Hippoglossoides dubius Schmidt, 1904
Япономорская палтусовидная камбала (Rus),
Flathead flounder
(Eng)
Синонимы:
Cynopsetta dubia Schmidt, 1902
Hippoglossoides dubius Schmidt, 1904
Hippoglossoides elassodon dubius Schmidt, 1915
Hippoglossoides katakurae Snyder, 1911
Hippoglossoides robustus dubius Bogdanov, 1981
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 6. (Pleuronectiformes). Г.У.Линдберг, В.В.Федоров 1993 г.
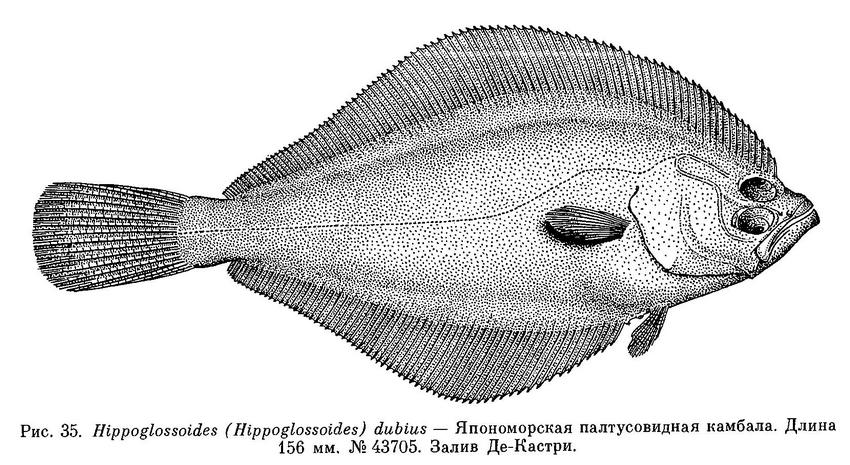
3. Hippoglossoides (Hippoglossoides) dubius Schmidt, 1904 — Япономорская палтусовидная камбала (рис. 35).
Cynopsetta dubia Шмидт, Изв. Имп. Рус. геогр. о-ва, 38, 5, 1902 : 521 (nomen nudum). — Jordan, Starks, Proc. U. S. Nat. Mus., 31, 1906 : 188 (описание).
Hippoglossoides dubius Шмидт, Рыбы восточных морей..., 1904 : 227, табл. 6, рис. 1 (Холмск и зал. Анива; исключая экз. из зал. Анива, принадлежащий к Hippoglossoides elassodon). — Hubbs, Proc. U. S. Nul. Mus., 48, 1915 : 466 (синонимия, описание). — Norman, Flatfishes (Heterosomata)..., 1934 : 301, fig. 222 (синонимия, описание). — Mоri, Mem. Hyogo Univ. Agri., 1, 3, (Biol. Ser. N 1), 1952 : 177. — Matsubara, Fish morph. a. hier., 2, 1955 : 1268. — Ueno, Fish. Hokkaido, 1965 : 66, fig. 6—A. — Chyung Moon Ki, Fish. Korea, 1977 : 566. — Фадеев, Изв. ТИНРО, 102, 1978a : 17 (частью: исключая экземпляры с жаберными тычинками более 15, лучами в спинном менее 80 и в анальном плавниках менее 60). — Sakamoto in Amaoka et al., Fishes North-Eastern Sea Japan..., 1983 : 160, 217, 306, 348, col. pl. 107, 187 (цветные фотографии). — Sakamoto, Mem. Fac. Fish. Hokkaido Univ., 31, 1—2, 1984 : 208 (положение в системе).
Hippoglossoides katakurae Snyder, Proc. U. S. Nat. Mus., 40, 1911 : 546 (Отару). — Snyder, Proc. U. S. Nat. Mus., 42, 1912a : 438, pl. 58, fig. 1 (рисунок). — Tanaka, Fig. descr. fish. Jap., 12, 1913b : 207, pl. 57, fig. 212 (описание).
