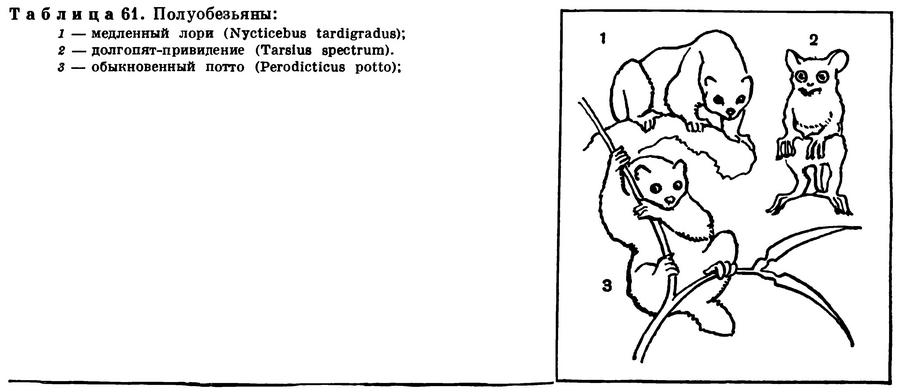
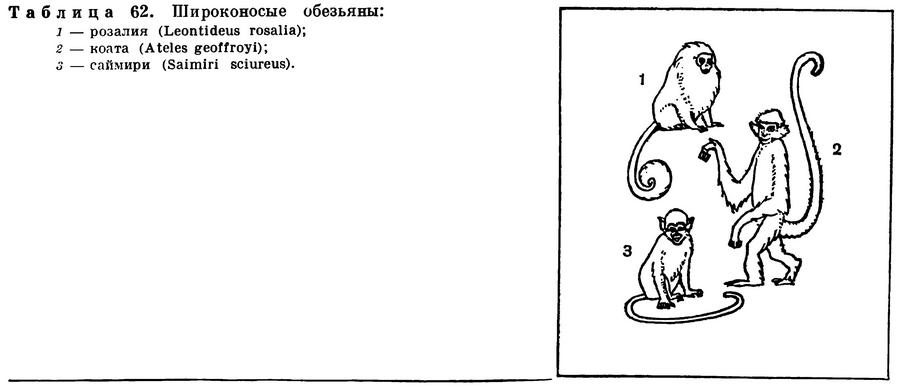
(Mammalia Linnaeus, 1758) Класс Маммалии, Млекопитающие, Звери, Class Mammalia Linnaeus, 1758 (Mammals) 29 отрядов
Жизнь животных. Том 6. Млекопитающие, или звери Под редакцией профессора Л.А.Зенкевича 1971 г.
ТИП ПОЗВОНОЧНЫЕ (VERTEBRATA)
КЛАСС МЛЕКОПИТАЮЩИЕ, ИЛИ ЗВЕРИ (MAMMALIA)
Краткая характеристика.
Млекопитающие — высший класс позвоночных животных, венчающий всю систему животного мира. Наиболее важными чертами общей организации этих животных служат: высокий уровень развития нервной системы, обеспечивающий сложные и совершенные формы приспособительного реагирования на воздействия внешней среды и слаженную систему взаимодействия различных органов тела; живородность, сочетающаяся (в отличие от других живородных позвоночных, например рыб и рептилий) с выкармливанием детенышей молоком. Она обеспечивает лучшую сохранность молодняка и возможность размножения в весьма разнообразной обстановке; совершенная система теплорегулирования, благодаря которой тело имеет относительно постоянную температуру, т. е. постоянство условий внутренней среды организма. Легко представить себе, какое огромное значение это имеет для распространения млекопитающих в разнообразных жизненных условиях.
Все это способствует почти повсеместному распространению млекопитающих по Земле, при котором они заселяют все жизненные среды: воздушно-наземную, водную и почвенно-грунтовую.
В строении млекопитающих можно отметить следующее. Тело их покрыто волосами, или шерстью (есть редкие исключения вторичного характера). Кожа богата железами, имеющими разнообразное и весьма важное функциональное значение. Особенно характерны млечные (молочные) железы, на которые нет даже намеков у других позвоночных.
Нижняя челюсть состоит лишь из одной (зубной) кости. В полости среднего уха имеются три (а не одна, как у амфибий, рептилий и птиц) слуховые косточки: молоточек, наковальня и стремечко. Зубы дифференцированы на резцы, клыки и коренные; сидят в альвеолах. Сердце, как и у птиц, четырехкамерное, с одной (левой) дугой аорты. Красные кровяные тельца без ядер, что повышает их кислородную емкость.
Ныне живущих млекопитающих насчитывают около 4500 видов.
Современная наука о млекопитающих предусматривает не только познание закономерностей их организации и жизнедеятельности, но и исследование биологических основ управления фауной.
Строение млекопитающих.
Внешний облик млекопитающих многообразен. Это объясняется удивительным разнообразием свойственной им жизненной обстановки — наземная поверхность, кроны деревьев, почва, вода, воздух. Очень сильно варьируют и размеры тела от 3,8 см при массе 1,5 г у карликовой белозубки до 30 м и даже более при массе около 150 т у синего кита, что соответствует массе 30 слонов или 150 быков.
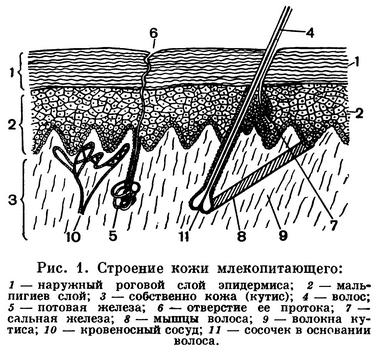
Кожа, как и у других позвоночных, состоит из двух слоев: наружного — эпидермиса и внутреннего — кутиса, или собственно кожи (рис. 1). Эпидермис в свою очередь подразделяется на два слоя: глубинный, ростковый (иначе мальпигиев), состоящий из живых, делящихся клеток, и наружный, представленный постепенно отмирающими, в связи с роговым перерождением, клетками. Наиболее поверхностно расположенные клетки полностью ороговевают и слущиваются в виде мелкой перхоти или целых лоскутов (у некоторых тюленей).
За счет деятельности эпидермиса возникают такие его производные, как волосы, ногти, когти, копыта, рога (кроме оленьих), роговые чешуи и кожные железы.
Собственно кожа сильно развита и состоит преимущественно из волокнистой ткани. Нижняя часть этого слоя рыхлая, и в ней откладывается жир — это так называемая подкожная жировая клетчатка. Собственно кожа особенно сильно развита у водных зверей — тюленей и китов, у которых она выполняет термоизолирующую роль и уменьшает плотность тела.
Общая толщина кожи у разных видов различна. Как правило, у наземных обитателей холодных стран, имеющих пышный волосяной покров, она меньше. Есть разница и в прочности кожи. Встречается своеобразная кожная хвостовая аутотомия у мышей, тушканчиков и особенно у сонь. Кожный хвостовой футляр у них легко обрывается и соскальзывает с хвостовых позвонков, что дает возможность схваченному за хвост зверьку уйти от врага. Такое же биологическое значение имеет и очень тонкая, непрочная и бедная кровеносными сосудами кожа зайцев.
Кожный покров млекопитающих имеет существенное терморегуляционное значение. Роль мехового покрова понятна, но нужно еще указать и на значение кожных кровеносных сосудов. При расширении их просветов, которые регулируются нервнорефлекторными механизмами, теплоотдача резко возрастает. У некоторых видов существенное значение имеет также испарение с поверхности кожи пота, выделяемого одноименными железами.
Кожные железы у млекопитающих, в отличие от рептилий и птиц, обильны и многообразны. Потовые железы — трубчатые, глубинные их части имеют вид клубка. Эти железы выделяют в основном воду, в которой растворены мочевина и соли. Не у всех видов млекопитающих потовые железы развиты одинаково. Их мало у собак, кошек; у многих грызунов они есть только на лапах, в паховой области и на губах. Вовсе нет потовых желез у китообразных, ящеров и некоторых других. Сальные железы гроздевидны, и протоки их открываются в волосяную сумку. Секрет этих желез смазывает поверхность эпидермиса и волосы, предохраняя их от снашивания и смачивания. Кроме того, выделения сальных и потовых желез придают животному и его следам специфический запах и этим облегчают общение между особями одного вида и между разными видами.
Пахучие железы представляют видоизменение сальных или потовых желез, а иногда их комбинации. Значение желез разнообразно. Их выделения помогают распознаванию особей разного пола, служат для мечения занятой территории, способствуют половому возбуждению, служат средством защиты от врагов. Таковы мускусные железы кабарги, выхухоли, землероек, ондатры, анальные железы хищных, копытные и зароговые железы коз, серн и других парнокопытных. Общеизвестны анальные железы скунса, секрет которых чрезвычайно едок и служит для защиты от врагов.
Млечные железы возникли как видоизменение потовых желез. У низших однопроходных млекопитающих они сохраняют простое трубчатое строение, и протоки их открываются на определенном участке кожи брюшной поверхности. Сосков в этом случае нет. У сумчатых и плацентарных млечные железы гроздевидны, и протоки их открываются на сосках. Расположение желез и сосков бывает различным. У летучих мышей и обезьян они располагаются на груди, и сосков бывает одна пара. У большинства копытных соски расположены, как и сами железы, в паховой области. У других зверей млечные железы и соски расположены на брюхе и груди. Число сосков в известной мере связано с плодовитостью вида. Максимальное число их 24 (опоссумы из сумчатых, тенреки из насекомоядных).
Волосяной покров — наиболее характерное эпидермическое образование млекопитающих. Отсутствие его у некоторых видов (например, у китообразных) — явление вторичное. Хотя волос — производное эпидермиса, при развитии его зачаток — волосяная луковица — глубоко внедряется в толщу собственно кожи, образуя волосяную сумку. В дно волосяной луковицы вдается сосочек кориума с кровеносными сосудами и нервами. Это так называемый волосяной сосочек. Формирование и рост волоса происходят за счет размножения и видоизменения клеток луковицы, а ствол волоса является уже мертвым образованием, неспособным к росту. В стволе волоса различают три слоя: верхнюю кожицу, корковый слой и сердцевину. Два верхних слоя состоят из плоских ороговевших клеток и, как правило, содержат красящий пигмент. Сердцевина состоит из высохших клеток, наполненных воздухом, что и сообщает шерсти малую теплопроводность.
Волосяной покров состоит из волос различного типа. Основными категориями их будут пуховые волосы, ость и чувствующие волосы, или вибриссы. У большинства видов преимущественное развитие получают пуховые волосы, формирующие подшерсток, или пух. Однако у некоторых животных, например оленей, кабанов, многих тюленей, подшерсток редуцирован, и волосяной покров состоит главным образом из остей. Наоборот, у подземных зверей (крота, слепыша, цокора и др.) остевых волос почти нет. У большинства видов волосы на поверхности кожи распределяются неравномерно, собраны в пучки, состоящие из остевого волоса, вокруг которого расположено несколько пуховых (от двух до двухсот).
Наблюдается периодическая смена волосяного покрова, или линька. У некоторых видов она бывает дважды в году — осенью и весной; таковы белки, некоторые зайцы, песцы, лисицы. У кротов бывает и третья, летняя линька. Суслики, сурки линяют один раз в году, в весенне-летнее время. У северных млекопитающих по сезонам меняется густота меха, а у некоторых и окраска. Так, у белки на 1 см2 на огузке летом бывает в среднем 4200 волос, а зимой — 8100. На зиму белеют горностаи, ласки, песцы, зайцы-беляки и некоторые другие.
Особую категорию волос представляют вибриссы — очень длинные жесткие волосы, выполняющие осязательную функцию. Они расположены на голове, нижней части шеи, груди, а у некоторых лазающих (например, у белки) — и на брюхе. Видоизменениями волос являются щетина и иглы.
Кроме волос, на коже млекопитающих бывают роговые чешуи. Наиболее сильно они развиты у ящеров. Роговые чешуйки наблюдаются также на лапах (мышевидные грызуны) и на хвосте (мышевидные, бобр, выхухоль, некоторые сумчатые).
Роговыми образованиями будут полые рога копытных, ногти, когти, копыта. Рога оленей также представляют придатки кожи, но развиваются они из кутиса и состоят из костного вещества.
Мышечная система млекопитающих весьма дифференцирована в связи с разнообразием телодвижений. Есть диафрагма — куполообразная мышца, отделяющая брюшную полость от грудной и имеющая важное значение при вентиляции легких. Хорошо развита подкожная мускулатура.
У ежей, ящеров и некоторых броненосцев она обеспечивает свертывание тела в клубок. Эта же мускулатура обусловливает ощетинивание зверей, поднятие игл у ежей и дикобразов, движение вибрисс. На лице — это мимическая мускулатура.
Череп характеризуется относительно крупными размерами мозговой коробки, что естественно связано с большим объемом головного мозга. Кости черепа срастаются поздно, и это обеспечивает увеличение головного мозга по мере роста животного. Характерно сращение ряда костей в комплексы. Так, четыре затылочные кости образуют одну; сращение ушных костей приводит к образованию единой каменистой кости; сложное происхождение имеют височная и основная кости. Специфической особенностью является строение нижней челюсти, состоящей только из зубной кости. Угловая кость дает начало характерной только для млекопитающих барабанной кости, расположенной в виде яйцевидного вздутия на дне мозговой части черепа. Сочленовная кость, которая у рептилий также входила в состав нижней челюсти, превращается в одну из слуховых косточек среднего уха — молоточек. Нижняя челюсть причленяется к черепу (к височной кости) непосредственно, так как квадратная кость, к которой причленяется нижняя челюсть у рептилий и птиц, также видоизменилась в слуховую косточку — наковальню.
В строении позвоночного столба характерны плоские сочленовные поверхности позвонков и четко выраженная расчлененность позвоночника на отделы: шейный, грудной, поясничный, крестцовый и хвостовой. Первые два шейных позвонка преобразованы в атлант и эпистрофей, а общее число шейных позвонков равно семи. Таким образом, длина шеи у млекопитающих, в отличие от птиц, определяется не числом позвонков, а их длиной. Исключение составляют только ленивцы и ламантины, у которых число шейных позвонков варьирует от шести до десяти.
Истинных крестцовых позвонков два, но обычно к ним прирастают еще два хвостовых позвонка.
Основой плечевого пояса служит лопатка, к которой прирастает рудиментарный коракоид, и только у однопроходных коракоид представлен самостоятельной костью. Ключица есть у видов, передние конечности которых совершают движения в разных плоскостях, например у обезьян. Виды, перемещающие эти конечности в одной плоскости, например копытные, ключиц не имеют.
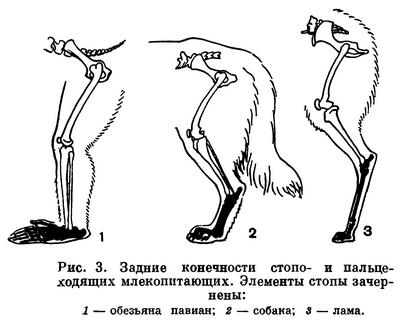
Конечности млекопитающих вполне типичны для наземных позвоночных, но число пальцев варьирует от пяти до одного. Сокращение числа пальцев или рудиментация крайних из них наблюдается у быстро бегающих видов, например копытных, тушканчиков. Сравнительно медленно перемещающиеся звери, например медведи, обезьяны, опираются при хождении на всю ладонь и ступню (стопоходящие виды); быстрые бегуны, например собаки, копытные, опираются только на пальцы (пальцеходящие виды) (рис. 3).
Пищеварительный тракт характеризуется большой длиной и хорошо выраженной расчлененностью на отделы. Он начинается преддверием рта, расположенным между мясистыми губами (свойственны только млекопитающим) и челюстями. У некоторых зверей расширение преддверия приводит к образованию больших защечных мешков (у хомяков, бурундуков, сусликов, некоторых обезьян). Мясистых губ нет у однопроходных и китообразных. В ротовую полость открываются протоки слюнных желез, секрет которых не только смачивает пищу, но и химически воздействует (фермент птиалин) на крахмал, превращая его в сахар. Слюна десмодов, питающихся кровью, обладает антикоагулирующим свойством, т. е. препятствует свертыванию крови. У некоторых насекомоядных слюна ядовита и используется для умерщвления добычи.
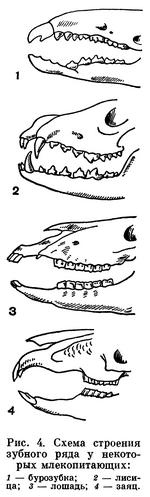
Зубы млекопитающих дифференцированы на группы, в зависимости от типа питания (рис. 4). Слабая дифференцировка свойственна малоспециализированным насекомоядным (бурозубкам). У зубатых китов дифференцировка зубов исчезла вторично.
Число зубов и распределение их по группам служат хорошим систематическим признаком. Для этого применяют зубную формулу, в которой группы зубов обозначают начальными буквами их латинских наименований: резцы — i (incisivi), клыки — c (canini), предкоренные — pm (praemolares) и истинно коренные — m (molares). Формулу пишут в виде дробей: в числителе — число зубов в верхней челюсти, в знаменателе — в нижней. Для сокращения указывают число зубов в одной половине челюсти.
Зубная формула волка такова:
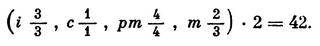
Желудок, обладающий многочисленными железами, имеет различный объем и внутреннее строение. Наиболее сложно устроен желудок жвачных копытных, поглощающих огромную массу малокалорийных и трудноперевариваемых кормов. У ящеров и муравьедов зубов нет, а желудок, как и у птиц, состоит из двух отделов: железистого и мускулистого. Сходство усиливается тем, что во втором обнаруживаются умышленно проглоченные камешки, обеспечивающие перетирание пищи.
Кроме тонкого, толстого и прямого отделов кишечника, некоторые млекопитающие имеют еще слепой отдел, в котором пища подвергается бактериальному сбраживанию. Особенно сильно слепая кишка развита у кормящихся грубой растительной пищей; длина ее достигает 1/3 длины кишечника. Длина толстого отдела кишечника по отношению к общей длине всего кишечника составляет в процентах: у грызунов — до 53, у насекомоядных — до 30, у хищных — до 22. Естественно, различна и общая длина кишечного тракта: у большинства летучих мышей он в 2,5 раза длиннее тела, у насекомоядных — в 2,5—4,2, у хищных — в 2,5—6,3, у грызунов — в 5,0—12,0, у копытных — в 12— 30 раз.
Протоки печени и поджелудочной железы впадают в переднюю часть тонких кишок.
Легкие имеют сложное ячеистое строение. Самые мелкие легочные ходы — бронхиолы заканчиваются пузырьками — альвеолами, в стенках которых ветвятся тончайшие кровеносные сосуды. Число альвеол даже у малоподвижных зверей (например, у ленивцев) равно 6 млн., а у весьма подвижных хищных оно достигает 300—500 млн. Механизм дыхания обусловливается изменением объема грудной клетки в результате движения межреберных мышц и диафрагмы.
Число дыхательных движений находится в зависимости от величины животного, определяющей разную интенсивность обмена веществ. Оно составляет (в 1 минуту): у лошади — 8—16, у черного медведя — 15—25, у лисицы —25—40, у крысы — 100—150, у мыши — около 200. Вентиляция легких не только обеспечивает газообмен, но имеет и терморегуляционное значение. При повышении температуры число дыханий увеличивается, а вместе с этим повышается и количество тепла, выводимого из организма. Так, у собаки отношение отдачи тепла при дыхании к общей его потере при температуре воздуха 8°С составляет (в процентах) 14, при 15°С — 22, при 30° — 46.
