свойственно лишь мурманской сельди. Другим признаком, по которому многопозвонковые и малопозвонковые сельди резко различаются, Л. А. Пономарева считает развитие зубов на сошнике. В подтверждение Л. А. Пономарева ссылается на П. Ю. Шмидта и Л. С. Берга, которые однако считают обеих этих сельдей лишь за подвиды. Со ссылкой на стр. 182 на рисунок «сошников сельдей группы harengus и группы pallasi, наглядно подтверждающий справедливость указаний, упомянутых автором», получилось явное недоразумение, так как этого рисунка в статье нет. На этом рисунке изображена карта распространения атлантической сельди. Что касается различий в числе поперечных рядов чешуй (перед спинным плавником), по которому «всегда легко отличить группу harengus от группы pallasi», то число их, как видно из табл. 3, также трансгрессивно: у форм первой колеблется от 13 до 15, у второй — от 9 до 13 (на стр. 189 от 10 до 13). Относительно различий в числе сегментов у личинок сказано выше. Следует лишь отметить, что в этой части статьи имеется также ряд ошибочных утверждений. Так, личинки беломорской сельди, по данным Каринского (1938), имеют не 53 сегмента, а 53 и более, анальное отверстие у них находится не на 43—44 сегменте, а на 42—45 и т. д.
Распространение. Северные части Атлантического и Тихого океанов и прилегающие моря. В Атлантическом океане у берегов Европы от Бискайского зал. до Шпицбергена, Новой Земли, в Балтийском, Белом и Карском морях, у берегов Сев. Америки от м. Гаттераса до Девисова прол. и Гренландии. В Тихом океане у берегов Азии от западного побережья Кореи в Желтом море и зал. Тояма в Японском, на север до Анадырского зал., в бухте Тикси и Оленёкском зал. у берегов Сибири, у берегов Сев. Америки от Калифорнии до Аляски (рис. 14).
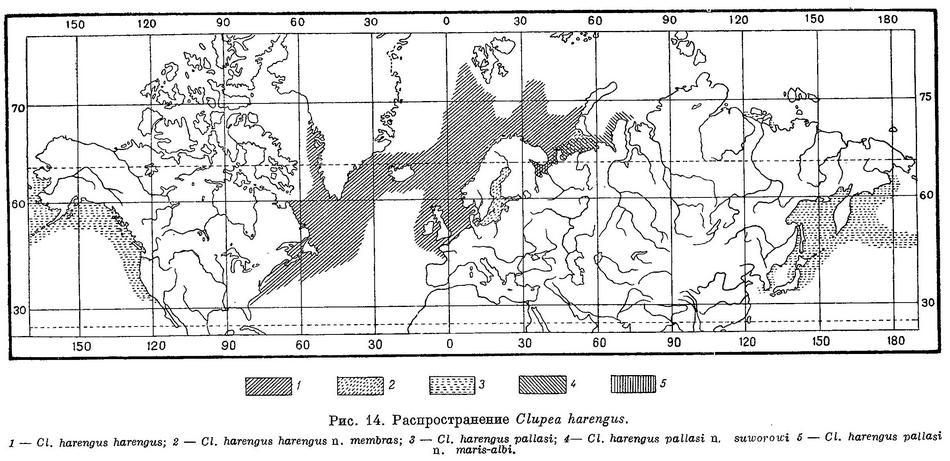
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
1 (4). Число позвонков 51—60, наичаще 55—57. Килевые чешуи более или менее развиты как позади, так и впереди основания брюшных плавников. Между брюшными и анальным плавниками 12—16 килевых чешуй. Зубы на сошнике сравнительно хорошо развиты. Северная часть Атлантического океана и прилегающие моря.
2 (3). Позвонков в среднем от 54.8 до 57.5. Грудные плавники короткие, менее 14% длины тела. Голова короткая и широкая, длина ее менее 23% длины тела, межглазничный промежуток более 16.5% длины головы. Глаза небольшие, менее 24% длины головы. Северная часть Атлантического океана и прилегающие моря — Северное, Баренцово и отчасти Белое. *la. Clupea harengus harengus L.
3 (2). Позвонков меньше, в среднем от 55.2 до 55.7. Грудные плавники
длиннее, не менее 13.5% длины тела. Голова длиннее и уже, длина ее более 23% длины тела, межглазничный промежуток менее 17.5% длины головы. Балтийское море. *lb. Clupea harengus harengus n. membras L.
4 (1). Число позвонков 49—57, наичаще 52—55. Килевые чешуи более или менее развиты лишь позади брюшных плавников. Между брюшными и анальным плавниками 10—14 килевых чешуй. Зубы на сошнике менее развиты. Северная часть Тихого океана и арктические моря.
5 (6). Голова короткая, менее 21% длины тела. Грудные плавники менее 14.5% длины тела. Глаза менее 23% длины головы. Крупные рыбы, в половозрелом состоянии длиной от 20 до 40 см и более. Северная часть Тихого океана и прилегающие моря. *lc. Clupea harengus pallasi Val.
6 (5). Голова длиннее, более 21% длины тела. Грудные плавники более 13% длины тела. Глаза более 23% длины головы. Размеры меньше, до 30—34 см, в половозрелом состоянии — от 10—20 см. Белое море, Мезенский зал., Чешская губа, Карское море на восток до Обской губы.
7 (8). Высота головы более 13% длины тела. Берг (1948 : 148) в определительной таблице в качестве отличительного признака между Clupea harengus pallasi n. maris-albi и Clupea harengus pallasi n. suworowi указывает длину основания анального плавника, меньшую у первого (около 10.5% длины тела), чем у второго (более 11%). Данные эти взяты из работ разных авторов. По нашим данным, длина основания анального плавника больше у Clupea harengus pallasi n. maris-albi (10.9%), чем у Clupea harengus pallasi n. suworowi (10.4%). Белое море. *ld. Clupea harengus pallasi n. maris-albi Berg
8 (7). Высота головы менее 14% длины тела. Мезенский зал., Чёшская губа, Карское море. *1е. Clupea harengus pallasi n. suworowi Rabinerson.
*la. Clupea harengus harengus L. — Атлантическая сельдь, многопозвонковая сельдь, также мурманская сельдь, норвежская сельдь и другие, чисто местные названия (табл. II).
— Clupea harengus Linnaeus, Syst. nat., ed. X, 1758 : 317 (Атлантический океан у берегов Европы).
— Cyprinus esca Wаlbaum, in: Artedi, Gen. pisc., 1792 : 36 (white bait, ex Pennant, Brit, zool., III, 1776 : 371, t. 69).
— Clupea halec Mitchill, Trans. Lit. a. Philos. Soc., I, 1815 : 451 (Нью-Йорк).
— Clupea vittata Mitchill, 1. c. : 456 (Нью-Йорк).
— Clupea caerulea (non Girard) Mitchill, l. c. : 457 (Нью-Йорк).
— Clupea elongata Le Sueur, Journ. Acad. Nat, Sci. Phila., I, 1817 : 234 (Массачузетс).
— Clupea latulus Сuvier, Regne animal, ed. II, II, 1829 : 318.
— Clupea alba Yarrell, Zool. Journ., IV, 1829 : 137, 465, pl. V, f. 2.
— Clupea leachii Yarrell, ibid., V, (1830), 1832 : 277, pl. XII (Англия).
— Clupea minima (Peck) Storer, Rept. fish. Mass., 1839 : 113 (Нью-Гемпшир).
— Clupea lineolata Cuvier et Valenciennes, Hist. nat. poiss., XX, 1847 : 256.
— Rogenia alba Cuvier et Valenciennes, l. c. : 341, pl. 508.
— Clupea harengus Smitt, Scand. fish., II, 1895 : 954 (частью); Heincke, Naturgesh. d. Herings, I, 1898 : VII, Tab. I, III—IV, VI—LXV, LXX— CXLI (частью: европейские моря; измерения рыб и черепов, расы).
— Clupea harengus harengus Берг, Рыбы пресных вод России, 1923 : 500 (Кольский зал.); Rabinerson, Zool. Jahrb., Abt. Syst., B.d. 58, 1929 : 294 (Мурманское побережье); Schnakenbeck, Zeitschr. f. Morphol. u. Okol. d. Tiere, Bd. 21, 1931 : 525, 529, f. 10, 16, 17, 19, 20 (банковские сельди Северного моря).
— Clupea harengus britannicus Schnakenbeck, l. c. : 526, 529, f. 11, 20, 23 В (сельди Ламанша).
— Clupea harengus atlanticus Schnakenbeck, l. c. : 526, 529, f. 33 (сельди южного входа в Ирландское море).
— Clupea harengus ivernicus Schnakenbeck, 1. с. : 526, 529, f. 39 А (клайдские сельди).
— Clupea harengus caledonicus Schnakenbeck, l. c. : 527, 529 (сельди западного побережья Шотландии).
— Clupea harengus scoticus Schnakenbeck, l. c.: 527, 529, f. 18 (зал. Фёрс оф Форс).
— Clupea harengus frisicus Schnakenbeck, l. c. : 528, 529, f.. 34, 35, 36 (зал. Зюдерзее).
— Clupea harengus cimbricus Schnakenbeck, l. c. : 259 (Ютландская банка).
— Clupea harengus scandicus Schnakenbeck, l. c. : 529 (норвежская сельдь, форма A).
— Clupea harengus norvegicus Schnakenbeck, l. c. : 529 (норвежская сельдь, форма В).
— Clupea harengus septemtrionalis Schnakenbeck, l. c. : 529 (фарерские сельди).
— Clupea harengus borealis Schnakenbeck, l. c. : 529 (весенняя сельдь Исландии).
— Clupea harengus islandicus Schnakenbeck, l. c.: 529 (летняя сельдь Исландии).
— Clupea harengus Lissner, Journ. Conseil, IX, 3, 1934 : 346, 364 (частью: атлантическая сельдь, разделение на два подвида — весенних и осенних сельдей); Lе Gall, Ann. Inst, oceanogr., XV, 1, 1935 : 19, 99, 155, 162—165 (частью: восточная половина северной части Атлантического океана; расы).
— Clupea harengus harengus n. murmanica Тихонов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 7, 1941 : 26 (Баренцово море).
—Clupea harengus harengus Расс, в : Промысловые рыбы СССР, 1949 : 120, табл. 30.
— Clupea harengus Пономарева, Учен. зап. Горьк. Гос. унив., XIX, 1951 : 177, рис. 1.