Hippoglossoides elassodon dubius, Schmidt, Ann. Mag. Nat. Hist., 8, 16, 1915 : 307 (подвидовой ранг). — Солдатов, Линдберг, Обзор рыб..., 1930 : 394 (синонимия, описание). — Шмидт, Рыбы Охотского моря, 1950 : 228 (синонимия, отличия). — Коваль, Богданов в: «Биохимическая и популяционная генетика рыб», 1979 : 100 (подвидовой ранг по данным биохимической генетики).
Hippoglossoides (Cynopsetta) dubius Hubbs, Annot. Zool. Jap., 9, 4, 1918 : 374 (принадлежность к подроду).
Hippoglossoides robustus dubius, Богданов и др., в: «Генетика и размножение морских животных», 1981 : 34 (подвидовой ранг или конспецифичность с Hippoglossoides robustus). — Коваль, Богданов, Вопр. ихтиол., 22, 4, 1982 : 685.
12366. Японское море, Татарский прол., Холмск. Глубина 84 м. 8 VI 1901. П. Ю. Шмидт. 1 экз. Лектотип. Второй экз. типовой серии (ЗИН 12367) оказался Hippoglossoides elassodon, поэтому ЗИН 12366 следует считать лектотипом Hippoglossoides dubius.
17900. Японское море, 50°38' с. ш., 141°05' в. д. 8 IX 1911. В. К. Солдатов. 1 экз.
17902. Японское море, мыс Поворотный. 6 IV 1913. ДВЭ. 1 экз.
17903. Японское море, 42°18' с. ш., 130°04' в. д. 8 IV 1913. ДВЭ. 1 экз.
17904. Японское море, зал. Петра Великого, п-ов Песчаный. 4 X 1912. ДВЭ. 1 экз.
17906. Японское море, Татарский прол., мыс Хой. 1 IX 1913. ДВЭ. 2 экз.
17908. Японское море, Татарский прол., мыс Сюркум. 25 VIII 1911. ДВЭ. 2 экз.
17910. Японское море, Татарский прол., Агнево. 5 VII 1913. ДВЭ. 2 экз.
17911. Японское море, 51°00'45" с. ш., 140°40' в. д. 17 VIII 1910. Ф. А. Дербек. 1 экз.
18599. Японское море, 48°27' с. ш., 140°42' в. д. 1 VII 1909. Ф. А. Дербек. 1 экз.
23739. Японское море, Пусань. 1930. 3 экз.
43705. Японское море, зал. Де-Кастри. 13 VI 1912. Ф. А. Дербек. 1 экз.
45179. Охотское море, зал. Анива, бухта Буссэ. 23 VIII 1901. П. Ю. Шмидт. 1 экз.
45412. Охотское море, мыс Хайрюзова. 28 VIII 1949. Э/с «Витязь». 1 экз.
48221. Охотское море, зал. Анива. Глубина 32 м. 23 IX 1947. КСЭ. 2 экз.
48222. Тихий океан, о. Итуруп, зал. Касатка. Глубина 299 м. 14 IX 1949. КСЭ. 3 экз.
48268. Тихий океан, о. Хонсю, зал. Сэндай. Глубина 540 м. 31 I 1981. Б. А. Шейко. 1 экз.
D 85; А 67; Р 11; V 6, l.l. 88; жаберных тычинок 4 + 14—18 (Шмидт, 1904 : 227).
D (79) 82—87 (89); А (60) 63—65 (69); l.l. 86—94; жаберных тычинок 2—3 + (11) 13—15 (16) = (13) 15—18 (19) / 2—3 (4) + 12—16 (17) = (14) 15—19 позвонков 13 + 29—32 = 42—45 (Hubbs, 1915 : 468).
D (79) 82—87 (90); А (62) 64 (69); Р 10—11; С 18 (10—12 ветвистых); l.l. 88—93, выше боковой линии около 38 рядов чешуи; жаберных тычинок ? + (11) 13—15 (16) (Norman, 1934 : 301).
