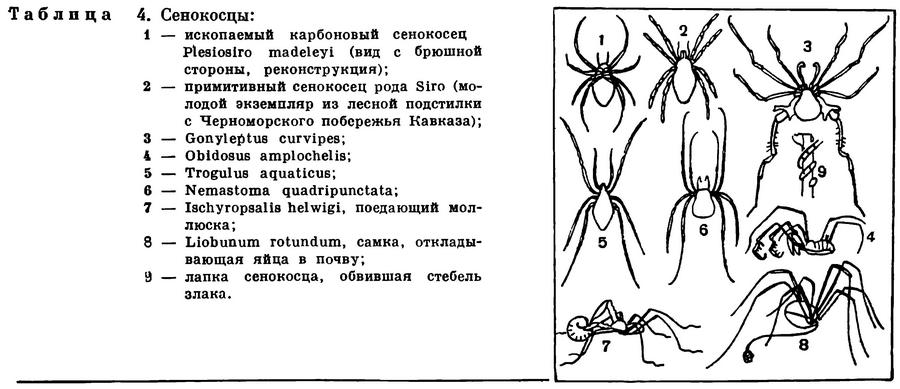
(Arachnida) Класс Арахниды, Паукообразные, Class Arachnida 11 отрядов
Жизнь животных. Том 3. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1969 г.
КЛАСС ПАУКООБРАЗНЫЕ, ИЛИ АРАХНИДЫ (ARACHNIDA)
Паукообразные, или арахниды (Arachnida), — это собрание всех наземных хелицеровых. Латинское название класса, в этой транскрипции теперь более принятое, прежде писалось Arachnoidea. Арахна — по-гречески «паук». В древнегреческих мифах это имя девушки, которая, по преданию, достигла такого высокого ткацкого искусства, что вызвала на состязание саму богиню Афину. Арахна выткала ткань не хуже Афины, но та в наказание за дерзость состязаться с богами не признала ее достоинств. В отчаянии Арахна хотела повеситься, тогда Афина превратила ее в паука, вечно ткущего свою паутину.
Их насчитывается около 35000 видов, и они очень различны по внешнему облику. Различают от 9 до 13 отрядов современных арахнид и несколько ископаемых. Среди них семь отрядов общеприняты: скорпионы (Scorpiones), кенении (Palpigradi), сольпуги (Solifugae), ложноскорпионы (Pseudoscorpiones), сенокосцы (Opiliones), рицинулеи (Ricinulei) и пауки (Aranei). Но в понимании нескольких группировок имеются разноречия. Это телифоны (Uropygi), фрины (Amblypygi) и тартариды (Tartarides), объединяемые в группу жгутоногих (Pedipalpi), и клещи (Acarina), на классификации которых мы остановимся в дальнейшем.
При большом разнообразии арахнид основные признаки хелицеровых свойственны им всем. Тело состоит из головогруди — просомы и брюшка — опистосомы, соединенных в области седьмого, предполового, сегмента. Усиков нет, глаза простые. Конечности головогруди — хелицеры, педипальпы и 4 пары ног — служат для захвата пищи и передвижения; конечности брюшка видоизменены, выполняют дыхательную и другие специальные функции и в значительной части атрофируются. Отличия арахнид от первичноводных хелицеровых обусловлены приспособлениями к жизни на суше. Главные из них: превращение жаберных ножек в легкие и далее замена их дыхательными трубками — трахеями; дальнейшая концентрация отделов тела; приспособление ног для передвижения по суше, а околоротовых конечностей для питания полужидкой пищей — содержимым жертвы, предварительно растворенным пищеварительными соками; ряд изменений жизненного цикла и общее уменьшение размеров.
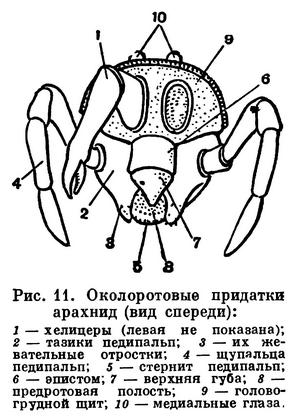
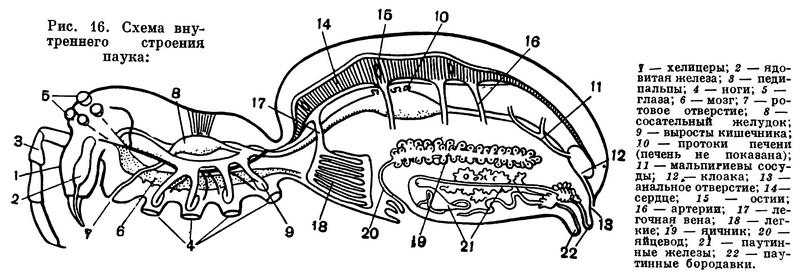
Строение головогруди (просомы) в общем однотипно. Обычно все 6 сегментов просомы слиты и она покрыта цельным головогрудным щитом. Но у сольпуг, кенений и части клещей слиты только четыре передних сегмента, отвечающие сегментам головы трилобитов. Они покрыты головным щитом (пропельтидием), а сегменты третьей и четвертой пар ног расчленены и имеют свои тергиты, — состояние, более примитивное, чем даже у меростомовых. Строение и функции околоротовых конечностей связаны со способом питания. Подавляющее большинство арахнид хищники, питающиеся живой добычей, главным образом насекомыми. При этом покровы жертвы разрываются и внутрь вводятся пищеварительные соки, которые обладают протеолитическим действием (способностью растворять белки). Затем разжиженное содержимое жертвы всасывается. Питание полужидкой пищей привело к тому, что у арахнид околоротовые конечности не приобрели характера челюстей в том виде, как у насекомых. Хелицеры служат для схватывания и разрывания добычи. Они обычно короткие, клешневидные; иногда концевой членик хелицер имеет вид когтя, на конце которого открывается проток ядовитой железы (например, у пауков), или хелицеры колющие, игловидные (у многих клещей). Тазики педипальп имеют отростки — эндиты, но они обычно не служат для пережевывания пищи, а ограничивают предротовую полость, на дне которой расположено ротовое отверстие (рис. 11). Верхняя стенка этой полости образована эпистомом с верхней губой. Изнутри на эндитах педипальп и в глотке имеются волоски, через которые фильтруется полужидкая пища. После питания твердые частички счищаются с волосков и выбрасываются. Щупальца педипальп служат органами осязания, но иногда они участвуют в передвижении (сольпуги, кенении), или они хватательные, с клешнями (скорпионы, ложноскорпионы) или когтевидными выростами (жгутоногие). В строении ног характерно образование членистой лапки с коготками — приспособление к хождению по суше. Жевательная функция ног у арахнид утрачена, но коксэндиты у примитивных форм частично сохраняются. Ноги, особенно передние, богато оснащены осязательными волосками и наряду со щупальцами педипальп имитируют исчезнувшие усики.
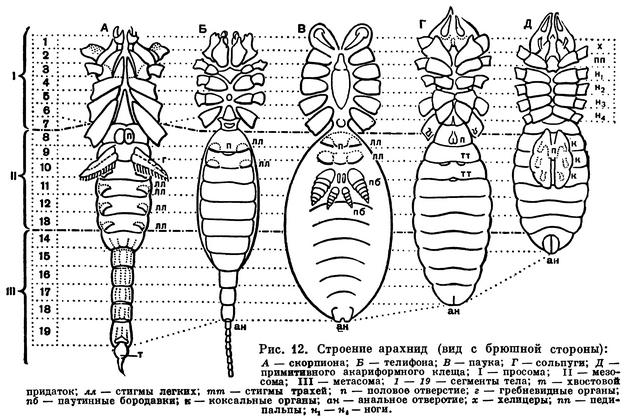
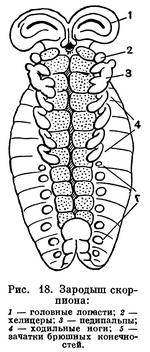
Наибольшее разнообразие наблюдается в строении брюшка (рис. 12). Число сегментов его заднего отдела — метасомы — постепенно сокращается, и брюшко укорачивается. Только у скорпионов метасома («хвост») состоит, как у эвриптерид, из 6 сегментов и концевого членика и общее число сегментов тела равно 19. У жгутоногих и пауков в метасоме 5 сегментов, у сольпуг, кенений, сенокосцев и рицинулей — 4, у ложноскорпионов — 3, у примитивных клещей — 2—1, у некоторых паразитических клещей исчезает не только метасома, но и несколько сегментов мезосомы. У некоторых отрядов оставшиеся сегменты метасомы имеют вид маленького стебелька (постабдомена), иногда с членистой хвостовой нитью. Степень развития соединительного предполового сегмента различна. У скорпионов, сенокосцев и рицинулей он не выражен, у ложноскорпионов и части клещей он, напротив, развит, как соседние. В обоих случаях брюшко широким основанием примыкает к головогруди. У других арахнид этот сегмент меньше остальных и иногда представлен узким стебельком, подвижно соединяющим головогрудь и брюшко. На этом основании арахнид иногда явно искусственно делят на сидячебрюхих и стебельчатобрюхих. Степень слияния сегментов брюшка различна. Обычно их границы хорошо различимы, но у большинства пауков и клещей границы сегментов исчезают. У многих клещей и головогрудь с брюшком сливаются в одно целое.
Конечности брюшка у арахнид превращены в легкие и другие специальные образования. Они имеются только на сегментах мезосомы. Наиболее полный набор видоизмененных брюшных конечностей сохранился у скорпионов: половые крышечки на восьмом сегменте, гребневидные органы на девятом, четыре пары легких на десятом — тринадцатом сегментах. У телифонов, фринов и четырехлегочных пауков по паре легких имеется на восьмом и девятом сегментах, у тартарид и двулегочных пауков — пара легких на восьмом сегменте, причем у последних на месте легких на девятом сегменте образуются трахеи. У всех пауков конечности десятого и одиннадцатого сегментов превращены в паутинные бородавки. У других арахнид легкие исчезают. Иногда на их месте открываются трахеи (сольпуги, сенокосцы), в других случаях трахеи не имеют отношения к легким. Рудиментами конечностей брюшка являются еще так называемые коксальные органы, имеющиеся на восьмом — десятом сегментах у кенений и части клещей, у которых на брюшке нет органов дыхания. Они имеют вид маленьких выпячивающихся мешочков, наполняемых гемолимфой, и, по-видимому, служат органами чувств, определяющими влажность (гигрорецепторы). Они приурочены к тазикам ножек и при утрате последних остаются на их месте. У кенений они расположены на брюшке открыто, а у части клещей входят в состав сложного наружного полового аппарата, указывая на участие в его образовании трех пар видоизмененных конечностей восьмого — десятого сегментов. Заметим, что система подобных коксальных органов наиболее полно развита у некоторых многоножек и низших насекомых. Наличие коксальных органов на брюшке кенений и низших клещей указывает на то, что у этих мелких форм легких никогда не было.
Покровы арахнид образованы хитинизированной кутикулой со слоем подстилающих плоских клеток — гиподермой. Щитки и членики конечностей образованы твердой кутикулой, в сочленениях между ними кутикула более тонкая, эластичная. Наиболее тверда кутикула коготков, пальцев хелицер. У некоторых скорпионов в кутикуле имеются отложения углекислого кальция. Строение наружного скелета арахнид различно и в той или иной мере отражает степень слияния сегментов. Головогрудь защищена сверху твердым щитом, между тазиками ног иногда имеется грудная пластинка (стернум). Тергиты и стерниты брюшка обычно посегментно расчленены. Но у части сенокосцев и многих клещей тергиты срастаются друг с другом и с головогрудным щитом, образуя цельный спинной щит, а иногда к ним примыкает и часть стернитов брюшка, так что тело заковано в панцирь. В других случаях, например у пауков, щиты имеются на головогруди, а брюшко покрыто эластичной кутикулой. Среди низших почвенных и эндопаразитических клещей есть формы с очень тонким покровом, лишенные щитков. Установлено, что кутикула арахнид имеет особые оптические свойства, она обладает двойным лучепреломлением, т. е. поляризует проходящий через нее свет. Благодаря этому в поляризационном микроскопе определенные элементы покрова светятся в темном поле. Французский исследователь Ф. Гранжан, впервые обнаруживший это явление у клещей, приписывает его наличию в кутикуле хитина особой структуры, названного им актинохитином или лучистым хитином. Распределение актинохитина в скелете закономерно. У одних отрядов он имеется только в толще щитков, у других только в волосках и щетинках, у третьих и там и тут.
Будучи хищниками, арахниды принуждены справляться подчас с сильной добычей. Мускулатура хорошо развита, особенно мышцы головогруди, приводящие в движение конечности.
Разнообразны железы покровного (гиподермального) происхождения: железы предротовой полости пауков, лобные и анальные железы жгутоногих, пахучие железы сенокосцев и др. К этой же категории относятся ядовитые и паутинные железы. Первые имеются у скорпионов в концевом членике брюшка, у пауков, у которых открываются на крючках хелицер, у ложноскорпионов, некоторых клещей. Ядовитый аппарат скорпионов и пауков служит весьма эффективным средством нападения и защиты. Паутинные железы имеются у ложноскорпионов, некоторых клещей и пауков. У последних они особенно развиты и открываются многочисленными отверстиями на брюшных паутинных бородавках.

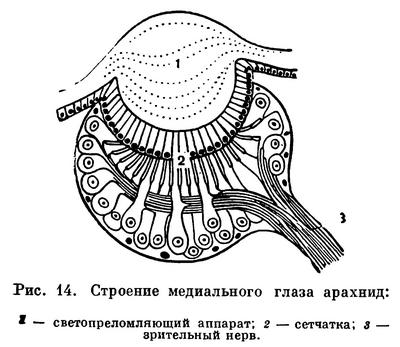
Органы чувств образуются путем дифференциации клеток покровного эпителия. Глаза имеются на просоме в разном числе: до 5 пар у скорпионов, обычно 4 пары у пауков и жгутоногих, 2—1 пара у большинства других; кенении, многие клещи, рицинулеи слепы. Глаза построены по типу простых глазков (оцелл). В глазу имеется диоптрический аппарат — хрусталик, образованный прозрачным утолщением кутикулы, и стекловидное тело, а под ним слой чувствительных клеток (ретина), связанных волокнами зрительного нерва с мозгом (рис. 14). Пара срединных (главных) глаз и боковые различаются в деталях строения. Зрительные возможности большинства арахнид ограниченны, они воспринимают вариации освещенности и движение. Лучше других видят сольпуги и бродячие пауки. Среди последних пауки-скакуны обладают предметным зрением, но различают форму на сравнительно близком расстоянии.
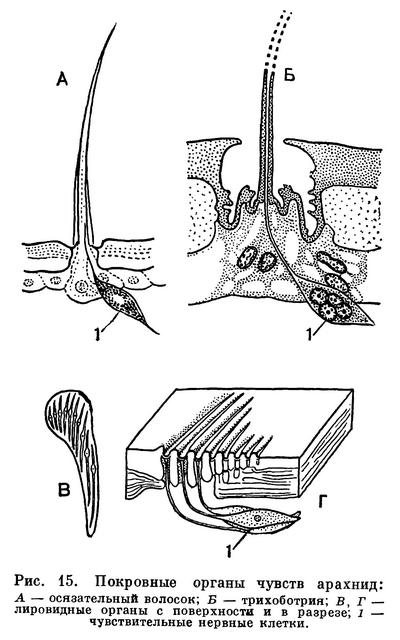
Слабое зрение компенсируется осязанием, играющим первостепенную роль в поведении арахнид. На теле и конечностях имеются многочисленные осязательные волоски, к основаниям которых подходят нервные окончания чувствительных клеток. По величине и форме эти волоски у арахнид чрезвычайно разнообразны. Кроме того, имеются специальные волоски, воспринимающие колебания, — трихоботрии. Эти своеобразные органы обычно в определенном количестве имеются на педипальпах и ногах, иногда на туловище (у части клещей). Длинный торчащий волосок, иногда утолщенный на конце, прикрепляется тонкой мембраной на дне воронковидного углубления. Малейшее сотрясение или дуновение воздуха приводит его в колебания, которые воспринимаются группой чувствительных клеток. У арахнид имеются и органы химического чувства, обонятельные и вкусовые. Первыми считают так называемые лировидные органы, многочисленные на туловище и конечностях. Это микроскопические щели в кутикуле, затянутые тонкой мембраной, к которой подходит окончание чувствительной клетки. Лировидным органам, правда, приписываются и другие функции, в частности механорецепторов, воспринимающих степень натяжения кутикулы. Более сложно устроены обонятельные тарзальные органы на лапках передних ног. Чувствительные вкусовые клетки найдены в стенках глотки у пауков.
Нервная система концентрированна. Отсутствие обособленной головы, усиков и сложных глаз привело к тому, что надглоточный ганглий (головной мозг), иннервирующий у членистоногих эти органы, в той или иной мере объединяется с головогрудной нервной массой. У скорпионов имеется парный надглоточный ганглий, связанный тяжами с подглоточным ганглиозным скоплением, и 7 ганглиев брюшной нервной цепочки. У сольпуг, кроме общей нервной массы, остается один брюшной узел; у большинства арахнид
вся нервная цепочка сливается в головогрудную массу.
Кишечник подразделяется на переднюю, среднюю и заднюю кишку. Ротовое отверстие ведет в расширение — снабженную мышцами глотку, которая служит для насасывания полужидкой пищи. Глотка переходит в тонкий пищевод, который у некоторых форм, например пауков, имеет еще расширение — сосательный желудок. Средняя кишка обычно образует несколько пар слепых выростов, увеличивающих ее вместительность и всасывающую поверхность. В брюшке слепые выросты кишечника хорошо развиты и образуют крупный железистый орган — печень. Клетки печени выделяют пищеварительные ферменты, и в них происходит внутриклеточное переваривание пищи. Задний отдел средней кишки образует клоаку, в которой скопляются экскременты и экскрет выделительных мальпигиевых трубок. Отбросы выводятся через короткую заднюю кишку и анальное отверстде. В кишечник арахнид в большинстве случаев попадает только жидкая пища, все крупные частички задерживаются фильтрами предротовой полости и глотки. Будучи прожорливыми хищниками, арахниды способны принимать большое количество пищи и потом долго голодать. Последнее возможно благодаря накоплению питательных веществ в запасной ткани, подобной жировому телу насекомых.
Выделительными органами служат коксальные железы и мальпигиевы сосуды. Первые, как говорилось, представляют остатки целомодуктов — посегментно расположенных выделительных органов предков членистоногих — кольчатых червей. Они состоят из экскреторного мешочка, извитого протока (лабиринта) и выводного канала и сохраняются обычно лишь в числе 1—2 пар, открываясь у оснований ног. Мальпигиевы сосуды арахнид — новообразование. Это 1—2 пары слепо замкнутых, иногда ветвящихся трубочек, которые открываются в кишечник близ клоаки. В клетках их стенок накапливаются экскреты, выводимые затем в клоаку. Выделительную функцию выполняют, кроме того, кишечник, печень, клоака и особые клетки — нефроциты, имеющиеся в полостях между органами. Главный продукт выделения арахнид — гуанин. Это вещество в организме находится в определенных биохимических отношениях с черным пигментом меланином, вместе с ним обусловливая окраску покровов.
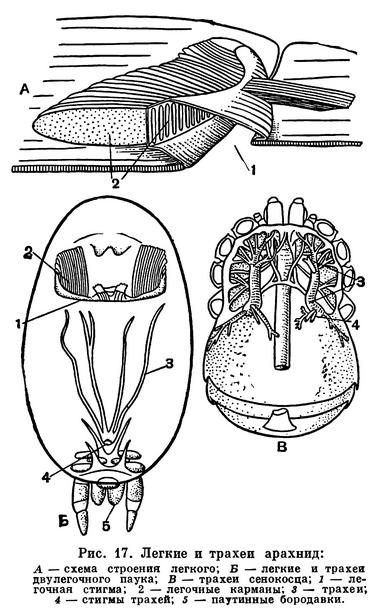
Строение дыхательной и кровеносной систем тесно связано. Органы дыхания арахнид двойственны по природе. Это органы локализованного дыхания — легкие, образовавшиеся из брюшных жаберных ножек водных форм, и органы диффузного дыхания — трахеи, возникающие вновь, как более совершенное приспособление для дыхания атмосферным воздухом (рис. 17). Каждый легочный мешок вдается внутрь от щелевидной стигмы. От его внутренней стенки отходят многочисленные листовидные карманы, сложенные подобно страницам книги. В карманах циркулирует кровь, а между ними проникает воздух. Трахеи представляют собой трубки, неветвистые или ветвящиеся, которые доставляют воздух непосредственно к органам и тканям. Их стенки образованы продолжением наружного покрова и выстланы кутикулой, обычно имеющей опорные утолщения: трахеи легко изгибаются, и стенки их не спадаются. Число пар легких, как говорилось, различно, и в ряде случаев они отсутствуют, заменяясь трахеями, а у некоторых мелких форм нет ни легких, ни трахей, и дыхание кожное (кенении, часть клещей). Число трахейных стволов также различно, причем они могут открываться стигмами в разных местах: на сегментах брюшка, по бокам головогруди, у оснований хелицер, что указывает на независимое их происхождение у разных арахнид. В некоторых случаях трахеи занимают место легких (у сольпуг, двулегочных пауков) и, по-видимому, возникли из них, хотя как органы не гомологичны легким. В целом у арахнид трахейная система гораздо менее развита, чем у насекомых, обычно не наблюдается у них и дыхательных сокращений брюшка, которые так характерны для многих насекомых.
Кровеносная система хорошо развита у крупных форм, дышащих легкими. Имеется пульсирующий спинной сосуд — сердце с несколькими парами боковых отверстий — остий, снабженных клапанами. От сердца отходят передняя и задняя аорты и несколько посегментных пар артерий, которые ветвятся. Кровь (гемолимфа) из сердца через артерии изливается в систему лакун — пространств между органами, собирается в легочные синусы, обогащается кислородом в легочных карманах, по легочным венам возвращается в околосердечное пространство и через остии — в сердце. По мере перехода от легочного дыхания к трахейному кровеносная система становится менее развитой, уменьшается число артерий и остий сердца. Так, у скорпионов и большинства жгутоногих остий 7 пар, у сольпуг — 6, у пауков — от 5 до 2, у сенокосцев — 2 пары, у клещей сердце в виде маленького мешочка с парой остий или оно отсутствует. Кровь обычно бесцветна, в ней имеются кровяные клетки нескольких типов.
Арахниды раздельнополы. Половые железы — яичники и семенники — расположены в брюшке и в исходном состоянии парны. В ряде случаев наблюдается объединение правой и левой гонад. Так, у самцов скорпионов семенники парные, каждый состоит из двух трубок, связанных перемычками; у самок яичник один и состоит из трех трубок, из которых средняя есть результат продольного слияния двух трубок. У многих арахнид парные гонады срастаются концами в кольцо. Парные яйцеводы и семепроводы открываются непарным половым отверстием на восьмом сегменте. Устройство выводной части половой системы и совокупительные приспособления разнообразны. У самок обычно имеется расширение яйцеводов — матка и семеприемники, в которых сохраняется сперма.
