Clupea pallasii pallasii Valenciennes, 1847
Тихоокеанская сельдь, Восточная сельдь, Восточная селедка, Малопозвонковая сельдь, Сельдь Паласса, Дальневосточная сельдь, Беринговоморская сельдь, Анадырская сельдь, Камчатская сельдь, Южнокамчатская сельдь, Северокамчатская сельдь, Восточнокамчатская сельдь, Гижигинская сельдь, Гижигинско-камчатская сельдь, Корфокарагинская сельдь, Олюторская сельдь, Жупановская сельдь, Корейская сельдь, Сельдь залива Петра Великого, Пластунско-нельминская сельдь, Декастринская сельдь, Южносахалинская сельдь, Южносахалинско-хоккайдская сельдь, Западносахалинская сельдь, Хоккайдская сельдь, Сахалино-хоккайдская сельдь, Охотская сельдь, Охотско-хоккайдская сельдь, Тохокуккайская сельдь, Восточносахалинская сельдь, Пенжинская сельдь, Чосенская сельдь, Калифорнийско-вашингтонская сельдь, Хинумская сельдь, Сельдь лимана Аккеси, Тоннайская сельдь, Тихоокеанская озёрная сельдь, Клупея Паласса, Клупея палласи палласи (Rus),
Pacific herring, North Pacific herring, Herring, Blue herring, Oriental herring, Blue back
(Eng).
Синонимы:
Clupea harengus pallasi Valenciennes, 1847 Тихоокеанская сельдь (Rus),
Clupea harengus pallasi mirabilis Rabinerson, 1928
Clupea inermis Basilewsky, 1855
Clupea lineolata Valenciennes, 1847
Clupea mirabilis Girard, 1854 Калифорнийско-вашингтонская сельдь (Rus),
Clupea pallasii Valenciennes, 1847 Восточная сельдь, малопозвонковая сельдь, сельдь Паласса (Rus),
Spratelloides bryoporus Cope, 1873
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Сельдь тихоокеанская — Clupea pallasi pallasi Valenciennes, 1847
Длина головы составляет менее 21% длины тела, отличается маленькими глазами. Зоопланктофаг. Образует ряд подвидов и локальных стад. Достигает длины до 50 см. Обычна в водах Северного Ледовитого и Тихого океанов от устья Лены до Кореи. Объект промышленного рыболовства.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
В океанариумах Японии представлены:
Сельдь Паласса — Clupea pallasi (Cuvier — Valenciennes, 1847), 25—32 см.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.2.1 (3). Clupea harengus pallasi Valenciennes, 1847 — Тихоокеанская сельдь (Св., 1952 : 145).
[1] русск. Белая рыбка — Гр. : 215. Бельчуг камч. — Кл. Восточная селедка — А. Ник. : 802. Восточная сельдь — Б., 1932 : 97. Дальневосточная сельдь — Г. Ник., 1954:114. Килька (мелк.) ДВ: р. Суйфуи — Дулькецт, 1925 : 68. Корейская сельдь — Шмидт, 1948 : 36. Малопозвонковая сельдь — Пром. р. : 127. Сельдь ДВ: р. Суйфун — Дулькейт, 1925 : 68. Тихоокеанская сельдь — Линдб. и Тар. : 226. Хоккайдская сельдь, Чосенская сельдь — Шмидт, 1948 : 35. ~ Бельчуг — САР. Белая рыбка, Бельчуч, Сельдь — Краш., 1 : 326, 346; Зуев : 69. [18] англ. Herring Америка — Пром. р. : 127. Pacific herring Канада — Ricker : 111. [70] ительм. Nepn-hep — Pallas : 204.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Восточная, или малопозвонковая, сельдь
(Clupea pallasi) распространена от
Белого моря на восток. Она обычна в юго-восточной части Баренцева моря, в
Чешской губе, в Печорском заливе; гораздо
менее многочисленна в южных районах
Карского моря. У берегов Сибири
известны небольшие популяции, приуроченные
к предустьевым пространствам рек.
В Тихом океане численность восточной
сельди очень велика. Сельдь является
здесь важным объектом промысла,
будучи распространена по азиатскому
побережью до Желтого моря, а по
американскому до Калифорнии (Сан-Диего).
Распространение ограничивается
прибрежными водами. Почти весь ареал этого
вида в зимнее время покрывается льдами.
В отличие от атлантической сельди
восточная размножается в пределах всего
ареала. В южных районах она
нерестится в самое холодное время года подо
льдом или же сразу после разрушения
его.
Восточная сельдь существенно
отличается по своей биологии от
атлантической. Размножение ее происходит на
мелководьях, иногда почти у самого
уреза воды, от глубины 0,5 м, главным
образом на глубине 3—4 м, и не глубже
10—15 м. Сельдь подходит для нереста
к берегам при температуре воды от
0,5° С (иногда даже при отрицательной
температуре) и до 8—10,7° С; основной
ход происходит при 3—9° С. Она откладывает икру преимущественно в
укрытых от ветра местах, на подводную
растительность — зостеру, фукусы и
другие растения. Плотность засева икрой
у Южного Сахалина обычно была 2—6 млн. икринок на 1 м2. Восточная
сельдь выносит значительное
опреснение, поднимаясь в устья рек и встречаясь
и в осолоненных озерах, но в совершенно
пресной воде погибает. Взрослая рыба не
совершает таких больших миграций,
как атлантическая сельдь, ограничиваясь
местными передвижениями
преимущественно к берегам из открытого моря и
от берегов. Характерно для восточной
сельди меньшее число позвонков, чем
у атлантической: обычно 52—55 (до 57).
У восточной сельди обычно зубы на
сошнике слабее развиты, чем у
атлантической.
Различают три подвида восточной
сельди: беломорскую сельдь,
чешско-печорскую сельдь и тихоокеанскую сельдь.
Эти подвиды, особенно беломорская
сельдь, распадаются на особые расы или
формы.
Тихоокеанская сельдь (Clupea pallasi pallasi) особенно многочисленна у
восточных берегов Камчатки, в Охотском море,
у берегов Южного Сахалина, у острова
Хоккайдо. По восточному побережью
сельдь имеет важное значение для
промысла в заливе Кука, в фиордах Южной
Аляски и у острова Ванкувер.
Тихоокеанская сельдь достигает длины
50 см, средние размеры нерестовой
рыбы — 24—38 см. Позвонков 41—57.
Распадается на ряд форм, среди которых
различают собственно морских,
размножающихся в море у берегов, и озерных, заходящих
для нереста в осолоненные озера и бухты
с пониженной соленостью. Всего
различают 10—12 местных форм, или стад,
морских сельдей и 3 формы озерных. Нерест
происходит в разных районах в разное
время: в Анадырском лимане в июле,
на севере Охотского моря с мая по июль,
у Восточной Камчатки в мае, в Северном
Приморье в мае — июне, в Южном
Приморье и у Южного Сахалина с марта по
май. У американских берегов нерест
происходит в несколько иные сроки: у о-ва
Кодьяк в мае — июне, у Юго-Восточной
Аляски в марте, у Британской Колумбии
и Калифорнии с декабря по апрель.
Весенние подходы сельди к берегам
происходят в несколько (2—4) последовательных
волн (ходов): вначале подходит более
крупная рыба, затем более молодая. По
окончании нереста сельдь отходит от
берега для нагула. Нагульная, или
жирующая, сельдь подходит к берегам для
откорма в летнее время, совершая здесь
суточные вертикальные миграции.
Различают периоды весеннего, или
преднерестового, откорма (апрель — май),
нерестового голодания (май — июнь), летнего
откорма (с конца июня по август) и
зимнего ослабления питания. Основу питания
составляют евфаузиевые рачки, каланусы,
черви-стрелки. Взрослая сельдь
нагуливает до 18,7—25,7% жира, мелкая — до
23—32% жира. Особой жирности — 20—33% жира — достигала крупная летне-осенняя (ловившаяся с июля по октябрь)
восточнокамчатская «жупановская» сельдь,
имевшая в длину 34—42 см.
Промысел тихоокеанской сельди
производится преимущественно неводами у
берегов.
Численность тихоокеанской сельди
подвержена еще более резким колебаниям,
чем численность атлантической сельди.
Например, очень большой численности
достигала в первой трети нашего века
раса сахалино-хоккайдской сельди.
Подходы сельди к берегам Сахалина
представляли грандиозное явление.
Обитатели бездны. Уильям Кроми 1971 г.
Типичный житель открытого моря, где обитают многие виды рыб, маленькая сельдь, пожалуй, самая многочисленная рыба в приповерхностных слоях воды. Атлантическая сельдь и ее тихоокеанская сестра Clupea pallasi передвигаются огромными многомиллиардными косяками, которые подчас достигают длины в несколько миль и такой же ширины. Иногда кажется, что море битком набито их серебристыми, длиной сантиметров в 30 телами.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
1. Clupea harengus pallasi Valenciennes, 1847 — Тихоокеанская сельдь (рис. 71).
Clupea harengus (non Linne) Pallas, Zoogr. rosso-asiat., 3, 1811 : 209, 210 (Камчатка).
Clupea pallasii Valenciennes in: Cuvier et Valenciennes, Hist. Nat. Poiss., 20, 1847 : 253 (Камчатка).
Clupea harengus pallasi, Световидов, Фауна СССР, Рыбы, 2, 1, 1952 : 145, табл. 3, фиг. 2 (синонимия, описание).
Clupea pallasi, Чжан и др., Рыбы зал. Бохай, Желт. море, 1955 : 46, fig. 31 (Чжифу). — Matsubara, Fish Morphol. a. Hierar., 1955 : 190. — Суворов, Промысловые рыбы СССР, 1949 : 127—130; атлас, табл. 32 (цветной рисунок).

D (III) IV (V) 13—15, A III (IV) 12—16, жаберных тычинок 63—73, килевых чешуй позади основания брюшных плавников 11—13, позвонков 53—55. Брюшные килевые чешуи более или менее развиты лишь позади основания брюшных плавников, впереди от них развиты слабо или
отсутствуют. Голова небольшая и широкая, длина ее 19.8% длины тела; межглазничный промежуток 17.2% длины головы. Глаза небольшие, 21.2% длины головы. Основание анального плавника сравнительно короткое, 10.2% длины тела (Антоново, западное побережье Сахалина — 50 экз.). (Световидов, 1952а : 145).
Пелагическая рыба, нерестующая у берегов Японского моря с марта (Посьет) до июня (Де-Кастри). Сводка по ее биологии дана в работе А. Н. Световидова (19526). За последние годы появились новые русские работы по размножению, поведению и распределению сельди (Веденский, 1957; Галкина, 1957, 1958, 1960а, 19606, 1961, 1962; Дружинин, 1957, 1959; Кагановский, 1954, 1955; Крыжановский, 1955, 1956; Моисеев, 1952, 1956; Пискунов, 1952а, 19526; Покровская, 1954а, 19546, 1955; Пробатов, 1954, 1956, 1957; Пробатов и Дарда, 1957; Пробатов и Козлов, 1954; Пробатов и Фридлянд, 1957; Световидов, 19526; Соколов, 1962; Фридлянд, 1949). Укажем также некоторые другие работы: Hirano, 1959; Kurata, 1959; Mikami а. Таmura, 1961; Tamura et all., 1954; Tchang, 1957; Yamamoto, 1955a, Yanagimachi, 1957a, 1957b, 1957c, 1957d, 1957e, 1957f, 1958.