Распространение, биология. Fulton, 9th Ann. Rept. Fish. Board. Scotl., III, 1891 : 267 (число икринок); Apstein, Schrift. Naturwissensch. Ver. Schleswig-Holst., XI, 2, 1898 : 222 (p. Шлей; нерест, плодовитость); Hinkelmann, Mitt. Deutsch. Seefischerei-Ver., XIV, 2, 1898 : 45; ibid., XV, 6, 1899; ibid., XVIII, 1, 7, 12, 1902 : 15, 362, 606 (Кильский канал; нерест); Jenkins, Wiss. Meeresunters. (N. F.), Abt. Kiel, VI, 1902 : 83 (плодовитость); Ehrenbaum, Fauna Arct., II, 1902 : 138 (Тромсе и Сёре-Сунд, Трольд-фиорд; личинки); Hinkelmann, Mitt. Deutsch. Seefischerei-Ver., XIX, 3, 1903 : 88 (Кильский канал; нерест); Нjоrt, Rapp. Proces-Verb., XX, 1914 : 14 (колебания уловов); Lеbour, Journ. Mar. Biol. Assoc. (n.s.), XII, 2, 1918 : 36 (питание личинок); Johansen, Meddel. Itomm. Havundersog., Fisk., V, 8, 1919 : 1 (Исландия, Фарерские о-ва, Норвегия; биометрика); Jespersen, ibid., VI, 1, 1920 : 4 (Исландия, Фарерские о-ва; личинки); Johansen, ibid., VI, 3, 1921 : 1 (Исландия; летняя сельдь, биометрика, размножение); ibid., VI, 4, 1921 : 1 (Фарерские о-ва; весенняя сельдь, биометрика, размножение); Lеbour, Journ. Mar. Biol. Assoc. (n. s.), XII, 3, 1921 : 278, 459 (питание личинок); ibid., XII, 4, 1922 : 644; ibid., XIII, 1, 1923 : 70 (значение личинок в питании планктонных организмов); ibid., XIII, 2, 1924 : 325, 402 (вертикальное распределение и питание личинок); Le Danois et Held, Notes et Mem. l'Office Sci. Techn. Peches Marit., 36, 1924 : 1 (южная часть Ирландского моря; расы); Johansen, Wiss. Meeresunters. (N. P.), Abt. Helg., XV, 1923—1924 : 1 (Каттегат; зимние сельди, расы, места и время нереста); Meddel. Komm. Havundersog., Fisk., VII, 5, 1924 : 1 (Ламанш, Северное море, датские проливы; расы и места нереста); Bigelow a. Welsh, Fish, gulf Maine, 1925 : 92 (берега Сев. Америки; места и время нереста, биология); Рабинерсон, Изв. Отд. прикл. ихт., III, 1, 1925 : 67; там же, IV, 1, 1926 : 121 (Кольский зал.; рост, отношение к беломорской и мурманской сельди); Johansen, Rapp. Proces-Verb., XXXIX, 1926 : 114—138 (берега Исландии; расовый анализ, нерест весенней и летней сельди); Lе Gall, No Lea et Mem. l'Office Sci. Techn. Peches Marit., 48, 1926 : 1 (берега Ирландии); Rapp. Proces-Verb., XLIV, 1927 : 115 (Ламанш; рост, зрелость); Johansen, Rept. Danish Biol. St., XXXIII, 1927 : 17 (прол. Б. Бельт; личинки зимних сельдей в начале марта, осенние сельди мечут икру в сентябре и октябре); Journ. Conseil., II, 1, 1927 : 3 (берега Норвегии, Исландии и Фарерских о-вов, Северное м., прол. Зунд; места нереста, миграции); Аверинцев, Тр. Инст. рыбн. хоз., II, 1, 1927 : 66 (Белое м.: Кандалакшский зал., близ устья р. Онеги, Двинский зал.); Lissnеr, Ber. Deutsh. Wiss. Komm. Meeresforsch. (N. F.), III, 2, 1927 : 47 (западпая часть Северного м.; расы); Schnakenbeck, ibid. : 95 (Северное м.; расы); Stоrrоw, Dove Mar. Labor., Cullercoat, Northumb. (n. s.), XVI, 1927 : 9; XVII, 1929 : 9; XVIII, 1929 : 9; XIX, 1931 : 9; XX, 1931 : 8; XXI, 1932 : 11 (размеры, возраст, рост, колебание численности возрастных групп, зрелость, нерест); Рабинерсон, Тр. Инст. изуч. Севера, 38, 1927 : 64 (рост); Карело-Мурманск. край, 1928, 10—11 : 47 (отношение мурманской сельди к норвежской); Макушок, Русск. зоолог. журн., VIII, 3, 1928 : 39; там же, VIII, 4, 1928 : 41 (отношение мурманской сельди к норвежской); Hodgson, Journ. Conseil, III, 2, 1928 : 224 (южная часть Северного м.; места нереста по икринкам из желудков пикши); Ford, Journ. Mar. Biol. Assoc. (n. s.), XV, 1,1928 : 237, 267, 279, 305; ibid., XVI, 1, 1929 : 1, 25 (Ламанш; количество позвонков, возрастной состав); Johansen, Rept. Danish Biol. St., XXXV, 1929 : 5 (Лимфиорд; весенняя и осенняя сельди); Lea, Journ. Conseil., IV, 1, 1929 : 1 (океаническая стадия сельди); Rapp. Proces-Verb., LIV, 1929 : 21 (годичные кольца на чешуе в связи с образом жизни); Мiеlсk, ibid. : 99 (Северное м.; личинки, ноябрь—апрель); Schnakenbeck, Ber. Deutsch. Wiss. Komm. Meeresforsch. (N. F.), V. 2, 1929 : 23 (развитие личинок); Петров, Изв. Гос. Инст. опытн. агроном., VII, 6, 1929 : 689 (отношение мурманской сельди к норвежской); Бородатова, Тр. Научн. инст. рыбн. хоз., V, 4, 1930 : 47 (отличие мурманской сельди от норвежской, личинки из Ура-губа); Lе Gall, Rap. Proces-Verb., LXIX, 1930 : 1 (сельди Ламанша и южной части Северного м.); Wood, Scient. Invest., Fish. Board Scotl., 1930, I : 1 (места и время размножения сельдей у берегов Шотландии); Schnakenbeck, Zool. Anz., 94, 1931 : 21 (р. Эльба у Гамбурга); Петров, Тр. Инст. изуч. Севера, 48, 1931 : 103; Суворов, Вадова и Сынкова, там же : 150 (отношение мурманской сельди к норвежской); Макушок, Тр. Гос. Океаногр. инст., I, 4, 1932 : 3 (отношение мурманской сельди к норвежской); Танасийчук, Карело-мурманск. край, 1932, 7—8 : 40 (отношение мурманской сельди к норвежской); Вадова, Сборн. научно-промысл. работ на Мурмане, 1932 : 159 (отношение мурманской сельди к норвежской); Poulsen, Meddel. Komm. Danmarks Fiskeri- og Havunders., Fisk., IX, 4, 1932 : 1 (Северное м., Скагеррак, Каттегат; места нераста); Watkin, Rapp. Proces-Verb., LXXXIV, 1933 : 41 (сельди южного входа в Ирландское м., зрелость, размеры, возраст, рост); Clark, ibid., LXXXV, 3, 1933 : 11 (берега Шотландии, Гебридских, Оркнейских и Шетландских о-вов; личинки); Ford, Journ. Mar. Biol. Assoc. (n. s.), XIX, 5, 1933 : 305 (Ламанш; места и время нереста, рост, расы); Runnström, Journ. Conseil, VIII, 2, 1933 : 235. (расовая неоднородность, сельди у берегов Норвегии); Макушок, За рыбн. индустр. Севера, 1933, 8 : 20 (западное побережье Канина: устье pp. Кии, Шойны и Торны); там же, 1934, 8 : 33. (восточное побережье Канина: устье р. Сев. Камбальницы; устье р. Индиги, открытая часть Чёшской губы); Saemundsson, Rapp. Proces-Verb., LXXXVI, 1934 : 4 (нерест у берегов Исландии); Ottestad, ibid., LXXXVIII, 1934 : 3 (берега Норвегии; северная и южная формы по темпу роста); Runnström, ibid., LXXXVIII, 1934 : 3 (берега Норвегии; личинки, места нереста); Clark, ibid., LXXXIX, 2, 1934 : 23 (Фарерские о-ва; личинки в мае и июне); Wood, ibid., LXXXIX, 3, 1934 : 57 (северная часть Северного м.; расы); Storrow a. Gowan, Dove Mar. Labor., Cullercoat, Northumb. (3), 2, 1934 : 9; 3, 1935 : 11; 4, 1938 : 10 (размеры, возраст, рост, колебание численности возрастных групп, зрелость, нерест); Awerinzew, Journ. Conseil, X, 1, 1935 : 66 (отношение мурманской сельди к норвежской); Нansen, Beretning, Vestgrønlands Sryrelse, 2, 1935 : 16 (Гренландия; расовый состав, нерест); Runnström, Rept. Norw. Fisher. Mar. Invest., V, 2, 1936 : 5 (строение годичных колец — кольца южного и северного типов, океанические и нерестовые); Tåning, Meddel. Komm, Danmarks Fiskeri- og Havunders., Fisk., X, 3, 1936 : 8 (Фарерские о-ва; личинки, время нереста весенних и летних сельдей); Wood, Scient. Invest., Fish. Board Scotl., 1936, III, 1 (расовый анализ весенних и осенних сельдей Шотландии); Tåning, Rapp. Proces-Verb., XCIX, 1936 : 1 (Исландия; личинки, места нереста); Andersson, ibid., С, 2, 1936 : 11 (Скагеррак, Каттегат, Зуид; расовый состав, нерест); Роulson, ibid. : 20 (Скагеррак, Каттегат, Зунд, Б. Бельт и западная часть Балтийского м.; расы и места нереста); Runnström, ibid. : 24 (атлантическо-скандинавские сельди; распространение); Tåning, ibid. : 32 (Фарерские о-ва, Исландия, Гренландия; расы и нерест); Агапов и Топорков, Пробл. Арктики, 1937, 2 : 104 (губы Грибова и Белушья, западное побережье Новой Земли); Marshall, Nicholls а. Оrr, Journ. Mar. Biol. Assoc. (n. s.), XXII, 1, 1937 : 245 (зал. p. Клайд; рост и питание личинок); Runnström, Journ. Conseil, XII, 2,1937 : 123 (различия между сельдями с разных нерестилищ у берегов Норвегии); Fаrran, ibid., XIII, 2, 1938 : 91 (размеры и количество икринок у сельди от берегов Ирландии); Еrdmаnn, Rapp. Proces-Verb., CVII, 3, 1938 : 31 (расовый анализ весенних сельдей Шотландии); Рыженко, Рыбн. хоз. СССР, 1938, 2 : 12 (личинки от Лофотенских о-вов до Мотовского зал., мальки в Мотовском и Кольском зал. и у Териберки, сеголетки на Гусиной и Канинской банках, отход их от берегов в открытое море); Юдкин, Тр. Моск. технич. инст. рыбн. хоз., 1, 1938 : 59 (Пала-губа, Териберка, Титовка; мальки); Глебов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 1, 1938 : 5 (условия захода летней и жирной сельди в губы); Карантонис, там же : 59 (строение и сочленение костей жаберной крышки и плечевого пояса, три «типа» сельдей); Мурашкинцева, там же : 74 (типы зимних колец на чешуе); Мантейфель и Болдовский, там же : 88 (причины заходов жирной сельди в губы); Болдовский, там же : 102 (питание в губах); Алеев, там же : 120 (распределение в Баренцевом море по нахождениям в желудках трески); Есипов, там же : 160 (библиография работ о мурманской сельди до 1937 г.); Коtthaus, Wiss. Meeresunters., Abt. Helg., I, 3, 1939 : 349; Sсhасh, ibid. : 359 (искусственное оплодотворение икринок, выращивание личинок, питание их); Rеdеkе, Journ. Conseil, XIV, 3, 1939 : 337 (Зюдерзее, исчезновение сельди после отделения его от моря); Jensen, Kgl. Danske Videnskab. Selsk., Biol. Meddel., XIV, 8, 1939 : 14 (у западных берегов Гренландии на север до 72.5° с.ш., у восточных — в Датском прол.; нерест и выметанные икринки в августе); Рыженко, Рыбн. хоз., 1939, 4 : 20 (Чёшская и Индигская губы, устья Индиги и Сев. Камбальницы, летом); Наумов, там же, 11 : 34 (одновременность икрометания); Рыженко, Сборн., посвящ. Книповичу, 1939 : 394 (личинки и мальки мурманской сельди); Марти, Сборн. «Биология и пром. Мурманск. сельди», 1939 : 54 (распределение в поверхностных слоях воды); Болдовский, там же : 41 (пищевые конкуренты); Тихонов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 4, 1939 : 5 (распределение в южной части Баренцова м. зимой); Мантейфель и Марти, там же : 41 (места и условия нереста мурманской сельди); Юданова, там же : 95 (содержание жира); Пчелкина, там же : 109 (распределение в связи с составом зоопланктона); Есипов, там же : 149 (литературная сводка по размножению у берегов Европы); Расс, там же, 6, 1939 : 93 (размножение, биология молоди); Дмитриев, Рыбн. хоз., 1940, 10 : 26 (Белое м.); Тихонов Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., 7, 1941 :3 (систематическое положение мурманской сельди); Марти, там же: 38 (жизненный цикл мурманской, сельди); Мантейфель, там же : 125 (распределение в связи с планктоном); Болдовский, там же : 219 (питание в море); Sund, Ann. Biol., I, (1939—1941), 1943 : 45 (возрастной состав уловов норвежской нерестовой сельди с 1907 по 1942 г.); Марти, Рыбн. промышл. СССР, 3, 1945 : 3 (современное состояние промысла); Дмитриев, биолог. и пром. сельди в Белом м., изд. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., 1946 : 41 (Белое м.); Тамбовцев, Рыбн. хоз., 1947, 6 : 30 (Белое м., западное побережье п-ова Канина); там же, 1949, 4 : 48 (Белое м., многочисленные заходы в 1942, 1944, 1946 и 1948 гг.); Rasmussen, Fiskeridirekt. Skrift., Havunders., IX, 7, 1950 : 3 (идентичность норвежской и исландской сельди); Fridriksson a. Aasen, ibid., IX, 11, 1950 : 7 (мечение, миграции между Норвегией и северной Исландией).
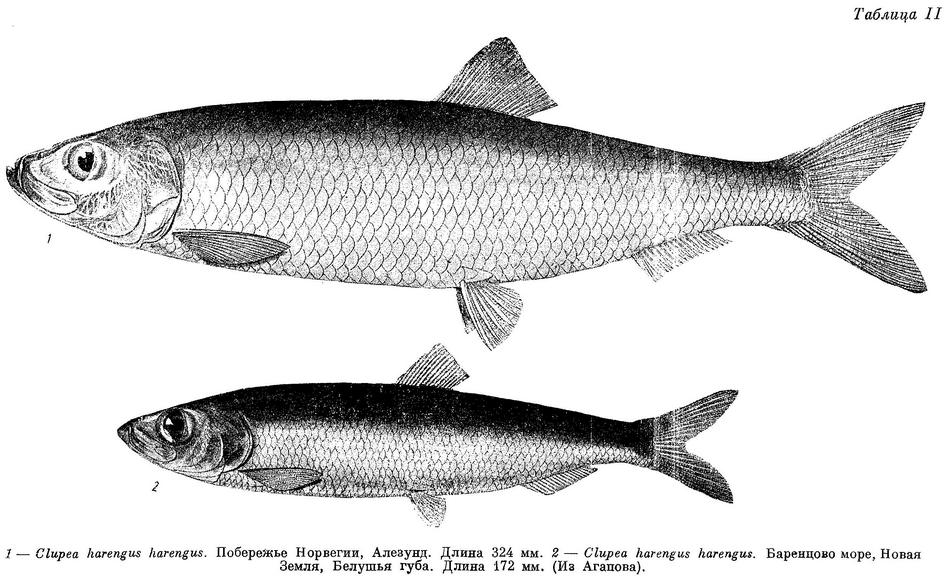
D (III) IV 14—17 (15.0), общим числом 17—21 (18.9), А III 13—17 (15.0), общим числом 16—20 (18.0), sp. br. 66—73 (70.1), килевых чешуй позади основания брюшных плавников 13—16 (14.3), vert. S. 56—59 (57.06), caud. vert. 31—34 (32.57). Брюшные килевые чешуи развиты как позади основания брюшных плавников, так и впереди от них. Зубы на сошнике сравнительно хорошо развиты. Голова укороченная и широкая, длина ее 20.2—22.8 (21.1)% длины тела, межглазничный промежуток 16.6—19.3 (17.7)% длины головы. Глаза небольшие, 20.8—23.9 (22.2)% длины головы. Основание анального плавника 9.0—11.1 (10.0)% длины тела. Длина до 36 см, у экземпляров от берегов Исландии — до 42 см. Описание дано по 38 экз. длиной от 28.9 до 34.4, в среднем 32.5 см, собранным 17 III 1947 у берегов Норвегии (Алезунд). По Гейнке (Heincke, 1898), позвонков 51—60, килевых чешуй 11—17; по Ле Галю (Le Gall, 1935), средние числа позвонков у форм подвида колеблются от 54.8 до 57.5, килевых чешуй от 13.3 до 15.6; по Тихонову (1941), у мурманской сельди позвонков 55—59, средние за ряд лет колеблются от 57.15 до 57.35.