Биология размножения разнообразна. Наружное оплодотворение, свойственное водным хелицеровым, сменяется на суше внутренним, сначала свободным сперматофорным, а далее различными способами копуляции. При сперматофорном оплодотворении сперматозоиды заключены в специальный мешочек — сперматофор, выделяемый самцом и защищающий сперму от высыхания. В наиболее примитивных случаях, у многих клещей, живущих во влажной почве, ложноскорпионов, самцы оставляют сперматофоры на субстрате, а самки захватывают их наружными половыми органами. При этом особи совершают характерные взаимные движения — брачные танцы. У многих арахнид самец тем или иным способом переносит сперматофор в половое отверстие самки, что чаще делается с помощью хелицер, имеющих для этого особые приспособления. Наконец, у ряда форм сперматофоров нет, и сперма вводится с помощью специальных копулятивных органов. Последние образуются либо в составе собственно наружного полового аппарата, либо для копуляции служат совсем иные органы, например концевые членики щупалец педипальп у самцов пауков, третьей пары ног у рицинулей. Копуляция сопровождается подчас очень сложным поведением партнеров и проявлением целой цепи инстинктов, особенно у пауков.
У некоторых клещей наблюдается партеногенез, т. е. развитие неоплодотворенных яиц. Иногда самцы появляются периодически, а в остальное время развитие партеногенетическое. Есть также формы, у которых самцы вообще неизвестны.
Арахниды в большинстве откладывают яйца, но есть и живородящие формы (часть скорпионов, некоторые клещи и др.). Плодовитость очень различна. Чаще она составляет несколько сотен яиц, но есть некоторые паразитические клещи, у которых самка откладывает до 30 000 яиц. Обычно яйца откладываются общей кладкой по нескольку десятков или сотен. В течение жизни бывает одна или несколько кладок, что зависит от биологии вида и длительности жизни самки. Среди мелких арахнид есть формы, у которых яйца созревают и откладываются по одному; общая плодовитость при этом обычно невелика. Яйца арахнид богаты желтком. В связи с этим основные процессы формирования организма протекают в эмбриональный период, и из яйца вылупляется особь, похожая на взрослую, только меньшего размера. Послезародышевое развитие прямое, без метаморфоза, сопровождается ростом и линьками. Этот тип развития унаследован арахнидами от водных хелицеровых, у которых, как говорилось, произошла эмбрионизация первичного метаморфоза. Только у клещей в связи с малыми размерами и меньшим запасом желтка в яйце послезародышевое развитие представляет своеобразный метаморфоз.
В связи с большим запасом желтка дробление яйца в большинстве случаев поверхностное: ядра, делясь, выходят на поверхность желтка, где образуется слой клеток (бластодерма). Желток при этом обычно не делится. Зародышевые листки арахнид были открыты впервые у скорпионов в 1870 году И. И. Мечниковым и в дальнейшем найдены у других форм. Изучение эмбрионального развития позволяет лучше понять строение взрослых форм. Например, в тех случаях, когда у взрослых исчезает сегментация, она бывает выражена у эмбриона (пауки и др.). В эмбриональном развитии удается проследить, как зачатки конечностей брюшка превращаются в легкие и другие органы и т. д. Большой интерес представляет эмбриональное развитие низших клещей, сохранившее примитивные черты, на чем мы остановимся в дальнейшем.
У многих арахнид наблюдается охрана потомства. Самка откладывает яйца в специально вырытую норку и остается при них. У пауков яйца оплетаются паутинным коконом, который обычно самка охраняет в гнезде или носит при себе. Вылупившиеся молодые особи обычно в первое время не питаются активно, пищей им служит эмбриональный желток, остающийся в кишечнике. Молодь в этот период держится в гнезде или на теле матери (у скорпионов, телифонов, ряда бродячих пауков и др.) и, только перелиняв, переходит к самостоятельной жизни.
По общему характеру жизненного цикла арахниды весьма различны. В этом отношении можно наметить два типа, между которыми имеются переходы. Один крайний тип представляют крупные долговечные формы, живущие по многу лет и периодически размножающиеся. Таковы, например, некоторые тропические скорпионы, жгутоногие, крупные пауки-птицеяды. Среди последних некоторые живут до 20 лет и всю жизнь не теряют способности линять. При этом типе жизненного цикла индивидуальное развитие длительное и половая зрелость достигается после долгого роста. Особи обычно не образуют массовых скоплений, и вообще численность таких форм в природе относительно невелика. Этот долговечный способ жизни, связанный с крупными размерами или даже гигантизмом и многократным периодическим размножением, очевидно, унаследован арахнидами от водных хелицеровых и вообще не характерен для наземных членистоногих. Среди водных форм меростомовые, а также многие крупные ракообразные по жизненному типу именно таковы. На суше этот тип удержался только у некоторых арахнид, живущих главным образом во влажных тропиках, где условия жизни, так сказать тепличные. Среди трахейнодышащих известную аналогию представляют некоторые гигантские тропические многоножки — кивсяки. Заметим, что среди наземных животных по пути долговечной жизни при крупных размерах особей пошли позвоночные, но у них имелись к тому свои особые биологические предпосылки.

Для большинства арахнид характерен другой, противоположный жизненный тип, который в своих крайних вариантах представлен у многих клещей. Эти мелкие арахниды недолговечны, но зато развиваются очень быстро, причем поколения следуют друг за другом, пока есть подходящие условия. Как только условия становятся неблагоприятными, все активные особи погибают, но остаются покоящиеся яйца или специальные формы (молодые или взрослые), способные переносить неблагоприятные условия (высыхание, низкую температуру, отсутствие пищи и т. п.). При наступлении подходящих условий покоящиеся формы пробуждаются, начинается активная жизнь, размножение, и в короткий срок численность восстанавливается. Этот эфемерный тип жизни, связанный с малыми размерами, высокой скоростью развития и обычно с наличием специальных переживающих стадий, очень характерен для наземных членистоногих вообще, в частности для насекомых. Он, несомненно, есть важнейшее биологическое приспособление к жизни на суше, где условия гораздо более изменчивы, чем в море. Помимо всякого рода случайных изменений среды, при выработке этого жизненного типа сказываются периодические сезонные явления, особенно резкие в умеренном климате. Большинство арахнид, например пауков, подобно многим насекомым, представлено односезонными формами, которые в течение лета успевают проделать одну генерацию. Зимуют обычно яйца или молодь, которая размножается в следующем году. Реже у арахнид бывает 2—3 генерации в год, и только некоторые клещи успевают проделать много генераций.
Не вызывает сомнений, что все арахниды произошли от водных хелицеровых. Как мы видели, переход к жизни на суше сопровождался выработкой многих приспособлений. Жаберное дыхание сменилось легочным, а далее стало дополняться и заменяться трахейным. Число сегментов тела сократилось, брюшко концентрировалось как единый отдел. Произошла дальнейшая специализация конечностей головогруди. Ноги потеряли жевательную функцию, лапки расчленились, и возникло стопохождение. Широкое распространение получило внекишечное разжижение пищи, и околоротовые конечности приспособились к этому своеобразному способу питания. Дифференцировалась сложная система кожных органов чувств, особенно осязательных. Изменения произошли и во внутреннем строении — концентрация нервной системны, дополнение и замена выделительных коксальных желез мальпигиевыми сосудами, сокращение кровеносной системы в связи с переходом к трахейному и кожному дыханию, особенно у мелких форм, и др. Изменилась биология размножения. Водный тип наружного оплодотворения сменился внутренним, сначала свободным сперматофорным, а далее разнообразными способами копуляции. В ряде случаев возникло живорождение, охрана потомства. Выработался эфемерный тип жизни, свойственный наземным членистоногим: способность заканчивать развитие в ограниченный срок, недолговечность и сравнительно малые размеры взрослой формы, наличие переживающих стадий. Так была решена задача перехода к сухопутной жизни.
Однако, как говорилось выше, предки арахнид были достаточно специализированными водными хелицеровыми, и при выходе на сушу новые приспособления могли развиваться лишь на основе уже сложившейся, весьма своеобразной организации водных форм, что создало ряд ограничений. И если взглянуть на арахнид не с обычной точки зрения — восхищения совершенством приспособлений к окружающей среде, а с противоположной — с точки зрения ограничений и трудностей, которые создались благодаря былой специализации и которые пришлось преодолевать или обходить, то многое в их эволюции станет более понятным. Очень показательно также сравнение с насекомыми — трахейнодышащими животными, наземными по природе. Так, дыхание с помощью легких, образовавшихся из жаберных ножек, у членистоногих с их незамкнутой кровеносной системой представляет гораздо менее совершенный способ газообмена, чем трахейное дыхание. Защита от пересыхания — главной опасности на суше — при локализованном легочном дыхании несовершенна, и действительно, большинство арахнид для дыхания нуждается в сильно увлажненном воздухе. Поскольку арахниды встали на путь легочного дыхания, трахейная система не развивалась в должной степени. Несмотря на многочисленные попытки в этом направлении, она не достигла такого совершенства, как у насекомых. Только сольпуги и сенокосцы по степени развития трахей несколько напоминают последних. Характерно, что мелкие тонкокожие арахниды (многие клещи, кенении), живущие во влажном почвенном воздуху вообще избавлены от противоречивого по природе легочно-трахейного аппарата и дышат через покровы. Многие ограничения в жизни на суше создались благодаря отсутствию обособленной подвижной головы с усиками и челюстями и в особенности атрофии сложных глаз. Арахниды были вынуждены пойти по пути совершенствования в основном осязания, имитируя усики конечностями, и ориентации в окружающем мире «на ощупь», что, помимо прочих неудобств, ограничивает эффективность охоты бродячего хищника. Вместо питания с помощью набора специальных ротовых конечностей — челюстей, приспособляемых для приема разнообразной пищи, что свойственно насекомым, у арахнид выработался очень однотипный способ питания разжиженным содержимым жертвы, т. е. почти поголовное хищничество. Только часть клещей сумела выбраться из этого однообразия. Прямое укороченное послезародышевое развитие, связанное с обилием желтка в яйце и поздним вылуплением, при всех преимуществах имело ту отрицательную сторону, что на его основе не могли возникнуть сложные формы метаморфоза, какие свойственны насекомым и открыли перед ними широчайшие возможности приспособления к различным условиям жизни. Только клещи с их своеобразным метаморфозом в этом отношении стали соперничать с насекомыми.
В том, как и в какой степени преодолевались или обходились эти исторически сложившиеся ограничения, отряды арахнид различны. Эволюционные возможности арахнид наглядно обнаруживаются при сопоставлении видового разнообразия и распространения отрядов. Из общего числа 35 000 видов львиная доля приходится только на пауков (20 000) и клещей (10 000). Из остальных 5000 видов 2500 составляют сенокосцы, 1100 ложноскорпионы, а прочие насчитывают по нескольку сотен или даже десятков видов. Такие соотношения не случайны. Маловидовые отряды — это как раз арахниды, в образе жизни и распространении которых отчетливо проявляются те ограничения, о которых только что говорилось. Все они теснейшим образом связаны с почвой и различными укрытиями, где воздух достаточно влажен. Это бродячие хищники, в большинстве ночные, которые ловят добычу «на ощупь» и прячутся днем в трещинах почвы, под камнями, в норах или постоянно живут под пологом растительности, в лесной подстилке, древесной трухе и т. п. В своем распространении эти отряды ограничены теплыми странами, многие формы не выходят за пределы тропиков. Только часть видов сенокосцев и ложноскорпионов встречается в умеренных широтах.
Иную картину представляют пауки и клещи. Среди арахнид по существу только они сумели вполне преодолеть или, вернее сказать, обойти историческую ограниченность своего класса. Немногочисленные примитивные представители этих групп — низшие норовые и бродячие пауки и примитивные клещи — по экологическому облику еще стоят в одном ряду с другими арахнидами, но дальнейшая судьба пауков и клещей совсем иная.
Решающее значение в эволюции пауков имела паутина, которая первоначально использовалась для устройства яйцевых коконов и выстилания убежищ, а далее стала применяться и для постройки ловчих сетей. В жизни высших тенетных пауков паутина — это все. Это убежище и ловушка. В убежище создается благоприятный микроклимат, особенно важный для дыхания, здесь паук подстерегает добычу, укрывается от врагов и непогоды. Добыча попадает в ловчую сеть, схватывается «на ощупь» при минимальном участии зрения и убивается с помощью хелицер, которыми впрыскивается яд. На паутине происходит спаривание, из нее сплетается яйцевой кокон, в ней укрывается неокрепшая молодь, на паутинках молодые паучки разносятся ветром и т. д. Обеспечив себя всем необходимым, пауки со своими паутинными приспособлениями проникли во всевозможные места обитания, широко заселили сушу и достигли небывалого расцвета. При довольно стереотипном общем облике высшие тенетные пауки исключительно разнообразны по местам обитания, форме и окраске, конструкциям ловчих сетей и повадкам. По сложности поведения и совершенству инстинктов пауки напоминают насекомых.
Совсем иным путем обошли затруднения клещи. Решающее значение в их эволюции имели микроскопически малые размеры. Прежде всего клещи заселили почву и гниющую растительную подстилку. Другие, более крупные арахниды либо роют норы, либо отыскивают достаточно объемистые временные укрытия, которые приходится покидать при выходе на охоту. Клещам этого не нужно. Они так малы, что для них пространства между частичками почвы — это готовая система пещер и галерей. А условия жизни здесь весьма подходящие: воздух влажен, пища — мелкие беспозвоночные и микрофлора — всегда в изобилии. В почве, лесной подстилке, скоплениях всевозможных гниющих остатков развился целый мир разнообразнейших клещей, который служил и продолжает служить источником новых форм в их дальнейшей эволюции. Выход из почвы происходил по нескольким направлениям. Одни устремились на растения, другие на животных в качестве паразитов, третьи заселили пресные воды и море. Паразитизм получил очень широкое распространение, и по своеобразию его форм клещи превзошли даже насекомых. Малые размеры и тут сыграли решающую роль. Наряду с наружными паразитами, сосущими кровь животных или сок растений, возникли такие своеобразные микроскопические формы, как чесоточные зудни, живущие в коже, железницы, живущие в волосяных сумках млекопитающих, паразиты трахей насекомых, галлообразующие клещи, обитатели вызываемых ими патологических разрастаний тканей растений — галлов, и многие другие.
Существенное значение в эволюции клещей имело разнообразие питания, возникшее уже на почвенной стадии их жизни. В почве клещам представился неограниченный выбор пищи, разнообразнейшая микрофлора, бактерии, дрожжи, низшие грибы, водоросли, всевозможные мелкие беспозвоночные. Для более крупных арахнид вся эта мелюзга не представляла бы ценности, но клещам пришлась по вкусу. Одни из них стали питаться растительной пищей, грибами, гниющими остатками с обильной микрофлорой, другие — беспозвоночными, мелкими червями, членистоногими, в том числе другими видами клещей. Разнообразие способов питания привело к специализации ротовых органов, возникли грызущие и колюще-сосущие формы. У большинства клещей хелицеры и педипальпы объединились в особый отдел — головку, или гнатосому, чем была обеспечена подвижность ротового аппарата как целого и в какой-то мере компенсировалось отсутствие подвижной головы, столь характерное для хелицеровых. Разнообразие питания отразилось и при переходе к паразитизму. Грызущие потребители органических остатков дали начало многочисленным паразитам птиц и млекопитающих, питающимся кожей, пером, волосом, кожными выделениями. Хищники, высасывающие мелких членистоногих, перешли к сосанию крови позвоночных. Сосущие растительноядные формы заселили высшие растения.
Как мы говорили, благодаря малым размерам яиц клещи развиваются с метаморфозом. По мере приспособления к новым условиям изменялась не только взрослая форма, но и способы метаморфоза, и это значительно расширило эволюционные возможности. В частности, возникли чрезвычайно быстро размножающиеся формы, способные в кратчайший срок достигать колоссальной численности, выработались специальные переживающие и расселительные стадии и т. п. По разнообразию и обилию в природе клещи превзошли пауков, хотя и уступают им по числу известных видов.
Таким образом, большинство отрядов арахнид оказалось ограниченным в освоении суши, и только пауки и клещи пошли значительно дальше и из бедных поселенцев превратились в завоевателей суши. Пауки и клещи распространены очень широко, от тропиков до полярных стран и высокогорий. Их можно встретить там, где жизнь скудна и почти нет даже насекомых. По численности в природе они не уступают последним. Не следует, однако, думать, что остальные, меньшие по числу видов отряды более похожи друг на друга. Напротив, каждый из них имеет свои неповторимые особенности и свои варианты приспособлений, вполне обеспечивающие жизнь в присущих ему условиях. Только эти приспособления более частного характера и не ведут к таким грандиозным эволюционным последствиям, как у пауков и клещей. Сравнивая отряды арахнид, можно как-то обрисовать лицо каждого.
Так, скорпионы — это древнейшие арахниды, по существу вышедшие на сушу эвриптериды. Минимум приспособлений к сухопутной жизни (легочное дыхание, стопохождение, арахнидный тип хищничества) сочетается у них с очень своеобразными чертами (ядовитый аппарат на конце метасомы, переход к живорождению, вынашивание молоди на себе и др.). По образу жизни и примитивности несколько напоминают скорпионов телифоны и фрины, но эти очень бедные видами отряды более узко приурочены к влажным теплым местам обитания, в основном к тропическим лесам, и отличаются по строению (иное число и положение легких, отсутствие ядовитого аппарата на метасоме и др.). Фрины в то же время имеют настолько много общего с пауками, что их считают беспаутинными родичами последних и называют иначе жгутоногими пауками.
Два отряда — сольпуги и сенокосцы — настолько выделяются по степени развития трахейной системы, что их можно назвать трахейнодышащими арахнидами. Главные трахейные стволы открываются стигмами на брюшке там, где у арахнид бывают легкие, и весьма вероятно, что трахеи здесь возникли из легких, в чем, может быть, и кроется причина такого мощного их развития. В остальном сольпуги и сенокосцы очень различны и далеки друг от друга. У сольпуг мощная трахейная система сочетается с примитивностью организации (полнота сегментации туловища, расчлененная просома, педипальпы, подобные ногам, и т. д.). Подобно большинству арахнид, сольпуги — ночные хищники, прячущиеся днем в укрытиях. Но распространены они в основном в сухих и жарких районах, чрезвычайно подвижны, и даже есть несколько видов, бегающих по песку под палящим солнцем в пустынях. Все это указывает на совершенство регуляции дыхания и водного обмена. Однако сама по себе трахейная система при прочих примитивных арахнидных свойствах, видимо, недостаточна для перехода к более совершенным формам открытой наземной жизни, и видовое разнообразие сольпуг невелико.
Сенокосцы по своему жизненному облику — это самые, если можно так выразиться, насекомоподобные арахниды. Наряду с развитым трахейным дыханием в этом отряде преобладает та панцирная жизненная форма, которая характерна для некоторых нелетающих или мало пользующихся крыльями насекомых, например жуков. Компактное тельце защищено кожистым или очень твердым панцирным покровом. Сегменты брюшка сомкнуты, а у многих форм их тергиты срастаются с головогрудным щитом в общий спинной щит. В то же время тело сенокосцев как бы подвешено на длинных ногах, которые при невысокой частоте движений обеспечивают большую скорость перемещения: шаг сенокосцев очень велик. Наряду с ночными хищниками среди сенокосцев много видов, активных днем, свободно разгуливающих на ярком солнце, даже и в сухих местностях. Не имея тех преимуществ, которые свойственны богатым видами отрядам, сенокосцы тем не менее широко распространились и достигли значительного разнообразия (2500 видов).
Несколько отрядов мелких арахнид — кенении, ложноскорпионы, рицинулеи — приспособились к скрытой жизни в естественных полостях и трещинах почвы, в лесной подстилке, древесных остатках и т. п. В этом отношении они напоминают клещей. Однако все они крупнее и не перешагнули ту ступепь измельчания, за которой возникла микроскопическая жизненная форма клещей с ее эволюционными возможностями. Кенении и рицинулеи представлены немногими редкими, в основном тропическими видами, ложноскорпионов известно 1100 видов и распространены они шире. Кенении — это типичные обитатели скважин почвы, одни из примитивнейших арахнид, напоминающие, с одной стороны, сольпуг в миниатюре, с другой — некоторых низших клещей. Ложноскорпионы также весьма примитивны, но имеют некоторые очень своеобразные черты: хватательные педипальпы с клешнями, как у скорпионов, чрезвычайно своеобразный способ живорождения и др. Они живут скрыто в лесной подстилке, древесной трухе, под отставшей корой, под камнями и могут расселяться, прикрепившись к насекомым. По-видимому, такой образ жизни способствовал довольно широкому распространению ложноскорпионов, хотя и они явно преобладают в тропиках. Об образе жизни рицинулей известно мало. Эти медлительные формы с очень твердым покровом замечательны тем, что в их развитии, как у клещей, имеется шестиногая личинка.
Смену мест обитания в эволюции арахнид можно иллюстрировать схемой (рис. 19). Выходя на сушу, арахниды были вынуждены ограничиться влажными местами обитания, в которых и до сего времени живут многие из них. Важнейшим условием выхода на сушу была наземная растительность. Многие нашли приют под ее пологом, другие, в особенности мелкие, заселили продукты разложения растений, органическую подстилку и почву. Выработавшаяся у арахнид способность устраивать логова и норки для себя и своего потомства в сочетании с ночной активностью заметно расширила возможности освоения суши и позволила выйти из-под покрова влажной растительности. Тесная связь арахнид с почвой на этой ступени их эволюции хорошо согласуется с представлениями М. С. Гилярова о переходной роли этой среды при смене водного образа жизни наземным, изложенными в его известной книге «Особенности почвы как среды обитания и ее значение в эволюции насекомых» (изд. АН СССР, 1949).
Прямое освоение открытой дневной поверхности суши, как мы видели, у арахнид явление редкое и связано с выработкой надежной защиты от высыхания, трахейным дыханием, уплотнением покровов (часть сенокосцев и немногие другие). Иными, обходными, способами порывают связь с почвой высшие пауки и клещи. Первые уходят от почвы со своими паутинными устройствами, причем наиболее широко заселяют растительность. Вторые благодаря микроскопическим размерам и разнообразию, достигнутому уже в почве, заселяют буквально все мыслимые места обитания, в том числе растения и животных в качестве их паразитов, а также вторично проникают в водоемы. Не имея крыльев, пауки и клещи проникают и в воздушную среду: молодь пауков на паутине разносится ветром, мелкие клещи поднимаются токами воздуха на огромные высоты, будучи паразитами птиц, совершают с ними дальние перелеты.