Тихоокеанская сельдь — важный промысловый объект. В настоящее время ее запасы значительно уменьшились. В СССР заготавливается в соленом и свежезамороженном виде. В Японии много выловленной тихоокеанской сельди идет на приготовление жира и тука (удобрение для полей) и заготавливается в сушеном виде.
Длина до 500 мм.
Распространение. В Японском море известен на юг до Пусаня, р-на Санин и о-вов Оки (Mori, 1952 : 31, 1956 : 5). Редко встречается в Желтом море у западных берегов п-ова Корея (Mori, 1952 : 31) и пров. Шаньдун — Чжифу (Чжан и др., 1955 : 46). Вдоль тихоокеанского побережья Японии проникает на юг до Хинума, пров. Хитати (Tanaka, 1928 : 844). Северная часть Тихого океана от Анадыря на юг до Японского и Желтого морей и Калифорнии. Северный Ледовитый океан. В Белом море раса maris-albi Berg, 1923.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1б. Clupea harengus pallasi Valenciennes — Тихоокеанская сельдь, восточная сельдь.
Clupea harengus (non L.) Pallas, Zoogr. Rosso-Asiat., III, 1811 : 209 (Камчатка). — Richardson, Fauna Bor.-Amer., 1836 : 231 (Bathurst Inlet в арктич. Америке около 67° с. ш. и 109° з. д.; описание экз. длиной 15 дюймов).
Clupea pallasi Cuvier et Valenciennes, Hist. Nat. Poiss., XX, 1847 : 253 (Камчатка; по экз. Палласа). — Jordan a. Evermann, Fish. N. America, I, 1896 : 422 (от Камчатки до Сан-Диего). — Schnakenbeck, 1931 : 539— 551, 561 и др. (число позвонков, килевых чешуек). — Rounsefell a. Dаhlgreen, Bull. U. S. Bur. Fish., XLVIII, 1935 : 119 (юго-вост. Аляска; расы, биология). — Tester, Journ. Biol. Board Canada, III, 2, 1937 : 108 (Канада; морфометрия, возраст).
Clupea mirabilis Girard, Proc. Ac. Nat. Sci. Phila., VII, 1854:138 (Сан-Франциско). — Turner, Contr. Nat. Hist. Alaska, 1886 : 111, pl. 14 (зал. Нортон; нерест в середине июня; хороший рис.).
Clupea harengus pallasi Шмидт, Рыбы вост. морей, 1904 : 258 (сходство с беломорской сельдью). — Амброз, Изв. Тихоок. научн. инст. рыбн. хоз., VI, 1931 : 313 стр. (зал. Петра Великого). — Агапов, Тр. Научно-иссл. инст. полярн. землед., сер. «Пром. хоз.», 15, 1941 : 101 (Анадырск. лиман; биометрика, биология). — Берг, 1948 : 155. — Суворов в изд.: Промысл. рыбы СССР, 1949 : 127, атлас табл. 32 (сводка, цветн. рис.). — Панин, Изв. Тихоок. научно-иссл. инст. рыбн. хоз. океаногр., XXXII, 1950 : 3—36 (вост. Камчатка; морфометрия, биология, промысел). — Световидов, 1952 : 145, табл. III (диагноз южносахалинск. сельди, библиография).
Clupea harengus pallasi. В последних выпусках «Известий Тихоокеанского института рыбного хозяйства и океанографии» (ТИНРО) опубликован ряд важных работ по биологии тихоокеанской сельди: А. Г. Кагановский и И. А. Полутов, Изв. ТЦНРО, XXXII, 1950 : 37—53 (Пенжинск. зал.). — А. П. Веденский, там же : 55— 63 (вост. Сахалин). — А. И. Фролов, там же : 65—71 (локальные формы сахалинской сельди). — В. Т. Богаевский, там же, XXXIV, 1951 : 5—24 (жирующая сельдь юго-зап. Сахалина). — А. Н. Пробатов и А. И. Варварин, там же : 25—39 (молодь в зал. Анива). — Г. И. Грищенко, там же : 41—46 (лов на свет). — К. И. Панин, там же : 257—259 (нагульная сельдь у вост. Камчатки). — И. Г. Фридлянд, там же, XXXV, 1951 : 105—145 (юго-зап. Сахалин; биология размножения). — И. А. Пискунов, там же, XXXVII, 1952 : 3—67 (юго-зап. Сахалин), и др.
Распространение, биология. Nelson, Nat. Hist. Coll. Alaska, 1887 : 320 (зал. Нортон; условия нереста). — Scofield, Fish. Arct. Alaska, 1899 : 494 (Порт-Кларенс; обычна после вскрытия льда). — Evermann a. Goldsborough, Fish. Alaska, 1907 : 232 (местонахождения на Аляске). — Сокольников, Тр. Общ. судох., Пром. отд., VI, 1911 : 52 (Анадырск. лиман). — Андрияшев, Иссл. морей СССР, 25, 1937 : 295 (Авачинск. зал.; в желудке стрелозубого палтуса, на глуб. 150 м). — Tester, Journ. Cons., XIII, 1, 1938 : 71 (число позвонков и температура воды во время развития). — Полутов, Костюченко и Панин, Рыбн. хоз., 3, 1941 :3 (Камчатка; промыслово-биологич. очерк). — Панин и др., Календарь рыбака Камчатки, 1945 : 13 (дл. до 42 см, вес до 800 г). — Clemens a. Wilby, Fish. Pacif. Coast Canada, 1946 : 79, fig. 33 (Канада). — Панин, Зоолог. журн., XXV, 5, 1946 : 469 (связь нерестовых подходов с приливами; см. также: Природа, 1, 1947 : 68). — Пирожников, Докл. АН СССР, LVI, 4, 1947 : 441 (близ бухты Тикси 3—26 августа; самки со зрелой икрой). — Пирожников, Задачи научно-исслед. организ. в четвертой пятилетке в развитии рыбн. хоз. Сибири, Изд. Главсибрыбпром, Новосибирск, 1948 : 9 (Оленекск. зал.). — Кун, Изв. Тихоок. научно-иссл. инст. рыбн. хоз. океаногр., XXIX, 1949 : 164 (питание). — Городничий, Рыбн. хоз., 1, 1949 : 36 (влияние потепления на уловы).
В последних выпусках «Известий Тихоокеанского института рыбного хозяйства и океанографии» (ТИНРО) опубликован ряд важных работ по биологии тихоокеанской сельди: А. Г. Кагановский и И. А. Полутов, Изв. ТИНРО, XXXII, 1950 : 37—53 (Пенжинск. зал.). — А. П. Веденский, там же : 55—63 (вост. Сахалин). — А. И. Фролов, там же : 65—71 (локальные формы сахалинской сельди). — В. Т. Богаевский, там же, XXXIV,
1951 : 5—24 (жирующая сельдь юго-зап. Сахалина). — А. Н. Пробатов и А. И. Варварин, там же : 25—39 (молодь в зал. Анива). — Г. И. Грищенко, там же : 41—46 (лов на свет). — К. И. Панин, там же : 257—259 (нагульная сельдь у вост. Камчатки). — И. Г. Фридлянд, там же, XXXV, 1951 : 105—145 (юго-зап. Сахалин; биология размножения). — И. А. Пискунов, там же, XXXVII, 1952 : 3—67 (юго-зап. Сахалин), и др.
D 17—20, среднее 18.7, А 15—19, ср. 17.1, жаберных тычинок 63—73, ср. 67.0, килевых чешуек позади основания брюшных плавников 11—13, ср. 11.8, позвонков 53—55, ср. 54.1 (по данным Световидова для 50 южносахалинских сельдей длиной 31.8—36.1 см). По Панину, проанализировавшему большой материал по восточнокамчатской сельди, позвонков (51) 52—55 (56), очень редко 49—50, как исключение 57 позвонков (у 3 сельдей из 3044); среднее число у сельди Авачинского зал. 54.2, у сельди с северо-восточного побережья Камчатки (заливы Уала и Корфа) 53.8—53.9. Еще меньше позвонков у анадырской сельди — 52—55, ср. у 50 экз. 53.1 (Агапов). Наиболее низкие цифры по числу позвонков дает калифорнийско-вашингтонская сельдь (mirabilis) — (50) 51-53 (54).
Килевые чешуйки более или менее развиты только позади основания брюшных плавников, впереди них они мало заметны или отсутствуют. Зубы на сошнике хуже развиты и менее многочисленны, чем у атлантической Clupea harengus; расположены в 1—2 коротких рядах (средний ряд зубов обычно не развит). Голова небольшая и широкая: ее длина, по Световидову, составляет 19.0—20.8, в среднем 19.8% L., а межглазничный промежуток — 15.7—18.6, в среднем 17.2% длины головы. Длина на Камчатке до 42 см и вес до 800 г, по данным Павленко — до 45—50 см.
Тихоокеанская сельдь, подобно атлантической, образует ряд локальных форм с изолированными нерестилищами. Только в западной части Тихого океана в настоящее время насчитывается не менее 10 рас морских сельдей и несколько озерных (подробнее см. у Световидова).
Распространение. Северная часть Тихого океана, частично арктические моря. От Берингова пролива (Порт-Кларенс) доходит до южн. Калифорнии (Сан-Диего); у азиатских берегов от западного побережья Кореи и Цусимского прол. до Анадырского лимана, где обычна. У берегов Чукотского полуострова до последнего времени известна не была, однако в коллекциях Тихоокеанского института рыбного хозяйства имеются 2 сельди из бухты Провидения (абсолютная длина 18.5 и 22 см), добытые П. Г. Никулиным 3 июля 1949 г. Экземпляров из Чукотского и Восточно-Сибирского (Ф. Н. Кириллов (со слов рыбаков) сообщил мне о нахождении близ дельты р. Индигирки трех рыб, «похожих на ряпушку, но без жирового плавника и с большим глазом»; это могла быть только тихоокеанская сельдь.) морей нет, однако в последние годы (1940—1946) найдена в море Лаптевых (бухта Тикси, Оленекский зал.); близкая, если не тождественная, форма обитает в Карском море (Обская и Карская губы), а также в юго-восточной части Баренцова моря (n. suworowi) и в Белом море (крупная раса). Относительно северных берегов Аляски достоверных данных мало; Скофилд, со слов местных жителей, указывает сельдь для устья Мэккензи, но сам он ее там не нашел. Карта А. Мика (A. Meek. The migrations of fish. 1916 : 104, fig. 38.), на которой сельдь также показана почти до Мэккензи, не документирована. Первым точным указанием для арктического побережья Сев. Америки следует считать сообщение Ричардсона, который во время первой полярной экспедиции Дж. Франклина 5 мая 1821 г. добыл (и описал) несколько экземпляров сельди близ устья р. Медных Рудников (109° з. д.). В этом же районе и западнее сельдь хорошо ловилась в 1914—1916 гг.
