Sardinops sagax (Jenyns, 1842)
Сардина-сардинопс, Тихоокеанская сардина, Тихоокеанский сардинопс, Тихоокеанская сардина-сардинопс, Южноамериканская сардина, Южноамериканский сардинопс, Южноамериканская сардина-сардинопс, Перуанская сардина, Перуанский сардинопс, Перуанская сардина-сардинопс, Чилийская сардина, Чилийский сардинопс, Чилийская сардина-сардинопс, Япономорская сардина, Япономорский сардинопс, Япономорская сардина-сардинопс, Дальневосточная сардина, Дальневосточный сардинопс, Дальневосточная сардина-сардинопс, Японская сардина, Японский сардинопс, Японская сардина-сардинопс, Иваси, Сардина иваси, Сельдь иваси, Калифорнийская сардина, Калифорнийский сардинопс, Калифорнийская сардина-сардинопс, Австралийско-новозеландская сардина, Австралийско-новозеландский сардинопс, Австралийско-новозеландская сардина-сардинопс, Австралийская сардина, Австралийский сардинопс, Австралийская сардина-сардинопс, Южноафриканская сардина, Южноафриканский сардинопс, Южноафриканская сардина-сардинопс, Сардинопс, Сардина, Пилчард, Сардинопс сагакс (Rus),
Pacific sardine, Pacific pilchard, Pacific American sardine, Pacific American pilchard, Peruvian Pacific sardine, Peruvian Pacific pilchard, Chilean sardine, Chilean pilchard, South American sardine, South American pilchard, Japanese sardine, Japanese pilchard, California sardine, Californian pilchard, Monterrey sardine, Sardina Monterrey, Australian sardine, Australian pilchard, Westralian sardine, Westralian pilchard, Southern African sardine, Southern African pilchard, Spotlined sardine, Spotlined pilchard, Blue sardine, Blue pilchard, Picton herring, Blue-bait, Bloater, Mulies, Smig, Sardina, Sardine, Pilchard
(Eng).
Синонимы:
Alausa californica Gill, 1862
Alosa musica Girard, 1855
Ambligaster melanostictum Jordan, Tanaka & Snyder, 1913
Arengus sagax (non Jenyns) Fowler, 1941
Clupanodon melanostictus Jordan & Snyder, 1900
Clupea advena Philippi, 1879
Clupea caerulea Mitchill, 1815
Clupea lata Richardson & Gray, 1843
Clupea melanosticta Temminck & Schlegel, 1846
Clupea neopilchardus Steindachner, 1879
Clupea ocellata Pappe, 1853
Clupea sagax Jenyns, 1842
Meletta caerulea Girard, 1854
Sardina sagax (non Jenyns) Regan, 1916
Sardinia melanosticta Jordan & Hubbs, 1925
Sardinia neopilchardus Steindachner, 1879
Sardinella melanosticta Jordan & Herre, 1907
Sardinella sagax (Jenyns, 1842) Тихоокеанская сардина (Rus), Pacific sardine, Pacific pilchard (Eng).
Sardinops melanosticta Kaganovskaya, 1935
Sardinops sagax coerulea Girard, 1854 Калифорнийская сардина (Rus), Californian pilchard, Blue-bait (Eng).
Sardinops sagax melanosticta (Temminck & Schlegel, 1846) Дальневосточная сардина, Иваси, Сардина иваси, Сельдь иваси, Япономорская сардина, Японская сардина (Rus), Japanese pilchard (Eng).
Sardinops sagax neopilchardus (Steindachner, 1879) Австралийско-новозеландская сардина, Австралийская сардина (Rus), Australian pilchard, Blue pilchard (Eng).
Sardinops sagax ocellata (Pappe, 1853) Южноафриканская сардина (Rus), Southern African pilchard (Eng).
Sardinops sagax sagax (Jenyns, 1842) Перуанская сардина, Чилийская сардина, Тихоокеанская сардина (Rus), Peruvian Pacific sardine, South American pilchard, Chilean sardine, Pacific sardine, Pacific pilchard (Eng).
Рыбы открытого океана. Н.В.Парин 1998 г.
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Известно, что в период высокой численности японского анчоуса и японской скумбрии в 50-е годы оба вида были весьма обычными и в открытых водах северо-западной части Тихого океана, в которых предполагалось даже существование их особых «стад» — в действительности, по всей вероятности, псевдопопуляций. В этом же районе в период высокой численности сардины иваси (Sardinops sagax melanosticta) в 30-е годы ее ареал расширялся вплоть до южной части Берингова моря (наблюдения в открытом океане отсутствовали), а современный подъем численности, начавшийся в 70-е годы (за первые 5 лет роста запас возрос не менее чем вдвое), привел к расширению области распространения в восточном направлении вплоть до центральной части океана, где, судя по некоторым сведениям, также образовывались временные псевдопопуляции.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Сардина дальневосточная (иваси) — Sardinops sagax melanosticta (Schlegel, 1846)
Тело в сечении округлое. Характерный признак рода — наличие на основании хвостового плавника крыловидных чешуй. Окраска типичная для сельдевых: на боках тела, ближе к спине, контрастные пятна черного цвета. Достигает длины 30 см. Океанический вид, но подходит к берегам. Обитает в морях Дальнего Востока, на север до Камчатки. Ценный объект промысла, известный под торговым названием сельдь-иваси.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
В роде сардинопс имеется только один
вид — сардина-сардинопс (Sardinops
sagax), состоящий из пяти подвидов.
Дальневосточная сардина (Sardinops sagax melanosticta) распространена у берегов Восточной
Азии от Сахалина до Южной Японии и
китайского побережья Желтого моря (Чифу). Калифорнийская сардина (Sardinops sagax
coerulea) обитает в водах тихоокеанского
побережья Северной Америки от
Северной Канады до Южной Калифорнии.