D 82—92; А 63—67; Р 9—11 / 9—10; V 6; С 18; l.l. 90—99; жаберных тычинок 2—4 + 13—15; позвонков 13 + 32 = 45 (Sakamoto, 1983 : 217, 348).
D 75—95; A 58—72; Р 8—12; V 6; l.l. 87—98; жаберных тычинок 1—4 + 11—17 (Masuda et al., 1984 : 336).
По 23 экз. длиной 111—289 мм коллекции ЗИН: D (77) 80—85 (87), ср. 82.54; А (62) 64—67, ср. 64.90; Р (10) 11—12, ср. 11.14 / (9) 10—11 (12), ср. 10.71; l.l. (91) 94—96, ср. 94.0, выше боковой линии (30) 32—38 (45), ср. 35.5 рядов чешуи, ниже боковой линии 30—49, ср. 37.67; жаберных тычинок (1) 2—3 (4), ср. 2.62 + (12) 13—14, ср. 13.15 = (13) 15—17, ср. 15.77.
В процентах стандартной длины: длина головы 26.9—28.8; наибольшая высота тела 33.0—45.6. В процентах длины головы: длина рыла 21.5—25.4; диаметр глаза 18.3—22.4; длина верхней челюсти глазной стороны головы 38.8—43.1, нижней челюсти глазной стороны головы 31.3—54.5, наибольшего луча спинного плавника 33—42.2, анального плавника 32.4—43.4, грудного плавника глазной стороны тела 41.0—52.8 (Sakamoto, 1983 : 217, 348).
По нашим материалам (23 экз. длиной 111—289 мм) в процентах стандартной длины: длина головы 27.8—29.7, ср. 28.76; наименьшая высота тела (хвостового стебля) 9.4—10.6, ср. 10.05; длина хвостового стебля 6.8—7.8, ср. 7.09, грудного плавника глазной стороны тела 12.2—13.9, ср. 13.41. В процентах длины головы: наименьшая высота тела (хвостового стебля) 33.8—37.8, ср. 34.94; длина хвостового стебля 23.0—28.0, ср. 24.67; длина грудного плавника глазной стороны тела 44.0—48.6, ср. 46.63.
Hippoglossoides dubius по счетным признакам близок к Hippoglossoides elassodon, но отличается от него меньшим числом жаберных тычинок на нижней части первой жаберной дуги, большим числом пор в боковой линии. Кроме этого, для него характерны наличие нескольких клыкообразных зубов впереди на каждой челюсти, узкое, покрытое 2—3 рядами чешуи, межглазничное пространство без гребня и суженное тело.
Окраска глазной стороны тела однотонная, коричневая, у молодых рыб с отдельными небольшими черными пестринами, лучи вертикальных плавников желтовато-коричневые, перепонка между ними прозрачная или желтая. Слепая сторона тела белесая. Отмечены нахождения левосторонних экземпляров, которые на слепой (правой) стороне имели темную окраску в задней половине тела и необычное строение оптической хиазмы (Honma, 1958 : 62; Okiyama, Tomi, 1970 : 84; Honma, Mizusawa, Kakimoto, 1979 : 20).
Проведены сравнительное изучение осевого (Hotta, 1961) и всего скелета (Sakamoto, 1984), количественный анализ нервных волокон в боковой линии (Tsukamoto, Yoshiko, 1957).
Исследовался состав липидов мышц, кишечника, печени и гонад (Igarashi et al., 1962, 1963а, 1963b, 1963с). Считается, что по соотношению ненасыщенных жирных кислот — моноеновых и полиеновых — можно судить о глубине обитания рыб. У видов, обитающих на больших глубинах, величина отношения больше 4 (Hayashi, Yamada, 1975).
