Trachurus picturatus (Bowdich, 1825)
Океаническая ставрида, канарская океаническая ставрида, азорская океаническая ставрида, западноафриканская шельфовая океаническая ставрида (Rus),
Blue jack mackerel
(Eng)
Синонимы:
Caranx cuvieri Lowe, 1838
Caranx suareus Risso, 1833
Seriola picturata Bowdich, 1825
Trachurus fallax Capello, 1867
Trachurus melanosaurus Cocco, 1839
Trachurus rissoi Giglioli, 1880
Рыбы открытого океана. Н.В.Парин 1998 г.
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Trachurus symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955г. в удалении до 1100 миль, а в 1972 г.— до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставри-ды (Trachurus symmetricus murphyi) в 40-х широтах южного полушария отмечен в 1985 г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45—60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2—3 лет). Таким образом, ати факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций), которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300—400 тыс. т в 1974—1976 гг. до 1100—1300 тыс. т в 1978—1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.
В целом возникновение новых самостоятельных популяций неритических видов в результате выселения в открытый океан представляется довольно маловероятным событием, но именно этим способом, по-видимому, возникли «баночные» популяции некоторых рыб, в частности ставриды (Trachurus picturatus) и рыбы-сабли (Lepidopus caudatus), на подводных поднятиях северо-восточной Атлантики в районе Азорских островов.
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus picturatus (Bowdich) - океаническая ставрида (рис.10)

Описание. D I, VIII, I+28-37, в среднем 33,0; А II, I+23-34, в среднем 27,9. L.l. 90-110, в среднем 100,1. Dll 3DII-10DII, наичаще 6DII-8DII. Жаберных тычинок на первой дуге 49-63, в среднем 56,7; на нижней части дуги 35-43, в среднем 40,0. Щитки боковой линии сравнительно мелкие. Высота наиболее высоких щитков передней (кривой) части боковой линии 15,7-18,2, в среднем 16,6% длины головы; высота наиболее высоких щитков задней (прямой) части боковой линии 15,7-19,1, в среднем 17,2% той же длины. Наиболее высокие щитки прямой части боковой линии несколько выше щитков ее кривой (передней) части или, реже, равны им по высоте. Кили на щитках хорошо развиты, шипы на килевых щитках сильные. Прямая (задняя) часть боковой линии, несущая киль, короткая; хорда дуги кривой части боковой линии 0,8-1,1, обычно 0,8-1,0 раза укладывается в длине ее прямой части. Ответвление каналов боковой линии на верхней стороне головы и на спине развиты очень сильно, образуя густую сеть. Тело прогонистое, кругловатое (у взрослых почти круглое), голова вытянута; наибольшая высота тела 16,0-29,0 в среднем 17,8% длины тела до конца средних лучей хвостового плавника. Глаз крупный, диаметр его 27,1-34,0, в среднем 31,1% длины головы. В процентах длины тела до конца средних лучей хвостового плавника: расстояние от вершины рыла до вертикали начала оснований брюшных плавников 29,0-32,6, в среднем 30,3; расстояние от вершины рыла до вертикали начала основания первого спинного плавника 32,0-35,0, в среднем 34,5; расстояние от вершины рыла до вертикали основания первого луча анального плавника 51,2-54,5, в среднем 52,6. Последние лучи второго спинного и анального плавников по сравнению с предпоследними значительно увеличены и заметно отставлены от остальной части плавника, хотя и соединены с ней перепонкой; расстояние между основаниями последнего и предпоследнего лучей анального плавника в 1,7-2,1 раза превосходит расстояние между основаниями предпоследнего и предшествующего ему (третьего сзади) луча того же плавника. Грудные плавники длинные, у взрослых оканчиваются близ вертикали основания первого луча анального плавника (слегка не доходят до этой вертикали или слегка заходят за нее). Спина темная, голубовато-зеленоватая, низ тела серебристый; на жаберной крышке черное пятно. Длина до 60 см [Алеев, 1957а; Шабонеев, 1973].