Перуанская сардина (Sardinops sagax sagax)
распространена у берегов Перу; австралийско-новозеландская (Sardinops sagax neopilchardus) —
в водах Южной Австралии и Новой
Зеландии; южноафриканская (Sardinops sagax ocellata) — в водах Южной Африки.
Обитатели бездны. Уильям Кроми 1971 г.
Воды у побережья Калифорнии буквально кишат анчоусами (Engraulis mordax), которых там такое количество, что, как полагают, можно ежегодно вылавливать до 1 миллиона тонн этой рыбы. Анчоусы по пищевым качествам не уступают тихоокеанским сардинам (Sardinella sagax), и их промысел помог бы восстановить запасы сардин, которые начиная с 40-х годов уменьшаются с катастрофической быстротой. Однако владельцы консервных заводов не заинтересованы в увеличении добычи мелких анчоусов, поскольку расфасовка их невыгодна из-за высокой стоимости рабочей силы. Использование анчоусов для изготовления рыбной муки или протеинового концентрата в Калифорнии затруднено законами об охране природы.
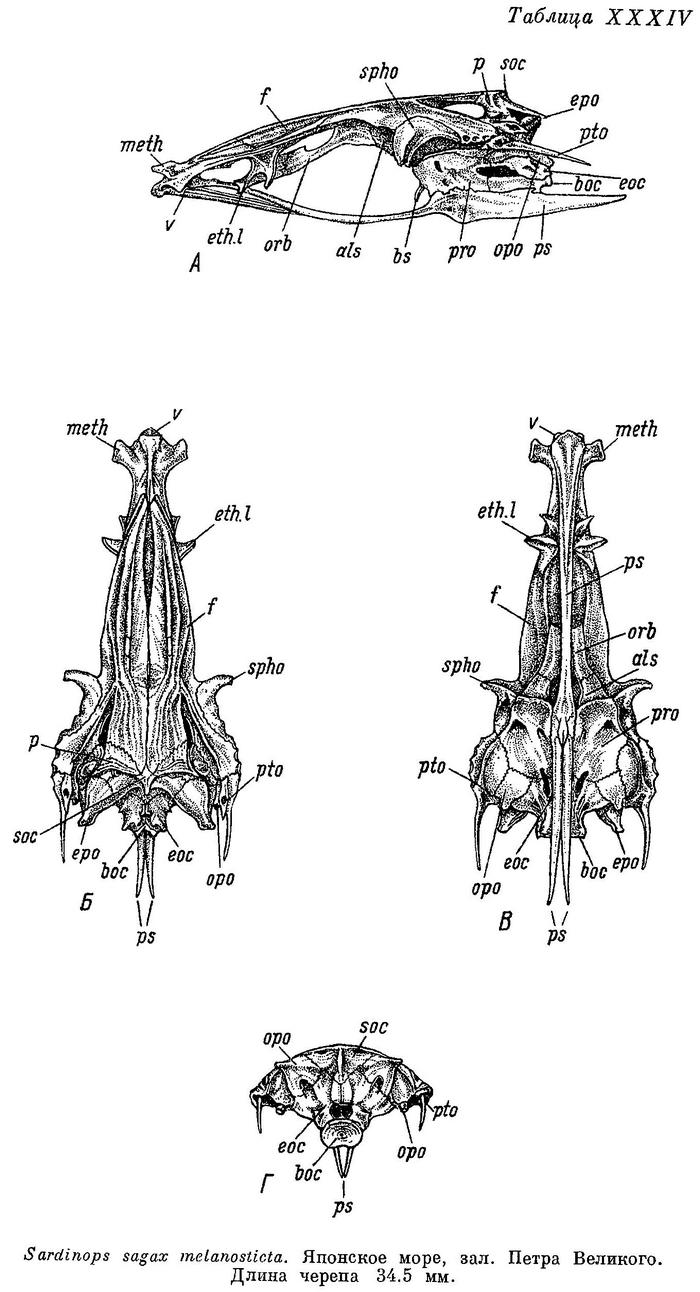
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
1. Sardinops sagax melanosticta (Schlegel, 1846) — Дальневосточная сардина, иваси (рис. 80).
Clupea melanosticta Temminck et Schlegel, Fauna Japonica, Poiss., 1846 : 237, tab. 107, fig. 3 (берега Японии).
Sardinops melanosticta melanosticta, Таранец, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океанограф., 14, 1938 : 69 (сравнение с калифорнийской сардиной).
Sardinops sagax coerulea n. melanosticta, Берг, Рыбы пресных вод, 1948 : 111, сноска 4.
Sardinops sagax melanosticta, Световидов, Фауна СССР, Рыбы, 2, 1, 1952 ; 178, рис. 21—23, табл. 6, фиг. 1 (синонимия, описание).
Sardinops melanosticta, Matsubara, Fish Morphol. a. Hierar., 1955 : 191 (Южный Сахалин, Приморье, п-ов Корея — по всем берегам, побережья китайских морей). — Кагановский, Промысловые рыбы СССР, 1949 : 77—82; атлас, табл. 19 (цветной рисунок).

D III—IV 14—16, А II—IV 15—18, жаберных тычинок 97—140 (в среднем 122), на нижней половине жаберной дуги 58—93 (78.4), позвонков 50—52 (50.73). Голова удлиненная и узкая, длина ее 21.1% длины тела, межглазничный промежуток 17.2% длины головы. Грудные плавники 13.3% длины тела. Тело удлиненное, наибольшая высота его 17.1%, высота головы у затылка 13.3% длины тела. (50 экз. из Японского моря и против северной части Корейского залива). Спина зеленовато-оливковая, бока серебристо-белые, конец рыла и задняя часть хвостового плавника черные, на боках тела с каждой стороны по ряду темных пятен числом от нескольких до 15, ниже которого иногда имеется второй и изредка третий ряд из меньшего числа более светлых пятен. (Световидов, 1952а : 180).
Длина до 292 мм.