Чтобы перейти к более подробному обзору отрядов арахнид, необходимо остановиться на некоторых вопросах классификации. Как говорилось, класс Arachnida есть собрание хелицеровых, перешедших к сухопутной жизни. Отряды арахнид очень различны. При глубокой общности их всех как представителей подтипа Chelicerata почти каждый отряд по сочетанию признаков уникален, и не только невозможно вывести его из какого-либо соседнего, но в ряде случаев затруднительно точно сказать, к каким из остальных отрядов он ближе. Такая уникальность отрядов объясняется, с одной стороны, различными вариантами приспособлений к сухопутной жизни, о которых говорилось выше. Но с другой стороны, признаки отрядов таковы, что их невозможно свести к одним только этим приспособлениям, они ведут куда-то глубже и заставляют думать, что арахниды более или менее независимо произошли от различных водных хелицеровых. Непосредственные предки большинства отрядов пока не обнаружены. Но в отношении одного отряда, а именно скорпионов, они теперь известны. Ряд ископаемых переходных форм, независимо от других арахнид, связывает скорпионов с некоторыми силурийскими эвриптеридами. Иначе говоря, класс Arachnida в его традиционном составе приходится считать искусственным. В связи с этим в последнее время не раз делались попытки сгруппировать отряды по их возможному происхождению и разделить арахнид на несколько классов. Но мнения зоологов расходятся, и работу по упорядочению классификации нельзя считать завершенной.
Как говорилось, большинство отрядов арахнид в качестве четких систематических группировок не вызывает сомнений. Разноречия имеются в отношении жгутоногих (Pedipalpi) и клещей (Acarina). С первыми обстоит несколько проще. Под жгутоногими понимают три достаточно четко разграниченные, хотя и близкие в некоторых отношениях группы: телифонов, фринов и тартарид. Большинство авторов вполне основательно считает телифонов и фринов самостоятельными отрядами. Тартарид одни оставляют в составе телифонов, другие, в том числе и мы, считают отдельным отрядом.
Гораздо сложнее обстоит дело с клещами. Клещи представляют огромное собрание мелких арахнид, очень различных по строению и образу жизни и в большинстве сильно изменившихся по сравнению с другими. До недавнего времени все это многообразие объединялось в один отряд Acarina со многими подотрядами и специальными более дробными подразделениями, (когортами, фалангами, сериями и т. п.), систематический состав которых различен у разных авторов. И, пожалуй, нет другой такой группы членистоногих, которая по запутанности и противоречивости классификации была бы подобна клещам. Клещи рассматривались как совсем особые арахниды, которые деградировали и настолько уклонились от исходного состояния, что их даже трудно сравнивать с остальными. Считалось, да и теперь пишется, особенно в учебниках зоологии, что всем клещам свойственны три основных признака, отличающие их от остальных арахнид. Во-первых, сегменты тела у клещей слились и границы между ними исчезли, а если и наблюдается разделение тела на отделы, то последние не соответствуют отделам тела других арахнид. Во-вторых, у клещей имеется особый подвижный передний отдел — головка, или гнатосома, в составе которой объединяются хелицеры и педипальпы. В-третьих, у клещей из яйца вылупляется шестиногая личинка, которая затем превращается в восьминогую форму.
Пока главное внимание исследователей было направлено на изучение отдельных наиболее распространенных групп клещей, особенно паразитических, действительно сильно измененных по сравнению с другими арахнидами, казалось, что по перечисленным признакам клещи сходны между собой и вполне отличимы от других паукообразных. Но со временем стали накапливаться данные о том, что далеко не все клещи так сильно видоизменены. В 30-х годах нашего века французским исследователем Ф. Гранжаном были детально описаны примитивные сегментированные клещи, вполне сравнимые с другими арахнидами. Эти работы привлекли внимание советского зоолога А. А. 3ахваткина, который в 1941 году в специальном исследовании показал, что отделы тела клещей вполне соответствуют таковым других арахнид, так что первый из упомянутых выше признаков потерял для клещей отличительное значение. Несостоятельным оказался и второй признак: головка, или гнатосома, свойственна не всем клещам, у примитивных форм строение околоротовых конечностей такое же, как у большинства арахнид. Шестиногая личинка была открыта в отряде рицинулей, так что и этот признак потерял для клещей уникальность. Автором этих строк были изучены низшие представители так называемых паразитиформных клещей, и выяснилось, что по строению и типу развития они резко отличны от остальных.
В результате создалось довольно странное положение: клещи — один из крупнейших отрядов арахнид — оказались лишенными общих признаков, по которым всех их представителей можно было бы отличить от других отрядов. Это наводило на мысль, что клещи — отряд сборный, что в нем искусственно объединены различные по происхождению арахниды. Анализируя данные о клещах, А. А. 3ахваткин в 1947 году пришел к выводу, что в действительности существует не один, а три самостоятельных отряда клещей, каждый из которых имеет определенные признаки, четко отличающие его от любого другого отряда. Это акариформные клещи (отряд Acariformes), паразитиформные клещи (отряд Parasitiformes) и клещи-сенокосцы (отряд Opilioacarina). Первый отряд сближается с такими примитивными арахнидами, как кенении и сольпуги, а два других более сходны с сенокосцами, рицинулеями и некоторыми ископаемыми отрядами. Основная масса клещей очень естественно и закономерно распределилась между первыми двумя отрядами. Третий отряд представлен лишь несколькими примитивными, в основном тропическими видами.
Дальнейшие исследования подтвердили правильность такого деления. Так, нами было показано, что строение головки (гнатосомы, или, как она теперь называется, гнатемы) различно и что это образование возникло в эволюции каждого отряда клещей независимо, своим особым способом. Исследование эмбрионального развития примитивных клещей показало, что в разных отрядах принципиально различны и способы образования шестиногих личинок, так что этот признак внешнего сходства при более внимательном изучении еще углубил различия. Каждый отряд клещей самобытен не только по своим исходным формам, но и по общему строю эволюции. Из двух многовидовых отрядов акариформные клещи замечательны, с одной стороны, большой примитивностью и древностью низших форм, которые были почти такими же, как сейчас, уже в девоне, а с другой — исключительным многообразием эволюционных ветвей, ведущих к различным паразитам животных и растений, водяным клещам и многим другим. Паразитиформные клещи несколько однотипнее, судя по низшим представителям отряда, они возникли из какого-то иного источника и, вероятно, позже, а главное направление их эволюции — переход от почвенных хищников к кровососам наземных позвоночных и частью сожителям и паразитам насекомых.
Несмотря на фактическую неопровержимость самостоятельности отрядов клещей, новая классификация вызывает различное отношение специалистов. Одни относятся к ней положительно, например, такой выдающийся зоолог и сравнительный анатом нашего времени, как В. Н. Беклемишев, приводит ее в своих «Основах сравнительной анатомии беспозвоночных» (издания 1962, 1964 гг.). Отношение других неопределенно, а некоторых отрицательно. Причины разноречий различны и, как ни странно, мало связаны с фактами. Сказывается главным образом сила традиции. Некоторые авторы пытаются найти выход в том, что, признавая три отряда клещей, объединяют их все в особый подкласс или даже класс. Так поступает, например, наш известный специалист по клещам В. Б. Дубинин в очерке хелицеровых, помещенном в капитальном академическом издании «Основы палеонтологии» (1962). Но такая операция по существу не меняет дела: возвышение ранга не придает естественности объединению клещей. С другой стороны, преобладает чисто формальное отношение к данному вопросу, что обусловлено самым характером изучения клещей. Дело в том, что благодаря многообразию клещей и сложности их исследования подавляющее большинство специалистов занимается отдельными систематическими группами. И для систематика, изучающего, например, только чесоточных или только галловых клещей, не так уж важно, относят ли их к отряду Acariformes или к отряду Acarina. А думать о клещах как о чем-то целом привычнее. Важно и то, что благодаря медицинскому и хозяйственному значению клещей возникла целая самостоятельная отрасль знания, наука о клещах — акарология, параллельная науке о насекомых — энтомологии, — отрасль знания со своими методами, своим кругом научно-практических проблем, сложнейшей терминологией, своими симпозиумами и конгрессами, своими традициями. Но если энтомология имеет объектом естественную группу членистоногих — класс насекомых, то акарология, при новом подходе к клещам, оказывается наукой всего лишь о нескольких разнородных отрядах мелких арахнид. Такое «упразднение» единого объекта целой отрасли знания подчас вызывает и чисто психологический протест.
Совсем иначе предстает разделение клещей на отряды, как только мы от частной и прикладной акарологии обратимся к акарологии общей, задача которой состоит в упорядочении всего огромного материала по клещам, по их строению, развитию, образу жизни, распространению и т. д., и в конечном счете в выяснении происхождения и эволюции клещей. Здесь пути и результаты анализа фактов всецело зависят от того, признаем ли мы клещей единой группой или тремя независимыми отрядами, не более родственными друг другу, чем арахниды вообще. В первом случае мы вынуждены изучать клещей как таковых, отвлекаясь на первых порах от других арахнид, и главные наши усилия направить на то, чтобы представить себе и по возможности найти исходную прототипическую форму для клещей в целом, проследить, как из этого прототипа возникло все многообразие клещей, а затем установить, каковы родственные отношения этого прототипа с другими отрядами. Во втором случае поиски единого прототипа клещей становятся бессмысленными. Мы должны изучать отряды клещей порознь и в каждом случае выяснить исходное состояние, пути эволюции каждого отряда и место каждого в общей эволюции арахнид. И весь фактический материал по клещам с полной убедительностью показывает, что единого прототипа клещей, если можно так выразиться, «проклеща», в природе нет и никогда не было. Традиционный подход к клещам как единой группе ничего хорошего не приносит. Достаточно раскрыть общие монографии по клещам, например наиболее известную объемистую сводку немецкого акаролога Г. Фицтума 1943 года, как мы наталкиваемся на груду фактов, бесконечное перечисление несвязуемых вариантов строения, развития, образа жизни и т. д. Попытки же свести эти данные к чему-то единому неизменно приводят к противоречиям, а подчас к таким фантастическим гипотезам, рассматривать которые здесь вряд ли уместно.
Иные результаты дает изучение клещей как трех независимых отрядов. При таком подходе выясняется, что многообразие клещей не хаотично. В пределах отрядов вырисовываются многочисленные естественные ряды форм, представляющие нам пути минувшего эволюционного процесса. Полнота этих рядов, сохранность в современной фауне исходных, переходных и конечных форм, особенно в отряде акариформных клещей, поистине поразительны, и среди членистоногих вряд ли найдутся другие подобные группы, где современное многообразие так наглядно отражает историю. Сравнительное изучение рядов форм подтверждает и глубокую самобытность отрядов клещей. Ведь если группа организмов едина по происхождению, то по мере того, как мы спускаемся от измененных ее представителей к примитивным, сходство неизменно должно возрастать и все ряды форм сходиться или по крайней мере сближаться. Внутри отрядов именно это и наблюдается: ряды форм сходятся к примитивам. Но при сравнении отрядов клещей друг с другом картина прямо противоположная: чем примитивнее, тем разительнее несходство. Иными словами, клещи представляют яркий пример конвергенции, т. е. внешнего сближения в эволюции нескольких самостоятельных отрядов арахнид. Степень конвергенции клещей очень велика и объясняется общностью их исторической судьбы, в которой, как мы говорили, решающими были малые размеры. Последние повлекли за собой внешне сходные изменения строения и развития (потеря сегментации, образование головки, своеобразный метаморфоз и др.), а также образа жизни (заселение почвы и других пористых субстратов, выход из почвы в качестве паразитов и т. д.).
Говоря о конвергенции клещей, не следует забывать и другую сторону этого явления. До сих пор мы говорили о разнородности клещей как трех отрядов. Но ведь все они хелицеровые и в этом смысле глубоко родственны, как и другие арахниды, так что явления конвергентного сближения отрядов клещей разыгрывались в эволюции на общей им всем арахнидной основе, и в этом также причина глубины конвергенции. Об этом приходится говорить еще потому, что некоторые ученые, отчаявшись понять своеобразие клещей, вообще отделяют их от паукообразных, что представляет другую крайность в вопросах классификации и абсолютно неприемлемо. Как нельзя соединить клещей в одну группу, так нельзя и выкинуть их из паукообразных. Клещи, или, точнее, клещеобразные арахниды, это три самостоятельных отряда, в такой же мере уникальные, как пауки, сенокосцы, сольпуги и другие, и в такой же мере относящиеся к собранию наземных хелицеровых, именуемому паукообразными.
Словом, клещи явились порядочной загадкой, разрешение которой только теперь, после разделения на отряды, встало на твердую почву. В этом отношении клещи служат прекрасным примером того, как классификация организмов есть не только средство их опознавания, или, как некоторые думают, условное «раскладывание по полочкам», но имеет гораздо более глубокое значение. Будучи сама выводом из некоторой, на первых порах ограниченной, группы фактов, естественная классификация придает правильное направление дальнейшему исследованию, избавляя науку от заблуждений и бесплодной траты времени.
Практическая значимость арахнид очень неодинакова. Действительно большое медицинское и хозяйственное значение имеют только клещи. Среди них имеются многочисленные паразиты, передающие человеку и животным возбудителей различных опасных инфекционных заболеваний — энцефалитов, клещевых тифов, чумы, туляремии и др. Многие виды повреждают культурные растения, запасы зерна, муки и других пищевых продуктов. Некоторые почвенные клещи служат промежуточными хозяевами ленточных глистов, поражающих сельскохозяйственных и ценных промысловых животных. Многообразное практическое значение клещей, как говорилось, вызвало к жизни самостоятельную отрасль знания — акарологию, которая по своим задачам параллельна науке о насекомых — энтомологии. Наряду с общими разделами эта наука включает и прикладные — медицинскую, ветеринарную и сельскохозяйственную акарологию. В последние десятилетия акарология развивается особенно интенсивно. Многое в этой области сделано советскими учеными. Во всех странах мира число акарологов, занимающихся разработкой только систематики и экологии отдельных групп клещей, по данным каталога, издаваемого в США, превышает тысячу. Значение остальных арахнид более ограниченно и сводится в основном к ядовитым укусам некоторых скорпионов и пауков. Среди последних обращают на себя внимание некоторые тропические виды, а в нашей стране — паук каракурт, распространенный в южных районах. Кроме того, пауки, будучи массовыми хищниками, уничтожающими в основном насекомых, несомненно играют определенную роль в жизни природных сообществ организмов — биоценозов, но они в этом отношении мало изучены.
ОТРЯД СКОРПИОНЫ (SCORPIONES)
Скорпионы — древнейший отряд не только среди арахнид, но и среди наземных членистоногих вообще. Как отмечалось, они представляют потомков палеозойских эвриптерид, и это редкий пример среди членистоногих, где по палеонтологическим материалам довольно полно прослеживается переход от водного образа жизни к сухопутному. Среди силурийских эвриптерид найдены формы, очень похожие на скорпионов, но жившие в воде и дышавшие с помощью брюшных жаберных ножек (табл. 3, 2). У наземных скорпионов последние превратились в легкие. Изменилось также строение ходильных ног. У водных форм они заканчивались остроконечным члеником (группа заостренноногих — Apoxipodes), у наземных ноги удлинились и их концевые части превратились в членистые лапки, приспособленные к хождению по суше (группа двукоготковых — Dionychopodes). Наземные формы, в общем подобные современным скорпионам, представлены уже в отложениях каменноугольного периода.
Скорпионы — средних размеров или крупные формы, обычно 5—10 см, некоторые до 20 см (табл. 3, 2). Во внешнем облике наиболее характерны крупные педипальпы с клешнями и членистая гибкая метасома («хвост») с ядовитым аппаратом на конце. По строению скорпионы наиболее близки к прототипу хелицеровых. Три отдела тела — про-, мезо- и метасома — хорошо выражены, каждый состоит из 6 сегментов. Головогрудной щит цельный, на нем имеется пара более крупных срединных глаз и до 5 пар мелких боковых. Хелицеры небольшие, клешневидные, педипальпы очень крупные с массивными клешнями. На тазиках педипальп и двух передних пар ног имеются жевательные отростки, направленные ко рту. Брюшко широким основанием примыкает к головогруди, предполовой (7-й) сегмент атрофирован. Передний отдел брюшка (мезосома) более широкий, его сегменты имеют обособленные тергиты и стерниты; видоизмененные брюшные конечности представлены полным набором: половые крышечки на восьмом сегменте, гребневидные органы на девятом, легочные мешки на десятом—тринадцатом. Сегменты заднего отдела (метасомы) узкие цилиндрические, тергит и стернит каждого сегмента слиты в цельное склеритное кольцо; первый сегмент метасомы конический. Метасома заканчивается вздутым хвостовым члеником, в нем помещается ядовитая железа, проток которой открывается на конце кривого острого жала. Щитки туловища и членики конечностей образованы очень твердой кутикулой, часто имеющей ребристую или бугорчатую скульптуру.
Скорпионы живут в странах с теплым или жарким климатом, причем встречаются в самых различных местах обитания, от влажных лесов и литорали морских побережий до бесплодных каменистых местностей и песчаных пустынь. Некоторые виды найдены в горах на высоте 3—4 тыс. м над уровнем моря. Принято различать гигрофильные виды скорпионов, живущие во влажных местах, и ксерофильные, встречающиеся в сухих районах. Но это деление в значительной мере условно, так как все они активны ночью, а днем прячутся в укрытиях, под камнями, под отставшей корой, в норах других животных или зарываются в почву, так что и в сухих районах находят места, где воздух достаточно влажен. Более отчетливы различия в отношении к температуре. Большинство видов теплолюбиво, но некоторые, живущие высоко в горах, а также у северных и южных границ области распространения скорпионов, хорошо переносят холодные зимы в неактивном состоянии. Некоторые виды найдены в пещерах, но они здесь случайные пришельцы. Скорпионы — нередкие посетители жилища человека, но настоящих сожителей человека (синантропов) среди них нет.
Образ жизни скорпионов изучался рядом исследователей, ценными сведениями мы обязаны Фабру. При содержании в неволе повадки скорпионов искажаются, и, как замечают некоторые авторы, нет более свободолюбивого существа, чем скорпион. Скорпионы в неволе нуждаются в достаточном разнообразии условий и возможности их свободного выбора: большая площадь садка, различная влажность почвы или песка в разных его участках, наличие укрытий, периодическая смена освещенности и температуры и т. д. При этом поведение скорпионов близко к естественному, в частности отчетливо выражен суточный ритм активности.
Скорпион выходит на охоту ночью и особенно активен в жаркое время. Он медленно идет с поднятым «хвостом», выставив вперед полусогнутые педипальпы с приоткрытыми клешнями. Он передвигается ощупью, главную роль при этом играют торчащие осязательные волоски (трихоботрии) педипальп. Скорпион очень чутко реагирует на прикосновение к подвижному объекту и либо схватывает его, если это подходящая добыча, либо отступает, принимая угрожающую позу: он круто загибает «хвост» над головогрудью и размахивает им из стороны в сторону. Добыча схватывается клешнями педипальп и подводится к хелицерам. Если она невелика, то сразу же разминается хелицерами и содержимое поглощается. Если добыча оказывает сопротивление, скорпион жалит ее один или несколько раз, обездвиживая и убивая ядом. Скорпионы питаются живой добычей, объекты охоты очень разнообразны: пауки, сенокосцы, многоножки, различные насекомые и их личинки, известны случаи поедания мелких ящериц и даже мышат. Скорпионы могут очень долго голодать, их можно сохранять без пищи по нескольку месяцев, известны случаи голодания до полутора лет. Большинство видов, вероятно, всю жизнь обходится без воды, но некоторые обитатели влажных тропических лесов пьют воду. При совместном содержании в небольших садках скорпион нередко поедает собрата.
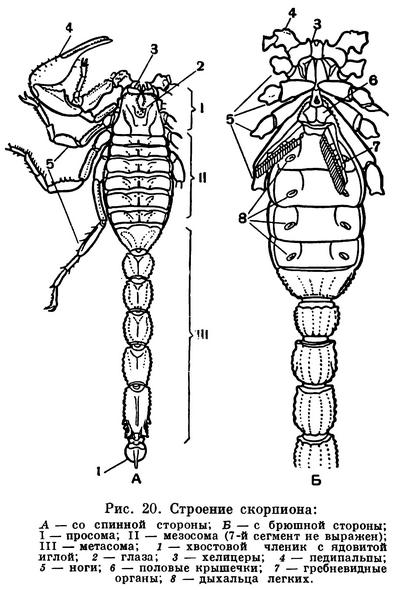
Биология размножения своеобразна. Спариванию предшествует «брачная прогулка». Самец и самка сцепляются клешнями и, подняв вертикально «хвосты», в течение многих часов и даже дней ходят вместе. Обычно самец, пятясь, влечет за собой более пассивную самку. Затем происходит копуляция. При этом особи прячутся в какое-либо укрытие, которое самец, не отпуская самку, быстро расчищает с помощью ног и «хвоста». Оплодотворение сперматофорное. Особи соприкасаются вентральными сторонами передних отделов брюшка, и самец вводит пакеты со сперматозоидами в половые пути самки, а затем выделяет особый секрет, которым заклеивается половое отверстие самки. Считают, что при спаривании какую-то роль играют гребешки — видоизмененные конечности девятого сегмента. Они снабжены многочисленными органами чувств. В покое гребешки прижаты к брюшку, при спаривании они оттопыриваются и колеблются. Но они оттопыриваются и при передвижении скорпиона, и им приписывают также роль органов равновесия и некоторые другие функции.
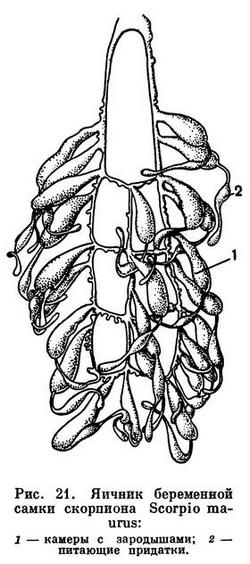
Скорпионы в большинстве живородящи, некоторые виды откладывают яйца, в которых зародыши уже развиты, так что вскоре вылупляется молодь. Это явление называется яйцеживорождением. Развитие эмбрионов в теле матери длительное; от нескольких месяцев до года и более. У одних видов яйца богаты желтком и зародыши развиваются в яйцевых оболочках, у других желтка почти нет и эмбрионы вскоре выходят в просвет яичника. По мере их роста образуются многочисленные вздутия яичника, в которых помещаются зародыши. Они питаются выделениями особых железистых придатков яичника (рис. 21). Зародышей бывает от 5—6 до нескольких десятков, реже около сотни. Маленькие скорпионы рождаются обернутыми эмбриональной оболочкой, которая вскоре сбрасывается. Они взбираются на тело матери и держатся на ней обычно дней 7—10. Скорпионы первого возраста не питаются активно, они беловатые, с гладким покровом и редкими волосками, лапки лишены коготков и имеют на конце присоски. Оставаясь на теле самки, они линяют, а через некоторое время покидают мать и начинают самостоятельно искать пищу. После линьки покровы твердеют и окрашиваются, на лапках появляются коготки. Скорпион становится взрослым через год-полтора после рождения, проделывая за это время 7 линек. Продолжительность жизни точно не установлена, но она обычно не менее нескольких лет. Встречаются интересные случаи аномалий, возникающих в эмбриональном развитии скорпионов, например удвоение «хвоста», причем особи жизнеспособны и дорастают до взрослого состояния («двухвостый скорпион» упоминается уже известным римским ученым Плинием Старшим в его «Естественной истории», I в. н. э.).
