Percis japonica (Pallas, 1769)
Японская лисичка (Rus),
Dragon poacher
(Eng)
Синонимы:
Agonus curillicus Tilesius, 1813
Agonus japonicus Bloch & Schneider, 1801
Agonus stegophthalmus Tilesius, 1813
Aspidophorus lisiza Lacepède, 1801
Aspidophorus superciliosus Cuvier, 1829
Cottus japonicus Pallas, 1769
Hippocephalus japonicus Jordan & Gilbert, 1882
Hippocephalus superciliosus Swainsоn, 1839
Phalangistes japonicus Pallas, 1814
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Лисичка японская — Percis japonicus (Pallas, 1769)
Тело удлиненное. Основания спинных плавников разделены промежутком, сопоставимым с длиной основания плавников. Костные щитки на теле, кроме тех, что лежат вдоль боковой линии, вооружены шипами, образующими продольные гребни. Число щитков в спиннобоковом ряду 34—40. D1 V—VI. D2 6—8. А 8—9. Окраска тела желтовато- или зеленовато-серая, у молоди с выделяющимися темными пятнами на теле и голове. Длина тела до 42 см. Обитает на дне в дальневосточных морях (преимущественно на песчаных грунтах).
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 5. (Scorpaeniformes). Г.У.Линдберг, З.В.Красюкова 1987 г.
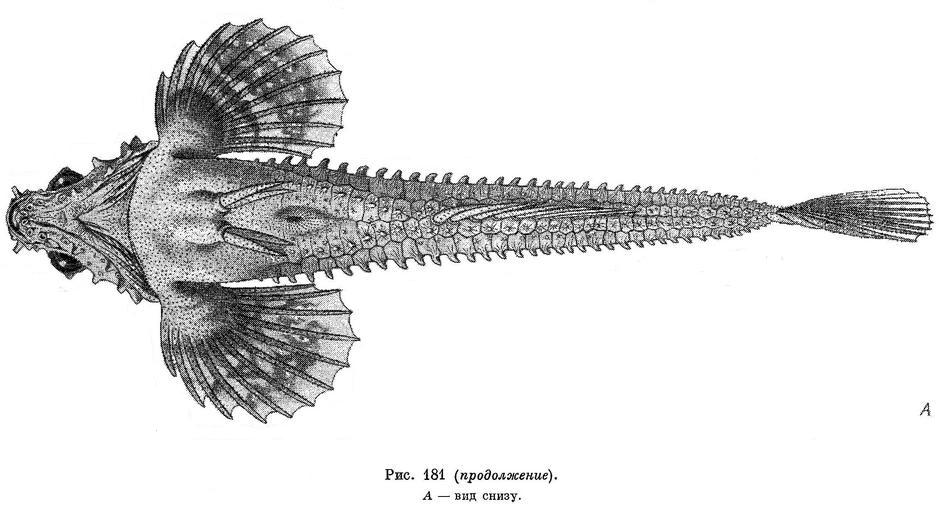
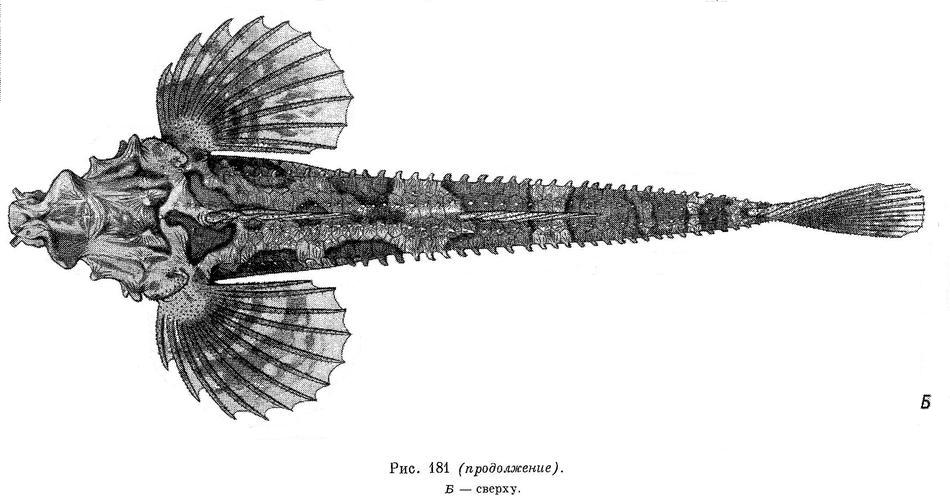
1. Percis japonicus (Pallas, 1769) — Японская лисичка (рис. 181, А, Б).
Cottus japonicus Pallas, Specilegia zoologica..., 1, 7, 1769 : 30, pl. 5, fig. 1—3 (сухой экземпляр с Курильских о-вов, добытый Стеллером в июне 1743 г.). — Gmelin, in: Linnaeus, Syst. Nat., cd. 13, 1788 : 1213 (по Палласу). — Walbaum, in: Artedi, Genera piscium, 3, 1792 : 387. — Tilesius, Mem. Soc. Nat. Moscou, 2, 1809 : 219 (no Палласу).
