Trachurus symmetricus (Ayres, 1855)
Калифорнийская ставрида, тихоокеанская ставрида (Rus),
Pacific jack mackerel
(Eng)
Синонимы:
Caranx symmetricus Ayres, 1855
Decapterus polyaspis Walford & Myers, 1944
Trachurus symmetricus simmetricus Калифорнийская ставрида (Rus),
Рыбы открытого океана. Н.В.Парин 1998 г.
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Trachurus symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955 г. в удалении до 1100 миль, а в 1972 г.— до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставриды (Trachurus symmetricus murphyi) в 40-х широтах южного полушария отмечен в 1985 г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45—60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2—3 лет). Таким образом, эти факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций), которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300—400 тыс. т в 1974—1976 гг. до 1100—1300 тыс. т в 1978—1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.
Цикличность процесса неизбежно предполагает грядущий спад численности и даже почти полное исчезновение этой рыбы из открытых для международного рыболовства вод. Правда, в те же сроки в каких-то иных участках Мирового океана могут появиться псевдопопуляции других видов. Прогнозировать ход этих пульсационных процессов — одна из важнейших, на мой взгляд, задач прикладной рыбохозяйственной науки.
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus symmetricus (Ayres) - калифорнийская ставрида
Описание. D I, VIII+I, 30-37, в среднем 33,4; A II+I, 25-32, в среднем 28,7. Всех щитков в боковой линии 92-110, в среднем 101; колючих щитков - 45-55, в среднем - 49,3. Голова 21,7-26,7% длины тела, в среднем 24,6%. Верхняя (спинная) ветвь боковой линии оканчивается под основанием пятого луча первого-второго спинного плавника - наичаще под первым лучом второго спинного плавника. Жаберных тычинок 51-60, в среднем 56,1; на нижней части жаберной дуги 36-45, в среднем 39,7. Щитки боковой линии средней величины; высота наиболее высоких щитков передней кривой) части боковой линии 7,7-14,7, в среднем 10,7% длины головы, высота наиболее высоких щитков задней (прямой) части боковой линии 12,7-18,7, в среднем 15% длины головы. Кили на щитках хорошо развиты, шипы на килевых щитках крепкие. Прямая часть боковой линии, несущая киль, составляет 35,2-43,7%, в среднем 39,4% длины тела, а хорда дуги кривой (передней части) боковой линии - 40,7-47,7%, в среднем 44% длины тела. Грудные плавники 18,2-25,2%, в среднем 22,2% длины тела. Тело овальное, его высота 16,2-20,7%, в среднем 18,7% длины. Горизонтальный диаметр глаза 18,7-25,7%, в среднем 21,4% длины головы. Верхняя челюсть 33,7-40,7, в среднем 36,4% длины головы. Изгиб боковой линии лежит под основанием 8-13-го, в среднем 10-11-го луча второго спинного плавника. Первый колючий щиток лежит под основанием 8-14-го, в среднем 11-го луча второго спинного плавника. Расстояние от вершины рыла до вертикали начала основания брюшных плавников 24,7-29,7%, в среднем 27,8% длины тела. Расстояние от вершины рыла до вертикали начала основания первого спинного плавника - 28,2-34,2% в среднем 31,3% длины тела. Расстояние от вершины рыла до вертикали начала основания анального плавника 53,2-59,7%, в среднем 56,4% длины тела. Отношение расстояния между последним и предпоследним лучом анального плавника к расстоянию между предпоследним и предшествующим ему (третьим сзади) лучом анального плавника больше в 1,3-2,1, в среднем в 1,6 раза. Ответвления каналов боковой линии на верхней стороне головы и на спине хорошо развиты. Спина и верхняя часть головы темные, бока и брюхо серебристые; грудные, спинные и хвостовой плавники серые; брюшные и анальный плавники - белые. Вершина рыла и основание грудных плавников - черные. На жаберной крышке черное пятно.
Описание дано по 302 экз. длиной от 27 до 59 см, средней длиной 44,2 см, пойманных нами за пределами 200-мильной зоны США.
