Pseudopleuronectes herzensteini (Jordan & Snyder, 1901)
Желтополосая камбала (Rus),
Yellow striped flounder
(Eng)
Синонимы:
Limanda angustirostris Jordan & Starks, 1906
Limanda angustirostris maculosa Pavlenko, 1910
Limanda herzensteini Jordan & Snyder, 1901
Limanda japonica Schmidt, 1904
Limandella angustirostris Jordan & Hubbs, 1925
Pleuronectes herzensteini (Jordan & Snyder, 1901)
Pleuronectes japonicus Herzenstein, 1890
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 6. (Pleuronectiformes). Г.У.Линдберг, В.В.Федоров 1993 г.
5. Pleuronectes (Pseudopleuronectes) herzensteini (Jordan et Snyder, 1901) — Желтополосая камбала (рис. 45).
Pleuronectes japonicus (non Houttuyn, 1782), Herzenstein, Mel. Biol. Bull. Acad. Imp. Sci. St.-Petersb, 13, 2 1890 : 130 (частью: только ЗИН № 8730; Владивосток и № 1583-в, с; Хакодатэ).
Limanda herzensteini Jordan, Snyder, Proc. U. S. Nat. Mus., 23, 1901c : 746 (новое название для экз. Герценштейна). — Jordan, Starks, Bull. U. S. Comm. Fish., 22, 1904 : 623. — Охрямкин, Вестн. ДВФ АН СССР, 20, 1936 : 127, fig. 1 (синонимия, описание). — Таранец, Краткий определитель..., 1937 : 145. — Matsubara, Fish morph. a. hier., 2, 1955 : 1272 (Курильские о-ва, о. Сахалин, пров. Фукуи, Внутреннее море Японии, восточная часть п-ова Корея, Уссурийский зал, Китай).
Limanda japonica, Шмидт, Рыбы восточных морей..., 1904 : 234 (частью: исключая ЗИН № 1583-а, Хакодатэ).
Limanda angustirostris Kitahara in Jordan, Starks, Proc. U. S. Nat. Mus., 31, 1906a : 208, fig. 15 (Аомори). — Snyder, Proc. U. S. Nat. Mus., 42, 1912a : 440. — Jordan, Tanaka, Snyder, J. Coll. Sci. Imp. Univ. Tokyo, 33, 1, 1913 : 326, fig. 275. — Jordan, Thompson, Mem. Carnegie Mus., 6, 1914 : 309, fig. 82. — Hubbs, Proc. U. S. Nat. Mus., 48, 1915 : 485 (о. Садо, Цуруга, о-ва Оки, Аомори). — Солдатов, Линдберг, Обзор рыб..., 1930 : 405 (синонимия). — Wu, Pois. Heterosomes Chine..., 1932 : 106.
Limanda yokohamae, Jordan, Starks, Proc. U. S. Nat. Mus., 31, 1906a : 209 (частью: в синонимии исключая Pteuronectus yokohamae). — Jordan, Tanaka, Snyder, J. Coll. Sci. Imp. Univ. Tokyo, 33, 1, 1913 : 327 (частью: в синонимии исключая Pleuronectes yokohamae Günther).
Limanda angustirostris var. maculosa Павленко, Рыбы зал. Петр Великий, 1910 : 58, рис. 12.
Limandella angustirostris, Jordan, Hubbs, Mem. Carnegie Mus., 10, 2, 1925 : 299.
Pseudopleuronectes herzensteini, Norman, Flatfishes (Heterosomata)..., 1934 : 342, fig. 252 (синонимия, описание). — Чжан и др., Рыбы зал. Бохай, 1955 : 289, рис. 179 (описание). — Чжу и др., Рыбы Восточно-Китайского моря, 1969 : 530, рис. 396 (описание).
Pleuronectes herzensteini, Sakamoto, Mem. Fac. Fish. Hokkaido Univ., 31, 1-2, 1984 : 209 (положение в системе).
1583. Хакодатэ. 1863. П. Ф. Максимович. 1 экз. Синтип.
6163. Хакодатэ. 1881. А. Григорьев. 1 экз. Синтип.
8730. Владивосток. 1889. 1 экз. Синтип.
12355. Холмск. 9 VI 1901. П. Ю. Шмидт. 2 экз.