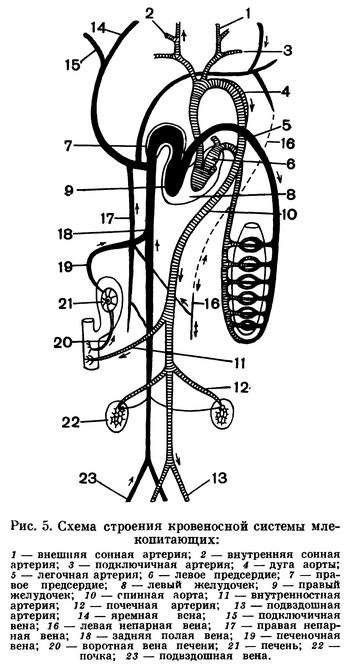
Кровеносная система млекопитающих сходна с таковой птиц (рис. 5). Сердце полностью разделено на два предсердия и два желудочка; от левого желудочка отходит одна дуга аорты (но не правая, как у птиц, а левая). Дуга аорты отсылает к голове сонные артерии и, огибая сердце, тянется под позвоночным столбом, по пути направляя разветвленные сосуды к системам органов. Венозная система характеризуется отсутствием воротного кровообращения в почках, которое в печени, как и у других позвоночных, хорошо развито. В печени происходит нейтрализация токсических продуктов белкового обмена.
Размеры сердца варьируют в зависимости от величины тела, образа жизни, а в конечном счете — в связи с интенсивностью обмена веществ. Масса сердца, выраженная в процентах к общей массе тела, составляет: у беззубых китов — 0,6—1,0, у дикого кролика — 3,0, у крота — 6,0—7,0, у летучих мышей — 9,0—15,0.
Подобная зависимость прослеживается и в отношении частоты сокращений сердца: у быка массой 500 000 г число сокращений сердца в 1 минуту составляет 40—45, у овцы массой 50 000 г — 70—80, у собаки массой 6500 г — 100—130, у мыши массой 25 г — 500—600.
Относительное количество крови и ее кислородная емкость у млекопитающих больше, чем у ниже стоящих классов. Это связано с количеством эритроцитов и их химической природой (см. табл.).
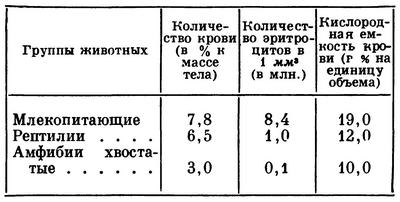
Плотность гемоглобина на 100 см3 крови у млекопитающих равна 10—15 г, у амфибий — только 5—10 г.
Все эти особенности обеспечивают млекопитающим более высокий уровень обмена веществ и общей жизнедеятельности.
Головной мозг очень велик; особенно велики полушария переднего мозга, которые покрывают сверху промежуточный и средний мозг. Головной мозг по массе в 3—15 раз больше спинного, в то время как у рептилий масса их примерно одинакова. Большое развитие получает серая кора полушарий, в которой располагаются центры высшей нервной деятельности. Это и обусловливает сложные формы приспособительного поведения млекопитающих. Кора передних полушарий несет многочисленные борозды, наибольшее число которых наблюдается у высших млекопитающих.
Мозжечок также сравнительно крупный и подразделен на несколько отделов.
Обонятельные органы характеризуются увеличенным объемом обонятельной капсулы и ее усложнением путем образования системы ответвлений — обонятельных раковин. Только у китообразных обонятельный аппарат редуцирован. Тюленям же свойственно достаточно острое обоняние.
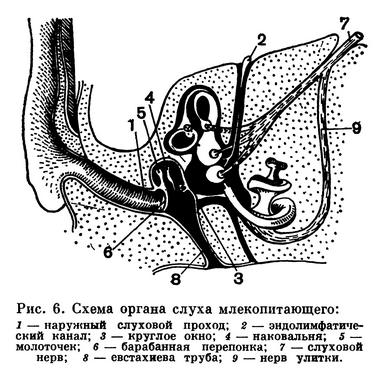
Органы слуха (рис. 6) у подавляющего большинства видов хорошо развиты. Они состоят из внутреннего и среднего уха, которые имеются и у предшествующих классов наземных позвоночных, а также из новоприобретений: наружного слухового прохода и ушной раковины, которая вторично исчезла у водных и обитающих в почве зверей. В полости среднего уха, которое отграничено от наружного слухового прохода барабанной перепонкой, находится не одна слуховая косточка — стремя, как у амфибий, рептилий и птиц, а еще две — молоточек (гомолог сочленовной кости нижней челюсти) и наковальня (гомолог квадратной кости). Молоточек упирается в барабанную перепонку, к нему причленена наковальня, которая в свою очередь сочленена со стремечком, упирающимся в окно перепончатого лабиринта (внутреннего уха). Вся эта система усиливает тонкость восприятия звуков. Кроме того, совершенство восприятия звуков обеспечивается и сильно развитой улиткой — извитым выростом перепончатого лабиринта, в котором расположено несколько тысяч тончайших волокон, резонирующих при восприятии звука.
Установлено, что у рукокрылых, мышевидных грызунов, землероек, китообразных выработался своеобразный механизм ориентировки путем звуковой локации: улавливание при помощи очень тонкого слуха отраженных предметами звуков высокой частоты, издаваемых голосовым аппаратом. Меняя частоту ультразвуков и улавливая их в отраженном виде, звери эти способны не только опознавать наличие предмета, но и чувствовать расстояние до него, а быть может, и его форму и другие качества. Такая эхолокация существенно помогает ориентировке в темноте или в воде.
Органы зрения не имеют принципиальных особенностей, и их значение в жизни зверей меньше, чем у птиц. На неподвижные предметы млекопитающие обращают мало внимания. Слабо развито или отсутствует у них цветное зрение. Так, кошки различают только 6 цветов, лошади — 4 цвета, крысы не различают зелено-желтый и сине-зеленый цвета. Только у высших приматов цветное зрение приближается к зрению человека.
Глаза водных зверей несколько похожи на глаза рыб: роговица уплощена, а хрусталик круглый, что свидетельствует о близорукости. У животных, ведущих подземный образ жизни, глаза рудиментарны, а у некоторых зверей (например, у слепого крота) они затянуты кожистой перепонкой.
Аккомодация по сравнению с птицами развита слабо и достигается только изменением формы хрусталика.
Характерной особенностью органов осязания является наличие осязательных волос, или вибрисс (рис. 7).

Почки у большинства млекопитающих бобовидной формы, с гладкой поверхностью. Только у китообразных, ластоногих и немногих других они состоят из нескольких долек. Основная в функциональном отношении часть почки — это ее наружный корковый слой, в котором располагаются извитые канальцы, начинающиеся боуменовыми капсулами, внутри которых находятся клубки кровеносных сосудов. В этих капсулах профильтровывается плазма крови, но не форменные ее элементы и белки. В почечных канальцах из фильтрата (первичной мочи) происходит обратное всасывание сахаров и аминокислот. Почечные канальцы впадают в почечную лоханку, от которой берет начало мочеточник. Число почечных канальцев велико: у мыши — 10 000, у кролика — около 300 000.
Половые органы самца состоят из семенников, семяпровода, придаточных желез и совокупительного члена. Семенники у большинства располагаются в мошонке, сообщающейся с полостью тела через паховый канал. У однопроходных, китообразных, слонов и некоторых других семенники находятся непосредственно в полости тела. К семеннику прилегает его придаток, от которого отходит семяпровод, впадающий у корня полового члена в мочеиспускательный (семяизвергательный) канал. Перед впадением в мочеиспускательный канал семяпроводы образуют парные семенные пузыри — железы, секрет которых принимает участие в образовании жидкой части спермы и благодаря клейкой консистенции препятствует обратному вытеканию спермы из половых путей самки (рис. 8). Рядом с семенными пузырями располагается вторая парная придаточная железа — предстательная, протоки которой впадают в начальную часть семяизвергательного канала. Секрет этой железы представляет основу жидкости спермы, в которой плавают сперматозоиды.
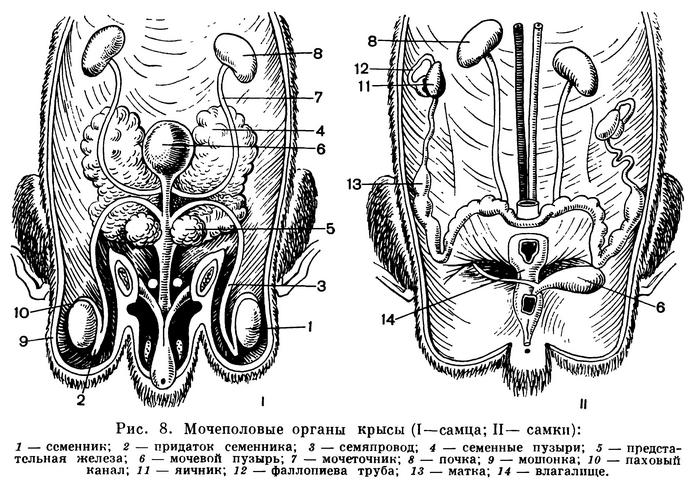
Совокупительный орган в основном состоит из пещеристых тел, которые заполняются перед совокуплением массой крови, что придает половому члену необходимую упругость. У ряда видов (например, у хищных, ластоногих) в толще совокупительного органа находится особая кость.
Парные яичники всегда лежат в полости тела. В непосредственной близости от них располагаются воронки парных яйцеводов, куда и попадают яйца после их выхода из яичника. Верхние извитые отделы яйцеводов представляют фаллопиевы трубы, в которых происходит оплодотворение яйца. Далее идут расширенные отделы матки, впадающие в непарное влагалище (у некоторых Сумчатых влагалище парное). Матка в простейшем случае парная, и во влагалище открываются два ее отверстия. У многих нижние отделы матки объединены, такая матка называется двураздельной (ряд грызунов, некоторые хищники). Слияние большей части маток приводит к образованию двурогой матки (некоторые хищники, китообразные, копытные). При полной утрате парности матка называется простой (некоторые рукокрылые, приматы).
Размер яиц у млекопитающих в большинстве случаев очень мал (0,05—0,4 мм) и только у однопроходных они в связи с наличием значительного количества желтка сравнительно велики (2,5—4,0 мм).
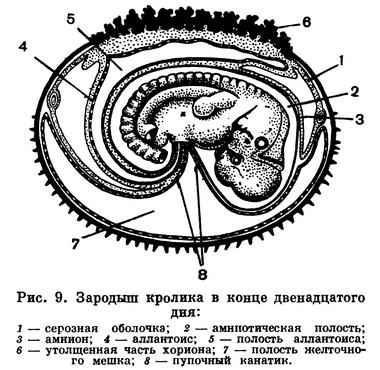
При развитии зародыша в матке у подавляющего большинства млекопитающих формируется плацента. Ее нет у однопроходных, а у сумчатых она зачаточна. Плацента возникает путем срастания двух наружных околоплодных оболочек (аллантоиса и прохориона), в результате чего формируется губчатое образование — хорион. Хорион образует выросты — ворсинки, которые внедряются или срастаются с разрыхленным эпителием матки. В этом месте происходит сплетение (но не сращение) кровеносных сосудов материнского организма и плода, что обеспечивает снабжение зародыша кислородом и питательными веществами и вынос продуктов обмена и углекислоты (рис. 9).
По способу распределения ворсинок хориона различают плаценты: диффузную — когда ворсинки распределяются равномерно (китообразные, некоторые копытные); дольчатую — когда ворсинки собраны в группы (жвачные); дискоидальную — когда ворсинки располагаются на участке хориона, имеющего вид обруча (насекомоядные, хищнее).
После родов в стенке матки на месте бывшего прикрепления плодов остаются темные, так называемые плацентарные пятна. По их количеству можно установить число детенышей в помете.
Половой диморфизм у млекопитающих выражен слабо (преимущественно в размерах и общем складе): самцы обычно несколько крупнее самок и более крепкого телосложения. Но у усатых китов, наоборот, самки заметно крупнее самцов. Многие парнокопытные отличаются по рогам. Самки всех оленей, кроме северного, безроги.
У самцов баранов рога более мощные, чем у самок, которые иногда бывают безрогими. Не имеют рогов самки многих антилоп. У самцов ушатых тюленей развивается подобие гривы.
Половые особенности в окраске свойственны меньшинству видов. Видимо, это связано с меньшим развитием у млекопитающих способности к зрительным (в частности, к цветовым) восприятиям. Разнополые особи различают друг друга главным образом по запаху.
Биология млекопитающах.
Млекопитающие распространены практически по всей Земле; их нет только на Антарктическом континенте, хотя у его побережья известны тюлени и киты. В районе Северного полюса бывают белые медведи, ластоногие, китообразные (нарвалы).
Широко распространены млекопитающие и по жизненным средам. Достаточно вспомнить, что наряду с наземными видами, которых большинство, ряд видов в той или иной мере связан с водной средой; многие активно летают по воздуху; кроме того, значительное число зверей обитает в почве, где проходит вся или большая часть их жизни. Ни один класс позвоночных не дал такого разнообразия форм, как млекопитающие.
Степень и характер взаимосвязей с жизненными средами различны. Некоторые животные обитают в весьма разнообразной обстановке. Например, обыкновенная лисица распространена в лесах, степях, пустынях и в горных районах. Волки обитают в еще более разнообразных условиях (кроме указанных зон для лисицы, волки встречаются и в тундре). Естественно, что у таких зверей нет ясно выраженных приспособлений к жизни в какой-либо определенной среде.
Наряду с этим специфическая среда обитания определяет многие особенности зверей. Среди обитателей тропических лесов выделяются, например, южноамериканские ленивцы, паукообразные обезьяны, некоторые южноазиатские медведи, вся или почти вся жизнь которых проходит в кронах деревьев. Здесь они кормятся, отдыхают и размножаются. В связи с таким образом жизни у видов этой группы развились разнообразные приспособления. Медведи и куницы лазают с помощью острых когтей, лемуры и обезьяны имеют хватательные лапы. У некоторых южноамериканских обезьян, древесных муравьедов и опоссумов развит цепкий хвост. Наконец, обезьяны гиббоны совершают прыжки, предварительно маятникообразно раскачиваясь на передних конечностях. Своеобразное приспособление к пассивному планирующему полету развито у летяг и южноазиатских шерстокрылов.
Среди обитателей лесов есть звери, ведущие строго наземный образ жизни (лось, настоящие олени, косуля, африканская окапи). Лес служит им укрытием, ветки и листья деревьев — отчасти кормом.
Животные, обитающие в открытых пространствах, имеют различные формы и степень специализации. У копытных и некоторых хищников сравнительно острое зрение, их быстрый бег помогает при больших переходах в поисках корма и воды, при преследовании. Скорость бега у некоторых животных такова (в км/ч): бизон — 40—45, жирафа — 45—50, дикий осел — 50—55, зебра — 60—65, газель Томсона — 75—80, а из хищных: лев — 75—80, гепард — 105—115. Жилищ или временных убежищ копытные не устраивают и рождают на поверхности земли развитых детенышей, способных уже в первые дни передвигаться вместе с матерью.
Весьма разнообразны виды прыгающих зверей, которые есть в различных систематических группах и в разных странах. Таковы пустынные азиатские тушканчики (грызуны), американские кенгуровые крысы (грызуны), африканские прыгунчики (насекомоядные), австралийские кенгуру (сумчатые). Характерная особенность этих зверей — «рикошетирующий» бег: прыжки вперед и из стороны в сторону, как бы отскакивание вследствие неровностей почвы. Длинный хвост играет роль балансира и руля. Скорость бега они развивают до 20 км/ч. От видов предшествующей группы они существенно отличаются тем, что устраивают, кроме кенгуру, постоянные убежища в виде нор. Новорожденные у всех слепые и голые.
Есть еще один тип зверей открытых пространств, условно обозначенный как «тип суслика» (рис. 10). Это мелкие либо средней величины звери, населяющие степи и горные луга с относительно густым травостоем, поэтому у них нет необходимости, а вместе с тем и возможности быстро перемещаться в поисках корма. Они сооружают постоянные норы, в которых размножаются и отдыхают; кроме того, сооружают временные норы, в которых лишь прячутся при опасности. Тело у этих зверей вальковатое, на коротких ногах, бегают они медленно. В связи с наличием нор они рождают беспомощных, голых детенышей. Кроме сусликов, к этой группе относятся сурки, хомяки, степные виды пищух.
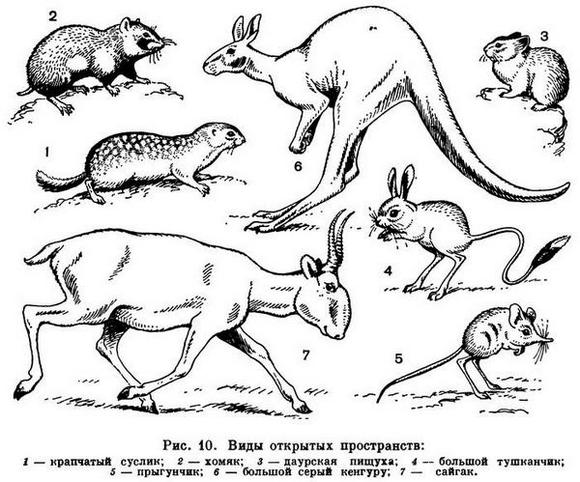
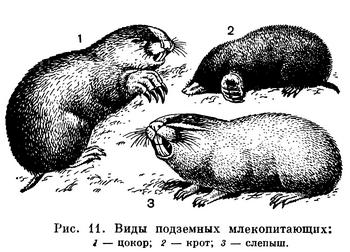
Подземные, звери — высокоспециализированная, но сравнительно немногочисленная группа. Представители ее есть среди разных отрядов и во многих странах. Таковы евразийские и североамериканские кроты (насекомоядные), евразийские слепыши и слепушонки (грызуны), африканские землекопы (грызуны), австралийские сумчатые кроты (рис. 11). Тело у них брускообразное, без ясно выраженной шеи. Мех низкий, бархатистый. Глаза небольшие, у некоторых они скрыты под кожей. Ушных раковин нет. Хвост рудиментарный. Способ прокладывания подземных ходов у разных видов различен. Кроты роют большими, вывернутыми наружу передними лапами, вращаясь вокруг оси тела. У слепышей и слепушонок лапы слабые, роют они резцами нижней челюсти, а лапами лишь выбрасывают землю. Система нор у этих животных очень сложная, но только небольшая часть их служит собственно жилищем, а остальная — ходами, образовавшимися при отыскивании пищи.
У водных зверей разная степень приспособления к жизни в воде. Один из видов хорьков — норка селится всегда по берегам водоемов, где устраивает норы, но корм добывает как в воде (рыбу, амфибий), так и на суше (мелких грызунов). Существенных изменений, сравнительно с наземными хорьками, в ее организации нет. Более тесно связана с водой речная выдра, добывающая корм главным образом в воде (рыбу, амфибий); наземные грызуны в ее рационе играют малую роль. Плавает и ныряет она хорошо, и при этом можно отметить такие приспособительные черты, как редукция ушных раковин, укорочение конечностей, наличие перепонок между пальцами.
Морская выдра, или калан, неправильно называемая морским бобром, обитает в северной части Тихого океана, корм добывает только в море (главным образом, морских ежей). На берег звери выходят для размножения, иногда для отдыха и при сильных штормах. Конечности у калана типа ластов, и все пальцы объединены толстой кожистой перепонкой.
Хорошо известны полуводные грызуны: бобр, ондатра, нутрия.
