Trisopterus luscus (Linnaeus, 1758)
Pouting
(Eng)
Синонимы:
Gadulus luscus Malm, 1877
Gadus barbatulus Linnaeus, 1758
Gadus barbatus Linnaeus, 1758
Gadus bibus Lacepède, 1800
Gadus colias Gronow, 1854
Gadus luscus Linnaeus, 1758
Morhua barbata Fleming, 1828
Morhua lusca Fleming, 1828
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
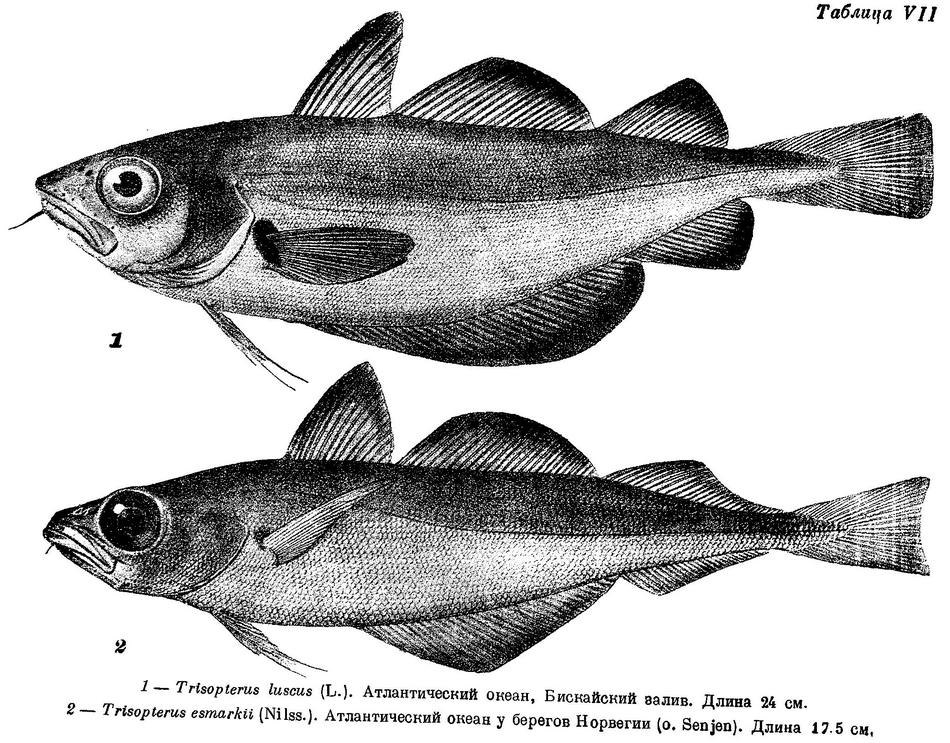
1. Trisopterus luscus (Linne) (табл. VII, XLII, LXX).
— Gadus luscus Lirme, Syst. nat., ed. X, 1758 : 252 (Oceano Europeo).
— Gadus barbatulus Linne, 1. c. : 252 (Oceano Europeo).
— Gadus bibus Lacepède, Hist. nat. poiss., II, 1800 : 365, 403 (Атлантический океан у берегов Европы).
— Morhua lusca Fleming, Hist. brit. animals, 1828 : 191.
— Morhua barbata Fleming, 1. c. : 191.
— Gadus colias Gronow ed. Gray, Cat. fish., 1854 : 131.
— Gadus minutus Steindachner, Sitzungsber. Akad. Wiss. Wien, math.-naturw. CI., LVII, 1, 1868 : 705 (ex parte: у берегов Пиренейского п-ова).
— Gadulus luscus Malm, Goteb. Bohusl. Fauna, 1877 : 482.
— Gadus luscus Williamson, 24th. Ann. Rept. Fish. Board Scotl., pt. 3, (1905), 1906 : 116 (остеология); Fage, Arch. Zool. Exper. Gener., (5), VI, 1911 : 257. (сравнение с близкими формами).
