Trachurus novaezelandiae Richardson, 1843
Новозеландская ставрида (Rus),
Yellowtail horse mackerel
(Eng)
Синонимы:
Trachurus mccullochi Nichols, 1920
Trachurus novaezelandiae Richardson, 1843
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus novaezelandiae Richardson - новозеландская ставрида
Описание. D I, VIII+I, 30-35; А II+I, 25-28. Щитков в боковой линии 74-85; колючих щитков - 40-45. Голова составляет 25,0-27,0% длины тела. Верхняя (спинная) ветвь боковой линии оканчивается под основанием начала второго спинного плавника. Щитки в боковой линии большие: высота наиболее высоких щитков передней части боковой линии 22,5-27,5 % длины головы; высота наиболее высоких щитков задней части боковой линии 25,0-28,5% той же длины. Кили на щитках хорошо развиты. Задняя часть боковой линии составляет 38,5-41,0% длины тела; хорда дуги передней (кривой) части боковой линии 38,0-39,0%. Длина грудных плавников 25,0-26,0% длины тела. Тело овальное, его высота 20,0-22,0% длины тела. Горизонтальный диаметр глаза 22,0-28,0% длины головы. Имеется жировое веко. Верхняя челюсть 36,0-40,0% длины головы. Первый колючий щиток лежит под основанием 10-го луча второго спинного плавника. Расстояние от вершины рыла до вертикали начала основания брюшных плавников составляет 31,0-33,0% длины тела. Расстояние от вершины рыла до вертикали начала основания первого спинного плавника 32,0-34,0% длины тела. Расстояние от вершины рыла до вертикали начала основания анального плавника - 55,0-57,0% длины тела. Отношение расстояния между последним и предпоследним лучами к расстоянию между предпоследним и предшествующим ему лучами анального плавника (третьего сзади) равно 1,42-1,70 раза [Алеев, 1957а; Носов, 1977; Stephenson, Robertson, 1976].
Сравнительные замечания. До недавнего времени Trachurus novaezelandiae считали синонимом Trachurus declivis [Линдберг и др., 1980). Однако анализ морфологических признаков и результаты анализа исследований гемоглобина, несомненно, показывают, что Trachurus declivis и Trachurus novaezelandiae представляют собой разные виды [Gauldie et al., 1977; Richardson, 1982].
Эти два вида легко различимы по месту окончания верхней ветви боковой линии: у Trachurus novaezelandiae ставриды она оканчивается примерно под 1-4-м лучом, а у Trachurus declivis - под 8-9-м лучом второго спинного плавника. Кроме того, передняя часть боковой линии у Trachurus declivis прямая, а у Trachurus novaezelandiae изогнута.
Распространение. Новозеландская ставрида обитает на шельфе Новой Зеландии и встречается в водах верхней части склона. Встречается на значительной площади шельфа от м.Рингс на севере до о-ва Стьюарт на юге, обычна во все сезоны года в водах о-ва Северный и северной оконечности о-ва Южный. В небольших количествах этот вид встречается также на банках в Тасмановом море. Область распространения новозеландской ставриды весьма значительна и включает воды различной структуры - от субтропических на севере до смешанных вод субтропической конвергенции на юге ареала. Северная граница распространения совпадает с изотермой 13,5°С, а южная - 12,5°С.
Биология. Наиболее значительные скопления новозеландской ставриды образуются в водах о-ва Северный и северной оконечности о-ва Южный. Здесь расположены основные нерестилища этого вида. Самые северные районы ареала новозеландской ставриды являются местом обитания теплолюбивой молоди, но нерест в этих районах не происходит [Носов, Платошина, 1975].
Взрослая часть популяции, по многолетним наблюдениям, представлена в уловах особями длиной от 12 до 52 см в возрасте от 2 до 16 лет, максимальная известная длина 55 см [Носов, Шурунов, 1975; James, 1975].
Размерно-возрастной состав новозеландской ставриды в каждом конкретном районе ежегодно меняется в зависимости от урожайности поколений, а также из-за разнокачественности исследуемых выборок, состоящих из групп особей, различающихся миграционной активностью. Последнее обстоятельство в известной степени определяется межгодовой изменчивостью условий среды, в которой находится популяция. Средняя длина и средний возраст ставриды, вычисленный по суммарному многолетнему ряду, соответственно 33,8 см и 7,1 года. Основу промысловой части популяции (около 61 %) составляют особи длиной 25-37 см (4-8 лет), доля наиболее крупной (более 38 см) новозеландской ставриды 9-16-летнего возраста - примерно 30% общей численности популяции [Носов, Шурунов, 1975].