Морская форма.
В пределах обширного ареала распадается на ряд локальных форм (рас, популяций), различающихся некоторыми морфологическими (количеством позвонков, килевых чешуй, размерами) и биологическими особенностями, в частности местами и временем нереста. По времени нереста среди форм атлантической сельди различают весенних, летних, осенних и зимних, из которых наиболее многочисленны весенние и осенние. Начало детальному систематическому изучению рас атлантической сельди в конце прошлого столетия положил Гейнке (Heincke, 1898), давший классификацию их для европейской части Атлантического океана. В дальнейшем исследования многих авторов способствовали более углубленному изучению рас сельди. На основе этих исследований новую классификацию их предложил Ле Галь (Le Gall, 1935), разделивший сельдей восточной половины Атлантического океана на следующие группы, среди которых он различает ряд популяций (= расам или локальным формам Heincke).
I. Атлантическо-скандинавские весенние сельди, куда относятся сельди Исландии, Фарерских о-вов, склонов; континентального плато Северного моря, Норвегии. К ним относится норвежская и наша мурманская сельдь. Средние числа позвонков выше 57.0 (от 57.0 до 57.5), килевых чешуй в среднем от 13.3 до 14.1. Характерны быстрым ростом и крупными размерами в половозрелом состоянии. Икрометание происходит весной при температуре воды от 4 до 8° и солености от 34.5 до 35.3‰. Атлантическо-скандинавские весенние сельди, судя по всему, есть и у берегов Сев. Америки, где, кроме того, имеются летние и осенние сельди; наибольшее значение здесь имеют весенние и осенние сельди, которые детально еще не изучены.
II. Зимние шотландские сельди — сельди северозападного побережья Шотландии, северного и западного побережья
Ирландии и края кельхийского континентального плато (восточного побережья Ирландии). Средние числа позвонков приближаются к 57.0 (56.9—57.0), килевых чешуй в среднем от 13.8 до 14.3. Характерны также быстрым ростом и крупными размерами. Икрометание в конце зимы и весной при температуре воды от 7 до 9° и солености от 34.6 до 35.3‰. К этой же группе провизорно относятся близкие по всем признакам, но размножающиеся позднее летние сельди Исландии и Фарерских о-вов. Сюда же, судя по всему, относится и летняя сельдь Гренландии.
III. Сельди Ламанша встречаются по всему проливу и в южной части Северного моря и представляют смешанную группу. Средние числа позвонков ниже 57.0 и выше 56.5 (от 56.6 до 56.8), килевых чешуй в среднем от 14.3 до 15.0. Растут медленнее и менее крупны, чем сельди предыдущих двух групп, половой зрелости достигают при длине около 25 см. Размножаются осенью и зимой при температуре воды от 7.5 до 12.5° и солености от 33.5 до 35.0‰.
IV. Сельди южного входа в Ирландское море (Smalls herring) близки к сельдям предыдущей группы. Среднее число, позвонков менее 57.0 (от 56.6 до 56.9), килевых чешуй 14.1. По темпу роста более близки к сельдям двух первых групп, чем к сельдям Ламанша. Нерестуют в зимне-весеннее время. Места, время и условия нереста точно, не установлены.
V. Осенние сельди Северного моря (банковские сельди) встречаются по всему Северному морю и временами в восточной части Ламанша. Среднее число позвонков близко к 56.5 (от 56.3 до 56.5), килевых чешуй в среднем от 14.75 до 15.25. Рост еще более медленный, чем у сельдей Ламанша. Размножаются осенью и в начале зимы при температуре воды от 9 до 13° и солености от 34 до 35‰.
VI. Сельди датских проливов представляют смешанный комплекс популяций, промежуточных по своим признакам между предыдущими группами и балтийской сельдью. Среди этих сельдей различаются следующие. 1) Зимние сельди Каттегата. Среднее число позвонков близко к 57.0 (от 56.75 до 56.90), среднее число килевых чешуй от 14.1 до 14.8. Размеры небольшие, значительно меньше сельдей двух первых групп того же возраста. Близки к атлантическо-скандинавским весенним сельдям. Нерестуют в конце зимы при низкой температуре (4—5°) и солености воды (29—34‰). 2) Осенние сельди северо-восточной части Каттегата. Среднее число позвонков немного менее 56.5 (от 56.1 до 56,4), килевых чешуй в среднем от 14.40 до 14.65. Размеры в среднем около 25 см. Близки к прибрежным сельдям Северного моря. Размножаются осенью при сравнительно высокой температуре (11—13°) и пониженной солености (29—34‰). 3) Осенние сельди южной части Каттегата и Зунда. Количество позвонков небольшое, близкое к числу их у балтийской сельди, в среднем около 55.5, килевых чешуй в среднем 14.2—14.5. Мелкие сельди. Икрометание осенью при высокой температуре воды (12—14°) и низкой солености (8—22‰). 4) Весенние сельди Скагеррака, Каттегата и Бельта. Среднее число позвонков близко к 56.0 (от 55.9 до 56.2), килевых чешуй в среднем 14.0—14.5. Мелкие сельди, размножающиеся весной при температуре воды от 6 до 12° и солености от 8 до 24‰.
VII. Сельди Зюдерзее — солоноватоводного залива Северного моря, образовавшегося в исторический период из пресноводного озера, отделенного от моря узким перешейком. Позвонков в среднем: от 54.8 до 55.3. Размеры небольшие, темп роста замедленный. Близки к балтийской сельди. Нерест весной при температуре воды от 7 до 13° и слабой солености — от 9 до 11‰. В 1932 г. залив был отделен от моря плотиной и превратился вновь почти в пресноводное озеро, в котором сельдь уже в 1933 г. была поймана в ничтожном количестве, а в 1934 г. исчезла совершенно.
Под названием атлантических сельдей Ле Галь объединяет сельдей первых двух групп (атлантическо-скандинавских весенних и зимних шотландских сельдей), живущих вблизи края континентального плато, для которых характерно среднее число позвонков, большее или близкое к 57.0, крупные размеры и быстрый темп роста, весенний нерест. В более обширную группу прибрежных сельдей им объединены следующие три группы (III—V), живущие ближе к берегам. Для них характерно среднее число позвонков ниже 57.0, средние размеры и более замедленный темп роста, осенний или зимний нерест. Под названием сельдей солоноватых вод Ле Галь объединяет сельдей Зюдерзее, а также фиордовых сельдей Норвегии и Швеции, сельдей дельт рек Эмс, Яд и Морле, для которых характерно среднее число позвонков меньше 56.2, мелкие размеры и замедленный темп роста, весенний нерест при пониженной солености. К этим сельдям относятся и балтийские сельди, рассматриваемые здесь в качестве особого подвида.
Ряд популяций Ле Галя и несколько более мелких подразделений внутри них, считая их за подвиды и дав им латинские наименования, охарактеризовал Шнакенбек (Schnakenbeck, 1931). Все подвиды Шнакенбека целиком включены в классификацию Ле Галя. Что касается подвидового значения подразделений Шнакенбека, то подвидом является сама атлантическая сельдь, и, следовательно, им следует придавать меньшее систематическое значение, считая их за nationes, каковые для атлантической сельди при современном состоянии изученности ее, по нашему мнению, выделять не следует.
Лисснер (Lissner, 1934) разделяет атлантических сельдей на два более крупных подразделения, предлагая считать их подвидами: 1) весенних сельдей, к которым он провизорно относит и зимних, и 2) осенних сельдей. Немногочисленные летние сельди, как указывалось выше, относятся к зимним, т. е. также к весенним сельдям.
Мурманская сельдь до сравнительно недавнего времени, по одним взглядам (Рабинерсон, 1926, 1927, 1928; Петров, 1929, 1931; Суворов, Вадова и Сынкова, 1931; Вадова, 1932; Аверинцев, 1935 и др. ), рассматривалась как неполовозрелая норвежская сельдь, заходящая в Баренцово море в молодом возрасте, по другим (Макушок, 1928, 1932; Бородатова, 1930; Танасийчук, 1932; Юдкин, 1938) — считалась отличной от нее. Позднее благодаря работам ряда советских исследователей, в особенности сотрудников Полярного научно-исследовательского института морского рыбного хозяйства и океанографии (Рыженко, 1938, 1939; Мантейфель и Марти, 1939; Расс, 1939; Марти, 1941), а также Рунстрем (Runnström, 1933, 1934, 1936, 1937), установлены места нереста мурманской сельди, распространение ее в Баренцовом море и отношение к норвежской. На основании сравнительно небольших отличий от последней мурманская сельдь была выделена как natio murmanica, что вряд ли является целесообразным, так как за natio следовало бы считать, если вообще нужно давать латинские названия этим группам, атлантическо-скандинавских весенних сельдей, а мурманская, норвежская и другие сельди этой группы систематически стоят ниже natio, и кроме того, эту сельдь, судя по всему, уже описал Шнакенбек. По исследованиям Карантониса (1938), сельдь, ловимая в Баренцовом море, по строению и сочленению костей жаберной крышки и плечевого пояса, разделяется на три так им называемых типа. Сельдь III типа предположительно относилась им к норвежской сельди, сельди II и I типов — к мурманской, причем сельдь I типа, согласно первоначальным исследованиям распространенная только в прибрежных частях Баренцева моря, принималась за мурманскую фиордовую сельдь, сельдь II типа, встречаемая как в прибрежных, так и в открытых частях моря, — за мурманскую океаническую. В дальнейшем было обнаружено, что сельдь I типа имеет такое же широкое распространение в открытых частях моря, как и сельдь II типа. Одновременно было констатировано, что различия между сельдями обоих этих типов в строении operculum (наличие или отсутствие выемки на заднем крае этой кости) обусловлены возрастной изменчивостью. Не было найдено реальных различий между сельдями этих двух типов и в счетных и пластических признаках, а также и в темпе роста (Тихонов, 1941). Несомненно в сильной степени варьирует и признак отличия норвежской сельди от мурманской (сочленение костей жаберной крышки).
Распространение. Северная часть Атлантического океана и прилегающие моря. На юг у берегов Сев. Америки до мыса Гаттераса, на север — в Девисовом прол., у восточных (до 66—67° с. ш.) и западных (приблизительно до 72.5° с. ш.) берегов Гренландии, у западных берегов Шпицбергена (до 77—78° с. ш.) и Новой Земли (до широты Маточкина Шара), на восток в Баренцовом море до 50—52° в. д., вокруг Исландии, Фаррерских, Шетландских и Оркнейских о-вов, Великобритании и Ирландии, в Скагерраке и Каттегате, в западной части Балтийского моря до о. Эланд — Данцигская бухта, на юг у берегов Европы до Бискайского зал. Заходит в Белое море (Терский берег, Кандалакшский, Онежский, Двинский заливы, Соловецкие о-ва), Мезенский зал. и летом в юго-восточную часть Баренцова моря к востоку от о. Колгуева, в Чёшскую и Индигскую губы, в западную часть Печорского зал. и в устья рек (Индиги, Сев. Камбальницы и др. (В р. Эльбе отмечена у Гамбурга (Schnakenbeck, 1931).)).