Agonus japonicus Bloch et Schneider, Syst. Ichthyol., 1, 1801 : 105 (по Палласу). — Tilesius, Mem. Acad. Sci., St.-Petersb., 4, 1813 : 416 (по Палласу).
Aspidophorus lisiza Lacepède, Hist. Nat., Poiss., 3, 1802 : 225 (по Палласу).
Phalangistes japonicus Pallas, Zoogr. Rosso-Asiat., 3, 1814 : 112. — Tilesius, Mem. Acad. Sci., St.-Petersb., 3, 1810 : 263 (по рукописи Палласа).
Agonus stegophtalmus Tilesius, Mem. Soc. Nat. Moscou, 2, 1809 : 219 (зал. Терпения); Mem. Acad. Sci. St.-Petersb., 4, 1813 : 427, pl. 12, fig. 1—3. — Крузенштерн, Путешествие вокруг света в 1803—1806 гг...., 1—3 части, Атлас, 1810—1813, табл. 87 (головач японский). — Günther, Catal. fishes Brit. Mus., 2, 1860 : 214. — Bean, Proc. U. S. Nat. Mus., 4, 1882 (1881) : 263 (Курильские острова, по Тилезиусу).
Agonus curilicus Tilesius, Mem. Acad. Sci., St.-Petersb., 4, 1813 : 416 (только название для Agonus japonicus).
Aspidophorus superciliosus Cuvier, in: Guvier, Valenciennes, Hist. Nat., Poiss., 4, 1829 : 215 (Курильские о-ва, по Палласу и Тилезиусу). — Bleeker, Scripsi Batavia Galensis, 1853 : 11 (Курильские о-ва), 1854—1857 : 14 (Курильские о-ва).
Hippocephalus superciliosus Swainsоn, Nat. Hist. Fishes..., 2, 1839 : 272 (по Guvier).
Hippocephalus japonicus Jordan a. Gilbert, Bull. U. S. Nat. Mus., 16, (1882) 1883 : 723 (по Guvier). — Сramer, Proc. Calif. Acad. Sci., 2nd ser., 4 (1893—1894) 1895 : 147, pl. X (экз. Краузе из Охотского моря). — Jordan, Evermann, Rep. U. S. Comm. Fish., 21 (1895) 1896 : 446 (по Крамеру).
Percis japonicus Sсоpоli, Introduct. hist. nat., 1777 : 454 (по Палласу). — Cramer, in: Jordan, Evermann, Bull. U. S. Nat. Mus., 47, 2, 1898 : 2034 (Охотское море, экз. Краузе, подробное описание). — Jordan, Gilbert, Fish. Bering sea, 1899 : 475. — Jordan, Snyder, Proc. U. S. Nat. Mus., 22, 1901 : 368 (Охотское море, о. Тюлений). — Шмидт, Изв. Рус. геогр. о-ва, 38, 5, 1903 : 516 (таблица, с.-з. часть Японского моря, Охотское и Берингово моря); Рыбы восточных морей..., 1904 : 125, рис. 7 в тексте, табл. 3, рис. 2 а, в (подробное описание). — Jordan, Starks, Proc. U. S. Nat. Mus., 27, 1904 : 57. — Гpацианов, Опыт обзора рыб..., 4, 1907 : 337 (распростр. по лит. данным). — Павленко, Рыбы зал. Петр Великий, 1910 : 68 (сев. часть Японского моря). — Gilbert, Burke, Bull. U. S. Bur. Fish., 30, 1912 : 65 (Авачинский зал.). — Jordan, Tanaka, Snyder, Catalogue..., 1913 : 295 (Ниигата). — Tanaka, Fig. a. Descr., 40, 1927 : 765, pl. 66, fig. 466 (Муроран). — Mori, Catal. fish. Korea, 1928 : 7 (Чхончжинь). — Линдберг, Таранeц, Список рыб..., 4, 21, 1929 : 256 (Японское море). — Солдатов, Линдберг, Обзор..., 1930 : 293 (зал. Петра Великого, Татарский прол.). — Rendahl, Ark. zool., 22А, 10, 1931 : 77 (Берингово м., м. Наварин). — Tanaka, J. Fac. Sci., Univ. Tokyo, 4, Zool., 3, 1, 1931 : 46 (сев. Япония). — Попов, Исслед. морей СССР, 19, 1933 : 146 (о. Фуругельма). — Mori, Uсhida, J. Chosen Nat. Hist. Soc., 19, 1934 : 19 (Чхончжинь). — Андрияшев, Исслед. морей СССР, 22, 1935 : 143 (на север до Анадырско-Лаврентьевского холодного мелководья); цит. соч., 25, 1937 : 314 (южная часть Анадырского зал.). — Таранец, Краткий определитель..., 1937 : 122 (Японское, Охотское и Берингово моря). — Шмидт, Рыбы Охотского моря, 1950 : 181 (синонимия, распространение, экология). — Matsubara, Fish morphol. a. hierar., 1955 : 1164, pl. 128, fig. 437 (зал. Тояма, Ниигата, Владивосток, Муроран, Курильские о-ва, Охотское и Берингово моря). — Abe, Enc. zool., 2, Fishes, 1958 : 63, fig. 181 (цветной рисунок). — Линдберг, Исслед. дальневост. морей СССР, 6, 2, 1959 : 254 (зап. поб. Южного Сахалина, зал. Анива, вост. поб. Южного Сахалина). — Chyung Moan Кi, The Fishes of Korea, 1977 : 549, pl. 122, fig. 1 (цветной рисунок).
