Arnoglossus kessleri Schmidt, 1915
Арноглосс Кесслера (Rus),
Scaldback
(Eng)
Синонимы:
Arnoglossus kessleri Schmidt, 1915
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
ARNOGLOSSUS KESSLERI SCHMIDT
Условия размножения. Нерест Arnoglossus kessleri в Черном море происходит в летнее время. Икринки этого вида встречаются у берегов Крыма и Кавказа с июня по август (Водяницкий, 1936; Овен, 1959). У побережья Болгарии нерест начинается в июне и продолжается до середины сентября (Георгиев и др., 1960).
По наблюдениям указанных исследователей, нерест Arnoglossus kessleri происходит повсеместно вблизи берегов над небольшими глубинами.
В опресненном Одесском заливе нахождение икринок Arnoglossus kessleri не отмечено (Зайцев, 1959а). Однако в Азовском море (в районе Казантипского маяка) единичные икринки были найдены нами при солености 11,55‰. Температура воды в районах размножения соответственно летнему режиму колеблется сравнительно в небольших пределах — от 18—19 до 24—25°.
По нахождению в планктоне икринок на разных этапах развития можно было уловить суточный ритм размножения этого вида. В утренние часы (05—06 ч) икринки Arnoglossus kessleri всегда находились в начале III этапа развития. Следовательно, вымет их, как и большинства рыб Черного моря, происходит ночью. Поскольку икринки Arnoglossus kessleri встречаются в планктоне редко, нам не удалось проследить последовательность развития суточных выметов.
Строение икринок. Икринки Arnoglossus kessleri отличаются от пелагических икринок всех других размножающихся летом рыб Черного моря своей очень маленькой величиной и соответственно наличием одной небольшой жировой капли. Желток гомогенный (рис. 60). По данным ряда исследователей, диаметр икринок Arnoglossus kessleri колеблется от 0,59 до 0,70 мм, жировой капли — от 0,10 до 0,13 мм (Водяницкий, 1936; Косякина, 1938; Водяницкий и Казанова, 1954; Овен, 1959; Георгиев и др., 1960; наши измерения).
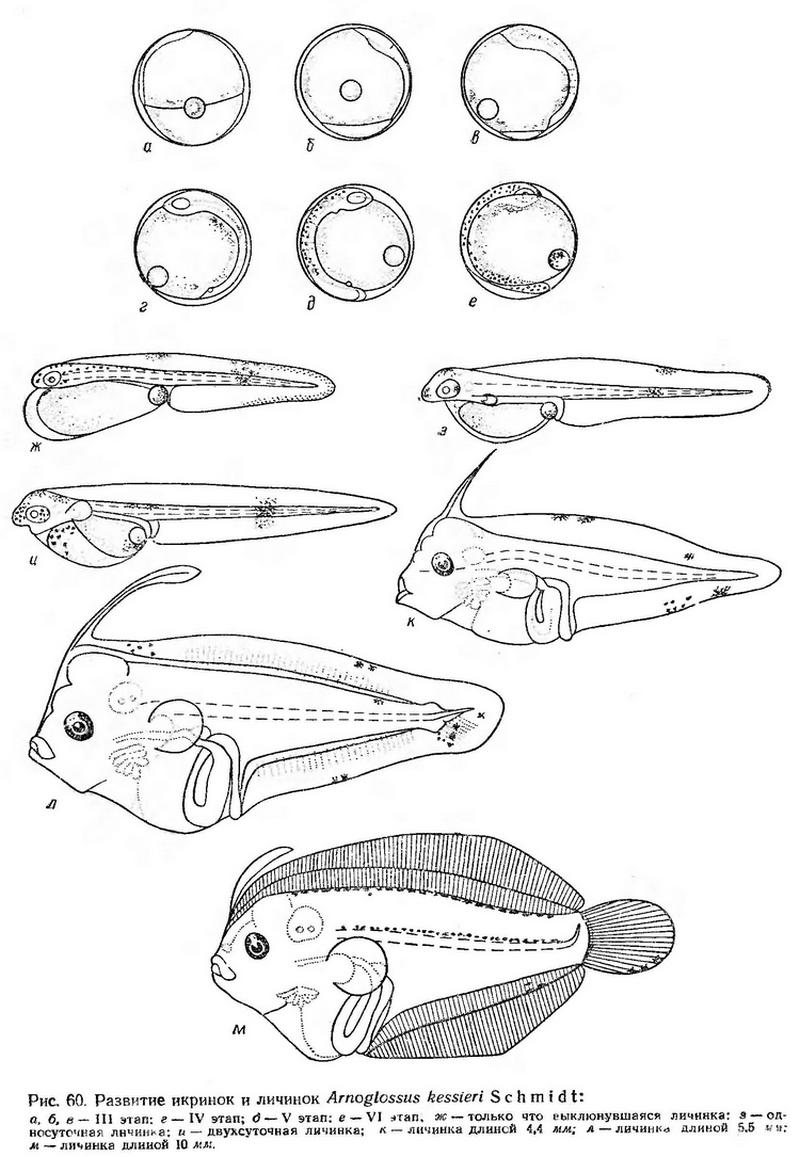
Эмбриональное развитие прослежено на икринках, собранных в ранние утренние часы, когда они находились на III этапе развития (начало обрастания желтка бластодиском).