Твердые покровы и ядовитый аппарат не всегда спасают скорпионов от врагов. С ними справляются крупные хищные многоножки, сольпуги, некоторые пауки, богомолы, ящерицы, птицы. Есть виды обезьян, которые лакомятся скорпионами, осторожно удалив «хвост». Но злейший враг скорпионов — человек. С глубокой древности скорпион был предметом отвращения и мистического ужаса, и, пожалуй, нет другого членистоногого, которое породило бы столько россказней и легенд. Скорпион фигурирует и в древних мифах египтян и греков, и в прописях средневековых алхимиков как магический атрибут «превращения» свинца в золото, и в астрологии, поскольку имя скорпиона носит одно из зодиакальных созвездий, и у христиан как типичный компонент «фауны» преисподней. Любопытны уверения, что скорпионы могут оканчивать жизнь «самоубийством»: если окружить скорпиона горящими угольями, то он, чтобы избежать мучительной смерти, будто бы убивает себя ударом жала. Это мнение не соответствует действительности, но имеет под собой известное основание. Дело в том, что скорпион, подобно некоторым другим членистоногим, под действием сильных раздражителей может впадать в неподвижное состояние — явление мнимой смерти (каталепсии, или танатоза). Будучи окружен горящими угольями, скорпион, конечно, мечется в поисках выхода, принимает угрожающую позу, размахивает «хвостом», а затем вдруг становится неподвижным. Эту картину и принимают за «самоубийство». Но через некоторое время такой скорпион «оживает», если только он не испекся от жара. Столь же необоснованно довольно распространенное мнение, что скорпион ночью специально отыскивает спящего человека, чтобы его ужалить. Там, где скорпионов много, они в жаркие ночи, совершая свои охотничьи прогулки, нередко посещают жилища и могут взобраться и на постель. Если спящий человек придавит скорпиона или коснется его, то скорпион может ударить «хвостом», но специальных поисков человека здесь, конечно, нет.
Укол скорпиона — средство нападения и защиты. На мелких беспозвоночных, служащих обычно пищей скорпиону, яд действует почти мгновенно: животное тотчас же перестает двигаться. Но более крупные многоножки и насекомые умирают не сразу и после укола живут день- два; есть и такие насекомые, которые, по- видимому, вообще малочувствительны к яду скорпионов. Для мелких млекопитающих яд скорпионов большей частью смертелен. Ядовитость разных видов скорпионов очень различна. Для человека укол скорпиона, как правило, не смертелен, но известен ряд случаев с очень тяжелыми последствиями, даже со смертельным исходом, особенно у детей и в жарком климате. При уколе появляются боль, опухоль, затем наступает сонливость, озноб, иногда температурная реакция. Обычно через день-два эти явления проходят, но могут и затягиваться. Все зависит от того, какой скорпион уколол, кого и где. У нас большинство случаев ужаления скорпионами наблюдается в Средней Азии и Закавказье, где скорпионы обычны и многочисленны.
Известно около 600 видов скорпионов, относящихся примерно к 70 родам и 6 семействам. Географическое распространение скорпионов представляет большой интерес для зоогеографии — науки о закономерностях распространения животных. Будучи древнейшими наземными членистоногими, скорпионы отражают в своем распространении геологические и климатические перемены и смену растительных и животных сообществ, многократно происходившие в истории Земли. Ограниченные способности расселения скорпионов придают особую ценность этим данным: в большинстве случаев те или иные формы присутствуют там, где сумели сохраниться с давних времен.
Разработке классификации и изучению распространения скорпионов посвящены труды ряда ученых. Очень ценны исследования А. А. Бялыницкого-Бирули, который в работе о скорпионах Кавказа (1917) произвел замечательный анализ материалов о распространении и эволюции скорпионов вообще. В настоящее время область распространения скорпионов опоясывает земной шар между примерно 50° северной и южной широты, но и в прошлые эпохи, вплоть до конца третичного периода, когда климат был более теплым и влажные леса простирались до высоких широт, скорпионы встречались на большей части суши.
По морфологическим признакам скорпионы распадаются на две большие группы: бутоидов, представленных семейством Buthidae (до 300 видов), и хактоидов (остальные семейства). Считают, что эти группы разделились в отдаленные времена, вероятно уже в силурийский период, и с тех пор каждая эволюционировала своим путем, по-своему отражая явления, влиявшие на распределение фаун (разобщение материков, изменения климата и др.). Распространение примитивных представителей этих групп подтверждает данные геологии о том, что мировая суша долгое время (с начала палеозойской вплоть до первой половины кайнозойской эры) была разделена морями на два комплекса материков — северный и южный. Так, древнее подсемейство бутоидов — Isometrinae — распространено преимущественно в Африке и Южной Америке, а для Южной Америки и одновременно Австралии характерно своеобразное семейство Bothriuridae. Древние хактоидные скорпионы семейств Chactidae и Vejovidae приурочены к субтропическому поясу северного полушария в Старом и Новом Свете и совершенно отсутствуют в Африке и Австралии.
Общая картина современного распространения скорпионов представляет результат сложного наслоения фаунистических элементов различных эпох и в целом подтверждает деление суши на зоогеографические области, установленное на основании распространения животных вообще. В семействе Buthidae подсемейства, а часто и роды строго приурочены к определенным зоогеографическим областям, т. е. обладают высокой степенью эндемизма. Так, подсемейства Centrurinae и Tityinae населяют северо- и центральноамериканский районы неотропической области. Роды Parabuthus и Babycurus из подсемейства Buthinae характерны для эфиопской области Африки (к югу от Сахары); род Grosphus встречается только на Мадагаскаре. Многие роды обитают по границам пустынь от Атлантического океана до Индии, представляя сахаро-индийские фаунистические элементы. Роды Isometrus и Isometroides характерны для австралийской фауны. Среди хактоидных скорпионов высокой степенью эндемизма обладают подсемейства и целые семейства. Семейство Scorpionidae в основном представлено эфиопскими формами, мадагаскарским родом Heteroscorpius и индо-малайским Heterometrus. В семействе Chactidae, как говорилось, эфиопских видов нет, подсемейства Chactinae — неотропическое, Cherilinae — индо-малайское, Scorpioninae — средиземноморское. Сходно распространение подсемейств семейства Vejovidae. Семейство Bothriuridae в основном южноамериканское, но есть виды, живущие в Австралии и на Суматре. Особенно богата скорпионами фауна Индии, где насчитывается свыше 80 видов. В фауне Палеарктики имеется около 100 видов, из них в СССР встречается около 15.
В Закавказье, Нижнем Поволжье и по всей Средней Азии обычен пестрый скорпион (Buthus eupeus, табл. 2), образующий ряд подвидов. Он буро-желтый с темными пятнами и продольными полосами на спине, до 6,5 мм длиной. В Крыму, особенно на Южном берегу, не редок крымский скорпион (Euscorpius tauricus), свойственный только Крыму. Он светло-желтый, клешни узкие, буроватые, длина 35—40 мм. В Западном Закавказье обычен мингрельский скорпион (Euscorpius mingrelicus), красновато-коричневый, более светлый снизу, до 40 мм длиной. На Черноморском побережье Кавказа живет еще италъянский скорпион (Euscorpius italicus), красно-бурый или почти черный, до 55 мм длиной.
ОТРЯД ТЕЛИФОНЫ (UROPYGI)
Телифоны — средних размеров или крупные паукообразные, до 75 мм длиной, с твердыми темноокрашенными покровами. Удлиненная головогрудь покрыта цельным щитом, на котором имеются медиальные глаза и три пары боковых. Короткие хелицеры имеют на конце когтевидные членики. Педипальпы очень крупные, хватательные, их концевые заостренные членики образуют с выростами предыдущих члеников подобие клешни. Ноги длинные, лапки полно расчленены, с двумя коготками; передние ноги длиннее и тоньше остальных, без коготков, выполняют роль усиков. Брюшко продолговатое, полно расчленено, три последних сегмента (16—18-й) образуют маленький стебелек (постабдомен) с длинной членистой хвостовой нитью на конце. Имеются две пары легких на восьмом и девятом сегментах.
Телифоны живут в теплых странах и особенно характерны для влажных тропических лесов, под пологом которых жарко, сыро и всегда царит полумрак. Они прячутся под лежащими стволами деревьев, под камнями и т. п. Есть виды, встречающиеся в муравейниках. Обычно телифоны в тех местах, где они скрываются, роют с помощью педипальп норки, иногда до полуметра глубиной, в которых прячутся в засушливое время года. Наиболее активны телифоны в период дождей. Это ночные хищники, выходящие на охоту в сумерки. Телифоны передвигаются на трех парах ног, выставив вперед педипальпы и передние ноги. Последними они поочередно прикасаются к субстрату, ощупывая дорогу. Питаются телифоны главным образом насекомыми, взрослыми и личинками, а также многоножками, червями, слизнями, некоторые виды нападают на мелких лягушек. Добыча схватывается педипальпами. В общем телифоны довольно пугливы и на крупных сильных насекомых не нападают. Те, кто содержал телифонов в садках, отмечают, что они крайне агрессивны к собратьям: два телифона, посаженные вместе, как правило, вступают в борьбу и калечат друг друга.
Своеобразным средством защиты от врагов служат едкие выделения анальных желез, открывающихся близ анального отверстия. Убегая, телифон поднимает хвостовую нить и с силой выбрызгивает секрет, мелкие капли которого образуют небольшое облачко. Некоторые крупные формы «выстреливают» едкую жидкость на расстояние до 30 см. Секрет содержит кислоты муравьиную, уксусную, у некоторых видов он пахнет хлором. У человека едкие выделения телифонов могут вызывать сильное раздражение слизистых оболочек, особенно при попадании в глаза.
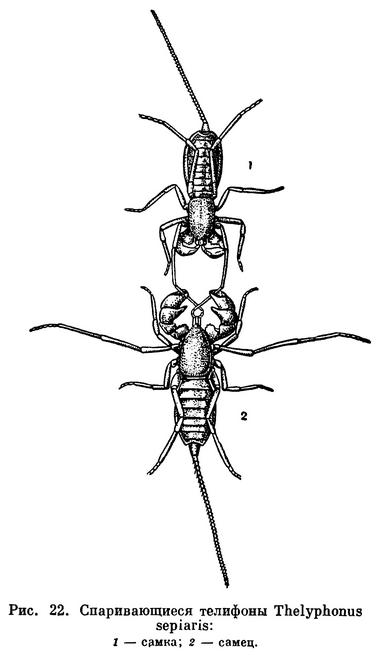
По биологии размножения телифоны напоминают скорпионов. Копуляции предшествует длительная «брачная прогулка». Самец держится хелицерами за перекрещенные передние ноги самки и, пятясь, влечет ее за собой (рис. 22). Оплодотворенная самка роет глубокую норку с расширением на дне. Здесь самка откладывает несколько десятков яиц. Яйца помещаются в клейком секрете внутри тонкой мешковидной оболочки, прикрепленной близ полового отверстия. Несколько недель самка остается в норке, охраняя кладку. Вылупляющиеся маленькие телифоны беловатые с тонким покровом, малоподвижны и не могут активно питаться. Они взбираются на самку и держатся на ее брюшке (табл. 3, 3). Дважды перелиняв, детеныши покидают мать, выбираются из норки и начинают самостоятельно охотиться. Дальнейшее развитие длительное, половая зрелость наступает обычно только на третий год. За это время телифон вырастает, линяя несколько раз.
Известно около 70 видов телифонов, относящихся к 10 родам семейства Thelyphonidae. Большинство их приурочено к Малайскому архипелагу и Центральной Америке. Роды Thelyphonus и Hypoctonus характерны для индо-малайской фауны. На Яве обычен телифон хвостатый (Thelyphonus caudatus), буро-черный, до 30 мм длиной. Виды рода Typopeltis распространены в Юго-Восточной Азии: и по побережью Китая доходят до Японии; один вид — телифон амурский (Typopeltis amurensis) — найден в Уссурийской области. Крупные телифоны рода Mastygoproctus встречаются в Бразилии и Мексике. В Австралии и Африке местных телифонов нет, но некоторые виды завезены в Западную Африку.
ОТРЯД ТАРТАРИДЫ, ИЛИ СХИЗОПЕЛЬТИДИИ (TARTARIDES, ИЛИ SCHIZOPELTIDIA)
Тартариды — небольшой отряд мелких арахнид (2—7 мм). Головогрудной щит разделен, пятый и шестой сегменты имеют свои тергиты. Педипальпы утолщены, с крючковидными концевыми члениками, без клешней. Передние ноги длиннее и тоньше других, играют роль усиков. Глаз нет. Брюшко веретеновидное с маленьким постабдоменом. Легких одна пара на восьмом сегменте.
Тартариды — тропические формы, живущие в почве, в скоплениях растительных остатков, под камнями. Они очень влаголюбивы и активны в дождливое время. Питаются мелкими насекомыми и другими беспозвоночными, обитающими в почве и лесной подстилке. Перед откладкой яиц самка вырывает в почве на глубине около 15 см небольшую камеру, стенки которой покрыты сцементированными частичками почвы. В камере самка откладывает несколько яиц, которые бывают прикреплены под основанием брюшка. До вылупления молоди самка находится в камере в своеобразной позе, с вертикально поднятым брюшком (табл. 3, 4). Описано до 30 видов этих арахнид, относящихся к трем родам семейства Schizomidae. Виды родов Schizomus и Trithyreus широко распространены в экваториальном поясе.
ОТРЯД ФРИНЫ, ИЛИ ЖГУТОНОГИЕ ПАУКИ (AMBLYPYGI)
Фрины (табл. 3, 5) — небольшой отряд тропических паукообразных средней величины или крупных (до 45 мм). Окраска темная, красноватая или желтоватая. Головогрудь покрыта цельным щитом, более или менее округлым по очертанию, с парой медиальных и 2—3 парами боковых глаз. Членистое брюшко компактное, без постабдомена и хвостовой нити. Оно подвижно соединено с головогрудью предполовым сегментом, суженным в стебелек. Легких две пары, на втором и третьем сегментах брюшка. Хелицеры короткие, с крючковидным концевым члеником, сгибающимся книзу. Педипальпы крупные, хватательные, вооруженные шипами, их концевые членики когтевидные. Ноги длинные, плоско расположенные, с расчлененными тонкими лапками. У некоторых форм, помимо пары коготков, на конце лапок развиты присоски, позволяющие передвигаться по гладким вертикальным поверхностям. Особенно длинны передние ноги, их лапки имеют вид очень длинных гибких многочлениковых жгутов, подобных антеннам насекомых. Как: отмечалось, фрины имеют много общих признаков с пауками и явно родственны им (расчленение тела, строение брюшка, устройство хелицер, число и положение легких и др.). Но у них нет ни ядовитых, ни паутинных желез, и этот бедный видами, отряд арахнид в известной степени дает нам представление о том, чем были бы пауки, не имей они яда и в особенности паутины.
Типичные места обитания фринов — влажные тропические леса. Эти ночные хищники днем скрываются под лежащими стволами деревьев, под отставшей корой, в трещинах скал и т. п., предпочитая в качестве укрытий разного рода плоские полости. Попадая под яркий свет, фрин замирает, распластавшись на субстрате, но, если его тронуть, поспешно убегает. Передвигаются фрины обычно вкось или боком, наподобие крабов. При этом передними ногами они непрерывно ощупывают дорогу в направлении движения. Пищей фринам служат главным образом насекомые, некоторые виды питаются в основном термитами, обильными в тропических лесах. Добыча схватывается педипальпами и разрывается когтевидными члениками хелицер и педипальп. Фрины не слишком прожорливы и могут долго голодать. Замечено, что они ищут и охотно пьют воду. Самки разных видов откладывают от нескольких до 60 яиц, которые носят под брюшком в пергаментоподобной оболочке, образующейся из особых выделений половых путей. Брюшко самки при этом уплощается и в виде свода покрывает сверху яйцевой пакет. Молодь малоподвижна, с прозрачными покровами, несколько дней остается под брюшком самки, линяет и затем расходится. Половая зрелость наступает обычно только на третий год жизни.
Описано около 50 видов фринов, принадлежащих 18 родам двух семейств — Charontidae и Tarantulidae, последнее с тремя подсемействами. Они распространены в основном в тропическом поясе и благодаря влаголюбивости почти не выходят за пределы лесов. Семейство Charontidae преобладает в индо-малайской фауне, часть видов живет в Африке. Представители этого семейства, так же как некоторые виды рода Phrynichus (подсемейство Phrynichinae), живут в пещерах, на сводах которых иногда скопляются большими группами. Среди пещерных есть белесые формы, утратившие боковые глаза. Некоторые виды встречаются в термитниках. Распространение подсемейств семейства Tarantulidae различно. Подсемейство Phrynichinae индо-африканское, подсемейство Damoninae в основном африканское, но есть виды, живущие в Южной Америке, подсемейство Tarantulinae центральноамериканское. Некоторые фрины в тропических странах поселяются в домах. Выходя ночью на охоту, они своим видом нередко приводят людей в ужас, особенно приезжих, незнакомых с этими безобидными животными. Таковы Phrynichus ceylonicus в Индии, Damon medius в Западной Африке и др.
ОТРЯД КЕНЕНИИ, ИЛИ ЩУПАЛЬЦЕХОДНЫЕ (PALPIGRADI)
Кенении — небольшой отряд мелких почвенных арахнид. Это очень своеобразные формы, и не случайно типичный представитель отряда назван кенения удивительная (Koenenia mirabilis). Среди паукообразных кенении демонстрируют примитивный биологический тип мелкого обитателя скважин почвы, приспособленного к жизни в насыщенном влагой почвенном воздухе, — тип, свойственный также низшим почвенным клещам. Благодаря такому образу жизни в организации кенений сохранились многие архаичные черты, чем они представляют немалый интерес для понимания эволюции арахнид. К сожалению, мы еще мало знаем об этих удивительных существах.
Тело кенении удлиненное, всего 0,5—2 мм длиной, очень полно расчленено. Сегменты хелицер, педипальп и двух передних пар ног покрыты сверху головным щитом, а сегменты двух задних пар ног расчленены и имеют самостоятельные тергиты. Расчленена и грудина (стернум). Веретеновидное брюшко состоит из 12 сегментов, предполовой сегмент хорошо выражен. Концевые сегменты брюшка постепенно уменьшаются и заканчиваются длинной гибкой членистой нитью, играющей роль осязательного церка при передвижении в скважинах почвы. Хелицеры длинные, 3-члениковые, с клешнями. Тонкие длинные педипальпы подобны ногам и участвуют в хождении, откуда и пошло название отряда «щупальцеходные». Жевательных отростков на тазиках педипальп нет, а рот расположен на вершине небольшого сосочка между основаниями хелицер — особенность, несвойственная другим арахнидам и, по-видимому, вторичная. Туловище и придатки богато оснащены чувствительными волосками, причем осязание чрезвычайно развито, что, очевидно, очень важно при передвижении между частичками почвы. Ряд примитивных черт сохранился во внутреннем строении кенений. Ясно выражена метамерия мускулатуры туловища, а также кишечника; выделительными органами служат коксальные железы.
Оформленных органов дыхания нет, кенении дышат через покровы, что связано с жизнью во влажном воздухе. Лишь у некоторых видов имеются зачаточные трахеи. Замечательная особенность кенений состоит в том, что на сегментах брюшка у них сохранился набор коксальных органов, представляющих собой рудименты брюшных конечностей (рис. 23, Б). Это парные выпячивающиеся мешочки на восьмом — двенадцатом сегментах, хорошо выраженные у молодых особей и, по-видимому, играющие роль органов чувств, воспринимающих влажность (гигрорецепторы). Наличие таких рудиментов брюшных конечностей указывает на то, что брюшные ножки здесь никогда не были превращены в легкие, т. е. что при выходе на сушу кенении сразу приспособились к жизни в почве и кожному дыханию, миновав стадию легочного дыхания, свойственную эволюции большинства других арахнид. Как мы увидим ниже, аналогичным путем шли клещи отряда Acariformes.
Кенений находят в почве, под камнями, погруженными в почву, некоторые виды обнаружены в пещерах. Они предпочитают воздух, насыщенный водяными парами. В то же время капельная влага для них губительна, так как покровы легко смачиваются. Во влажное время года они могут появляться на поверхности почвы, но большую часть времени проводят скрыто, а по мере просыхания почвы уходят вглубь. Будучи слепы, кенении тем не менее резко отрицательно относятся к свету и поспешно убегают в затененные укрытия. Питание кенений недостаточно изучено. Известно, что они могут высасывать яйца мелких членистоногих: ногохвосток, камподей, многоножек-сколопендрелл, которые также обитают в скважинах почвы и по жизненному облику подобны кенениям. Своеобразие ротового аппарата указывает на какой-то особый способ питания, и, быть может, кенении действительно представляют собой специфических «яйцеедов», тем более что в их кишечнике желток находили не раз. Кладка яиц не сезонна и многократна, яйца откладываются по одному, причем в теле самки обычно видно только два зрелых яйца. Вылупляющаяся молодь похожа на взрослое животное, послезародышевое развитие непродолжительное, сопровождается незначительным ростом, несколькими линьками и изменениями в деталях строения (число волосков, брюшных мешочков и т. п.).
Прежде кенений включали в сборный отряд жгутоногих (Реdipalpi) в качестве подотряда Microthelyphonida. По некоторым признакам кенении действительно напоминают этих арахнид, особенно мелких тартарид, но в то же время сходны с акариформными клещами и отчасти с сольпугами, так что их самостоятельность как отряда не вызывает сомнений. Описано около 20 видов кенений, относимых к 4 родам семейства Koeneniidae. Они распространены в странах с теплым и влажным климатом, найдены в Южной Европе (Южная Франция, острова Средиземного моря), в Африке, Юго-Восточной Азии, Северной и Южной Америке, Австралии. Известны находки в теплицах ботанических садов Западной Европы, куда они попадают с почвой при транспортировке тропических растений. В СССР кенении не отмечены, но их нахождение в южных районах в природе, а также завоз в ботанические сады не исключены.