Новозеландская ставрида относится к порционно-нерестующим рыбам. Созревание половых продуктов у различных размерно-возрастных групп происходит неравномерно, поэтому нерестовый период растянут. Размножается новозеландская ставрида в весенний и весенне-летний период, т.е. в теплое время года. Массовый нерест наблюдается в наиболее благоприятный летний период. К началу нереста косяки ставриды, обитающей на окраинных участках шельфа (глубина 100-150 м), начинают мигрировать в сторону берега к местам нерестилищ. Первой, как правило, для нереста подходит крупная ставрида. Установлено, что репродуктивный район новозеландской ставриды ограничен водами о-ва Северный и северной оконечности о-ва Южный. Основные нерестилища расположены в мелководных частях заливов западного и восточного побережий о-ва Северный. Для нереста ставрида предпочитает хорошо прогретые воды с температурой 16-23°С (оптимальная 18-20°С ). Более южные районы ареала малопригодны для нормального развития икры, и размножение ставриды в них не происходит. Нерест продолжается в течение восьми месяцев [Носов, Платошина, 1975].
Нерестовая часть популяции новозеландской ставриды состоит из большого числа возрастных групп: у самок из 10-11-летних особей, у самцов - из 13-14-летних. Многовозрастным является не только остаток (рыбы повторно нерестующие), но и пополнение. В половозрелой части популяции ставриды соотношение полов обычно близко 1:1. Среди молоди наблюдается некоторое преобладание самцов, массовое созревание которых происходит в более раннем возрасте и более быстрыми темпами. В возрасте 13-14 лет обнаруживается значительное, а в возрасте 15-16 лет абсолютное преобладание самцов. Однако количество особей 15-16-летнего возраста составляет всего 0,5-0,6%, поэтому в нерестовых скоплениях в целом сохраняется соотношение полов 1:1.
Специалисты Тихоокеанского научно-исследовательского института рыбного хозяйства и океанографии (ТИНРО) считают, что за один нерестовый сезон одна особь новозеландской ставриды может трижды принимать участие в нересте. Средняя плодовитость самок составляет 430 тыс. икринок [Носов, Платошина, 1975].
Новозеландская ставрида наиболее интенсивно растет на первом году жизни. Сеголетки от ранненерестующих родителей (нерест в ноябре - декабре) вырастают к зиме южного полушария (июль - август) в среднем до 10 см, а от поздненерестующих (нерест в феврале - марте) - до 4 см.
Следовательно, существенные различия в темпе роста сеголетков закладываются в связи с очень большой продолжительностью нереста. Кроме абсолютной разницы во времени, составляющей около 90-120 суток, различия в темпе роста сеголетков в значительной степени определяются условиями среды в течение лета. Наиболее благоприятные условия складываются для мальков старших генераций. Их переход на экзогенное питание совпадает с периодом, когда в репродуктивной части ареала начинают усиленно развиваться мелкие формы кормового фито- и зоопланктона [Носов, 1977]. В значительно худших условиях находятся сеголетки позднего нереста, когда с осенним похолоданием количество и разнообразие кормовых форм планктона начинает уменьшаться [Бархатов, 1976]. Разница в скорости роста, обозначившаяся на первом году жизни, с возрастом не только не сокращается, но даже увеличивается. Быстрорастущие особи становятся в массе половозрелыми на два года раньше тугорослых.
Для характеристики естественной смертности новозеландской ставриды построена кривая возрастного состава, которая показывает, что преобладающей группой являются шестигодовики (17,1%); с возрастом количество рыб уменьшается: семигодовики составляют 13%, восьмигодовики - 10,5% и т.д. В необлавливаемой популяции это обусловлено естественной смертностью особей. Разница между численностью каждой данной и предшествующих возрастных групп характеризует естественную убыль в этом возрасте. Общий характер изменения численности с увеличением возраста наиболее достоверно описывается уравнением параболической кривой [Соколовский, 1973; Носов, 1977; Аксютина, 1986]:
у = 0,01912х2 - 0,2732х + 1,0136
Молодые особи более теплолюбивы и концентрируются в более северных водах Новой Зеландии. По мере движения с юга на север доля крупных рыб в уловах уменьшается, а мелких увеличивается: в зал. Ахипара мода длины составляет 15,0 см, в зал. Пленти - 22,7 см, в зал. Хок - 25,0 см, в зал. Таранаки - 28,2 см, в зал. Тасман - 39,9 см, в зал. Карамса - 45,3 см [Шунтов, 1970].
Анализ распределения новозеландской ставриды во все сезоны убеждает, что она активный мигрант. Перемещения ее на шельфе и вдоль побережий хорошо прослеживаются как визуально (поверхностные нерестовые косяки), так и при сопоставлении уловов в смежных районах. Северные участки новозеландского шельфа лежат в зоне более теплых вод, но с обедненной кормовой базой. Южные районы холоднее, но зато продуктивнее, особенно в теплое время года. В связи с этим новозеландская ставрида использует южные районы для нагула, совершая весьма протяженные миграции [Носов, 1975а,б].