Пелагическая рыба, обитающая в водах при температуре от 15 до 25°. Сводка по ее биологии дана в работе А. Н. Световидова (1952а). Дополнительно укажем следующие русские работы: Богаевский, 1955; Кагановский, 1955; Перцева-Остроумова, 1955; Парин, 1959; Пробатов, 1953; Токарев, 1955, а также некоторые иностранные работы: Ito et all., 1954, 1960, 1961; Nishimura, 1960; Ronguillo, 1959; Tamura, 1952; Yasuda a. Ogura, 1950; Yoneda a. Yoshida, 1955. Особо следует отметить весьма обстоятельную работу японского ихтиолога Накаи (Nakai, 1962) по выяснению причин колебания уловов японской сардины за последние 50 лет.
Важная промысловая рыба. В СССР добывалась с 1925 по 1942 г. В настоящее время в наших водах встречаются лишь отдельные экземпляры (район пос. Яблочное, Южный Сахалин и зал. Анива; Пробатов, 1951 : 147).
Распространение. В Японском море встречается преимущественно в юго-восточной, а единичные экземпляры в северо-восточной частях. В Желтом море известен у западного побережья п-ова Корея, о. Чечжудо (Mori, 1952 : 31) и района Чжифу (Коо, 1933 : 146). В Японии вдоль тихоокеанского побережья к югу от Хоккайдо; Тайвань (Fowler, 1941 : 623). В годы потеплений встречалась на север до лимана Амура, восточного Сахалина (Световидов, 1952а : 181), восточной Камчатки (Авачинский и Кроноцкий заливы: Панин, 1936а : 41, 19366 : 149) и у Командорских островов (Линдберг, 1935а : 47).
Наши коллекции из Японского моря — из устья Тумыньцзяна, зал. Петра Великого, бухты Сяуху и Квандагоу, Хакодатэ, а также из Кагосимы и Нагасаки.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*1. Sardinops sagax (Jenyns).
Clupea sagax Jenyns, Zool. «Beagle», Fish., 1842 : 134 (Лима, о. Сан-Лоренцо, Перу).
В отдельных изолированных частях обширного ареала распадается на ряд подвидов: Sardinops sagax sagax (Jenyns, 1842; тихоокеанские берега Южн. Америки), Sardinops sagax melanosticta (Schlegel, 1846; берега восточной Азии), Sardinops sagax ocellata (Рорре, 1853; берега южной Африки), Sardinops sagax caerulea (Girard, 1854; тихоокеанские берега Сев. Америки), Sardinops sagax neopilchardus (Steindachner, 1879; вокруг Новой Зеландии, у южных, восточных и западных берегов Австралии и северных Тасмании). Все эти формы рядом авторов признаются идентичными, за исключением последней, или самостоятельными видами. Сравнительно-морфометрическим изучением Sardinops sagax melanosticta и Sardinops sagax caerulea обнаружено (Таранец, 1938), что между этими двумя формами имеются различия, но незначительные, и поэтому их следует правильнее считать подвидами. Приблизительно таковы же, судя по всему, различия и между остальными формами. Несколько более отличается лишь Sardinops sagax neopilchardus (Regan, Ann. Mag. Nat. Hist. (8), XVIII, 1916 : 11—14; Thompson, Fish. a. Game Comm. Calif., Fish. Bull. 11, 1926 : 8—17. Сведения по биологии, главным образом по возрасту и росту, этой формы см.: Blackburn М. Age, rate of growth, and general life-history of the Australian pilchard (Sardinops neopilchardus) in New South Wales waters. Commonwealth Sci., Ind. Res. Org., Australia, Melb., Bull. 242, 1949 : 1—86, 8 pis.), но, судя по имеющимся данным, и эта форма, подобно другим, резких отличий от остальных не имеет. Поэтому все формы провизорно, до детального сравнительного изучения их на большом материале, правильнее всего считать подвидами.
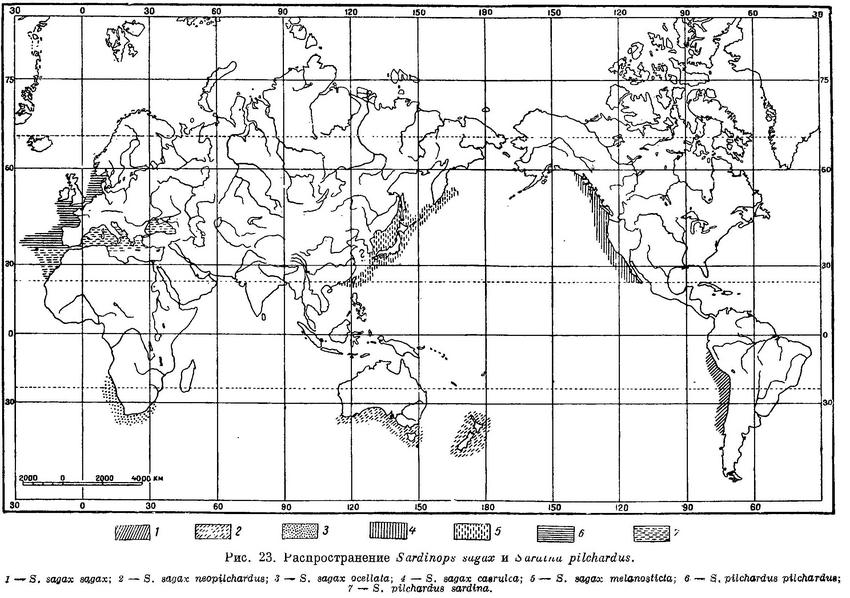
Распространение. Тихий океан по обе стороны в северном и южном полушариях, Индийский океан. У берегов восточной Азии от Кантона, вокруг Японии, по обе стороны почти до северной оконечности Сахалина, у берегов Сев. Америки от Калифорнии до м. Омманей (юго-восточная Аляска), у берегов Южн. Америки, вокруг Новой Зеландии, у Тасмании и южных берегов Австралии от Квинслэнда до западных берегов, у южной оконечности Африки от бухты Столовой и Фольс до Наталя (рис. 23).