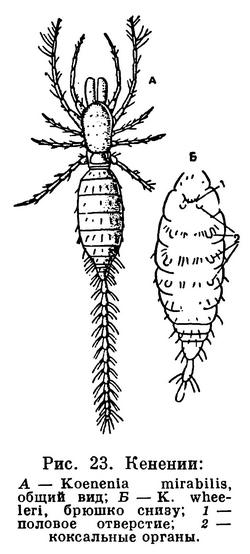
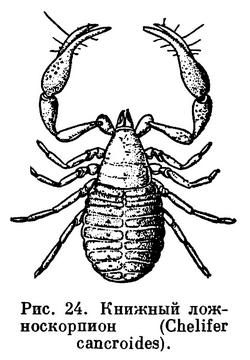
ОТРЯД ЛОЖНОСКОРПИОНЫ (PSEUDOSCORPIONES, ИЛИ CHELONETHI)
Ложноскорпионы — мелкие паукообразные, обычно не превышающие 2—3 мм в длину, в редких случаях до 7 мм. Они довольно широко распространены в природе, но малозаметны, так как живут скрыто, в мхах, почвенном гумусе, под камнями, под отставшей корой, в норах и гнездах позвоночных животных, в пещерах. Есть среди них и обитатели человеческого жилища. Ложноскорпионы получили свое название благодаря тому, что у них педипальпы, как у скорпионов, снабжены клешнями и служат для схватывания добычи. В остальном они заметно отличаются от скорпионов, хотя по ряду признаков так же примитивны. Головогрудь покрыта щитом, в переднебоковых частях которого имеются простые глаза в числе 1—2 пар; некоторые виды слепы. Сегмент четвертой пары ног обычно в той или иной степени обособлен и на головогрудном щите отделен бороздой. Брюшко широкое, сзади закругленное, состоит из 11 сегментов, снабженных тергитами и стернитами. Характерно, что предполовой сегмент, соединяющий головогрудь и брюшко, здесь развит, как соседние. Маленькие хелицеры имеют клешни с гребневидными выростами по внутренним краям. На концах подвижных пальцев хелицер открываются протоки пары паутинных желез, расположенных в головогруди. Педипальпы, помимо хватательной функции, играют роль органов осязания, на клешнях имеются длинные торчащие чувствительные волоски (трихоботрии). Ложноскорпионы очень чутко реагируют на прикосновение, сотрясение субстрата или колебания воздуха. Они прижимают конечности к туловищу и замирают на короткое время.
Все 4 пары ног служат для передвижения. Лапки снабжены парой коготков и маленькой присоской, позволяющей взбираться по гладким поверхностям. Ложноскорпионы в большинстве медлительны, но есть виды и весьма проворные. При движении ложноскорпион держит педипальпы на весу перед собой, изогнув их клешнями внутрь, и поводит ими из стороны в сторону. Будучи потревожен, он убегает, причем может двигаться боком наподобие краба или поспешно пятиться. Дышат ложноскорпионы с помощью трахей, которые открываются 2 парами стигм по бокам второго и третьего сегментов брюшка. От каждой стигмы внутрь отходит короткая трубочка с пучком длинных тонких неветвящихся трахей на конце. Наружный половой аппарат весьма сложен, что связано со своеобразным способом оплодотворения. Во внутреннем строении ложноскорпионов сохраняется ряд примитивных черт: посегментные парные выросты средней кишки и яичников у самок, развитые выделительные коксальные железы и др.
Ложноскорпионы — хищникиу объекты питания разнообразны и: малы: низшие бескрылые насекомые, особенно ногохвостки, мелкие жуки, двукрылые и их личинки, молодь пауков, почвенные клещи, нематоды и др. Жертва схватывается клешнями педипальп, а затем клешнями хелицер, так что педипальпы тотчас освобождаются. Это важно, так как они служат основным органом ориентации и защиты от врагов. Жертва прокалывается хелицерами и высасывается. После этого ложноскорпион долго чистит ротовые органы, трет хелицеры друг о друга, счищает частички пищи с жевательных отростков педипальп, эпистома и верхней губы.
Биология размножения и развитие ложноскорпионов очень своеобразны, и детально изучены французским исследователем М. Вашоном. Оплодотворение происходит без копуляции, с помощью сперматофора, который самец оставляет на субстрате, совершая при этом характерный брачный танец перед пассивной самкой (рис. 25). Особи располагаются передними концами друг к другу, самец высоко поднимается на ножках, вибрирует телом, выпячивает свои цилиндрические органы (пара трубчатых придатков полового аппарата, наполняемых гемолимфой) и производит характерные как бы плавательные движения педипальпами. Во время танца он приближается к самке и, прикасаясь брюшком к субстрату, выделяет каплю вязкой жидкости, вытягивая ее в стержень, на конце которого помещается мешочек со спермиями. Последний снабжен полыми рожками с отверстиями на концах. Затем самка подходит к сперматофору, располагается над ним и вводит рожки сперматофора в отверстия семеприемников. В это время особи сцепляются педипальпами, самец сильно трясет самку и сперма изливается через рожки в семеприемник. Яйца созревают примерно через месяц после этого. К этому времени спермин проникают в просвет яичника и происходит собственно оплодотворение.
Оплодотворенные яйца не откладываются наружу, а выталкиваются в мешковидное выпячивание — выводковую камеру, в которой самка вынашивает молодь. Эта камера представляет собой выпячивание половых путей и связана с яичником. Число яиц варьирует: их бывает обычно 2—3 десятка, у некоторых видов более 50, у других всего 2—3. Развивающиеся эмбрионы прикреплены к стенке камеры. На стадии формирования придатков зародыш одевается первой кутикулой и образуется личинка первого возраста. Она поглощает большое количество желтка, поступающего в камеру из яичника, сильно увеличивается и, разрывая оболочку яйца и стенку выводковой камеры, выходит наружу (рис. 25, В), но остается прикрепленной к камере ротовой областью, через которую продолжается питание желтком. Личинка первого возраста мешковидная и непохожа на взрослого ложноскорпиона. Линяя, она превращается в личинку второго возраста, или протонимфу, похожую на взрослую особь. Она сбрасывает первую кутикулу, покидает мать, выходит из гнезда, в котором находится самка в период размножения, и начинает активно добывать пищу. Далее следуют еще 3 линьки и соответственно превращения в дейтонимфу, тритонимфу и взрослую особь. При каждой линьке ложноскорпион строит гнездо, в котором около недели находится в состоянии покоя. При последней линьке образуется наружный половой аппарат и половые пути открываются наружу. Половая зрелость достигается через год; живут ложноскорпионы 2—3 года; яйцекладок в течение жизни несколько, но число их точно неизвестно.
Гнезда строятся под корой, под камнями, в трещинах почвы и т. п. Гнездо имеет вид колокола диаметром несколько миллиметров. Стенки делаются из растительных остатков, песчинок и т. п., которые прочно переплетаются и склеиваются беспорядочными нитями паутины. В начале постройки ложноскорпион входит внутрь колокола и постепенно наглухо замыкается в нем. Он протягивает в гнезде несколько поперечных нитей и подвешивается на них. Гнездо, как отмечалось, служит для размножения и линьки, а также для зимовки и в каждом случае строится заново. При выходе ложноскорпион отделяет край колокола от субстрата, пустое гнездо остается приподнятым.
У ложноскорпионов наблюдается пассивное расселение на теле других животных — членистоногих и позвоночных. Это явление, называемое форезией, объясняется различно. В некоторых случаях ложноскорпион в поисках пищи схватывает слишком крупную подвижную добычу, которая уносит его с собой. Чаще всего это бывает при попытках нападения на мух (табл. 3, 6). В других случаях ложноскорпионы попадают на тело насекомых, например жуков, или мелких млекопитающих и птиц при охоте за клещами и другими членистоногими, которые паразитируют на этих животных. В результате ложноскорпионы переносятся хозяевами этих паразитов. Самки ложноскорпионов особенно активно ищут пищу в период размножения, и в это время ярко проявляется склонность к форезии.
Ложноскорпионы распространены во всех частях света. Их описано около 1100 видов, которые группируются примерно в 200 родов и 20 семейств. Наиболее обильны и разнообразны они в тропиках. Размещение по различным местам обитания довольно закономерно. Так, виды рода Neobisium живут главным образом в мхах и гумусе, виды Chthonius — под камнями, в трещинах скал, частью в пещерах, а виды Chernes — под корой и в древесной трухе. Есть галофилы, приуроченные к берегам морей, например Garypus beauvoisi, один из наиболее крупных представителей отряда, до 7 мм длиной, живет в скоплениях водорослей по берегам Средиземного моря и выдерживает более или менее длительное затопление морской водой. Многие ложноскорпионы встречаются в норах и гнездах млекопитающих и птиц, а также в муравейниках. Правда, большинство их не специфично в этом отношении, в то время как некоторые очень характерны для таких местообитаний, например: Lasiochernes pilosus в норах крота, мирмекофильный вид Myrmochernes africanus, живущий в муравейниках южноафриканских муравьев рода Camponotus, и др. Относительно многочисленны ложноскорпионы, живущие в пещерах, — троглобионты. Их облик очень характерен: тело стройное, педипальпы и ноги тонкие, длинные, с многочисленными чувствительными волосками, глаз нет (табл. 3, 7). Пещерные виды относятся к родам Blothrus, Chthonius и др. В жилых помещениях и хозяйственных постройках встречается несколько видов ложноскорпионов, среди которых имеются настоящие синантропы, в частности наиболее обычный обитатель жилищ книжный ложноскорпион (Chelifer cancroides), распространенный по всему свету. Этот вид поселяется в книжных шкафах среди книг и бумаг, под отставшими обоями, среди белья и т. п. Он коричневато-бурый, 3—4 мм длиной. Клешни педипальп относительно крупные, массивные. Он медлителен в движениях и одинаково хорошо двигается вперед, боком и вспять. Он живет скрыто и редко попадается на глаза. Питается книжный ложноскорпион главным образом сеноедами, в частности книжным сеноедом, а также хлебными клещами, мелкими личинками жуков. Он полезен, так как в большом количестве уничтожает этих нежелательных обитателей жилища. Синантропными видами считаются также Allochernes italicus, распространенный на юге Европы, и Cheiridium museorum, иногда встречающийся наряду с книжным ложноскорпионом в коллекциях насекомых и гербариях.
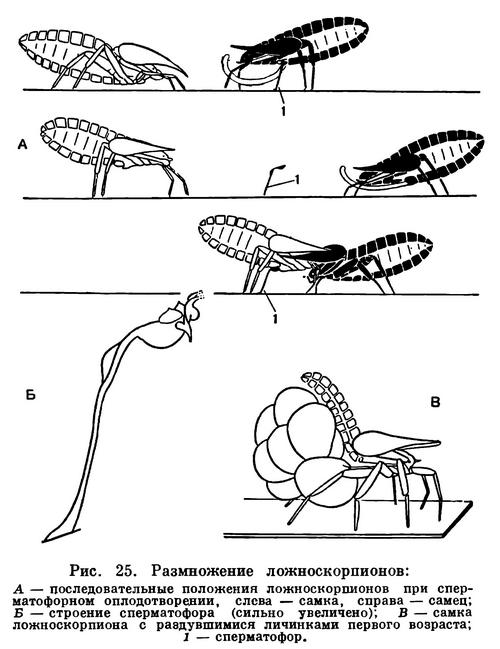
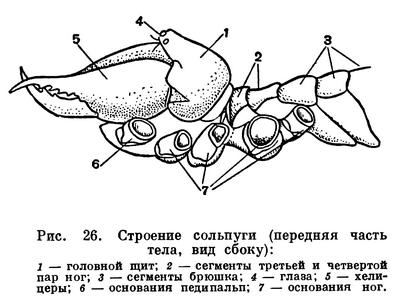
ОТРЯД СОЛЬПУГИ (SOLIFUGAE)
Сольпуги, иначе называемые фалангами, очень своеобразный отряд. Название «фаланги» неудобно тем, что звучит так же, как одно из латинских названий отряда сенокосцев — фалангид (Phalangida). Название «сольпуги» наиболее употребительно, но в принятой латинской транскрипции (Solifugae) оно означает «убегающие от солнца», что не вполне точно, так как среди сольпуг есть и солнцелюбивые виды. Есть еще третье название этого отряда — «бихорки», но оно употребляется редко. В строении и образе жизни сольпуг примитивные черты сочетаются с признаками высокого развития. Наряду с примитивным типом расчленения тела и строения конечностей они имеют наиболее развитую среди арахнид трахейную систему. Будучи в большинстве ночными хищниками, сольпуги распространены в сухих и жарких странах, очень подвижны, и есть виды, активные в дневные часы.
Сольпуги — крупные формы, 50—70 мм длиной, лишь немногие виды не превышают 10—15 мм. Окраска буро-желтая, песчано-желтая, белесоватая, реже пестрая или темная. Головогрудь расчленена, ее передний отдел, несущий хелицеры, педипальпы и передние ноги, покрыт крупным головным щитом, а сегменты второй — четвертой пар ног расчленены и имеют свои тергиты (рис. 26). У переднего края головного щита хорошо заметен глазной бугорок с парой выпуклых глаз; боковые глаза недоразвиты. Нижняя поверхность головогруди занята треугольными тазиками педипальп и ходильных ног. Хелицеры очень крупные со вздутыми основными члениками и мощными клешнями, направленными вперед. Клешни раскрываются в вертикальной плоскости, их пальцы по внутренним краям вооружены крепкими зубцами. Педипальпы большие, по строению подобны ногам, но лишены коготков и имеют на конце особый мягкий осязательный придаток. Функции педипальп разнообразны, они участвуют в передвижении, служат осязательными органами, используются при схватывании и удержании добычи, чему способствуют крепкие щетинки на их члениках; педипальпами самец удерживает самку во время копуляции. Ноги несколько различаются по строению. Передние тоньше и короче других и выполняют главным образом осязательную функцию. Остальные служат для передвижения, их лапки расчленены, снабжены коготками, иногда с маленькой присоской между ними. Задние ноги длиннее других, на их тазиках и расчлененных вертлугах снизу прикреплены 2—5 пар своеобразных органов, так называемых привесок (малеол), имеющих форму треугольных пластинок, подвешенных на стебельках. Назначение этих образований неизвестно, по-видимому, это какие-то органы чувств. Их края снабжены многочисленными сенсиллами с чувствительными клетками, отростки которых объединяются в нерв, проходящий через стебелек каждой пластинки. Опыты по удалению привесок не приводили к изменению поведения сольпуг, в том числе проявления половых инстинктов.
Брюшко крупное, веретеновидное, состоит из 10 расчлененных сегментов с тергитом и стернитом на каждом. Оно сочленяется с головогрудью суженным предполовым сегментом. Половое отверстие имеет вид продольной щели на первом брюшном сегменте, прикрытой боковыми створками. Характерная особенность сольпуг — мощно развитая трахейная система (рис. 27). Основные трахейные стволы открываются парными дыхальцами на брюшке у задних краев второго и третьего сегментов, ближе к средней линии. Дыхальца прикрыты волосками. Кроме этого, имеется непарное дыхальце на четвертом сегменте брюшка и пара дополнительных дыхалец по бокам головогруди. Трахеи, отходящие от дыхалец, соединяются в мощные продольные стволы, которые связаны перемычками и посылают многочисленные сильно ветвящиеся трахеи ко всем органам и тканям. Хорошо выражены спиральные утолщения стенок трахей.
Туловище и придатки сольпуг покрыты многочисленными волосками и щетинками, частью мягкими, частью утолщенными, шиповидными, частью очень длинными, торчащими поодиночке. Лохматый щетинистый покров сольпуг в сочетании с крупными размерами и молниеносными движениями придает им устрашающий облик. Сольпуги бегают с большой быстротой, легко взбираются по вертикальным поверхностям, могут прыгать на значительное расстояние (крупные виды более метра). При встрече с врагом сольпуга принимает угрожающую позу: передний отдел тела поднят, хелицеры с раскрытыми клешнями направлены вперед, педипальпы и передние ноги подняты и направляются в сторону врага. При этом некоторые виды издают пронзительные звуки, писк или стрекотание трением хелицер друг о друга.
Сольпуги в большинстве активны ночью. Днем они прячутся в различных укрытиях, под камнями, в норах грызунов и других животных, или сами роют норы с помощью хелицер, отбрасывая землю ногами. Некоторые используют одну и ту же нору длительное время, другие каждую ночь устраивают новое логово. Ночные виды привлекаются различными источниками света. В жарких пустынных районах сольпуги нередко приходят на свет костра, скопляются под фонарями, проникают в освещенные жилые помещения. Особенно привлекает сольпуг, как и многих ночных насекомых, ультрафиолетовое излучение ртутной лампы. В литературе можно встретить мнение, что сольпуг привлекают насекомые, скопляющиеся у источника света. Хотя сольпуга, прибежавшая на свет, действительно тут же начинает хватать подвернувшихся насекомых, но привлекается она, вероятно, самим источником света, по тем же не вполне ясным для нас побуждениям, которые неудержимо влекут к свету ночных насекомых. Отношение ночных видов сольпуг к дневному свету, напротив, резко отрицательное. Но есть среди сольпуг и дневные, явно солнцелюбивые формы. В Испании сольпуги даже получили местное название «солнечных пауков». У нас к дневным видам принадлежит среднеазиатская сольпуга солнцелюбивая (Paragaleodes heliophilus). Эта небольшая белесовато-золотистая сольпуга бегает днем по степи.
Сольпуги чрезвычайно прожорливы и поедают самых различных животных, с которыми только могут справиться, главным образом насекомых, а также многоножек, пауков, мокриц и др. Они вылавливают термитов, прогрызая стенки их построек; некоторые виды известны в Калифорнии как опустошители ульев. Крупные сольпуги нападают на небольших ящериц, птенцов мелких птиц, детенышей грызунов. В схватках со скорпионами при равных размерах противников обычно побеждает сольпуга. Добыча молниеносно схватывается, крепко удерживается, разрывается и разминается хелицерами. Обильно смоченное пищеварительным соком содержимое добычи всасывается. Отталкивающую картину обжорства сольпуг можно воспроизвести при содержании их в неволе. Если сольпуге предоставить неограниченное количество пищи, например подносить ей пинцетом насекомых, то она наедается настолько, что брюшко раздувается и может даже лопнуть. Такая обреченная на гибель сольпуга тем не менее продолжает схватывать и поедать подносимую ей пищу, пока хелицеры не перестанут двигаться. В природе такие случаи, по-видимому, исключены: наевшаяся сольпуга с увеличенным брюшком теряет способность гоняться за добычей, раньше чем насытится чрезмерно.
Ядовитых желез у сольпуг нет. Специальные опыты показали, что и пищеварительный сок, изливаемый при питании, не ядовит. Однако необоснованное мнение о том, что сольпуги ядовиты и очень опасны для человека, довольно распространено среди неспециалистов. Правда, некоторые крупные сольпуги с мощными хелицерами могут поранить кожу человека до крови. Остающиеся на хелицерах частички пищи и развивающиеся на них гнилостные микроорганизмы могут при этом попасть в ранку и вызвать воспаление. Но большинство видов сольпуг, по-видимому, вообще неспособно прокусить кожу человека.
Спаривание происходит обычно ночью. Самец отыскивает самку по запаху. При удалении педипальп, на которых имеются обонятельные органы, самец перестает привлекаться самкой. При спаривании самец очень активен, самка, напротив, впадает в неподвижное состояние, причем самец иногда перетаскивает ее с места на место. Оплодотворение сперматофорное. Самец выпускает на поверхность почвы клейкий сперматофор, в котором заключены спермии, подхватывает его хелицерами и переносит в половое отверстие самки. При этом определенную роль играют специальные копулятивные придатки (так называемые флажки), имеющиеся на хелицерах самца. Весь процесс длится обычно всего несколько минут. Оплодотворенная самка вскоре становится подвижной и весьма агрессивной по отношению к самцу, который поспешно убегает, рискуя иначе быть тут же съеденным. Поведение самца при спаривании представляет цепь рефлекторных актов, которая, раз начавшись, продолжается даже в том случае, если убрать самку или сперматофор.
Оплодотворенная самка особенно прожорлива. После периода питания она вырывает норку с небольшим расширением у дна, где откладывает яйца (от 30 до 200 у разных видов). Эмбриональное развитие происходит еще в яйцеводах самки, так что из отложенных яиц вскоре вылупляются маленькие сольпуги. Они неподвижны, покрыты тонкой прозрачной кутикулой, лишенной членистости и волосков. Через 2—3 недели происходит линька, новые покровы расчленяются и твердеют, на теле появляются волоски, сольпуга начинает двигаться. Самка остается при молоди, пока она не окрепнет; есть сведения, что мать доставляет детенышам пищу. Общее число линек и продолжительность жизни сольпуг точно неизвестны. В жизни сольпуг выражены сезонные явления. Зимой они находятся в спячке, некоторые виды исчезают и в самые сухие летние месяцы.
Сольпуги — обитатели степных и пустынных стран тропического, субтропического и отчасти умеренного поясов. Лишь немногие виды встречаются в более влажных лесных местностях. Всего известно до 600 видов, распределяющихся примерно в 30 родов и 10 семейств (число родов очень различно у разных авторов). Преобладают они в Старом Свете, где насчитывается 8 семейств и около 500 видов. В Южной и Северной Америке встречаются представители двух семейств — Eremobatidae и Ammotrechidae, свойственных только Новому Свету. В Старом Свете сольпуги распространены почти по всей Африке (исключая Мадагаскар, где они не найдены), в странах Передней, Центральной и Южной Азии, но в юго-восточной части Азии их почти нет. Нет их в Австралии и на островах Тихого океана. В Палеарктике по числу видов преобладают семейства Galeodidae, Rhagodidae, Karschiidae и Daesidae. В Южной Африке наряду с другими имеются два эндемичных, свойственных только этой части континента семейства. Одно из них — семейство Hexisopodidae — представлено своеобразными роющимися почвенными формами с компактным телом, очень мощными хелицерами и уплощенными ногами (табл. 3, 8). В Западной Европе сольпуги встречаются на Балканском и Пиренейском полуостровах.
Данные о сольпугах СССР сведены в монографии А. А. Бялыницкого-Бирули («Фауна СССР», 1938). В нашей фауне отмечено до 50 видов 10 родов четырех упомянутых выше палеарктических семейств. Сольпуги очень обычны в Средней Азии, встречаются также на Кавказе, в Закавказье (за исключением гор Главного хребта и Черноморского побережья), в Крыму, на юге степной полосы от низовий Дона до реки Урала. Северная граница их распространения проходит от 47° с. ш. на западе до 52° с. ш. на реке Урал и далее по параллели к границам Монголии.
К роду Galeodes принадлежат наиболее крупные длинноногие, быстро бегающие формы. В Европейской части СССР (в Крыму, на Кавказе, в степях юго-востока) широко распространена обыкновенная сольпуга (Galeodes araneoides, табл. 2), бледно-желтая, до 5 см длиной. В Средней Азии наиболее распространенный вид — закаспийская сольпуга (Galeodes caspius), буровато-рыжая с сероватым брюшком, несущим темные полоски. Она бывает до 6,5 см в длину и уступает по размерам только черно-бурой дымчатой сольпуге (Galeodes fumigatus), достигающей 7 см и распространенной в песках Туркмении. Закаспийская сольпуга местами очень обычна. Она нередко забегает в дома на свет лампы и приводит в ужас жителей. Как и все ночные сольпуги, она к утру забирается в какое-либо укрытие и может неожиданно оказаться в снятой обуви, складках висящей одежды и т. п.