Ластоногие — настоящие водные звери, но размножаются только вне водной среды — на льдах или на берегу.
Полностью водные звери — китообразные и сиреновые, способные к размножению в воде. Характеристика их дана в соответствующих местах тома. Здесь же важно подчеркнуть, что среди современных зверей можно найти весь ряд переходов от наземных, через околоводных и полуводных до полностью водных форм.
Наконец, надо упомянуть о видах млекопитающих, перемещающихся по воздуху. И в этом случае можно выявить ряд зверей от перепрыгивающих с дерева на дерево до способных к активному полету. У белок в какой-то мере роль парашюта, определяющего начальные элементы планирования, играет распластанное при прыжке тело с вытянутыми конечностями и расчесанное надвое опушение хвоста. Более специализированы летяги и шерстокрылы, обладающие кожистыми складками между передними и задними лапами; у шерстокрыла складки расположены и между задними лапами и хвостом. Такое приспособление не только позволяет планировать при спуске по отлогой линии, но и служит для поворотов в воздухе и даже для незначительного поднятия. Летяги могут планировать на расстояние 30—60 м, а шерстокрылы — на 130—140 м.
К активному полету (с помощью крыльев) из млекопитающих способны только рукокрылые. Они подробно охарактеризованы в соответствующей части тома. Здесь же важно подчеркнуть удивительное разнообразие приспособлений млекопитающих к жизни в различных средах и к различным формам поведения. Все это, несомненно, расширило возможность их адаптивного расхождения, приведшего к поразительному многообразию форм зверей.
Размножение млекопитающих, характеризующееся большим разнообразием, имеет все же общие черты: внутреннее оплодотворение, живорождение (за редким и неполным исключением), выкармливание новорожденных молоком, а также устройство большинством видов специальных гнезд для деторождения.
В простейшем случае, у однопроходных, настоящего живорождения нет и самки откладывают яйца. Однако яйца они откладывают после довольно длительного развития в половых путях самки, и длительность инкубации яиц после их откладывания равна менее половины того срока, который проходит от времени их оплодотворения до откладывания. Таким образом, у однопроходных бывают «преждевременные» роды или незавершенное яйцеживорождение.
У сумчатых период беременности короткий и без образования настоящей плаценты. Мелкие виды сумчатых (например, опоссумы) носят детенышей всего 8—13 дней, и новорожденные имеют массу всего 0,003% массы матери. Для сравнения укажем, что у зайцев масса новорожденного составляет 3 % массы матери. Недоразвитые детеныши сумчатых завершают развитие, будучи прикрепленными к соскам, чаще открывающимся в полость кожистой сумки.
Длительность беременности, а в этой связи и развитость новорожденных у разных видов плацентарных зверей существенно отличны. Минимальная длительность беременности характерна для серого хомячка (11—13 дней), максимальная — для индийского слона (более 500 дней). Продолжительность внутриутробного развития зависит от совокупного действия трех обстоятельств.
Во-первых, это связано с величиной животного. Так, крошечная рыжая полевка (длина ее тела около 10 см) носит детенышей всего 18 дней, более крупная обыкновенная серая полевка (длина тела до 13 см) — 20 дней, еще более крупная крыса пасюк — 22 дня, хомяк (длина тела до 35 см) — 30 дней, сурок байбак (длина тела до 59 см) — 40 дней, лисица (длина тела до 90 см) — 52 дня.
Во-вторых, имеют значение условия деторождения, точнее, условия, в которых оказываются новорожденные. Все указанные выше млекопитающие рождают в надежно устроенных норах, и новорожденные у них голые, слепые, беспомощные.
Дикие кролики также котятся в норах, беременность у них всего 30 дней, и детеныши такие же слепые и голые. Сходной величины зайцы (мелкие расы беляка и песчаники), не устраивающие нор или гнезд, носят около 50 дней, и рождают они зайчат зрячих, покрытых шерстью и способных в первые же дни бегать. Близкая по размерам к сурку нутрия (длина тела до 60 см) приносит детенышей в очень примитивно устроенной норе или в наземном гнезде. Беременность у нее в три раза продолжительнее, чем у сурка (около 130 дней), и новорожденные развиты так же хорошо, как и у зайцев.
Велика длительность беременности у копытных, не имеющих каких-либо убежищ. У горных баранов она достигает 150 дней, у оленей и лосей — 240—250 дней, у зебры — 340—365 дней. Новорожденные у этих видов хорошо развиты и в первые дни после рождения уже могут следовать за матерями.
В-третьих, продолжительность беременности зависит от длительности промежутка между оплодотворением яйца и прикреплением зародыша к стенке матки, т. е. образованием плаценты. Есть виды, у которых этот промежуток очень велик, и он в основном влияет на срок беременности. Так, у соболя и куницы начавшееся развитие яйца прерывается фазой покоя, когда развитие почти полностью прекращается (латентная фаза беременности). Период этот длится несколько месяцев, и в итоге у этих сравнительно мелких зверьков (длина тела около 50 см), рождающих в хорошо защищенных гнездах, беременность продолжается в течение 230—280 дней. При этом детеныши рождаются слепыми и беспомощными.
Латентная фаза беременности свойственна и другим куньим (горностаю), а также росомахе, барсуку, тюленям, косуле и др. При этом бывает и так, что сложная беременность наблюдается только у части особей данного вида. Например, у барсуков, спаривающихся осенью, беременность длится 150 дней, а у спаривающихся летом — 220—240 дней. У горностаев беременность обычно длится 300—320 дней и большая часть этого срока падает на латентную фазу. Но бывают случаи, когда латентная фаза почти или полностью выпадает и период внутриутробного развития длится лишь 42—70 дней.
В связи с условиями деторождения, различной длительностью беременности и, главным образом, с наличием или отсутствием латентной фазы сроки спаривания не имеют такого же единообразия, как, например, у птиц. Волки и лисицы спариваются в конце зимы, хорьки, зайцы — весной, соболи, куницы — летом, олени — осенью. Но замечательно, что роды у подавляющего большинства приурочены к наиболее благоприятному для воспитания молодняка времени — концу весны и началу лета.
Скорость размножения разных видов существенно различается. Это различие определяется тремя обстоятельствами: временем полового созревания, длительностью промежутка между рождениями, числом детенышей в выводке.
Вариация скорости полового созревания очень велика, что видно из следующих примеров. Рыжие полевки достигают половой зрелости в возрасте около одного месяца, ондатра — не ранее трех месяцев, зайцы, лисицы — около года, соболи, волки — около двух лет, бурые медведи — около 3—4 лет, моржи — в 3—6 лет, зубры — 5—6 лет, слоны — 10—15 лет, носороги — около 20 лет. Таким образом, этот показатель варьирует в пределах нескольких сотен раз.
Сильно варьирует и частота повторяемости рождений. Мелкие грызуны, например домовая мышь и обыкновенная полевка, при благоприятных температурных и кормовых условиях могут приносить в течение года до 6—8 пометов. Зайцы-русаки на юге плодятся круглый год и приносят до 4 пометов, зайцы-беляки — до 3 пометов. Только один раз в году размножаются хорьки, куницы, соболи, лисицы, волки, олени и многие другие. Бурые медведи и верблюды рождают, как правило, один раз в два года. Один раз в два-три года обычно приносят приплод моржи, белые медведи, тигры, некоторые усатые киты, зубры. Этот показатель скорости размножения варьирует в десятки раз.
Существенно различна и величина выводка, варьирующая в пределах от 1 до 20 детенышей, чаще до 12—15. Наименее плодовиты виды, которым свойственны постоянные и подчас длительные перемещения и которые не устраивают даже примитивных временных жилищ. Таковы слоны, лошади, зебры, ослы, ламы, верблюды, жирафы, олени, тюлени, киты, сирены.
Наибольшая величина выводка свойственна мелким норным грызунам: полевкам, мышам, некоторым сусликам. У этих видов нередко бывает 10 или несколько более детенышей. В отдельных случаях отмечали даже до 20 эмбрионов. Значительна плодовитость некоторых зайцев (до 12 детенышей), кабанов (до 12) и норных хищников мелкой и средней величины (лисица — до 12, песец — до 18, горностай — до 18). Крупная величина выводка у этих видов обеспечивается надежностью жилищ, в которых укрыты новорожденные, а также обилием и доступностью пищи (хотя бы в некоторые годы).
Промежуточное положение занимают крупные хищники и некоторые крупные грызуны. По 2—3 детеныша приносят львы, пумы, тигры, рыси, бурые медведи.
Добывать корм для молодняка этим животным труднее, чем предыдущим; большинство их не строит надежных жилищ при выводе детенышей.
Если обобщить все сказанное о скорости размножения, то можно видеть, что, как правило, виды, медленно созревающие, имеют большие промежутки между рождениями и наименьшее число детенышей. Наоборот, виды млекопитающих, у которых половое созревание наступает рано, плодятся часто и выводок у них большой.
Замечено, что скорость размножения находится в довольно тесной зависимости от продолжительности жизни: медленно размножающиеся живут значительно дольше высокоплодовитых. Примерная продолжительность жизни в природной обстановке такова: слон индийский — 70—80 лет, зубр — до 50 лет, олень марал — до 25 лет, киты усатые — около 20 лет, медведь бурый — 30—50 лет, волк — около 15 лет, лисица — 10—12 лет, соболь — 7—8 лет, зайцы — 5—7 лет, полевки — 14—18 месяцев. В итоге скорость обновления населения у разных видов очень различна.
Характер брачных отношений разнообразен. Большинство видов — полигамы: самцы у них оплодотворяют по нескольку самок. Таковы насекомоядные, большинство грызунов, многие хищные, копытные, китообразные. Самцы большинства полигамных видов в период спаривания не удерживают около себя самок, и в видовом строе населения не возникает в это время принципиальных особенностей (грызуны, насекомоядные, полигамные хищники). Самцы становятся лишь более подвижными , разыскивая неоплодотворенных самок.
Иначе протекает брачный период у ушатых тюленей и многих копытных. Котики образуют гаремы — скопления самок (иногда до нескольких десятков) около одного самца-производителя. Гарем существует более месяца, и самцы ревниво оберегают своих подруг. Самцы диких лошадей образуют косяки, самцы оленей — табуны.
Моногамия имеет разную выраженность. Самец настоящих тюленей кроет только одну самку, но пара образуется лишь на время совокупления. Шакалы, лисицы, песцы, ондатры образуют пары на сезон размножения, и самцы у этих видов принимают участие в охране гнездового участка, а у некоторых (например, у песцов) и в воспитании молодых. Сильные самцы волков удерживают около себя самку в течение нескольких лет. Только немногие виды обезьян иногда образуют пары на всю жизнь.
Время существования семьи и другие моменты размножения весьма различны у разных млекопитающих. Они определяются в основном частотой деторождения, обилием корма и в этой связи возможностью перехода молодых к самостоятельному образу жизни, степенью трудности самостоятельного перезимовывания. Очень короткий семейный период у многоплодных мелких мышевидных грызунов; например, у полевок он длится около 2—3 недель. Кормов для них обычно достаточно, и добыча их не требует особых навыков. Рано, в возрасте около одного месяца, переходят на самостоятельное существование суслики. Примерно в два раза дольше длится семейный период у белок, условия жизни которых более сложные. Еще дольше существует семья у хищников. Самостоятельная добыча корма в этом случае требует больших навыков. Так, у хорька семья распадается при возрасте молодых в 3 месяца, у лисицы — в 3—4 месяца, у волка — в 10—11 месяцев. Семьями зимуют и сурки. Почти 2 года находятся при матери бурые медведи, а тигрята — 2—3 года.
Распад семей и расселение молодняка есть несомненное приспособление, предотвращающее истощение кормовых средств близ гнезда. Так, у малого суслика в богатых травостоем степях Ростовской области молодые расселяются в возрасте 45—50 дней, а в полупустынных степях Заволжья — в 20—25 дней.
Бывает, наконец, и временный распад семьи, связанный с гоном. У некоторых оленей теленок ходит с матерью до осени, и при начале гона самка покидает его, но нередко после гона самка находит теленка и держится с ним почти до отела.
Приспособления млекопитающих к переживанию неблагоприятных в кормовом и погодном отношении периодов года более разнообразны и совершенны, чем у ниже стоящих классов. К зиме или засушливому лету в организме накапливаются резервные энергетические вещества, помогающие пережить тяжелый сезон. Помимо накопления гликогена в печени, многие виды в той или иной мере жиреют. Например, малый суслик весной имеет массу около 100—150 г, а в середине лета — до 400 г. У сурка-байбака подкожный и внутренний жир в июне составляет 10—15 г, в июле — 250—300 г, в августе — 750—800 г. У некоторых особей до 25% общей массы тела составляет жир.
Сезонные приспособления выражаются и в миграциях. Осенью, по мере ухудшения кормовых условий, основная масса песцов и северных оленей откочевывает из тундры на юг, в лесотундру и даже в тайгу, где легче добыть корм из-под снега. Вслед за оленями мигрируют к югу и тундровые волки. В северных районах тундры зайцы-беляки в начале зимы предпринимают массовые кочевки на юг, весной — в обратном направлении. Горные копытные к лету поднимаются в верхние пояса гор с их богатым травостоем, зимой же, по мере увеличения глубины снегового покрова, они спускаются вниз. И в этом случае наблюдаются совмещенные с копытными миграции некоторых хищников, например волков.
Закономерные сезонные миграции пустынных копытных тоже зависят от сезонных изменений растительного покрова, а местами — и от характера снегового покрова. В Казахстане сайгаки летом держатся чаще в северных глинистых полупустынных степях; зимой они откочевывают к югу, в область менее снежных полынно-типчаковых и полынно-солянковых полупустынь.
Сезонные кочевки наблюдаются у летучих мышей, китообразных и ластоногих.
В целом у млекопитающих миграции свойственны относительно меньшему числу видов, чем у птиц и у рыб. В наибольшей мере они развиты у морских зверей, рукокрылых и копытных, в то время как среди видов наиболее многочисленных групп — грызунов, насекомоядных и мелких хищников — их практически нет.
Третье сезонное приспособление — это спячка, свойственная многим зверям из отрядов: однопроходных, сумчатых, насекомоядных, рукокрылых, неполнозубых, хищных, грызунов. Нет видов, впадающих в спячку, среди тех отрядов, которым в наибольшей мере свойственны сезонные миграции: китообразные, ластоногие, копытные. Исключение составляют некоторые летучие мыши, у которых спячки чередуются с миграциями.
По степени глубины спячки различают два основных варианта.
1. Сезонный сон, или факультативная спячка. В этом случае температура тела, число дыхательных движений и общий уровень обменных явлений снижаются мало. При изменении обстановки или при беспокойстве сон легко может быть прерван. Характерен он для медведей, енотов, енотовидных собак, отчасти для барсуков. У белого медведя в берлогу залегают только беременные самки и неполовозрелые. Бурые медведи и барсуки в южных частях ареала в спячку не ложатся. О состоянии спящего зимой американского черного медведя представление дают такие данные. При температуре воздуха — 8°С на поверхности кожи отмечена температура +4°, в прямой кишке +22°, в ротовой полости +35° (против +38° в период бодрствования). Число дыханий сократилось до 2—3 в минуту (против 8—14 при бодрствовании).
Сроки залегания в зимний сон и его длительность изменчивы не только географически, но и по годам. Известны случаи, когда при оттепелях, особенно в малокормные годы, еноты, енотовидные собаки и бурые медведи выходят из убежищ и ведут активный образ жизни.
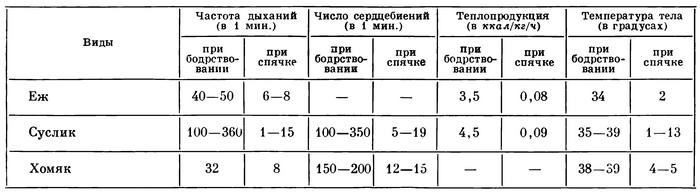
2. Настоящая непрерывная сезонная спячка. Она характеризуется потерей способности к терморегуляции (состояние гетеротермии), резким сокращением числа дыхательных движений и сокращений сердечной мышцы, падением общего уровня обмена веществ. Следующая таблица дает представление о сдвигах в состоянии зверей, находящихся в зимней спячке (в условиях эксперимента):
Как видно, полного прекращения обмена веществ во время спячки не бывает, и животные существуют за счет расходования энергетических запасов своего тела. В первую очередь расходуются жир и гликоген печени, а также вещества и некоторых других органов. В 1857 г. Валентин установил следующий расход тканей у сурка, находящегося в спячке:

Не у всех особей и не всегда расход бывает таким большим. В ряде случаев наблюдали сурков, вышедших из нор после спячки с вполне заметным отложением жира.
Наряду с зимней спячкой бывает и летняя спячка, вызванная также сезонным ухудшением кормовой базы. Чаще всего она наблюдается у грызунов, лишенных летом полноценного и богатого водой корма. Таковы в основном суслики. Наиболее рано впадает в спячку желтый, или песчаный, суслик Средней Азии (в июне — июле). У сусликов летняя спячка обычно без перерыва переходит в зимнюю.
Летняя спячка наблюдается и у обитателей тропической зоны. Сенегальские ежи впадают летом в спячку, длящуюся три месяца, а примитивные мадагаскарские насекомоядные — тенреки спят летом в течение четырех месяцев.
Четвертое (новое, сравнительно с предшествующими классами) основное приспособление , обеспечивающее переживание неблагоприятных сезонных условий жизни, — собирание запасов корма. Оно в разной мере свойственно разным систематическим группам млекопитающих. Не запасают корм классические кочевники — номады: китообразные, ластоногие, летучие мыши, копытные и млекопитающие, впадающие в спячку. В зачаточной форме это наблюдается у насекомоядных. Некоторые землеройки, например североамериканские короткохвостые землеройки (Blarina), делают лишь небольшие запасы из беспозвоночных. Наши выхухоли складывают в норах некоторое количество двустворчатых моллюсков. Местами кроты собирают запасы земляных червей. Некоторое время черви остаются живыми, так как укусы кротов в головном отделе червей лишают их способности двигаться. В ходах обнаруживали запасы по 100—300, а в отдельных случаях и до 1000 червей.
Более распространено захоронение излишней добычи у хищных. Ласки и горностаи собирают по 20—30 полевок и мышей, черные хори складывают подо льдом по нескольку десятков лягушек, норки — по нескольку килограммов рыбы. Более крупные хищники (куницы, росомахи, кошки, медведи) прячут остатки добычи в укромных местах, под упавшими деревьями, под камнями. Леопарды нередко укрывают часть добычи в ветвях деревьев.
Характерной особенностью запасания корма хищниками служит то, что для его захоронения не сооружается специальных кладовых, запасом пользуется только одна особь, соорудившая его. В целом запасы служат лишь малым подспорьем для переживания малокормного периода, и они не могут предотвратить вдруг наступившую бескормицу.
По-иному запасают корм различные грызуны и пищухи, хотя и в этом случае наблюдается разная степень совершенства запасания и его значения. Летяги собирают по нескольку десятков граммов концевых веточек и сережек ольхи и березы, которые они складывают в дупла. Белки захоранивают в опавших листьях, в дуплах и в земле желуди и орехи. Они развешивают также грибы на ветках деревьев.
Одна белка в темно-хвойной тайге запасает до 150—300 грибов, а в ленточных борах Западной Сибири, где кормовые условия хуже, чем в тайге, до 1500—2000 грибов, преимущественно маслят. Сделанные белкой запасы используются многими особями этого вида.
В Восточной Сибири и на Камчатке большие запасы корневищ, клубней и семян (до 15 кг) делает маленький зверек полевка-экономка.