12861. Охотское море, восточный берег Сахалина, севернее м. Терпения. 1899. В. К. Бражников. 3 экз.
12862. Японское море, мыс Поворотный. 10 V 1900. П. Ю. Шмидт. 1 экз.
17450. Японское море, зал. Петра Великого. Глуб. 68 м. 7 IV 1913. ДВЭ. 1 экз.
17451, 17452. Японское море, Татарский прол., западный берег Сахалина, мыс Хой. Глуб. 19 м. 5 VII и 1 IX 1913. ДВЭ. 2 экз.
23707. Охотское море, западный берег Камчатки. 3 VII 1915. ГЭВО. 2 экз.
25140. Японское море, Приморье, к востоку от зал. Владимира. 31 V 1931. 1 экз.
28956. Японское море, зал. Петра Великого. 29 IX 1930. Штейнберг. 1 экз.
31794. Охотское море, восточный берег о. Сахалин, бухта Низменная. Глуб. 127 м. 9 IX 1947. КСЭ. 1 экз.
33856. Берингово море, Олюторский зал. 25 VI 1952. Ин-т океанологии АН СССР. 1 экз.
33857. Берингово море, Камчатский зал. 23 V 1952. Ин-т океанологии АН СССР. 1 экз.
33858. Берингово море, Олюторский зал. 25 VI 1952. Ин-т океанологии АН СССР. 1 экз.
34373. Берингово море, Анадырский зал. Глуб. 53 м. 27 VIII 1933. К. И. Панин. 2 экз
35363. Охотское море, зал. Шелихова. 29 VII 1915. ГЭВО. 1 экз.
38833. Охотское море, 46°39' с. ш., 143°43' в. д. 5 VII 1960. Л. А. Лисовенко и В. В. Барсуков. 2 экз.
43247, 43248. Японское море, зал. Петра Великого. 7—15 VIII 1958. М. И. Легеза и Е. А. Дорофеева. 4 экз.
42484. Охотское море, 46°26' с. ш., 143°41' в. д. Глуб. 110 м. КСЭ. 1 экз.
42485. Охотское море, 46°58' с. ш., 143°20' в. д. Глуб. 80—93 м. IX 1947. КСЭ. 1 экз.
42486. 43487. Курильские о-ва, о. Парамушир, тихоокеанское побережье. Глуб. 130— 135 м. 13—14 VII 1954. М. И. Легеза. 2 экз.
44789. Курильские о-ва, о. Итуруп, тихоокеанское побережье у мыса Иодного. Глуб. 198—213 м. 27 IX 1949. КСЭ. 1 экз.
44790. Охотское море, восточный берег о. Сахалин, зал. Мордвинова. Глуб. 33—60 м. 2 IX 1947. КСЭ. 1 экз.
44791. Охотское море, зал. Анива. Глуб. 87—92 м. 23 VIII 1947. КСЭ. 2 экз.
46141, 46142. Курильские о-ва, о. Итуруп, зал. Рока. Глуб. 80—126 м. 7 IX 1948. КСЭ. 2 экз.
46206. Курильские о-ва, о. Шумшу. Глуб. 114 м. 2 VIII 1954. М. И. Легеза. 1 экз.
46207. Курильские о-ва, о. Парамушир. Глуб. 176 м. 8 VIII 1954. М. И. Легеза. 1 экз.
46221. Берингово море, 55°58' с. ш., 167°24' в. д. Глуб. 135 м. 25 VI 1969. В. В. Федоров и В. М. Макушок. 2 экз.