Представители рода Rhagodes встречаются у нас в Закавказье и в Туркмении. Это формы с короткими ногами, пестро окрашенные или темные. Они живут обычно в глинистых и полынных пустынях и редко заходят в населенные места. Виды рода Karschia — небольшие желтые сольпуги — живут в горных местностях Закавказья и Средней Азии. Некоторые из них найдены на высоте до 3000 м над уровнем моря.
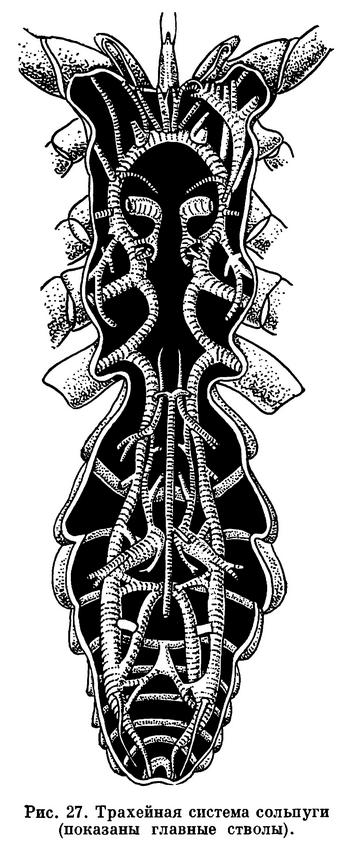
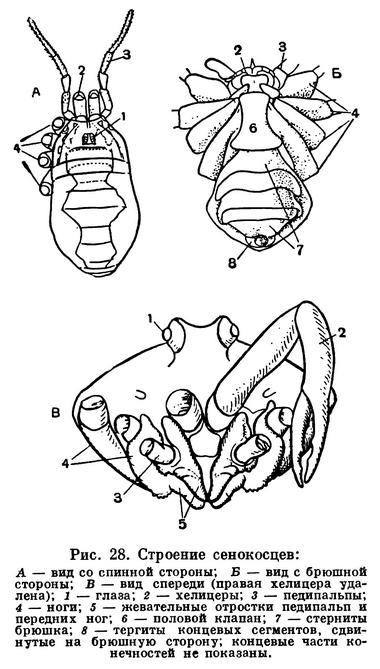
ОТРЯД СЕНОКОСЦЫ (OPILIONES, ИЛИ PHALANGIDA)
Сенокосцы на первый взгляд напоминают длинноногих пауков, и в научной литературе их иногда называют ложнопауками. В действительности родство этих отрядов ограничивается лишь принадлежностью их обоих к арахнидам. Отличить сенокосца от паука легче всего по строению брюшка, которое у сенокосцев расчленено и соединяется с головогрудью широким основанием, а не стебельком, как у пауков. Облик сенокосца с непомерно длинными ногами, на которых подвешено компактное тельце, может показаться странным, даже нелепым, но, как мы увидим, это есть одна из совершенных форм приспособления к жизни на суше.
Туловище сенокосцев небольшое, 1—10 мм длиной, редко около 2 см, овальное, иногда угловатое или уплощенное. Длинные ноги свойственны большинству сенокосцев, но не всем. Иногда ноги чрезвычайно длинны, например при величине туловища 0,6 см ноги имеют по 16 см в длину, т. е. занимают окружность более 30 см в диаметре. В большинстве случаев покровы очень твердые, панцирные, у части форм более эластичные. Головогрудь покрыта щитом, на котором иногда сохраняются границы сегментов третьей и четвертой пар ног. В передней части щита имеется пара медиальных глаз, нередко расположенных на возвышении.
Брюшко короткое, состоит из 9—10 плотно сомкнутых сегментов. Укорочение брюшка сенокосцев обусловлено тем, что тергиты задних сегментов смещены на брюшную сторону, а стерниты и анальное отверстие сдвинуты вперед, т. е. продольная ось брюшка искривлена. Часто наблюдается срастание тергитов друг с другом и с головогрудным щитом, так что образуется общий спинной щит, который загибается краями на боковые и заднюю поверхности туловища. У ряда форм почти все щитки срастаются в сплошной твердый панцирь, гладкий или бугорчатый. Окраска сероватая, буроватая или черная, но есть пестрые формы с красивым цветным рисунком, серебристыми пятнами, металлическим блеском.
Хелицеры 3-члениковые, с клешнями. Обычно они невелики, но у некоторых: видов, питающихся наземными моллюсками, они длиннее туловища. Педипальпы или небольшие щупальцевидные, или более массивные, хватательные, с когтем на конце и шипами на члениках. Тазики ног расположены звездообразно и занимают почти всю нижнюю поверхность головогруди. Жевательные лопасти имеются не только на тазиках педипальп, но и на тазиках передних ног. Членики, ног очень длинны, а лапки вторично расчленены, гибкие, нередко жгутиковидные, причем число члеников лапок может превышать сотню. На лапках имеются маленькие коготки, иногда двойные.
Интересен принцип работы таких многочлениковых лапок. Сгибание производится мышцей, подтягивающей длинное сухожилие, которое проходит сквозь членики к концу лапки, а разгибание — давлением гемолимфы, заполняющей лапку, т. е. мышечно-механический принцип сочетается с гидравлическим. Выгода такого сочетания понятна, иначе возникла бы необходимость образования огромного числа отдельных крошечных мышц, двигающих каждый членик, что, помимо чисто анатомических неудобств, неизбежно снизило бы общее усилие сгибания. Лапки могут обвиваться вокруг стеблей трав и других тонких предметов, крепко удерживая сенокосца.
У большинства длинноногих сенокосцев ноги легко отрываются (так называемая автотомия). Если схватить сенокосца за ногу, то она остается в руках, а сенокосец убегает. Нога отделяется в месте сочленения тазика и вертлуга, причем ранка тотчас затягивается и гемолимфа не вытекает. Это явление рассматривают как средство спасения от врагов, чему, впрочем, нет прямых доказательств. Оторвавшаяся нога продолжает довольно долго, до получаса, ритмически сокращаться, откуда, говорят, и пошло название сенокосцев. Не исключено и другое толкование: сенокосцы бывают обильны на лугах в конце лета и бросаются в глаза во время сенокоса.
Органы чувств, осязательные и обонятельные, сосредоточены у сенокосцев на педипальпах и ногах. При движении субстрат ощупывается главным образом ногами второй пары, обычно наиболее длинными. Отрицательное отношение сенокосцев к запахам некоторых летучих веществ установлено экспериментально. Зрение в большинстве случаев развито слабо, но у некоторых форм медиальные глаза очень крупные, выпуклые, расположены на возвышении. Вообще же реакции на различные раздражители у сенокосцев более вялы, чем у большинства арахнид, и ответом нередко бывает не бегство, а замирание (танатоз).
Наружный половой аппарат устроен иначе, чем у других арахнид. Половое отверстие у самки открывается на конце трубчатого втяжного яйцеклада, расположенного в основании брюшка и во втянутом состоянии прикрытого половым щитком. Яйцеклад выдвигается из-под щитка вперед и нередко бывает значительно длиннее туловища. У самца имеется трубчатый копулятивный орган, расположенный также в основании брюшка. Половой диморфизм обычно не резкий и проявляется в общих размерах, строении хелицер, пропорциях ног и т. п.
Дышат сенокосцы с помощью трахей, хорошо развитых (рис. 17, В). В основании брюшка, по сторонам от полового щитка открывается пара дыхалец, по строению очень похожих на дыхальца насекомых. Они снабжены решетчатым фильтрационным аппаратом и могут плотно замыкаться. От каждого дыхальца отходит крупный трахейный ствол, посылающий многочисленные ветвящиеся трахеи во все части тела. Стенки трахей имеют спиральные утолщения, препятствующие их опаданию при сгибах.
Сенокосцы распространены очень широко, от тропических до полярных стран, и встречаются в самых различных местах обитания, от влажных лесов и лугов до сухих степей и пустынь; некоторые виды поднимаются высоко в горы до границ вечных снегов. Сенокосцы нередки в поселках и больших городах. Большинство сенокосцев активно в сумерки или ночью, но среди них много и дневных форм, в том числе не боящихся палящего солнца. Обычно они днюют или ночуют (в зависимости от времени активности) где придется, не заботясь слишком об укромном убежище. Правда, среди сенокосцев, особенно мелких и не очень длинноногих, немало таких, которые явно предпочитают сильно увлажненные места обитания и постоянно живут в лесной подстилке, влажных мхах, близ водоемов и т. п.
Сенокосцы сравнительно быстро передвигаются на своих длинных ногах, легко взбираются по скалам, стволам деревьев, по стенам, где их можно видеть замершими в характерной позе с плоско раскинутыми ногами. Они очень ловко пробираются среди травянистой растительности, особенно среди параллельно стоящих стеблей злаков. Обвивая стебли своими гибкими лапками, сенокосец легко движется в этой гладкоствольной чаще. Есть среди сенокосцев и очень медлительные формы, иногда настолько, что их легка принять за неживых.
По типу питания сенокосцы заметно отличаются от большинства арахнид. Они: по преимуществу также хищники, питающиеся мелкими членистоногими, главным образом насекомыми; некоторые виды специализировались как потребители наземных моллюсков. Но поглощается не только жидкое содержимое жертвы, но и твердые частички. При этом многие виды наряду со свежей охотно питаются разложившейся животной пищей, а нередко и растительной, вплоть до гниющих остатков с обильной микрофлорой, кусочков плодов и т. п. Сенокосец орудует при этом хелицерами, отрывает ими кусочки пищи и подносит ко рту. Дальнейшее размельчение пищи производится жевательными отростками педипальп и передних ног, которые у сенокосцев бывают отчленены и функционируют как челюсти, — случай среди арахнид редкостный.
По биологии размножения сенокосцы также отличаются от большинства арахнид, скорее напоминая насекомых. При спаривании наблюдается настоящая копуляция без образования сперматофоров. Пара удерживается хелицерами и сплетается ногами, соприкасаясь передними концами, или самец взбирается на спину самки, причем копулятивный орган вводится в расширение половых путей самки (вульву). Не наблюдается и специфических брачных танцев, сопровождающих у других арахнид сперматофорное оплодотворение. Зато нередки жестокие драки самцов за самку.
Яйца откладываются в почву, во влажные мхи, под опавшую листву и т. п. Самка обычно долго ощупывает субстрат яйцекладом и, выбрав подходящее место, погружает его на всю длину (табл. 4, 8). Некоторые тропические сенокосцы с коротким яйцекладом откладывают яйца более поверхностно. Яйца обволакиваются клейкими выделениями. Их число в кладке варьирует от нескольких десятков до нескольких сотен. Кладки могут быть повторными, причем общая плодовитость одной самки может достигать 600 яиц. Известны случаи девственного размножения, но развитие партеногенетических яиц обычно приостанавливается. Вылупляющиеся маленькие сенокосцы похожи на взрослых. По мере роста они периодически линяют 5—7 раз, после чего становятся способными размножаться. У некоторых сенокосцев молодь сильно отличается от взрослых по форме тела, строению панциря и другим признакам, так что развитие протекает с элементами метаморфоза.
Большинство сенокосцев, по крайней мере в умеренных широтах, имеет одногодичный жизненный цикл. Они появляются в начале лета, размножаются в июле — августе, зимуют яйца и молодь. Некоторые виды успевают проделать две генерации в сезон, и зимует молодь осеннего поколения. Известны случаи появления маленьких сенокосцев в зимние оттепели на снегу вместе с некоторыми видами пауков. При всей обычности сенокосцев численность большинства видов не бывает велика, и, может быть, в этом причина того, что они не составляют сколько-нибудь существенного компонента питания ни насекомых, ни позвоночных. Есть также мнение, правда экспериментально не проверенное, что защитную роль играют пахучие головогрудные железы сенокосцев, секрет которых имеет иногда довольно резкий иодистый запах. К тому же сенокосцы в природе малозаметны, особенно среди растительности, и бросаются в глаза лишь в необычной для них обстановке, когда это длинноногое существо, например, окажется на белой стене или бежит в городе по асфальту.
Жизненная форма сенокосцев интересна во многих отношениях. По общему характеру приспособлений к жизни на суше они более, чем любой другой отряд арахнид, напоминают насекомых, не летающих или мало пользующихся крыльями, например некоторых жуков. Здесь также компактное туловище и плотные, нередко панцирные покровы, служащие защитой от потери влаги и механических повреждений, сочетаются с хорошо развитым трахейным дыханием. Сходство наблюдается и в других признаках. Настоящая копуляция без сперматофоров, откладка яиц в толщу субстрата с помощью яйцеклада, многоядность, способность питаться не только жидкой, но и твердой пищей, в том числе растительной, пережевывание пищи — все это признаки, распространенные у насекомых, а не у паукообразных. Иначе говоря, создается определенное впечатление, что сенокосцы в своей эволюции на суше избрали пути, параллельные тем, которыми шли насекомые, и уподобились им в такой мере, в какой позволила их арахнидная природа
В освоении суши насекомыми существенную, если не решающую, роль сыграл полет. У сенокосцев крыльев, конечно, нет, но зато очень длинные ноги обеспечивают им весьма экономный и совершенный способ движения и активного расселения. Главный смысл этого способа состоит в том, что при относительно небольшой частоте мышечных сокращений и минимальной трате энергии скорость перемещения достаточно велика. Сенокосцев правильнее назвать не бегающими, а шагающими формами, не бегунами, а скороходами. Выгода этого способа движения особенно отчетлива в умеренном климате, где в силу более низкой температуры, особенно ночью, развить высокий ритм мышечных сокращений трудно. И действительно, большинство наиболее длинноногих форм (например, семейство Phalangiidae) — жители умеренных широт, активные к тому же до глубокой осени. В жарких и сухих местностях у дневных форм длинноногость, вероятно, имеет и другое значение — как средство защиты от перегрева; туловище сенокосца высоко поднято над поверхностью почвы, раскаляющейся в дневные часы. Они более воздушные, чем наземные животные. При этом сильно возрастает, конечно, угроза потери влаги через покровы, но как раз почти все жители жарких стран (подотряд Laniatores) — формы панцирные.
Жизненная форма сенокосцев оказалась весьма эффективной, о чем свидетельствует их широкое распространение и видовое разнообразие. Если отвлечься от пауков и клещей, имеющих свои, особые пути эволюции, то сенокосцы по числу видов оказываются на первом месте среди остальных отрядов. Их насчитывается более 2500 видов, причем эта цифра явно меньше действительной. Сенокосцы, особенно тропические, изучены очень неполно, и появляются многочисленные описания новых форм.
В последнее время сенокосцы приобрели практический интерес. Установлено, что секрет головогрудных желез некоторых сенокосцев содержит антибиотические вещества, действующие на микроорганизмы. Вещество, обладающее такими свойствами (гонилептидин), выделено из секрета латеральных желез головогруди южноамериканских сенокосцев семейства Gonyleptidae.
Сенокосцы представляют интерес еще в одном, несколько неожиданном отношении, а именно техническом. Дело в том, что они удивительно похожи на один из вариантов разрабатываемых сейчас автоматов, предназначенных для обследования поверхности других планет. Такой автомат мыслится как шагающее устройство, которое имеет ряд преимуществ перед автоматом на колесном или гусеничном ходу, особенно в смысле ориентации и преодоления препятствий. Аппаратура, предназначенная для исследовательских работ и движения, сосредоточена в герметизированной капсуле, возвышающейся на нескольких коленчатых длинных ногах. На концах ног и в суставах имеются датчики, посылающие информацию о касании ноги к субстрату и ее относительных положениях, в соответствии с чем в каждый момент программируются дальнейшие согласованные движения. Шагающий автомат сможет ориентироваться «на ощупь», обходить и преодолевать препятствия. Хотя конструкторы автоматов, очевидно, не имели в виду сенокосцев, но технические принципы действия совпадают даже в деталях. Например, считают, что для приведения в движение ног наиболее целесообразно то самое сочетание механического и гидравлического принципов, которое наблюдается в лапке сенокосца: сгибание за счет тяжа, проходящего через членики к концу ноги, и разгибание давлением жидкости, заполняющей ногу. Быть может, изучение сенокосцев как живых моделей шагающих самоориентирующихся устройств помогло бы в решении некоторых технических задач, т. е. эти арахниды стали бы одним из объектов бионики — новейшей отрасли знания, использующей принципы функционирования живых существ в технике.
Отряд Opiliones делится на три подотряда: Cyphophthalmi, Laniatores и Palpatores.
Небольшой подотряд Cyphophthalmi (одно семейство Sironidae с 10 родами и 40 видами) — наиболее примитивные сенокосцы. Они малы, 2—3 мм в длину, с продолговато-овальным туловищем и короткими ногами, по облику напоминают некоторых клещей. Покровы очень твердые. По бокам головогруди имеются два выроста с отверстиями пахучих желез.
Cyphophthalmi — древнейшие сенокосцы, известные из каменноугольных отложений. Здесь найдены некоторые похожие на них формы (ископаемый отряд Architarbi и др.), дышавшие с помощью легких (табл. 4, 2). В те времена длинноногих сенокосцев, по-видимому, еще не было.
Подотряд Laniatores (табл. 4, 3,4) почти целиком тропический, самый большой и наименее изученный. Известно более 1500 видов и 500 родов, группируемых в 6 семейств. Эти сенокосцы имеют хватательные педипальпы с когтями и шипами и очень твердые покровы. Тергиты брюшка и головогрудной щит спаяны в панцирь. Брюшко короткое, иногда малозаметное. Задние ноги обычно утолщены, особенно у самцов. Туловище и придатки бугристые, нередко с причудливыми выростами. Многие виды пестро окрашены.
Подотряд Palpatores включает подавляющее большинство сенокосцев нашей фауны. У них педипальпы тонкие, не бывают хватательными. Покровы более эластичные, но бывают и панцирные. Половой клапан крупный, выступающий между тазиками ног. Яйцеклад самок обычно очень длинный. Ноги в большинстве случаев очень длинные, тонкие. Сюда относится около 900 видов и 130 родов, группирующихся в 4 семейства. Эти сенокосцы распространены по всему свету, но наиболее многочисленны в Голарктической области.
Наиболее распространенный обыкновенный сенокосец (Phalangium opilio) — эвритопный вид: его можно встретить и на лугу, и в лесу, и близ жилья, на стенах, заборах, нередко в больших городах. Туловище буроватое, у самца 4—7 мм, у самки до 9 мм длиной. Ноги у самца длиннее, чем у самки, до 54 мм. Хелицеры самца с крупными коническими выростами на втором членике. Обыкновенный сенокосец наиболее активен в сумерки и в первую половину ночи, но его нередко можно видеть разгуливающим и днем или распластавшимся на стене под лучами солнца. Он питается главным образом насекомыми, но не отказывается от растительной пищи. Размножение происходит с июля по октябрь, зимуют яйца и молодь. Распространен этот вид в Евразии и Северной Америке. Сходный по биологии вид Opilio parietinus обычен в степной полосе.
ОТРЯД РИЦИНУЛЕИ (RICINULEI)
Очень своеобразный и малочисленный тропический отряд, насчитывающий всего около 15 видов. Это небольшие арахниды, 5—10 мм длиной, с твердым панцирным покровом. Головогрудной щит цельный, но его передний отдел превращен в подвижную крышечку, прикрывающую ротовые конечности. Брюшко короткое, внешне на нем различимы границы трех сегментов, два-три передних сегмента недоразвиты. На конце брюшка имеется маленькое заднебрюшье (остаток метасомы) с анальным отверстием. Хелицеры короткие 2-члениковые, со своеобразной клешней. У самцов ноги третьей пары преобразованы в копулятивные органы. Дышат хорошо развитыми трахеями. Из яйца выходит шестиногая личинка, которая затем превращается в восьминогую форму, чем рицинулеи напоминают клещей.
Распространены рицинулеи в Западной Африке и Южной Америке. Известно, что они влаголюбивы, живут скрыто, под опавшими листьями, под корой, некоторые найдены в пещерах. Двигаются рицинулеи очень медленно, ощупывая дорогу передними ногами, на прикосновение отвечают замиранием и долго остаются неподвижными. Имеется одно семейство Ricinoididae с двумя современными родами, близкими к некоторым ископаемым карбоновым формам.
ОТРЯД ПАУКИ (ARANEI)
Латинское название отряда идет от Линнея, который в 1735 г. в своей «Системе Природы» объединил пауков в род Araneus и отнес их со всеми другими членистоногими к классу насекомых (Insecta). Клерк в 1757 г. уже рассматривал пауков как особую группу, отличную от насекомых, и называл их Aranei. В дальнейшем латинское название отряда писалось различно (Araneides, Araneina, Araneae и др.). На Международном зоологическом конгрессе в 1948 г. было решено восстановить приоритет Клерка и называть пауков Aranei.
Пауки — самый большой отряд арахнид. Их описано более 20000 видов, причем специалисты считают, что эта цифра в дальнейшем значительно возрастет, так как фауна пауков земного шара изучена очень неравномерно и неполно. Пауками заселена вся суша. Подобно насекомым и клещам, они живут повсюду, и в природе буквально не найдется уголка, где не было бы тех или иных видов пауков.
Чтобы перейти к более обстоятельному рассмотрению пауков, попробуем сразу же уяснить природу этого огромного отряда и особенности его разнообразия. Мы вкратце уже упоминали о том, что решающее значение в эволюции пауков имела паутина. Действительно, во всех главных жизненных проявлениях, поддерживающих существование вида, — добывании пищи, размножении, расселении и переживании неблагоприятных условий — пауки пользуются паутиной. Из нее делается убежище и ловчее устройство, с ее помощью происходит сложная процедура спаривания, из нее плетется яйцевой кокон и зимовочный мешок, на ней молодь разносится ветром и т. д. Паук взаимодействует с окружающим миром не столько непосредственно, как другие животные, сколько через свои паутинные приспособления, которые у каждого вида соответствуют его жизненным нуждам и той конкретной обстановке, в которой он обитает. Иначе говоря, взаимоотношения с окружающей средой осуществляются у пауков через паутинную деятельность (Этот термин кажется нам наиболее подходящим. В иностранной литературе деятельность пауков, связанную с паутиной, принято называть индустрией — словом, которое в русском языке имеет более узкий смысл применительно к деятельности человека и потому вносит ненужный антропоморфизм, мешающий пониманию поведения животных.), которая, как и все поведение пауков, имеет в своей основе инстинкты. Сравнительное изучение пауков показывает, что эволюция паутинной деятельности, эволюция инстинктов есть то ведущее направление эволюционного развития пауков, на котором этот своеобразный отряд достиг небывалого расцвета.