Биология. Жизненный цикл тихоокеанской сельди больше, чем у атлантической Clupea harengus, связан с прибрежными и опресненными водами. Она не только часто заходит в сильно опресненные заливы и устья рек, но и образует местные формы, постоянно зимующие и нерестующие в озерах-лагунах (озера Халыгер и Нерпичье на Камчатке, оз. Тоннай на Сахалине, озера на о. Хондо и др.). Нерест происходит по всему ареалу от Кореи и Калифорнии до самого Берингова пролива; сельдь, недавно обнаруженная в море Лаптевых, также имела зрелую икру. Все формы тихоокеанской сельди относятся к биологическому типу весенне-нерестующих, разница же в сроках нереста в Тихом океане с декабря по июль, в арктических морях и в августе) связана главным образом с географическим положением и температурными и ледовыми условиями на нерестилищах. Икра откладывается на растительность (как исключение на гальку) в узкой прибрежной зоне (от уреза воды до глубины 5—10 м). В разных широтах Берингова моря размножение происходит с мая по июль: у северо-восточных берегов Камчатки (Анапка, Уала) — во второй половине мая (Панин, 1950), в Анадырском лимане сельдь с текучими половыми продуктами регулярно ловится в июле, когда, по предположению Агапова, нерест заканчивается. В зал. Нортон, омываемом идущим вдоль берегов Аляски теплым течением, нерест происходит немного раньше, — подход нерестовых стай отмечался с 9 по 15 июня (1877—1879 гг.). В море Лаптевых близкие к нересту особи ловились в августе (Пирожников). Размножение у восточных берегов Камчатки детально описано Паниным (1950). Нерестовые подходы здесь тесно связаны с приливо-отливными явлениями; массовый подход, по данным Панина (1946), «происходит в период развития сизигийной фазы больших колебаний уровней в начале—середине мая и протекает во времени от первой четверти луны через полнолуние до последней четверти». Нерест происходит в прибрежной зоне (от уреза воды до 3—4 м) при температуре 3—6° и значительном опреснении (20—23‰). Икра выметывается за один прием (не порционно), как правило на заросли зостеры. Развитие икры при температуре 6—8° продолжается около 23 дней; выклюнувшиеся личинки имеют длину 7—8 мм. Массовый нерест в зал. Нортон, по наблюдениям Тэрнера, происходит в сходных условиях. Икра откладывается в прибрежной зоне на водоросли (фукоиды?) при сравнительно высокой температуре (в разгар нереста в июне при 6.7°). Отход икры в результате приливо-отливных колебаний уровня достигает 25%. Плодовитость камчатской сельди колеблется от 12.6 до 122.8 тыс. икринок, в среднем в возрасте 4 года — 34 тыс., у 6-годовалых — 57 тыс., у 8-годовалых — 77 тыс., у 10-годовалых — 101 тыс. икринок. Половозрелости камчатская сельдь достигает в основном в возрасте 4—5 лет, основную массу нерестующих особей составляют 5—8-годовалые особи (Панин, 1950). О молоди и периоде ее роста данные для беринговоморской сельди недостаточны. Повидимому, она не совершает значительных меридиональных миграций, ограничиваясь отходами в море и подходами к берегам (молодь, жирующая сельдь, нерестовые подходы и т. д.). Рост у северо-восточных берегов Камчатки (Анапка, Уала) приводим по данным Панина (1950) за 1939—1948 гг. (длина до конца средних лучей С в см, вес в г):

Продолжительность жизни камчатской сельди 15 лет, в уловах обычно преобладают 5—10-летние сельди (средняя длина до выемки С 33.5 см, средний вес 419 г). В питании основную роль играют Calanoida и Euphausiidae, а также Sagitta, Themisto, Mysidae и др. М. С. Кун различает следующие периоды в питании сельди (Татарский пролив): 1) весенний или преднерестовый откорм (апрель—май), 2) нерестовое голодание (май— июнь), 3) интенсивный летний откорм (конец июня—август) и 4) осеннее ослабление питания (зимний период не изучен). Наибольшей жирностью отличаются крупные камчатские сельди (например так называемые жупановские сельди), имеющие в летне-осеннее время до 33% жирности.
Хозяйственное значение меньше, чем атлантической сельди, хотя уловы в урожайные годы превышают 6 млн ц. Наибольшие уловы дает нерестовая сельдь южн. Сахалина. Промысел на Камчатке развился лишь в последние 10—12 лет (150—200 тыс. ц) и имеет значительные перспективы для роста. Некоторое промысловое значение сельдь может иметь и в Анадырском лимане, где она обычна с конца июня по август. Агапов сообщает, что близ р. Туманской (южнее Анадырского лимана) в 1936 г. в кетовый ставной невод было поймано 31.5 ц сельди.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*lc. Clupea harengus pallasi Val. — тихоокеанская, восточная сельдь (табл. III).
— Clupea harengus Pallas, Zoogr. rosso-asiat., III, 1811 : 209, 210 (частью: «in Oceano lotam Camtschatcam alluente et in sinu Ochotensi»; p. Камчатка).
— Clupea pallasii Cuvier et Valenciennes, Hist. nat. poiss., XX, 1847 : 253 (Камчатка).
— Clupea mirabilis Girard, Proc. Acad. Nat. Sci. Phila., L, 854 : 138 (Сан-Франциско).
— Clupea inermis Basilewsky, Nouv. Mem. Soc. Nat. Moscou, X, 1855 : 242 (Шаньдун в Чжилийском зал. Экземпляр сельди, доставленный Базилевским, хранится в коллекциях Зоологического института Академии Наук СССР (№ 5655, China sept.).).
— Spratelloides bryoporus Cope, Proc. Amer. Phil. Soc., XIII, 1873 : 25 (Ситка, молодь).
— Clupea harengus var. pallasi Шмидт, Рыбы восточных морей, 1904 : 258.
— Clupea harengus pallasi Берг, Рыбы пресных вод России, 1923 : 502.
— Clupea pallasii Нubbs, California Fish a. Game Commiss., Fish Bull. 8, 1925 : 5 (Брит. Колумбия, Калифорния; количества позвонков); Fujitа а. Коkubo, Proc. Third Pan-Pacific Sci. Congr., Tokyo, 1926 : 2253; Bull. Scool Fish., Hokkaido Imp. Univ., Sapporo, I, 1, 1927 : 1—127 (берега Японии; расы).
— Clupea harengus pallasi Рабинерсон, Изв. Тихоокеанск. научно-промысл. ст., I, 1, 1928 : 143, 156 (зал. Петра Вел.; биометрика).
— Clupea harengus pallasi mirabilis Рабинерсон, там же: 156 (берега Сев. Америки).
— Clupea harengus pallasi Rabinerson, Zool. Jahrb., Abt. Syst., Bd. 58, 1929 : 292, 301.
— Clupea harengus pallasi mirabilis Rabinerson, l. c. : 293.
— Clupea pallasii Rounsefell, Bull. U.S. Bureau Fisher., XLV, (1929), 1930 : 227 (берега Сев. Америки; расы, места нереста, рост, уловы).
— Clupea harengus pallasi Амброз, Изв. Тихоокеанск. научн. инст. рыбн. хоз., 6, 1931 : 1 (зал. Петра Вел.; возраст, рост, нерест, миграции, питание).
— Clupea pallasii Rounsefell a. Dahlgrеn, Bull. U. S. Bureau Fisher., XLVIII, 1935 : 119 (юго-восточная Аляска; расы, количества позвонков, рост, мечение); Tester, Journ. Biol. Board Canada, III, 2, 1937 : 108 (берега Брит. Колумбии; число позвонков, измерения, возраст); Коkubо а. Теzuka, Sci. Rept. Tohoku Imp. Univ., Ser. 4 (Biol.), XII, 4, 1938 : 639 (озерная сельдь оз. Обучи, сравнение с сельдью оз. Хинума; биология).
— Clupea harengus pallasi Берг, Рыбы пресных вод, I, 1948 : 155; Суворов, в: Промысловые рыбы СССР, 1949 : 127, табл. 31; Фролов, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 32, 1950 : 65, рис. 1—4 (западное побережье Сахалина, зал. Анива; измерения, просчеты).
— Clupea pallasi Пономарева, Учен. зап. Горьк. Гос. унив., XIX, 1951 : 179, рис. 2.
Распространение, биология. Steller, Beschr. Kamtschatka, 1774 : 167, 175 (Берингово м. от Авачинской губы до Олюторского зал., Вилючинское оз., устье р. Камчатки, о. Карагинский); Крашенинников, Описание земли Камчатки, 1755 : 326 («водятся в Восточном море, а в устьях рек, текущих, в Пенжинское море, бывают гостем», «В осень заходят они в великие озера, и там плодятся и зимуют, а весною выплывают в море»); Turner, Contrib. nat. hist. Alaska, II, 1886 : 111, pl. 14 (о. Михаила; нерест в середине июня); Nelson, Rept. nat. hist, collections Alaska, III, 1887 : 320 (о. Михаила; условия размножения); Гребницкий, Вестн. рыбопр., XII, 6—7, 1897 : 339 (Командорские о-ва); Heincke, Naturgesch. d. Herings, I, 1898 : LXVI (восточные берега Японии); Слюнин, Охотско-камчатск. край, 1900 : 335 (р. Пятибратная, в 1893 г. зашло руно, которое потом уснуло); Шмидт, Морск. пром. о-ва Сахалина, СПб., 1905 : 50—93 (биология); Evermann a. Goldsborough, Fish. Alaska, 1907 : 232 (Аляска, от Порт-Кларенс к югу); Сокольников, Тр. Общ. судох., Промысл. отд., VI, 1911 : 52 (Анадырский лиман); Jordan a. Metz, Mem. Carnegie Mus., VI, 1, (1913), 1914 : 6 (Пусан, Нампхо в северо-западной Корее); Павленко, Мат. позн. русск. рыбол., III, 10, 1914 : 1—144 (зал. Петра Вел. и Татарский прол.; биология, р. Суйфун в 15 в. от устья); Берг, Ежегодн. Зоолог. муз. Акад. Наук, XIX, 4, (1914), 1915 : 555 (р. Тумень-ула, в 3 в. выше устья); Fraser, Contrib. Canad. Biol., VI, (1921), 1922 : 105 (берега Канады; биология); Державин, Русск. гидробиолог. журн., II, 8—10, 923 : 193 (оз. Нерпичье в устье р. Камчатки; единичный заход); Jordan a. Hubbs, Mem. Carnegie Mus., X, 2, 1925 : 121 (Кусиро, Хакодате); Амброз, Произв. силы Дальн. Вост., IV, 1927 : 61 (зал. Петра Вел.; биология); Рабинерсон, Изв. Отд. прикл. ихт., VIII, 1928 : 47 (зал. Петра Вел.; возрастной состав, рост); Правдин, Изв. Тихоокеанск. иаучно-промысл. ст., I, 1928 : 192 (Авачинская губа, Усть-Большерецк); Тanaka, Fish. Japan, XLIII, Tokyo, 1928 : 844, f. 484 (у берегов Японии на юг в Японском м. до Тояма, в Тихом океане до Хинума, пров. Хитати); Линдберг и Дулькейт, Изв. Тихоокеанск. научно-промысл. ст., III, 1929 : 7 (Шантарские о-ва; единично); Курнаев, Рыбн. хоз. Дальн. Вост., 1930, 2 : 34 (западная Камчатка; химический состав); Амброз, там же, 1930, 2 : 30 (Амурский лиман, Охотск, Тауйск, Гижига, Ича на западном берегу Камчатки, Авачинская губа, заливы Карагинский, Олюторский, Корфа и Анадырский); там же, 1930, 7—8 : 38 (зал. Де-Кастри; возраст); там же, 1930, 9—10 : 55 (оз. Халыгер на Камчатке, Авачинская губа, Болынерецкий район); Соц. реконстр. рыбн. хоз. Дальн. Вост., 1931, 5—7 : 129 (заливы Ныйво и Чайво на Сахалине; возраст, размеры); там же, 1931, 8—10:3 (Приморье; возрастной состав, возраст, рост); Rounsefell, Bull. U. S. Bureau Fisher., XLVII, 2, 1931 : 15 (юго-восточная Аляска; колебание уловов); Rounsefell a. Dahlgren, ibid., XLVII, 9, 1932 : 263 (Аляска; размеры, возрастной состав, колебание уловов); Journ. Conseil, VIII, 3, 1933 : 371 (юго-восточная Аляска; мечение); Кагановский, Вестн. Дальневост. фил. АН СССР, 1—2—3, 1933 : 139 (Анадырский лиман); Mori a. Uсhida, Journ. Chosen Nat. Hist. Soc., 19, 1934 : 4 (Сейсин, Вонсан, Пусан); Hart a. Tester, Trans. Amer. Fish. Soc., LXIV, Wash., 1934 : 307 (берега Брит. Колумбии; количественный учет икры на нерестилищах, плодовитость); Rounsefell, Рас. Fisherm., 32, 3, Febr. 1934 : 31 (юго-восточная Аляска; расы, миграции); Wailеs, Journ. Biol. Board Canada, I, 6, 1935 : 477 (берега Брит. Колумбии; питание); Кuragami а. Кawana, Bull. Jap. Soc. Sci. Fish. Tokyo, V, 2, 1936 : 93 (зависимость интенсивности нереста и уловов от солнечных пятен); Dahlgren, Journ. Conseil, XI, 2, 193 : 229 (юго-восточная Аляска; мечение); Hart a. Tester, It opt. Prov. Fish. Dept., Brit. Colombia, (1936), 1937 : 55 (берега Брит. Колумбии; мочение); Tester, Journ. Biol. Board Canada, III, 2, 1937 : 145 (там же; возрастной состав, половое созревание); Aikawa, Bull. Japan. Soc. Sci. Fish. Tokyo, VI, 2, 1937 : 59 (о. Хоккайдо; возрастной состав уловов, запасы); Schaefer, Copeia, 1937, 1 : 57 (процесс нереста); Андрияшев, Исслед. морей СССР, 25, 1937 : 295 (Авачинский зал.); Tester, Journ. Conseil, XIII, 1, 1938 : 71 (зависимость количества позвонков от температуры воды в период ранних стадий развития личинок); Писарев, Рыбн. хоз., 1938, 8 : 25 (содержание жира); Кагановский, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 14, 1938 : 19 (Лионское м.; возрастной состав); Агапов, Тр. Научно-исслед. инст. полярн. землед., животн. и промысл. хоз., сер. промысл. хоз., 15, 1941 : 101 (Анадырский лим.; нерест, возрастной состав, измерения); Полутов, Костюченко и Панин, Рыбн. хоз., 1941, 3 : 3 (берега Камчатки; подходы, питание, уловы); Кизеветтер, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 21, 1942 : 58 (химический состав, содержание жира); МсНuрh, Journ. Fish. Res. Board Canada, V, 4, 1942 : 347 (ненормальности строения позвонков); Clemens a. Wilby, Bull. Fish. Res. Board. Canada, LXVIII, 1946 : 79 (берега Канады); Панин, Рыбн. хоз., 1946, 8 : 31 (северо-восточные берега Камчатки; условия подходов, состав уловов); Зоолог. журн., XXV, 5, 1946 : 459; Природа, 1947, 1 : 68 (там же; связь подходов с приливами); Варварин, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 22, 1946 : 3 (Татарский прол.; жирующая сельдь, биология); Пирожников, Рыбн. хоз., 1946, 10—11 : 59; Докл. АН СССР, н. с., LVI, 4, 1947 : 441 (м. Мастах у юго-восточной оконечности Быковского п-ова в бухте Тикси, 3, 21 и 26 VIII 1945, 3 экз.); Аюшин, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 25, 1947 : 3 (северо-западная часть Охотского м.; подходы, нерест, возраст); Пискунов, там же : 230 (зал. Де-Кастри; возраст, рост); Пирожников, Задачи научно-исслед. организаций в четв. пятилетке в обл. развит. рыбн. хоз. Сибири, изд. Главсибрыбпром, Новосибирск, 1948 : 9 (Оленёкский зал.; дважды в 1946 г.); Пискунов, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 29, 1949 : 182 (западное побережье южной части Сахалина; возрастной состав); Кун, там же : 164 (северная часть Татарского прол. у берегов Сахалина; питание); Городничий, Рыбн. хоз., 1949, 1 : 36 (колебание уловов в связи с потеплением); Голенченко, там же, 1949, 2 : 18, рис. 1 (западное побережье Сахалина; летнее и осеннее распределение); Пискунов, там же, 1949, 5 : 37 (западное побережье южной части Сахалина; ход на нерест, возрастной состав); там же, 1949, 7 : 27 (лов на свет); Веденский, там же : 36 (южные Курильские о-ва); Фридлянд, там же, 1949, 11 : 35 (западное побережье южной части Сахалина; половое созревание, плодовитость, развитие икры); Tester, Journ. Fish. Res. Board Canada, VII, 7, 1949 : 43 (западный берег о. Ванкувер; число позвонков из ряда пунктов за 1927—1938 гг.); Фридлянд, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 31, 1949 : 193 (западное побережье южной части Сахалина у Антоново; личинки с начала мая по половину июня); Кун, там же : 198 (Амурский лиман; зимнее питание молоди); Панин, там же, 32, 1950 : 3 (северо-восточное побережье Камчатки; биология, промысел); Кагановский и Полутов, там же : 37 (Пенжинский зал.; биология); Веденский, там же : 55 (восточное побережье Сахалина; биология); Богаевский, там же : 155 (о. Монерон; промысел); Фридлянд, там же : 157 (западное побережье южной части Сахалина; личинки); Пробатов, Рыбн. хоз., 1950, 2 : 24 (западное побережье южной части Сахалина; возрастной состав); Леванидов, там же : 37 (химический состав); Богаевский, там же, 1950, 3 : 43 (о. Монерон; преднерестовые и нагульные подходы, нерест вокруг почти всего острова до 1926 г.); Козлов, Фролов и Шнель, там же, 1950, 6 : 45 (восточное побережье северной части Сахалина; возможность увеличения уловов); Богаевский, там же, 1950, 9 : 14 (район о. Монерон; лов нагульной сельди); там же, 1951, 3 : 25, рис. (западное побережье южной части Сахалина; летнее распределение); Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 34, 1951 : 5, рис. 1—6 (юго-западные берега Сахалина; летнее и осеннее распределение); Пробатов и Варварин, там же : 25, рис. 1—5 (зал. Анива; подходы молоди); Грищенко, там же : 41 (влияние электрического света); Панин, там же : 257, рис. 1 (восточное побережье Камчатки; распределение нагульной сельди); Пробатов и Шелегова, Докл. АН СССР, LXXVIII, 3, 1951 : 601, рис. 1 (зал. Анива; причины осенних подходов к берегам); Пробатов, там же, LXXX, 5, 1951 : 825, рис. 1 (южная часть Сахалина; причины изменения уловов); Аюшин, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 35, 1951 : 81 (северная часть Охотского м.; нагульная сельдь); Кун, там же: 87, рис. 1—4 (северная часть Охотского м.; питание); Пробатов и Фролов, там же: 97, рис. 1 (оз. Тоннай; отличие от морской сельди, нерест, плодовитость, размеры, возраст, рост); Фридлянд, там же: 105, рис. 1—11 (юго-западное побережье Сахалина; нерест, плодовитость, развитие икры, личинки и мальки); Пискунов, Зоолог, журн., XXXI, 1, 1925 : 115 (западный берег Сахалина; плодовитость).

D (III) IV (V) 13—15 (14.6), общим числом 17—20 (18.7), A III (IV) 12—16 (14.1), общим числом 15—19 (17.1), sp. br. 63—73 (67.0), килевых чешуй позади основания брюшных плавников 11—13 (11.8), vert. S. 53—55 (54.1), caud. vert. 27—32 (29.7). Брюшные килевые чешуи более или менее развиты лишь позади основания брюшных плавников, впереди от них развиты слабо или отсутствуют. Зубы на сошнике менее развитые, чем у Clupea harengus harengus L.. Голова небольшая и широкая, длина ее 19.0— 20.8 (19.8)% длины тела; межглазничный промежуток 15.7—18.6 (17.2)% длины головы. Глаза небольшие, 19.7—22.6 (21.2)% длины головы. Основание анального плавника сравнительно короткое, 9.2—11.2 (10.2)% длины тела. Размеры до 50 см, у берегов Камчатки в уловах от 24 до 38 см, средние размеры нерестовой сельди в ряде промысловых районов Камчатки от 29.1 до 34.1 см, в Пенжинском зал. — 24 см, в зал. Петра Великого — 24 см. Описание дано по 50 экз. длиной от 31.8 до 36.1, в среднем 34.0 см, собранным 12 III 1949 у западных берегов южной части Сахалина в районе Антоново. У экземпляров от азиатских берегов позвонков 49—57, средние колеблются от 52.78 до 54.67; у экземпляров от американских берегов позвонков (45) 47—57, средние колеблются от 51.68 до 54.79.
Морская форма, живущая в воде с океанической соленостью, но размножающаяся как в морской, так и в солоноватой воде.