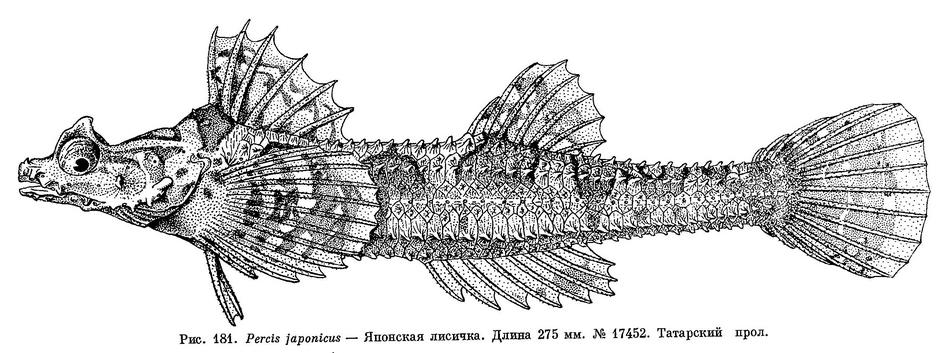
D V (3) — VI (27), 6 (2) — 7 (27) — 8 (1); А 8 (27) — 9 (3); Р 11 (1) — 12 (29); V I 2; С 11 (2) — 12 (28) основных лучей; l.l. 36—40; жаберных лучей 6; позвонков 12—13 + 27—28 = 39—41. (По 30 экз. с абсолютной длиной тела от 35 до 392 мм).
Все костные пластинки-щитки, покрывающие тело, за исключением пластинок, расположенных вдоль боковой линии, вооружены шипами, образующими продольные гребни. Число щитков в спиннобоковом ряду 34—40, чаще 36—38; срединноспинной гребень отсутствует, так как гребни спиннобоковых рядов позади спинного плавника не сливаются друг с другом, а, заходя за него, продолжаются до основания хвостового плавника; верхнебоковой ряд щитков с хорошо выраженным гребнем начинается на вертикали первой поры боковой линии и представлен 36—42, чаще 39 пластинками; нижнебоковой ряд начинается на вертикали 6—й поры боковой линии и состоит из 34—40, чаще 34—37 пластинок, гребни этого ряда хорошо развиты; брюшнобоковой ряд содержит 33—43, чаще 33—40 пластинок, и его гребень различим до основания хвостового плавника, и так как он не сливается с гребнем правой стороны тела, то срединнобрюшной гребень не образуется.
Из-за отсутствия срединноспинного и срединнобрюшного гребней тело в поперечном сечении в области туловища и хвостового стебля 8—гранное. Характер отношения частей тела в процентах к длине тела без С различен у особей с большой (210—392 мм) и малой длиной тела, что представлено в табл. 1.
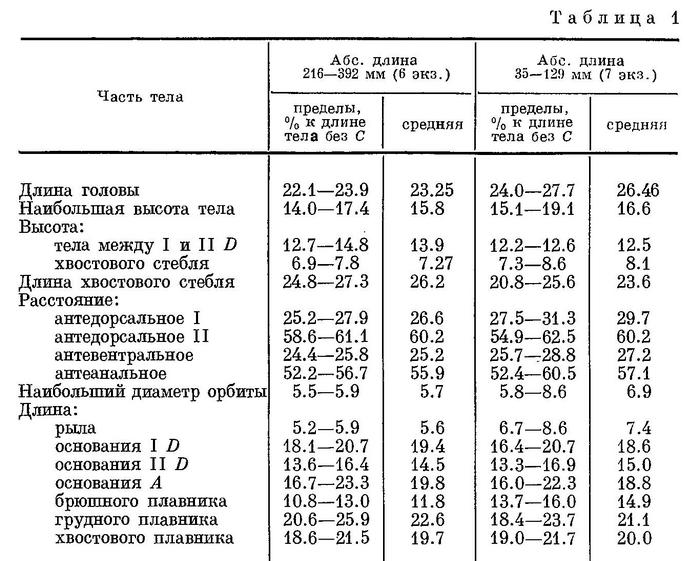
Тело удлиненное, сжатое с боков, наиболее широкое в области грудного пояса, где превышает наибольшую ширину головы; наибольшая высота тела находится на вертикали начала первого спинного плавника, который расположен сразу же за затылком. Высота тела между первым и вторым спинными плавниками 12.2—14.8 % длины тела без С, высота хвостового стебля 6.9—8.6 % этой длины, а его длина 21—27 %. Анальное отверстие находится ближе к основанию брюшных плавников, чем к началу анального, примерно в передней трети расстояния между этими плавниками. Промежуток между началом брюшного и началом анального плавников составляет около 32 % длины тела без С, а расстояние от ануса до начала анального плавника 21 % этой же длины. Голова небольшая, в среднем ее длина составляет 22.1—27.7 % длины тела без С, приплюснута, ее высота заметно меньше ее наибольшей ширины. Межглазничный промежуток у более молодых особей вогнут, с возрастом эта вогнутость становится все меньше заметной. Затылочные бугры также лучше выражены у молодых рыб, а с возрастом становятся едва заметны (при длине рыбы 400—430 мм). У экземпляров до 200 мм длиной надглазничные гребни с заостренной вершиной и направлены в стороны и вверх, а у более крупных рыб, 400 мм и более, они расположены почти горизонтально. Рот маленький, конечный, челюсти равной длины, и их задний край достигает вертикали переднего края орбиты. На челюстях и сошнике имеются мелкие волосковидные зубы, нёбные кости без зубов. Жаберные перепонки широко соединены между собой и свободны от межжаберного промежутка. Задний край предкрышечной кости с 3—4 крупными шипами; 1 шип имеется на щеке. Ноздри передней пары трубковидно удлинены. Продольный диаметр орбиты 5.5—8.6 % в длине тела без С. Спинные плавники отстоят друг от друга на расстояние, равное длине основания первого спинного плавника; начало анального плавника заходит вперед, вертикали начала спинного. Расстояние между вторым спинным и хвостовым плавниками превосходит расстояние между спинными плавниками примерно в 1.25 раза, расстояние между анальным и хвостовым плавниками в 1.1 раза более расстояния между вторым спинным и хвостовым плавниками.