Наглядным подтверждением служит характер разнообразия пауков. Паутинные приспособления представляют эволюционные ряды от очень простых до чрезвычайно сложных и совершенных, будь то яйцевые коконы, логовища и гнезда или ловчие сети. Одновременно усложняются цепи инстинктов, направленных на создание паутинных устройств. Замечательно, что общий тип строения паука при этом стойко сохраняется. Очень различны размеры пауков, окраска, внешняя форма, изменяется строение отдельных органов, но все это бесконечное многообразие заключено в рамки некоего стереотипа. Паук — всегда паук. Единство выдерживается и в ряде особенностей биологии, типе питания, индивидуального развития и т. д.
Показательно сравнение с другими арахнидами. Мы видели, что в каждом отряде также имеется свой устойчивый тип организации, однако при этом видовое разнообразие большинства отрядов не идет в сравнение с паучьим. Если же обратиться к другой группе многовидовых отрядов — клещам, то там обилие видов зиждется на глубоких и различных изменениях организации и развития подчас настолько, что сразу и не узнаешь, клещ это или что-то другое. Иначе говоря, клещи, приспособляясь к новым условиям, сильно менялись сами, пауки же изменяли свои паутинные приспособления, сохраняя тип организации.
В чем же главные особенности организации пауков и почему паутина приобрела такое значение в их жизни и эволюции?
Паутинный аппарат образовался у пауков, если можно так выразиться, в счастливом пункте — на нижней стороне брюшка. В брюшке нашлось достаточно места для развития объемистых паутинных желез, обеспечивших обилие паутины. Функцию выведения секрета желез наружу взяли на себя брюшные конечности, которые превратились в паутинные бородавки и сохранили подвижность, что очень важно при пользовании паутиной. Очень подвижным стало и брюшко, которое у всех пауков соединяется с головогрудью тонким стебельком — сузившимся предполовым сегментом. Таким образом, целый отдел тела — брюшко — превратился в подвижную паутинную «фабрику». Хорошо развитые, расчлененные и весьма подвижные конечности головогруди, свойственные паукам, как и всем арахнидам, оказались весьма подходящими для прядения самых различных паутинных приспособлений. Добавились лишь некоторые детали: гребенчатые коготки, ряды щетинок для расчесывания паутины и т. п. Важная особенность, дополняющая жизненный облик паука, — это ядовитые железы, открывающиеся на когтевидных концевых члениках хелицер. Ядовитый аппарат как средство нападения и защиты возник у пауков еще до того, как они стали устраивать ловчие приспособления, у бродячих форм, но наибольшего развития ядовитые железы достигают у тенетных пауков. Трудно представить более совершенный способ убивания добычи, попавшей в тенета, чем впрыскивание яда хелицерами. Ведь иначе схватки с добычей, бьющейся в тенетах, очень быстро приводили бы последние в негодность.
Заметим, что паутиной пользуются и некоторые другие арахниды — ложноскорпионы, паутинные клещи. Но у них паутинные железы расположены иначе, например в хелицерах, и по одному этому не могли получить такого развития, как у пауков. По обилию секрета с пауками могут сравниться только гусеницы бабочек с их шелком, кстати, близким к паутине по химическому составу. Однако бабочки имеют полное превращение, шелковые железы не переходят от гусениц к взрослым бабочкам, и применение шелка ограничивается плетением кокона. У пауков же, развивающихся без метаморфоза, паутинные железы сохраняются на всех активных стадиях. Паутина используется на протяжении всей жизни, вплоть до размножения и заботы о потомстве.
Как возникли пауки, мы не знаем. Это очень древний отряд, известный из девонских и каменноугольных отложений, но уже в те отдаленные времена пауки были похожи на современных, правда наиболее примитивных. Можно только сказать, что наиболее характерная особенность пауков — паутинный аппарат образовался у их предков в самом процессе выхода на сушу, а быть может, еще в воде. Доказательство этого — паутинные бородавки. Ведь у всех хелицеровых при выходе на сушу брюшные жаберные ножки либо превращаются в легкие и иные специальные органы, либо атрофируются. Жаберные ножки как таковые на суше немыслимы. Поэтому паутинные бородавки могли оформиться только у водных или амфибиотических форм. Они образовались у пауков из ножек десятого и одиннадцатого сегментов, а ножки восьмого и девятого превратились в легкие. Все это показывает, что пауки вышли на сушу своим путем, независимо от других арахнид. Даже наиболее близкий к паукам примитивный отряд — фрины не могут считаться их предками, хотя бы потому, что ножки, соответствующие паутинным бородавкам, у них атрофированы. Каково было назначение паутинного аппарата первоначально? Вероятнее всего, паутина использовалась для яйцевых коконов, как у тех современных пауков, у которых в остальном паутинная деятельность еще мало развита. В дальнейшем паутина стала все более входить в жизнь пауков. С ее помощью выработались своеобразнейшие формы спаривания, стали устраиваться логовища, гнезда, а затем и ловчие приспособления самых различных конструкций.
Признаки, по которым можно безошибочно отличить паука: резкое разделение головогруди и брюшка, хелицеры с когтевидным концевым члеником, небольшие щупальцевидные педипальпы, паутинные бородавки на брюшке.
Форма тела большинства пауков довольно однотипна. Головогрудь, как правило, меньше брюшка, спереди сужена и притуплена, брюшко яйцевидное. Уклонения многочисленны, встречаются в разных семействах. Есть пауки сильно вытянутые, с длинными тонкими ногами, которые они вытягивают вдоль тела, делаясь незаметными среди узких листьев злаков и осок, где они живут. У других, живущих под корой, на стволах деревьев, на цветах, тело уплощено, брюшко нередко расширено. У ряда тропических пауков на брюшке бывают длинные шипы, причудливые выросты. Иногда изменяется форма головогруди, например глаза подняты на длинном выросте и т. п.
Размеры варьируют в широких пределах. Самые мелкие пауки — 0,8 мм, самые крупные тропические птицеяды бывают до 11 см в длину, а с ногами до 20 см. У нас крупные тарантулы достигают 3,5 см, а у самок одного дальневосточного вида крестовиков брюшко бывает с грецкий орех.
Окраска очень разнообразна (табл. 6, 7), особенно у тропических пауков. Наряду с темными и невзрачными формами имеется множество ярко окрашенных, одноцветных или со сложным рисунком. Нередок желтый цвет в комбинации с белым. Есть ярко-красные формы или черные с красным брюшком. Многие виды, живущие среди растений, окрашены в различные оттенки зеленого, иногда в комбинации с желтым, бурым и другими тонами. Есть виды с золотыми и серебряными пятнами, металлически блестящие, перламутровые.
Головогрудь пауков покрыта цельным твердым щитом, в передней части которого расположены глаза, обычно четыре пары.
Хелицеры короткие, 2-члениковые. Их когтевидный или серповидный концевой членик, подгибаясь, входит в бороздку на основном членике подобно лезвию перочинного ножа. На его конце открывается проток ядовитой железы. У лифистио- и мигаломорфных пауков хелицеры расположены горизонтально, а концевые членики отгибаются книзу, у высших аранеоморфных пауков основные членики направлены вниз, а концевые — внутрь, навстречу друг другу. Хелицерами паук схватывает и убивает добычу, разминает и разрывает ее, защищается от врагов, разрезает нити паутины, перетаскивает яйцевые коконы, в ряде случаев самец удерживает хелицерами самку при спаривании, норковые виды копают хелицерами землю и т. д.
Педипальпы похожи на ноги, но короче и с одним коготком. Их тазики обычно снабжены лопастями, которые ограничивают предротовую полость и покрыты волосками, служащими для процеживания жидкой пищи. Щупальца педипальп служат органами осязания и в передвижении, как правило, не участвуют. У половозрелых самцов педипальпы видоизменены в связи с их совокупительной функцией, их концевые членики превращены в копулятивные органы, обычно очень сложно устроенные.
Ноги 7-члениковые, их тазики прикрепляются вокруг цельного грудного щитка. На лапках имеются два серповидных, обычно гребенчатых коготка. Между ними расположен непарный придаток (эмподий), также когтевидный, или в виде липкой подушечки. Пауки, делающие тенета, имеют еще добавочные коготки, образуемые щетинками, нередко зазубренными. Ноги покрыты волосками, частью простыми, прилегающими, частью торчащими, различными по форме и размерам. Относительные размеры ног сильно варьируют в зависимости от образа жизни. Иногда отдельные пары ног отличаются от остальных, например хватательные передние ноги некоторых пауков-скакунов. Функции ног разнообразны, помимо передвижения, они могут служить для выкапывания норок, удержания добычи, яйцевого кокона и т. п. С помощью ног плетутся различные паутинные устройства. Ногами паук натягивает и разрывает паутинные нити, отмеривает расстояние между радиусами и оборотами спирали тенет, расчесывает паутину и т. д. Ноги богато оснащены органами чувств, осязательными и обонятельными.
Брюшко у подавляющего большинства пауков нечленистое, его сегменты слиты. Покровы брюшка эластичные, обычно густо опушенные волосками. Лишь в редких случаях на брюшке образуются вторичные склериты. У примитивных лифистиоморфных пауков, в отличие от всех остальных, расчленение брюшка хорошо выражено, все его сегменты имеют тергиты, а на вентральной стороне разделены поперечными бороздками. Следы сегментации брюшка сохраняются у высших пауков в строении мускулатуры, иногда в рисунке брюшка. Сегменты хорошо выражены у зародыша, у молодых паучков желточная масса, заполняющая кишечник, также сегментирована. Судя по всем этим данным, в составе брюшка пауков объединено 11 сегментов, причем тергальные участки развиты полнее, чем стернальные, а несколько задних сегментов в той или иной степени атрофируется. На брюшке открывается половое отверстие, расположены органы дыхания — легкие и трахеи — и паутинные бородавки.
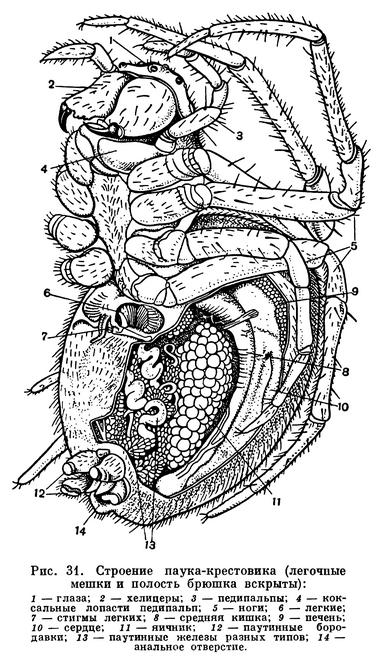
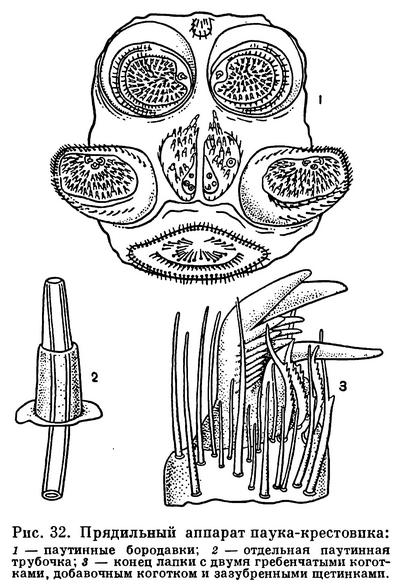
Как говорилось, паутинные бородавки — это видоизмененные брюшные ножки. Они расположены на брюшке снизу перед бугорком с анальным отверстием. На концах бородавок имеются многочисленные хитиновые паутинные трубочки (видоизмененные волоски), которыми открываются протоки паутинных желез (рис. 32).
Объекты питания очень разнообразны. Обычно это самые различные насекомые, подходящие по размеру. В сеть тенетных пауков попадают главным образом летакющие формы, чаще других двукрылые. Паук, высасывающий муху, — картина для каждого само собой разумеющаяся. Видовой состав потребляемых насекомых зависит от местообитания и времени года. Многие пауки, живущие близ почвы или в норках, питаются в основном жуками и прямокрылыми. Некоторые наряду с насекомыми затаскивают в норку и поедают дождевых червей и улиток. Есть пауки, специализировавшиеся в выборе добычи, охотящиеся только на муравьев или только на пауков других видов (сем. Mimetidae). Водяной паук Argyroneta охотится за личинками водных насекомых, рачками, мальками рыб. Громадные тропические пауки-птицеяды при случае убивают и съедают небольших птиц, хотя в основном питаются насекомыми. В неволе птицеяды охотно поедают небольших ящериц, лягушек, мышей. Бразильский птицеяд Grammostola питается молодыми змеями и уничтожает их в большом количестве. Амфибиотические пауки (Dolomedes и др.), бегающие по воде, вылавливают небольших рыбок, головастиков и личинок насекомых.
По способу питания пауки в наиболее законченной форме представляют арахнидный тип. Фильтрующий аппарат предротовой полости и глотки, узкий пищевод, мощный сосательный желудок — все это приспособления для питания жидкой пищей. Поймав и убив добычу, паук разрывает и разминает ее хелицерами, изливая при этом пищеварительный сок, растворяющий внутренние ткани. Выступающая жидкость всасывается. Выделение сока и всасывание капель пищи чередуются, паук поворачивает жертву, обрабатывая ее с разных сторон, пока не останется сморщенная шкурка. Пауки, питающиеся насекомыми с твердым покровом, например жуками, наносят хелицерами рану в сочленовную перепонку, чаще между головой и грудью. Через это отверстие проникает пищеварительный сок и высасывается содержимое.
Общая продолжительность жизни пауков очень различна. Большинство видов заканчивает жизненный цикл в течение года. У форм умеренного климата обычно зимуют яйца в коконе или молодь. Половозрелость достигается в конце лета, и после откладки яиц взрослые пауки гибнут. В этом случае нередко наблюдается осенне-зимняя диапауза, развитие яиц осенью приостанавливается, несмотря на то что в природе еще достаточно тепло, и возобновляется только следующей весной, после длительного зимнего охлаждения яиц. Среди пауков южных, особенно тропических, районов есть немало таких, которые могут размножаться периодически в течение всего года. В зависимости от длительности жизни взрослых особей поколения либо следуют непрерывно друг за другом, либо накладываются во времени. Таковы и некоторые наши домовые формы, например Tegenaria (сем. Agelenidae), живущие по нескольку лет. Наиболее долговечны крупные пауки-птицеяды, которые живут не меньше 7—8 лет, а самка Eurypelma californica жила в неволе даже 20 лет.
Большинство пауков живет оседло в своих постройках — логовищах и тенетах, но немало и бродячих, не имеющих постоянных убежищ. Эволюция паутинной деятельности пауков наглядно проявляется в конструкциях построек.
Логовища делят на несколько типов: норки, трубки, покрышки, колпачки и др. Норки выкапываются в почве и служат постоянным жильем или только убежищем, в котором паук проводит день, выходя ночью на охоту. Известны все переходы от простых углублений в почве до сложно устроенных норок, что хорошо прослеживается у мигаломорфных пауков, у ликозид и ряда других. Одни виды делают небольшие углубления или норки, несовершенные еще по отделке, другие устраивают правильные вертикальные глубокие норки, выстланные паутиной, третьи строят у входа в норку различные дополнительные приспособления из паутины и почвы. Последние совершенствуются в двух направлениях. У одних форм норка удлиняется над входом за счет паутинной надстройки, другими делается подвижная дверца-крышечка, закрывающая вход. Паутинные надстройки бывают в форме небольшой закраины, вертикальной трубочки или воронки, либо паутинная выстилка норки продолжается в длинную надземную трубку (табл. 5, 4,5,11).
Крышечки (табл. 5, 1,2,3) бывают с шарниром и без него. Нередко свободный край паутинной крышечки утяжеляется частичками почвы, так что она сама захлопывается. У некоторых мигаломорфных пауков крышечка утолщена и затыкает норку наподобие пробки. Обычно паук днем сидит в норке, удерживая крышечку хелицерами, а ночью либо выходит наружу, либо подстерегает добычу, высунувшись из норки. Снаружи крышечка бывает замаскирована частицами почвы, растительными остатками и неотличима от окружающего фона.
У мигаломорфных пауков норки нередко усложняются: имеются ответвления, запасный выход с крышечкой и др. Паук Rhytidicolus structor, живущий в Венесуэле, устраивает сложную норку из трех переходящих друг в друга камер с тремя шарнирными крышечками (табл. 5, 6). Защищаясь от врагов, паук удерживает хелицерами наружную дверцу, но, если это не удается, переходит в другую камеру, запираясь следующей дверцей. Замечательны защитные приспособления некоторых пауков семейства Ctenizidae (табл. 5, 7,8). У них брюшко сзади как бы обрублено и образует твердый диск такого же диаметра, как вход в норку. Последняя имеет расширения. Паук сидит вниз головой, закрывая вход в норку диском брюшка, а будучи потревожен, уходит вглубь и закрывает диском вход в следующий узкий отрезок норки. Считают, что совершенствование всех этих защитных приспособлений в значительной мере направлено против ос-помпилов, среди которых ряд видов специально охотится на мигаломорфных пауков (табл. 5, 13).
Морфологические приспособления для рытья и жизни в норках выражены у пауков в разной степени, нередко жизнь в норке почти не сказывается на строении паука. Норки роются хелицерами, которые у пауков хорошо развиты и легко приспосабливаются к этой новой функции; при отделке норок паук пользуется паутиной. Наиболее отчетливы приспособления к жизни в норках у мигаломорфных пауков. Их тело компактное, ноги мощные, на хелицерах имеются зубцы для рытья — так называемые «грабли» (табл. 5, 12). У некоторых форм пара передних глаз расположена так, что их оси совпадают с осью норки.
Паутинные трубки располагаются на почве, среди камней, на коре деревьев, под корой и т. п. Трубка по существу есть паутинная выстилка норки, только вынесенная из нее. Мигаломорфный паук Stothis astuta в зависимости от твердости грунта то роет U-образную норку с двумя крышечками, то строит трубку, покрытую частичками почвы (табл. 5, 9). Типичные жилые трубки встречаются в семействах Agelenidae, Dysderidae, Clubionidae и др., а также у многих мигаломорфных пауков. Нередко трубка продолжается в ловчую сеть, имеющую вид воронки или полога.
Пауки — одни из самых распространенных животных. Наиболее богаты пауками области с обильным растительным покровом, но встречаются они во всех ландшафтных и климатических зонах, от полярных областей и высоких гор до сухих степей и раскаленных пустынь. Пауки найдены в Гренландии близ ледников и на антарктических островах, многие виды обычны в горах на высоте 2—3 тыс. м, а один вид скакунов найден на Эвересте на высоте 7 тыс. м.
Места обитания пауков чрезвычайно разнообразны. Они живут в почве и на ее поверхности, в лесной подстилке, во мху, на травянистой и древесной растительности, под корой, в дуплах, под камнями, в трещинах скал, в пещерах, в норах и гнездах других животных, в жилище человека. Только водная среда почти не освоена пауками. Единственный пример — пресноводный паук Argyroneta, который проник в воду благодаря паутине, дышит атмосферным воздухом и не выработал особых морфологических приспособлений к жизни в воде. Правда, пауков, живущих в очень сырых местах, близ воды, свободно бегающих по ее поверхности, немало. Некоторые виды живут на литорали морских побережий, прячутся во время прилива в своих логовищах из непромокаемой паутины, а при отливе охотятся за мелкими рачками и другими беспозвоночными.
Географическое распространение пауков представляет большой интерес для зоогеографии. Состав фауны пауков очень характерен для различных зоогеографических областей. Различные роды, подсемейства и семейства пауков обладают разной степенью эндемизма, т. е. приуроченности в своем распространении к тем или иным территориям суши, что дает ценный материал для выяснения происхождения фаун различных областей земного шара. Огромное разнообразие мировой фауны пауков не позволяет, однако, в рамках этого издания рассмотреть закономерности их распространения, так как для этого пришлось бы оперировать названиями многочисленных родов и семейств, незнакомых читателю.
Практическое значение пауков при современном уровне наших знаний о них фактически сводится к вреду, приносимому ядовитыми укусами некоторых форм. Однако многие пауки, несомненно, полезны как истребители вредных насекомых, но эта их роль пока точно не оценена. Есть и некоторые перспективы технического использования пауков.
Попытки изготовления ткани из паутины делались с давних времен. Исключительная по прочности, легкости и красоте ткань из паутины известна в Китае под названием «ткани восточного моря». Полинезийцы употребляли паутину крупных тенетных пауков в качестве ниток для шитья и плетения рыболовных снастей. В начале XVIII в. во Франции из паутины крестовиков были изготовлены перчатки и чулки, представленные в Академию наук и вызвавшие всеобщее восхищение. Известно, что нить можно выматывать на катушку прямо из паутинных бородавок крестовика, заключенного в маленькую клеточку, причем от одного паука получается сразу до 500 м нити. В 1899 году из паутины крупного мадагаскарского паука Nephila пытались получить ткань для покрытия дирижабля и удалось изготовить образец роскошной ткани длиной 5 м. Однако попытки акклиматизировать этих пауков во Франции кончились неудачей. Производство паутинного шелка неизменно сталкивается с трудностью массового разведения пауков, прежде всего прокормления этих хищников. Возможно, что разработка искусственных питательных сред позволит решить эту проблему, тем более что искусственное питание гусениц шелкопрядов уже практикуется в Японии. Пока паутина находит применение в оптике для изготовления визиров (перекрещивающихся нитей) в окулярах различных приборов.
Классификация пауков очень сложна, что связано с огромным числом видов при относительной однотипности строения. Деление пауков на три подотряда — Liphistiomorphae, Mygalomorphae и Araneomorphae — наиболее принято. Но в отношении систематического состава родов, семейств и надсемейственных группировок среди аранеологов имеются значительные расхождения. По данным Милло, приводимым А. В. Ивановым, число описанных видов пауков фауны мира приблизилось к 21000. Эти виды объединяются примерно в 2100 родов и 63 семейства. Некоторые из них приведены ниже. По данным Д. Е. Харитонова, к 1953 году в фауне СССР насчитывалось около 1200 видов пауков. Наиболее богата пауками лесная зона, где встречается около 900 видов.
ПОДОТРЯД ЛИФИСТИОМОРФНЫЕ, ИЛИ ЧЛЕНИСТОБРЮХИЕ, ПАУКИ (LIPHISTIOMORPHAE)
Это самые примитивные пауки, известные уже из карбоновых отложений. Брюшко членистое, легких две пары, паутинных бородавок обычно 8, они еще не сдвинуты на конец брюшка. Хелицеры направлены вперед, ядовитые железы небольшие, помещаются внутри хелицер. Педипальпы крупные, почти как ноги. Размеры тела 11—35 мм. К ним относится одно семейство Liphistiidae с двумя родами и 8 видами, встречающимися в Юго-Восточной Азии. Живут в норках с крышечкой-дверцей. Один вид живет в пещерах, где делает на стенах паутинные трубки.