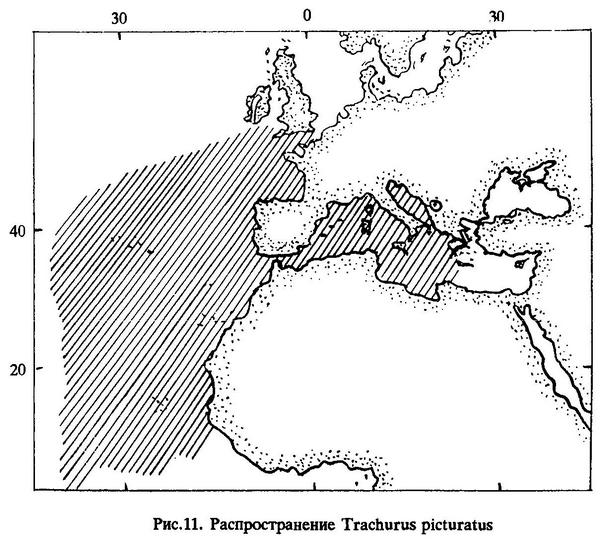
Распространение. Северо-восточная часть Атлантического океана от Бискайского залива до Азорских островов и берегов западной Африки; Средиземное море, за исключением его самых восточных частей. Океаническая ставрида в массовых количествах встречается как на шельфе, так и в открытой части Атлантического океана [Gail, 1955; Шабонеев, 1980] (рис.11).
В районе к северо-востоку от экономической зоны Азорских островов в августе 1982 г. в научно-поисковом рейсе РТМС "Патриот" на большой площади была обнаружена и облавливалась молодь океанической ставриды длиной 12-17 см, уловы которой достигали 0,5 т за траление.
Скопления океанической ставриды во все сезоны года встречаются на банках Геттисберг, Жозефин, Ампер, Сен, Дасия, Консепсьон и других, которые расположены примерно в 200-250 милях западнее Португалии, и на подводных горах системы Азорского архипелага.
Trachurus picturatus, обитающий у берегов южной Африки, представляет, по-видимому, особый подвид, поскольку в приэкваториальных районах восточной Атлантики Trachurus picturatus неизвестен, что указывает на географическую изоляцию североатлантических популяций от южноатлантических. О степени морфологических отличий южного подвида за неимением достаточных материалов в настоящее время судить затруднительно. Экземпляр от мыса Доброй Надежды, хранящийся в коллекциях Зоологического института Российской Академии Наук (N 9887), заметно отличается от экземпляров из района о-вов Мадейра и Средиземного моря более многочисленными (106) и более мелкими щитками боковой линии, а также тем, что у него наиболее высокие щитки передней части боковой линии по высоте равны щиткам ее задней (прямой) части, высота их составляет около 15,5% длины головы) [Алеев, 1957а].
Биология. Океаническая ставрида постоянно, на протяжении всего года, встречается как на континентальном шельфе у побережья северо-западной Африки, так и в открытой части Атлантического океана.
Район обитания океанической ставриды характеризуется сложным рельефом дна с многочисленными подводными возвышенностями и горами, являющимися областями активного топографического циклогенеза. Наличие над вершинами гор разномасштабных и разнонаправленных круговоротов способствует выносу биогенных элементов в верхние слои воды и формированию зон высокой биологической продуктивности.
Особенности рельефа дна и гидрологической ситуации в районах обитания океанической ставриды создают условия для концентрации и удержания ее икры, личинок и взрослых особей над отдельными подводными горами, расположенными в районах Азорского и Канарского архипелагов, а значительные по протяженности пространства океана с большими глубинами между отдельными участками ареала являются изолирующими барьерами, преодолеть которые неритической форме удается далеко не всегда. В результате образуется сложная пространственная и внутривидовая структура вида.
После наступления темноты скопления ставриды поднимаются над грунтом и обычно наблюдаются в слое 130-220 м над глубинами 180-360 м. Регистрируются такие скопления в виде "размытой дорожки" средней плотности с вертикальным развитием 15-40 м.