Обитатель шельфа и материкового склона от 20- до 1200-метровых глубин. Совершает четкие сезонные миграции. Осенью перемещается на большие глубины. Зимует на материковом склоне: основные концентрации распределяются на глубинах 150—800 м при температуре придонных слоев воды от — 1 до 1 °С; в это время рыбы не питаются, малоподвижны; скопления отличаются наибольшей плотностью населения. В отдельные годы в зал. Петра Великого к югу от о. Аскольд такие скопления облавливались рыбаками даже на глубинах до 1200 м. Весной начинается обратное перемещение на шельф, где летом на глубинах 25—130 м происходит нагул, а на глубинах 25—85 м — нерест. Наибольшие летние концентрации отмечены на глубинах 50—100 м при температуре придонных слоев воды от —1 до 13 °С, плотность скоплений небольшая, так как рыбы занимают на шельфе значительную площадь.
Сроки нереста в зал. Петра Великого в апреле-июне, у берегов Приморья — в апреле-июле, в Татарском прол. — в мае-августе, у о. Хоккайдо — в феврале-апреле. Нерест происходит на глубинах 30—100 м при температуре придонных слоев воды от —0.45 до +8 °С, пик — при 2—3 °С. Икра пелагическая, с большим перивителлиновым пространством, диаметр ее колеблется от 2.02 до 2.94 мм; развивается при температуре поверхостных слоев воды в зал. Петра Великого от —0.22 до +5.5 °С, у берегов Приморья — от 3.42 до 8.2 °С, в Татарском проливе — от 1.30 до 4.72 °С. Плодовитость колеблется от 211 до 560 тыс. икринок. Икрометание порционное, в 2—3 приема. Выклюнувшиеся предличинки имеют длину 2.6—3.13 мм через 4—5 сут после оплодотворения.
В уловах исследовательских судов преобладают рыбы длиной 28—42 см и массой 0.18—0.67 кг.
Половое созревание наступает при длине самцов 19—32 см, самок — 20—37 см, в возрасте самцов 3—4 года, самок 4—5 лет.
Питание как типично донными (Pandalidae, Bivalvia, Ophiuroidae), так и пелагическими организмами (Hiperiidae, Euphausiidae, Sagittidae). Зимой интенсивность питания резко снижается, отмечается до 50 % рыб с пустыми желудками. В зал. Вакаса в желудках Hippoglossoides dubius отмечены Glyptocephalus stelleri, а молодь Hippoglossoides dubius поедается взрослыми особями Theragra chalcogramma и Glyptocephalus stelleri. Дополнительные сведения по биологии можно найти в ряде работ отечественных (Микулич, 1952, 1954; Моисеев, 1953а; Перцева-Остроумова, 1955, 1956, 1961; Иванков и др., 1972; Иванков, Курдяева, 1973; Иванков, Иванкова, 1974; Фадеев, 1984, 1987) и зарубежных (Okada, 1955; Hayase, Hamai, 1974; Minami, 1986) ихтиологов.
Промысел начат в зал. Петра Великого в конце 20-х годов, в 1935—1936 гг. вылов уменьшился почти в 4 раза. Поэтому лов крупнотоннажным флотом был прекращен, но развивался летний промысел мелких судов. Hippoglossoides dubius в уловах камбаловых отмечался в виде прилова. Увеличение интенсивности промысла при непрерывной модификации судов, и орудий лова позволило к 40-м — началу 50-х годов довести общий вылов камбаловых до 10.2—12.9 тыс. т. За этим последовало быстрое ухудшение состояния запасов: снизились с 277 до 90 кг уловы на усилие, общий вылов к 1964 г. упал до 2.7 тыс. т, произошло резкое уменьшение размеров и омоложение камбал, замена основных промысловых видов малоценными камбалами. Hippoglossoides dubius в составе уловов вышел на первые места. С целью восстановления запасов с 1967 г. был введен запрет на промысел камбал. За 5 лет (1968—1972) уловы на усилие, по данным контрольных уловов, увеличились почти в 3 раза, возросла относительная численность рыб старших возрастов. Все эти изменения позволили с 1974 г. организовать лимитированный промысел с жестким соблюдением комплекса охранных мероприятий. Допустимое изъятие было оценено в 4 тыс. т.
