Engraulis japonicus Temminck & Schlegel, 1846
Японский анчоус (Rus),
Japanese anchovy
(Eng)
Синонимы:
Atherina japonica Houttuyn, 1782
Engraulis japonicus Temminck & Schlegel, 1846
Engraulis zollingeri Bleeker, 1849
Stolephorus celebicus Hardenberg, 1933
Рыбы открытого океана. Н.В.Парин 1998 г.
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Известно, что в период высокой численности японского анчоуса и японской скумбрии в 50-е годы оба вида были весьма обычными и в открытых водах северо-западной части Тихого океана, в которых предполагалось даже существование их особых «стад» — в действительности, по всей вероятности, псевдопопуляций.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Кроме хамсы, в советских водах
встречается еще японский анчоус, довольно
обычный в водах Приморья, но в
особенно большом количестве обитающий у
берегов Кореи и Японии. Эта рыба также
живет всего два-три года и достигает
в длину не более 16 см. Нерест идет
повсеместно в пределах обширного ареала,
простирающегося от Тайваня до Южного
Сахалина. Икра и мальки могут от
берегов Японии переноситься в потоке Куро-Сиво на северо-восток, так что отдельные
особи попадаются даже в удалении от
прибрежных вод. Во многих районах
Дальнего Востока японский анчоус
образует крупные скопления, составляя
значительную часть общей продукции
пелагических рыб.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
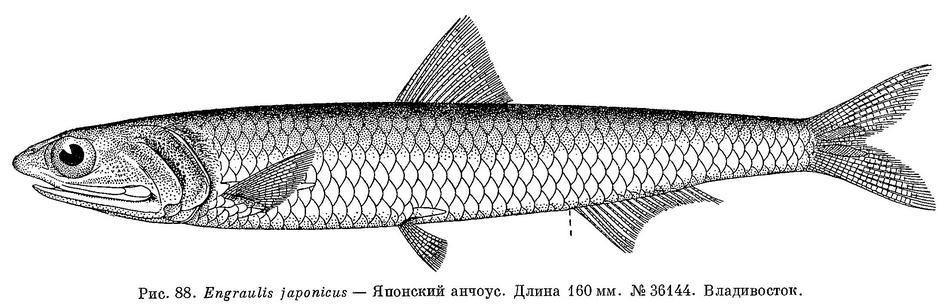
1. Engraulis japonicus (Houttuyn, 1782) — Японский анчоус (рис. 88).
Atherina japonica Houttuyn, Verh. Holland Maatsch. Haarlem, 20, 1782 : 340 (Япония).
Engraulis japonicus, Schlegel, Fauna Japonica, Poiss., 1846 : 239, pl. CVIII, fig. 3 (южная Япония). — Fowler, Bull. U. S. Nat. Mus., 100, 13, 1941 : 694. — Matsubara, Fish Morphol. a. Hierar., 1955 : 195. — Pасс, Промысловые рыбы СССР, 1949 : 144, табл. 35 (цветной рисунок).
Engraulis encrasicholus, Wang. Contr. Biol. Lab. Sci. Soc., China, 9, 1, 1933 : 8 (Чжифу).
D II 13, A 14—17, squ. 42—44, жаберных тычинок 34—35 (3 экз. из Тумынь-цзяна, 100 16791).
Характеристика дана при описании рода. (Тело частью цилиндрическое. Нет брюшных килевых чешуй. Верхнечелюстная не достигает жаберного отверстия. Жаберных тычинок на нижней половине первой жаберной дуги 36—49. Позвонков 46—47. Удлиненные чешуйки на основании хвостового плавника (alae) большие. Анальный плавник короткий, начало его основания далеко позади начала спинного. (Fowler, 1941).)
Используется в пищу и в качестве живой наживки для ловли бонито и настоящих тунцов. Нерест у берегов Японии с марта по июль. Икринки
эллиптической формы, мелкие (по длинной оси 1.08—1.60 мм, по короткой 0.55—0.72 мм), без жировой капли (Okada, 1955 : 45). Длина до 130 мм и более.
По биологии, морфологии и распределению анчоуса имеется значительное число работ, из которых отметим некоторые (Амброз, 1930; Бирман, 1958в; Введенский, 1949; Дружинин и Фридлянд, 1951; Казанова, 1959; Линдберг, 1935а; Моисеев, 1953а; Перцева-Остроумова, 1955; Пробатов, 1951, 1953; Расс, 1959а; Фридлянд, 1949; Храпкова, 1960) и некоторые японские (Hayashi a. Kondo, 1957; Hayashi a. Suzuki, 1957; Itazawa, 1954; Kondo, 1957; Kubo a. oth., 1954; Maekawa a. Yatsuyanagi, 1951; Shimomura, 1957; Shimomura a. Fukataki, 1957; Tamura, 1958; Tatsuki a. Furakawa, 1952).
Распространение. В Японском море анчоус обычен вдоль западного берега от Пусаня до Унги (Mori, 1952 : 33) и далее на север до зал. Петра Великого и зал. Ольги; вдоль берегов Японии до северной оконечности Хоккайдо (Honma, 1952 : 141). В годы потепления встречается у западного побережья Сахалина — поселок Антоново (Пробатов, 1951 :146) и может доходить даже до Александровска (Попов, 1933 : 140; Таранец, 1937а : 40). В Желтом море известен у западных берегов п-ова Корея (Mori, 1952 : 33) и вдоль побережья Китая на юг до Циндао (Чжан и др., 1955 : 51). Южная часть Охотского моря у мыса Ноторо (Hikita, 1952 : 3); берега западной Камчатки (Полутов, 1954 : 99). В 1929 г. проник до Авачинской бухты на Камчатке (Линдберг, 1935а : 47; Панин, 19366 : 149). Указывается для берегов Юго-Восточной Африки и о-вов Алдабра (Smith, 1950, 1955).
Наши экземпляры из Японского (Воньсань (№ 12591), зал. Петра Великого (№№ 16791, 16958, 17017, 22175, 31134, 31525), зал. Сяуху (№ 25178), зал. Ольги (№ 7861), Александровск-на-Сахалине (№№ 25501, 33205).) и Желтого морей (Чанчунь (Синьцзин, № 35582).), Петропавловска-на-Камчатке (№ 25110) и Кагосимы (№ 22391).