17720, 17722, 17723, 17725. Японское море, зал. Петра Великого. 5 VI — 29 IX 1913. ДВЭ. 11 экз.
17721, 17724, 17726, 17728. Японское море, зал. Петра Великого. 14 IV — 16 X 1912. ДВЭ. 8 экз.
19072. Японское море, 51°23' с. ш., 142°00' в. д. 29 VIII 1911. ДВЭ. 2 экз.
19149. Японское море, о. Хоккайдо, зал. Исикари. 16 XI 1914. С. Хатта. 2 экз.
23799. Тихий океан, о. Хонсю, зал. Муцу, Асамуси. 1930. 1 экз.
26217. Японское море, Приморье, бухта Преображения. 30 X 1934. Эксп. ЗИН. 1 экз.
33315. Японское море, Владивосток, базар. 1—10 IV 1907. Н. А. Пальчевский. 1 экз.
35611. Желтое море. Дальний. 18 V 1956. 1 экз.
41413. Охотское море, Южный Сахалин, оз. Изменчивое. 27 VI11 1970. 1 экз.
43746. Тихий океан, бухта Авача, Петропавловск-Камчатский. 6 VIII 1907. Н. Смирнов, А. Г. Вегак. 1 экз.
45163. Японское море, зал. Петра Великого, Широкая Падь. 20 VIII 1933. А. Кузнецов. 2 экз.
45164. Дальний Восток. Д. И. Охрямкин. 1 экз.
45165. Тихий океан, Южно-Курильский прол. 29 VI 1950. ИОАН СССР. 1 экз.
45592. Японское море, Татарский прол, г. Чехов. Глубина 62 м. 8 VIII 1947. КСЭ. 1 экз.
45593. Японское море, зал. Петра Великого. 29 VIII 1958. М. И. Легеза, Е. А. Дорофеева. 1 экз.
47062. Японское море 44°44'8 с. ш., 136°31'5 в. д. Глубина 80—140 м. 21 III 1982. Б. А. Шейко. 5 экз.
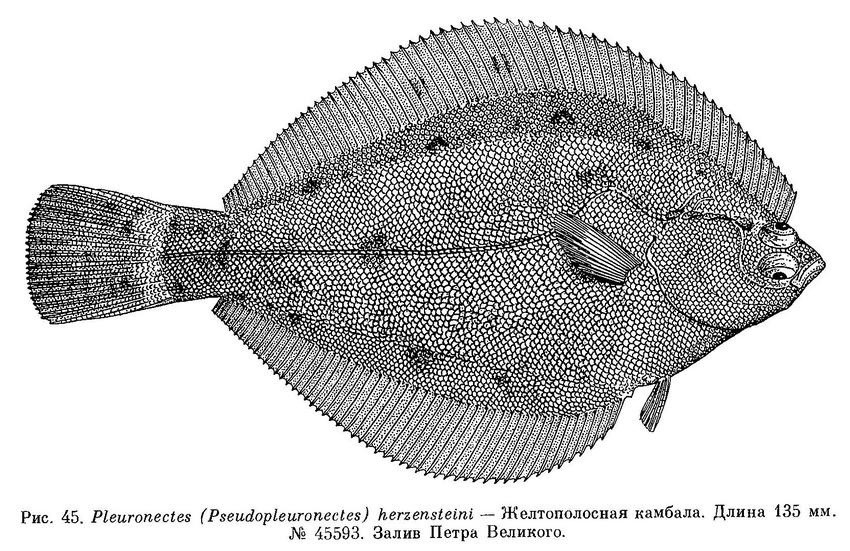
D 68—73; А 51—57 : Р 11; С 17; l.l. 75—78, выше боковой линии 22—25 рядов чешуи, ниже — 24—30; зубов 0 + 19—25 / 7—8 + 21—31; пилорических придатков 3+1 (Herzenstein, 1894 : 130; Шмидт, 1904 : 234; Охрямкин, 1936 : 126).
D 68—74; А 52—55 : l.l. 74—78, ниже боковой линии 21—24 ряда чешуи; зубов 0 + 19—22 / 5—6 + 22—24; жаберных тычинок ? + 6 (Kitahara in Jordan, Starks, 1906a : 208).
D 70; A 54; l.l. 75 (Павленко, 1910 : 58).