Характерно, что в западных частях ареала этого вида, где зима менее продолжительная и мягкая, эта полевка запасов не делает. То же наблюдается и у водяной полевки, делающей большие запасы на востоке ареала (в Якутии) и почти не делающей их в западных районах распространения. Большие запасы на зиму корма в виде травы и луковиц лука делает обитатель континентальных степей Забайкалья и Северной Монголии — полевка Брандта.
Большая песчанка и пищухи запасают траву или высушенное сено, складывают их в норы, под наземные укрытия или на поверхности земли стожками. Бобры собирают на зиму запасы веточного корма, который чаще убирают в воду около гнезда, реже складывают вне воды. Находили запасы веток объемом до 20 м3.
Характерными особенностями запасания служат множественность запасов, обеспечивающих зверей кормом в голодный период, устройство специальных хранилищ для запасенного корма и коллективное, чаще семейное его употребление.
Укажем в заключение, что корм запасают и немногие виды зверей, впадающих на зиму в спячку. Таковы бурундуки и сибирские длиннохвостые суслики. Собранный в местах спячки корм используется этими видами весной, когда пробудившиеся зверьки еще не обеспечены вновь появившейся пищей.
В целом еще раз следует учесть, что приспособления млекопитающих к переживанию сезонных неблагоприятных условий жизни более разнообразны и совершенны, чем у других позвоночных. Особого внимания заслуживает собирание запасов корма. Из числа других позвоночных только немногие филогенетически молодые и прогрессивные группы птиц (воробьиные, совы, дятлы) собирают на зиму корм, но размеры запасов и приспособительное значение этой деятельности у них сравнительно с млекопитающими ничтожно.
Численность видов млекопитающих по годам меняется в широких пределах. Особенно значительные изменения наблюдаются у массовых, быстро размножающихся видов. Таковы многие грызуны, зайцеобразные, некоторые хищники. Кроме того, в больших пределах изменчива численность некоторых копытных: кабана, косули, северного оленя, сайгака и некоторых других. У сравнительно редких и медленно размножающихся видов, как, например, у крупных хищников (медведи, крупные кошки), резких изменений численности по годам нет или они менее очевидны.
В основе непостоянства численности зверей лежат многообразные и не всегда ясные причины. Можно утверждать, что различие в обилии особей по годам возникает в результате изменения интенсивности размножения и скорости отмирания особей видов.
Одной из наиболее распространенных причин колебаний численности млекопитающих служит изменчивость обилия кормов и его доступности. Периодически повторяющиеся неурожаи семян хвойных ставят белку в очень трудные условия, в результате которых сокращается размножение и повышается смертность. Подобного рода зависимость численности от изменчивости обилия кормов установлена для песца, лисицы, горностая и других зверей. Хорошо известно, что численность мышевидных грызунов, служащих основным кормом для упомянутых и других хищников, по годам меняется в больших пределах, что в свою очередь связано с разнообразными изменчивыми условиями среды.
Второй важной причиной непостоянства численности служат заболевания, принимающие в определенных условиях эпизоотический характер. Показательно, что эпизоотии чаще возникают среди видов, обеспеченность которых кормами по годам существенно не меняется.
Таковы многие виды зайцев, ондатра, водяная полевка и др. Природа эпизоотий разнообразна. Широкое распространение находят гельминтозы, например легочно-глистная болезнь, заражение печени плоскими червями трематодами, болезни, вызываемые простейшими (кокцидиоз, пироплазмоз, токсоплазмоз), разнообразные бактериальные и вирусные заболевания (туляремия, псевдотуберкулез, некробацилез, сибирская язва, чума плотоядных и др.).
Известны случаи, когда заболевание распространяется одновременно среди поголовья нескольких видов млекопитающих. Так бывает при туляремии, псевдотуберкулезе, чуме плотоядных. Однако выяснено, что не у всех видов, восприимчивых к той или иной инфекции, заболевание протекает одинаково остро. Например, к туляремии восприимчивы 50 видов млекопитающих нашей страны, но в острой форме болезнь протекает примерно у 40 видов, в том числе и у промысловых: зайцев, ондатры, водяной полевки, хомяков, кротов и некоторых других. Суслики, белка, бурундук, речной бобр, нутрия, выхухоль болеют менее остро, и болезнь обычно не приводит к смерти. К вирусу «дикования» весьма восприимчивы песцы, менее — лисицы; а волки, как предполагают, вовсе не восприимчивы.
Следует учитывать, что массовые заболевания не только приводят к непосредственной гибели зверей, но они снижают плодовитость, а также облегчают преследование жертвы хищниками и делают животных менее стойкими к пагубному воздействию неблагоприятных условий погоды.
Немаловажное значение для многих видов имеют отклонения условий погоды. Весенние заморозки с дождями и снегопадами губят ранние выводки зайцев, главным образом зайца-русака. Глубокие снега ставят в тяжелое положение многих копытных: кабанов, сайгаков, джейранов, маралов. Из хищников от глубокоснежья и гололеда особенно страдают корсаки, лисицы, енотовидные собаки и ряд других. Выпадающие иногда зимой дожди вызывают массовую гибель мышевидных грызунов, погибающих от затопления нор и невозможности достать корм из-под ледяной корки.
При отсутствии снегового покрова или недостаточной его мощности сильные морозы приводят к гибели в промерзшей почве кротов и землероек.
Значение хищников в изменениях численности «жертв» различно для разных видов. Зайцы-беляки, бурундуки, водяные полевки хотя и страдают вполне ощутимо от хищников, все же хищники обычно лишь усиливают скорость отмирания, которая в основе вызвана другими причинами. Однако в отдельных случаях могут возникать такие ситуации, в которых высокая численность хищников при низкой численности «жертв» может надолго задержать восстановление последних. Для медленно размножающихся копытных урон, наносимый хищниками, как правило, бывает большим, чем для многоплодных грызунов.
Замечено, что у мелких грызунов, например у лесных полевок, занимающих более или менее определенные гнездовые участки, по мере роста численности зверьков размножение заметно сокращается. Это происходит в силу нехватки убежищ и беспокойства, создаваемого многочисленностью зверьков.
Динамика численности млекопитающих проявляется во времени и пространстве с известной закономерностью. У ряда видов она имеет некоторую, хотя и не строго выдержанную, периодичность. У песца цикл изменения численности обычно равен 3—4 годам, у таежного зайца-беляка — 10—12 годам. В южных частях ареала последнего вида подъемы и спады бывают чаще, а интервалы между ними менее определенны. Кроме того, здесь меньше размах колебаний. Изменение численности вида в ту или иную сторону охватывает одновременно не всю площадь ареала, а лишь отдельные ее части. Пределы пространственного распространения этого явления в большей мере зависят от степени разнообразия, мозаичности природных условий: чем они однотипнее, тем на больших пространствах наблюдаются сходные изменения. Наоборот, в условиях разнообразного ландшафта распределение численности имеет весьма пестрый характер.
Практическое значение млекопитающих.
Значение млекопитающих в жизни человеческого общества весьма разнообразно. Это определяется, с одной стороны, обилием и разнообразием видов зверей, с другой — разнообразием хозяйственной деятельности человека. В целом класс млекопитающих представляет огромное природное богатство, заслуживающее бережного отношения. В настоящее время это весьма актуально, так как за предшествующие столетия уже полностью истреблено более 100 видов. Среди истребленных животных такие уникальные, как морская корова, бык тур, дикая лошадь тарпан и др.
Сейчас во многих странах ученые заняты важной проблемой — сохранением и увеличением численности редких животных, из млекопитающих — в первую очередь некоторых видов китов и тюленей, белого медведя, азиатского льва, азиатского носорога, лошади Пржевальского, кулана, горала, дикого пятнистого оленя и других.
Опыт показывает, что при надлежащей постановке дела восстановление численности видов вполне реально. Достаточно вспомнить замечательные результаты, которых удалось добиться при восстановлении поголовья сайгака, лося, соболя.
Особенное значение охрана полезных видов зверей приобрела после принятия Закона об охране природы в РСФСР и аналогичных законов, принятых в других республиках Советского Союза. Закон об охране природы предусматривает необходимость охраны и регулирования использования полезных диких животных как ресурсов охотничьего хозяйства, как объектов возможного одомашнивания. Закон предусматривает охрану редких и исчезающих видов животных как ценных памятников природы.
Итоговая оценка значения ряда видов иногда встречает затруднения, связанные с тем, что один и тот же вид в разной природной и хозяйственной обстановке играет разную роль. Многие виды мелких грызунов вредны для полевых культур. Местами они мешают лесонасаждению. Вместе с тем ими питаются пушные хищники, мех которых представляет большую товарную ценность. Лесные мышевидные грызуны составляют основу кормового благополучия для таких ценных зверей, как, например, соболь, куница, норка, колонок. Но в известной ситуации они представляют опасность для здоровья человека как хранители инфекций и прокормители клещей — передатчиков болезней. Лисица нежелательна близ птицеферм и на территории охотничьих хозяйств, где разводят фазанов или других птиц — объектов охоты. Этот же вид в таежной полосе никакого реального вреда не приносит и представляет большую ценность как пушной зверь.
Вблизи посевов, несомненно, вредны суслики и полевки. Они вредят посевам на разных стадиях произрастания их: поедают высеянные семена, портят всходы, уничтожают семена созревших растений. Местами грызуны вредят травяному покрову на пастбищах. Бывают случаи уничтожения в травостое более 50% самых ценных растений, например злаков и бобовых.
Некоторые грызуны (большая песчанка и др.) мешают пескоукрепительным работам, так как растаскивают заделанные в землю семена кустарников и древесных растений.
Слепыши, цокоры, местами водяные полевки и кроты, выбрасывая при сооружении нор землю на поверхность, затрудняют сенокошение.
Некоторые млекопитающие, в основном грызуны, имеют существенное эпидемическое значение, так как являются хранителями и передатчиками опасных для человека болезней. Кроме того, они служат прокормителями клещей и некоторых насекомых, распространяющих эти заболевания, называемые антропозоонозными. Сурки, суслики, песчанки, крысы — бациллоносители и распространители страшного заболевания людей — чумы. Чумные палочки передаются человеку при непосредственном контакте или через питающихся на больных зверьках блох. Микробоносителями туляремии среди млекопитающих служат преимущественно грызуны, в первую очередь водяная полевка, обыкновенная полевка, домовая мышь, суслики, зайцы. Возбудитель передается человеку через кровососущих насекомых, клещей, через воду или при непосредственном контакте с больным зверьком, например при снятии с него шкурки. При эпизоотиях пастбищных сыпнотифозных лихорадок возбудители (риккетсии) передаются от больных грызунов клещами. При вирусных энцефалитах (тяжелое поражение центральной нервной системы) возбудителей от больных зверьков также передают клещи.
С грызунами — вредителями сельского хозяйства или опасными в эпидемическом отношении — ведется планомерная борьба.
Диких млекопитающих добывают для получения разнообразных продуктов, а также ведут на них спортивную охоту, которая до настоящего времени не потеряла значения.
По промыслу диких зверей СССР занимает первое место в мире. Примерно из 350 видов, свойственных нашей фауне, около 150 потенциально могут быть объектами охоты. Такие виды есть во всех отрядах, кроме рукокрылых и непарнокопытных. Больше всего их среди хищников (около 40 видов) и грызунов (около 30 видов); менее всего среди насекомоядных (5 видов). Для получения пушнины и мехового сырья добывают более 50 видов, но основную массу промысла составляют всего около 20 видов. Наибольшее значение имеют белка, соболь и ондатра. Долевое значение каждого из этих видов в общей стоимости всей пушнины примерно 15%. На втором месте стоят лисица, песец и суслики (все их виды) — примерно по 10%. Далее идут крот (все виды), водяная полевка, куница (2 вида) — примерно по 5%. Долевое значение прочих видов равно 1—2%, а часто и того менее.
Приведенные данные характеризуют среднее значение видов за последнее время.
В связи с резкими изменениями численности ряда из них в конкретные годы удельное значение их может быть несколько иным. Особенно это характерно для белки, ондатры, зайцев.
Реальное практическое значение имеет также промысел копытных, при добыче которых используются мясо и кожа, а у некоторых и другие продукты. Практически повсеместно в лесной полосе добывают лося. В Нижнем Поволжье и в Казахстане существует регулярный промысел сайгака. Местами добывают северного оленя, марала, косулю, кабаргу, кабана. Добывание ценных пушных (например, соболя) и копытных зверей (например, лося, сайгака) тщательно регулируется и осуществляется только после выдачи специальных разрешений. Промысел массовых видов, особенно вредных в том или ином отношении (например, сусликов), поощряется.
В Советском Союзе охотничий промысел проводится на научной основе и предусматривает не только добывание зверей, но и их охрану и разведение. Уже проведена значительная работа по обогащению фауны восстановлением отечественных и акклиматизацией новых видов.
Очень больших успехов удалось добиться в восстановлении поголовья одного из наиболее ценных пушных зверей — соболя. Это было достигнуто многолетним запретом добычи, тщательной охраной и искусственным расселением. В настоящее время численность соболя выше, чем она была сто лет назад. Значительно увеличилась численность лесной куницы. Энергично ведется реакклиматизация речного бобра путем искусственного его расселения. Он обитает теперь в 50 областях, краях и АССР РСФСР, в Белоруссии и в ряде областей Украины. Местами уже начат выборочный промысел бобра на шкурку. Замечательные результаты получены при восстановлении численности лося и сайгака.
Проведено искусственное переселение некоторых отечественных видов пушных зверей в новые районы, куда они самостоятельно не могли расселиться. Так, соболь поселен в лесах Тянь-Шаня, дальневосточная енотовидная собака — в Европейской части СССР, белка — в лесах Кавказа, Крыма и Тянь-Шаня, заяц-русак — в степях Сибири, выхухоль — в водоемах Западной Сибири, пятнистый олень — в некоторых областях Европейской части СССР, здесь же местами поселен кабан.
Одновременно были поставлены опыты акклиматизации в СССР пушных видов из других стран. Наибольший эффект был получен при расселении североамериканского полуводного грызуна ондатры, распространенной теперь в бассейнах всех крупных рек нашей страны. Ондатра стала важнейшим в СССР пушным видом. Местами в Сибири и на Дальнем Востоке успешно акклиматизировалась ценная американская норка, добыча которой имеет существенное промысловое значение. В Закавказье ведется добыча акклиматизированных там североамериканского енота-полоскуна и южноамериканского полуводного грызуна нутрии.
Акклиматизация чужеземных видов зверей проводилась и в других странах. Так, в Западной Европе широко расселена лань, местами распространен корсиканский муфлон и ондатра. В Англии была акклиматизирована серая американская белка, которую вскоре пришлось истреблять, так как она оказалась вредной. В Северную Америку завезены заяц-русак, кабан, благородный олень, пятнистый олень, индийские олени замбар и аксис, индийская антилопа нильгау. Были случаи побега с ферм разводимых там муфлонов, африканских антилоп гну и зебр. Местами эти виды теперь существуют в природной обстановке.
Около 20 видов зверей (чаще преднамеренно, реже случайно) завезено в Новую Зеландию, где местных млекопитающих практически нет. Умышленно акклиматизированы 3 вида австралийских сумчатых (чернохвостый кенгуру и 2 вида кузу), благородный олень, американский олень вапити, лань, индийский олень замбар, американский белохвостый олень, американский лось, гималайский козел тар, серна, хорек, горностай, ласка, кролик, заяц-русак, еж. Случайно завезены домовая мышь и 2 вида крыс. Общий облик новозеландской фауны после акклиматизации зверей и птиц (24 вида) и одичания некоторых домашних животных, например свиней, изменился до неузнаваемости.
Известны случаи, когда завоз и акклиматизация зверей приносили не пользу, а вред. Так, например, обстоит дело с акклиматизацией европейского кролика в Австралии, где он оказался страшным вредителем сельского, главным образом пастбищного, хозяйства. Кролика не могли истребить завезенные и акклиматизированные хищники (лисицы, хорьки) и даже распространение среди кроликов особо опасной для них болезни — миксоматоза.
Приручение и одомашнивание млекопитающих, начатое в глубокой древности, продолжается и сейчас, хотя виды зверей находятся на разной степени одомашнивания и приручения. Полностью одомашненными надо считать те виды, которые хорошо подчиняются человеку и свободно размножаются в неволе. Таковы, например, собака, лошадь, рогатый скот и многие другие. Однако бывает и так, когда указанные два условия не совпадают. Тысячелетия назад были приручены слоны, послушно выполняющие разнообразные работы. Но в неволе слоны практически не размножаются. Львы и тигры хорошо размножаются в неволе, но не приручаются к человеку и не могут считаться одомашненными животными.
В малой мере одомашнены некоторые пушные звери, хотя в клетках они успешно размножаются. Таковы песцы, лисицы, норки, соболи и некоторые другие. В условиях полной свободы их, в отличие, например, от собак, кошек, содержать нельзя.
Пушное звероводство возникло менее ста лет назад, и число видов, разводимых в неволе, продолжает увеличиваться. Сравнительно недавно стали разводить норок и шиншилл. Однако в результате направленного подбора и отбора, зная законы наследования признаков, за короткий срок удалось вывести ряд новых пород. Таковы, например, серебристо-черная лисица и разнообразные цветные породы норок.
По сравнению с дикими предками мало изменились одомашненный северный олень, верблюд, як и южноазиатские быки. Это обусловлено тем, что условия содержания их мало отличаются от обстановки, в которой живут дикие родичи. Кроме того, не регулируется в должной мере подбор пар и отбор потомства. Наоборот, большинство домашних зверей образует большое разнообразие пород, отличных не только по внешнему виду, но и по хозяйственному использованию.
Это определилось в основном селекционной работой, при которой человек отбирал животных по каким-либо нужным для хозяйства признакам. Так возникли многочисленные породы лошадей, коров, овец, собак.
Однако нужно учитывать, что некоторые домашние животные возникли, видимо, не от одного, а от нескольких диких предков. Полагают, что одни породы собак произошли от волка, другие — от шакалов. Сложное происхождение имеют породы домашних свиней, возникшие в результате одомашнивания нескольких видов диких кабанов, потомки которых в последующем подвергались скрещиванию. Также обстоит, видимо, и с происхождением пород овец.
В целом история одомашнивания и породообразования во многом еще остается невыясненной, и предстоят многообразные палеонтологические, археологические и генетические исследования. Вместе с тем несомненны большие перспективы приручения и одомашнивания новых видов млекопитающих с разнообразными хозяйственными и эстетическими целями. Наряду с акклиматизацией диких зверей это обеспечит увеличение разнообразия окружающих нас видов.
Происхождение млекопитающих.
Предками млекопитающих, несомненно, были древние палеозойские рептилии, еще не утратившие некоторых черт строения амфибий: кожные железы, расположение сочленений в конечностях (между голенью и предплюсной и между предплечьем и запястьем) и, быть может, двойной затылочный мыщелок. Такой предковой группой считают подкласс звероподобных (Theromorpha).
Особенно близок к млекопитающим один из отрядов этого подкласса — отряд зверозубых (Theriodontia), у представителей которого зубы были дифференцированы на те же категории, что и у млекопитающих, т. е. на резцы, клыки и коренные, и сидели они в альвеолах. Имелось вторичное костное нёбо — признак весьма характерный для млекопитающих. Квадратная кость и сочленовная кость нижней челюсти были сильно уменьшены. При этом сочленовная кость не была приращена к нижней челюсти. Зубная кость, наоборот, была очень крупной. Здесь следует учесть, что у млекопитающих нижняя челюсть состоит только из зубной кости, а сочленовная кость превращена в одну из слуховых косточек — молоточек.