Распространение, биология. Smitt, Scand. fish., I, 1893 : 495, pl. LXXII, f. 2 (от южных берегов Скандинавского до Пиренейского п-ова, Средиземное море); Hjort, Rapp. Proces-Verb., X, 1909 : 60, f. 16 (нерест); Damas, ibid. : 27 (биология); J. Schmidt, ibid. : 76, 144 (нерест); Chevey, Rev. Trav. Off. Peches Marit., II, 1, 1929 : 73 (сводка данных по биологии, распространение); Tortoneze, Boll, pesca, piscicult., idrobiol., XI, 2, 1935 : 232 (у берегов Италии).
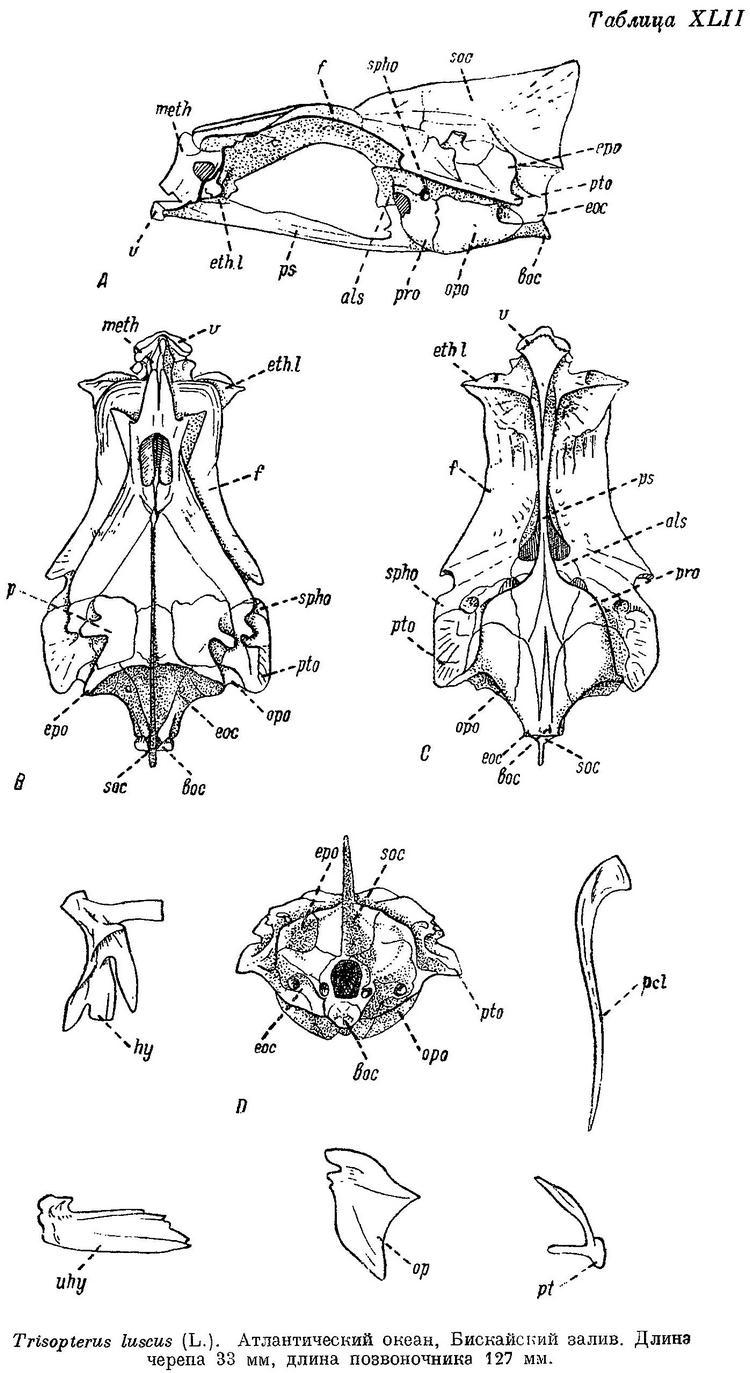
I D 11—14, II D 20—24, III D 18—20, I A 30—34, II A 19—22, sp. br. 18—23, vert. 47 (По Williamson'у, 48—49.). Mesethmoideum спереди спускается довольно круто, передний конец его почти над передним концом vomer. Передний конец frontalia почти над передним концом ethmoidalia lateralia. В межглазничной области череп широкий, около 3—4 раз в длине его. Рот нижний. Спинные и анальные плавники соприкасаются, анальные при основании часто друг с другом слиты. 1-й анальный плавник длинный, 35.7—37.8%: длины тела, начало его расположено под началом или несколько позади 1-го спинного плавника, но не далее его середины. Тело высокое, боковая линия в передней части тела значительно изогнута. В процентах к длине тела: антеанальное расстояние 29.2—33.8, антедорсальное расстояние 26.6—29.3, длина грудного плавника 15.9—19.3, длина брюшного плавника 12.3—15.7, основание 1-го спинного плавника 11.4—14.0, основание 2-го спинного 23.9—27.7, основание 3-го спинного 13.7—15.8, основание 1-го анального 35.7—37.8, основание 2-го анального 15.4—17.1, высота 1-го спинного 16.2—17.4, высота 1-го анального 12.9—14.8, высота хвостового стебля 4.9—5.9, длина его 10.2—12.8, длина головы 22.7—23.8. В процентах к длине головы: длина рыла 26.9—28.8, продольный диаметр глаза 28.1—30.9, длина верхней челюсти 40.0—43.4, длина нижней челюсти 46.2—49.1, ширина лба 17.9—18.1, длина усика 23.1—28.6. Верхняя часть