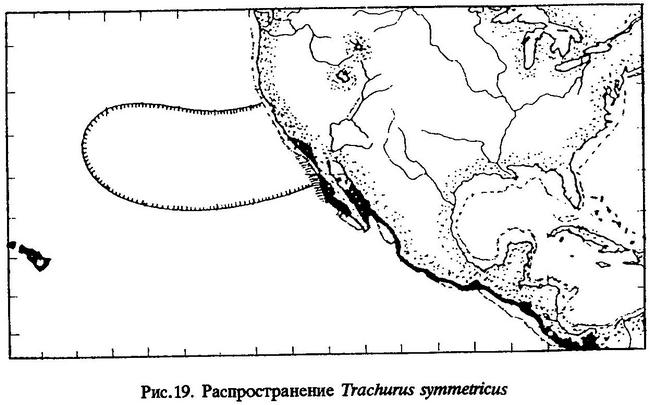
Распространение. Калифорнийская ставрида (рис.19) - обитатель субтропических и умеренных вод, обитающий в эпипелагиали как над шельфом и банками, так и в открытом океане далеко от континента. Ареал калифорнийской ставриды охватывает обширный регион северо-восточной части Тихого океана от залива Аляска до южно-калифорнийских вод у побережья Мексики (мыс Сан- Лукас) [Степаненко, 1980]. Самое южное нахождение этой ставриды отмечено в июле 1957 г. у побережья Коста-Рики, под 10°01'с.ш. и 85°55'з.д. [MacGregor, 1966]. Западная граница распространения находится на расстоянии 400-700 миль [Степаненко, 1980], а по некоторым данным - до 1500 миль [Squire, 1983; Atlas..., 1972] от побережья штата Калифорния. В августе 1955 г. икра и личинки калифорнийской ставриды были обнаружены на 150°з.д., что составляет около четверти расстояния от США до Японии [Alhstrom, 1956]. Однако наибольшая ее численность отмечена в водах у побережья Калифорнии между мысом Консепшен и Калифорнийским заливом; наибольшее количество икры и личинок обычно на 30-36°с.ш. в районе между 80 и 300 милями от берега [MacGregor, 1966; Степаненко, 1980].
Сравнительные замечания. Decapterus polyaspis Wolford et Myers, 1944 следует рассматривать как синоним Trachurus symmetricus [см. MacGregor, 1966].
Биология. Сезонное распределение калифорнийской ставриды сравнительно хорошо исследовано у младшевозрастной части популяции (особи длиной 20-30 см).
Ставрида этих размеров в течение всего года обитает на подводных возвышенностях (банках) среди многочисленных островов от м.Сан-Лукас вдоль п-ва Калифорния и до м.Консепшен. По результатам мечения установлено, что ставрида в этом районе нагуливается, совершая лишь незначительные миграции между островами от берега к берегу и вдоль побережья на север и юг [Tasto, Gregory, 1975]. Особенностью ее распространения является то, что в течение всего года здесь обитают исключительно мелкие особи длиной не более 40 см, в основном от 16 до 30 см [Mais, 1974], тогда как максимальная длина калифорнийской ставриды достигает более 60 см.
Для крупноразмерной ставриды в различные сезоны года характерны протяженные миграции. В зимний сезон ставрида обитает на шельфе США и Мексики и предматериковом склоне на удалении до 50 миль от береговой линии. В весенне-летний сезон ее скопления широким фронтом перемещаются, выходя за 200-мильную экономическую зону, в северную часть ареала - к побережьям штатов Вашингтон, Орегон и в воды Британской Колумбии, заходя в зал. Аляска [Blunt, 1969; Hart, 1973].
Распределение калифорнийской ставриды связано с явлением Эль-Ниньо. Как известно, Калифорнийское течение относится к восточным пограничным течениям. Оно проходит вдоль берегов Северной Америки от 48° до 23°с.ш. и переносит с севера на юг холодную воду. Его ширина составляет в среднем 700 км. Наиболее значительные изменения в системе Калифорнийского течения, как правило, происходит в годы проявления Эль-Ниньо в экваториальной зоне Тихого океана. Вследствие этого существенно изменяется распределение калифорнийской ставриды, что отрицательно сказывается на ее уловах США [Амаров и др., 1985].