Такие известные высшие зверозубые, как иностранцевия (из пермских отложений на Северной Двине) и циногнатус (из триасовых отложений Южной Африки), не могли быть предками зверей, так как были высокоспециализированными и при этом крупными (2—3 м длиной) животными. Малые размеры древнейших млекопитающих дают основание искать предков зверей среди более примитивных и мелких зверозубых. В этом отношении представляет интерес иктидозавр (из верхнего триаса Южной Африки). Это было животное размером с крысу, скелет которого удивительно сходен с таковым млекопитающих; только нижняя челюсть у него состояла из нескольких костей. Очевидно, от таких мелких зверозубых и произошли млекопитающие.
Зверозубые просуществовали недолгое время, лишь до верхнего триаса.
Находки древнейших млекопитающих приурочены к юрским отложениям. Это были разнообразные и в ряде случаев высокоспециализированные животные; следовательно, возникновение млекопитающих надо отнести к более раннему времени — к триасу. С конца этого периода и до первой половины третичного времени существовали так называемые многобугорчатые. Это были мелкие, размером с крысу, зверьки, и только немногие достигали размеров сурка. Свое название они получили в связи с наличием на коренных зубах многочисленных бугорков. Клыков у них не было, а резцы развивались, наоборот, очень сильно.
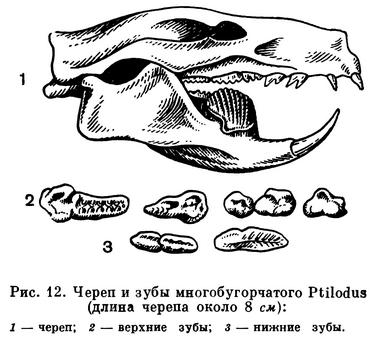
Многобугорчатые представляли специализированных растительноядных зверей, и их нельзя считать прямыми предками последующих групп млекопитающих. Лишь предположительно допускают, что наиболее ранние формы могли дать начало однопроходным. Основанием для такого предположения служит сходство строения их зубов с зубами зародыша утконоса (рис. 12).
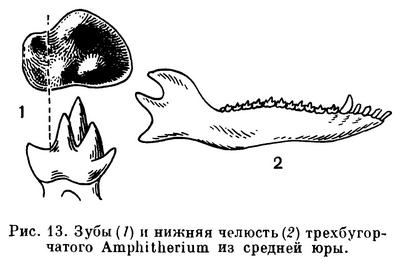
В отложениях с середины юры и до мела обнаруживаются многообразные трехбугорчатые, открывшие подлинную историю класса млекопитающих. Зубная система у них менее специализированная, чем у многобугорчатых; зубной ряд сплошной (рис. 13). Это были мелкие зверьки, близкие к насекомоядным; питались они частью животной, частью растительной пищей. Трехбугорчатые, в частности пантотерии, — наиболее вероятные предки сумчатых и плацентарных.
Сумчатые появились, видимо, в начале мелового периода. Однако первые ископаемые остатки их известны из верхнемеловых отложений Северной Америки. Это были мелкие зверьки типа опоссумов с бугорчато-режущими коренными и крупными клыками. В отложениях нижнетретичного времени они известны и в Евразии. Таким образом, родиной сумчатых следует считать северное полушарие. Однако еще до конца третичного времени они были вытеснены более высокоорганизованными плацентарными млекопитающими и в настоящее время сохранились только в Австралии, Новой Гвинее, Тасмании, Южной Америке, т. е. в южном полушарии, и лишь отчасти в Северной Америке (1 вид) и на острове Сулавеси (1 вид). Только в Австралии, куда из плацентарных проникли лишь рукокрылые и мышевидные грызуны, сумчатые дали большое разнообразие форм зверей, приспособленных к существованию в весьма различной жизненной обстановке.
Высшие, плацентарные млекопитающие, как и сумчатые, произошли от трехбугорчатых в начале мелового периода и развивались самостоятельно, параллельно с сумчатыми. Первые ископаемые остатки, относимые к насекомоядным, известны из верхнемеловых отложений Монголии. Плацентарные образовали 31 отряд, из которых 17 распространены в настоящее время, а 14 полностью вымерли.
Древнейшим отрядом, давшим, по-видимому, начало всем остальным отрядам, будут насекомоядные, известные из позднемеловых отложений. Это были мелкие зверьки, ведшие частью наземный, частью древесный образ жизни. Постепенное приспособление некоторых древесных насекомоядных вначале к планированию, а затем и к полету положило начало отряду рукокрылых. Увеличение плотоядности и переход на питание все более крупными животными послужили предпосылкой для формирования в начале третичного времени отряда древних хищников, или креодонтов. Но уже в олигоцене, когда медлительных травоядных начала третичного времени заменили более подвижные формы, креодонты были вытеснены новыми специализированными хищниками типа современных (Carnivora). В олигоцене уже существовали предковые группы вивер, куниц, собак, кошек. В миоцене и плиоцене были обыкновенны оригинальные саблезубые кошки с огромными изогнутыми клыками.
Очень рано, на грани между эоценом и олигоценом, хищники разделились на две неравноценные по обилию видов ветви: большую — наземных хищников и меньшую — ластоногих.
От древних хищников — креодонтов, перешедших на питание растительностью, в самом начале третичного времени (в палеоцене) возникли первичные копытные, или кондилятры. Это были сравнительно мелкие (размером от куницы до волка) всеядные животные с умеренно длинными клыками, бугорчатыми коренными зубами, с пятипалыми конечностями, в которых средний палец был развит сильнее остальных, а первый и пятый пальцы ослаблены. От кондилятр возникли две ветви современных копытных: непарнокопытные и парнокопытные. В эоцене возникли хоботные (корни их происхождения неизвестны). Также неясно происхождение и китообразных. От древних насекомоядных берут начало грызуны, неполнозубые, трубкозубые и приматы.
Наиболее древние ископаемые обезьяны найдены в палеоцене. Древесные обезьяны нижнего олигоцена (проплиопитекусы) дали начало гиббонам и крупным обезьянам из миоцена Индии (рамапитекусам), близким к антропоидам. Остатки человекообразных обезьян (орангутана, гориллы, шимпанзе) известны из плиоценовых отложений Африки, Индии, Китая. Некоторые из них (например, австралопитек, плезиантропус и парантропус) по строению стояли близко к человеку, и какая-то ветвь их дала начало всем его современным расам.
Следует подчеркнуть большую самостоятельность двух основных ветвей в филогении млекопитающих. К одной из них относятся только виды однопроходных (утконос, ехидна и проехидна). Эта ветвь берет начало от упоминавшихся ранее многобугорчатых и никогда не имела большого развития. К другой ветви, корнями уходящей к трехбугорчатым, относятся все остальные млекопитающие (сумчатые и плацентарные).
Современное представление о системе млекопитающих выглядит так:
Класс Млекопитающие (Mammalia)
Подкласс Первозвери (Prototheria)
Отряд Однопроходные (Monotremata)
Подкласс Настоящие звери (Theria)
Инфракласс Низшие звери (Metatheria)
Отряд Сумчатые (Marsupialia)
Инфракласс Высшие звери (Eutheria)
17 отрядов современных и
14 отрядов вымерших.
ПОДКЛАСС ПЕРВОЗВЕРИ (PROTOTHERIA)
ОТРЯД ОДНОПРОХОДНЫЕ, или ЯЙЦЕКЛАДУЩИЕ (MONOTREMATA)
Однопроходные (или яйцекладущие) — наиболее примитивные среди современных млекопитающих, сохранившие ряд архаичных особенностей строения, унаследованных от рептилий (откладка яиц, наличие хорошо развитой коракоидной кости, не соединенной с лопаткой, некоторые детали сочленения костей черепа и др.). Развитие у них так называемых сумчатых костей (небольших косточек таза) также рассматривается как наследие пресмыкающихся.
Присутствием отчетливых коракоидных костей однопроходные отличаются от сумчатых и других млекопитающих, у которых эта косточка стала простым выростом лопатки. Вместе с тем, волосяной покров и млечные железы — это два взаимосвязанных признака, характерные именно для млекопитающих. Однако млечные железы яйцекладущих примитивны и аналогичны по строению потовым железам, в то время как млечные железы сумчатых и высших млекопитающих имеют гроздевидную форму и похожи на сальные железы.
Довольно многочисленные черты сходства однопроходных с птицами — черты скорее адаптивные, чем генетические. Откладка яиц этими животными больше сближает однопроходных с пресмыкающимися, чем с птицами. Однако в яйце желток у однопроходных гораздо менее развит, чем у птиц. Ороговевшая скорлупа яиц состоит из кератина и также напоминает скорлупу яиц рептилий. Напоминают птиц и такие черты строения, как некоторая редукция правого яичника, наличие в пищеварительном тракте карманов, напоминающих зоб птиц, отсутствие внешнего уха. Однако эти черты сходства имеют скорее приспособительный характер и не дают права говорить о каком-то родстве непосредственно однопроходных и птиц.
У взрослых яйцекладущих зубы отсутствуют. В 1888 г. были обнаружены молочные зубы у детеныша утконоса, которые у взрослого животного исчезают; эти зубы разнообразны по строению, как у высших млекопитающих, а два самых крупных зуба на каждой челюсти имеют расположение и вид коренных. По температуре тела однопроходные занимают промежуточное положение между пойкилотермными (пресмыкающимися) и настоящими теплокровными (млекопитающими и птицами). Температура тела у ехидны колеблется около 30°, у утконоса — около 25°. Но это только средние цифры: они меняются в зависимости от температуры внешней среды. Так, температура тела ехидны при изменении температуры среды с +5° до +30° С повышается на 4—6°.
В настоящее время отряд однопроходных имеет 5 живых представителей, относящихся к двум семействам: утконоса и 4 вида ехидн. Все они распространены только в Австралии, на Новой Гвинее и Тасмании.
ПОДКЛАСС НАСТОЯЩИЕ ЗВЕРИ (THERIA)
ИНФРАКЛАСС НИЗШИЕ ЗВЕРИ (METATHERIA)
ОТРЯД СУМЧАТЫЕ (MARSUPIALIА)
Сумчатые млекопитающие, за исключением американских опоссумов, распространены на материке Австралии, на Новой Гвинее и близлежащих островах. К этому отряду относится около 200 видов из 9 семейств. Среди сумчатых есть насекомоядные, хищные и растительноядные формы. Сильно различаются они и по размерам. Длина их тела, включая длину хвоста, может колебаться от 10 см (сумчатая мышь Кимберли) до 3 м (большой серый кенгуру).
Сумчатые более сложно организованные животные, чем однопроходные. Температура тела у них выше (в среднем +36°). Все сумчатые рождают живых детенышей и выкармливают их молоком. Однако по сравнению с высшими млекопитающими у них много древних, примитивных черт строения, которые резко отличают их от остальных зверей.
Первая характерная черта сумчатых — наличие так называемых сумчатых костей (особых косточек таза, которые развиты и у самок, и у самцов). Сумка для вынашивания детенышей есть у большинства сумчатых, однако не у всех она развита в одинаковой степени; есть виды, у которых сумка отсутствует. У большинства примитивных насекомоядных сумчатых нет «законченной» сумки — кармана, а есть лишь небольшая складка, ограничивающая млечное поле. Так, например, обстоит дело у многочисленных сумчатых мышей, или мышевидок. У желтоногой сумчатой мыши — одного из наиболее архаичных сумчатых — есть лишь слабое поднятие кожи, как бы бордюр вокруг млечного поля; у близкой к ней жирнохвостой сумчатой мыши есть две боковые складки кожи, которые несколько разрастаются после рождения детенышей; наконец, у мышевидки-крошки есть уже нечто похожее на сумку, открывающуюся назад, к хвосту. У кенгуру, сумка которых совершеннее, она открывается вперед, к голове, наподобие кармана передника.
Вторая характерная особенность сумчатых — это особое строение нижней челюсти, нижние (задние) концы которой загнуты внутрь. Коракоидная кость у сумчатых слита с лопаткой, как у высших млекопитающих, — это отличает их от однопроходных.
Строение зубной системы — важный классификационный признак отряда сумчатых. По этому признаку весь отряд делится на 2 подотряда: многорезцовых и двурезцовых. Количество резцов особенно велико у примитивных насекомоядных и хищных форм, имеющих в каждой половине челюсти вверху по 5, а внизу по 4 резца. У растительноядных форм, напротив, не более одного резца с каждой стороны нижней челюсти; клыки у них отсутствуют или недоразвиты, а коренные зубы с притуплёнными бугорками.
Характерно строение млечных желез сумчатых; они имеют соски, к которым прикрепляются только что родившиеся детеныши. Протоки млечных желез открываются у края сосков, как у обезьян и у человека, а не во внутренний резервуар, как у большей части млекопитающих.
Однако основное отличие сумчатых от всех остальных млекопитающих — это особенности их размножения. Процесс размножения сумчатых, наблюдения за которым очень сложны, лишь совсем недавно выяснен до конца.
Детеныши в сумке матери сперва так малы и недоразвиты, что у первых наблюдателей возник вопрос: не родятся ли они непосредственно в сумке? Ф. Пельсарт, голландский мореплаватель, в 1629 г. впервые описал сумчатое животное. Он, как и многие более поздние натуралисты, думал, что детеныши у сумчатых рождаются прямо в сумке, «из сосков»; по этим представлениям, детеныш растет на соске, как яблоко на ветке дерева. Казалось невероятным, что полусформировавшийся зародыш, инертно висящий на соске, мог сам забраться в сумку, если он родился вне ее. Однако уже в 1806 г. зоолог Бартон, изучавший североамериканского опоссума, установил, что новорожденный может сам передвигаться по телу матери, залезать в сумку и прикрепляться к соску. Для австралийских животных это было подтверждено в 1830 г. хирургом Колли. Несмотря на эти наблюдения, знаменитый английский анатом Р.Оуэн в 1833 г. вернулся к уже высказывавшейся мысли, что новорожденного переносит в сумку мать. По мнению Оуэна, она губами берет детеныша и, придерживая лапами отверстие сумки, кладет его внутрь. Авторитет Оуэна более чем на полвека закрепил в науке эту неверную точку зрения.
Зародыш у сумчатых начинает развиваться в матке. Однако со стенками матки он почти не связан и в значительной мере является лишь «желточным мешком», содержимое которого быстро истощается. Задолго до того, как зародыш полностью сформируется, ему уже нечем питаться, и его «преждевременное» рождение становится необходимостью. Продолжительность беременности очень мала, особенно у примитивных форм (например, у опоссума или у сумчатых кошек от 8 до 14 дней, у коала она достигает 35, а у кенгуру — 38-40 дней).
Новорожденный очень мал. Его размеры не превышают 25 мм у большого серого кенгуру — самого крупного представителя отряда; у примитивных насекомоядных и хищников он еще меньше — около 7 мм. Масса новорожденного от 0,6 до 5,5 г.
Степень развития эмбриона к моменту рождения несколько различна, но обычно детеныш почти лишен шерсти. Задние конечности развиты слабо, согнуты и закрыты хвостом. Напротив, рот широко раскрыт, а передние лапки развиты хорошо, на них ясно видны коготки. Передние конечности и рот — это органы, которые в первую очередь понадобятся новорожденному сумчатому.
Как бы недоразвит ни был детеныш сумчатых, нельзя сказать, что он слаб и лишен энергии. Если отделить его от матери, он может прожить около двух суток.
У кенгуровых крыс и некоторых поссумов бывает только один детеныш; у коала и бандикутов иногда рождаются двойни. У большинства насекомоядных и хищных сумчатых детенышей много больше: 6—8 и даже до 24. Обычно число детенышей соответствует числу сосков матери, к которым они должны прикрепиться. Но часто детенышей больше, например у сумчатых кошек, у которых на 24 детеныша приходится лишь три пары сосков. В этом случае могут выжить лишь первые прикрепившиеся 6 детенышей. Бывают и противоположные случаи: у некоторых бандикутов, имеющих по 4 пары сосков, число детенышей не превышает одного-двух.
Чтобы прикрепиться к соску, новорожденный должен попасть в сумку матери, где его ожидают защита, тепло и пища. Как происходит это перемещение? Проследим его на примере кенгуру.
Новорожденный кенгуру, слепой и недоразвитый, очень скоро выбирает нужное направление и начинает ползти прямо к сумке. Он движется с помощью передних лапок с коготками, извиваясь, как червяк, и вертя головой по сторонам. Пространство, по которому он ползет, покрыто шерстью; это, с одной стороны, мешает ему, но, с другой, помогает: он крепко цепляется за шерсть, и стряхнуть его очень трудно. Иногда детеныш ошибается в направлении: доползет до бедра или груди матери и поворачивает назад, ищет до тех пор, пока не найдет сумку, ищет непрерывно и неутомимо. Найдя сумку, он тут же влезает внутрь, находит сосок и прикрепляется к нему. Между моментом рождения и временем, когда детеныш прикрепляется к соску, у крупных кенгуру проходит обычно от 5 до 30 минут. Прикрепившись к соску, детеныш теряет всю свою энергию; он снова на долгий срок становится инертным, беспомощным зародышем.
Что же делает мать, пока ее детеныш ищет сумку? Помогает ли она ему в этот трудный момент? Наблюдения за этим еще неполны, и мнения довольно разноречивы. В течение времени, необходимого новорожденному, чтобы добраться до сумки, мать занимает особое положение и не двигается. Кенгуру обычно сидят на хвосте, проходящем между задними лапами и направленном вперед, или ложатся на бок. Голову мать держит так, как будто все время наблюдает за детенышем. Часто она облизывает его — сразу после рождения или во время движения к сумке. Иногда она вылизывает свою шерсть по направлению к сумке, как бы помогая детенышу двигаться в нужную сторону. Если детеныш заблудится и долго не может найти сумку, мать начинает беспокоиться, чесаться и вертеться, при этом она может ранить и даже убить детеныша. В общем, мать скорее свидетель энергичной деятельности новорожденного, чем его помощник.
Первоначально сосок имеет удлиненную форму. Когда к нему прикрепляется детеныш, на его конце развивается утолщение, связанное, по-видимому, с выделением молока; это помогает детенышу держаться на соске, который он все время с силой сжимает ртом. Отделить его от соска, не разорвав ему рот или не повредив железы, очень трудно.
Детеныш пассивно получает молоко, количество которого регулирует мать с помощью сокращений мускулатуры млечного поля. Например, у коала мать снабжает детеныша молоком в течение 5 минут через каждые 2 часа. Чтобы он не захлебнулся этим потоком молока, имеется особое устройство дыхательных путей: воздух проходит непосредственно из ноздрей в легкие, так как нёбные кости в это время еще не до конца сформировались, и надгортанный хрящ продолжается вперед до носовой полости.
Оберегаемый и снабжаемый пищей, детеныш быстро растет. Развиваются задние лапы, становясь обычно длиннее передних; открываются глаза, и через несколько недель неподвижность сменяется сознательной деятельностью. Детеныш начинает отрываться от соска и высовывать голову из сумки. Первое время, когда он хочет вылезти наружу, его не пускает мать, которая может регулировать размер выходного отверстия сумки. Разные виды сумчатых проводят в сумке различный срок — от нескольких недель до нескольких месяцев. Пребывание детеныша в сумке кончается, как только он становится способен кормиться не молоком, а другой пищей.
Мать обычно заранее подыскивает гнездо или логово, где первое время дети живут под ее наблюдением.