Окраска рыб, фиксированных формалином, светлая коричневато-серая или фиолетовая. Верх головы и бока тела с темными коричневатыми пятнами неправильной формы; спинные, грудные и хвостовой плавники с большим числом темных пятен и пятнышек, сливающихся в темные полоски, брюшные и анальный плавники светлые, желтоватые, с небольшим числом темных пятнышек. Окраска молодых экземпляров при жизни желтовато- или зеленовато-серая с ярко выделяющимися пятнами на туловище и на голове (Шмидт, 19046 : 126).
Широкая плоская грудь, относительно невысокое тело и окраска характеризуют рыб этого вида экологически как обитателей придонных слоев воды, питающихся бентосными организмами. Как показали наблюдения, во время КСЭ, они обладают способностью к довольно быстрому передвижению.
В литературе имеются незначительные отрывочные данные по экологии и биологии. Указывается нахождение на глубине от 32 до 143 м, на каменисто-галечном, песчаном и илистом грунте, при температуре, близкой к 0°, но не отрицательной (Андрияшев, 1937 : 314; Шмидт, 1950 : 18).
Анализ материалов КСЭ дает возможность расширить и уточнить сведения по экологии данного вида для Южного Сахалина и южных Курильских островов. Встречается обычно единично, в трал за 1 ч траления попадает до 10 экз.
Температурный фактор при распределении этого вида играет ведущую роль. В районе западного побережья Южного Сахалина Percis japonicus встречен в августе при температуре +2.9 °С, в Приморье в том же месяце при +3.0 °С. В зал. Анива в августе и сентябре обнаружен при температуре воды от —0.3 до —1.0 °С; вдоль восточного побережья Южного Сахалина в сентябре и октябре от —1.5 до +3.6 °С.
Распределение по глубине носит закономерный характер и связано с температурным режимом данного района. В районе западного побережья Южного Сахалина в августе Percis japonicus встречен на глубине от 77 до 103 м. В Приморье взрослые экземпляры в августе обнаружены на глубине 68 и 105 м, у мыса Сюркум на глубине 21 м и температуре воды +14.7 °С пойман молодой экземпляр длиной 43 мм. Молодь держится ближе у берега и обитает при сравнительно высокой температуре. Средняя глубина, на которой обнаружен Percis japonicus в Татарском прол., 87 м. В зал. Анива в июле, августе и сентябре этот вид взят с глубины от 37 до 103 м. По-видимому, в этом участке Percis japonicus может быть встречен и на меньшей глубине, чем у западного берега Сахалина, что в свою очередь связано с температурным режимом зал. Анива, где отрицательной температурой характеризуются глубины примерно 25—30 м. В районе восточного побережья Южного Сахалина в сентябре и октябре этот вид обнаружен на глубине от 48 до 163 м, а в районе о. Итуруп (в октябре) от 90 до 206 м. Таким образом, в зал. Анива и у восточного берега Южного Сахалина Percis japonicus может встречаться на незначительных глубинах по сравнению с западным побережьем Сахалина и тихоокеанским берегом о. Итуруп, где этот вид не встречен на глубине менее 77 м. Подобного рода распределение по глубинам связано с температурным режимом каждого из этих участков.
Распределение по грунтам не носит закономерного характера. У западного побережья Южного Сахалина и в Приморье Percis japonicus встречен на песчаном с гравием и галькой и илисто-песчаном грунте. В зал. Анива в июле, августе и сентябре обнаружен на илисто-песчаном и илистом грунте. Вдоль восточного побережья Южного Сахалина этот вид в сентябре и октябре также обнаружен на смешанных грунтах — песчаном и илисто-песчаном. У тихоокеанского берега о. Итуруп встречен на песчаном грунте.