В наших водах один подвид.
*la. Sardinops sagax melanosticta (Schlegel) — Япономорская сардина, дальневосточная сардина, иваси (табл. VI).
— Clupea melanosticta Schlegel, in: Temminck et Schlegel, Fauna Jap., Pisces, 1846 : 237, t. CVII, f. 3 (берега Японии); Günther, Cat. fish., VII, 1868 : 430 (Китай, Япония).
— Clupea sagax (non Jenyns) Günther, l. c.: 430 (частью: Япония).
— Clupanodon melanostictus Jordan a. Snyder, Proc. U. S. Nat. Mus., XXIII, 1900 : 349 (Токио).
— Clupea melanosticta Kishinouye, Journ. Imp. Fish. Bureau, Tokyo, XIV, 3, 1907 : 71, 94, 97, t. XVlI, f. 1—2, t. XXI, f. 1 (описание черепа, второй ряд пятен на теле).
— Sardinella melanosticta Jordan а. Herre, Proc. U. S. Nat. Mus., XXXI, 1907 : 632 (Хакодате, Токио, Иокогама, Цуруга, Аомори, Мисаки, Саме, Татейяма, зал. Матцусима, Ваканоура, Кобе, Ономичи, Хаката, Нагасаки).
— Ambligaster melanostictum Jordan, Tanaka a. Snyder, Cat. fish. Japan, 1913 : 37 (берега Китая, Кореи, Японии, Отару, Мисаки, Сагами)
— Sardina sagax (non Jenyns) Regan, Ann. Mag. Hist. (8), XVIII, 1916 : 13 (частью: Япония).
— Sardinia melanosticta Jordan a. Hubbs, Mem. Carnegie Mus., X, 2, 1925 : 121 (Мисаки, Микава, Нагоя, Токио, Тоба, Кусиро, Такасима).
— Ambligaster melanostictus Fowler, Proc. Acad. Nat. Sci. Phila., LXXXI, (1929), 1930 : 590 (Токио).
— Sardinops melanosticta Кагановская, Рыбн. хоз. Дальн. Вост., XIII, I, 1935 : 38 (систематическая характеристика); Таранец, там же : 40 (сравнение с калифорнийской сардиной); Кагановская, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., XIV, 1938 : 45 (систематическая характеристика).
— Sardinops melanosticta melanosticta Таранец, там же : 69 (сравнение с калифорнийской сардиной).
— Arengus sagax (non Jenyns) Fowler, U. S. Nat. Mus., Bull. 100, v. 13, 1941 : 622 (частью: Китай, Тайвань, Корея, Япония).
— Sardinops sagax melanosticta Кагановский, в: Промысловые рыбы СССР, 1949 : 79, табл. 19.
Распространение, биология. Сноу, Зап. Общ. изуч. Амурского края, VIII, 1, 1902 : 50 (о. Кунашир — южный остров Курильской гряды); Franz, Abh. Bayer. Akad. Wiss., IV, Suppl.-Bd. 1, 1910 : 5 (зал. Сагами, Абуратсубо, Фукуура); Snyder, Proc. U. S. Nat. Mus., XLII, 1912 : 402 (Отару, Мороран, Томакомай, Хакодате, Мисаки); Павленко, Мат. позн. русск. рыбол., III, 10, 1914 : 145 (устье р. Тумень-ула); Берг, Ежегодн. Зоолог. муз. Акад. Наук, XIX, 4, (1914), 1915 : 555 (там же, 24 и 26 IX 1913); Jordan a. Hubbs, Mem. Carnegie Mus., X, 2, 1915 : 121 (Мисаки, Микава, Нагоя, Токио, Тоба, Кусиро, Такасима); Солдатов и Линдберг, Обзор рыб дальневосточн. морей, 1930 : 38, табл. 3; Амброз, Рыбн. хоз. Дальн. Вост., 1930, 3—4 : 33 (причины появления в наших водах); Инясевский, там же : 5 (промысел); Шмидт, Изв. АН СССР, отд. мат. и ест. наук, 1931 : 103 (Обама); Тр. Тихоок. комит. АН СССР, II, 1931 : 19 (Нагасаки, Кагосима); Сhu, Index pise. Sinens., 1931 : 14 (Кантон, Китай, Корея, берега южн. Японии); Mori a. Uсhida, Journ. Chosen Nat. Hist. Soc. 19, 1934 : 4 (Вонсан, Пусан); Кагановский, Рыбн. хоз. Дальн. Вост., 1934, 1—2 : 11 (наблюдения над ходом); Кагановская, там же : 34 (биология); Гайл, там же : 52 (питание); Линдберг, Природа, 1935, 5 : 47 (появление у берегов Камчатки в 1933 и 1934 гг., Командорские о-ва в 1934 г., Приморье в 1923 г.); Кагановский, Рыбн. хоз. Дальн. Вост., XIII, I, 1935 : 35 (биология); Андрияшев, Исслед. морей СССР, 22, 1935 : 141 (Авачинская губа, Кроноцкий зал., Командорские о-ва); Берг, Пробл. физич. геогр., II, 1935 : 77 (причины появления у берегов Камчатки); Бродский и Янковская, Вестн. Дальневост. фил. Акад. Наук, 13, 1935 : 103 (питание); Бродский, там же, 18, 1936 : 159 (питание); Янковская, там же : 63 (питание); Панин, Докл. АН СССР, III (XII), 1 (96), 1936 : 41 (восточные берега Камчатки: Кроноцкий зал., 1933 г.; Кроноцкий зал., бухта Моржовая, Авачинский зал. и Авачинская губа, 1934 г.); Рauin, Zool. Anz., 115, 5 6, 1936 : 149 (там же); Amеmiуa, Japan. Journ. Zool., VI, 4, 1936 : 79 (изменение количества позвонков у берегов Японии); Udа а. Оkamоtо, Journ. Imp. Fish. Exp. St., 7, 1936 : 47 (связь с температурой); Кimura Bull. Jap. Soc. Sci. Fish. Tokyo, v. 6, 2, 1937 : 69 (сезонные изменения коэффициента упитанности); Оya, Usui a. Sukegawa, ibid., v. 5, 5, 1937 : 308 (сезонные изменения жирности); Таранец, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 12, 1937 : 33 (Охотское м. у восточных берегов Сахалина: зал. Ныйво и Чайво); Кагановская, там же : 115 (расчисление роста); Евстафьев, Лямин и Маконогова, Учен. зап. Ленингр. Гос. унив., III, 15, 1937 : 302 (Приморье; уловы, размеры, возраст, возрастной состав, рост); Андрияшев, Исслед. морей СССР, 25, 1937 : 294 (восточные берега Камчатки); Кагановский, Дальневост. сардина, изд. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 1939 : 3—39; промысл. разведка иваси, там же, 1939 : 9 и сл. (биология, промысел); Румянцев и Голубев, Рыбн. хоз., 1940, 3 : 24 (ход в Приморье в 1939 г.); Aikawa, Bull. Jap. Soe. Sci. Fish. Tokyo, v. 8, 5, 1940 : 227 (возраст, количество позвонков, миграции, места нереста); Rec. oceanogr. works Jap., XI, 2, 1940 : 81 (возраст); Кизеветтер, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 21, 1942 : 98 и сл. (содержание жира); Шмидт, Рыбн. промышл., 1, 1945 : 3 (причины исчезновения в 1941 г. и следующих годах); Кагановский, там же, 3, 1945 : 8 (причины исчезновения в те же годы); Clemens a. Wilby, Fish. Res. Board Canada, Bull. LXVIII, 1946 : 77 (берега Сев. Америки); Румянцев, Зоолог. журн., XXVI; 1, 1947 : 47 (время появления в наших водах); Городничий, Рыбн. хоз., 1949, 1 : 36 (колебание уловов в связи с изменением температурного режима моря); Кизеветтер, Изв. Тихоокеанск. научно-исслед. инст. рыбн. хоз. и океаногр., 31, 1949 : 209 (содержание жира); Пробатов, Докл. АН СССР, LXXVII, 1, 1951 : 147 (западный берег южной части Сахалина в районе Яблочное, 1 экз., 1, Х 1949).

D III—IV 14—16 (15.4), общим числом 18—20 (19.3), А II—IV 15— 18 (15.8), общим числом 17—21 (18.7), sp. br. 97—140 (122.0), на нижней половине жаберной дуги 58—93 (78.4), vert. S. 50—52 (50.73), caud. vert. 30—33 (31.13). Голова удлиненная и узкая, длина ее 19.9—22.5 (21.1)% длины тела, межглазничный промежуток 15.6—18.6 (17.2)% длины головы. Грудные плавники 12.3—14.3 (13.3)% длины тела. Тело удлиненное, наибольшая высота его 15.6—18.9 (17.1)%, высота головы у затылка 12.5—14.2 (13.3)% длины тела. Спина зеленовато-оливковая, бока серебристо-белые, конец рыла и задняя часть хвостового плавника черные, на боках тела с каждой стороны по ряду темных пятен числом от нескольких до 15, ниже которого иногда имеется второй и изредка третий ряд из меньшего числа более светлых пятен (Сардин с тремя рядами пятен встречал также А. Г. Кагановский.). Размеры до 292 мм, наименьшая длина в наших водах за ряд последних лет 158 мм, наичаще 155—240 мм. Описание дано по 50 экз. длиной от 199 до 263 в среднем 220.1 мм, собранным в июне—июле 1947 г. в Японском море против северной части Корейского зал. По исследованиям Кагановской (1938), у 158—233 экз. длиной 190—290, в среднем 228.1 мм (до конца хвостового плавника, до конца средних лучей его в среднем 208.3 мм) из б. Троица D III—IV 14—17 (15.2), A III—IV 14—17 (15.8), жаберных тычинок 126—174 (144.6), килевых чешуй 16—19 + 13—16 (17.9 + 14.1), рядов чешуй 50—57 (52.3), позвонков 48—53 (50.5), пилорических придатков 75—120 (99.0); по исследованиям Таранца (1938), у 61—76 экз. длиной 180—240, в среднем 205.1 мм (до конца средних лучей хвостового плавника) из зал. Ольги и бухты Преображения жаберных тычинок 120—170 (142.4), поперечных рядов чешуй 52—57 (54.0), позвонков 50—51 (50.6); по данным К. И. Панина (Андрияшев, 1937), у 223 экз. длиной от 140 до 200 мм (какой не указано) от восточных берегов Камчатки жаберных тычинок 108—139 (123.5); по Ригэну (Regan, 1916), жаберных тычинок на нижней половине жаберной дуги 70—100.
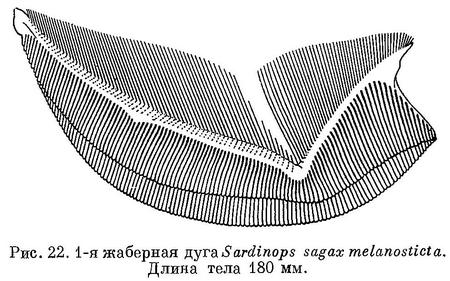
На протяжении вытянутого в меридиональном направлении ареала не однородна: у сардины, локализованной в северной части ареала, позвонков несколько больше, чем у сардин из южной части ареала. Так, например, вдоль обоих берегов Японии ловится сардина с количеством позвонков от 50.72 (о. Хоккайдо) до 50.48 (о. Кю-сю) (Amemiya, 1936). Равным образом и в наших водах у берегов Приморья в начале ловится сардина с большим количеством позвонков — в среднем 49.73, по наблюдениям в 1932 г. в бухтах Преображения и Троица (Кагановская, 1938), которую в дальнейшем на протяжении хода сменяет сардина с меньшим количеством позвонков — в среднем 49.66, причем параллельно наблюдается изменение и величины антедорсального расстояния в среднем от 43.88% в начале хода до 42.86% длины тела (до конца средних лучей хвостового плавника) в конце. (Об изменении длины тела и пр. см. ниже в разделе «Биология»). Обусловлено это тем, что при летних миграциях происходит смещение ареала с юга на север, и поэтому в наших водах сперва появляется сардина, локализованная в северной части ареала, которую в дальнейшем сменяет сардина более южной части ареала. Изменение количества позвонков и пр. происходит в общем постепенно и довольно закономерно, но каких-либо обособленных группировок не обнаружено. Сходным образом с севера на юг изменяется количество позвонков и у Sardinops sagax caerulea, что особенно наглядно видно у сеголеток: север Калифорнии (к северу от 35° с. ш.) 51.77, юг Калифорнии, (к северу от 32°50' с. ш.) 51.75, север Нижней Калифорнии (к северу от 27° с. ш.) 51.62, юг Нижней Калифорнии (к северу от 24°30') — 51.30, м. Сан-Лукас и Калифорнийский зал. 51.05 (Clark, Fish. Bull., 65, 1947).