Распадается на ряд еще слабо изученных форм. Наиболее существенно делится на две группы — морских или береговых сельдей, размножающихся в море у берегов, и озерных, живущих в море, но для икрометания заходящих в осолоненные озера и в заливы и бухты с пониженной соленостью. Морские сельди у берегов Камчатки, по исследованиям К. И. Панина, характеризуются низким, прогонистым и округлым телом, спинным и отчасти анальным плавниками, отнесенными далее назад, быстрым темпом роста. У озерных сельдей тело более высокое и плоское, спинной и отчасти анальный плавники расположены ближе к концу рыла, темп роста замедленный. Сходным образом от морских сельдей отличается сельдь и оз. Тоннай (Пробатов и Фролов, 1951). В пределах каждой из этих групп имеется ряд локальных форм или стад, характеризующихся обособленностью нерестилищ, некоторыми морфологическими особенностями, различным возрастным составом в уловах и темпом роста. В восточной части Тихого океана по направлению с юга на север у западных берегов Японского моря, по исследованиям Амброза (1931), Фуджита и Кокубо (Fujita a. Kokubo, 1927), среди морских сельдей в настоящее время различают следующие локальные формы, или расы, с обособленными нерестилищами, отличающиеся главным образом особенностями роста: 1) корейская сельдь, нерестящаяся у восточных берегов Кореи (Хоко, Пусан); 2) сельдь зал. Петра Великого, с наиболее быстрым ростом из всех форм, встречающихся в наших водах; 3) пластунско-нельминская сельдь, с более медленным ростом, размножающаяся в зал. Пластун — б. Нельма; 4) декастринская сельдь, с самым медленным ростом из всех япономорских сельдей (см. ниже), основные нерестилища которой по материковому берегу Татарского прол. расположены в зал. Де-Кастри, бухтах Сущева и Чихачева и далее на север до м. Лазарева; по сахалинскому побережью эта сельдь размножается у открытого берега в районе Черной речки, в полузащищенных бухтах в районе Александровска — Половинки, м. Танги — м. Хоэ и далее до м. Погиби. Эта сельдь, по некоторым указаниям (Фролов, 1949), в нерестовый период заходит и южнее Черной речки — до Широкой Пади и Углегорска. Декастринская сельдь близка к сельди Охотского моря. Остается неясным вопрос, является ли самостоятельной формой сельдь от западных берегов Хондо. У берегов Хоккайдо и южной части Сахалина установлена 5) южносахалинско-хоккайдская сельдь (называемая также у нас западносахалинской, у японцев — хоккайдской), куда относятся сельди, размножающиеся у западных и отчасти восточных берегов Хоккайдо и у западных и отчасти южных (западное побережье зал. Анива) Сахалина. Ареал этой, наиболее многочисленной из всех форм тихоокеанской сельди у западного побережья Сахалина уточнен исследованиями Тихоокеанского научно-исследовательского института морского рыбного хозяйства и океанографии (Пискунов, Фролов и др.). Основные места размножения здесь расположены от м. Кузнецова до м. Усу и в западной части зал. Анива (в апреле); в годы больших подходов к южной части западного побережья Сахалина она распространяется и далее к северу — до Углегорска и Широкой Пади. У восточных берегов Сахалина и северных Хоккайдо, по Фуджита и Кокубо, встречается смешанная группа этой сельди и охотской, так называемая охотско-хоккайдская сельдь (см. ниже). У тихоокеанских берегов Японии локализована: 6) тохокуккайская сельдь, размножающаяся у восточного побережья Хондо (Тасиродзима, пров. Мияги). Наименее изучены сельди Охотского моря. У восточных берегов Сахалина указывается: 7) восточносахалинская сельдь, свойственная зал. Терпения и северной части зал. Анива, но изученная на материале лишь из последнего (Фролов, 1949). В зал. Анива эта сельдь нерестует в мае, после нереста южносахалинско-хоккайдской сельди. Южносахалинско-хоккайдская сельдь, судя по ряду указаний, проникает и к восточному берегу южной части Сахалина. Сельди от Шантарских о-вов и охотского побережья далее к северу изучены мало. По размерам, возрастному составу и темпу роста к ним близка декастринская сельдь. В северной части Охотского моря указывается: 8) пенжинская сельдь, с более медленным ростом и менее крупными размерами, чем южносахалинско-хоккайдская сельдь; эта сельдь встречается и размножается в Пенжинском зал. По исследованиям Кагановского и Полутова (1949), сельдь Пенжинского зал. разделяется на два стада — пенжинское и гижигинское. Гижигинская сельдь нерестует раньше (со второй половины мая до второй половины июня) и при более высокой температуре воды (3—10°, массовый ход при 4—7°), чем пенжинская (с середины июня, при температуре от 7—8°, массовый ход после повышения температуры воды до 10—12°). Нерестовая пенжинская сельдь мельче (в среднем 23.8 см) и более молодого состава, чем гижигинская сельдь (в среднем 26.2 см). В Беринговом море у восточных берегов Камчатки, по исследованиям К. И. Панина, изучавшего меристические признаки и биологические особенности, и Р. А. Костюченко, исследовавшего рост, различаются две локальные формы или стада: 9) южнокамчатская сельдь, с медленным ростом, размножающаяся в Авачинской губе, в бухтах юго-восточного побережья до м. Лопатка, а также, возможно, у юго-западного побережья в районе зал. Камбального на север до устья р. Озерной; 10) северокамчатская сельдь, с более быстрым ростом, чем предыдущая, нерестящаяся в зал. Олюторском, Корфа и в бухтах северной части прол. Литке (Анапка, Уала). Сельди более северных районов Берингова моря не изучены. У берегов Сев. Америки констатированы отдельные формы сельди у Калифорнии, южной части Брит. Колумбии, ряд форм у юго-восточной Аляски, в зал. Вильяма, в зал. Качмак — прол. Чуяк, в бухте Чигник, у Шумагинских о-вов, Уналашки и в зал. Головина. В западной половине Тихого океана сельди южных частей ареала имеют меньшее число позвонков, чем из северных. Так, например, в Японском море корейская сельдь имеет в среднем 53.80 позвонков, сельдь зал. Петра Великого 54.21—54.67. У южносахалинско-хоккайдской сельди позвонков в среднем 54.20—54.62 (Хоккайдо; по Фуджито и Кокубо); у экземпляров от западных берегов южной части Сахалина, по Фролову, 54.16. У сельди Охотского моря в южной части ареала позвонков также меньше: у восточносахалинской (зал. Анива) позвонков в среднем 53.85, в северной части моря у пенжинской сельди 54.11 (в Кичигинском зал. 54.82). В Беринговом море, наоборот, количество позвонков больше у сельдей из южных частей его: у южнокамчатской сельди позвонков в среднем 54.18—54.35, у северокамчатской 53.28—53.98. У берегов Сев. Америки среднее число позвонков увеличивается с юга на север: от 50.68 (Сан-Диего) до 53.22 (Уналашка). В аналогичной зависимости находится и рост, более быстрый у сельдей из южных частей ареала, чем из северных (см. ниже).
Среди озерных сельдей различаются (Fujita a. Kokubo, 1927): 1) хинумская сельдь, на восточном побережье Хондо (нерест в оз. Хинума); с этой сельдью по всем признакам идентична сельдь, заходящая для нереста в оз. Обучи (Obuchi, пров. Аомори) на о. Хондо (Kokubo а. Tezuka, 1938); 2) сельдь лимана Аккеси, на восточном побережье Хоккайдо (нерест в лимане Аккеси, находящемся в глубине залива того же названия); 3) тоннайская сельдь, на восточном побережье южной части Сахалина (нерест в оз. Тоннай, относительно этой сельди см. Пробатов и Фролов, 1951). По восточному побережью Сахалина озерного типа сельди есть и в северной его части, в зал. Ныйво и Чайво. У восточных берегов Камчатки озерные сельди, заходящие с осени в осолоненные й связанные с морем протоками прибрежные озера, известны из оз. Халыгер и Нерпичье.
Некоторые из перечисленных форм, изученных, как указывалось, еще недостаточно, в дальнейшем, подобно атлантическим, несомненно будут объединены. Так, судя по всему, следует рассматривать южно- и северокамчатскую сельдь, подобно тому как рассматривают норвежскую и мурманскую сельдей атлантическо-скандинавской сельди.
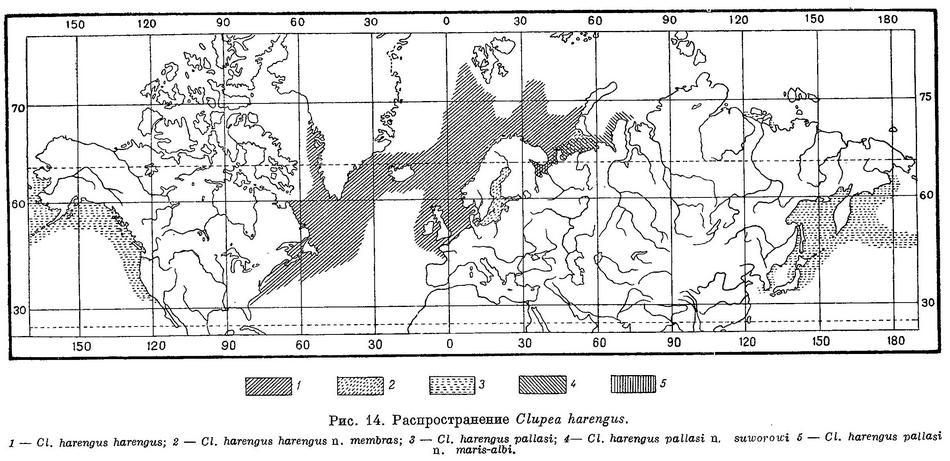
Распространение. Северная часть Тихого океана и прилегающие моря. У берегов Сев. Америки на юг до Калифорнии (Сан-Диего), на север до Порт-Кларенс на Аляске. В более ранних работах сельдь у берегов Сев. Америки указывалась и далее к северу: в 80 милях к западу от устья р. Мекензи (Scofield. List fishes Arctic Alaska, 1899 : 494, со слов местных жителей) и еще далее — из зал. Батурст (Bathurst inlet, 67° с. ш., 109° в. д.; Richardson. Fauna Boreali-Americana, 1836 : 230). У азиатских берегов на юг до западного побережья Кореи в Желтом море. Согласно Базилевскому, есть и далее, в Чжилийском зал. В Японском море вдоль всего западного побережья, вдоль восточного побережья на юг до зал. Тояма на о. Хондо. У тихоокеанских берегов Японии на юг до Хинумы (пров. Хитати) на о. Хондо, далее к северу вдоль Курильских и возможно Алеутских о-вов. По всему побережью Охотского моря, в Беринговом море вдоль восточных берегов Камчатки и далее к северу до Анадырского зал. В последние годы отмечена у северных берегов Сибири в бухте Тикси близ дельты Лены и в Оленёкском зал. В бухте Тикси сельдь была обнаружена в 1940 (Берг, 1948 : 154) и в 1945 гг. (Пирожников, 1947). Один из 3 экз. Пирожникова хранится в коллекциях Зоологического института АН СССР (№ 31143). В 1946 г. сельдь дважды была поймана в южной части Оленёкского зал. (Пирожников, 1948). Один из этих экземпляров также хранится в коллекциях Зоологического института СССР (№ 31154, п. Станнах-Хочо, 27 VII 1946). Указание наше (1944) на нахождение сельди в Чаунской губе и у о. Ляховского недостоверно и основано на недоразумении. По сообщению Ф. Н. Кириллова, она есть также, повидимому, близ дельты Индигирки. Нерест происходит в ряде мест почти по всему ареалу. У северных берегов Сибири нерестует, повидимому, в бухте Тикси (Пирожников, 1946). У берегов восточной Азии нерест отмечен в Анадырском лимане (Агапов, 1941), в б. Уала, Анапка, зал. Корфа, южнее — в районе Авачинской губы и в бухтах к югу от нее (Жировая, Ахомтэн) у восточных берегов Камчатки, в районе р. Озерной по западному берегу (по И. А. Полутову и Р. А. Костюченко), вдоль берегов Гижигинской и Пенжинской губ (Кагановский и Полутов, 1950), в заливах Ныйский, Даги, Чайво, Пильтун по восточному берегу северной части Сахалина, в заливах Терпения и Анива и по западному побережью Сахалина, в ряде мест (зал. Петра Великого и др.) у берегов Приморья (Амброз, 1931), у западных и отчасти восточных берегов Хоккайдо. У берегов Сев. Америки нерест происходит в зал. Сан-Диего, Сан-Франциско, а также в ряде мелких заливов севернее — у берегов США, Брит. Колумбии и юго-восточной Аляски. Среди последних наибольшее нерестилище расположено в зал. Ситка с наружной стороны о. Баранова, меньшее — в зал. Сан-Альберта на о. Принца Уэльского (у входа в проливчик Клауок), небольшое — в прол. Стефенса (у Juneau), непостоянное и уменьшающееся — в проливчике Kootznahoo. Севернее, у берегов Аляски, места нереста отмечены в зал. Принца Вильяма, Кука, у о. Кодьяка и в Беринговом море у Уналашки. Для нереста заходит также в солоноватые озера, стоящие в связи с морем, или в изолированные части заливов, бухт и пр. Камчатки, Сахалина и японских островов. Для берегов Сев. Америки нереста в озерах не указано.