Соленость на местах поимок этого вида варьировала мало: от 32.45 до до 33.68 ‰.
Данные по размножению, которыми мы располагаем, незначительны. У западного побережья Сахалина 18 и 20 августа были пойманы 3 самки длиной от 350 до 365 мм, половые продукты которых находились в IV—V стадии зрелости. У самок Percis japonicus яичник непарный, расщеплен на переднем конце. Окраска овариальных икринок желтая и, чем крупнее икра, тем окраска ее становится менее интенсивной. Икринок коричневого цвета, о котором пишет П. Ю. Шмидт (1904 : 126), у особей этого вида нами не обнаружено. Икринки имели диаметр 1.0—3.0 мм и незначительное количество икринок было диаметром 0.3—0.5 мм. Нерест у Percis japonicus единовременный, летний. Возможно, летне-осенний. Плодовитость 3637—4935.
Percis japonicus может быть охарактеризован как морская донная рыба, обитающая на глубине от 37 до 213 м на смешанных грунтах, главным образом песчаном и илисто-песчаном, при температурах как отрицательных (—1.5 °С), так и положительных (до +3.6 °С). В фондовой коллекции ЗИН имеются экземпляры, добытые с еще меньшей глубины — 19 и 33 м (№№ 17452, 44790). Ведущий фактор при распределении этого вида — температурный; в участках, где низкие температуры, Percis japonicus встречаются на сравнительно небольших глубинах (зал. Анива и восточное побережье Южного Сахалина).
В обследуемых районах была выявлена определенная приуроченность рыб данного вида к тем или иным донным группировкам животных и растений, которые имели неоднозначный состав. Так, у западного берега Южного Сахалина предпочтительными оказались группировки 3 и 4. Донные группировки животных и растений в районах, обследованных КСЭ, выявлены и описаны З. И. Кобяковой (Гурьянова, Кобякова, 1955 : 74), их номера, приводимые нами, соответствуют номерам, приданным этим группировкам указанным автором в атласе.
3 — группировка моллюсков, морской козочки и змеехвостки; показатели — двустворчатый моллюск макома (Macoma calcarea), рачок морская козочка (Capreila sp.) и змеехвостка амфиоплус (Amphioplus macraspis). Кормовая ценность очень велика; здесь много червей (Maldanidae, Sternaspis, Onuphis conchylega, Сhone cincta и др.), мелких моллюсков (Yoldiella derjugini, Verticordia, Yoldia thraciaeformis, Yoldia johanni, Yoldia lanceolata, Crenella, Acila, Axinopsis, Serripes, Polynices, Admete и др.), змеехвосток (Amphioplus, Ophiura leptactenia, Amphiodia rossica, Amphiodia craterodmeta, Ophiura sarsi и др.), многолучевых звезд-солнечников. (Solaster dawsoni, Solaster pacificus), пятилучевых звезд (Pseudarchaster parellii), плоских морских ежей (Echinarachnius parma), бокоплавов (Ampelisca, Socarnes), кумовых раков (виды родов Diastylis, Campylaspis, Brachydiastylis), морских тараканов (Synidothea, Arcturus), звездчатых червей и др. Эта донная группировка характерна для чистых и илистых песков на глубинах 50—100 м только у западных берегов Сахалина. Летом является местом скопления камбал (главным образом большеротых), колючей акулы, скатов, терпугов, здесь кормовые поля камчатского краба.
4 — группировка морской лилии, змеехвостки и горгоноцефала; показатели — морская лилия (Heliometra glacialis maxima), змеехвостка Сарса (Ophiura sarsi) и горгоноцефал (Gorgonocephalus caryi); у восточного берега и в прол. Лаперуза также змеехвостка коротколучевая (Stegophiura brachyactis). Кормовая ценность ниже, чем 3 предыдущих группировок, хотя здесь порядочно червей (Onuphis conchylega, Pectinaria moorei, Terebellides strömi, Sternaspis, Maldanidae), моллюсков (Acila, Nucula, Yoldiella, Leda pernula, Pandora glacialis, Astarte banksi, Solariella, Bela, Cylichna, Dentalium и др.), змеехвосток (Ophiura quadrispina, Ophiopenia vicina), коротколучевых звезд (Ctenodiscus crispatus), кумовых раков (Diastylis bidentata, Brachydiastylis hexoceros), креветок, бокоплавов и др. Эта группировка характерна для сильно заиленных песков и илов на глубинах 60—200 м. Образует кормовые поля для некоторых более холодноводных камбал; встречаются треска, камчатский краб.
В зал. Анива предпочтительной оказалась группировка 7 и в меньшей степени группировка 2.