Отряд сумчатых (Marsupialia) делится на 2 подотряда: многорезцовых (Polyprotodontia) и двурезцовых (Diprotodontia). К первым относятся более примитивные насекомоядные и хищные особи, ко вторым — растительноядные сумчатые. Промежуточное положение между многорезцовыми и двурезцовыми занимает малоизученная группа ценолестовых, которых некоторые зоологи считают отдельным подотрядом. Группа ценолестовых включает одно семейство и три рода. Это небольшие зверьки, напоминающие американских опоссумов и встречающиеся в Южной Америке.
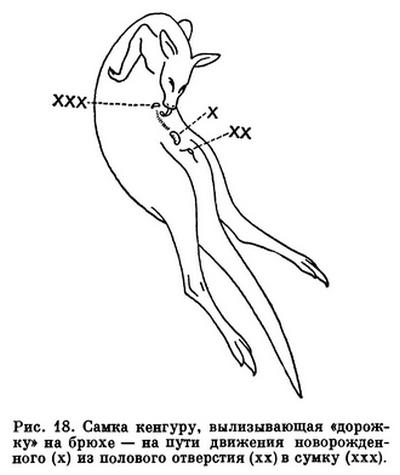
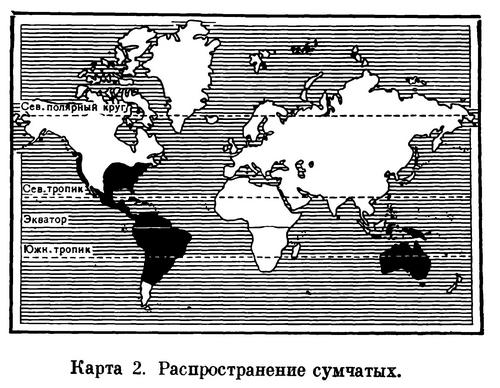
ПОДОТРЯД МНОГОРЕЗЦОВЫЕ (POLYPROTODONTIA)
Подотряд многорезцовых подразделяется на 5 семейств. Первое из них — семейство американских опоссумов (Didelphidae). Остальные 4 семейства предоставлены австралийскими животными (карта 2).
Семейство Американские опоссумы (Didelphidae)
К этому семейству относятся самые примитивные из сумчатых. Все ныне живущие его представители населяют Америку. Ископаемые формы известны из третичных отложений Европы.
Семейство Хищные сумчатые (Dasyuridae)
Хищные сумчатые примитивны и наиболее близки к американским опоссумам. Они имеют архаичную зубную систему с полным рядом резцов. Для них типично примитивное строение задних конечностей: они пятипалые, все пальцы хорошо развиты и обособлены один от другого. Зубная система, строение ног и размеры этих животных позволяют предположить, что один из наиболее примитивных представителей семейства — желтоногая сумчатая мышь — очень напоминает древнюю исходную форму, из которой когда-то развились все сумчатые.
Семейство хищных сумчатых (правильнее было бы называть его семейством хищных и насекомоядных) насчитывает 3 подсемейства, из них наиболее богатое видами примитивное подсемейство сумчатых мышей, или мышевидок, подсемейство собственно хищных сумчатых и подсемейство сумчатых волков.
Семейство Намбаты, или Сумчатые муравьеды (Myrmecobiidae)
Намбаты, или сумчатые муравьеды, происходят от хищных сумчатых семейства Dasyuridae. Они были впервые описаны в 1836 г. зоологом Уотерхаузом. Намбаты встречаются на западной и восточной окраинах Центральной австралийской пустыни. В начале колонизации Австралии они были довольно широко распространены в Западной и Южной Австралии. У западных зверьков спина покрыта черными полосками, а у восточных намбатов более однотонная окраска.
Семейство Сумчатые кроты (Notoryctidae)
По форме тела и по образу жизни сумчатый крот чрезвычайно похож на обыкновенных кротов, точнее, на африканского златокрота. Сходство их так велико, что даже у анатомов возникал вопрос: нет ли между этими двумя животными действительно близкого родства? Бесспорно, что сумчатый крот и обычные кроты — насекомоядные — не родственники. Их сходство всего лишь пример самой совершенной конвергенции, усугубленной специфическим образом жизни. По некоторым особенностям строения зубов и конечностей можно предполагать,что по происхождению сумчатый крот близок к группе бандикутов, о которой будет сказано ниже. В настоящее время его выделяют в особое семейство.
Два известных вида сумчатых кротов живут в резко засушливых районах, по окраинам пустынь.
Семейство Австралийские бандикуты (Peramelidae)
Под именем бандикутов со времен путешественника Басса объединяют сумчатых с заостренной мордой, длинноухих, размером от крысы до кролика. Это семейство представляет собой как бы переход от хищных к растительноядным сумчатым. Бандикуты главным образом насекомоядные и хищные животные. Однако у многих есть тенденция к смешанному питанию, а один из видов — свиноногий бандикут почти полностью растительнояден. К растительноядным сумчатым их приближает также строение задних конечностей, второй и третий пальцы которых срослись, как у поссумов или кенгуру; в результате образовался особый двойной коготь, с помощью которого животные освобождаются от паразитов.
ПОДОТРЯД ДВУРЕЗЦОВЫЕ (DIPROTODONTIА)
В подотряд двурезцовых объединяются растительноядные сумчатые, имеющие не более трех резцов на каждой стороне верхней челюсти и обычно по одному — на нижней. В процессе адаптации при переходе от насекомоядного типа питания к растительноядному у них исчезли мелкие передние зубы, типичные для первого типа, а размер каждого зуба увеличился, как это обычно бывает у травоядных животных. Подавляющее большинство этих сумчатых растительноядны, хотя некоторые архаичные виды питаются насекомыми.
Вторая характерная черта подотряда — строение задних конечностей: у всех его представителей, кроме коала, второй и третий пальцы срослись и имеют вид одного пальца с раздвоенным когтем. Поссумы, вомбаты и кенгуру, как и бандикуты, применяют двойной коготь для чесания и чистки меха.
Подотряд двурезцовых делится на 4 семейства: семейство древесноживущих поссумов (самые мелкие и низкоорганизованные из растительноядных сумчатых; питание смешанное: насекомые, нектар, пыльца и другая растительная пища); семейство коала (высокоспециализированные бесхвостые сумчатые с единственным ныне живущим видом); семейство вомбатов (роющие животные; отличаются от остальных сумчатых грызуноподобной зубной системой; коренные не имеют корней и растут в течение всей жизни); семейство кенгуру (высокоспециализированная группа животных, приспособившихся к передвижению прыжками; почти все виды — наземные животные). К этому же подотряду относилось вымершее семейство Diprotodontidae, исчезнувшее уже в четвертичном периоде.
ИНФРАКЛАСС ВЫСШИЕ ЗВЕРИ (EUTHERIA)
ОТРЯД НАСЕКОМОЯДНЫЕ (INSECTIVORA)
В отряде преобладают мелкие и очень мелкие зверьки. Голова у большинства клиновидно заостренная, с удлиненным в виде хоботка носом. Тело у большинства зверьков покрыто ровным густым бархатистым мехом, у немногих — жесткими щетиновидными волосами или короткими иглами. Конечности устроены по-разному, в зависимости от образа жизни. Для многих насекомоядных характерны пахучие (мускусные) железы. Зубная система примитивная. Число зубов от 44 до 26. Набор из 44 зубов считается исходным (самым примитивным) для высших зверей. Предкоренные зубы по строению отчетливо делятся на две категории — малые предкоренные и большие, или крупные, предкоренные. Малых предкоренных у представителей разных родов и семейств бывает разное число, поэтому они имеют большое значение в систематике, а крупных предкоренных только по одному с каждой стороны в верхней и нижней челюстях. Большие коренные примыкают к малым коренным, по размерам всегда крупнее малых и имеют по 2 или 3 корня, часто по нескольку острых вершин. На коренных зубах между острыми вершинами проходят режущие гребни (комиссуры), образующие узор, похожий на букву W или V. Зубы такого строения называют зубами насекомоядного типа.
Головной мозг с относительно большим обонятельным отделом, а размеры полушарий невелики, у многих без борозд и не покрывают сверху мозжечок. Из органов чувств наибольшего развития достигают органы обоняния и осязания. Органы зрения почти у всех развиты слабо, у некоторых глаза вообще скрыты под кожей. В связи со слабым развитием головного мозга условные рефлексы у них вырабатываются медленно и быстро утрачиваются (затухают).
Насекомоядные распространены почти по всему свету за исключением Антарктиды, Австралии и большей части Южной Америки. К северу они идут до берегов и больших островов Ледовитого океана, в горах поднимаются почти до нижнего края снежников и ледников (карта 3).
Насекомоядные — одна из древнейших групп высших зверей. Ископаемые остатки их известны из верхнемеловых отложений мезозойской эры. Время, прошедшее с тех пор до наших дней, составляет около 135 миллионов лет. Их происхождение связывают с юрскими звероподобными существами подкласса пантотериев, отряда тритуберкулят.
Систематика насекомоядных до сих пор остается недостаточно разработанной. Учитывая мнения авторов систематических сводок и материалы, изученные при разработке системы насекомоядных фауны СССР, мы объединяем в этот отряд следующие семейства:
Ежовые (Erinaceidae).
Гимнуровые (Echinosoricidae).
Тенрековые (Tenrecidae).
Златокротовые (Chrysochloridae).
Выдровоземлеройковые (Potamogalidae).
Выхухолевые (Desmanidae).
Кротовые (Talpidae).
Землеройковые (Soricidae).
Щелезубые (Solenodontidae).
Прыгунчиковые (Macroscelididae).
ОТРЯД ШЕРСТОКРЫЛЫ (DERMOPTERA)
Шерстокрылы имеют плотную, покрытую мехом перепонку, которая соединяет шею, все конечности и хвост. Глаза этих зверьков очень крупные, с круглым зрачком. Все конечности пятипалые и снабжены прочными острыми когтями. Зубов — 34; у верхних клыков и наружных верхних резцов по два корня, что представляет редкое исключение среди зверей. Нижние резцы гребнеобразно зазубрены, причем на каждом зубе бывает до двенадцати зубчиков; возможно, шерстокрылы пользуются ими при расчесывании волос. В современной фауне известно всего 1 или 2 вида шерстокрылов.
ОТРЯД РУКОКРЫЛЫЕ (CHIROPTERA)
Рукокрылые — мелкие или средних размеров зверьки, способные к настоящему длительному полету. Передние конечности их видоизменены в крылья: предплечье, пястные (метакарпальные) кости и фаланги всех пальцев, кроме первого, сильно удлинены; между плечом, предплечьем, пальцами, боками тела и задними конечностями натянута тонкая эластичная летательная перепонка. Задние конечности вывернуты так, что колени обращены дорзально. Ушные раковины обычно большие, иногда относительно величины тела огромные, у многих с хорошо развитым кожным выступом — козелком. Хвост у большинства видов длинный, полностью или частично заключенный в межбедренную перепонку; свободный край этой перепонки поддерживается парной хрящевой или костной шпорой, отходящей от пятки. Вдоль основания шпоры у многих видов тянется своеобразная кожная лопасть — эпиблема.
Межчелюстные кости черепа всегда недоразвиты или даже отсутствуют. В зубной системе есть все категории зубов. Средняя пара верхних резцов всегда отсутствует. Нижние резцы очень мелкие. Клыки большие. Коренные зубы делятся на 3 естественные группы: малые предкоренные, крупные (или большие) предкоренные и задние (или собственно) коренные. Наиболее полная зубная формула выглядит так:
i 2/3, c 1/1, pm 2/2, pmp 1/1, m 3/3 = 38
Количество резцов и особенно малых предкоренных имеет большое значение в родовой систематике рукокрылых. Молочные зубы не только по размерам, но и по форме резко отличаются от постоянных.
Около 1000 видов рукокрылых группируются в 2 подотряда: 1) крылановых (Pteropoidei) с одним семейством (Pteropidae) и 2) кожановых, или летучих мышей (Vespertilioidei), с 14 семействами; одно из них — семейство клейконогих (Natalidae) — некоторые систематики делят на 3 семейства. В фауне СССР представлены 40 видов из 3 семейств только второго подотряда.
ОТРЯД НЕПОЛНОЗУБЫЕ (EDENTATA)
К этому небольшому, но крайне своеобразному отряду относятся ленивцы, муравьеды и броненосцы, резко различающиеся как внешне и по размерам (длина тела их от 12 см до 1 м 20 см, масса от 90 г до 55 кг), так и по образу жизни. Однако все они характеризуются либо полным отсутствием зубов (муравьеды), либо упрощенным строением их. Зубы лишены эмали и корней, резцы и клыки отсутствуют (за исключением двуполых ленивцев, у которых есть клыкообразные зубы), все зубы однородные, с постоянным ростом. На последних грудных и на поясничных позвонках есть совершенно своеобразные придаточные сочленения, дополнительно укрепляющие опору конечностей. На передних лапах обычно меньше пяти пальцев, но два или три из них развиты чрезвычайно сильно и вооружены мощными когтями. Полушария переднего мозга развиты слабо и почти лишены борозд, но хорошо развиты обонятельные центры мозга. У большинства есть хороша развитая крупная воронья кость, поздно срастающаяся о лопаткой.
Отряд неполнозубые включает 3 современных семейства, состоящих из 14 родов и около 30 видов. Неполнозубые распространены в Южной и Центральной Америке и на юге центральных и юго-восточных штатов США (карта 4).
В ископаемом состоянии неполнозубые известны только из западного полушария, где обитало в прошлом примерно в 10 раз больше видов, чем в настоящее время (7 семейств вымерло полностью).
Древнейшие неполнозубые известны из позднего палеоцена Южной Америки, но отделялись они от Palaeodonta, очевидно, еще в мелу, поскольку даже наиболее примитивные из них были уже типичные» представители отряда с костным панцирем и безэмалевыми зубами. В течение почти всего третичного периода (многих миллионов лет) неполнозубые были изолированы в Южной Америке и дали там несколько групп, из которых немногие только в позднем плиоцене проникли в Северную Америку.
Среди ископаемых неполнозубых интересны гигантские ленивцы, или тихоходы (Gravigrada), например мегатерии, достигавшие величины слона и кормящиеся, подобно современным ленивцам, листьями. Видимо, они пригибали деревья к земле, пользуясь своей огромной силой и тяжестью. Другие тихоходы, такие как милодон и мегалоникс, были также тяжеловесные и толстоногие животные, размером с быка. В толще кожи у этих гигантских ленивцев найдены кожные окостенения, что делает их как бы промежуточным звеном, связывающим современных ленивцев с броненосцами. Тихоходы населяли кустарниковые саванны. Они вымерли лишь в позднем голоцене, т. е. 10—12 тысяч лет назад, и были современниками древнего человека. В пещерах Патагонии найдены не только многочисленные скелеты и помет этих животных, но и кожа со следами разрезов, сделанных человеком. Есть сведения о том, что человек держал некоторые виды их в особых загонах, как домашних мясных животных, и один вид ископаемого тихохода даже получил название Mylodon domesticum.
Другая интересная группа ископаемых неполнозубых — глиптодонты, или гигантские броненосцы (Glyptodontida), включала крупных, иногда размером с носорога, броненосцев с мощным костным панцирем. У некоторых видов окостенения на спине срастались, образуя подобие панциря черепах (Glyptodon); у других на конце хвоста развивались мощные костные шипы, образуя как бы булаву, служившую, видимо, орудием защиты (Doedicurus).
В настоящее время у себя на родине, в Южной Америке, ряд видов неполнозубых вполне обычны и даже многочисленны. Пищей им служат листья деревьев (ленивцам), муравьи и термиты (муравьедам и многим броненосцам), реже — другие растительные и животные корма. Среди неполнозубых много видов, узкоспециализированных по питанию. Для представителей этого отряда типично длительное развитие зародышей, обычно со стадией покоя (латентной), и рождение однояйцовых близнецов.
Мясо большинства неполнозубых используется местными жителями в пищу. Некоторые виды стали редкими, главным образом, из-за преследования их человеком и подлежат охране (большой муравьед, ошейниковый ленивец, гигантский, трехпоясной шаровой, большой и малый плащеносные броненосцы).
ОТРЯД ЯЩЕРЫ, ИЛИ ПАНГОЛИНЫ (PHOLIDOTA)
В этот отряд объединены своеобразные млекопитающие, напоминающие огромную еловую шишку благодаря серо-коричневым роговым чешуям, которые, налегая друг на друга, покрывают тело животного.
Нижняя поверхность тела и лап, а также морда покрыты волосами. Передние лапы с сильными роющими когтями. Морда вытянута, рот сужен, челюсти беззубые, язык длинный (до 40 см), очень большие слюнные железы выделяют клейкий секрет. Питается почти исключительно муравьями и термитами. Как показали недавние исследования, слюна ящеров обладает привлекательным для насекомых медовым запахом; ватка, смоченная этой слюной, особенно привлекает термитов и муравьев. Желудок, в котором измельчается пища, выстлан ороговевшим эпителием, и в его полость вдается особая складка, вооруженная роговыми зубами. Животные заглатывают камешки, что способствует измельчению пищи. Очень специализированы по питанию. Одни ящеры (например, длиннохвостый) предпочитают муравьев, а другие, например белобрюхий, — термитов. В желудках панголин находили от 150 до 2000 г термитов или муравьев (преимущественно 2—3 видов). Другую пищу эти животные почти не употребляют, поэтому их очень трудно содержать в неволе.
Все ящеры относятся к одному семейству и одному роду; 7 видов из них отличаются друг от друга размерами, длиной хвоста, расположением, формой и числом роговых чешуй.
ОТРЯД ЗАЙЦЕОБРАЗНЫЕ (LAGOMORPHA)
Еще сравнительно недавно зайцеобразных, к которым, кроме общеизвестных зайцев, принадлежат мелкие пищухи, или сеноставцы, включали в отряд грызунов в качестве подотряда. Но уже в 1912 г. Гидлей учел многие специфические черты организации и истории зайцеобразных и предложил выделить их в самостоятельный отряд, некоторое сходство которого с грызунами имеет только внешний (конвергентный) характер. Этот взгляд в настоящее время следует считать общепринятым.
От грызунов зайцеобразные принципиально отличаются тем, что в верхней челюсти у них не одна, а две пары резцов. Вторая пара резцов развита слабее и располагается позади основной пары; вершины их далеко не доходят до вершин основных (передних) резцов. Кроме того, костное нёбо у зайцеобразных устроено очень своеобразно. Оно имеет вид узкого поперечного мостика между левым и правым рядом коренных зубов. У грызунов костное нёбо представляет сплошную площадку, доходящую впереди до резцов (рис.83, 84). Есть существенные отличия в организации желудка. У зайцеобразных он состоит (физиологически, но не морфологически) из двух отделов, в одном из которых происходит бактериальное брожение пищи, в другом, ближнем к выходу кишки, пища переваривается пепсином.
Основное сходство зайцеобразных и грызунов состоит в том, что у них отсутствуют клыки, а резцы и коренные зубы отделены широким, лишенным зубов пространством — диастемой.
Размножаются быстро, принося обычно по нескольку пометов в год. Половая зрелость наступает рано, на следующее после рождения лето.
Зайцеобразные возникли в начале третичного времени, и ископаемые их остатки известны из отложений палеоцена. Распространены по всему земному шару, за исключением южной части Южной Америки и Мадагаскара; в Австралии, Новой Зеландии и на многих океанических островах акклиматизированы. Всего современных видов около 60, объединенных в 2 семейства: зайцы (Leporidae) и пищухи (Lagomyidae).