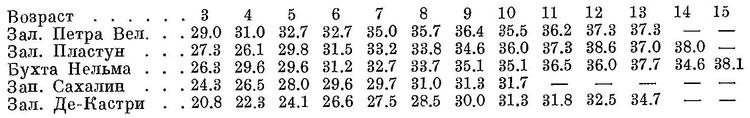
Биология. Время нереста находится в зависимости от географического положения нерестилищ и длится в общем с декабря по июль включительно. В Анадырском лимане нерест, судя по нахождению сельди с текучими половыми продуктами в конце июля, происходит в июле. Южнее, в бухтах Ананка и Уала у восточных берегов Камчатки, нерест начинается с середины мая и заканчивается в конце его (Панин, 1950). У северо-западных берегов Охотского моря нерест начинается с середины мая — начала июня и заканчивается в начале июля (Аюшин, 1947). У берегов Приморья нерест происходит раньше — в зал. Де-Кастри начинается со второй половины мая — начала июня (Ахошин, 1947) и заканчивается в конце июня; южнее, в зал. Посьета, — с первой половины марта по конец апреля — начало мая. У восточных берегов Сахалина икрометание начинается с конца мая — начала июня, местами вследствие плавающих льдов — позднее — в конце июня и начале июля (Амброз, 1931), у западных берегов южной части Сахалина — с начала — середины апреля по середину мая. У берегов Сев. Америки икрометание происходит значительно раньше, чем в соответствующих широтах у берегов Азии. В южных частях ареала, у Сан-Диего, нерест отмечен в середине декабря, в зал. Сан-Франциско — со второй половины декабря по середину апреля. В северных частях ареала нерест происходит позднее — у берегов юго-восточной Аляски с середины по конец марта, в зал. Кука в середине мая, у о. Кодьяка с конца мая по начало июня (Rounsefell, 1930). Общая продолжительность нереста составляет около 12—15 суток (Панин, 1950). У берегов Брит. Колумбии продолжительность нереста меньше — на каждом нерестилище всего лишь несколько (2—3) суток (Hart a. Teater, 1934). Икра выметывается на мелководьях, на глубине 3—4 м, максимально, повидимому, до 10—15 м, у западных берегов южной части Сахалина не дальше глубин 6—9 м, иногда почти у самого уреза воды на глубине 0.5—1.5 м, обычно в укрытых от действия ветра местах: в заливах, бухтах, часто в предустьевых пространствах рек или у открытого берега, на подводную растительность — зостеру, фукус и др., с каменистым или илисто-песчаным дном, покрытым мелкими камнями, галькой и пр. (Амброз, 1931), в исключительных случаях при отсутствии подводной растительности икра в небольшом количестве откладывается на камень и гальку (Аюшин, 1947; Фридлянд, 1951). Икра, как и у других форм вида, клейкая. Температурные условия нереста, как и у других малопозвонковых подвидов океанической сельди, довольно низкие, иногда нерест начинается еще подо льдом. У восточных берегов Камчатки (б. Уала) нерестовые подходы начинаются при температуре воды от 2.2 до 7° и заканчивается при повышении среднесуточной температуры до 8°, основные подходы и нерест происходят при температуре от 3 до 6° (Панин, 1950). В зал. Петра Великого нерестовые подходы начинаются даже при отрицательной температуре, но основной ход происходит при 4—9° (Амброз, 1931). У западных берегов южной части Сахалина подходы к берегам для нереста начинаются при 2° (иногда ниже, даже при отрицательных температурах, или выше, до 4.9°), заканчиваются при 10°, основная масса подходит обычно при 3—6° (Пискунов, 1949; Фридлянд, 1951). У северо-западных берегов Охотского моря нерестовая сельдь подходит при температуре воды от 0.5 до 10.7° (Аюшин, 1947). Подобно другим малопозвонковым подвидам, выносит значительные опреснения, заходя в устья рек и с приливом подымаясь высоко вверх по течению (в р. Тумень-ула в низовья, в р. Суйфун на 15 км от устья; Павленко, 1914; Берг, 1915), но полного опреснения не переносит и, попав в пресную воду, погибает, как это отмечено в р. Пятибратной (Слюнин, 1900). Еще большую склонность к опреснениям обнаруживают озерные сельди, размножающиеся в осолоненных озерах или частях заливов и бухт с пониженной соленостью. В озера Камчатки эти сельди заходят с осени и, перезимовав, весной после нереста уходят в море. Плодовитость у сельди из зал. Петра Великого колеблется (длина рыб 21—37 см) от 10.8 до 134.1, в среднем 72.2 тыс. икринок (Амброз, 1931), от западных берегов южной части Сахалина — от 25.1 до 85.2 тыс. икринок (Фридлянд, 1949), по другим указаниям (Пискунов, 1952; длина рыб 23—37 см) — от 17.6 до 106.4, в среднем 50.6 тыс. икринок, от западных берегов северной части Сахалина у Широкой Пади (1940 г., длина рыб 23—34 см) — от 20.8 до 95.5, в среднем 44.9 тыс. икринок, там же (1941 г., длина рыб 20—32 см) — от 13.9 до 77.8, в среднем 38.4 тыс. икринок, там же (1946 г., длина рыб 21—32 см) — от 13.8 до 117.8, в среднем 49.8 тыс. икринок, у Александрова (1940 г., длина сельдей 18—30 см) — от 8.7 до 46.5, в среднем 27.3 тыс. икринок (Пискунов, 1952), от берегов Камчатки (северная часть прол. Литке, длина рыб 27—38 см) — от 12.6 до 122.8, в среднем 65.9 тыс. икринок (Панин, 1950), от берегов Брит. Колумбии — от 18.2 до 29.5, в среднем около 20 тыс. икринок (Hart a. Tester, 1934), по другим указаниям (Fraser, 1922) — от 12 до 35 тыс. икринок, из оз. Тоннай — от 7.5 до 29.0, в среднем 17.0 тыс. икринок (Пробатов и Фролов, 1951). Диаметр оплодотворенных икринок: 1.3—1.6 мм (Амброз, 1931). Икра выметывается, судя по указаниям ряда авторов, одновременно. Продолжительность развития икры при средних температурах воды около 9.2° равна 15 суткам, при 8.1° — 24 суткам, при 5.8° — 23 суткам, при 3.7° — 38 суткам, при 2.7° — 50 суткам (Амброз, 1931; Панин, 1950). В естественных условиях при средней температуре воды 6° развитие икры длится около 26 суток, при 2.9° — 30 суток. Температура 17—18° и выше для развивающейся икры гибельна. Развитие икры может происходить при довольно широких пределах колебания солености — от 10.66 до 33.33‰. Плотность засева икрой субстрата крайне густа, у западных берегов южной части Сахалина обычно составляет 2—6 млн. икринок на 1 м2, максимально достигает до 17.5 млн. икринок, у северной части в 2—3 раза меньше. Среди развивающихся икринок у западных берегов южной части Сахалина мертвых обнаружено в зависимости от субстрата от 2 до 48% (Фридлянд, 1949, 1951), у берегов Брит. Колумбии от 1.4 до 10.5% (Hart a. Tester, 1934). Личинки по выходе из икринок имеют в длину около 6.0—6.2 мм. Подросшие мальки озерных сельдей выходят из озер в море, не отходя далеко от этих мест. Личинки морских сельдей первоначально держатся на местах нереста, постепенно отходя от берегов и из заливов и бухт (Амброз, 1931). У западных берегов южной части Сахалина личинки ко времени рассасывания желточного мешка отходят от берегов до зоны глубин 28—30 м; отход личинок от берегов заканчивается к концу мая (1949 г.) или середине июня (1948 г.). Позднее, до конца ноября сформировавшиеся мальки держатся повсюду вдоль западного побережья южной части Сахалина к северу от м. Лопатина (Фридлянд, 1949). К северо-восточному побережью Хоккайдо (пров. Китами) и в зал. Анива молодь, выведшаяся, судя по всему, у западных берегов Хоккайдо, подходит с середины июня. Взрослые больших миграций, меридиональных и вдоль берегов, такой протяженности, как у атлантической сельди, не совершают; все передвижения имеют местное значение в пределах ограниченного пространства, преимущественно к берегам из открытого моря и от берегов. Лишь местами, например, у западных берегов южной части Сахалина, какая-то часть половозрелой сельди отходит вдоль берегов южнее. Зимний период жизни не изучен. Судя по имеющимся наблюдениям и опытам мечения у берегов Сев. Америки (Dahlgren, 1936), держится вблизи мест икрометания, иногда даже в тех же заливах, где размножается, например в зал. Петра Великого. По наблюдениям Н. И. Куличенко, у западных берегов южной части Сахалина, судя по нахождениям в желудках трески, пойманной в октябре, ноябре, феврале и марте на глубине 100—150 м, какая то часть сельди держится в открытом море против мест икрометания на глубинах. Во второй половине марта сельдь появляется в районе о. Монерон и затем образует преднерестовые скопления в 10—12 милях от Сахалина. Преднерестовая сельдь во второй половине марта держится на глубине 40—50 м, затем по мере прогревания воды поднимается в поверхностные горизонты и подходит к берегам (Пискунов, 1949). Подходы имеют обычно фронтальный характер — из открытых частей моря к берегам и лишь в узко прибрежной области вдоль берегов. Весенние подходы сельди к берегам начинаются в южных районах, раньше чем в северных. В разных пунктах Приморья по направлению с севера на юг начало весенних подходов приходится на следующие сроки: зал. Петра Великого — 17—20 марта, зал. Пластун — 20—22 апреля, бухта Нельма — 1—10 мая, зал. Де-Кастри — 17—24 мая (Амброз, 1931). У западных берегов южной части Сахалина начало нерестовых подходов наступает с 5 по 22 апреля (Богаевский, 1951). У восточных берегов Камчатки в Авачинской губе весенние подходы сельди начинаются с середины апреля; севернее, в бухтах Уала, Анапка и зал. Корфа в середине или во второй половине мая. Продолжительность весенних подходов обычно не превышает 20 дней, в отдельных случаях (бухты Уала, Анапка) они заканчиваются в 5—6 дней (Панин, 1950). На подходы сельди к берегам для размножения существенное влияние имеют приливо-отливные течения, особенно в сизигийные фазы — ко времени полнолуния или новолуния. В частности, в бухтах Анапка и Уала у восточных берегов Камчатки подходы сельди происходят в сизигийные фазы больших колебаний уровней в начале или в середине мая от первой четверти луны через полнолуние до последней четверти (Панин, 1946). Подходам сельди к берегам благоприятствуют береговые ветра, выносящие прогретые прибрежные воды в открытое море и тем способствующие подходам. Сильные и длительные береговые ветра, вызывающие выход на поверхность холодных вод нижележащих горизонтов, по наблюдениям К. И. Панина, на подходы действуют отрицательно. Весенние подходы к берегам происходят в несколько (2—3—4) последовательных волн: в первых подходах преобладают крупные сельди старших возрастов, в конце — более мелкие младших возрастов (Амброз, 1931; Панин, 1950 и др.). Крупные сельди начала весенних подходов идут на нерест, мелкие и более жирные сельди конца подходов для питания. Сельди более старших возрастов начинают подходить даже при отрицательных температурах воды, трехгодовалые — при 3—4°, двухгодовалые — в массовом количестве при 5—9°. По окончании нереста сельдь из прибрежных областей отходит и начинает усиленно откармливаться. После весенних нерестовых подходов летом подходит к берегам для питания нагульная или жирующая сельдь. К берегам Приморья жирующая сельдь подходит в первой половине июня (Амброз, 1931). Против западных берегов южной части Сахалина первые косяки отнерестовавшей сельди появляются во второй половине апреля у о. Монерон, сначала к северо-востоку и в некотором отдалении (3—10 миль) от него, в конце апреля и начале мая приближаются (до 0.5 мили) и держатся здесь до конца июня. Вначале сельдь здесь в дневное время в поверхностных слоях воды держится сравнительно редко, чаще всплывая к поверхности после заката солнца и ночью; в июне всплывание в поверхностные слой воды происходит реже, в начале