По многолетним наблюдениям американских исследователей, основное место обитания мелкоразмерных ставрид - шельф п-ва Калифорния. Здесь скопления калифорнийской ставриды представлены в уловах особями средней длиной 19 см (максимальная - 30 см). Другие материалы также показали, что у побережья п-ва Калифорния обитают особи средней длиной 20 см (максимальная до 30 см), а несколько севернее - на шельфе штата Калифорния, - наряду с рыбами средней длиной 19 см в большом количестве присутствуют рыбы средней длиной 23 и 27 см. На Вашингтоно-орегонском шельфе (северная граница ареала), напротив, в уловах не встречаются рыбы длиной менее 37 см. Основу уловов здесь составляют ставриды средней длиной 54,0 см [Пащенко, 1983].
Интересно распределение калифорнийской ставриды по размерному составу в океаническом районе. Еще в 1957-1958 гг. американскими исследованиями установлено, что в удалении на 80-200 миль от побережья между 27 и 35° с.ш. обитает ставрида в основном двух размерных групп - с модами 39 и 51 см. При этом обращает на себя внимание тот факт, что здесь совершенно отсутствуют в уловах мелкоразмерные группы рыб: особи длиной менее 36 см не встречались. Исследования в открытом океане в 1977-1979 гг. также показали, что в уловах преобладали ставриды крупных размеров - с модами 53-57 см, а рыбы длиной менее 31 см не наблюдались [Пащенко, 1983]. Аналогичную картину наблюдали и мы в 1984 г. в рейсе на СТ "Возрождение" и в 1985 г. на НИС "Профессор Месяцев".
По нашим наблюдениям, в марте днем ставрида держалась в слое 50-150 м, редко - чуть выше и отличалась очень высокой активностью. Ночью ставрида опускалась на глубину 180-240 м, не утрачивая активности, и практически не облавливалась судами типов РТМС и БАТМ. Такое распределение калифорнийской ставриды отмечено несколькими судами экспедиции Минрыбхоза СССР. Но 5 апреля было отмечено изменение в поведении ставриды: днем она стала держаться в слое 30-110 м, а ночью - в слое 40-90 м. И лишь к середине апреля косяки ставриды ночью стали подниматься к поверхности (эхолотом они фиксировались в слое 5-70 м), а днем опускаться на глубину 40-120 м. В мае и последующие летние месяцы в районе 32°37'с.ш. в целом поведение ставриды не изменялось. Вечером она поднималась к поверхности и фиксировалась на глубине от 5 до 50 м, а днем опускалась на большую глубину.
Поведение калифорнийской ставриды на подводных возвышенностях и в открытых частях океана различается. Над большими глубинами косяки ставриды прослеживаются и днем, но глубже, чем ночью. На банке Кобб в светлую часть суток ставрида рассеивается. Например, 8 апреля над банкой Кобб ночью были зафиксированы отдельные плотные косяки ставриды высотой 10-25 м и протяженностью 30-40 м в слое температурного скачка (25-50 м). С наступлением рассвета ставрида рассеялась, записи исчезли. Это был начальный момент подхода нагульной калифорнийской ставриды, количество косяков над банкой быстро увеличйвалось. Они сливались в ленты протяженностью до 1100 м. Плотность косяков была настолько высокой, что при их записи срабатывала "белая лента" гидролокатора. Скопления наблюдались в радиусе 3 миль от центра банки Кобб. К концу октября ставрида с банки исчезла. Следует отметить, что обследования в это время акватории, прилегающей к банке Кобб (до 290 миль в северном и южном направлениях и до 145 миль к западу), положительных результатов не дали. Из этого было сделано предположение, что ставрида ушла в экономическую 200-мильную зону. Правда, в некоторые годы, в частности в 1983 и 1984гг., штучно ставрида была отмечена в уловах на банке Кобб в декабре и даже в январе. Следует также отметить, что в отличие от 1981-1982 гг. косяки ставриды в 1984 г. располагались на грунте и держались там круглые сутки, не рассеиваясь днем.
Большинство ставрид на шельфе в промысловых уловах имеют возраст менее шести лет и представлены в основном двух-четырехгодовиками [Пащенко, 1983].
Между самцами и самками нет различия в темпе роста. Данные разных авторов по росту калифорнийской ставриды хорошо согласуются между собой. Еще Фитч [Fitch, 1956] установил, что калифорнийская ставрида в возрасте двух лет достигает длины 25 см, а в три года - 35 см. Максимальный возраст, по Фитчу, равен 30 годам. Степаненко (1980) отмечал очень быстрый рост калифорнийской ставриды на первом году жизни - до 20 см; затем рост замедляется, и в возрасте четырех лет рыбы достигают длины 35 см. Более подробно рост калифорнийской ставриды изучал В.М.Пащенко (1983), который обратил внимание на различный темп роста в популяциях, обитающих в различных частях ареала [Пащенко, 1983; Blant, 1969; Wine, Knaggs, 1975] (табл.38).