Нерест атлантическо-скандинавской весенней сельди происходит: 1) вдоль берегов северной Норвегии к северу от Вест-фиорда, главным образом вблизи о. Рёст, у западных берегов о-вов Лофотенских и Вестеролен — на банках против о-вов Вествогей, Лангей, Андей, Квалей, Рингвассе и фиорда Маланген (мурманская сельдь, с начала февраля до середины мая, основной нерест в конце февраля до середины апреля); к северу от Тромсе, вдоль Финмаркена и Мурмана, нерест, судя по имеющимся данным, происходит в очень малых размерах в Мотовском (Малая Лица, Вичанское озерко) и Кольском заливах (губы Полярная Гавань, Варламова, Сайда, Ваенга, Рослякова), губе Орловка (у Териберки), где найдены только личинки, предположительно относимые к мурманской фиордовой сельди (Мурманская фиордовая сельдь изучена севершенно недостаточно; единого мнения о реальности ее существования нет (Расе, 1939; Тихонов, 1941). Судя по имеющимся данным, оснований для признания ее нет.) (с марта—апреля по май—июнь, в более северных и восточных районах позднее: у Финмаркена — в мае, в Кольском зал. — в мае—августе); 2) у берегов Норвегии к югу от 63° с. ш., наиболее интенсивно между 58 и 60° с. ш. у юговосточных (в Скагерраке) и юго-западных (от Листер-фиорда до Алезунда), между 62 и 63° с. ш. в районе Мёре и на банке Викингов (норвежская сельдь, с февраля по май); в районе между нерестилищами мурманской и норвежской сельдей нерест происходит в крайне незначительном размере, единичные нахождения личинок известны только от Тронделаген, в Бейстад-фиорде и других фиордах к северу до Склина-банки (норвежская фиордовая сельдь); 3) вблизи Шетландских о-вов (с января по апрель); 4) в некоторых южных фиордах Фарерских о-вов (весенние сельди Фарерских о-вов, с марта, возможно с февраля, по май включительно); 5) у южных берегов Исландии от м. Рейкьянес на западе до V. и Е. Horn на востоке (весенние сельди Исландии, в марте—апреле); 6) у берегов Гренландии в Датском прол. (были находимы близкие к зрелости сельди в июле, что указывает на возможность нереста здесь весенних сельдей). Зимние шотландские сельди мечут икру: 1) в районе Гебридских о-вов и к северу от Шотландии (сельди западного побережья Шотландии, с января по март включительно); 2) на банках против устья р. Клайд у побережья Айршайр и юго-западного побережья о-ва Аррэн (клайдские или клондайкские сельди, конец февраля или начало марта); 3) у южных берегов Ирландии и в западном входе в Ламанш (сельди кельтийского плато, с января по апрель включительно); 4) у Фарерских о-вов, места точно не известны (летние сельди Фарерских о-вов, в августе—сентябре); 5) у южных и юго-западных берегов Исландии (летние сельди Исландии, с конца июня до середины августа); 6) у берегов Гренландии вблизи мыса Фэруэл (летние сельди Гренландии, недавнего, повидимому исландского происхождения, август). Сельди Ламанша нерестуют по всем берегам пролива и в южной части Северного моря — у берегов Англии в Плимутском зал. и у Сёссекса, у берегов Франции в Сенской бухте и от м. Антифер до м. Гри Нэ (с сентября по март включительно с максимумами в ноябре—декабре и в декабре—январе, в Плимутском зал. в декабре—феврале). Места нереста сельдей южного входа в Ирландское море точно не известны (с января по март—апрель зимне-весенние и с октября по декабрь осенние). Осенние сельди Северного моря (банковские сельди) размножаются во всей восточной части Северного моря, на Доггер, Малой Рыбачьей, Ютландской банках и на банке Герман Байт (с июня по декабрь, по направлению к южной части моря позднее, чем в северной). Сельди датских проливов, ведущих в Балтийское море, нерестятся: 1) в восточной части Каттегата и в северной части Б. Бельта (зимние сельди Каттегата, с января по март включительно) ; 2) в северо-восточной части Каттегата (район Кобер-Грунд — к юго-западу от о. Лесё, осенние сельди северо-восточной части Каттегата, в сентябре—декабре); 3) в южной части Каттегата, в Зунде, Б. Бельте и в западной части Балтийского моря к югу от о. Лоланда и у о. Борнгольм (осенние сельди южной части Каттегата и Зунда, в августе, сентябре и октябре); 4) в южной части Скагеррака, в западной части Каттегата, в Бельте и в западной части Балтийского моря к юго-западу и югу от о. Лоланд до Рюгена и к западу от него (весенние сельди Скагеррака, Каттегата и Бельта, вблизи берегов, места нереста изменяются, с марта по июнь включительно). Сельди Зюдерзее до отделения его от Северного моря размножались в наиболее глубоких частях залива (с апреля по май включительно). Особо следует сказать о нереете сельдей в Лим-фиорде, пересекающем Ютландский п-ов в его северной части. Здесь размножаются две формы: 1) восточная, древнего происхождения, проникшая сюда из Каттегата, которая размножается весной, с апреля по начало июня, и 2) западная, недавнего происхождения, зашедшая сюда после прорыва в 1825 г. перешейка, отделявшего фиорд от Северного моря, и нерестующая осенью; последняя сельдь близка к осенним сельдям Северного моря, в частности к размножающимся на банках Ютландской и Герман Байт. Более позднего происхождения сельди, размножающиеся весной и осенью в нескольких местах Кильского канала, который с 1896 г. соединяет Северное море с западной частью Балтийского через Ютландский п-ов у его основания. Нерест у берегов Сев. Америки происходит с апреля по ноябрь. В зал. Св. Лаврентия нерестует весенняя и осенняя сельди, в зал. Фёнди — весенняя и поздне-летняя, у западного побережья Новой Шотландии — весенняя и осенняя, южнее до м. Код — позднелетняя и осенняя, причем нерест здесь происходит позднее, чем в более северных районах.
Биология. Икрометание у подвида в целом, как видно из всего сказанного выше, происходит круглый год, причем даже у сельдей одной и той же группы по сезону нереста, например у весенних или какой-либо другой группы, сроки его значительно изменяются в зависимости от географической широты положения нерестилищ. Нерестилища располагаются вблизи берегов, в фиордах и заливах, в проливах между островами или отделяющих острова от материка, на банках среди моря, на глубине от нескольких метров до 200 м, различной для разных форм сельдей. На сравнительно небольшой глубине мечут икру сельди, размножающиеся в проливах, заливах и вблизи берегов: сельди датских проливов (осенние сельди Каттегата и Зунда на глубине от 8 до 22 м, весенние сельди Скагеррака, Каттегата и Бельта — от 8 до 24 м, осенние сельди северо-восточной части Каттегата и зимние сельди Каттегата — от 29 до 34 м), клайдские сельди (от 15 до 40 м) и весенние сельди Фарерских о-вов (до 50 м). На еще меньшей глубине вблизи берегов нерестует еще мало изученная летняя сельдь Гренландии (0.5 м). На наибольшей глубине расположены нерестилища сельдей, размножающихся в отдалении от берегов, в частности осенних (банковских) сельдей Северного моря (до 200 м). Приблизительно на такой же глубине нерестует мурманская сельдь. Норвежская сельдь размножается на глубине до 100—120 м, по более ранним указаниям, на значительно меньшей — от 0 до 70 м. Икра, в отличие от икры преобладающего большинства сельдевых, клейкая, приклеивающаяся к различному субстрату на дне моря, откладывается на дно преимущественно с твердыми, песчанистыми, гравиевыми и каменистыми грунтами, где она располагается тонким слоем по поверхности дна — по камням, битой ракуше, приклеивается к песчинкам, иногда к подводной растительности. На илистых и песчано-илистых грунтах сельдь, как правило, икру не откладывает, за исключением редких случаев, в частности, когда дно покрыто растительностью. В отношении солености нерестилища, расположенные в Атлантическом океане и в Северном море, довольно однообразны — соленость на них близка к нормальной океанической. Наиболее широки пределы колебания содержания солей на нерестилищах мурманской сельди (31.9—35.9‰), покрывая пределы колебаний солености на нерестилищах остальных, размножающихся в Атлантическом океане и Северном море форм. При значительно более низком и изменчивом содержании солей происходит размножение сельдей в проливах, ведущих в Балтийское море, и в некоторых заливах. Осенние сельди северо-восточной части Каттегата и зимние сельди Каттегата мечут икру при солености 29—34‰. Осенние сельди южной части Каттегата и Зунда нерестуют при еще более низком содержании солей (от 8 до 22‰). При наиболее низкой солености размножались сельди Зюдерзее (от 9 до 11‰). Температурные условия нереста подвида в целом довольно широки, с амплитудой колебаний от 4 до 14°, но для разных форм значительно уже и в общем довольно постоянны, с колебаниями в несколько градусов. Весенние сельди нерестятся при более низкой температуре воды, чем сельди с летним и осенним икрометанием. При наиболее высокой температуре воды размножаются осенние сельди датских проливов (осенние сельди северо-восточной части Каттегата при 11—13°, осенние сельди южной части Каттегата и Зунда при 12—14°). При наиболее низкой температуре происходит размножение зимних сельдей Каттегата — от 4 до 5°. Плодовитость для подвида в целом, по исследованиям ряда авторов, колеблется в довольно широких пределах. Наименьшее количество икринок указывается в 10 тыс., наибольшее — до 100 тыс. Пределы колебаний, указываемые разными авторами, значительно уже, средние значения существенно отличаются, что обусловлено принадлежностью исследованных сельдей к разным формам и неодинаковыми размерами их. Так, например, сельди от восточных берегов Шотландии имеют от 21.5 до 47.5 тыс., в среднем 31 768 икринок (Fulton, 1891), сельди из устья р. Шлей — от 21 до 95 тыс., в среднем 44.5 тыс. икринок (Apstein, 1898), осенние сельди из Кильской бухты — от 13 до 65.4 тыс. икринок, весенние тех же мест — от 26 до 40 тыс. икринок (Jenkins, 1902). Мурманская сельдь длиной от 23 до 25 см имеет от 14.8 до 23.3 тыс. икринок (Есипов, 1939). Икра выметывается одновременно (Наумов, 1939). Диаметр икринок у сельди из района р. Шлей колеблется от 1.125 до 1.350 мм, в среднем равен 1.238 мм (Apstein, 1898). У сельди, размножающейся в Кильском канале, икринки крупнее, от 2 до 3 мм (Hinkelmann, 1899). У берегов Ирландии икринки весенних сельдей имеют диаметр от 0.8 до 1.48 мм, осенних мельче, 0.36—1.06 мм, причем у рыб одного размера икринки весенних сельдей приблизительно на 25% тяжелее икринок осенних (Farran, 1938). Период развития икры в зависимости от температуры воды длится от 6 до 50 суток, при 14—19° он продолжается 6—8 суток, при 0—1° — от 47 до 50 суток, при наиболее обычных в естественных условиях температурах 9—10° — 12 суток, при 3—4° — 15 суток. При выклевывании из икры личинки имеют размеры от 5 до 8 мм, причем вышедшие из икры, развивавшейся более длительный срок при более низкой температуре воды, крупнее вышедших из икринок, развитие которых происходило в более короткий период при высокой температуре воды. Благодаря этому личинки сельдей, размножающихся в холодные сезоны в северных частях ареала, имеют более крупные размеры, чем личинки сельдей, нерестующих в теплые сезоны или в южных частях ареала. Личиночная стадия с желточным мешком длится от 3 до 21 суток, причем срок развития тоже определяется температурой воды. В районе Плимута желточный мешок всасывается, начиная с размеров личинки в 8 мм, наичаще при 10 мм, реже при несколько больших, до 12 мм (Lebour, 1921). По всасывании желточного мешка личинки переходят к активному питанию, но указываются случаи питания личинок и до всасывания желточного мешка (Lebour, 1922; Marshall, Nicholls and Orr, 1937). По выходе из икры большинство личинок сразу же поднимается в поверхностные слои воды, причем они совершают вертикальные суточные миграции, встречаясь в большем количестве у поверхности ночью. В поверхностных слоях воды держится преобладающее большинство питающихся личинок, в придонных — большая часть еще не питающихся; в промежуточных слоях воды и те и другие личинки встречаются в значительно меньших количествах (Lebour, 1924). Течениями, в сфере действия которых находятся места размножения, личинки и мальки некоторых форм сельди далеко уносятся от нерестилищ. В частности, личинки и мальки норвежской сельди в прибрежной ветви Североатлантического течения перемещаются к северу и в возрасте 1—2 лет достигают берегов Финмаркета (Johansen, 1927; Lea, 1929; Расс, 1939). Сходным образом переносятся течением с мест нереста личинки и мурманской сельди. Единичные экземпляры близких к нересту мурманских сельдей найдены в юго-западной части Баренцова моря зимой и весной, наиболее часто в январе и феврале, реже в декабре, марте, апреле и июле, еще реже в другие месяцы. Сельдь со зрелыми половыми продуктами найдена у северных берегов Норвегии в апреле, в Кольском заливе в мае и июне—августе. Отнерестившиеся сельди единично найдены у берегов северной Норвегии, на Финмаркенской и Мурманской банках весной, в феврале и апреле, и еще реже летом, в июне и июле (Расс, 1939). Икринки мурманской сельди найдены 11 апреля в желудке трех пикш, пойманных на глубине 209 м в районе о-ва Вествогей (Мантейфель и Марти, 1939) и, кроме того, в большом количестве в желудках пикши, ловимой в апреле 1939 г. на Малангенской банке, где одновременно поймана и нерестующая сельдь (Расс, 1939). Личинки с желточным мешком за период исследований с 1899 по 1938 гг. найдены рядом авторов у северных берегов Норвегии от Вест-фиорда на западе до Трольд-фиорда на северо-востоке (Ehrenbaum, 1902; Runnström, 1934; Рыженко, 1938, 1939; Мантейфель и Марти, 1939). В наибольшем количестве личинки в этой стадии обнаружены на банках против островов Вествогей, Лангней, Андей, Квалей, Рингвассе и фиорда Маланген, преимущественно с внешней стороны островов; со стороны материковой, в фиордах, шхерах и проливах, личинки встречаются единично или (в Вест-фиорде) отсутствуют совершенно (Мантейфель и Марти, 1939). У Финмаркена и Мурмана нерест, судя по имеющимся данным, более