июля уловы прекращаются. В течение всего этого периода здесь происходит усиленное питание сельди, желудки ее у преобладающего большинства до предела наполнены пищей; с пустыми желудками встречается сравнительно небольшое количество (18% в 1948 г. и 28% в 1949 г.). Позднее, в июне в зависимости от температурных условий в начале или конце этого месяца, жирующая сольдь появляется у западных берегов южной части Сахалина в районе м. Слепиковского. В 1946 г. сельдь появилась здесь 28 июня, в 1947 г. — 10 июня, в наиболее теплом 1948 г. — 4 июня. Большая часть сельди и начале держится преимущественно в верхних слоях на глубине 3—4 м от поверхности и подходит близко к берегам в зону ставных неводов. И этот период сравнительно хорошо упитанная сельдь питается менее интенсивно; с пустыми желудками встречаются большее количество ее (54% в 1948 г. и 52% в 1949 г.), чем у о. Монерон. Исчезновение сельди у о. Монерон по времени совпадает с появлением ее у м. Слепиковского и, судя также по размерам, возрастному составу, упитанности и жирности, это одна и та же сельдь, переместившаяся из одного района в другой. В зависимости от температуры воды, в середине или конце июля, с образованием сильно прогретого поверхностного слоя воды и избегая температуры воды выше 10°, сельдь от берегов отходит. Осенью, со второй половины сентября до начала октября, с похолоданием сельдь вновь держится в этом районе. В период от июля до половины сентября местопребывание сельди остается невыясненным (Богаевский, 1951). У западных берегов Камчатки нагульная сельдь подходит с июня по сентябрь, у восточных — в течение всего теплого сезона. Подходы отмечены во всех заливах — от Авачинского до Олюторского, вдоль побережья от Олюторки до Анадыря, у юго-восточных берегов от м. Лопатки до м. Поворотного. В нагульный период сельдь держится здесь в относительна прогретых водах с колебаниями температуры от 6 до 14°, наичаще в верхних слоях воды (до 20 м) при температуре от 4.5 до 7.5° (Панин, 1951). Озерные сельди, по наблюдениям К. И. Панина, в озера заходят с осени, держатся здесь всю зиму и после нереста выходят в море (см. также: Крашенинников, 1755, и Пробатов и Фролов, 1951). В промысловых уловах встречаются экземпляры от двух- до тринадцати- и пятнадцатигодовалых. Возрастной состав уловов и размеры сельди вследствие обособленности стад значительно различаются даже и в близлежащих районах. В северных районах восточного побережья Камчатки в весенних подходах с середины мая в 1938—1943 гг. преобладали семи-, восьми- и девятигодовалые; во втором подходе (конец мая — начало июня) преобладали младшие возрастные группы, в основном четырех- и пятигодовалые. Урожайными поколениями в уловах этих лет было поколение 1935 г., появившееся впервые в промысле в 1939 г. и составлявшее основную массу уловов до 1943 г. У западных берегов Камчатки (Кихчик—Кольский район) в 1934 г. основную массу уловов крупных сельдей составляли восьми- и девятигодовалые, среди мелких сельдей — четырех- и пятигодовалые (Панин, 1946). В зал. Петра Великого в 1926 г. в уловах преобладали трех- и четырехгодовалые сельди, в бухте Нельма — семи- и восьмигодовалые, в зал. Де-Кастри в 1929 г. — восьми- и девятигодовалые (Амброз, 1930), в 1938—1946 гг. — более младшие возрастные группы (Пискунов, 1947). У западных берегов южной части Сахалина и у Хоккайдо преобладающими в уловах в последние, годы были урожайные поколения 1939 и 1940 гг., которые составляли основную или значительную массу уловов, начиная с 1942—1943 гг. до 1951 г. включительно в возрасте от трех-четырех- до одиннадцати- двенадцатигодовалых. Половой зрелости у северных берегов Приморья достигает в трех-, отчасти в двухгодовалом возрасте и мечет икру, повидимому, до предельного возраста, судя по имеющимся данным, ежегодно {Амброз, 1931). У северо-восточных берегов Камчатки впервые начинает метать икру в трехгодовалом, как правило в четырехгодовалом возрасте.
В Авачинской губе встречаются половозрелые в двухгодовалом возрасте; возможно, что это озерные сельди, зимующие в оз. Вилюй. В Пенжинском зал., по наблюдениям К. И. Панина, среди мелкой нерестовой сельди встречаются половозрелые и в двухгодовалом возрасте. У западных берегов южной части Сахалина впервые начинает метать икру в трехгодовалом, в основном в четырехгодовалом возрасте; судя по нарастающему количеству поколения одного и того же года в более старших возрастных группах, часть сельди достигает половой зрелости и в более старшем возрасте — в пяти-, шести- и, возможно, как у мурманской сельди, в семигодовалом. У берегов Брит. Колумбии впервые начинает метать икру также в трех- и отчасти двухгодовалом, некоторые в четырехгодовалом возрасте (Tester, 1937). У берегов юго-восточной Аляски (прол. Стефенса) зрелые среди трехгодовалых составляют 84%, четырех- и пятигодовалые зрелые все; севернее, у берегов центральной Аляски (зал. Качмак), среди трехгодовалых зрелых 52%, среди четырехгодовалых — 83%, пятигодовалые зрелые все (Rounsefell, 1931). Сельдь, нерестующая в оз. Обучи (о. Хондо), в 1936 г. состояла из трех- и четырехгодовалых (Kokubo a. Tezuka, 1938), в оз. Тоннай — от двух- до десятигодовалых (Пробатов и Фролов, 1951). По темпу роста сельди разных локальных форм, как указывалось выше, существенно различаются, причем в восточной половине Тихого океана в северных частях ареала рост более замедленный, чем в южных. В Японском море в наших водах наилучший рост имеет сельдь зал. Петра великого, более замедленный — пластунско-нельминская сельдь, затем сельдь от западного берега южной части Сахалина и самым медленным — декастринская сельдь [поАмброзу (1930, 1931) и Фролову; см]:
У берегов Камчатки рост сельди еще более замедлен, причем здесь, наоборот, сельдь северной части прол. Литке (северокамчатская) имеет более быстрый рост, чем сельдь из Авачинской губы (южнокамчатская); сельдь Пенжинского зал. имеет еще более медленный рост (1940 г., по Р. А. Костюченко; см):
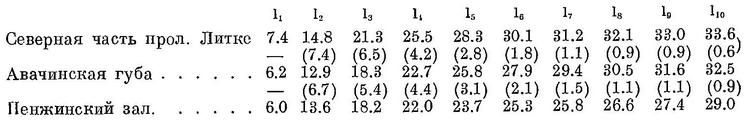
Годичные приросты у северокамчатской сельди (цифры в скобках) в первые годы выше, чем у южнокамчатской; с четвертого года жизни, наоборот — южнокамчатские имеют больший прирост. Более быстрым; ростом, как и у берегов Камчатки, обладают сельди из северных частей ареала, чем из южных, а также от берегов Сев. Америки. Так, например, шестигодовалые сельди от Уналашки на 6.5 см больше сельдей того же возраста от юго-восточной Аляски (прол. Стефенса). Озерные сельди при одном и том же возрасте имеют значительно меньшие размеры, чем морские из близких мест, как это видно из нижеследующих данных (по К. И. Панину; см):

Питание тихоокеанской сельди в северной части Татарского прол. у западного побережья Сахалина на протяжении года по интенсивности распадается на следующие периоды: 1) весеннего или преднерестового откорма (апрель—май), 2) нерестового голодания (май—июнь), 3) интенсивного посленерестового летнего откорма (с конца июня по август включительно), 4) осеннего ослабления питания и 5) мало изученный зимний период, когда сельдь или не питается совершенно, или питается слабо. Основу питания здесь составляют Euphausiidae Thysanoessa inermis, Thysanoessa raschii (82.1% по весу), Calanus finmarchicus (5.12%) и Sagitta elegans (9.04%). Весной, частично осенью и иногда летом в пище обычно преобладают Euphausiidae; Calanus finmarchicus наичаще встречается в желудках в июле, августе, иногда в сентябре (Кун, 1949). У западных берегов южной части Сахалина в период нагула сельди различают периоды: 1) усиленного питания после нереста, накопления жировых отложений и восстановления гонад после нереста (апрель—июнь); 2) уменьшения интенсивности питания, продолжения накопления жировых отложений и начала роста тела и развития гонад (июнь—август); 3) повышения интенсивности питания, замедления и окончания роста тела, продолжения развития гонад (август—октябрь) (Богаевский, 1951). В зал. Петра Великого взрослые поедают Copepoda, Mysidae, Amphipoda, реже икру рыб. Сеголетки и годовалые питаются также ракообразными, но более мелкими — мелкие формы Copepoda, zoea крабов, 2 раза в желудках были находимы личинки сельди (Амброз, 1931). В зимнее время здесь же (Амурский зал.) молодь в основном питается ракообразными и пр. — Calanus cristatus (47.9%), Stylomysis sp. (32.4%), Sagitta sp. (14.9%). У восточных берегов Камчатки, по исследованиям К. И. Панина, основной пищей служат веслоногие ракообразные; в Авачинском зал. основу питания составляют Eucalanus elongatus (61.2%), несколько меньшее Calanus cristatus (31.6%) и ничтожное Calanus tonsus (1.8%) и Thysanoessa sp. (4.3%); в Кроноцком зал. большое значение имеет Calanus cristatus (64.6%), значительно меньшее Eucalanus elongatus (22.9%) и незначительное Calanus tonsus (1.3%) и Thysanoessa sp. (7.5%); в Петропавловском ковше сельдь питается, помимо Copepoda, донными организмами (Polychaeta, Amphipoda, случайно моллюск Tellina calcarea, молодь рыб). В северной части Охотского моря поедает наиболее массовые формы планктона — Metridia sp. (40—50% по весу), Themisto libelulla (местами до 90%), Calanus finmarchicus (10—15%), Pseudocalanus elongatus и Acartia longiremus (25—30%) (Кун, 1951). В Пенжинском зал. по исследованиям И. А. Полутова, поедает в основном Thysanoessa sp. У берегов Брит. Колумбии основную пищу взрослых составляют Copepoda, Euphausia pacifica и случайно икра сельди: весной главное значение имеет Calanus tonsus, в другие времена года большую роль в питании играет Euphausia pacifica (Wailes, 1935). Конкурентами в питании тихоокеанской сельди являются горбуша, скумбрия, колючая акула, навага и некоторые другие рыбы, а также в некоторые годы в августе медузы и, как у атлантической сельди, гребневики. В местах большого скопления гребневиков, а также и медуз в период летнего откорма, подобно атлантической сельди, встречается в меньшем количестве. Массовое развитие фитопланктона в период летнего откорма оказывает отрицательное влияние на распределение тихоокеанской сельди; весной, во время нереста, скопления фитопланктона не оказывают на нее влияния, и она проходит их (Кун, 1949). Нерестовая сельдь значительно менее жирна, чем более мелкая, не нерестующая. У западных берегов Камчатки, по данным К. И. Панина, нерестовая сельдь длиной 32—36 см имеет в теле 18.6—18.7% жира (на сухой вес), мелкие сельди, не нерестующие, длиной 24—26 см — от 23.7 до 32.4% жира. У берегов Приморья наибольшее количество жира содержит в теле летняя сельдь — 14.4—17.9% в мае—июле, нерестовая сельдь содержит жира значительно меньше — 7.4% в марте (Кизеветтер, 1942). Наибольшей жирностыо отличаются крупные (34—42 см длиной) сельди, вылавливаемые в летнее время (июль—август) в Кроноцком зал. и Авачинской губе у восточных берегов Камчатки, а также в Олюторском зал. осенью (сентябрь—октябрь) (жупановская сельдь), содержание жира в теле которых колеблется от 20.2 до 33.2% (Писарев, 1938; Панин). Значительно большим содержанием жира отличается нагульная летняя сельдь (до 25.7%) по сравнению с нерестовой (1.1—9.1%) и у западного побережья Сахалина (Леванидов, 1950).