Распространение. Западная часть Тихого океана и прилегающие моря. На юг у берегов Китая до Тайвана и Кантона. Восточно-китайское (Нагасаки) и Японское моря. В Японском море вдоль обоих берегов до самого севера, вдоль западных от Кореи (Пусан, Вонсан) до м. Лазарева в Татарском прол., иногда до южной части Амурского лимана. Вдоль восточных берегов Японии до Курильских о-вов, единично встречалась в Охотском море у восточных берегов Сахалина (зал. Ныйво и Чайво), в Беринговом море до берегов Камчатки (Авачинская губа, Кроноцкий зал.) и Командорских о-вов. В северных частях ареала (к северу от южной части о. Хондо) в зимнее время отсутствует. Икрометание в Японском море от южной оконечности о. Кю-сю до зал. Тояма на о. Хондо, вдоль тихоокеанских берегов Японии к югу от зал. Сагами (о. Хондо); по материковому берегу Японского моря в наших водах, как правило, нереста нет, но единично зрелые и текучие встречались в южном Приморье.
Биология. Нерест с декабря по начало мая, в южных частях ареала раньше, чем в северных: у южной оконечности о. Кю-сю (Кагосима) — в конце декабря — начале января, у о. Хондо (у берегов пров. Фукуи) — в феврале, в районе зал. Тояма — в марте — начале апреля, на восточном побережье Японии несколько раньше, чем на западном (Aikawa, 1940). В годы с низкой температурой в январе—марте на юге Японского моря зрелые и недавно отнерестовавшие, по наблюдениям А. Г. Кагановского, встречались до августа месяца у берегов северной части Приморья. Икрометание происходит в бухтах и заливах, по некоторым указаниям (Uda a. Okamoto, 1936), в прибрежных областях открытого моря (против о. Хондо), при температуре воды 13—15°, но наблюдалось и при более низкой, 11—12°, точно так же, как и при более высокой, до 20°. Плодовитость от 27 до 84, в среднем 45 тыс. икринок. Диаметр икринок неодинаковый, что указывает на возможность порционного икрометания (это отмечено и у Sardinops sagax caerulea). Развитие икры при 18° длится 2—4 дня. Молодь в течение первого года совершает, судя по всему, ограниченные миграции, но в некоторые годы заходит даже в Приморье. Взрослые в зимнее время держатся в южных частях Японского моря и в прилегающих Восточно-китайского. Миграции на север начинаются с марта, отчасти с февраля, когда температура воды подымается до 8° и выше. Массовые подходы к берегам происходят при температуре воды от 10°, при прогревании воды до 19—20° отходит, в водах Приморья встречалась при 23—24°, а в южной части Японского моря и при 25°, так же как при похолодании встречается при 6° и даже 4°. При быстром похолодании погибает уже при 6°, при постепенном понижении температуры воды выживает и при 4°, но становится вялой, малоподвижной. В зал. Петра Великого обнаружена в декабре при 2°. Молодые, неполовозрелые более теплолюбивы, температурные условия миграции и подходов для них выше, повидимому 15—26°. Массовый отход при обратных миграциях в южном Приморье начинается при 9—10°, в северном при 10—11°. По отношению к колебаниям солености гораздо более чувствительна. Обычно держится при 29.9—35.0‰ содержания солей, но в Амурском зал., по указанию А. Г. Кагановского, в дождливое время, когда соленость значительно снижается, встречалась и при 20‰; известны случаи захода в устья рек, вероятно с приливами. В Японском море из Цусимского прол. на север продвигается по обе стороны его: вдоль берегов Кореи и далее Приморья, с одной стороны, и Японии и Сахалина, с другой, причем часть сардины через проливы Сангарский и Лаперуза уходит в Тихий океан. Обратные миграции на юг происходят теми же путями, причем значительная или большая часть сардины от восточного берега Японского моря, от Сахалина, переходит к западному и двигается к местам зимнего пребывания вдоль берегов Приморья. С тихоокеанской стороны Японии одна часть сардины продвигается вдоль берегов (неретический путь), другая — в открытых частях океана (океанический путь) в струях Куро-сиво (Aikawa, 1940). В начале миграций идет с юга на север с водами теплого течения и при установлении благоприятных температурных условий устремляется к берегам из прилегающих участков моря, где держится вблизи берегов, а не по всей ширине моря, так как в открытых частях его вода, повидимому, менее богата планктоном, чем воды прибрежные. Это подтверждается наблюдениями в районе зал. Петра Великого и к югу от него и с самолета, причем сардина здесь наблюдалась на расстоянии не далее 20—30 миль от берега. В восточной половине Японского моря в начале хода идет не далее 8 миль от берега и лишь к северу, у северной части о. Хондо (против Акита) до 30 миль и у берегов Кореи иногда на 80—90 миль от берега (Uda a. Okamoto, 1936); в северной части Приморья (к северу от бухты Нельма) и у южной оконечности Сахалина идет, судя по всему, несколько в большем отдалении от них. Обратные миграции на юг, по указанию А. Г. Кагановского, происходят на большем расстоянии от берегов, так как осенью у берегов после штормов наступает резкое похолодание воды, между тем как вдали от берегов находится более теплая вода. При этом в годы с ослабленным холодным течением и близким прохождением к берегу теплого течения миграции начинаются раньше и рыба идет ближе к берегу. В годы усиленной деятельности холодного течения, отжимающего