обычен в шхерах (Сёре-Сунд) и фиордах (Кольский зал.). В наибольшем количестве личинки пойманы в марте и апреле, в мае — единично. Наибольшее количество личинок попадается над глубинами от 50 до 150 м и наичаще от 150 до 200 м, куда простирается сфера действия Североатлантического течения, а вообще найдены над глубинами от 7 до свыше 700 м, но над глубинами менее 50 и свыше 300 м обнаружены лишь в единичных случаях. Судя по нахождению личинок с желточным мешком, в марте сельдь нерестится на несколько большей глубине, чем в апреле. Температура придонных слоев воды в местах нахождения личинок колеблется от 5.6 до 6.9°, в местах нахождения личинок над большими глубинами (600—700 м) — ниже, до 2.3°. Соленость в придонных слоях воды здесь колеблется от 34.87 до 35.95‰ (Расс, 1939; Мантейфель и Марти, 1939). Грунт на местах нереста песчаный и гравиевый. Личинки мурманской сельди первых двух стадий по всасывании желточного мешка размерами до 35 мм в южной, где дрейф наиболее изучен, и в северной ветвях Североатлантического течения уносятся от мест нереста вдоль берегов северной Норвегии и Финмаркена на север до 72° с. ш. и на восток до 33° в. д. Мальки в литоральной стадии размерами 35—60 мм, в возрасте около 5 месяцев найдены у самых берегов Мурмана от губы Титовка (Мотовский зал.) до губы Вороньей (35°50' в. д.), главным образом в местах впадения рек, пройдя, таким образом, около 800 миль от мест нереста, что близко согласуется со скоростью Нордкапской ветви. Вполне сформировавшиеся мальки размерами свыше 60 мм (до 90—100 мм) осенью и зимой (с середины октября и почти до конца марта) обнаружены вдоль берегов от Мотовского зал. до восточной части Мурмана и в открытом море до Канинской и Гусиной банок, до 47°50' в. д. на восток и 71°51' с. ш. на север, опускаясь в этот период в придонные слои воды (Расс, 1939). На втором и третьем году жизни мурманская сельдь занимает наиболее широкий ареал в Баренцовом море, летом активно распространяется в центральной и восточной частях моря на север до 76°48' с. ш., проникая на восток до п-ова Канина, о. Колгуева и заходя в Белое море, зимой отходит в центральные и западные районы моря. В этом возрасте сельдь обнаруживает усиленный рост в длину, но жировых отложений не накапливает и поэтому почти не прекращает питаться и зимой. Летом, начиная с половины мая по июнь, одно- и двухгодовалые с небольшой примесью трех- и четырехгодовалых заходят в губы Мурманского побережья (так называемая летняя сельдь), причем в одних стаях преобладают одногодовалые, в других двухгодовалые. Возрастной состав летней сельди для отдельных лет, по данным Глебова (1938), таков (в процентах):
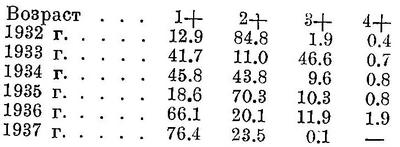
Заходы летней сельди в губы не носят регулярного характера — в некоторые годы она заходит в большом количестве, в иные — в незначительном. Большим колебаниям подвержены и сроки начала заходов сельди. Летние подходы сельди к берегам и заходы ее в губы обусловлены в основном большим прогреванием их вод по сравнению с открытым морем. Заходы летней сельди в губы начинаются при наличии более, высокой температуры воды поверхностных слоев в прибрежной области и в губах, чем в открытом море. Особенностями термического режима прибрежных вод в отдельные годы обусловливается и нерегулярность подходов летней сельди. В годы с более ранним прогреванием поверхностных слоев воды в прибрежной области и в губах сельдь заходит раньше. Наибольшие заходы сельди бывают в годы с более резко выраженным повышением температуры поверхностных слоев воды по направлению из моря в губы. Летняя сельдь заходит главным образом в губы Кольского зал., в губы Мотовского зал. заходит обычно в значительно меньшем количестве и позднее, что обусловлено различиями в термическом режиме этих заливов — поверхностные слои воды в более узком Кольском зал. нагреваются быстрее и больше, чем в Мотовском зал. Кроме того, на термический режим Кольского зал. существенное влияние оказывает поступающая в него в большем количестве пресная вода, летом более теплая и значительно отепляющая поверхностные слои его воды. Подходам летней сельди в прибрежные воды и заходу ее в губы благоприятствуют слабые береговые ветра, выносящие дальше в море прогретые прибрежные воды. Сильные ветра, перемешивающие воду, на подходы сельди действуют отрицательно (Глебов, 1938). Начиная с четвертого года жизни, сельдь не имеет такого широкого распространения в Баренцовом море на север и на восток, как в предыдущие возрасты, за лето накапливает в мышцах и на внутренностях жировые отложения. С наступлением осени, сокращая ареал по сравнению с летним, сельдь перемещается из восточных районов в западные, сперва в район южного склона Мурманской банки, позднее на северо-западный, весной (в марте) в район Нордкапской и Пордкинской банок. Зимой опускается в придонные слои воды и обычно перестает питаться. С наступлением лета вновь распространяется к востоку — до северного склона Мурманского мелководья и меридиана Святого Нoca. В этом, а также в несколько меньшем и большем возрастах (2+ — 6+) сельдь (так называемая жирная сельдь) в массе заходит в прибрежную область и в губы Мурманского побережья, составляя основу прибрежного промысла. В отличие от летней сельди жирная сельдь, подходя в прибрежную область ежегодно, в губы может и вообще не заходить, если она в течение лета не накапливает в теле в достаточном количестве жира, продолжая отчасти питаться и зимой, хотя наполнение желудков бывает большей частью незначительным (Болдовский, 1938). В прибрежные воды заходит лишь достаточно упитанная и жирная сельдь. Заходы жирной сельди в прибрежную область и в губы обусловлены термическим режимом, причем, в отличие от заходов летней сельди, происходят при более низкой температуре воды здесь, чем в открытом море. Вопрос об условиях захода жирной сельди в губы все же не совсем ясен, если принять во внимание, что при отходе в западные районы моря та же жирная сельдь ориентируется на более высокую температуру воды. Следует также отметить, что массовые заходы жирной сельди за время с 1930 по 1941 гг. наблюдались только для урожайного поколения 1930 г. в 1933 и 1934 гг. (Марти, 1941 : 117). Подходам сельди благоприятствуют сгонные ветра, вызывающие выход на поверхность холодных нижних слоев воды. Первые подходы жирной сельди в прибрежную область наблюдаются с августа, иногда даже с июня. Массовые заходы в губы начинаются с октября — начала декабря, оканчиваются с начала января по середину марта. В небольшом количестве жирная сельдь заходит в губы и раньше, в августе и сентябре. В отличив от летней жирная сельдь заходит преимущественно в губы Мотовского зал. и значительно реже в губы Кольского (Глебов, 1938). Заходы атлантической сельди в Белое море аналогичны заходам ее в губы Мурманского побережья. Старше трехгодовалого возраста в Белом море атлантическая сельдь не встречается, и основную массу в уловах составляют особи с одним и двумя зимними кольцами на чешуе. В наибольшем количестве встречается в горле и центральной части моря, составляя в отдельных уловах до 50 и 100%. Заходы в последние годы (1938, 1942, 1944, 1946, 1948) в связи с потеплением возросли (Тамбовцев, 1947, 1949). Выход подросшей атлантической сельди из Белого моря в Баренцово совпадает с превращением мелкой сельди в жирную (Дмитриев, 1946). В открытом море осенью сельдь держатся при температуре воды 5—7°. Зимой жирная сельдь в губах, в прибрежной области и в море держится при температуре воды около 2—3.6°, опускаясь в море в придонные слои (Мантейфель и Болдовский, 1938). В море в поверхностных слоях воды в наибольших концентрациях мурманская сельдь держится на Финмаркенской банке и на северо-западном склоне Мурманской банки, а в июле—августе, кроме того, на северо-восточном и юго-западном склонах последней и в прибрежной области. В сентябре—октябре и позднее в значительных количествах сельдь была обнаружена на Мурманской, Финмаркенской и Рыбачьей банках, во вторую половину зимы — к западу от Финмаркенской банки и в районе нерестилищ (Марти, 1939). В придонных слоях воды в ноябре—декабре основные скопления сельди обнаружены в восточных районах — на Гусиной банке, в Центральном районе, на Мурманском мелководье и на Святоносской банке при температуре воды 3.4—4.8° у дна и 3.5—4.9° у поверхности; в прибрежных районах держится крупная сельдь, более мелкая — далее в открытом море. В январе—феврале крупная сельдь встречается в западной части моря — на Мурманской, Финмаркенской, Нордкинской, Нордкапской и Демидовской банках откуда зрелые идут к местам нереста, а неполовозрелые с прогреванием верхних слоев воды (с апреля) поднимаются к поверхности (Тихонов, 1939). На пятом году значительная часть мурманской сельди впервые становится зрелой. С наступлением осени созревающая сельдь начинает отходить на запад, в октябре—ноябре, в некоторые годы в январе держится по восточному склону Мурманской банки и вблизи северных склонов Рыбачьей банки. В марте сельдь с совершенно зрелыми половыми продуктами держится далее к западу, в районе Нордкапской банки. Сельди, нерестующие впервые, размножаются главным образом в районе от Малангенской банки до о. Сёре, сельди более старших возрастов идут для нереста в более южные части нерестилищ. После первого нереста мурманская сельдь в шести- семигодовалом возрасте держится предположительно на границе Баренцова и Норвежского морей к югу от о. Медвежий, в восточную часть моря не заходит. Сельдь более старшего возраста (восьми- двенадцатигодовалая) мигрирует для питания после нереста по Шпицбергенской ветви Североатлантического течения на север до широты западных берегов Шпицбергена (77—78° с. ш.) (Марти, 1941). Схема жизненного цикла мурманской сельди по имеющимся данным в общих чертах такова. Личинки с нерестилищ у о-вов Лофотенских и Вествогей в Нордкапской ветви Североатлантического течения дрейфуют на север и восток и проходят литоральную стадию в возрасте 4—5 месяцев у берегов Финмаркена и Мурмана. По окончании метаморфоза мальки в струях течений активно расселяются в центральных и восточных частях Баренцова моря. На втором и третьем годах жизни сельдь в летнее время имеет в Баренцовом море наиболее широкий ареал, зимой, отходя в центральные и западные районы моря, занимает меньший ареал, летом заходит в губы Мурманского побережья. Начиная с четвертого года жизни, ареал сельди смещается к западу. Летом сельдь этого возраста распространяется в восточном направлении, осенью и зимой отходит к западу и подходит в прибрежную область, в некоторые годы в массе заходя в губы Мурманского побережья. Созревающая сельдь, начиная с осени, в течение зимы из западной части моря постепенно отходит к местам икрометания и впервые размножается в северных частях нерестилищ. После первого икрометания пяти- и шестигодовалые сельди отходят с нерестилищ в западную часть Баренцова моря и, повидимому, держатся на границе его с Норвежским. Повторно нерестующие сельди (восьмигодовалые и выше) размножаются в более южных частях нерестилищ и после икрометания отходят к северу, до Шпицбергена (Марти, 1941; Расс, 1939). Схема жизненного цикла норвежской сельди в общих чертах сходна с таковой мурманской сельди, отличаясь в некоторых деталях. Личинки норвежской сельди с нерестилищ у южных берегов Норвегии прибрежной ветвью Североатлантического течения сносятся вдоль берегов к северу, достигая в возрасте 1—1.5 лет Финмаркена, где ловятся в виде мелкой сельди (smaasild, small-herring). У берегов Финмаркена сельдь отходит от берегов и мигрирует обратно к югу, подходя вновь к берегам средней части Норвегии и в возрасте 2.5—4 лет ловится здесь в виде жирной сельди (fetsild, fat-herring). С началом созревания половых продуктов сельдь вновь отходит от берегов, на этот раз значительно дальше и в течение одного или нескольких лет в возрасте 2—7 лет проходит в открытом море так называемую океаническую стадию жизни, отсутствующую у мурманской сельди. Созревающие сельди в возрасте 4—7 лет подходят к местам нереста и ловятся здесь в виде так называемой крупной (storsild, large-herring) и весенней (vaarsild, spring-herring). По окончании нереста норвежская сельдь отходит далеко к северу, заходя в западную часть Баренцева моря и достигая о. Медвежьего и широты южной оконечности Шпицбергена, откуда через год возвращается для нереста вновь и мечет икру в более южных частях нерестилищ, чем сельдь, размножающаяся впервые (Lea, 1929; Runnström, 1937; Расс, 1939). Однако последующими исследованиями (Марти, 1941) установлено, что по Шпицбергенской ветви течения к северу совершает миграции лишь мурманская сельдь. В последние годы получены данные, свидетельствующие о сходстве между норвежской сельдью и сельдью от северных берегов Исландии (Rasmussen, 1950), а также о миграциях сельди между Норвегией и северной Исландией (Fridriksson a. Aasen, 1950). Вследствие различия в условиях существования в южных и северных частях вытянутого почти в меридиональном направлении ареала норвежская сельдь в первые годы жизни (мелкая сельдь) по характеру роста и строению зимних колец на чешуе существенно различается. Сельдь, проведшая первые годы жизни в южной части западного побережья Норвегии, имеет неясно выраженные зимние кольца (так называемого южного типа зимние кольца). Сельдь, попавшая к северному побережью Норвегии, имеет на чешуе резко очерченные зимние кольца (так называемого северного типа). Сельдь, ловимая у западных и средней части северных берегов Норвегии, по строению зимних колец представляет смесь обоих типов. Количество сельдей с кольцами южного типа увеличивается к югу, с кольцами северного типа — к северу. Кольца северного типа имеются в количестве 3—6, южного — не более чем 3. В течение океанической стадии на чешуе сельди откладываются 1—2 менее резко выраженных, чем северного, — но более резко, чем южного