Хозяйственное значение очень велико. Общий улов в 1936—1939 гг. колебался от 4 до 6.4 млн ц. Основная масса его добыта у южных берегов Сахалина (0.7—2.4 млн ц, около 3.7 млн ц в 1934 г.), у о. Хоккайдо (0.13—1.29 млн ц, 3.7 млн ц в 1934 г.), у берегов Кореи (0.3—0.5 млн ц), Сев. Америки (1.4—1.8 млн ц). В водах СССР в 1936—1940 гг. уловы составляли 425—523 тыс. ц (без южного Сахалина), из них в Японском море добыто 221—250 тыс. ц, в Охотском море — 147—195, в Беринговом море — 75—97 тыс. ц. Уловы в связи с колебаниями численности претерпевают большие изменения. В частности, у берегов южной части Сахалина уловы в 1931 г. достигли наибольшей величины — 5.3 млн. ц, в 1937—1938 гг. снизились до 0.7 млн ц, к 1940—1941 гг. возросли до 1.6—1.9 млн ц, к 1945 г. вновь уменьшились до 1.4 млн. ц. У берегов Хоккайдо наибольший улов до 9.7 млн ц был в 1897 г.; в дальнейшем уловы держались в среднем на несколько меньшем уровне. В двадцатых годах уловы резко снизились, в 1930 г. достигли всего лишь 2.8 млн ц, а к 1938 г. снизились до 0.13 млн ц, после чего вновь увеличились, не достигнув, однако, размеров высоких уловов предыдущих лет. Уловы в ряде промысловых районов могут быть увеличены путем освоения новых промысловых участков и лова в другое время года кроме весны, в частности жирующей летней сельди. Ловится во время нереста, в Приморье в марте—мае, в Амурском лимане в июне, у охотского побережья и западного Камчатки — в июне—июле, у восточных берегов Камчатки в мае—июне. Нагульную (жирующую) сельдь ловят в Приморье в июле и в сентябре—январе, у Сахалина и Камчатки — в июне—августе.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Clupea harengus pallasi Valenciennes. — Восточная сельдь
(У японцев — нисин (nishin), на Камчатке бельчуч, белая рыбка.)
сельди, на Камчатке бельчуч, белая рыбка Крашенинников, Описание Земли Камчатки, I, 1755, ч. 2, гл. VIII (в устьях рек, текущих в Охотском море, «бывают гостем, так что мне не более десяти рыб случилось видеть»; Берингово море: «в осень заходят они в великие озера и там плодятся и зимуют, а весною выплывают в море; примечания достоин лов их бывает в Вилючинском озере, которое от моря саженях только в 50, и посредством истока имеет с ним сообщение». В нижнем Камчатском остроге варят жир из бельчучей).
Hering, казаки на р. Камчатке Beeltschitschi Stеller, Beschreibung von dem Lande Kamtschatka, 1774, p. 167 (Берингово море от Авачинской бухты до Олюторского залива; спасаясь от преследований китообразных, заходят в реки и озера; в Вилючинском озере их ловят с марта по май; жители Нижнекамчатска ловят в устьях р. Камчатки в июле; о. Карагинский), р. 175 (с 1730 г. сельди стали подходить лишь единичными экземплярами к устью р. Камчатки).
Clupea harengus Pallas, 1. с., 1811, p. 209 (частью: in Oceano totam Camtschatcam alluente et in sinu Ochotensi), p. 210 (p. Камчатка).
Clupea pallasi Cuv.-Val., Hist. nat. des poissons, XX, 1847, p. 253 (Камчатка).
Clupea inermis Basilewsky, Nouv. Mem. Soc. Natur. Moscou, X, 1855, p. 242 (Шаньдун в Печилийском заливе; длина 216 мм).
Clupea pallasi Гребнидкий, Вестн. рыбопр., 1897, стр. 339 (Командорские острова, 1895 г., «появление ее совпало с приходом большого количества китов»).
Heringе der Ostküste Japans Heincke, 1. c., 1898, p. LXVI.
Clupea harengus var. pallasii Шмидт, Рыбы восточных морей, 1904, стр. 258; Морские пром. острова Сахалина. СПб., 1905, стр. 50—93.
Clupea pallasi Evermann and Goldsborough, Fishes of Alaska, 1907, p. 232 (Аляска от Pt. Clarence к югу).
Clupea harengus Павленко, Мат. к позн. русск. рыбол., III, вып. 10, 1914, стр. 141 (от зал. Посьета до Сахалинского залива, Петропавловск), стр. 1—140 (биология сельдей зал. Петра Великого и Татарского пролива).
Clupea pallasi Jordan and Metz, Mem. Carnegie Mus., VI, 1913—1914, p. 6 (Фузан, Chinnampo = Цзын-нам-пхо в сев.-зап. Корее).
Clupea harengus pallasi Берг, Рыбы пресн. вод России, М., 1923, стр. 502. — Амброз, Производ. силы Дальн. Востока, IV, 1927, стр. 61 (зал. Петра Великого; возраст, биология). — Рабинерсон, Изв. Отд. прикл. ихтиол., VIII, 1928, стр. 47 (зал. Петра Великого; возраст, размеры); Изв. Тихоокеанск. ст., I, вып. 1, 1928, стр. 143 (зал. Петра Великого; расовая характеристика). — Правдин, там же, стр. 192 (Усть- Болыперецк, Авачинская губа). — Линдберг и Дулькейт, Изв. Тихоокеанск. ст., III, вып. 1, 1929, стр. 7 (Шантарские острова, единично, 16—21 см). — Амброз, Рыбн. хоз. Дальн. Востока, № 2, январь 1930, стр. 30—34 (Лиман Амура, Охотск, Тауйск, Гижига, Ича на западном берегу Камчатки, Авачинская губа, Карага, Олютора, зал. Корфа, Анадырский залив); Изв. Тихоокеанск. инст. рыбн. хоз., VI, 1931 (зал. Петра Великого).
Clupea pallasi Tanaka, Fishes of Japan, fasc. XLIII, Tokyo, 1928, p. 844, fig. 484 (южные пределы в Японии: Toyama в Японском море, Hinuma prov. Hitachi на тихоокеанской стороне).
Позвонков столько же, сколько и у беломорской, наичаще 53—54, пределы колебаний 50—56; в среднем, по Павленко, 53.4 (118 экз.), по Рабинерсону, 53.6 (74 экз., зал. Петра Великого); по Амброзу; 53.2—53.7 (для разных проб из зал. Петра Великого, всего 423 экз). Жаберных тычинок на 1-й дуге 56—70, в среднем 63.9 (у 225 экз.). Спинной плавник несколько ближе к голове, чем у мурманской сельди: антедорсальное расстояние: самцы 43.3, самки 43.9% длины всего тела (у весенних сельдей из зал. Петра Великого, каковые имеются в виду и ниже); спинной и анальный плавники несколько длиннее, чем у мурманской: длина D самца 11.1, самки 11.0 %, Длина А самца 11.1, самки 10.5. Среднее число лучей в D и А такое же, как у беломорских, но несколько меньше, чем у мурманских. Килевых чешуй за V 11—12, впереди V они обычно отсутствуют или слабо развиты. На сошнике слабые зубы. Как и вообще тихоокеанские рыбы, сельдь эта крупнее и более скороспела, чем ледовитоморская и атлантическая; средние размеры ста самок, пойманных в марте—апреле 1926 г. в зал. Петра Великого, были в среднем 350 мм (абс. длина), ста самцов в среднем 335 мм; самки, пойманные там же в декабре 1911 г., имели в среднем 258 мм (абс. длина), самцы 256 мм. Павленко упоминает об экземплярах длиной 400—450 мм и до 500 мм (абс. длина).
Корея (западный и восточный берег), западный берег Японского моря, Татарский пролив, Сахалинский залив, Шантарские острова, все берега Охотского моря, берега Сахалина, сев. Япония, по берегу Японского моря на юг до зал. Тояма (36.66° с. ш.), на тихоокеанской стороне на юг до Hinuma, пров. Hitachi. Камчатка как по западному (немного), так и по восточному побережьям, Олюторский залив и севернее вплоть до Анадырского залива; изредка на Командорских островах. В Америке от Берингова пролива (Порт Кларенс) — на юг до Калифорнии (Сан-Диего). Тихоокеанская сельдь гораздо больше приспособлена к пресной воде, чем атлантическая. Она, подобно беломорской, массами подходит к устьям рек и иногда даже входит в устья. Так, сельдь осенью ловится в р. Суйфуне в 15 км от устья. Входит она также в низовья р. Тумень-ула (Л. Берг, Ежегодн. Зоол. муз. Акад. Наук, XIX, 1914, стр. 555.). Паллас (1. с., со слов Стеллера, см. также у Крашенинникова) сообщает, что на Камчатке она заходит в пресные озера и прежде массами входила в р. Камчатку. И в настоящее время наблюдается заход сельдей в чуть солоноватое Нерпичье озеро, что в устье р. Камчатки (А. Н. Державин, Русск. гидробиол. журн., II, 1923, стр. 193. По устному сообщению И. И. Лагунова (1939), в настоящее время в Нерпичьем озере вылавливают много сельди; она представляет озерную форму.).
Продолжительность жизни тихоокеанской сельди до 17—18 лет. Нерестится частью в возрасте 2 лет; трехгодовалые нерестятся все. Икрометание в зал. Петра Великого с начала марта до начала июня и даже июля. Рост сельдей, взятых в Уссурийском заливе (зал. Петра Великого) в марте—мае 1927 г., по данным непосредственных измерений, таков (длина до конца средних лучей хвостового плавника) в миллиметрах:
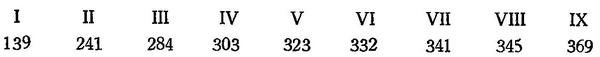
Преобладают трехгодовалые (1922 экз.), длина которых колебалась между 250 и 328 мм.
Темп роста весенней сельди зал. Петра Великого, по Рабинерсону (1928), таков в миллиметрах (абс. длина):

Сельдь, распространенная по американскому побережью от Аляски; до Калифорнии (Clupea mirabilis Girard 1854), тоже относится к группе pallasi; у сельдей из Британской Колумбии чаще всего 53 позвонка (в среднем 52.8), у калифорнийских чаще всего 52 (в среднем 51.8): (Hubbs). Такие же цифры приводит и Rounsefell (1930, р. 254), если прибавить позвонок hypuralia.
Clupea harengus pallasi. П. Л. Пирожников впервые наблюдал этих сельдей близ дельты Лены в августе 1945 г. (не 1946 г., как сказано у меня). В 1946 г. они ловились в Оленекском заливе (см. Пирожников, 1948).