7 — группировка горгоноцефала, крупных брюхоногих и краба-стригуна; показатели — горгоноцефал (Gorgonocephalus caryi), крупные брюхоногие моллюски букциниды (Buccinium bayani, Argobuccinum oregonensis, Neptunea lirata, Neptunea eulimata) и краб-стригун (Chionoecetes opilio), по преимуществу его мелкие особи, иногда образующие большие скопления. Кормовая ценность очень высокая (бимасса до 1 кг/м2); масса червей (Maldanidae, Polynoinae, Тerebellidae, Sternaspis), мелкие моллюски (Leda, Nucula. Macoma calcarea, Solariella, Margarites, Trichotropis), мелкие голотурии (Chiridota pellucida, Myriotrochus), звездчатые черви (Phascolosoma hudsonianum), креветки, масса мелких крабов-пауков и др.
Характерна для сильно заиленных песков и илов на глубинах 50—120 м только у восточных берегов Сахалина, в зал. Анива и в прол. Лаперуза. Весной здесь скопления камчатского краба. Летом имеется много волосатых крабов промыслового размера; из рыб встречаются треска и минтай.
2 — группировка морских звезд и крупных брюхоногих; показатели — пятилучевая амурская звезда (Asterias amurensis), многолучевые звезды-солнечники (Solaster pacificus и Crossaster papposus) и крупные брюхоногие моллюски (Neptunea lirata, Buccinum bayani и Buccinum verkrüzeni). Кормовая ценность группировки очень велика; здесь обильны черви (Sigalion mathildae, Magelona, Ampharete, Pectinaria, Scalibregma, Glycera, Travisia и др.), мелкие моллюски (Yoldia johanni, Axinopsis, Liocyma fluctuosa, Nucula, Macoma, Solen, Siliqua, Spisula, Serripes grönlandicus, Polynices и др.), змеехвостки (Amphiodia rossica, Amphiodia craterodmeta, Ophiura sarsi vadicola и др.), креветки, раки-отшельники, кумовые раки, бокоплавы и др.
Характерна для песчаных грунтов мелководий (глубина 25—60 м). Здесь основные кормовые поля камбал, отчасти камчатского и синего краба и колючей акулы; промысловые скопления морской кубышки; камчатский краб образует основные скопления весной.
У восточного берега Сахалина была обнаружена предпочтительность Percis japonicus также к донной группировке 7, в меньшей степени к группировке 2 и 6.
6 — группировка гидроидов, губок, мшанок и аномии; показатели — прикрепленные формы (мелкие губки, гидроиды, мшанки) и двустворчатый, прикрепленный к камням моллюск аномия (Anomia macrochisma). Кормовая ценность низкая. В состав группировки входят многочисленные многощетинковые черви (Serpulidae, Polynoinae, Glycera, Stylarioides, Sabellaria cementarium и др.), моллюски (Astarte, Musculus, Arvella, Saxicava, Lepeta concentrica, Scala, Trichotropis insignis, морские хитоны и др.), раки-отшельники, мягкохвостые десятиногие раки (Dermaturus mandti, Hapalogaster grebnizkii), креветки (Sclerocrangon, Euatus, Nectocrangon crassa и др.), морские желуди (Balanus balanus, Balanus rostratus), морские тараканы (Arcturus, Tecticeps renoculus), бокоплавы (Stenothoidae, Ericthonius tolli, Melita), морские звезды (Henricia, Lethasterias nanimensis, Crossaster papposus и др.), змеехвостки (Ophiopholis aculeata, Ophiura maculata и др.), морские ежи, сложные асцидии.
Характерна для скал и каменистых грунтов на глубинах от 0 до 200 м.
У тихоокеанских берегов о. Итуруп японская лисичка обнаружена в донных группировках 13 и 17.
13 — группировка морских звезд, красных губок и раков-отшельников; показатели — пятилучевая амурская звезда (Asterias amurensis) и красные губки с живущими в них раками-отшельниками (Suberites domuncula + Pagurus pectinatus). Кормовая ценность очень велика; здесь изобилуют черви (Glycera, Travisia, Goniada, Magelona, Onuphis iridescens, тепловодные Sigalion mathildae, Lumbriconereis latreilli japonica), моллюски (Cardita, Mactra, Cardium californiense, Solen krusensternii, Soletellina olivacea, Macoma calcarea, тепловодные Turbo sangarensis, Tellina lutea venulosa и др.), крабики (Pinnixa, Oregonia gracilis, Doclea bidentata), креветки, различные виды раков-отшельников, бокоплавы; очень характерно обилие тепловодных морских звезд (Patiria pectinifera, Distalasterias nipon, Distalasterias elegans, Lysastrosoma anthosticta), масса амурских пятилучевых звезд (Asterias amurensis), змеехвосток (Amphiodia rossica, Amphiodia periercta) и т. д.