типа, кольца (так называемого океанического типа). Наконец, на чешуе нерестовой сельди откладываются более определенные и резко очерченные кольца нерестового типа (Lea, 1929; Runnström, 1936). Чешуя мурманской сельди по строению зимних колец в преобладающей массе относится к северному типу; сельдей с кольцами южного типа у ней не обнаружено. Колец океанического типа у мурманской сельди также не найдено. Однако на чешуе части сельди урожайного поколения 1930 г. у 74% общего количества исследованных рыб этого поколения обнаружены 1—2 зимних кольца, значительно более широких, чем кольца северного типа, похожие на океанические кольца норвежской сельди, но значительно менее широкие, чем последние. Сельди с чешуёй такого типа (названного морским) ловились в течение нескольких лет, начиная с конца лета или осенью, т. е. в то время, когда у берегов Мурмана появляется норвежская сельдь, и, повидимому, относятся к последней. Судя по более быстрому темпу роста лет с морским типом колец, сельдь эта держится вдали от берегов Мурмана. Среди норвежских сельдей, заходящих в Баренцово море, сельдей с кольцами южного типа, по исследованиям последних лет, не найдено (Мурашкинцева, 1938). Годичный цикл питания мурманской сельди таков: наиболее интенсивно сельдь питается летом, главным образом в июне и июле; с августа по октябрь интенсивность питания снижается, в ноябре становится крайне незначительной; с декабря по март сельдь почти совсем не питается, за исключением сельди, зимующей в открытом море, которая понемногу начинает питаться с января. Одно- и отчасти двухгодовалые сельди в море не прекращают питаться и зимой. Неполовозрелая одно- пятигодовалая сельдь в открытом море наиболее интенсивно питается летом, поедая в это время в массе Calanus finmarchicus. Весной и осенью основными компонентами питания сельди в море являются Euphausiidae и отчасти Calanus finmarchicus, кроме августа, когда сельдь поедает преимущественно молодь рыб (Болдовский, 1941). В прибрежной области в период перед зимними заходами в губы интенсивность питания сельди слабая, поедаются почти исключительно Euphausiidae (Thyssanoessa inermis и Thyssanoessa raschii), в некоторые годы Limacina retroversa. В период зимних заходов в губы, с октября по апрель, желудки сельдей всех возрастов (1+ — 5+) пустые. По окончании зимней концентрации сельди в губах она начинает понемногу питаться, поедая главным образом Euphausiidae, Sagitta и Calanidae (Calanus finmarchicus, Calanus hyperboreus, Metridia longa, Metridia luceus). Во время летних заходов в губы сельдь питается менее интенсивно, чем в открытом море. В июне основу питания летней сельди в губах составляют Calanus finmarchicus и отчасти личинки Euphausiidae и некоторые другие ракообразные. В июле интенсивность питания сельди в губах снижается еще более, основной пищей в это время являются Calanus finmarchicus и, кроме того, мелкие Copepoda, Cladocera, личинки Cirripedia и Lamellibrachiata. В августе сельдь в губах питается еще менее интенсивно поедая в основном Euphausiidae, мальков рыб и отчасти Calanus finmarchicus и мелких Copepoda, Cladocera и др. За весь период пребывания сельди в губах основу питания ее составляют личинки Cirripedia (37.6%), Calanus finmarchicus (34.2%) и Euphausiidae (13.9%), которые в общем итоге дают 95% общего компонента по весу. В желудках мальков литоральной стадии, размерами 30—45 мм в губах (Пала-губа) Мурманского побережья, пойманных в начале августа 1933 г., обнаружены ювенальные стадии и nauplii Copepoda (Oithona similis), Microstella norvegica, личинки Lamellibranchiata и единично Peridinium depressum. Сеголетки в губах не прекращают питаться и зимой (Болдовский, 1938). Только что выклюнувшиеся личинки у берегов Великобритании вначале, еще до всасывания желточного мешка, поедают личинок моллюсков и nauplii Copepoda, затем по мере роста, после всасывания желточного мешка, пища личинок состоит из взрослых Copepoda (Pseudocalanus elongata clausii, Corycaeus angilis и др.), диатомей (главным образом Paralapia sulcata и др., заглатываемых отчасти вместе с организмами, питающимися ими), инфузорий и др. (Lebour, 1918, 1921, 1924; Marshall, Nicholls a. Orr, 1937). Личинки сельдей поедаются рядом рыб (треской, пикшей и др., даже самой сельдью) и некоторыми беспозвоночными (медуза Rathkea octopunctata, ктенофора Pleurobrachia pileus, червь Sagitta bipunctata и др.; Lebour, 1922, 1923). Существенными пищевыми конкурентами мурманской сельди являются гребневики (Bolina, Beroe), питающиеся планктоном и имеющие массовое развитие с августа, иногда с июня, причем сельдь в период интенсивного питания (в июне и июле) в местах массового скопления гребневиков держалась в меньшем количестве, чем в местах, свободных от гребневиков. В периоды менее интенсивного питания сельди массовое скопление гребневиков обычно сопровождается большими концентрациями и сельди (Болдовский, 1939). На распределение сельди в море отрицательное влияние оказывает периодическое массовое развитие водорослей, отмеченное рядом авторов и в Северном море. В Баренцовом море массовое развитие водорослей (Phaeocystis, Chaetoceros) происходит в мае в центре южной части моря в поверхностных слоях воды (до глубины 25—50 м), вследствие чего сельдь здесь в это время доржится на глубине 50—100 м. В июне области массового развития водорослей перемещаются к востоку от Мурманской банки, а затем к северо-западу, в связи с чем сельдь поднимается в поверхностные слои воды, скапливаясь у границ распространения водорослей и перемещаясь по мере отодвигания их. Существенное влияние на распределение сельди оказывает развитие водоросли Rhizosolenia, наблюдаемое нередко в западных частях Баренцова моря в июле, в связи с чем сельдь из этих районов отходит (Болдовский, 1939; Мантейфель, 1941). Половой зрелости преобладающее большинство форм атлантической сельди достигает в трех- четырехгодовалом возрасте. Норвежская сельдь впервые мечет икру и трех- восьмигодовалом возрасте, причем массовое созревание (46.8%) происходит в пятигодовалом возрасте, несколько меньшее (21.7%) — и четырехгодовалом. Мурманская сельдь впервые начинает размножаться в четырех- семигодовалом возрасте, массовое созревание (43.5 %) происходит в шестигодовалом возрасте, несколько меньшее (31.4%) — в пятигодовалом, в четырехгодовалом — в незначительном количестве (7.1%), трехгодовалые еще неполовозрелые (Тихонов, 1941). Рост мурманской и норвежской сельди по приводимым для последней Тихоновым (1941) данным Рунстрема для категории 1930 г. происходит следующим образом (см):
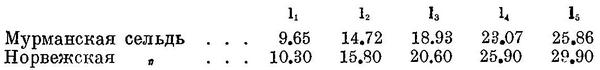
Возрастной состав, как и у других промысловых рыб, в отдельные годы различен. Колебания возрастного состава обусловливаются различной урожайностью отдельных поколений, причем урожайное поколение имеет преобладающее значение в промысловых уловах в течение ряда лет. Наиболее урожайное поколение норвежской сельди 1904 г. впервые стало ловиться по достижении половой зрелости в трехгодовалом возрасте в 1907 г. (1.6% улова), преобладало в уловах в 4—14-годовалом возрасте в 1908—1918 гг. (78.1—31.6%), т. е. в течение 11 лет, составляло значительную часть улова в 15—18-годовалом возрасте в 1919—1922 гг. (24.8—9.6%) и встречалось еще в уловах до 23-годовалого возраста в 1923—1927 гг. (3.9—1.5%) (Sund, 1943). У мурманской сельди одним из урожайных было поколение 1930 г., которое являлось основным в уловах летней сельди в губах в 1931 и 1932 гг. и зимой 1933—1934 и 1934— 1935 гг. Далее урожайным было поколение 1934 г., которое особой роли в уловах летней сельди в губах не играло, но имело существенное значение в прибрежных дрифтерных уловах в 1936 г. и особенно в морском дрифтерном промысле в 1937 и 1938 гг. Вследствие того что вблизи берегов Мурмана держится неполовозрелая сельдь в возрасте до 6 лет включительно, влияние урожайных поколений у мурманской сельди в нашем промысле сказывается в течение только нескольких лет. Количество жира, отлагаемого в теле и на внутренностях сельди, сильно изменяется в зависимости от возраста и сезона лова. С возрастом жирность сельди возрастает. Наибольшее содержание жира в мясе бывает после интенсивного летнего питания, достигая в августе у двухгодовалых сельдей (длина 17 см) около 9%, у четырех- пятигодовалых (длина около 24—25 см) — до 23—26 %. В дальнейшем, с понижением, а позднее почти с прекращением питания происходит уменьшение жирности до апреля месяца, когда содержание жира падает в среднем до 1.28% . У мелкой сельди после уменьшения жирности в сентябре наблюдается небольшое повышение ее в октябре, причем, судя по имеющимся данным, до декабря количество жира уменьшается мало. Наибольшее содержание жира на внутренностях у крупных сельдей приходится на сентябрь, у мелкой — на октябрь—ноябрь. Общее содержание жира в теле у крупных сельдей (длиной около 22 см) в августе достигает 22.6%, в мае падает до 4.7%. У мелкой сельди (17 см длиной) общее содержание жира достигает максимума в октябре—ноябре (12.3— 12.7%), снижаясь в апреле до 2.3% (Юданова, 1939).
Хозяйственное значение. Занимает по уловам второе место. Общие уловы составляют 15—16 млн. ц, т. е. 40—50% улова Зап. Европы. В наибольшем количестве вылавливается в Северном море (9.1—10.3 млн ц), у берегов Норвегии (2.2—2.5 млн. ц) и Исландии (1.5— 2.3 млн. ц). Уловы СССР в Баренцовом море в годы наилучших подходов достигали до 975 тыс. ц (1935 г.), в обычные годы менее, в 1939 г. 162 тыс. ц. Уловы в наших водах могут быть значительно увеличены. В море у Терского берега Белого моря в июне—июле ловится молодь сельди (так называемая килька) и мелкая летняя сельдь; в Баренцовом море у восточных берегов Мурмана и в центральных и западных районах в августе—ноябре ловят жирную сельдь, в западных районах в декабре—марте — преднерестовую сельдь, в Медвежинско-Шпицбергенском районе в июле—октябре — крупную сельдь. В заливах и губах Мурманского побережья в декабре—июне ловят молодь (килька) и мелкую летнюю сельдь, в ноябре—феврале — мелкую жирную и жирную сельдь.
*1b. Clupea harengus harengus n. membras L. — Балтийская сельдь, салака (табл. III).
— Clupea harengus β membras Linne, Fauna suecica, ed. II, 1761 : 128 (Stromling, vero minor ex mari Bothnico); Pallas, Zoogr. rosso-asiat., III, 1811 : 211 (частью: Балтийское м., у берегов Лифляндии и у о. Эзеля).
— Clupea harengus Кесслер, Рыбы СПб. губ., 1864 : 185 (частью : близ устья Наровы, у Кронштадта); Smitt, Scand. fish., II, 1895 : 954, 961 (частью: var. membras).
— Clupea harengus var. membras Heincke, Naturgesch. d. Herings, I, 1898 : LXIY, Tab. CXLII—CLXIX, Taf. I, II, V, VI, IX, X, XIV, XVII, XVIII (восточная часть Балтийского м. к востоку от Пиллау и о. Готланда, Ботнический и Финский заливы; измерения рыб и черепов); Суворов, Тр. Балтийск. эксп., II, 1913 : 44 (Балтийское м.; малоглазая и большеглазая формы).
— Clupea harengus membras Rabinersоn, Zool. Jahrb., Abt. Syst., Bd. 58, 1929 : 300 (Финский зал.); Берг, Рыбы пресных вод, I, 1932 : 95 (Балтийское м.).
— Clupea harengus Lе Gall, Ann. Inst. oceanogr., XV, 1, 1935 : 155—161 (частью: Балтийское м.).
— Clupea harengus membras Телегин, в: Промысловые рыбы СССР, 1949 : 124, табл. 31.
Распространение, биология. Möbius u. Heincke, Fische d. Ostsee, 1883 : 265 (частью: var. membras); Reuter, Meddel. Soc. Fauna Flora fennica, XX, 1894 : 52 (пресноводные озерки на островах Абосского архипелага); Schneider u. Levander, Acta Soc. Fauna Flora fennica, XX, 1, 1900 : 5, 35 (питание, развитие икры, инкубационный период); Nordquist, ibid., XX, 7, 1901 : 8 (нерест); G. Schneider, Rapp. Proces-Verb., IX, 1908 : 66 (Балтийское м. к востоку от южной конечности о. Эланд и Данцигской бухты, в пресноводном озерке пров. Кальмар; нерест, возраст, литература); Михин, Изв. Отд. прикл. ихтиол., III, 1, 1925 : 40 (подходы к берегам); Рабинерсон, там же, III, 2, 1925 : 134 (Копорская губа, у Шепелева; измерения, рост); Неsslе, Publ. Circonst., 89, 1925 : 1 (расы, миграции, питание, размеры); Journ. Conseil, II, 2, 1927 : 169 (осенние сельди, возрастной состав); Lundbeck, Mitl. Deutsch. Seefischerei-Ver., 46, 1930 : 310 (рост, возрастной состав, распределение); Неsslе, Journ. Conseil, VI, 1, 1931 : 21 (соотношение в уловах осенних и весенних сельдей); Кanеrva, Fiskeritidskr. f. Finland, 1931 : 117 (возраст, нерест); Михин и Антипова, Изв. Всесоюзн. Научно-исслед. инст. озерн. и речн. рыбн. хоз., XV, 1932 : 28, 92 (подходы к берегам, размеры, рост, возрастной состав, уловы); Andersson, Rapp. Proces-Verb., С, 2, 1936 : 11 (частью: Балтийское м. у берегов Швеции; расы, нерест); Неsslе, Svenska Hydrogr.-Biol. Kommiss. Skrifter (n. s.), Biol., II, 2, 1937 : 1 (питание, половозрелость, нерест, миграции); Andеrssоn, Rapp. Proces-Verb., CVIII, 1, 1938 : 67 (осенние сельди, рост); VI annsfeld, Korr.-Bl. Naturf.-Ver. Riga, LXIII, 1939 : 43 (p. Латвийская Aa в 9 км от устья); Берг, Изв. Всесоюзн. Научно-исслед. инст. озерн. и речн. рыбн. хоз., XXIII, 1940 : 6 (Финский зал.; уловы, нерест, биология); Alander, Ann. Hiol., I, (1939—1941), 1943 : 177; ibid., II, (1942—1945), 1947 : 172 (южная и центральная части Балтийского м., Ботнический зал.); Cieglewicz a. Posadzki, ibid. : 159 (Данцигская бухта; нерест, возраст, рост); Дементьева, Рыбн. хоз., 1947, 8 : 23 (Рижский зал. и балтийское побережье СССР; нерест, возрастной состав); Маркова, Изв. Всесоюзн. Научно-исслед. инст. озерн. и речн. рыбн. хоз., XXIX, 1949 : 106 (влияние солености и температуры на газообмен); Бризинова, там же : 143 (интенсивность пищеварения в зависимости от температуры); Кожевников, там же : 172 (подходы у Койвисто); Левиева, Рыбн. хоз., 1950, 5 : 34 (содержание жира); Дрягин, Вестн. Ленингр. унив., 8, 1950 : 27 (общий улов в некоторые годы свыше 1 млн. ц, запуск в военные годы не повлиял ни на размеры, ни на возрастной состав).