Сравнение материала по морфометрии океанической ставриды из шельфовых вод западной Африки и из районов канарских и азорских банок показало, что по ряду пластических и некоторым меристическим признакам существуют различия между этими тремя группировками [Шабонеев, Рязанцева, 1977]. Результаты анализа морфологических признаков позволили выделить внутри вида две биотопически разъединенные группировки, одна из которых приурочена в своем распространении к водам материкового шельфа, а другая - к банкам и подводным возвышенностям в открытой части океана.
Биотопическая группировка, обитающая на материковом шельфе от Бискайского залива до Мавритании, в свою очередь может быть подразделена на географически изолированные части с ареалами, охватывающими шельфовые воды северо-западной Африки и побережья Западной Европы. По-видимому, самостоятельной популяцией следует считать форму этой ставриды в Средиземном море.
У биотопической группировки, распространенной в открытых водах океана, имеет место более сложная структура иерархических уровней [Шабонеев, Рязанцева, 1977].
Для уловов океанической ставриды характерны значительные колебания размерного состава по сезонам и районам. На континентальном шельфе западной Африки наиболее мелкая рыба отмечена в районе мыса Кап-Блан, наиболее крупная - у побережья Марокко. Более стабилен размерный состав уловов ставриды у мыса Бохадор - центра ареала шельфовой популяции. Для этого района характерно наличие относительно мелкой рыбы в зимние месяцы (средняя длина 22,0-23,4 см) и последующее увеличение средних размеров (до 25,2 см) в весенне-летний и осенний периоды.
Несколько иная картина наблюдается на банках Канаро-Мадейрского архипелага, где основу составляют особи длиной 18-24 см и только зимой и в летние месяцы в уловах в заметных количествах появляются крупные экземпляры.
Наиболее резкие различия размерного состава, которые проявляются как в средних значениях, так и в годовой динамике, характерны для ставриды Азорского архипелага.
Присутствие океанической ставриды на каждой из обследованных банок и в районах континентального шельфа в течение всего года, различия в динамике размерного состава подтверждают правильность ранее сделанных выводов о сложной пространственной и внутривидовой структуре вида. В то же время значительные сезонные колебания размерного состава ставриды в уловах свидетельствуют о наличии местных миграций и, следовательно, о возможности обнаружения ее концентраций в эпипелагиали за пределами неритических зон.
Океаническая ставрида относится к порционно-нерестующим видам рыб с непрерывным типом созревания половых продуктов. Общий ход созревания половых продуктов и динамика жирности ставриды укладываются в обычную для субтропических видов схему, хотя наличие ряда популяций, обитающих при различных условиях, вносит в эту схему свои коррективы. В ноябре-декабре половозрелые особи обычно имеют половые продукты в основном на III-IV стадиях зрелости. В апреле океаническая ставрида уже находится в преднерестовом и нерестовом состоянии. К летнему периоду (июнь-июль) все рыбы находятся в посленерестовом состоянии.
Обращает на себя внимание значительное отставание процессов развития половых продуктов ставрид шельфовых популяций по сравнению с океаническими. Стабильность условий среды, в которых обитают океанические популяции, способствует удлинению периода нереста, который начинается у них в декабре, а заканчивается в июне. Более суровые условия на шельфе вынуждают ставриду проводить нерест в сжатые сроки. Соотношение полов близко 1:1.
Поскольку процесс созревания половых продуктов неразрывно связан с энергетическими затратами, то существует и различие между океаническими и шельфовыми популяциями ставрид в динамике жиронакопления. Так, если у азорских популяций к январю практически весь внутриполостной жир расходуется на развитие гонад, то у канарских примерно 30% особей еще имеет степень ожирения внутренностей 1-2 балла. Особенно большие различия имеют место между азорскими и шельфовыми популяциями. У последних в январе более 70% особей имеют значительные запасы внутриполостного жира.