D (67) 69—75; A (51) 54—57; P 10—11 (5—8 ветвистых); С 19 (13 ветвистых); l.l. 68—75, выше боковой линии 20—26 рядов чешуи; жаберных тычинок ? + 6—8; зубов 0 + 14—23 / 2—6 + 19—24; пилорических придатков 3+1 (Norman, 1934 : 342).
D (65) 68—74; А (51) 52—58; Р 10—11; V 6; С 19 (13 ветвистых); l.l. 68—78, выше боковой линии 22—27 рядов чешуи; жаберных тычинок ? + 6—8; зубов 0 + 14—25 / 4—8 + 17—31; пилорических придатков (3) 4 (5); позвонков 37 (38) (Охрямкин, 1936 : 129).
D 66—70 (73), ср. 68.77; А 50—54 (58), ср. 52.74; Р (9) 10—11, ср. 10.26; l.l. (69) 71—75 (80), ср. 73.41; жаберных тычинок (8) 9—10 (11), ср. 9.61; позвонков 11 + (26) 27 (28), ср. 27.07 = (37) 38 (39), ср. 38.07 (Моисеев, 1937б : 148).
D 70—77; А 54—58; Р 10—11; V 6; С 18—20; l.l. 73—83; жаберных тычинок 3 + 6—7; зубов 0 + 15—22 / 3—8 + 20—24 (Чжан и др., 1955 : 289).
D 69; А 54; Р 10; V 6; С 20; l.l. 76; жаберных тычинок 3+6; зубов 0 + 14—23 / 2—8 + 19—24 (Чжу и др., 1963 : 530).
По нашим материалам (9 экз. длиной 111—277 мм): D 66—73, ср. 69.78; А 52—56, ср. 54.33; Р 9—11 (12), ср. 10.22 / 9—11 (12), ср. 9.89; С 3 + 7 (8) + 6 (7) + 3 = 19 (21), ср. 19.22; l.l. 72—80, ср. 75.33; выше боковой линии 20—25, ср. 23,0 рядов чешуи, ниже — 25—28, ср. 26.11; жаберных тычинок 3 (4), ср. 3.22 + (5) 6 (7), ср. 6.0 = 9 (10), ср. 9.22.
В стандартной длине: наибольшая высота тела 1.9—2.3 раза; длина головы 3.5—4.2. В длине головы: диаметр верхнего глаза 4.5—6.1 раза; длина грудного плавника 1.5—1.8 раза; длина верхней челюсти 4.1—4.9 раза, нижней челюсти 2.6—3.0 раза; длина дуги боковой линии 1.3—1.8 раза (Охрямкин, 1936 : 129).
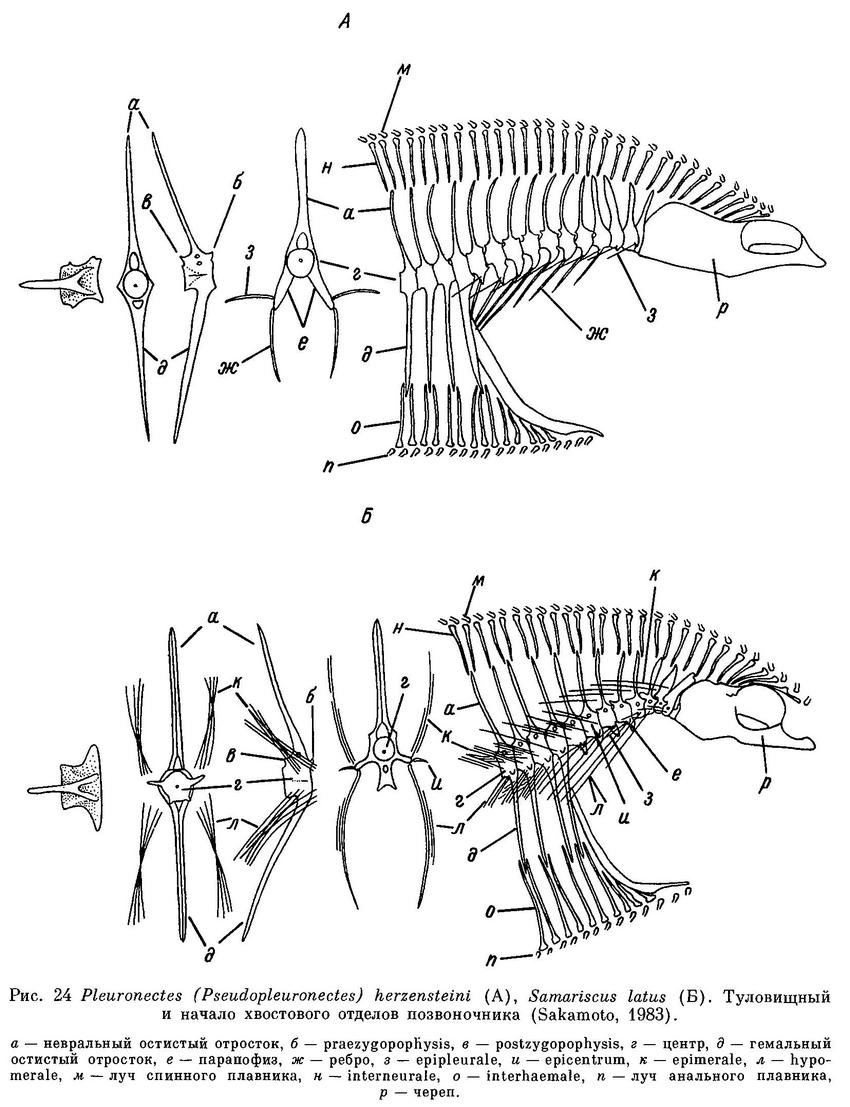
Рыло образует угол с линией спины над верхним глазом. Верхний глаз немного позади нижнего. Межглазничное пространство узкое, иногда покрыто рудиментарными чешуйками. От орбиты верхнего глаза назад до начала боковой линии тянется шероховатый костный гребень. Верхнечелюстная кость достигает переднего края нижнего глаза и немного заходит за передний край орбиты. Нижняя челюсть немного выдается. Зубы близко сидящие, резцевидные, не образующие сплошного режущего края. Жаберные тычинки широкие в основании, довольно короткие, с сильно заостренными кончиками. Нижнеглоточные зубы на несливающихся площадках, в 3 ряда, тупобугорчатые. Чешуя впереди не налегает друг на друга, циклоидная и только в последней трети тела более или менее черепицевидная, слабоктеноидная, с 1—7 ктениями. Боковая линия с высокой дугой впереди. Длина хвостового стебля 1.1 раза в его высоте (Охрямкин, 1936 : 128).