D (III) IV 14—16 (15.0), общим числом 17—21 (18.9), A III 12—16 (14.0), общим числом 15—19 (17.0), sp. br. 60—71 (66.0), килевых чешуй позади основания брюшных плавников 12—14 (13.1), vert. S. 54—57 (54.7), cand. vert. 29—33 (30.5). Брюшные килевые чешуи развиты как позади брюшных плавников, так и впереди них. Зубы на сошнике сравнительно хорошо развитые. Голова крупнее, чем у предыдущей формы, 23.4—25.9 (24.9)% длины тела; межглазничный промежуток несколько уже, 14.4—17.5 (16.1)% длины головы. Глаза крупнее, 24.7—28.7 (26.7)% длины головы. Основание анального плавника короче, 8.3—10.8 (9.7)% длины тела. Размеры меньше, чем предыдущей формы, в среднем 14—16 см, но гигантские стремлинги (см. ниже) достигают размеров до 37.5 см. Описание дано по 30 экз. длиной от 12.1 до 18.7 см, в среднем 14.9 см, собранным 22 V 1948 в районе Мерсрагс. По Гейнке (Heincke, 1898), позвонков 54—57, килевых чешуй 11—15, по Гессле (Hessle, 1925), средние числа позвонков у форм подвида колеблются от 55.2 до 55.7, килевых чешуй от 13.3 до 14.2, т. е. и позвонков и килевых чешуй в среднем несколько меньше, чем у атлантической сельди.
Морская форма, живущая и размножающаяся в воде с пониженным содержанием солей.
Балтийская сельдь является такой же локальной формой, как и атлантическо-скандинавские сельди или более близкие к ней сельди датских проливов, и рассматривается здесь в качестве особой формы лишь вследствие ее большей, чем других форм Clupea harengus harengus, географической обособленности.
Распадается на ряд еще мало изученных форм. По времени нереста различают (Hessle, 1925) две большие группы: осенние и весенние сельди. Осенние сельди характеризуются средним числом позвонков более 55.40 (от 55.40 до 55.71), туловищных позвонков более 25.25 (от 25.25 до 25.85), килевых чешуй более 13.50 (от 13.55 до 14.17); рост резко замедленный после наступления половой зрелости, мигрирующие, нерест преимущественно во внешних частях архипелага островов и вдоль побережья моря. Весенние сельди в архипелаге островов у берегов Швеции образуют ряд форм, среди которых различаются: 1) весенние сельди открытого моря — среднее число позвонков около 55.50 (от 55.37 до 55.73), туловищных позвонков около 25.00 (от 24.80 до 25.10), килевых чешуй меньше 13.60 (от 13.32 до 13.60), рост резко замедленный после наступления половой зрелости, мигрирующие, нерест вдоль берегов у архипелага островов; 2) весенние фиордовые сельди средней части моря — среднее число позвонков меньше 55.40 (от 55.17 до 55.32), туловищных позвонков меньше 24.80 (от 24.52 до 24.77), килевых чешуй меньше 13.60 (от 13.10 до 13.53), рост равномерно замедленный, локальные, нерест в архипелаге островов; 3) ледовые сельди южной части Ботнического зал. — среднее число позвонков больше 55.50 (от 55.70 до 55.74), туловищных позвонков больше 25.00 (от 25.09 до 25.25), килевых чешуй больше 13.50 (от 13.66 до 13.77), рост равномерно замедленный, мигрирующие, нерест в архипелаге островов. Кроме того, по величине глаз у наших берегов установлены среди осенних сельдей две формы: большеглазая (диаметр глаза 5.7—6.0% длины тела, экз. от Либавы) и малоглазая (диаметр глаза 5.0—5.2% той же длины, экз. от Домеснеса). В Финском зал. обе формы встречаются вплоть до его восточной части. Среди весенних и осенних сельдей встречаются крупные, так называемые гигантские стремлинги (Riesenstromlinge), достигающие свыше 300 (до 330 и даже 375) мм длины. Гейнке (Heincke, 1898) и Шнейдер (Schneider, 1908) рассматривают гигантских стремлингов как старых особей салаки, так как никаких морфологических отличий они не имеют. По основным признакам гигантские стремлинги ближе всего к весенним фиордовым сельдям, причем в значительной степени питаются рыбой, почему Шнейдер и считает их старыми особями с хищным характером питания. Однако из сопоставления роста салаки и гигантского стремлинга явствует, что последний представляет особую форму, обладающую более быстрым ростом (Kanerva, 1931; Берг, 1940; см. также ниже).
Распространение. Балтийское море к востоку от линии, проходящей к западу от южной оконечности о-ва Эланд (Карлскруна) и Данцигской бухты (Возможно, далее к западу, в частности для осенних сельдей включая побережье о. Борнгольма.), в Ботническом зал. на север до самого конца его, в Финском зал. на восток до Сестрорецка, заходит в Нарвскую, Лужскую и Копорскую губы. Заходит и живет в совершенно пресной воде — в небольшом пресном озере в Швеции (пров. Кальмар, близ Ytterhult, к северу от устья p. Virboa), соединенном с морем небольшим мелким ручьем, в некоторых небольших пресных озерках на архипелаге островов Турку (Або), в р. Латвийской Аа при северных ветрах зашла на 9 км выше устья. Нерест вдоль берегов моря и заливов. Осенние сельди мечут икру у берегов средней части моря (с августа по конец сентября), на банках у берегов Ботнического зал. (с июля по ноябрь, наиболее интенсивно в августе и сентябре), у берегов Латвии к западу от Домеснеса (в августе и начале сентября), во внешних частях шхер по северному берегу Финского зал. к западу от Ловизы (в августе и сентябре), здесь же изредка и во внутренних частях шхер, восточнее у островов Нарви и Секкер (до сентября), у Курголова (половозрелые наблюдались в конце августа). Весенние сельди нерестятся вдоль берегов средней части моря (весенние сельди открытого моря, с мая по начало июля), в фиордах между островами западного берега Ботнического зал. (весенние фиордовые сельди, с апреля по начало июля, реже по август, наиболее интенсивно в июне), в архипелаге островов у берегов Ботнического зал. (ледовые сельди Ботнического зал., июнь), в Финском зал. во внутренних (в апреле—мае и в начале июня) и внешних частях шхер (в конце мая и начале июня) северного берега к западу от Ловизы, восточнее (у Бьёркэ) близ берегов, в заливах, и позднее дальше от берегов (с конца мая по начало июня), у южных берегов к востоку от Нарвы (в конце мая — начале июня). Позднее, летом, нерест у открытого моря в незначительных размерах происходит во внешних частях шхер у Сиббо (к востоку от Хельсинки, в июне и июле) и на границе Копорской и Лужской губ и в Копорской губе дальше от берегов, чем весной (в конце июня—начале июля).
Биология. Нерест, как видно из всего сказанного выше, происходит весной, осенью и отчасти летом. Время нереста находится в зависимости от географического положения; например, в северных частях моря, в Ботническом зал., осенние сельди размножаются с июля, в средних частях моря — с августа. В северных и восточных частях моря весенняя сельдь размножается позднее, осенняя — раньше. Осенний период нереста более растянут, чем весенний. Нерест происходит вблизи берегов, в заливах, во внешних и внутренних частях шхер. Весенние сельди размножаются ближе у берегов, чем осенние, как во внешних, так и во внутренних частях шхер, на глубине от 3—4, даже 2, наичаще 7—10 м. В Финском зал. весенний нерест начинается раньше ближе к берегам, во внутренних частях шхер, позднее — в отдалении от берегов, во внешних частях шхер, в Ботническом зал. наоборот. Осенние сельди нерестятся дальше от берегов, у открытого моря, во внешних, реже во внутренних частях шхер, на глубине от 5 до 20 м. Летний нерест происходит также дальше от берегов, у открытого моря, во внешних частях шхер. Икра откладывается, в местах с твердым грунтом, каменистых и гравиевых. Нерест происходит при довольно низкой солености. При наиболее низком содержании солей размножаются весенние сельди, нерестящиеся ближе к берегам (фиордовые и ледовые сельди) — при 4—10‰. При более высокой солености (12—20‰) мечут икру весенние сельди открытого моря и осенние сельди. Осенние сельди нерестятся при более высокой (11—14°) температуре воды, чем весенние (6—10°). Период развития икринок при 14—15°равен 6—8 дням, при 15° — 5—7 дням. Выклюнувшиеся в последнем случае личинки имели размеры 6.0—6.5 мм. Таковы же были размеры личинок, выведенных из икринок, пойманных во внешних частях шхер северного берега Финского зал. к югу от Порккала 17 VII 1900 на глубине 4—10 м (Nordquist, 1901; Schneider, 1908). В зимнее время держится вдали от берегов. К берегам подходит весной, в Финском зал. начиная с середины мая или начала июня и после нереста отходит на большие глубины. В конце июня — начале июля вновь подходит к берегам, но не так близко, как весной (Михин и Антипова, 1932). В районе Койвисто (внешняя часть Выборгского зал.), кроме весеннего нерестового подхода, наблюдается осенний со второй половины августа по начало ноября. В этот период держится на глубине от 7—8 до 20 м при температуре до 15—17° (в сентябре) и солености до 5‰. Кроме того, салака подходит зимой с февраля, отчасти с января и держится по март, в некоторые годы до начала апреля. В это время она держится на склонах подводных гряд с придонной температурой воды 1—2° и соленостью 3—4‰ (Кожевников, 1949). В весенних подходах (в мае и июне) в восточной части Копорской губы в 1932 г. встречались рыбы в возрасте от одно- до восьмигодовалых, преобладали четырех- и пятигодовалые, в Курголове в весеннее время (в мае), а также и в начале осени (в августе) того же года преобладали трех- и четырехгодовалые, двух-, пяти- и шестигодовалых встречалось мало. При осенних подходах в Лужскую (Усть-Луга) и Копорскую губу в 1934 г. преобладали двухгодовалые, трехгодовалые встречались в небольшом количестве, в еще меньшем количестве — другие возрастные группы (Берг, 1940, по Михину). В восточной части Финского зал. основу питания составляет Eurytemora affinis var. hirundoides и ее яйца и сперматофоры, отчасти Bosmina obtusirostris var. maritima, Corophium, Mysis, Limnocalanus, Cyclops (Берг, 1940, по Шевчене). В желудках гигантских стремлингов, помимо беспозвоночных, из средней части моря находили также рыб (колюшку, молодь салаки или шпрота) длиной до 12 см (Schneider, 1908). В Финском зал. (у Бьёркэ) гигантские стремлинги весной питаются ракообразными и салакой длиной до 45 мм, осенью — колюшкой (преимущественно Gasterosteus aculeatus, реже Pungitius pungitius) длиной до 30 мм (Kanerva, 1931). В Ботническом зал. осенние сельди наиболее интенсивно питаются с июня по сентябрь включительно. Основу питания составляют Copepoda (Limnocalanus grimaldii), в средней части моря — Acartia longiremis, Acartia bifilosa и Temora longicornis и отчасти мизиды. В архипелаге островов против Стокгольма главную роль в питании играют Eurytemora hirundoides, Bosmina maritima и Daphnia cucullata; первый составляет основу питания зимой, два последних — летом и осенью. В меньших количествах в желудках были находимы Limnocalanus grimaldii и Mysis relicta. Кроме того, некоторую роль в питании крупных сельдей играют донные животные — Gammarus locusta, Chiridothea entomon и мелкие рыбы (Gasterosteus aculeatus и молодь Zoarces viviparus; Hessle, 1937). Половой зрелости большая часть весенних сельдей в архипелаге островов против Стокгольма достигает в двухгодовалом возрасте, причем большинство мелких сельдей в этом возрасте (длиной около 12 см) как правило еще незрелые. Неполовозрелых двухгодовалых встречается больше в возрастных группах урожайных лет (например 1932 г.), чем в менее урожайных (1933, 1934 и 1935 гг.; Hessle, 1937). У берегов Швеции размеры сельдей в уловах уменьшаются с юга на север. В северной части Ботнического зал. пяти- шестигодовалые сельди имеют размеры около 16 см, в южной части — около 17 см, у северных берегов средней части моря — около 18 см, у южных — около 19.5 см (Hessle, 1925). Осенние сельди составляют основу промысла, причем в средней части моря они в уловах обычно составляют больший процент, доходящий в нормальные годы до 90% и выше, чем в Ботническом зал.; весенние сельди — наоборот. В некоторые годы, однако, весенние сельди ловятся в значительном количестве, составив, например, в 1929 г. в Ботническом зал. свыше 30% улова (Hessle, 1931). В Финском зал. более многочисленны весенние сельди, чем осенние. В начале нерестовых подходов преобладают крупные сельди старших возрастных групп, которых сменяют более мелкие младших возрастных групп (Дементьева, 1947). После икрометания от берегов отходят, причем осенняя сельдь держится ближе к берегам, на мелководьях. Ранней весной и поздней осенью короткое время держится над большими глубинами, зимой опускается в придонные слои воды, в места с крутыми обрывами склонов или уходит в глубокие впадины шхер (Lundbeck, 1930). В Финском и Рижском заливах и по балтийскому побережью весенние и осенние сельди ловятся в двух- семигодовалом возрасте, преобладают трех- и четырехгодовалые, иногда двухгодовалые. Половой зрелости достигает к концу второго года, но в основной массе к концу третьего.
Рост весенней и осенней сельди в Финском зал. происходит следующим образом (Михин и Антипова, 1932; мм):


В Данцигской бухте рост обеих сельдей происходит следующим образом (Gieglewicz a. Posadzki, 1947; мм):
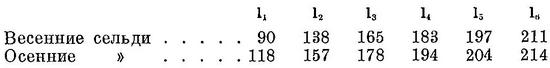
Рост в Рижском, а также и в Финском зал. более замедленный, чем в открытых частях моря (Дементьева, 1947). На границе распространения (у Карлсгамна) рост происходит следующим образом (Schneider, 1908; мм):
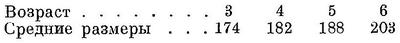
Гигантские стремлинги, пойманные у входа в Ботнический зал. (у Эрегрунд), имели такие размеры (Schneider, 1908; мм):

Пятигодовалые гигантские стремлинги, как видно из этих данных, значительно превосходят размеры салаки того же возраста. То же самое указывается и для Финского зал. Пятигодовалая зрелая самка гигантского стремлинга из Лужской губы имела размеры 317 мм, в то время как салака из этой губы в этом же возрасте достигает в среднем 174 мм (Берг, 1940). Четырехгодовалые гигантские стремлинги из окрестностей Бьёркэ имели размеры 240 мм (Kanerva, 1931). Содержание жира уменьшается к маю (до 3.2—4.1%), наибольшее наблюдается в сентябре (8.4—9.8%) (Левиева, 1950).