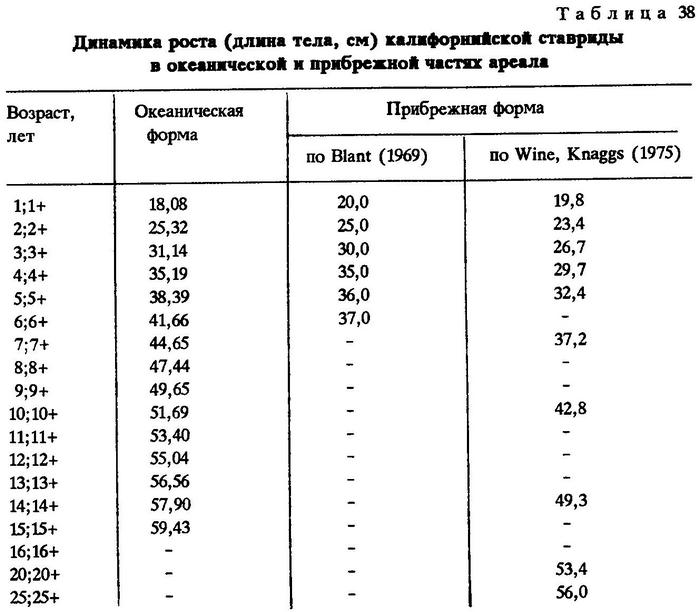
Р.Лонг [Leong, 1984] описал результаты непосредственных наблюдений за ростом калифорнийской ставриды в лабораторных условиях. По уравнению Берталанфи было подсчитано, что средняя длина (по Смитту) посаженных в бассейн ставрид, равная 243 мм, соответствовала возрасту 2,2 года. За два года наблюдений рыбы достигли длины в среднем 408 мм, т.е. прирост составил 165 мм. По уравнению Вайна-Наггса [Wine, Knaggs, 1975], в естественных условиях калифорнийская ставрида при стартовой длине 243 мм достигает длины 408 мм через 6,5 лет. Следует отметить также и значительное увеличение массы ставриды в лабораторных условиях по сравнению с рыбами той же длины, пойманными в море.
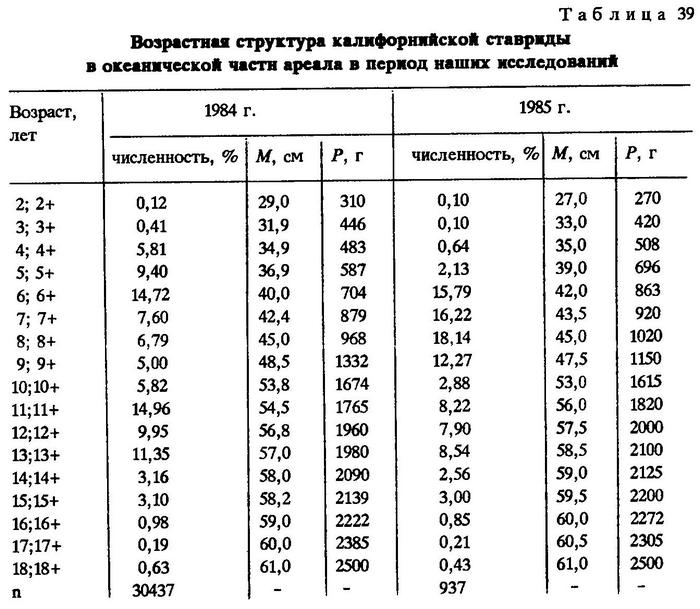
Возрастная структура калифорнийской ставриды в открытой части океана во время исследований и в предшествующие им годы мало различалась. Основу уловов составляли особи 5-14 лет в 1984 г. и 6-15 лет в 1985 г.(табл.39).
Половой зрелости 50% самок достигают при длине 25 см и в возрасте двух лет, 100% - при длине 35 см в возрасте трех лет [Hart, 1973; MacGregor, 1966]. В.Вайн и Е.Наггс (1975) установили, что половозрелость у ставриды, обитающей у берегов Южной Калифорнии, наступает в возрасте одного года при длине 19,8 см, а в двухлетнем возрасте половозрелые самки имеют длину 24,4 см.