тела красновато-бурая, нижняя часть светлее, с голубовато-серым оттенком, переходящим на брюхе в беловатый; бока серебристые, боковая линия золотисто-желтая. Почти все тело покрыто мелкими черновато-коричневыми точками. Плавники голубовато-черные, к вершине темнее. Поперек тела иногда проходят поперечные полосы беловато-серого цвета у верхнего края основания грудного плавника черное пятно. Длина обычно 15—20 см, редко превосходит 30 см, но может достигать и 45 см длины, (11 экз.).

Распространение. Европейские берега Атлантического океана от самых южных берегов Марокко и даже южнее (у мыса Бланко уже нет), в Бискайском заливе, наичаще вокруг Великобритании и Ирландии, в Ламанше, у западных берегов Шотландии, в Скагерраке, на восток до мыса Скаген и южных берегов пров. Bohuslan. У Исландии и Фарерских о-вов нет. Единично в западной части Средиземного моря вдоль европейских и африканских берегов (Марокко и Алжир, Fage, l. c. : 267).
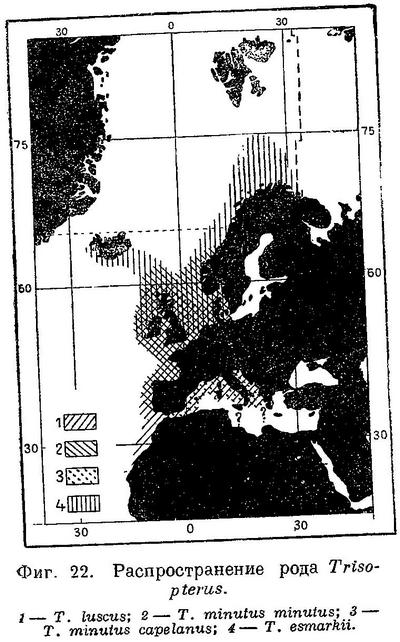
Биология изучена мало. Держится обычно на глубине 250—300 м, реже встречается и на неглубоких местах у берегов. Нерестится главным образом в Ламанше, где встречаются наибольшие количества икры и пелагических личинок, а также в Бискайском заливе, у западных и северных берегов Великобритании, вокруг Ирландии, в небольшом количестве в южной части Северного моря, в Средиземном море. Нерест происходит на сравнительно небольших глубинах в пределах 100-метровой изобаты, преимущественно на глубине 50—100 м, при довольно высокой температуре (не ниже 8—9° С) и нормальной океанической солености — 32.00—35.35‰. Главное время нереста весна, март—апрель, в небольшом количестве нерестится и летом в июне—июле и даже августе; начало — с января. Половой зрелости достигает в 1 год. В 1 год имеет длину 21—25 см, в 2 года — 23—27 см, в 3 года — 28—33 см, в 4 года до 30—31 см. Питается моллюсками, ракообразными, мелкой рыбой.