Окраска глазной стороны тела имеет 2 варианта: однотонная светло-коричневая без отметин или со следами темных отметин. На слепой стороне тела белая, обычно при жизни на теле вдоль спинного и анального плавников видны широкие золотисто-желтые полосы, которые у некоторых экземпляров могут заходить на голову, очень редко встречаются особи, совсем без полос.
Аномалии окраски и строения описаны японскими учеными (Nishimura, Ogawa, 1963; Imaoka, Nishimura, 1964; Honma et al., 1979).
Прибрежный вид, совершающий сезонные миграции. Летом распределяется на шельфе до глубин 70—80 м, скопления отмечаются на глубинах 20—50 м при температурах воды у дна от 1 до 19 °С. Осенние миграции начинаются в ноябре и заканчиваются в декабре-январе. Зимует на глубинах более 100—150 м при температуре воды у дна от 0 до 1 °С, но на меньших глубинах наиболее часто встречается при отрицательных температурах. Весенние миграции на шельф начинаются в апреле и к середине мая уже полностью заканчиваются. Молодь заходит в опресненные районы эстуарий и пресные озера.
В уловах поисковых судов преобладают рыбы длиной 24—33 см, массой 180—450 г, в возрасте 4—6 лет.
Нерест в Японском море в зал. Петра Великого — с мая по июль, сразу же после прекращения весенней миграции, обычно на глубинах 10—70 м при температуре воды у дна 2.6—18 °С. В Татарском прол. нерест более продолжительный — с мая по сентябрь с пиком в июне-июле. У берегов Японии в зал. Тояма нерест сдвинут на более ранние сроки: начало — во второй половине февраля, конец — в начале мая, пик — в марте-апреле. Икрометание порционное. Плодовитость 174—265 тыс. икринок. Икра пелагическая, диаметр 0.78—0.92 мм. Выклев первых предличинок через 4 сут и 10 ч после искусственного осеменения, конец — 4 сут и 22 ч. Длина выклюнувшихся предличинок 2.02—2.90 мм. Метаморфоз происходит при длине 9.0—10.4 мм, после чего личинки оседают на дно.