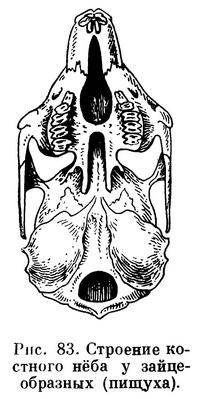
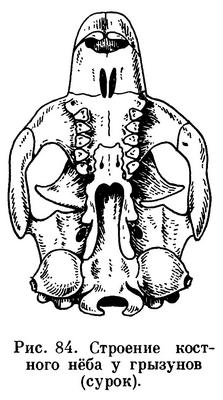
ОТРЯД ГРЫЗУНЫ (RODENTIA)
Зубная система — наиболее характерная особенность, по которой звери объединяются в этот отряд и отличаются от животных любых других отрядов. Резцы грызунов, расположенные по одному с каждой стороны верхней и нижней челюсти, очень велики, лишены корней и постоянно растут. Их свободный конец долотообразно заострен. Стачиваются резцы неравномерно и всегда остаются острыми. Клыков у грызунов нет и коренные зубы отделены от резцов широким беззубым промежутком — диастемой. Общее число коренных зубов (вместе с предкоренными) с каждой стороны: в верхней челюсти — от 5 до 1, в нижней челюсти — от 4 до 1, а у грызунов фауны СССР соответственно от 5 до 3 и от 4 до 3. Уменьшение количества зубов у грызунов идет сначала за счет передних предкоренных, соответствующих малым предкоренным зубам насекомоядных и рукокрылых, а затем за счет самых задних коренных. Коренные зубы у грызунов имеют широкую жевательную поверхность из рядов тупых бугорков (исходное строение), невысоких поперечных гребней, образовавшихся в результате соединения бугорков, или их жевательная поверхность становится складчатой. У некоторых грызунов образуются хорошо обособленные корни, но у большинства корни не формируются, и в таком случае зубы становятся постоянно растущими. Количество коренных зубов и строение их жевательной поверхности — важные признаки, используемые для разделения грызунов в разные семейства, подсемейства и роды, а иногда и в диагностике видов.
Грызуны населяют все материки, кроме Антарктиды, и большинство островов Мирового океана, на многие из которых они были завезены человеком.
В отряд объединяется более одной трети современных млекопитающих, населяющих земной шар (около 2500 видов). Они группируются в 32 семейства и 3 надсемейства (дикобразовые, белковые и мышиные), выделяемые некоторыми систематиками в ранг подотрядов.
ОТРЯД КИТООБРАЗНЫЕ (СЕТАСЕА)
Китообразные — резко отклонившаяся группа водных млекопитающих, внешне похожих на рыб, но отличающихся от них наличием теплокровности, легочного дыхания, а также внутриутробным развитием, выкормом детенышей молоком и многими другими чертами.
70 миллионов лет назад наземные предки китообразных переселились в воду. Здесь они, спасаясь от наземных врагов и конкурентов, первоначально питались на мелководьях, а затем в поисках пищи все смелее стали отходить от берегов и, наконец, утратили связь с сушей. Так как сопротивление воды при передвижении в ней в 800 раз больше, чем в воздухе, то у китообразных развивалась хорошо обтекаемая торпедовидная форма тела. В новой среде за долгую эволюцию в корне менялось их строение и образ жизни. На теле исчезло все, что мешало его скольжению: шерстный покров, задние ноги, ушные раковины. Большая упругость, эластичность и несмачиваемость голой неороговевающей кожи помогали китообразным преодолевать сопротивление трения при быстром плавании. Передние конечности превратились в грудные, уплощенные жесткие плавники — это рули высоты, поворотов и торможения; они же обеспечивали движение тела назад. Сжатая с боков и мускулистая хвостовая часть тела китообразных очень гибка, подвижна, заканчивается широкими горизонтальными лопастями. У большинства видов появляется спинной плавник, придающий устойчивость телу в воде. Грудные, спинной и особенно хвостовой плавники китообразных, как открыли в 1968 г. советские ученые (С. В. Першин, А. С. Соколов, А. Г. Томилин), имеют переменную упругость, которая меняется в зависимости от скорости плавания и регулируется специфическими комплексными кровеносными сосудами (рис. 138), открытыми А. Г. Томилиным в 1947 г. Такой регулируемый гидроупругий эффект в плавниках — важнейшее приспособление к различному режиму плавания. Приспособления, обеспечивающие высокую скорость передвижения китообразных (до 50 км/ч), в настоящее время интересуют судостроителей, стремящихся создать наиболее выгодную (мягкую) обшивку и форму кораблей.
Все плавники выполняют также роль терморегуляторов, через которые в первую очередь отдается избыток тепла при перегреве тела. Потовых и сальных желез нет. Толстый подкожный слой жира защищает тело от охлаждения и используется как энергетический запас во время сезонных голодовок китообразных, поэтому он сильно колеблется по сезонам года.
Под слоем жира в задней половине или трети тела расположены две млечные железы, каждая с соском. Соски у китообразных помещаются в двух продольных кожных карманах по бокам мочеполовой щели и только у кормящих самок выступают наружу.
Окраска китообразных бывает однотонной, противотеневой (темной сверху и светлой снизу) или расчленяющей, с резкими светлыми участками кожи и пятнами на ней.
Скелет китов губчатый, пропитанный жиром. В позвоночнике от 41 до 98 позвонков, образующих четыре отдела: шейный (очень короткий, но всегда из семи свободных или срощенных позвонков), грудной, поясничный и хвостовой. Грудной отдел несет 10—17 пар ребер, из которых только первые 2—8 пар сочленяются с грудиной. Межпозвоночные диски придают позвоночнику, особенно его хвостовой части, большую подвижность. Утрата задних конечностей, крестцового отдела позвоночника и таза увеличивает свободу движения хвостового стебля и позволяет рождать очень крупных и развитых детенышей. К паре небольших косточек, оставшихся от таза, у самцов прикрепляются пещеристые тела копулятивного органа, а у самок — мышцы, расширяющие влагалище. Плоские грудные плавники поддерживает очень короткая плечевая кость, две короткие кости предплечья и многочисленные косточки кисти, в которой 4 или 5 пальцев с увеличенным числом фаланг. Ключица исчезает, лопатка веерообразной формы (рис. 139).
Череп приспособлен к тому, чтобы дыхание совершалось при выставлении ноздрей из воды без изгибания шеи (ноздри смещены на темя). Верхнечелюстные, межчелюстные и нижнечелюстные кости удлинены в связи с развитием цедильного аппарата (китового уса) или многочисленных одновершинных зубов. Носовые кости уменьшены, теменные сдвинуты в бока так, что верхнезатылочная кость соприкасается с лобными. Дыхало — одно или два наружных носовых отверстия — расположено на вершине головы и открывается лишь в момент короткого дыхательного акта выдоха — вдоха, производимого сразу же после выныривания. В прохладную погоду при выдохе взлетает вверх конденсированный пар, образуя так называемый фонтан, по которому китобои различают вид кита. Иногда с этим паром взлетают и распыленные брызги воды. Все остальное время, пока длится дыхательная пауза и животное ныряет, ноздри плотно закрыты клапанами, которые не пропускают воду в дыхательные пути. Вследствие особого строения гортани воздухоносный путь отделен от пищевого. Это позволяет безопасно дышать, если вода или пища находятся в ротовой полости. Носовой канал большинства видов соединен с особыми воздушными мешками и вместе с ними выполняет роль звукосигнального органа.
Легкие весьма упруги и эластичны, приспособлены к быстрому сжатию и расширению, что обеспечивает очень короткий дыхательный акт и позволяет обновлять воздух за одно дыхание на 80—90% (у человека только на 15%). В легких сильно развиты мускулатура альвеол и хрящевые кольца даже в мелких бронхах, а у дельфинов — и в бронхиолах.
Китообразные могут долго (кашалоты и бутылконосы до 1,5 часов) находиться под водой с одним и тем же запасом воздуха: большая емкость легких и богатое содержание мышечного гемоглобина позволяют им уносить с поверхности повышенное количество кислорода, который расходуется очень экономно: во время ныряния деятельность сердца (пульс) замедляется более чем вдвое и ток крови перераспределяется так, что кислородом снабжаются в первую очередь мозг и сердечная мышца. Эти органы при длительном погружении получают кислород также с артериальной кровью из запасов «чудесной сети» — тончайшего разветвления кровеносных сосудов. Менее чувствительные к кислородному голоданию ткани (особенно мышцы тела) переводятся на голодный паек. Мышечный гемоглобин, придающий мускулатуре темный цвет, снабжает мышцы кислородом во время дыхательной паузы. Пониженная чувствительность дыхательного центра к накоплению углекислоты в крови позволяет китообразным удлинять дыхательную паузу и использовать ее для кормежки.
ОТРЯД ХИЩНЫЕ (CARNIVORA)
Звери, входящие в отряд хищных, выделяются среди остальных групп млекопитающих необычайным разнообразием внешнего вида, размеров тела, биологических особенностей, приспособлений к среде обитания, способов передвижения и т. д. Достаточно сказать, что к отряду хищных принадлежат и миниатюрная ласка, и могучий тигр, и огромный белый медведь. Большинство хищных зверей ведет наземный образ жизни, но отдельные виды, вроде норок и выдр, стали обитателями пресных водоемов, а калан — морским животным. Вопреки своему названию некоторые хищники предпочитают питаться не мясом, а насекомыми, водными беспозвоночными и даже растительной пищей. Соответственно этому, они сильно различаются по своей биологии, давая широкий спектр приспособительных типов. Тем не менее всех этих, казалось бы, столь не похожих друг на друга зверей мы с полным основанием относим к одному отряду.
Тому причиной сходство морфологических особенностей (главным образом в строении черепа и зубной системы) и историческое родство.
Длина тела у хищных колеблется от 14 см до 3 м, масса от 100 г до 1000 кг. Форма тела изменяется от вытянутой, гибкой до массивной, иногда неуклюжей. У одних зверей высокие, стройные конечности, у многих других — короткие, неуклюжие. На каждой лапе насчитывается не менее четырех пальцев, а у медведей и собак их по пять. Они вооружены когтями, особенно острыми у кошек, у которых (за исключением гепарда) могут втягиваться (втягивающиеся когти есть также у некоторых виверр). Напротив, у отдельных видов выдр и у калана когти превратились в подобие ногтей. У большинства хищников имеется длинный, нередко пушистый хвост и лишь у медведей, большой панды и ряда других он мал и скрыт в меху. Представители двух родов енотовых и виверр обладают хватательным хвостом. Наружные ушные раковины у большинства видов хорошо развиты, заострены, у фенека и большеухой лисицы необычайно велики, тогда как у песца, горностая, ласки и других едва выдаются из окружающего меха, а у калана недоразвиты.
У всех хищных зверей хорошо развит волосяной покров, варьирующий по густоте, длине, пышности, а также окраске. Многим видам свойственна пестрая окраска меха (пятнистая, полосатая и другая), достигающая наибольшей яркости у южных форм. У отдельных северных видов наблюдается сезонная смена окраски — побеление меха зимой (ласка, горностай, песец) или его значительное посветление (полярный волк).
В соответствии с характером питания череп у большинства видов обладает сильно развитыми гребнями, широко расставленными скуловыми дугами, а иногда также крупными отростками в затылочной части, служащими для прикрепления мощной мускулатуры. Количество зубов колеблется от 28 до 48. Среди них обращают внимание хорошо развитые, более или менее изогнутые, заостренные клыки. Жевательная поверхность предкоренных и коренных зубов обычно бугорчато-режущего типа, но иногда становится тупо-бугорчатой и даже почти уплощенной (калан). Последний верхний предкоренной и первый нижний коренной у большинства видов превратились в особые хищнические зубы, отличающиеся величиной и острыми, режущими бугорками. Напротив, резцы относительно невелики.
ОТРЯД ЛАСТОНОГИЕ (PINNIPEDIA)
Ластоногие, как и китообразные, водные (преимущественно морские и океанические) млекопитающие. Однако связанность с водной средой у них не столь велика, как у китов. Они не могут спариваться и рождать в воде и в период размножения выходят на лед или на сушу. Смена мехового покрова у ластоногих проходит вне воды. Наконец, ряд видов ластоногих выходит из воды и для периодического отдыха. Надо указать также, что пищу они добывают только в воде, чем подчеркивается полнота связи с водой как с обязательной средой обитания.
Ластоногие — немногочисленная (около 30 видов) и очень четко выраженная группа. Сравнительно малые масштабы различий между составляющими ее видами, очевидно, связаны с относительным однообразием жизненной обстановки, в которой возникли и обитают ластоногие, и, быть может, с молодостью отряда, возникшего, видимо, в середине третичного периода. Филогенетически наиболее близкие к ластоногим наземные хищники (отряд Carnivora) имеют значительно более длинную историю, они обитают в крайне разнообразных природных условиях и поражают огромным разнообразием форм.
В большинстве ластоногие — это крупные или средней величины звери, длина тела которых 1,2—6,0 м, а общая масса тела от 40 кг до 3,5 т. Тело удлиненное, веретенообразной формы, округлое в сечении и суживающееся к головному и хвостовому отделам. Характерна форма шеи, которая не отграничена резко от туловища и головы. Шея очень толстая, особенно в задней своей части. По направлению к голове она постепенно суживается, но остается шире последней. У большинства видов подвижность шеи невелика, исключение составляют ушатые тюлени (например, морской лев).
В результате длительного приспособления к жизни в водной среде весьма существенны особенности строения конечностей и их соотношение с туловищем. Значительная часть конечностей скрыта в кожном туловищном мешке, и наружу выступают у передней пары конечностей не более чем половина предплечья и кисть, у задней пары их — только задняя треть голени и стопа, а у некоторых — только стопа. С указанным сочетается резкое укорочение плеча, предплечья, бедра и голени. Наоборот, кисть и стопа заметно удлинены. Особо характерно наличие толстой кожистой перепонки, связывающей пальцы конечностей, в результате чего они преобразованы в ласты. При этом пальцы кисти снаружи не расчленены, хотя скелет их развит полностью. Пальцы задних конечностей (стопы) снаружи, наоборот, явственно видны. Когти развиты у разных видов неодинаково: у ушатых тюленей и моржей они малы, иногда рудиментарны; у настоящих тюленей когти развиты хорошо, особенно на пальцах передних ластов.
В отличие от своих ближайших родственников — наземных хищников — ластоногие используют конечности только для передвижения и не могут удерживать ими кормовые объекты. Наиболее полно для гребли приспособлены конечности настоящих тюленей. Только передними ластами они могут опираться о твердый субстрат, а задние у них вытянуты назад параллельно продольной оси тела. Наоборот, моржи и ушатые тюлени способны опираться о субстрат и задними ластами. У всех видов основным органом движения в воде служат задние ласты. Вытянутые при плавании назад, они то сближены между собой, то несколько раздвинуты и действуют как более или менее единое целое. Совершая в основном колебательные движения вправо и влево и одновременно несколько изгибаясь сверху вниз, задние ласты своей работой отчасти напоминают гребной винт. Именно они сообщают телу поступательное движение. При работе задних ластов их собственная мускулатура играет сравнительно небольшую роль и основную нагрузку несут мышцы задней части туловища, производящие энергичное колебательное движение позвоночника.
ОТРЯД ТРУБКОЗУБЫЕ (TUBULIDENTATA)
К отряду Трубкозубые (Tubulidentata) относится одно семейство (Orycteropodidae) с одним родом и единственным современным видом — африканский трубкозуб (Orycteropus afer) (рис. 219). Он распространен по всей Африке к югу от Сахары и на этом большом ареале образует 3 подвида, отличающихся окраской волос хвоста и размерами.
Свое название трубкозуб получил благодаря своеобразному строению зубов, в виде сросшихся дентиновых трубочек, без эмали и корней, с постоянным ростом. У эмбрионов закладываются многочисленные зубы, но у взрослых нет резцов и клыков, а появляется лишь 2 предкоренных и 3 коренных зуба в каждой половине челюсти. Строением зубов и мозга трубкозуб сходен с древними копытными. Его сходство с муравьедами из отряда неполнозубых чисто внешнее и связано с тем, что он, как муравьед, кормится главным образом термитами и муравьями.
Внешне трубкозуб напоминает свинью с длинной мордой, ослиными ушами и сильным мускулистым хвостом, откуда и произошло его местное название аардварк, что значит «земляная свинья». Длина тела трубкозуба 100—158 см, хвоста — 44—61 см, высота в плечах — 60—65 см, длина ушей 15—21 см, масса до 82 кг, чаще 50—70 кг. Мощные передние лапы с 4 пальцами; задние — пятипалые. Все пальцы, особенно на передних лапах, вооружены сильными когтями, которые по своему строению сходны с копытами. Толстая кожа покрыта редкими, щетинистыми волосами тусклого коричнево-серого или желто-серого цвета, у одного подвида волосы на хвосте белые. На ногах волосы обычно темнее, чем на туловище. На морде много жестких, длинных вибрисс. Особенно длинные они (до 2,5—2 см) около глаз и в круглых ноздрях, расположенных на свинообразном пятачке.
ОТРЯД ХОБОТНЫЕ (PROBOSCIDEA)
Ископаемые предки современных слонов начиная с эоцена населяли почти все континенты мира (за исключением Австралии и Антарктиды). Это были животные различной величины, нередко не превышавшие размеров лошади или такие гиганты, как плиоценовый южный слон высотой 5 м. Жили слоны преимущественно в лесах, саваннах и по речным долинам. Один вид — хорошо известный мамонт — при общем похолодании климата приспособился даже к суровым условиям тундры. К настоящему времени, однако, сохранилось лишь два вида, относящихся к одному семейству, но представляющих два самостоятельных рода.
В строении слонов особенно поражает хобот, аналогов которому нет в животном мире. Это не нос, как иногда думают, а верхняя губа, сросшаяся с носом. Благодаря сложной системе мускулов и сухожилий хобот обладает удивительной подвижностью и огромной силой. Вместе с тем особое хватающее устройство, которым он заканчивается, позволяет ему поднимать с земли мельчайшие предметы. В зоопарках, например, слоны легко собирают с пола монеты или пуговицы. Хобот для слона гораздо больше, чем рука для человека. С его помощью слоны дышат, добывают пищу, пьют, защищаются, выражают свои эмоции, избавляются от паразитов, купаются. Слон, лишившийся хобота (а такие случаи в Африке сейчас не редкость — слоны часто именно хоботом попадают в петли, расставляемые браконьерами), обречен на голодную смерть. Правда, они пытаются тогда пастись на коленях, но долго так жить не могут. Единственное время, когда слону при еде не нужен хобот — раннее детство: материнское молоко слоненок сосет прямо ртом. Кстати, млечные железы и два соска у слонихи располагаются на груди, между передними ногами, как у приматов, сирен и летучих мышей.
Замечательно устройство ноги слона: на подошве, под кожей, имеется особая желеобразная пружинящая масса, которая позволяет ступать совершенно бесшумно. Кроме того, когда слон опирается на ногу, подошва расширяется, как бы набухает, опорная поверхность увеличивается. Но стоит ему разгрузить ногу , как она принимает первоначальную форму. Поэтому слон легко преодолевает болотистые топи и не вязнет, даже погрузившись в трясину по брюхо.
Своеобразна и зубная система слонов. У них нет клыков. То, что обычно называют клыками, в действительности резцы, которых у слонов лишь пара в верхней челюсти. Кроме того, на каждой челюсти расположено по две пары предкоренных и одна пара коренных зубов с широкой жевательной поверхностью и невысокой коронкой. Всего 26 зубов.
Кожа слонов толстая, почти лишена волосяного покрова и изрезана частой сетью морщин. Только у мамонта тело было покрыто длинной и довольно густой рыжей шерстью.