У самки длиной 229 мм и массой 171 г в гонадах общей массой 6,14 г общее количество икринок составило 52600, т.е. на 1 г массы рыбы приходилось 308 икринок диаметром 0,40-0,58 мм [MacGregor, 1966]. Следовательно, у самок длиной 40 см количество икринок может быть близким к 225 тыс., у самок длиной 50 см - 425 тыс. и у самок длиной 60 см - 700 тыс. икринок.
Икрометание у калифорнийской ставриды порционное. Обычно нерест происходит весной. По данным Д.А.Фарриса [Farris, 1961], в 50-е годы в водах США пик икрометания имел место в мае, хотя в остальные годы наиболее интенсивный нерест наблюдался в марте. На скорость развития половых продуктов большое влияние оказывает ход температуры вод в преднерестовый сезон, т.е. в конце зимы и начале весны. Ускоренное созревание половых продуктов и более раннее икрометание наблюдаются, когда в преднерестовый сезон идет быстрое потепление. Относительно быстрое нарастание температуры происходит в годы, когда в первые зимние месяцы термический режим вод холоднее обычного. Например, такие условия наблюдались в нерестовый сезон 1951 г. [Farris, 1961], когда в феврале температура воды была ниже среднемноголетней, а в апреле - выше. Наиболее быстрое потепление в период с февраля по апрель, когда температура повысилась в нерестовой части ареала до 15,3°С, пик икрометания был отмечен в марте, хотя нерест ставриды начался только в феврале. В смежные годы, близкие по гидрологическим условиям к среднемноголетним показателям, нерест начинался в январе - феврале и наиболее интенсивно проходил в мае.
По многолетним данным, в мае - июне центр икрометания обычно смещается в прибрежные воды и на север, в район Северной Калифорнии. Вероятно, в этом районе, обычно на расстоянии 50-100 миль от побережья, нерестится младшевозрастная ставрида [Farris, 1961; Wine, Knaggs, 1975].
Кроме отмеченных фактов нереста калифорнийской ставриды у побережья штата Калифорния и п-ова Калифорния икра и личинки ее зарегистрированы также в августе у побережья штатов Вашингтон и Орегон на расстоянии 100-1000 миль от берега [Ahlstrom, 1959].
Соотношение полов в целом близко 1:1, хотя в уловах среди рыб старших возрастов преобладают самцы.
Ихтиопланктонные съемки, проведенные в весенний период на большой акватории с удалением от берега до 300 миль по схеме CalCOFI [Ahlstrom, 1969], позволили выявить районы нерестовых концентраций ставриды. Наиболее интенсивный нерест в 1977 г. отмечен в районе 30-34°с.ш. на расстоянии 21-300 миль от берега. Здесь же в марте - апреле облавливались в большом количестве икра и личинки. Нерестовые скопления в этом районе состояли из крупных рыб длиной 42-64 см, причем в части нерестового района доля младшевозрастных особей была больше. В районе 34-35°с.ш. средняя длина нерестившихся рыб составляла 56 см, а в районе 30-32°с.ш. - 52 см. В конце марта - начале апреля скопления нерестовой ставриды были обнаружены на 34-35°с.ш. на расстоянии 230 миль от берега. В пределах 200-мильной прибрежной зоны нерест ставриды также зарегистрирован, но интенсивность его была небольшой. Личинки ставриды встречались, как правило, ближе к берегу, куда они переносились течением [Степаненко, 1980].
Полагают, что основной нерест калифорнийской ставриды происходит между 27° и 37°с.ш. на расстоянии до 400-600 миль от берега с января по ноябрь с максимумом в марте - июле. Около 69% популяции ставриды нерестится в 200-мильной зоне. Личинки дрейфом выносятся на расстояние до 400 миль от берега [Atlas of the living..., 1972]. Некоторые авторы считают, что за пределами 200-мильной зоны количество нерестующих рыб, вероятно, лишь немногим меньше, чем в ее пределах [Беренбейм, 1979].