Когда края бластодиска заходят за середину желтка, очертания зародышевой полоски еще довольно расплывчатые. Головной конец слабо расширен, достигает анимального полюса (рис. 60, а). В конце III этапа перед закрытием бластопора намечаются глаза, в туловищном отделе появляются первые миотомы. Пигмента на теле нет (рис. 60, б, в). При температуре воды 21,6° процесс обрастания желтка продолжался около 6 ч. После замыкания бластопора число миотомов в туловищном отделе быстро увеличивается, намечаются зачатки внутренних органов. Тело остается бесцветным; икринки совершенно прозрачные (рис. 60, г). IV этап при указанной температуре продолжается около 4 ч.
Когда эмбрион охватывает примерно половину поверхности желтка, появляется хвостовая почка и начинается быстрый рост хвостового отдела. Продолжается дифференцировка туловищных миотомов, появляются зачатки слуховых капсул, развиваются заложенные зачатки внутренних органов. В конце этапа намечаются зачатки грудных плавников, появляются мелкие клеточки оранжевого пигмента на спинной стороне зародыша и на жировой капле на стороне, обращенной к зародышу (рис. 60, д). V этап при температуре 22° продолжается 2,5—3 ч.
Когда хвост эмбриона немного заходит за середину желтка, начинается слабая пульсация сердца; эмбрион время от времени подергивается. На VI этапе продолжается формирование внутренних органов, усиливается оранжевый пигмент на теле (рис. 60, е). При температуре 22—21,5° VI этап продолжается 5—7 ч.
Постэмбриональное развитие. Соответственно небольшим размерам икринок личинки выклевываются очень маленькими с большим грушевидным желточным мешком. Длина только что выклюнувшихся личинок 1,8—1,9 мм (Водяницкий и Казанова, 1954; наши данные). Тело их стройное, прогонистое, окаймлено узкой плавниковой каймой, края которой в постанальной части тела на дорсальной и вентральной сторонах имеют пильчатую зазубренность. В отличие от личинок многих других нерестующих летом рыб с пелагической икрой, только что выклюнувшиеся личинки Arnoglossus kessleri имеют зачатки грудных плавников, которые довольно быстро развиваются и уже к трехсуточному возрасту приобретают подвижность. Жировая капля находится в заднем верхнем углу желточного мешка. Анус открывается сразу за желточным мешком; антеанальное расстояние составляет около 50% длины тела. На спинной стороне туловищного отдела и на голове разбросаны клеточки оранжевого пигмента. Такой же пигмент в виде отдельных пятен имеется на дорсальной плавниковой оторочке, в средней части постанального отдела и на жировой капле (рис. 60, ж). Личинки плавают брюшной стороной вверх. Движения порывистые, но кратковременные, осуществляются благодаря сильным изгибам тонкого стройного тела. Периоды покоя длительные — до 1,5—2 мин.
В течение первых суток развития происходят заметные изменения в строении личинок. Голова освобождается от желтка, намечается ротовая ямка, формируется кишечник. Увеличиваются зачатки грудных плавников. Заметно удлиняется хвостовой отдел; антеанальное расстояние сокращается до 44—45% длины тела (рис. 60, з). Движения личинок становятся более быстрыми, но по-прежнему остаются очень непродолжительными (до 2 сек).
К двухсуточному возрасту личинки достигают 2,3 мм длины. Желточный мешок сильно сокращается, значительно уменьшается также жировая капля. Хвостовой отдел почти в два раза длиннее туловищного (антеанальное расстояние равно 39% длины тела). Грудные плавники большие, веерообразные. Рот прорезался (рис. 60, и). Периоды покоя значительно сократились; личинки быстро и стремительно плавают.
Дальнейшее краткое описание личинок дано по фиксированному материалу.