теплое течение к востоку, в начале хода держится вдали от берегов. Скорость движения по мере продвижения на север увеличивается, что обусловлено более быстрым прогреванием воды в северных районах. У западных берегов Японского моря вдоль Кореи (от Кейхоку до Конхоку), по указанию А. Г. Кагановского, идет со скоростью около 15.2 км (от 11.0 до 18.5 км), вдоль берегов Приморья (от зал. Посьета до бухты Светлой) — со скоростью 26 км. У восточных берегов Японского моря (Uda a. Okamoto, 1936) вдоль о. Хондо (от Фукуи до Аомори) идет со скоростью 13.6 км (от 9.0 до 18.0 км), вдоль о. Хоккайдо (от Осима до Китами) — со скоростью 23.9 км (от 13.0 до 26.0 км). Массовые подходы в южной части Приморья (Хасанский район) на протяжении 10 лет (1929—1938 гг.) начинались от 19 мая по 24 июня, в северной части (Советский район) — от 1 июля по 15 августа. При обратных миграциях на юг массовые подходы на протяжении тех же 10 лет в северной части Приморья (Советский район) заканчивались от 18 октября по 5 декабря, в южной части (Хасанский район) — от 31 октября по 19 ноября. Таким образом, продолжительность периода массовых подходов в южной части Приморья значительно больше, чем в северной: в Хасанском районе, по данным А. Г. Кагановского, период массовых подходов на протяжении того же десятилетия колебался от 132 до 178 дней, между тем как на севере, в Советском районе, продолжительность его колебалась от 65 до 126 дней. Вертикальное распределение на протяжении хода неодинаково. В начале лета (в июне), когда нагрет лишь сравнительно небольшой поверхностный слой воды, держится на небольшой глубине (от 5 до 12 м). При дальнейшем прогревании воды на большие глубины занимает большую толщу воды. Глубокая гомотермия с температурой воды выше 20° вызывает отход в более холодные участки моря. В начале хода (июнь—июль) обычно идет рыба более крупных размеров и большего веса, более старшего возраста и более быстрого темпа роста, затем более мелкая, имеющая минимальные размеры в сентябре. Параллельно с изменением размеров, веса и пр. изменяется и коэффициент упитанности. Такая же последовательность хода наблюдается и у берегов Японии (Uda a. Okamoto, 1936). В конце хода (октябрь—ноябрь), при обратных миграциях на юг, мелкую рыбу сменяет опять более крупная, но не достигающая размеров, которые наблюдались в начале хода, причем в некоторые годы, вследствие того что во время обратного хода рыба идет дальше от берегов, крупная сардина в конце хода не ловится, что видно из следующих данных А. Г. Кагановского:

Таким образом, в северном Приморье, куда не доходит мелкая сардина, зимующая и размножающаяся в самых южных частях ареала и идущая в конце, средние размеры ее крупнее, чем в южном. Сардина из южных частей ареала при одинаковом возрасте имеет меньшие размеры и при одинаковых размерах меньший вес, чем из северных (Кагановский, 1939). Основной пищей является зоопланктон, в частности Copepoda, которые составляют главную массу содержимого желудков в период усиленного кормления. Значительную роль в питании играет фитопланктон, но, повидимому, в качестве вынужденного корма. Большое количество растительных организмов встречается в желудках обычно в период менее интенсивного кормления, когда желудки бывают слабо наполнены или пусты. Постоянными компонентами пищи из Copepoda являются: Oithona similis, Paracalanus parvus, Microstella norvegica, nauplii и juv. Copepoda, встречаемые в течение почти всего периода времени с июня по октябрь. Из растительных организмов таковыми являются: Coscinodiscus sp., Chaetoceras sp., Thalassionema nitzschoides, Skeletonema costatum, Bacteriastrum sp., Dinophysis acuta. В период усиленного кормления в сильно наполненных желудках в массе встречались Centropages mcmurrichi, Calanus finmarchicus, Pseudocalanus elongatus. Наиболее интенсивно, по наблюдениям, питается осенью (сентябрь, октябрь), летом (июль, август) желудки наполнены мало (Бродский и Янковская, 1935; Бродский, 1936; Янковская, 1936). Показателем отсутствия сардины является наличие гребневиков, медуз, а также и сельди, которые держатся в более холодной воде (Кагановский, 1939). Половой зрелости достигает в основной массе в двухгодовалом возрасте, после чего мечет икру ежегодно. Семи- и восьмигодовалые самки часто оказываются стерильными. У годовалых и мелких двухгодовалых в начале хода пол невооруженным глазом почти не различим. Взрослые летом в наших водах имеют половые продукты во II стадии зрелости, единичные — во II—III; изредка в июне, июле и даже в августе попадается еще не отнерестовавшие, со зрелыми половыми продуктами. В исключительных случаях, иногда в значительном количестве, рыба со зрелыми половыми продуктами встречается в июне и июле в Сангарском прол. (Aikawa, 1940). У двухгодовалых количество самцов и самок приблизительно равно, затем у более взрослых количество самок возрастает, у старших возрастных групп количество самок всегда больше количества самцов. В уловах сетей количество самок в среднем несколько больше количества самцов (в среднем 63.5% первых и 56.5% вторых), в уловах неводов ловятся в большом количестве самцы. В уловах преобладают трех- и четырехгодовалые, последние в начале хода, первые в конце, двухгодовалые встречаются редко, но в некоторые годы заходят в значительном количестве, одногодовалые встречаются единицами процентов, пятигодовалые — в значительном количестве, в некоторые годы в большом, шести- и семигодовалые — в небольшом количестве, последние не каждый год, совсем редко встречаются восьмигодовалые. Возрастной состав в уловах различных орудий лова таков (в процентах):