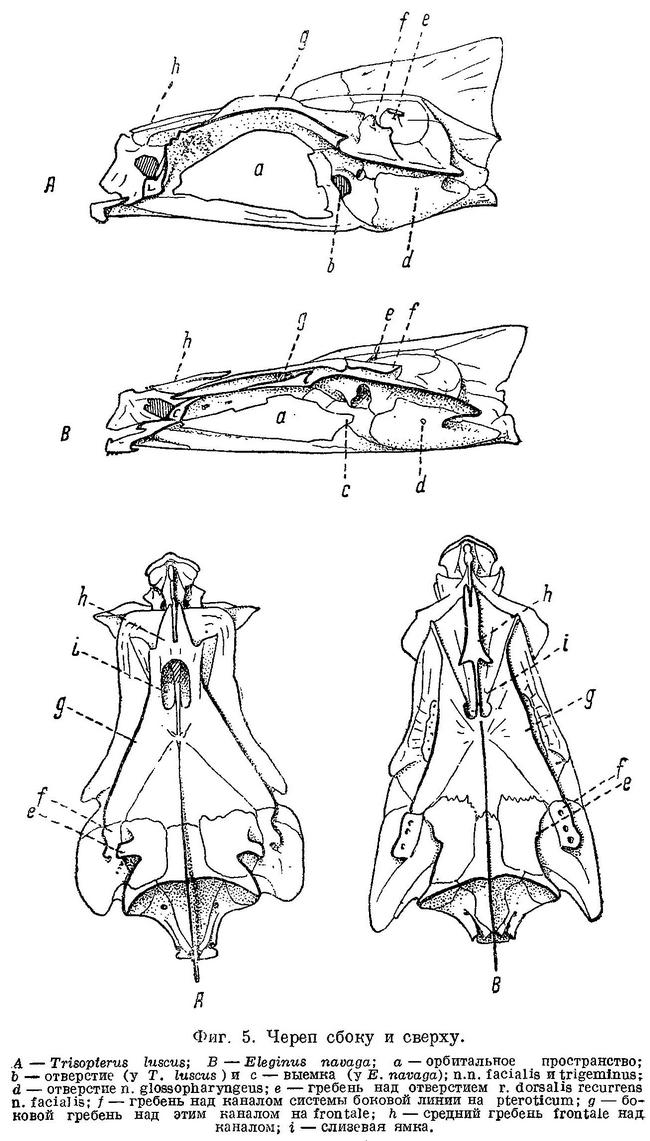
N. facialis выходит из черепа у тресковых вместе с n. trigeminus через отверстие или различной глубины выемку в передней части prooticum сбоку черепа (фиг. 5,b,с). У Brosme brosme нерв этот выходит через отверстие, а у следующего за ним по ряду Gaidropsarus mediterraneus — через глубокую выемку в prooticum, края которой настолько сближены, что почти замыкают ее спереди. У Ciliata mustela и далее следующих за ним в ряду рыб выемка становится спереди более широкой и менее глубокой. У Lota lota выемка так же глубока и края спереди сужены почти так же, как у Gaidropsarus mediterraneus, а у всех видов Molva и Merluccius n. facialis из черепа выходит опять через отверстие. Через отверстие n. facialis выходит и у следующих далее в ряду видов рода Trisopterus. При этом ширина костной перемычки у Trisopterus luscus несколько шире, чем у Trisopterus minutus, а у последней шире, чем у Trisopterus esmarkii. Odontogadus merlangus и далее следующие за ним представители семейства имеют в prooticum выемку, которая последовательно становится спереди более широкой и менее глубокой. У обеих форм Gadiculus argenteus выемки почти нет. Далее, у видов родов Eleginus, Microgadus, Boreogadus, Arctogadus и Theragra выемка постепенно углубляется, а края ее становятся более сближенными. Micromesistius имеет сбоку черепа отверстие. Таким образом, по этому признаку ряд тресковых, расположенных по степени дифференциации их непарных плавников, разделяется в свою очередь на два параллельных ряда, соответственно делению семейства на подсемейства. Такой же закономерности подчиняется изменение и некоторых других признаков.