Хозяйственное значение. Главная промысловая рыба Балтийского моря, уловы которой составляют около половины улова этого моря и до 60% улова в водах СССР. Общий улов в некоторые годы превышал 1 млн. ц, в 1937 г. достигал 786.8 тыс. ц. В пределах СССР в 1936—1939 гг. уловы колебались от 210—220 тыс. ц, из них у берегов РСФСР добыто 39—53.5 тыс. ц, Эстонии — 85.6—101.9 тыс. ц, Латвии — 65.6—76.5 тыс. ц, Литвы — около 0.4 тыс. ц. У берегов Швеции добывается около 303—325 тыс. ц, Финляндии — 139—179 тыс. ц. Ловится круглый год, главным образом у берегов, ставными сетями, ставными и закидными неводами. Уловы могут быть увеличены организацией лова в открытом море.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Clupea harengus harengus Linne. — Мурманская сельдь, норвежская сельдь, атлантическая сельдь
(Норвежцы — sild, шведы — sill, немцы — Hering, англичане — herring, французы — hareng.)
Clupea harengus Linne, 1. с., p. 317 (Атлантический океан у берегов Европы).
Frühjahrsheringe von Norwegen Неinсkе, Naturgeschichte des Herings, 1898, p. LIX.
vaarsild норвежцев.
Clupea harengus harengus Берг, Рыбы пресн. вод России, М., 1923, стр. 500 (Кольский залив). — Рабинерсон, Изв. Отд. прикл. ихтиол., III, вып. 1, 1925, стр. 77 Кольский залив; расовая характеристика); там же, IV, вып. 1, 1926, стр. 127 (Кольский залив; возраст).
океаническая раса сельдей Аверинцев, Тр. Инст. рыбн. хоз., II, вып. 1, 1927, стр. 66 (Белое море: Кандалакша, близ устьев р. Онеги, Двинский залив).
сельдь Суворов, Изв. Гос. инст. опытн. агрон., VII, № 2, 1929, стр. 221 (Мурман: от Титовки на западе до Иоканги на востоке).
многопозвонковая сельдь, Clupea harengus Макушок, «За рыбную индустрию Севера», 1933, № 8, стр. 20 (западное побережье Канина, устья рек Кии, Шойны и Торны); там же, 1934, № 8, стр. 33 (устье р. С. Камбальницы на восточном побережье Каннна, устье р. Индиги, открытая часть Чошской губы).
Clupea harengus harengus Есипов, Промысловые рыбы Баренцова моря, Л., 1937, стр. 22. — Агапов и Топорков, Проблемы Арктики, № 2, 1937, стр. 104, рис. (Нов. Земля: Грибова губа, 4 сентября 1936 г.; Белужья губа, 29 сентября 1936 г.). — Есипов (ред.), Биология и промысел мурманской сельди, Л., 1939, 100 стр. — См. также ряд статей: Тр. Полярн. инст. морск. рыбн. хоз., I, 1938; II, 1938; IV, 1939; VI, 1939; VII, 1941. — Н. Дмитриев, Биология и промысел сельди в Белом м., М., 1946, стр. 3, 41 (Белое м.; длина 12—15 см, возраст l+, 2+).
Позвонков 56—59, наичаще 57 (в среднем 57.3). Брюшные килевые чешуи заметны как позади основания брюшных плавников, так и впереди. На сошнике сравнительно сильные зубы. Жаберных тычинок на 1-й дуге 61—72, в среднем 67.
Килевых чешуй между брюшными плавниками и анальным отверстием наичаще 13—14. В спинном плавнике наичаще 18 лучей (считая с неветвистыми), в анальном наичаще 17 (считая с неветвистыми). Антедорсальное расстояние в среднем 44.7% длины всего тела, длина D 10.2, длина А 10.7. Половозрелые норвежские сельди у берегов Норвегии достигают 300 мм и больше в длину; в Кольском зал. бывают сельди до 334 мм длиной.
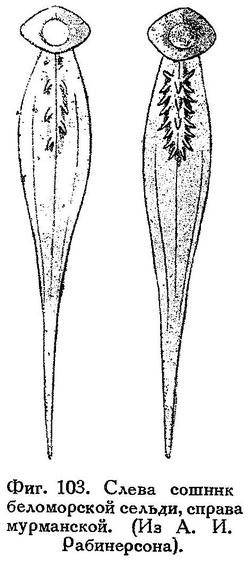
В. Н. Тихонов (Тр. Поляр, инст. рыбн. хоз., вып. 7, 1941, стр. 26) считает мурманскую сельдь за особое племя (natio murmanica Tichonov) атлантической сельди.
Мурманская сельдь распространена в Баренцевом море на восток до Чошской губы, устья р. Индиги, Колгуева и даже западной части Печорского залива (В. К. Есипов, 1937, стр. 25.); в 1936 г. она обнаружена у западного берега Новой Земли в районе Маточкина шара. В значительных количествах она встречается в Баренцовом море от Мурмана на север до 72° с. ш. и между 30° и 51° в. д. Исследованиями Полярного института обнаружено, что молодые особи мурманской сельди встречаются в открытой части Баренцова моря на север до 76° 48' с. ш. В области шпицбергенской ветви Гольфштрома эта сельдь достигает 77—78° с. ш. Мурманская сельдь нерестится на севере Норвегии — от Лофотенских островов до о. Сере, откуда течением мальков заносит на восток, в заливы Мурмана. (М. Рыженко, Личинки и мальки мурманской сельди, Рыбн. хоз., 1938, № 2, стр. 12—14. — Б. Мантейфель и Ю. Марти, Исследования нереста мурманской сельди, Тр. Полярн. инст. морск. рыбн. хоз., IV, 1939, стр. 41—92 (основная масса сельди Баренцова моря мечет икру на лофотенских нерестилищах, но, возможно, что, кроме того, есть менее значительные нерестилища в фиордах Норвегии и в заливах Мурмана). — Биология и промысел мурманской сельди, 1939, стр. 27—28.) Заходит также в Белое море (мелкие неполовозрелые экземпляры, в среднем длиной 14 см до конца средних лучей хвостового плавника), где встречается по Терскому берегу, в Кандалакшском, Онежском, Двинском заливах, а также у Соловков и в Мезенском заливе. Во время прилива сельдь эта в больших количествах входит в устья рек западного Канина (Макушок).
В Эльбу Clupea harengus иногда входит до Гамбурга (летом 1911 г.), т. е. на 70 км выше границы солоноватых вод (W. Schnakenbeck, Zool. Anz., vol. 94, 1931, p. 21.). В р. Шлей (Schley) в Шлезвиге сельди даже мечут икру (1. с., р. 23).
У сельдей типа Clupea harengus А. И. Рабинерсон при счете позвонков включал в число их и уростиль (Изв. Отд. прикл. ихт., III, вып. 1, 1923, стр. 70), как делал и Гейнке. Напротив А. И. Амброз (Изв. Тихоок. инст. рыбн. хоз., VI, 1931, стр. 12) уростиль не считал. Поэтому данные этого автора следует увеличить на 1: вместо 53.2—53.7 нужно 54.2—54.7.
Clupea harengus membras Linne. — Салака
(Ижоры — aili, финны—haili, silli, эсты — räim, немцы — Strömling, шведы — strömming.)
Clupea harengus β membras Linne, Fauna suecica, ed. 2, Stockholm, 1761, p. 128 («Strömming; vero minor ex mari Bothnico»). — Pallas, Zoogr. rosso-asiat., III, 1811, p. 211 (частью: Балтийское море, особенно у берегов Латвии и у о. Эзеля).
Clupea harengus Кесслер, Рыбы СПб. губ., 1864, стр. 185 (частью: близ устья Наровы, Кронштадт). — О. Rеutеr, Meddel. Soc. fauna et flora fenn., XX, 1894, p. 52 (пресные озерки на островах Абоского архипелага).— Smitt, Scandin. fish., II, 1895, p. 954 (частью: «var. membras», p. 960).
Frühjahrsheringe der östlichen Ostsee oder Strömlinge Heineke, Naturgeschichte des Herings, 1898, p. LXIV (восточная часть Балтийского моря), рисунки сельдей из Стокгольма и Мемеля.
Clupea harengus membras G. Schneider, Rapp. et proc.-verb. du Conseil perm. pour l'explor. de la mer, IX, 1908, p. 77 (к востоку от меридиана о. Готланда). — Суворов, Тр. Балт. эксп., II, 1913, стр. 44. — Рабинерсон, Изв. Отд. прикл. ихтиол., III, вып. 2, 1925, стр. 134 (Копорская губа, Шепелево). — Михин и Антипова, Изв Инст. озерн. и речн. рыбн. хоз., XV, 1932, стр. 28 (восточная часть Финского залива; в промысловых количествах от меридиана западной оконечности о. Котлина), стр. 92—109. — Берг, там же, XXIII, 1940, стр. 6 (Финский залив).
Салака отличается своей малой величиной, обычно менее 200 мм (абсолютной длины). В восточной части Балтийского моря половозрелые самцы наблюдались, начиная от 132 мм абсолютной длины, самки от 143 мм (Суворов, стр. 68). Средняя длина салаки из Копорской губы зимней 158, летней 168 мм (абс.).
Впрочем иногда попадаются так наз. гигантские стремлинги (Riesenströmlinge), достигающие 330 и даже 375 мм (такие особи ловятся и в Финском заливе и, между прочим, близ Кронштадта и устья Наровы). Эти гигантские сельди, по Гейнке и Шнейдеру, представляют собой лишь особей обыкновенной салаки (восточно-балтийской сельди). Если это справедливо для гигантских салак, ловимых у Хельсинки, то в отношении крупных сельдей восточной части Финского залива дело, повидимому, обстоит иначе. Салака со зрелой икрой, длиной 317 мм (абс.), добытая О. П. Антиповой в Лужской губе у Усть-Луги 10 мая 1930 г., имела 5 лет. В этом возрасте обычная салака из Лужской губы достигает в среднем всего 185 мм и даже в возрасте 7 лет имеет менее 200 мм. Таким образом, гигантская салака есть особая быстрорастущая раса. Такого же мнения держится и Канерва (R. Kanerva, Finlands Fiskerier, XI (1930), 1931, p. 114—117.), исследовавший гигантских салак из окрестностей Бьёркё, где они изобилуют: они тоже отличаются быстрым ростом, достигая в 3-летнем возрасте такой же длины, как обыкновенные салаки в 6-летнем; четырехгодовалая гигантская салака имела в длину 240 мм. У Бьёркё эти салаки мечут икру весной, в мае. Осенью, у Бьёркё, они подходят к берегу, и здесь их промышляют совместно с трехиглой колюшкой, которою, как установлено путем вскрытий, гигантская салака в это время питается.
Кроме небольшого роста салака отличается малым числом позвонков, в среднем по Гейнке 55.5—55.0 (25 сельдей, пойманных в апреле у Хельсинки, имели в среднем длину 184 мм (абс.) и 55.3 позвонка: Heincke, табл., р. 174; 60 сельдей из шхер у Стокгольма 55.1 позвонков: G. Schneider, р. 81; 50 сельдей, добытых в первой половине мая 1930 г. в Лужской губе у д. Усть-Луга О. П. Антиповой, имели длину (абс.) 135—242 мм, позвонков 54—56 (в среднем 55.15). Число килевых чешуй за брюшными плавниками мало, в среднем от 12.4 до 14.0. Голова и хвост длинные. Жаберных тычинок наичаще 65—69.
В восточной части Финского залива есть несколько рас, различимых по времени нереста. У одних икрометание происходит в конце мая — начале июня, когда эта сельдь массами подходит к берегу (Михин и Антипова). Помимо этой расы, есть салака, нерестующая в конце июня — начале июля. Наконец, в последних числах августа 1932 г. С. М. Сорокин наблюдал в Кургалове (южный берег Финского залива) половозрелых салак.
Среди салаки можно отличить две крайних формы, большеглазую (диаметр глаза 5.7—6.0% длины всего тела) и малоглазую (5.0—5.2%).
Салака (стремлинг) распространена в восточной части Балтийского моря, к востоку от линии: южная оконечность о. Эланд — Данцигская бухта, а также в Финском и Ботническом заливах до крайних концов их. В Финском встречается на восток до Сестрорецка. Есть в заливах Нарвском, Лужском и Копорском. Здесь в 1930 г. преобладали особи в возрасте 3+, длиною в среднем 140—160 мм (абс.), хотя встречались особи от 2+ до 7+ (В. С. Михин).
Иногда живет и в совершенно пресной воде. Так, по данным Рейтера (Reuter), она водится в некоторых пресных озерках на островах Абоского архипелага (именно, Björkö в Korpo, Medelby в Houtskär, Högsar в Nagu). О таких фактах упоминают также Mobius и Heincke. Со слов Трибома (Trybom) G. Schneider (p. 77—78) передает, что в Швеции, в провинции Kalmarlän, к северу от устья р. Вирбо, близ Ytterhult, салака живет в пресном озере, соединенном ручьем с морем. Во время северных ветров была находима в р. Латвийской Аа в 9 км выше устья (W. Mannsfeld, Korr.-Bl. Naturf. Ver. Riga, LXIII, 1939, p. 43.).
Нерестится на твердом грунте, на глубине чаще всего 7—10 м. Плодовитость весенней салаки в восточной части Финского залива в среднем 10.6 тыс. икринок. Половозрелость наступает в возрасте 2—3 лет. Основную массу в уловах составляют особи 2—4 лет, длиной 14—16 см, в среднем 15.4 см, весом 24 г (П. В. Тюрин).
В 1935 г. во всем Финском заливе добыто 150 тыс. ц салаки.