Все основные нерестовые скопления и главные миграционные пути наблюдаются в районе обширного мелководья между Северным и Южным островами. Именно в этот район мигрирует значительная часть ставриды из северных участков шельфа западного и восточного побережий о-ва Северный после вымета первой порции икры [Носов, Платошина, 1975; Носов, Шурунов, 1975]. С началом зимнего охлаждения южных районов ареала крупная новозеландская ставрида из мест нагула в продуктивных водах мигрирует на север, образуя основные скопления на обширном участке между Северным и Южным островами. В холодные годы новозеландская ставрида заканчивает нагул у о-ва Южный в мае и быстро мигрирует в теплые северные воды. Основная ее масса через район зал. Тасман идет в мае - июне. В теплые годы ставрида в мае и июне продолжает нагул; миграции на север происходят постепенно до конца августа. В годы промежуточные по гидрологическим условиям ставрида мигрирует во второй половине июня - начале июля. К этому времени снижается интенсивность ее питания и она распределяется в широком диапазоне глубин - от прибрежной зоны до материкового склона, образуя концентрации наибольшей плотности в слое 50-125 м. В этот период тяготение ставриды к более глубоководным участкам объясняется двумя основными факторами - гидрологическим и кормовым. На мелководье наблюдается быстрое и сильное охлаждение вод. Для относительно теплолюбивой ставриды такие условия неблагоприятны. Кроме того, в прибрежной зоне значительно снижается (по сравнению с летним периодом) биомасса планктона, тогда как в присклоновой части шельфа и над материковым склоном в это время наблюдаются значительные скопления макропланктона, что создает благоприятные кормовые условия для ставриды, продолжающей интенсивно питаться зимой.
Летом наиболее плотные скопления ставриды образуются в верхних и средних слоях воды над шельфом, в особенности на глубине 25-50 и 75-100 м. В верхнем из них происходит массовый нерест, а глубже концентрируются рыбы, отходящие с нерестилищ на нагул. К середине января, после завершения нереста, часть крупной новозеландской ставриды мигрирует на юг в район зоны субтропической конвергенции для нагула. Другая часть крупной ставриды остается в репродуктивной зоне у о-ва Северный, смещаясь с мелководья в нижние участки шельфа. Зависимость короткопериодной изменчивости вертикальных миграций ставриды от синоптических условий в летний сезон представляет полную противоположность характеру ее поведения зимой. Поверхностные косяки крупной и средней половозрелой ставриды летом при ухудшении погодных условий стремятся уйти в придонные горизонты. Мелкие неполовозрелые рыбы в этих условиях рассредоточиваются в пелагиали [Носов, 1970; Шунтов, 1975].
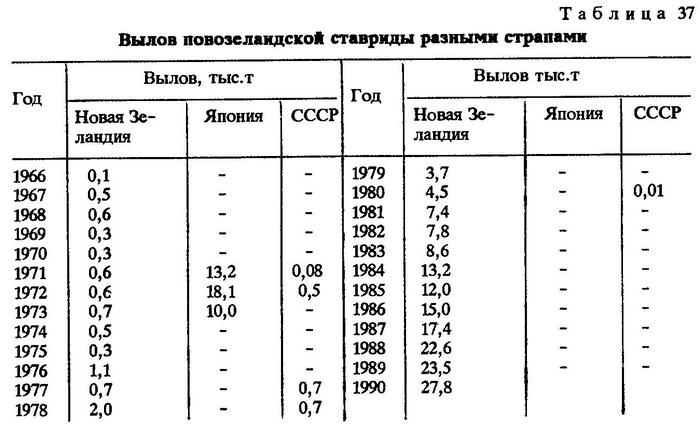
Местный специализированный промысел новозеландской ставриды развит слабо. Имеются сведения о добыче этого вида японскими рыболовными судами. Ежегодный вылов ставриды в водах Новой Зеландии колеблется от 0,1 до 19,2 тыс.т и, несмотря на высокую численность вида, занимает одно из последних мест среди добываемых Новой Зеландией морских объектов (табл. 37) [Yearbook..., 1980, 1984, 1985].
Добывают новозеландскую ставриду тралами и кошельковыми неводами. Наиболее благоприятным для промысла является осенне-зимний период, когда рыба более упитана и ее распространение не связано с прибрежной зоной.
Для количественной оценки промыслового запаса новозеландской ставриды на основе результатов контрольных тралений в экспедициях ТИНРО был использован метод изолиний [Аксютина, 1968], дающий возможность при получении среднего взвешенного улова за одно часовое траление по зонам разной мощности скоплений вычислить общую промысловую массу рыбы. Уловистость тралов принимали за 0,2. Средняя биомасса, по данным траловых уловов 1974 г., вычисленная способом площадей, составила 594 тыс.т [Носов, 19756]. Другим методом [Соколовский, 1973] определена абсолютная численность (биомасса) популяции ставриды в 760 тыс.т. Отсюда следует, что при соблюдении норм вылова с учетом естественной убыли по возрастным группам при 15-18% изъятия (при данном уровне запаса) ежегодно можно вылавливать до 140 тыс.т новозеландской ставриды.
В целом запасы новозеландской ставриды в районе Новой Зеландии, по оценкам международных экспедиций, весьма значительны, однако используются они далеко не полностью, поэтому в настоящее время воды, омывающие Новую Зеландию, рассматриваются как один из районов, где возможно расширение морского рыболовства.