Е.Альстром и О.Болл (1954) отмечали, что у калифорнийской ставриды икра пелагическая, неклейкая, сферическая, 0,98 мм (0,90-1,08) в диаметре. Установлено [Ahlstrom, 1959], что 97% икры и 88% личинок калифорнийской ставриды располагаются в слое воды до 50 м, остальные икра и личинки - до 100 м, ниже 140 м они не были обнаружены.
Интересно распределение личинок калифорнийской ставриды по акватории. Данные по количественному распространению ставриды, выловленной на станциях с различным удалением от берега, выглядят таким образом (в %): от побережья до 80 миль - 9,9; от 81 до 160 миль - 32,7; от 161 до 240 миль - 39,9; от 241 до 320 миль - 15,0; от 321 до 400 миль - 2,5.
Е.Альстром и О.Болл (1954) дали детальное описание эмбрионального развития калифорнийской ставриды. Оно типично для большинства видов рода Trachurus. Инкубационный период длится двое суток при температуре воды 17,9°С; трое суток - при температуре 15,9°С и четверо суток - при 14,5°С [MacGregor, 1966]. Личинки, перешедшие на активное питание, растут со скоростью 0,3-0,45 мм в сутки. При этом у них уже через 4-6 сут после перехода на активное питание потребность в пище возрастает в семь - восемь раз, потому что личинки размером 3,0-3,4 мм способны потреблять пищевые организмы только в очень узком размерном диапазоне (от 0,075 до 0,150 мм), что делает их весьма уязвимыми и может приводить к массовой гибели в этот период. Но уже личинки размером 5-6 мм потребляют пищевые организмы в более широком размерном диапазоне (от 0,125 до 1,125 мм). Расширение размерного диапазона кормовых организмов улучшает обеспеченность пищей личинок, способствует более быстрому их росту, что, в свою очередь, обусловливает ускоренное прохождение критического периода в жизни личинок. Совокупность этих факторов можно рассматривать как особый вид приспособления личинок для выживания [Липская, 1985].
Мелкие, ювенильные, особи калифорнийской ставриды питаются копеподами, эуфаузиидами и птероподами [MacGregor, 1966], взрослые - макропланктоном (в основном крупным). Свыше 90% всего состава пищи составляют копеподы, птероподы и эуфаузииды, среди которых преобладают Euphausia pacifica [Carlisle, 1971]. Крупноразмерные особи ставриды, обитающие в мористой части ареала, потребляют эуфаузиид, кальмаров и миктофид, а также креветок, сальп, медуз, парусника-велеллу.
Как уже указывалось, в основном калифорнийская ставрида питается в светлое время суток. Об этом говорит относительно низкий балл наполнения желудков, но высокий - кишечника. Видимо, ставрида в период нереста достаточно активно питается, и не только в промежутках после вымета одной из порций, но и во время икрометания. Исследователи неоднократно обнаруживали пищу в желудках у особей со зрелыми и текучими половыми продуктами. Рыба питалась даже в тех случаях, когда держалась у поверхности. Так, например, в желудках ставрид были обнаружены парусники- велеллы, которые во взрослом состоянии обитают только на поверхности.
Некоторые исследователи давно предполагали наличие двух самовоспроизводящихся популяций калифорнийской ставриды [Gregory, Tasto, 1976]. На это указывали значительные расхождения в возрасте, при котором наступает половая зрелость рыб. По Д.Харту [Hart, 1973], 50% самок ставриды достигает половой зрелости в возрасте двух лет при длине 25-28 см и 100% - в возрасте трех лет при длине тела 31 см. Вайн и Наггс (1975) установили, что половозрелость у мелкой ставриды наступает в возрасте одного года при длине 19,8 см, а в двухлетнем возрасте половозрелые самки имеют длину тела 24,4 см. Следовательно, судя по данным о темпе роста [Wine, Knaggs, 1975] и возрасте наступления половой зрелости, мелкую ставриду, обитающую у п-ова Калифорния, можно отнести к скороспелым рыбам. Общеизвестно, что рыбы в основном становятся половозрелыми, достигнув половины своей максимальной длины. Это значит, что предельная длина мелкой (прибрежной) формы калифорнийской ставриды должна быть примерно 40 см. Именно такую длину имеет ставрида в уловах кошельковыми неводами в водах п-ова Калифорния.