ОТРЯД ДАМАНЫ (HYRACOIDEA)
Этот отряд объединяет одно современное семейство Procavidae, включающее 3 рода и около 10 видов.
Внешне даманы немного похожи на кролика, бесхвостого сурка или очень крупную сеноставку. Длина их тела от 30 до 60 см, хвоста нет, или он всего 1—3 см длиной, масса зверька от 1,5 до 4,5 кг. Морда короткая, с раздвоенной верхней губой; уши небольшие, у некоторых видов почти скрытые в шерсти; ноги короткие, но сильные. Передние лапы четырехпалые с уплощенными когтями, напоминающими копыта; задние лапы трехпалые, внутренний палец несет длинный изогнутый ноготь, а другие имеют копытообразные когти, как на передних лапах. На голых подошвах есть подушечки, а центральная часть свода подошвы может подниматься специальными мышцами при опоре ее на субстрат, что создает вакуум, и лапа присасывается к поверхности камня или ствола дерева. Железы на подошвах, выделяющие резиноподобный секрет, способствуют крепкому присасыванию подошвы к субстрату. Благодаря такому приспособлению даманы могут бегать, по вертикальным скалам и стволам деревьев вверх и вниз головой с большой ловкостью и быстротой. Молочных зубов — 28, постоянных — 34—38. Единственная пара верхних резцов с постоянным ростом лишена эмали на внутренней поверхности и напоминает резцы грызунов. Широкая диастема отделяет резцы от одной пары клыков (последние могут отсутствовать). Предкоренные (4/4) и особенно коренные (3/3) зубы сходны с зубами копытных. Желудок разделен на 2 отдела. На спине даманов есть крупное секретирующее железистое поле из 7—8 долей — спинная железа, значение которой неясно. У молодых она слабо развита, и у самок — меньше, чем у самцов. При испуге или возбуждении волосы, покрывающие железу (они иного цвета, чем волосы на всей спине), взъерошиваются, оголяя железу, из которой выделяется пахучее вещество.
Шерсть даманов густая, имеет мягкий подшерсток и жесткие ости. На теле (особенно на морде над глазами и на шее) — пучки длинных вибрисс. Окраска меха чаще коричнево-серая с разными оттенками, но на спинной железе всегда есть пятно светлых или черных волос.
Даманы населяют Африку, Юго-Западную Азию (Аравийский полуостров). Наземные виды даманов обитают на скалах, поднимаясь по склонам гор до высоты 4500 м над уровнем моря, либо среди камней и кустарников на сухих равнинах. Древесные даманы населяют леса. Они растительноядны, но большинство их ест также насекомых и их личинок. Даманы размножаются круглый год. Беременность их продолжается 7—7,5 месяца. Молодые рождаются хорошо развитые, зрячие, покрытые шерстью и скоро становятся самостоятельными.
ОТРЯД СИРЕНЫ (SIRENIA)
Сирены — чисто водные растительноядные млекопитающие тропических и субтропических широт.
Тело сирен веретеновидное, заканчивающееся горизонтальным хвостовым плавником округлой или грубо треугольной формы. Передние конечности превращены в плавники, а задних нет, имеются лишь рудименты бедра и таза. Спинного плавника тоже нет. Голова небольшая, подвижная, спереди притуплённая, без ушных раковин, с маленькими глазками, направленными чуть вверх. Парные ноздри на кончике морды плотно замыкаются клапанами и открываются только на момент выдоха — вдоха.
Внешне сходные с китообразными, сирены сохраняют более четкие черты наземных предков: грудные плавники их достаточно подвижны в плечевом и локтевом суставах; подвижны даже сочленения кисти, поэтому плавники лучше называть ластами. На теле растут одиночные щетинки, а на морде многочисленные вибриссы. Мясистыми подвижными губами сирены рвут водоросли и перетирают их уплощенными коренными зубами либо нёбной и нижнечелюстной роговыми пластинами (лишь у морских коров зубов нет совсем). В связи с растительноядностью резцы рано исчезают, кроме дюгоней, развивается емкий двухкамерный желудок с парой мешковидных придатков и длинный кишечник с крупной слепой кишкой. В скелете характерны толстые тяжелые кости и толстостенный массивный череп.
Флегматичные и беззащитные сирены скрытно живут среди густых водорослей вблизи морских берегов и в устьях тропических рек. Они обладают чутким слухом, а также, судя по крупным обонятельным долям мозга, хорошим обонянием. Глаза их покрыты студенистой массой. Однако зрение при жизни в зарослях водорослей или в мутных реках не может быть хорошо развитым. Выпуклые млечные железы, с одним соском каждая, расположенные на груди между ластами или почти под ними, набухают в период выкорма детенышей. Это обстоятельство, дополненное воображением моряков средневековья, послужило основой для рассказов о морских девах — сиренах. Кормящихся
детенышей они прижимают ластами к своей груди.
Сирены — вымирающая группа млекопитающих. Они произошли от наземных хоботных животных, на это указывает их ископаемый предок — эотерий. Сирены сохранили общие со слонами признаки: грудные млечные железы, смену коренных зубов в течение всей жизни, бивнеподобные резцы (у дюгоней), плоские, ногтевидные копытца на ластах ламантинов и т. п.
Отряд включает 3 семейства, одно из которых (морские коровы) истреблено 200 лет назад.
ОТРЯД НЕПАРНОКОПЫТНЫЕ (PERISSODACTYLA)
Этот отряд объединяет копытных, характеризующихся тем, что наибольшего развития у них достигает третий палец, через который проходит ось конечности, другие же пальцы развиты слабо или совсем отсутствуют. У непарнокопытных нет ключиц; желудок простой. Пара сосков только одна (в паховой области).
Появились непарнокопытные в нижнем эоцене, предками их были примитивные хищники — креодонты (Creodonta). Они быстро развивались, дав более 500 видов, объединенных в 12 семейств, из которых 9 вымерло в позднетретичное время и до наших дней дожили только три: тапиры, носороги и лошади, насчитывающие всего 16 видов. Дикие представители отряда населяют Азию, Африку и Южную Америку. Количество непарнокопытных за последнее время резко уменьшилось, в связи с чем в той или иной мере они находятся под охраной.
ОТРЯД МОЗОЛЕНОГИЕ (TYLOPODA)
Отряд мозоленогие (Tylopoda) объединяет верблюдов Старого Света и лам, или безгорбых верблюдов, Америки. До недавнего времени этих животных рассматривали как подотряд отряда парнокопытных, однако новейшие исследования показали, что мозоленогие настолько своеобразны, что должны быть выделены в особый отряд.
У мозоленогих нет копыт, а на двупалых конечностях имеются лишь тупые искривленные когти. Мозоленогие опираются не на концы пальцев, как копытные, а на совокупность фаланг пальцев. Нижняя поверхность ступни образована расширенной парной или непарной эластичной мозолистой подушкой. Эритроциты крови овальные, а не дисковидные, что отличает мозоленогих от всех остальных млекопитающих. Желудок трехкамерный, а рубец и сычуг особого строения и сильно отличаются от таковых у жвачных. Слепая кишка короткая. Плацента диффузная, примитивнее, чем у копытных; зародышевые оболочки (амнион, аллантоис) и желточный мешок резко отличаются по строению и развитию от соответствующих органов копытных. Строение половых органов мозоленогих имеет ряд особенностей, присущих только представителям этого отряда. В телосложении мозоленогих обращает на себя внимание не включенное в контуры туловища свободное бедро и очень длинная шея. Рогов нет. Зубов — 30—34. Мозоленогие появились в эоцене Северной Америки, откуда (в плиоцене или в плейстоцене) расселились в Азию, Северную Африку и Европу, а также в Южную Америку. В настоящее время 1 дикий вид (двугорбый верблюд) распространен в Центральной Азии и 2 диких вида (гуанако и викунья) — в Южной Америке. Кроме того, отряд включает домашнего одногорбого верблюда и домашних ламу и альпаку. Все они объединяются в 2 рода и одно семейство верблюдов (Camelidae), обитают в засушливых областях и горных странах, растительноядны; бегают иноходью, рождают одного (очень редко двух) детеныша.
ОТРЯД ПАРНОКОПЫТНЫЕ (ARTIODACTYLA)
Отряд объединяет копытных, характеризующихся наибольшим развитием III и IV пальцев, между которыми проходит ось конечности. Другие боковые пальцы (у большинства видов два) развиты слабо и расположены выше средних. Концевые фаланги всех пальцев одеты копытами. Ключицы нет. Желудок у большинства видов сложный, состоящий из нескольких отделов, редко простой.
Появились парнокопытные в нижнем эоцене, вероятно, из одного корня с непарнокопытными. Их предками были примитивные хищники креодонты (Creodonta). Уступая в середине третичного периода непарнокопытным по числу видов, в верхнетретичное время они достигли расцвета и стали преобладающей группой копытных, главным образом за счет представителей подотряда жвачных. В настоящее время парнокопытные населяют все материки, кроме Антарктики (в Австралию завезены человеком), и объединяют 9 семейств, около 85 родов и примерно 200 видов.
Систематика парнокопытных очень сложна, и количество семейств, подсемейств, родов и видов у различных авторов сильно отличается. В настоящее время принято подразделять отряд парнокопытных на два подотряда: нежвачные (Nonruminantia) и жвачные (Ruminantia).
ПОДОТРЯД НЕЖВАЧНЫЕ (NONRUMINANTIA)
Подотряд объединяет три современных семейства: свиней (Suidae), пекари (Tajassuidae) и бегемотов (Hippopotamidae), включающих 8 родов и 12 видов. Все нежвачные характеризуются массивным туловищем, короткими четырехпалыми конечностями (у пекари задние ноги трехпалые). Боковые (II и IV) пальцы развиты довольно хорошо и касаются земли. Животные не способны к быстрому бегу. Желудок относительно простой, состоит из одного, двух или трех отделов. Коренные зубы бугорчатые, клыки большие с постоянным ростом. Большинство представителей кормится как растительной, так и животной пищей.
ПОДОТРЯД ЖВАЧНЫЕ (RUMINANTIA)
Подотряд объединяет многочисленных парнокопытных животных, имеющих сложный желудок, состоящий из четырех (у оленьков из трех) отделов: рубца, сетки, книжки и сычуга. Пища из пищевода поступает в рубец, отсюда в сетку. При отрыгивании обильно смоченная слюной пища в виде жидкой кашицы поступает прямо в книжку, а оттуда в сычуг (у оленьков нет книжки). Коренные и задние предкоренные зубы имеют лунчатое строение. В верхней челюсти резцов нет. Пальцев 2 или 4, в последнем случае боковые пальцы малы и не касаются земли. Пястные и плюсневые кости (метаподии) третьих и четвертых пальцев слиты в одну кость — канон. Боковые метаподии и кости пальцев тонкие и короткие. У большинства жвачных копытных на лобных костях имеются рога.
Жвачные представляют собой самую разнообразную и процветающую современную группу парнокопытных. Появились они в виде оленьков еще в верхнем эоцене. В нижнем олигоцене появились олени, и в нижнем миоцене — полорогие, а в миоцене произошло чрезвычайно быстрое развитие всех жвачных.
Подотряд жвачных обычно разделяют на 6 семейств, 76 родов, объединяющих около 180 видов. Систематика надвидовых групп (надсемейств, подсемейств, подродов) очень сложна.
ОТРЯД ПРИМАТЫ (PRIMATES)
Из всех млекопитающих приматы (обезьяны и полуобезьяны) отличаются, пожалуй, наибольшим разнообразием и богатством форм. Но, несмотря на внешние различия, их объединяют многие общие черты строения тела, которые выработались в процессе эволюции в условиях древесного образа жизни.
Приматы обладают хорошо развитой пятипалой, хватательной конечностью, приспособленной к лазанью по ветвям деревьев. Для всех приматов характерно наличие ключицы и полное разделение лучевой и локтевой костей, что обеспечивает подвижность и разнообразие движений передней конечности. Большой палец подвижен и у многих видов может противопоставляться остальным пальцам. Концевые фаланги пальцев снабжены ногтями. У тех форм, которые обладают когтевидными ногтями или имеют коготь на отдельных пальцах, большой палец всегда несет плоский ноготь. При передвижении по земле приматы опираются на всю стопу.
Приматы в основном питаются смешанной пищей с преобладанием растительной, реже — насекомоядны. В связи со смешанным питанием желудок у них простой. Зубов четыре типа — резцы, клыки, малые (премоляры) и большие (моляры) коренные; моляры с 3—5 бугорками. Имеет место полная смена зубов — молочные и постоянные.
Значительные вариации отмечаются в размерах тела приматов — от маленьких мышиных лемуров до горилл ростом 180 см и выше. Волосяной покров густой, с подшерстком у полуобезьян, у большинства обезьян он развит слабо. У многих видов шерсть и кожа ярко окрашены, глаза карие или желтые. Хвост длинный, но имеются короткохвостые и бесхвостые формы.
Размножаются приматы круглый год, у самки рождается обычно один (у низших форм — иногда 2—3) детеныш. Как правило, приматы живут на деревьях, но есть наземные и полуназемные виды. Образ жизни приматов дневной, стадный, реже парный или одиночный; они обитают главным образом в тропических и субтропических лесах Африки, Азии и Америки, встречаются и в высокогорных областях.
Современных приматов известно около 200 видов. Они объединяются в 57 родов, 12 семейств и 2 подотряда — полуобезьяны (Prosimii) и обезьяны (Anthropoidea).
Согласно наиболее распространенной в настоящее время классификации в отряд приматов включаются тупайи, которые в качестве самостоятельного семейства Tupaiidae вместе с лемурами и долгопятами составляют подотряд полуобезьян. Через лемуров они связывают насекомоядных животных с приматами, напоминая о древнейших предках последних.
В нашем описании приматов мы придерживаемся наиболее распространенной классификации, принятой Дж. и П. Нейпирами (J. R. Napier and P. Н. Napier. A Handbook of Living Primates. London — New-York, 1967).
Полагают, что предками приматов были примитивные насекомоядные млекопитающие, весьма сходные с современными тупайями. Остатки их обнаружены в верхнемеловых отложениях Монголии. Эти древнейшие приматы, по всей вероятности, расселились из Азии в другие места Старого Света и Северной Америки, где дали основу для развития лемуров и долгопятов. Исходные формы обезьян Нового и Старого Света, вероятно, получили начало от примитивных долгопятов (некоторые авторы предками обезьян считают древних лемуров). Американские обезьяны возникли независимо от обезьян Старого Света. Предки их проникли из Северной Америки в Южную, здесь развивались и специализировались, приспосабливаясь к условиям исключительно древесной жизни.
Человек по многим анатомическим и биологическим чертам относится к высшим приматам, где составляет отдельное семейство людей (Hominidae) с родом человек (Homo) и одним видом — современный разумный человек (Homo sapiens recens).
ПОДОТРЯД ПОЛУОБЕЗЬЯНЫ (PROSIMII)
Этот подотряд включает наиболее примитивных представителей приматов — тупаий, лемуров, долгопятов.
Иногда тупаий и лемуров объединяют в группу стрепсириновых приматов, которые обладают ноздрями в виде запятых, открывающимися на оголенную часть кончика носа. Верхняя губа у этих приматов гладкая, неподвижная и без волос. Напротив, долгопяты и обезьяны составляют группу гаплориновых приматов, с ноздрями более округлыми, окаймленными стенками носа и открывающимися на подвижную, с развитым мускульным слоем, и обволошенную верхнюю губу.
Подотряд полуобезьян объединяет 6 семейств, 26 родов и около 50 видов с большим количеством подвидов.
ПОДОТРЯД ЧЕЛОВЕКОПОДОБНЫЕ ВЫСШИЕ ПРИМАТЫ (ANTHROPOIDEA)
В подотряд высших приматов входят обезьяны широконосые (Platyrrhina), или американские, и узконосые (Catarrhina), или африканско-азиатские. Это подразделение основывается на различии в строении их носа. У большинства обезьян Нового Света хрящевая носовая перегородка широкая, а ноздри широко разделены и обращены наружу. У обезьян Старого Света более узкая носовая перегородка и ноздри, как и у человека, обращены вниз. Но правильнее говорить о степени выраженности данного признака, так как толщина носовой перегородки и положение ноздрей у разных форм широконосых и узконосых обезьян может варьировать.
У всех обезьян на пальцах плоские ногти (у игрунковых — когтеобразные ногти); глаза обращены вперед, и глазница полностью отделена от височной ямы костной перегородкой; мозг, за исключением игрунковых, богат бороздами и извилинами; верхние резцы не разделены промежутком.
Для обезьян характерна редукция обонятельного аппарата и специальных осязательных органов на лице, где сохранились лишь три пары вибрисс — надглазничные, верхнечелюстные и подбородочные. С редукцией же вибрисс связано прогрессивное развитие осязательных кожных гребешков на ладонных и подошвенных поверхностях. Только у эдиповых игрунок и в большей степени у ночных обезьян на ладонях и подошвах еще встречаются участки кожи без гребешков. У остальных низших и высших обезьян ладонные и подошвенные поверхности покрыты кожными гребешками сплошь, как и у человека.
В подотряде 3 надсемейства: Ceboidea, Cercopithecoidea и Hominoidea.
Надсемейство Широконосые обезьяны (Ceboidea)
Животные мелких и средних размеров. Защечные мешки и седалищные мозоли отсутствуют. Хвост длинный, у некоторых цепкий, хватательный. Ведут древесный, дневной и стадный образ жизни. Обитают в Центральной и Южной Америке. Размножаются круглый год.
В составе американских обезьян 2 семейства: Callitrichidae (с 5 родами) и Cebidae (с 11 родами).
Надсемейство Собакоподобные, или Низшие узконосые, обезьяны (Cercopithecoidea)
Низшие узконосые обезьяны, в отличие от американских обезьян, обладают узкой носовой перегородкой и выступающим лицевым отделом черепа. Хвост варьирует от короткого (черный павиан, мандрил, дрил, свинохвостый макак) до длинного, никогда не бывает хватательным. Конечности равной длины или передние чуть короче задних; кисти короче стоп; самый длинный палец третий; большой палец кисти обычно способен противопоставляться остальным. Имеются седалищные мозоли — оголенные участки кожи с большой жировой подкладкой в области седалищных костей. Защечные мешки есть у всех, кроме азиатских тонкотелов.
К этому надсемейству относится одно семейство мартышкообразных обезьян (Cercopithecidae) с двумя подсемействами.
Надсемейство Человекоподобные приматы (Hominoidea)
Это надсемейство объединяет высших приматов — человекообразных обезьян, или антропоидов (гиббонов, орангутанов, горилл, шимпанзе), и человека.
Человекообразные обезьяны характеризуются относительно большим мозгом с хорошо развитыми бороздами и извилинами. Объем его у гиббонов 100—150 см3, у крупных человекообразных обезьян 350—600 см3. Лицевой отдел черепа сильно выступает вперед (прогнатизм); у самцов горилл и орангутанов на черепе имеются хорошо развитые гребни. У орангутана 12 пар ребер, у остальных антропоидов 13 пар. Передние конечности значительно длиннее задних. Хвост, защечные мешки и седалищные мозоли отсутствуют. Но у гиббонов седалищные мозоли есть. Червеобразный отросток (аппендикс) слепой кишки хорошо развит и достигает в длину 20—25 см. Горловые мешки, служащие для усиления звуков, хорошо развиты (кроме собственно гиббонов). Как и у человека, имеются все четыре группы крови.
В настоящее время большинство представителей антропоидов занесены в международную Красную книгу.