Сверху черепа тресковых на каждом parietale расположено по отверстию, через которое выходит r. dorsalis recurrens n. facialis (фиг. 5,е и рис. В на табл. XXI—LXVII). У одних тресковых каждое из этих отверстий прикрыто у каждого вида в различной степени развитыми гребнями; у других гребень не развит, и отверстие сверху открыто. У Brosme brosme и далее следующих за ним в ряду Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius гребни, прикрывающие эти отверстия, имеются. У Raniceps ranius и всех видов родов Phycis и Urophycis отверстия r. dorsalis recurrens n. facialis не прикрыты, так как гребни у них имеют вид едва заметных выростов, расположенных несколько сзади отверстий. У Lota, всех видов Molva и Merluccius гребни развиты и прикрывают отверстия. В пределах подсемейства Gadinae гребни на parietalia развиты сильнее. У всех видов родов Trisopterus, Odontogadus, Pollachius, Gadus и Melanogrammus гребни развиты, размеры их уменьшаются от Trisopterus к Melanogrammus. Нет гребней у Gadiculus. Далее гребень есть, постепенно увеличиваясь к концу ряда, у Eleginus, Microgadus, Boreogadus, Arctogadus, Theragra и Micromesistius, причем у одних особей Eleginus navaga гребни есть, у других они не развиты.
N. glossopharyngeus (Stensiö, 1921 : 155) выходит из черепа через небольшое отверстие на opisthoticum (фиг. 5,d и рис. А на табл.XXI—LXVII). Величина этого отверстия, изменяясь в пределах ряда, подчиняется той же закономерности. У Brosme brosme отверстие почти не заметно. У Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius, Raniceps raninus оно небольшое, у видов родов Phycis и Uropkycis несколько больше, у Lota и Molva большое и у Merluccius вновь небольших размеров. Далее, у всех видов рода Trisopterus отверстие едва заметно, размеры его постепенно увеличиваются до Gadus и Melanogrammus и, начиная с Gadiculus, уменьшаются вновь.
Боковая линия у тресковых образует на голове с каждой стороны каналы: предкрышечно-нижнечелюстной, подглазничный и надглазничный с короткой комиссурой, соединяющей правый и левый каналы. Сама боковая линия имеет на голове надвисочную часть, над которой образуется окостенение из 4—5 косточек ossa tabularia (= supratemporale). Каждый канал системы боковой линии на голове прикрыт сверху почти на всем своем протяжении гребнем тех костей, на которых эти каналы расположены (frontale, pteroticum, sub- и praeorbitale, nasale, praeoperculum, tabulare). Среди этих гребней наиболее существенное значение имеет средний гребень frontale (фиг. 5, h) и боковые гребни frontale и pteroticum (фиг. 5, f и g). Межглазничная комиссура надглазничного канала расположена в особом углублении frontale — слизевой ямке (фиг. 5, i). Слизевая ямка у одних представителей Gadidae спереди замкнута сросшимися краями боковых передних гребней над каналами системы боковой линии на frontale, у других широко открыта, так как края боковых и средних гребней frontalia укорочены и не срастаются впереди слизевой ямки. У Brosme brosme края боковых и средних гребней frontalia настолько сближены, что почти замыкают спереди небольших размеров слизевую ямку. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopis cimbrius слизевая ямка спереди открыта, очень небольших размеров. Raniceps raninus имеет слизевую ямку спереди открытой. У одних видов родов Phycis и Urophycis слизевая ямка спереди открыта, у других закрыта; оба эти рода представляют в этом отношении небольшое отклонение от общего правила, о причинах чего будет сказано ниже. У Lota lota слизевая ямка спереди почти замкнута сближенными краями боковых и средних гребней frontalia, которые у видов рода Molva, срастаясь, образуют хорошо замкнутую ямку. Слизевая ямка у всех видов рода Merluccius представляет собой обширное углубление посередине frontalia и имеет несколько уклоняющийся от обычного для тресковых вид. Слизевая ямка замкнута далее, начиная с Trisopterus и кончая Gadiculus. У всех далее следующих в ряду видов слизевая ямка спереди открыта и замкнута вновь у стоящих в конце его Theragra chalcogramma и Micromesistius poutassou и Micromesistius australis. При этом следует отметить, что закрытая или открытая спереди слизевая ямка обычно свойственна всем видам любого рода тресковых, за исключением, как указывалось выше, родов Phycis и Urophycis. Кроме того, есть один вид — Eleginus navaga, — у которого сближенные края боковых и средних гребней frontalia иногда бывают сращены с одной стороны, а на одном из исследованных черепов и с обеих сторон. Таким образом, эта особенность находится в неустойчивом состоянии в первом случае в пределах рода, во вторых — в пределах вида, стоящих в середине каждого из параллельных рядов.