Характерна для песчаных грунтов на глубинах 30—40 м в Южно-Курильском проливе. Здесь кормовые поля камбал, зимой промысловые скопления колючего и волосатого краба, весной — камчатского краба, летом много колючей акулы, крупные промысловые скопления морской кубышки и японского морского гребешка (Cucumaria japonico и Pecten jessoensis).
17 — группировка бокаловидных губок, кораллов и аномий; показатели — крупные губки (Semisuberites), роговые кораллы (горгонарии), гидрокораллы (Allopora и Errinopora), одиночные кораллы (Caryophyllina clavus) и двустворчатый прикрепленный моллюск аномия (Anomia macrochisma). Много червей (Serpulidae, Sabellidae, Polynoinae, Travisia, Curratulus, Nereis, Sigalion и др.), морских тараканов (Synidothea, Arcturus, Tecticeps), бокоплавов (Ericthonius, Anonyx), раков-отшельников, креветок (Pandalus borealis eous, Sclerocrangon), моллюсков (Admete, Margarites, Polynices, Nucula, Lyonsia, Astarte, Pecten erythrocomatus), змеехвосток (Ophiopholis aculeata, Ophiacantha bidentata), морских звезд (Henricia, Pteraster, Evasterias), гидроидов, мшанок, брахиопод и актиний.
Характерна для каменисто-скалистых грунтов у о. Итуруп на глубинах 30—160 м. Летом здесь были отмечены промысловые скопления колючего, камчатского и волосатого краба; встречаются палтус, треска, минтай, палтусовидная камбала. В феврале—марте образуются нерестовые скопления трески.
Таким образом, для всех обследованных участков донными группировками животных, в которых были встречены японские лисички, были 2, 3, 4, 6, 7, 13 и 17. Из них высокой кормовой ценностью обладают группировки 2, 3, 4, 7 и 13, в которых в изобилии представлены черви, мелкие моллюски, креветки и бокоплавы, и биомасса достигает 1 кг/м2.
Длина до 420 мм (экз-ры КСЭ).
Распространение. В Японском море известен у берегов п-ова Корея (Mori, 1952 : 170), о. Фуругельма (Попов, 1933 : 146), в зал. Петра Великого (Солдатов, Линдберг, 1930 : 294; №№ 17450, 43247, 43248), у южного Приморья (№ 12862), у обоих побережий Татарского прол. (Солдатов, 1913 : 10; Линдберг, 1959 : 254; 1971 : 94; №№ 17451, 25140), у западного побережья о. Хоккайдо (Ueno, 1971 : 94), у Ниигаты (Matsubara, 1955 : 1164), у о. Садо (Honma, 1952 : 3), в зал. Тояма (Katayama, 1940 : 19), в р-не Санин (Mori, 1956а : 29). В Охотском море встречен в зал. Анива (Линдберг, 1959 : 254; № 44791), у мыса Левенорна (№ 43484), в зал. Мордвинова (№№ 43485, 44790), в зал. Терпения (№ 12861), у мыса Низкий (№ 31794), в северной части Охотского моря (№ 35363), у западного побережья Камчатки (№ 23707); Курильские о-ва (Kuronuma, 1943 : 111; Линдберг, 1959 : 254; Ueno, 1971 : 94; №№ 38833, 43486, 43487, 46141, 46142, 46206, 46207), упомянут для северного побережья о. Хоккайдо («Oshoro Maru» Cruis, 1969 : 390; Ueno, 1971 : 94). В Беринговом море встречается только по западному побережью, на север до мыса Наварин и южной части Анадырского зал., причем в последний не заходит (Андрияшев, 1937 : 314). По мнению А. П. Андрияшева (1935), продвижение далее к северу ограничено Анадырско-Лаврентьевским холодным мелководьем. Тихоокеанское побережье о. Хоккайдо от Аккэси (Sato, 1937 : 33) и Кусиро («Oshoro Maru» Cruis, 1967 : 161) до Муроран (Matsubara, 1955 : 1164).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
По отношению к температуре агоновые рыбы относятся к типичным холодолюбивым рыбам. В период гидрологического лета в районе Южного Сахалина и Курильских островов агоновые рыбы ловились при температуре у дна в среднем от 0,5 до 10,5°С, но нередки были поимки некоторых видов (Percis japonicus, Aspidophoroides bartoni, Podothecus gilberti) в водах с отрицательной температурой (до —1,5° С); максимальные температуры не превышали 17,3° С (Tilesina gibbosa).
Размеры агоновых рыб небольшие —
от 10 (Bothragonus) до 40 см (Percis
japonicus, Podothecus sachi).
Нерест агоновых рыб
обычно летний или летне-осенний,
единовременный (Percis japonicus, Leptagonus
decagonus), но бывает и весной, в марте
(Agonus cataphractus). Икра клейкая,
крупная, с толстой оболочкой.