Личинки длиной 4,4 мм (рис. 60, к) имеют очень характерную листовидную форму тела. Высокая голова и широкое туловище переходят в суживающийся постанальный отдел. Кишечник образует несколько петель. Между перикардиальной областью и кишечником расположена массивная печень. Имеется довольно большой плавательный пузырь. Отношение наибольшей высоты тела к длине составляет 30—31%. На голове расположен заостренный вырост, появляющийся из продольного расщепления (расслаивания) наружного края спинной плавниковой каймы (Kyle, 1913). Такой вырост имеется уже у личинок этого вида длиной 3,43 мм (Водяницкий и Казанова, 1954).
Первоначально головной вырост, по наблюдениям Кайла (Kyle, 1913), располагается вдоль края спинной плавниковой каймы. Затем, по мере роста личинки, он принимает все более и более вертикальное положение, и у некоторых видов этого рода по мере развития вырост снова приобретает горизонтальное положение, выступая впереди головы.
Личинки длиной 5,5 мм (рис. 60, л) имеют совершенно симметричное листовидной формы тело. Высота тела увеличивается до 38% по отношению к длине. Начинается дифференцировка непарных плавников. Вырост на голове становится массивным, верхний конец его расширяется.
У личинок размером 9,2 мм наибольшая высота тела достигает 42—43% длины. Личинки остаются симметричными, внешних признаков метаморфоза нет. Плавательный пузырь сохраняется. Вырост на голове заметно уменьшается (Калинина, 1960).
Личинки длиной 10—11 мм имеют вполне сформированные непарные плавники. Внешних признаков метаморфоза по-прежнему нет. Туловище очень высокое (до 50% длины тела), тело сильно сплющено с боков. Вырост на голове редуцируется; он сохраняется в виде небольшого язычка, расположенного вдоль спинного плавника (рис. 60, м). По наблюдениям Кайла (1913), головной вырост у всех видов рода Arnoglossus внезапно исчезает, как только начинается миграция глаза, тогда как у видов рода Bothus он исчезает задолго до начала метаморфоза. Сопоставляя развитие личинок, относящихся к двум указанным родам, Кайл (1913) приходит к выводу, что вырост на голове предохраняет передние доли мозга и, следовательно, выполняет защитную функцию. Указание Э. М. Калининой (1960) со ссылкой на Кайла о гидростатической функции этого выроста ошибочно.
Данные о распределении и численности икринок и личинок. Икринки и личинки Arnoglossus kessleri встречаются в Черном море в прибрежных районах и обычно в небольшом количестве. В. А. Водяницкий (1936) отмечает нахождение единичных икринок в районе Батуми на глубине 10—15 м. По наблюдениям Е. Г. Косякиной (1938), они закономерно встречались в планктоне Новороссийской бухты. Сравнительно большое количество икринок этого вида было обнаружено в районе Гудаут в июле 1948 г. (Дехник и Павловская, 1950). У побережья Болгарии икринки Arnoglossus kessleri обычно встречаются единичными экземплярами. Максимальное количество (26 икринок на лов) было выловлено в июне 1957 г. в Несебрском заливе (Георгиев и др., 1960).
Наши многолетние наблюдения также свидетельствуют о сравнительной малочисленности икринок Arnoglossus kessleri в планктоне. Они закономерно встречаются в июле и августе в разных районах Черного моря, но обычно не более 5—7 экз. на один горизонтальный 10-минутный лов.
Личинки этого вида, по наблюдениям З. М. Пчелиной (1940), вылавливались в Новороссийской бухте с конца июля до середины сентября. Чаще всего они ловились в районе Новороссийск — Анапа. Представляет интерес нахождение личинки этого вида длиной 2,5 мм на расстоянии 30 миль от берега (Дехник и Павловская, 1950), что свидетельствует о рассеивании личинок благодаря течениям на большой площади моря. У берегов Болгарии личинки Arnoglossus kessleri встречаются редко и единичными экземплярами (Георгиев и др. 1960).
В наших сборах в районе Камышовой бухты личинки этого вида встречались довольно часто в июле и августе, но не более 1—2 экз. в одном горизонтальном десятиминутном лове.
По горизонтам лова все имеющиеся в наших сборах личинки распределяются следующим образом: на 0 м поймано 4 личинки, на 10 м — 13 и на 20 м — 2 личинки.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
У побережья Черного моря и в Средиземном
море повсеместно обитает арноглосс Кесслера (Arnoglossus kessleri), маленькая рыбка,
длиной 5 см.