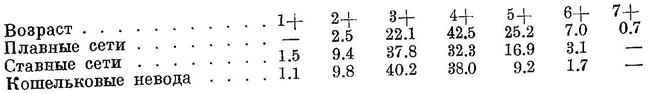
Неводами ловится более мелкая рыба размерами наичаще 15.5—19.0 (14—17.5 до конца средних лучей хвостового плавника) и весом 30—75 г, в сети попадает более крупная рыба длиной наичаще 19.5—24 см (18—22 до конца средних лучей хвостового плавника) и весом 70—120 г. Размеры ловимой сардины из года в год меняются; наиболее крупная рыба ловилась в 1928—1930 гг., в 1931 г. — менее крупная, к 1933 г. более крупная, с 1935 по 1938 гг. наблюдалось уменьшение средних размеров и веса ловимой сардины с сопутствующим увеличением среднего возраста. Рост, по исследованиям А. Г. Кагановского, происходит следующим образом (зал. Посьета; см, длина до конца средних лучей хвостового плавника):

По содержанию жира стоит на одном из первых мест среди представителей семейства, уступая в этом отношении лишь немногим видам. Содержание жира в теле сардины из различных районов наших вод за период времени с 1929 по 1940 г. колебалось от 11.32 до 28.27%, среднее — 19.16%. Крупные сардины содержат жировых отложений больше, чем мелкие, как это можно видеть из нижеследующего: длина тела 20.5 см — содержание жира 17.53%, 21.5 см — 18.93%, 22.5 см — 19.45%, 23.5 см — 19.98%, выше 24 см — 20.49% (Кизеветтер, 1942).
Появление сардины в наших водах ставилось в связь с общим потеплением (Берг, 1935), с землетрясением 1923 г., колебанием запасов сельди (Амброз, 1930) или полагалось (Линдберг, 1935; Кагановский, 1939), что она встречалась постоянно и ранее, и нормальной северной границей ее ареала следует считать южные воды восточного побережья Камчатки, где она была обнаружена в сентябре 1933 г. в Кроноцком зал., и в августе, сентябре, октябре и начале ноября 1934 г. в Кроноцком. зал., бухте Моржовой и в Авачинском зал. и Авачинской губе (Панин, 1936), а также у Командорских о-вов (Андрияшев, 1937), где она указывалась и ранее (Сноу, 1902). В Приморье сардина была находима в зал.. Петра Великого в 1910 г., в устье р. Тумень-ула — в 1913 г. (Павленко, 1914; Берг, 1915), в прол. Стрелок ловилась в 1912 г., в ставные невода попадалась с 1919 г., в 1922 г., поданным А. Г. Кагановского, в Хасанском районе был организован небольшой лов ее, а в 1923 г. в бочонке, полученном для исследования сельди, оказалась сардина (Линдберг, 1935). Отсутствие более частых и больших по размерам уловов объяснялось тем, что ко времени появления в наших водах сардины промысловый лов уже заканчивался и орудия лова снимались (Кагановский, 1939). Все эти факты свидетельствуют о том, что сардина несомнденно встречалась в наших водах и ранее 1925 г., когда начался промысловый лов ее, однако вряд ли в значительных количествах. Несомненно также, что возникновение промысла сардины в какой-то мере было связано с изменением температурного режима моря, так как в 1941 г., при менее благоприятных температурных условиях прибрежных вод Японского моря, чем в предыдущие годы, сардина пришла в наши воды в значительно меньшем количестве, и улов ее в этом и следующем году был ничтожным, а в 1948 и 1944 гг.. ловились лишь единичные экземпляры {Шмидт, 1945). Падению уловов сардины в наших водах сопутствовало снижение уловов ее в 1942 г. и у берегов Японии, у о. Хондо в 3 раза, севернее — у о. Хоккайдо во много раз большее (Кагановский, 1945), причем тенденция к снижению уловов у берегов Кореи и Японии обнаружилась еще в 1936—1937 гг. (Ясугава Macao, 1944; цит. по Шмидту, 1945). С изменением температурного режима моря так или иначе связывается падение уловов и согласно всем трем взглядам, высказанным в объяснение причин этого падения (Шмидт, 1945, Кагановский, 1945; Ясугава Macao, цит. по Шмидту, 1945 и Кагановскому, 1945). Следует также отметить, что увеличение численности сардины сопровождалось понижением уловов нерестовой сельди у берегов Приморья (Городничий, 1949) и у Хоккайдо и южной части Сахалина. Падение уловов отмечается и у калифорнийской сардины, но несколько позднее. Первые признаки сокращения уловов ее были отмечены после 1944 г., а к 1947 г. снизились в 5 раз.
Хозяйственное значение. Важнейшая промысловая рыба, стоявшая по уловам мирового промысла на первом месте. Общий улов рассматриваемого подвида в 1936—1939 гг. составлял 21.7—27.9 млн ц, если же принять во внимание уловы других подвидов, то общий улов вида в те же годы достигал 27.3—34.8 млн ц, из них 5.5— 7.3 млн ц добыто у берегов Сев. Америки (Sardinops sagax caerulea). В наших водах промысел возник в 1925 г., в 1931—1940 гг. уловы составляли от 0.959 до 1.397 млн ц. У берегов Японии, где возникновение промысла относится к 1854—1859 гг., с 1911—1912 гг. промысел начал быстро возрастать, в 1936—1939 гг. поймано от 10.9 до 16.3 млн ц, у берегов Кореи — от 9.8 до 13.9, у южной части Сахалина — от 0.5 до 4.89 млн ц. Лов происходил у восточных берегов Кореи, вдоль берегов Приморья и по обе стороны Японии. Ловят как в открытом море, так и у берегов.