Максимальная длина крупноразмерной формы ставриды составляет 64-66 см. Принимая во внимание вышеуказанную закономерность, логично предположить, что крупноразмерная ставрида должна созревать при длине 32-33 см. Именно на такие размеры полового созревания указывает Д.Харт (1973). На нерестилищах в открытой части океана встречаются исключительно половозрелые особи, минимальная длина которых 30-32 см. Отсюда следует, что по крайней мере существуют две группы ставрид - скороспелая (или быстросозревающая) и позднесозревающая с длинным жизненным циклом. Это позволило предположить наличие двух самовоспроизводящихся популяций - прибрежной и океанической. Исследования, проведенные в водах северовосточной части Тихого океана, подтверждают факт наличия двух популяций калифорнийской ставриды. Установлено, что популяции существенно отличаются темпом роста, возрастом полового созревания, сроками нереста, районами размножения и нагула, миграциями, численностью. Отличия обнаружены также и по числу мягких лучей во втором спинном и анальном плавниках и по ряду пластических признаков [Пащенко, 1983].
В наших исследованиях в 1984 г. на СТ "Возрождение" и в 1985 г. на НИС "Профессор Месяцев" в открытом океане, от северной границы ареала до южной, ставрида была представлена различными размерными группировками с размахом ряда от 24 до 64 см, при хорошо выраженных двух модальных классах со средней длиной 40 и 57 см. Что касается младших возрастных групп ставриды, то, как было показано ранее, они имеют более тесную связь с пришельфовой зоной, тогда как старшевозрастная часть популяции занимает наиболее мористые участки ареала. Нагул этой части популяции происходит в зоне субарктического фронта с постепенным смещением в зимнее время к югу, к месту нереста. Отсюда можно заключить, что старшевозрастная часть популяции приходит на нерест не с шельфа, а из более мористой части ареала, хотя и не обязательно только из пределов 200-мильной зоны. После нереста эта часть популяции, пополняясь средневозрастной группировкой, мигрирует на север, в район летнего положения субарктического фронта, где и нагуливается. Среднеразмерная группировка, по-видимому, занимает промежуточное положение и соприкасается в большей мере с рыболовной зоной. Мелкоразмерная группировка постоянно находится в пределах рыболовной зоны. По мере вступления в нерестовую фазу часть этой группировки нерестится на юге ареала у побережья Калифорнии, главным образом к югу от м. Консепшен и у северной части п-ова Калифорния. Молодь обитает в прибрежной зоне у побережья п-ова Калифорния, а также на подводных возвышенностях этого района, расположенных в 120-150 милях от берега. Анализ многолетних материалов показал стабильность расположения концентраций молоди в междугодовом аспекте. По мере роста и созревания ставрида отходит от берегов и обитает в океанических водах, где, по-видимому, находит более богатую кормовую базу. В посленерестовый период крупная половозрелая ставрида мигрирует на север вплоть до северо-восточной части зал.Аляска, однако основные нагульные концентрации ее располагаются в прибрежной продуктивной зоне у штатов Орегон и Вашингтон. Часть скоплений ставриды в отдельные периоды может наблюдаться и в океанической части вашингтоно-орегонского района, где в летний период развивается обильный зоопланктон. В октябре крупная ставрида начинает мигрировать в южном направлении и в преднерестовый период обитает в рассеянном состоянии главным образом в прибрежной 150-мильной зоне. Самые крупные экземпляры (76 см - зоологическая длина) встречаются в прибрежных уловах только в течение одного или двух летних месяцев [Fitch, 1956; Степаненко, 1980].
Плотные нерестовые концентрации ставрида образует непосредственно перед нерестом. Фактором, определяющим место и сроки образования концентраций, является, по-видимому, температура. Икрометание ставриды происходит в очень узком диапазоне температуры.
Хозяйственное значение. Промысел калифорнийской ставриды освоен давно, но уловы ее долгие годы были незначительными - 200-1500 т и носили смешанный характер, так как она вылавливалась вместе с сардиной и скумбрией. Большое промысловое значение ставрида начала приобретать с конца 40-х годов, когда резко сократились уловы сардины, и вплоть до 1970 г. она являлась основным промысловым объектом Калифорнийского района [Каратаева, 1985а]. Сейчас ни в США, ни в Мексике значительного промысла ставриды нет. С 1950 г. у южного побережья Калифорнии существует небольшой промысел рыб младших возрастных групп (моложе шести лет), но эта отрасль, как считает А.Д.Мак-Колл [MacCall, 1983], сталкивается с трудностями в реализации продукта. Значение калифорнийской ставриды в промысле невелико и в последние годы. Так, из 2,5 млн.т вылова рыбы в северо-восточной части Тихого океана в 1983 г. на долю ставриды пришлось всего 18672 т.