Типичный бентофаг. В зал. Петра Великого в июне основная пища состоит из оболочников и офиур, в сентябре — офиуры 47.9 %, полихеты 41.6 %, ракообразные 6.7 %, моллюски 3.5 %, прочие 0.3 %. Прекращение питания — с декабря по март. В Татарском прол. у берегов о. Сахалин большое значение в пище имеют моллюски (главным образом Spisula vladivostokensis) — 47.4 %, ракообразные (премущесвенно Pinnixa rathbuni) — 24.8 %, полихеты — 11.5 %, иглокожие (преобладает Amphiodia rossica) — 9.0 %, асцидии — 5 % и актинии — 2.3 %. В районе Южно-Курильского мелководья главную роль в пище играют иглокожие (преимущественно Amphiodia rossica) — 63.4 %, полихеты — 14.0 %, ракообразные (в основном Tecticeps glaber) — 13.9 % и моллюски — 6.6 %. Подробные сведения об особенностях биологии имеются в работах отечественных и зарубежных авторов (Мищенко, 1938; Демидова, 1939; Моисеев, 1941б, 1953а; Yusa, 1953; Микулич, 1956; Hikita, 1956; Фадеев, 1957, 1984, 1987; Дехник, 1959; Sato, 1960; Перцева-Остроумова, 1961; Hikita, Shibata, 1964; Wada, 1970а, 1970b; Иванков и др., 1972; Иванков, Курдяева, 1973; Иванков, Иванкова, 1974, 1975; Tominaga, Maeda, 1984; Kawasaki et al., 1985; Choi et al., 1986; Takahashi et al., 1987a).
Исследованы жирнокислотный состав нейтральных липидов (Hayashi, Yamada, 1975), свойства и специфичность ДНК печени (Rasskazov et al., 1968), биохимический полиморфизм (Коваль, Богданов, 1978, 1979), строение хромосом (Fukuoka, Niiyama, 1970; Sakamoto, Nishikawa, 1980) и паразитофауна (Nagasawa, Maruyama, 1987).
Длина 430 мм (Фадеев, 1987 : 115).
Распространение. В Японском море известен вдоль континентальных берегов от Пусаня (Mori,. Uchida, 1934 : 33), Пхоханя, Чхончжинь (Mori, 1952 : 181), в зал. Петра Великого (Моисеев, 1937б : 132) и вдоль Приморья (Солдатов, Линдберг, 1930 : 405) до северной части Татарского прол. (Фадеев, 1987 : 76), у островных берегов Японии вдоль о. Хонсю в р-не Санин (Mori, 1956а : 32; Minami, Takomaki, 1980 : 159), в заливах Вакаса (Tomiyama, Morino, 1970 : 387) и Тояма (Katayama, 1940а : 21), у берегов преф. Ниигата (Ito, 1970 : 34), о. Садо (Honma, 1956а : 54; Honma, Kitami, 1978 : 63), на банке Мукозе (Ouchi, 1963 : 129), о. Хоккайдо (Ueno, 1971 : 99), о. Сахалин (Николаев и др., 1958 : 5—7). Желтое море (Wu, 1932 : 106; Wang, 1933 : 50). В южной части Охотского моря в зал. Анива (Линдберг, 1959 : 256), у берегов о. Хоккайдо (Ueno, 1971 : 99; Hikita, 1981 : 77), где молодь заходит в устья рек (Hikita, 1956 : 163) и озера (Hikita, Shibata, 1964 : 30). В Южно-Курильском прол. и у тихоокеанских берегов о-вов Шикотан и Итуруп (Линдберг, 1959 : 256). У тихоокеанских берегов Японии во Внутреннем море (Matsubara, 1955 : 1272), у берегов и в эстуариях в преф. Миэ (Okada, Kurota, 1956 : 214) и далее на север у Аомори в зал. Муцу (Kitahara in Jordan, Starks, 1906 : 209) и вдоль берегов о. Хоккайдо (Ueno, 1971 : 99). В Восточно-Китайском море около Амакусы (Kikuchi, 1970 : 45).