В водах Северного Приморья (мыс Белкина — Советская Гавань) и Татарского прол. (Александровская банка) запасы камбал были открыты в конце 30—х годов. В первом районе промысла не было, хотя по результатам исследовательских траловых съемок допустимое изъятие всех камбаловых оценено в 5.5 тыс. т. Во втором районе лов был начат с 1943 г. В первые три года вылов достиг 10.1 тыс. т, затем с 1947 г. по настоящее время промысел был развит слабо в связи с переловом в предыдущие годы. В середине 60—х годов отмечалось вмешательство иностранного (КНДР, Япония) промысла. В этом районе допустимое изъятие всех камбаловых оценено в 2.4 тыс. т. Сведения о промысле и промысловых запасах заимствованы из работ отечественных авторов (Павленко, 1920; Кривобок, 1931; Моисеев, 1937б, 1946а, 1946б, 1946в, 1953а; Тычкова, 1946; Минева, 1971а, 1971б, 1972; Фадеев, 1971б, 1984, 1987; Иванкова, 1975, 1976, 1979; Стрельцов и др., 1985).
Длина 520 мм (Фадеев, 1987 : 115).
Распространение. В Японском море известен повсеместно: у материкового побережья у Пусаня (Шмидт, 1931а : 318), Пхоханя (Hubbs, 1915 : 466), Воньсаня (Mori, 1952 : 177), многочислен в зал. Петра Великого (Павленко, 1910 : 56, 71) и у берегов Приморья на север до Амурского лимана (Охрямкин и др., 1936 : 12). Вдоль островных берегов в р-не Санин (Yanai, 1950 : 20, Mori, 1956а : 31; Minami, Takomaki, 1980 : 159), у о-вов Оки (Hubbs, 1915 : 466), на банке Ямато (Mori, 19566 : 32), в заливах Вакаса (Hubbs, 1915 : 466; Takegawa, Morino, 1970 : 357; Minami, Nakamura, 1978 : 33) и Тояма (Katayama, 1940a : 21), у преф. Ниигата (Honma, 1952 : 224; Honma et al., 1979 : 20), у о. Садо (Honma, 1963 : 26;. Honma, Kitami, 1978 : 63), около о. Хоккайдо у Таджимы (Katayama, 1952 : 5, 58), в зал. Исикари (Snyder, 1911 : 546; Hubbs, 1915 : 466; Kobayashi, 1962 : 263), вдоль о. Сахалин на север до Широкой Пади (Шмидт, 1904 : 227; Hubbs, 1915 : 466; Таранец, 1937б : 38). В Охотском море известен в южной части: в зал. Анива (Шмидт, 1904 : 227, 1950 : 228; Hubbs, 1915 : 466) и у берегов о. Хоккайдо (Ueno, 1971 : 99; Hikita, 1981 : 76), и только 1 экз. (ЗИН 45412) был пойман в северо-восточной части моря у мыса Хайрюзова. У южных Курильских о-вов (Линдберг, 1959 : 256). У тихоокеанских берегов Японии: вдоль побережья о Хоккайдо у Кусиро (Jordan, Hubbs, 1925 : 298), около Хидака (Hubbs, 1915 : 466), в зал. Утиура (Hikita, 1950 : 127, 1951b : 312, 1951а : 203; Sato, Kobayashi, 1956 : 16), у Хакодатэ (Линдберг, 1947 : 197); вдоль побережья о. Хонсю у Аомори (Jordan, Hubbs, 1925 : 298), около р-на Санрику (Ueno, 1971 : 99), у о. Кинкадзан (Sakamoto, 1983 : 217), в зал. Сэндай (ЗИН 48268) и около Абуроцубо в зал. Сагами (Franz, 1910 : 63).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Различают четыре вида. Три вида собственно
палтусовидных камбал — охотоморская
(Hippoglossoides elassodon), япономорская (Hippoglossoides dubius)
и северная (Hippoglossoides robustus) — живут в морях
Тихого океана.