Вылов ставриды по годам распределялся следующим образом, (в т): 1979 г. - 16573; 1980 г. - 20135; 1981 г. - 15930; 1982 г. - 26270; 1983 г. - 18672; 1984 г. - 10859; 1985 г. - 9663; 1986 г. - 11813; 1987 г. - 12320 г. - 1988 г. - 10290; 1989 г. - 13090; 1990 г. - 4160 [Yearbook..., 1985, 1989, 1992].
Известно, что до 70-х годов вылов калифорнийской ставриды кошельковыми неводами США и Мексики достигал значительных величин. В удачные сезоны ее вылавливали до 73 тыс.т, причем 67% из этого улова добывалось в 80-мильной прибрежной зоне. Наибольший вылов калифорнийской ставриды приходится на воды, прилегающие к району Лос-Анджелеса. Калифорнийская ставрида хорошо ловится в прибрежной зоне, а наименьшие ее уловы приходятся на весну и начало лета; вылов подвержен значительным годовым колебаниям [MacGregor, 1966].
Вылов калифорнийской ставриды отечественными судами в 1983 г. составил всего 600 т, в 1984 г. - 181 т [Yearbook..., 1985]. Некоторая часть этой величины приходится на ставриду, выловленную за 200-мильной зоной.
Снижение вылова калифорнийской ставриды в 1983 и 1984 гг. некоторые авторы считают связанным с явлением Эль-Ниньо. В результате воздействия Эль-Ниньо 1982-1983 гг. в водах Калифорнийского течения значительно снизилась первичная продукция. Биомасса фитопланктона в марте 1983 г. упала вдвое по сравнению с апрелем 1982 г.- до 0,4 мг/м3. Существенно изменилось распределение объектов промысла, что в конечном счете сказалось на эффективности рыболовства. Общий вылов рыбы и нерыбных объектов США уменьшился на 25% по сравнению с 1982 г. и на 32% по сравнению со среднегодовым значением за последние шесть лет. Наиболее сильно снизился вылов пелагических рыб [Fiedler, 1984; Амаров и др., 1985].
Тем не менее имеющиеся материалы показывают, что запасы калифорнийской ставриды позволяют вылавливать значительно больше рыбы. По данным американских исследователей, численность калифорнийской ставриды в последние два десятилетия находится на высоком уровне. Биомассу ставриды рассчитывают по учету икры и личинок на основе ихтиопланктонных съемок в районе CalCOFI. В 1955-1957 гг. запасы калифорнийской ставриды в прибрежной полосе до 1,5 тыс.км от берега оценивались в 2 млн.т [Каратаева, 1985а]. В 60-х годах величина пополнения колебалась в широких пределах, причем годы высокого пополнения сменялись несколькими годами низкого. В 1968 г. запас ставриды был определен в 2,1 - 4,8 млн.т [Ahlstrom, 1968]. В течение последних двух десятилетий отмечался высокий уровень пополнения в 1976 и 1982 гг. Несмотря на значительные запасы, промысел ставриды ведется лишь в южных водах Калифорнии в районах нереста. Предполагают, что в будущем возможно расширение акватории промысла [MacCall, 1983]. В 1983 г. нерестовая часть популяции оценивалась в 2 млн.т [MacCall, Staufer, 1983], что может обеспечить вылов от 200 до 420 тыс.т в зависимости от численности поколений.
Следует, однако, отметить, что появились работы, уточняющие эти расчеты, в результате чего оценка биомассы калифорнийской ставриды несколько снизилась и составляет 0,6-1,2 млн.т [Fishery..., 1979].
Как видно, запасы калифорнийской ставриды достаточны для того, чтобы увеличить ее вылов по сравнению с существующим по крайней мере на порядок. Промысловые ресурсы ставриды за 200-мильной зоной в весенне-летний период в отдельные годы также позволяет организовывать промысел, хотя о его устойчивости и масштабах говорить пока не представляется возможным.
Некоторые исследователи считают, что в водах Мексики запасы калифорнийской ставриды практически не затронуты, но их состояние до настоящего времени не изучено (Alvares, 1984).