Основными компонентами питания взрослой океанической ставриды служат мелкие рыбы (миктофиды, красноглазка и др.) и эвфаузииды. Ставриды на ювенильной стадии, как показали наблюдения, в основном питаются миктофидами.
У экземпляров из района Мадейры темп роста сравнительно высокий; годовалые особи имеют длину около 12-13 см, двухгодовалые - около 15-17 см, трехгодовалые - 20-23 см. Пятилетний экземпляр из района Мадейры имел длину 25,0 см. На шельфе океаническая ставрида в уловах представлена особями длиной 14-34 см, а на банках в открытом океане - 10-49 см.
Ретроспективный анализ долгопериодных колебаний численности океанической ставриды позволил связать, во-первых, появление этого вида на шельфе Марокко в 1952 г. с аномально большими градиентами температуры между западной и восточной областями океана в 50-е гг.; во-вторых, отсутствие ее в уловах в 1960-1969 гг. с однородностью в распределении температуры на акватории океана, когда температурный градиент не превышал 1°С и, в-третьих, обнаружение промысловых концентраций этой ставриды после 1970 г. с возрастанием градиентов поверхностной температуры. Иначе говоря, биомасса этого вида хорошо коррелирует с общей динамикой вод Северного субтропического антициклонического круговорота и непосредственно связана с переносом тепла Северо-Атлантическим течением из западной области океана в восточную. При этом не исключено образование условий, способствующих резкому увеличению численности популяций, приуроченных к отдельным банкам Азорского и Канарского архипелагов, расширению их ареалов и образованию промысловых скоплений в эпипелагиали за пределами неритических зон [Шабонеев, 1985].
Хозяйственное значение невелико; промысловой статистикой этот вид часто не отделяется от Trachurus trachurus. В уловах на шельфе северо-западной Африки порой составляет от 10 до 80% улова всех видов ставрид.
В августе 1970 г. на банке Консепсьон работало несколько промысловых БМРТ, вылов которых на судо-сутки находился в пределах 33-40 т, а океаническая ставрида составила в общем вылове около 30%. Эффективность работы судов на банке Консепсьон оказалась выше по всем показателям, чем в традиционном районе рыболовства на шельфе у северозападного побережья Африки. Промысел на банках Канаро-Мадейрского архипелага продолжал развиваться и достиг апогея в 1973 г., когда общий вылов составил 46,5 тыс.т, из которых океаническая ставрида составила 11,2 тыс.т. Параллельно проводились поисковые работы и началось промышленное освоение запасов океанической ставриды на банках Азорского архипелага, где в 1973 г. было выловлено 11,4 тыс.т.
В дальнейшем промысел на банках открытой части океана не получил развития. Причина этого - в установлении 200-мильных экономических зон, удаленности районов от традиционных мест лова, сложности материально-технического обеспечения судов. Однако часть банок Канаро-Мадейрского и особенно Азорского архипелага оказалась за пределами 200-мильных экономических зон. В 1975 г. только на банках Ампер и Жозефин было выловлено 10 тыс.т рыбы, из которых основу составляла океаническая ставрида.
Результаты работ по лову ставриды на банках свидетельствуют о том, что тактика облова ее концентраций должна быть достаточно качественно отработана. Днем при облове скоплений ставриды траления на банке должны выполняться с меньших глубин на большие, что снижает вероятность аварии орудий лова. Обратным курсом в светлое время суток тралить не рекомендуется. Поскольку днем рыба ведет себя очень активно перед тралом и резко "садится" на грунт, положительный результат дают лишь те траления, которые выполняются на скорости не менее 4,5 уз. в 1-3 м от грунта. Ночью, когда рыба распределяется в пелагиали, основная задача сводится к точному выходу судна на скопления. В это время суток лучшие "заходы" в трал обычно регистрируются при ведении верхней подборы на горизонте верхней границы скоплений.
Результаты проведенных исследований дают основание считать целесообразным возобновление отечественного крупномасштабного промысла океанической ставриды в открытом океане.