ПОДОТРЯД МИГАЛОМОРФНЫЕ ПАУКИ, ИЛИ ПАУКИ-ПТИЦЕЯДЫ В ШИРОКОМ СМЫСЛЕ (MYGALOMORPHAE)
К этому подотряду принадлежат самые крупные пауки (табл. 7, 1—3). Брюшко внешне не сегментировано, на его конце расположено от 2 до 6 паутинных бородавок. Легких четыре пары. Хелицеры направлены вперед. Ядовитые железы небольшие. Глаза сближены на переднем крае головогруди. Размеры от средних до очень крупных. К подотряду принадлежит более 1500 видов, группируемых в 9 семейств. Это ночные бродячие и более оседлые формы, обитатели норок или паутинных трубок (табл. 5), распространенные главным образом в тропических странах.
ПОДОТРЯД АРАНЕОМОРФНЫЕ ПАУКИ (ARANEOMORPHAE)
У аранеоморфных пауков хелицеры направлены вниз, а их крючки — внутрь. Передняя пара органов дыхания — легкие, задняя — трахеи, за исключением примитивного семейства Hypochilidae с четырьмя легкими и нескольких семейств высших пауков, дышащих только трахеями. К этому подотряду относится большинство пауков фауны мира, которые группируются примерно в 53 семейства. Последние объединяются в несколько надсемейств, триб и секций, состав которых несколько различен у разных авторов.
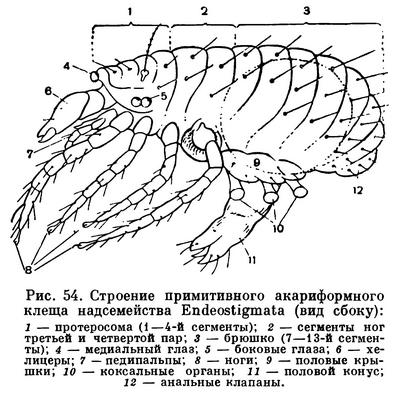
ОТРЯД АКАРИФОРМНЫЕ КЛЕЩИ (ACARIFORMES)
Переходя к рассмотрению клещей, или, точнее, клещеобразных арахнид, напомним, что они представляют собой три неродственных отряда, в такой же мере самостоятельных и обособленных, как и другие отряды арахнид. Два из них, акариформные клещи и паразитиформные клещи, включают огромное число чрезвычайно разнообразных форм. Третий отряд — клещи-сенокосцы, напротив, мал и однотипен.
Как мы говорили, замечательная особенность больших отрядов клещей состоит в том, что их громадное и на первый взгляд хаотичное разнообразие в действительности представляет естественные ряды форм от примитивных исходных до сильно изменившихся в разных направлениях. Поэтому сравнительное изучение современного разнообразия каждого отряда позволяет представить пути и результаты его длительной эволюции и таким образом понять строение, индивидуальное развитие и образ жизни его многочисленных и подчас совсем непохожих друг на друга представителей. Кратко и популярно изложить данные о клещах в такой сравнительной форме нелегко, но другого пути для понимания природы клещей нет. При рассмотрении каждого отряда мы сначала остановимся на наиболее примитивных его представителях, служащих прототипами отрядов, а далее попробуем проследить, как изменялись образ жизни, строение и развитие в пределах отрядов. Конечно, мы сможем коснуться лишь некоторых основных группировок клещей в такой мере, в какой это необходимо для общей картины.
Акариформные клещи — это самый большой отряд клещеобразных, насчитывающий более 6000 видов, причем эта цифра далека от действительной. Благодаря интенсивному изучению клещей, особенно в последнее время, ежегодно появляются описания сотен новых видов. Разнообразие акариформных клещей очень велико. Отряд делится на два подотряда: саркоптиформные клещи (Sarcoptiformes) и тромбидиформные клещи (Trombidiformes). К первому относятся такие обширные и разнообразные группы, как панцирные, тироглифоидные клещи и ряд паразитических — перьевые, волосяные, чесоточные и др. Второй подотряд включает очень много группировок, в том числе паутинных, водяных клещей, краснотелок и ряд других. В этом отряде немало практически важных клещей — вредителей растений и сельскохозяйственных продуктов, паразитов животных и человека. Но переносчиков инфекционных болезней здесь сравнительно немного.
Классификация отряда сложна и находится в стадии разработки. В обоих подотрядах сейчас насчитывается более 170 семейств, а общее число родов трудно указать, настолько оно различно у разных авторов. Современная классификация, несомненно, есть результат чрезмерного дробления систематических группировок, в чем отражается аналитический этап изучения клещей, на котором находится наука о них — акарология. Одно из основных систематических подразделений — семейство обычно понимается акарологами в очень суженном смысле. Есть немало «семейств» акариформных клещей всего с одним-двумя десятками видов, а семейства по нескольку сотен видов составляют скорее исключение. Если в этом отношении сравнить клещей с насекомыми, то семействам последних более соответствуют надсемейства клещей, которыми мы и будем в основном оперировать в дальнейшем.
Наиболее примитивные акариформные клещи — это низшие представители подотряда Trombidiformes — небольшое надсемейство Endeostigmata (рис. 54, табл. 8, 1). К ним близки самые примитивные представители подотряда Sarcoptiformes — клещи-палеакариды надсемейства Palaeacaroidea (рис. 57, 1). Эти два надсемейства служат прототипом отряда в целом, а их близость указывает на единство его происхождения. Эти малоизвестные формы изучались французским акарологом Гранжаном, а у нас — А. А. Захваткиным и автором этого раздела. В последние годы удалось изучить биологию и эмбриональное развитие этих клещей.
Это очень древние клещи, найденные в ископаемом состоянии в среднедевонских отложениях и дожившие до нашего времени почти без изменений. Они живут в почвенном гумусе, и, вероятно, в этом одно из условий их эволюционного консерватизма. Почва — среда сравнительно мало менявшаяся на протяжении геологических эпох, и эти «живые ископаемые» нашли в ней такие условия, в которых смогли сохраниться.
По плану строения примитивные Acariformes соответствуют арахнидам вообще, чем опровергается ходячее мнение о том, что все клещи крайне видоизменились и деградировали. Более того, они сохранили ряд таких примитивных черт, которые давно утрачены другими арахнидами. Эти клещи очень малы, обычно всего 0,2—0,3 мм. Покровы тонкие, лишены щитков. Туловище хорошо расчленено (рис. 54), причем сегментировано не только брюшко, но и головогрудь. Сегменты хелицер, педипальп и двух передних пар ног разделены спинными бороздами и, как у кенений и сольпуг, образуют передний отдел, отвечающий голове трилобитов и называемый у клещей протеросомой. По сторонам этого отдела имеются боковые глаза, а спереди парный по происхождению медиальный глаз. Сегменты двух задних пар ног расчленены. Членистое брюшко состоит из шести сегментов и соединяется с головогрудью хорошо развитым предполовым сегментом. У других акариформных клещей сегментация обычно исчезает. Сохраняется борозда между протеросомой и задним отделом туловища, который образуется путем присоединения сегментов двух задних пар ног к брюшку и называется гистеросомой. Такой тип расчленения тела очень характерен для всего отряда.
Важную роль приобретают волоски, щетинки и другие кожные органы. Благодаря малым размерам клещей эти органы у них бывают относительно очень велики и обычно строго постоянны по числу и положению на сегментах туловища и придатках. Часть из них выполняет функции органов чувств — осязательных, воспринимающих колебания (трихоботрии на переднем отделе тела), обонятельных. Последние представлены лировидными органами и так называемыми соленидиями на ногах. Это тонкостенные замкнутые на конце трубочки со спиральной бороздой, пронизанной мельчайшими канальцами; внутрь соленидия входит отросток чувствительной нервной клетки. Щетинки нередко служат дополнительной защитой, особенно у тонкокожих форм, и бывают очень разнообразны по величине и форме — шиповидные, перистые, плоские, покрывающие клеща, как черепица, и т. п.
Хелицеры и педипальпы у низших Acariformes свободные, не соединяются в «головку», что считалось характерным для всех клещей. Хелицеры обычно 3-члениковые, с клешней, но есть формы, у которых можно различить до шести члеников хелицер, чем последние очень похожи на ноги. Тазики педипальп подвижные и имеют жевательные лопасти. Ноги с одним когтевидным эмподием, еще лишены добавочных коготков, которые в дальнейшем образуются из щетинок.
Строение ротовых органов в деталях довольно разнообразно уже у низших форм. Как удалось выяснить, это связано с разнообразием питания. Одни из них жующие растительноядные, потребляют гифы и споры грибов, почвенные водоросли, дрожжи и т. п. Другие ловят ногохвосток, других клещей, прокалывают их хелицерами и высасывают. Третьи высасывают яйца ногохвосток и т. д.
Замечательно строение наружного полового аппарата — сложного образования, занимающего большую часть нижней поверхности брюшка и сходно устроенного у обоих полов. Как теперь выясняется, в его состав входят конечности трех передних сегментов брюшка. Половое отверстие открывается на конце крупного выпячивающегося полового конуса на восьмом сегменте. Ножки восьмого, девятого и десятого сегментов срастаются и образуют половые крышки, прикрывающие половой конус. Коксальные органы брюшных ножек, имеющие вид выпячивающихся мешочков, наполняемых гемолимфой, в числе трех пар расположены под крышками. Они называются у клещей половыми присосками. Это органы чувств, играющие определенную роль при сперматофорном оплодотворении и откладке яиц, по-видимому, воспринимающие влажность. Благодаря микроскопическим размерам низших клещей их сперматофоры и особенно яйца, будучи малы абсолютно, очень велики относительно взрослой особи, с чем и связана громоздкость и сложность наружного полового аппарата. Яйца созревают и откладываются по одному, так как в теле самки может уместиться обычно только одно готовое к откладке яйцо.
Примитивные Acariformes лишены специальных органов дыхания и дышат кожей. В сочетании с тем, что только что говорилось о судьбе брюшных ножек, это приводит нас к выводу, что предки этого отряда клещей никогда не имели легких: из воды они прямо перешли к жизни во влажной почве и кожному дыханию, в чем сходны с кенениями. В дальнейшем у акариформных клещей образовались трахеи, причем в разных группах независимо, в разных местах. Примитивные черты обнаруживаются во внутреннем строении низших форм, например, имеется три пары выделительных коксальных желез, функционирующих у взрослых клещей, сохраняется сегментация кишечника и т. п.
Индивидуальное развитие низших Acariformes представляет большой интерес. Оплодотворение сперматофорное. Самец оставляет на субстрате клейкие сперматофоры на ножке, а самка захватывает их наружными половыми органами. Но у некоторых видов даже нет еще сперматофора, а просто выделяются капли семенной жидкости, которые самка, идя за самцом, всасывает половым конусом. Яйца очень малы (0,1—0,2 мм) и бедны желтком. Благодаря этому индивидуальное развитие сохранило архаичные черты, утраченные другими арахнидами. Дробление яйца полное, равномерное, по некоторым признакам напоминает дробление яйца кольчатых червей. Зародыш проходит четырех сегментную стадию (рис. 55, А), сходную с личинкой трилобитов — протасписом (см. В описании типа Arthropoda).
Клещи этого отряда выходят из яйца на значительно более ранней стадии развития, чем все остальные арахниды. Образование кутикулы и подготовка к вылуплению начинается в то время, когда у зародыша еще не развита четвертая пара ног, нет зачатков брюшных ножек и недостает трех концевых сегментов брюшка. Поэтому вылупляется шестиногая личинка с коротким брюшком, лишенным трех последних сегментов и половых створок. Первая кутикула, одевающая зародыш, снабжена шипиками или упругими щетинками («яйцевыми зубами»), которые служат для разрыва яйцевых оболочек. Это так называемая предличинка. У большинства акариформных клещей под этой первой кутикулой в яйце вскоре образуется кутикула собственно личинки, которая, вылупляясь, сбрасывает яйцевые оболочки и предличиночный покров. Но у низших представителей отряда предличинка вылупляется сама, хотя обычно бывает неподвижна. Недавно автору этого раздела удалось найти вид низших клещей, у которых предличинка подвижна и хорошо бегает, так что по существу сохранилось два активных личиночных возраста, разделенных линькой.
Личинка (рис. 55, В) превращается в нимфу, которая, трижды линяя, становится взрослым клещом. Соответственно различают три нимфальных возраста: протонимфу, дейтонимфу и тритонимфу. В течение нимфального периода появляются сегменты брюшка и конечности, недостававшие у личинки (рис. 55, Г—Е). Замечательно, что новые сегменты образуются один за другим вперед от анальной зоны роста и одновременно на лежащих впереди сегментах появляются конечности. Иначе говоря, здесь частично сохранился анаморфоз, т. е. тот первичный способ развития, который мы видели у предков хелицеровых — трилобитов. При превращении личинки в протонимфу появляются ноги четвертой пары, еще не вполне развитые и почти лишенные щетинок; на конце брюшка, перед створками, прикрывающими анальное отверстие, образуется новый, одиннадцатый по общему счету сегмент; на восьмом сегменте появляются видоизмененные ножки — зачатки половых клапанов с парой коксальных органов под ними. У дейто- и тритонимфы последовательно появляются двенадцатый и тринадцатый сегменты брюшка, а на девятом и десятом сегментах — видоизмененные ножки. Последние одни за другими прирастают сзади к ранее появившимся, образуя половые крышки с тремя парами коксальных органов под ними. При последней линьке и превращении тритонимфы во взрослого клеща новых сегментов и конечностей уже не появляется, а образуется только половой конус.
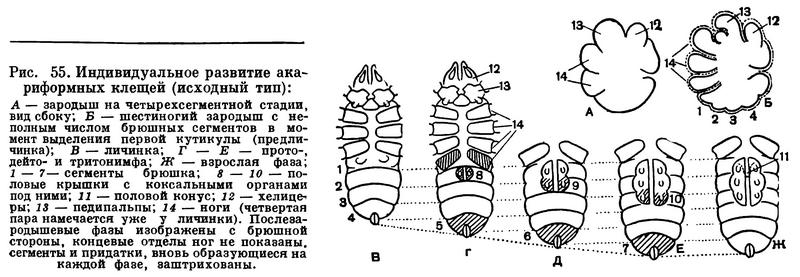
Жизненная форма низших Acariformes очень своеобразна и не меняется от личинки до взрослого клеща. Всю жизнь эти клещи обитают во влажной почве, в перегнивших растительных остатках, в пространствах между частичками, и не выработали каких-либо специальных расселительных фаз, свойственных другим клещам. Они чрезвычайно влаголюбивы и могут жить только в воздухе, насыщенном водяными парами. Извлеченные из почвы, они быстро высыхают и гибнут. Но не менее опасна и капельная влага. Если клещ прилипнет к поверхностной пленке, он намокает и гибнет. Условия жизни этих крошечных дышащих кожей существ лежат на грани воды и пара. С одной стороны, угроза высохнуть, с другой — безнадежно прилипнуть к воде и размокнуть. Эти клещи так малы, что силы поверхностного натяжения воды для них резко преобладают над земным тяготением. Их стихия — силы молекулярного сцепления. Отсюда понятны и некоторые приспособления — несмачивающиеся покровы и прыгательные задние ноги у целого ряда форм. Последнее требует пояснений. Наблюдая этих клещей под микроскопом на поверхности воды, можно видеть, как они беспомощно перебирают ногами, скользя и не двигаясь с места. А прорвать коготками поверхностную пленку опасно: мениски намертво схватят за ноги, и тогда конец. Но вот удар задними ногами — и клещ легко отрывается, перескакивая на новое место.
Заметим, что, вероятно, в том же состоит истинный смысл прыгательного аппарата низших бескрылых насекомых — отряда ногохвосток (Collembola), которые обитают совместно с низшими клещами и представляют ту же жизненную форму мелкого тонкокожего несмачиваемого прыгающего жителя влажных пористых субстратов. Интересно, что некоторые виды тех и других выходят из почвы на поверхность стоячих водоемов. Низшие акариформные клещи и ногохвостки живут бок о бок с незапамятных времен, они найдены вместе в девонских отложениях, которым 300 миллионов лет, и, вероятно, были одними из первых членистоногих, заселивших почву на нашей планете.
Таково исходное состояние отряда Acariformes. Выше мы говорили о роли малых размеров в эволюции клещей. В судьбе данного отряда роль малых размеров особенно отчетлива и двояка. С одной стороны, благодаря малым размерам эти древнейшие клещи остановились на более низком эволюционном уровне, чем другие арахниды. Обогащение яиц желтком, столь характерное для хелицеровых вообще, здесь затормозилось малой величиной самки. В результате сохранился архаичный способ развития (шестиногая личинка, анаморфоз) и примитивные черты строения (полнота сегментации и др.), о чем свидетельствуют низшие представители отряда.
Но, с другой стороны, акариформные клещи в течение огромного периода времени их жизни на суше максимально использовали приспособительные возможности, которые открылись перед ними благодаря малым размерам. Они в изобилии заселили почву и всевозможные пористые органические субстраты, из почвы устремились на растения и животных в качестве паразитов, проникли даже в водоемы. При этом замечательно, что архаичный метаморфоз, о котором мы только что говорили, явился той основой, на которой возникли приспособления к новым условиям. По мере необходимости отдельные фазы метаморфоза брали на себя те или иные жизненные функции: одни стали питательными, другие расселительными или переживающими неблагоприятную обстановку; даже размножение, в зависимости от условий жизни, оказалось возможным и целесообразным на разных фазах индивидуального развития. Благодаря малым размерам взрослого клеща и отсутствию необходимости длительного роста возникли чрезвычайно быстро размножающиеся формы — тот скоростной тип развития, о котором мы упоминали.
Таким образом, огромное разнообразие акариформных клещей возникло на основе различных приспособительных изменений их жизненного цикла, причем эти приспособления складывались на более низком эволюционном уровне, чем тот, на котором находятся другие арахниды. На столбовой дороге эволюции, по которой прошли хелицеровые животные, акариформные клещи раньше других остановились и рассыпались по боковым тропкам.
Поскольку в основе эволюции клещей лежат приспособительные изменения индивидуального развития, мы составили сравнительную схему жизненных циклов для данного отряда в целом, к которой нам постоянно придется обращаться по ходу дальнейшего изложения материала (рис. 56). Следует сразу же отметить, что при сравнительном рассмотрении жизненных циклов клещей, помимо специализации отдельных: фаз развития, усложнения метаморфоза, а также живорождения, мы столкнемся с такими несколько непривычными явлениями, как уподобление взрослой фазы молодым (педоморфоз), сдвиг половой зрелости на ранние фазы цикла и отпадение поздних (неотения), выпадение промежуточных фаз, сверхраннее вылупление из яйца (дезэмбрионизация) и размножение в полуэмбриональном состоянии и т. п.
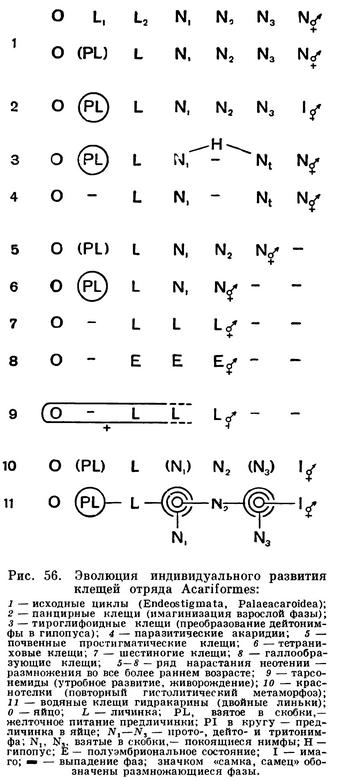
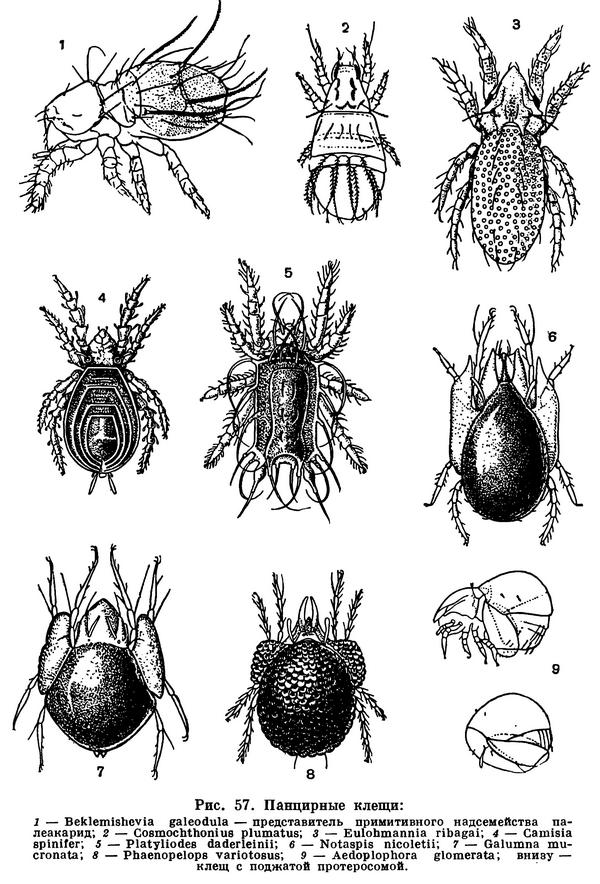
ПОДОТРЯД САРКОПТИФОРМНЫЕ КЛЕЩИ (SARCOPTIFORMES)
Представителей этого подотряда можно назвать грызущими или жующими акариформными клещами, так как большинство их обладает клешневидными хелицерами с крепкими клешнями жующего типа. Питаются они главным образом твердой растительной пищей, а переходя к паразитизму на позвоночных животных, питаются кожей, пером, волосом, выделениями кожных желез. В этот подотряд входят две большие серии семейств — панцирные клещи, или орибатиды (Oribatei), и акаридии (Acaridiae). В пределах каждой серии семейства объединяются в ряд надсемейств. Серии отражают два главных направления эволюции жизненного цикла в этом подотряде. У орибатид взрослая фаза развития становится панцирной и, помимо размножения, выполняет функцию расселения и переживания неблагоприятных условий. У свободноживущих акаридий такой расселительной и переживающей фазой становится дейтонимфа — так называемый гипопус, свойственный относящимся сюда тироглифоидным клещам (надсем. Tyroglyphoidea). При переходе акаридий к паразитизму переживающая дейтонимфа обычно выпадает. Таковы перьевые клещи (надсем. Analgesoidea), чесоточные клещи (надсем. Sarcoptoidea) и ряд других.
Панцирные клещи (Oribatei) — это самая обширная группа среди почвенных клещей, включающая, по разным авторам, от 40 до 95 семейств. Главная особенность орибатид, благодаря которой они распространились повсюду и достигли громадного видового разнообразия, состоит в том, что у взрослой фазы образовались твердый панцирь и трахеи, взрослый клещ стал устойчивым к внешним воздействиям, долговечным и способным активно переселяться. Предшествующая часть жизненного цикла сохранилась при этом почти без изменений (рис. 56, 2). Личинка и нимфы лишены панциря и трахей и вынуждены жить во влажном воздухе в толще субстрата.
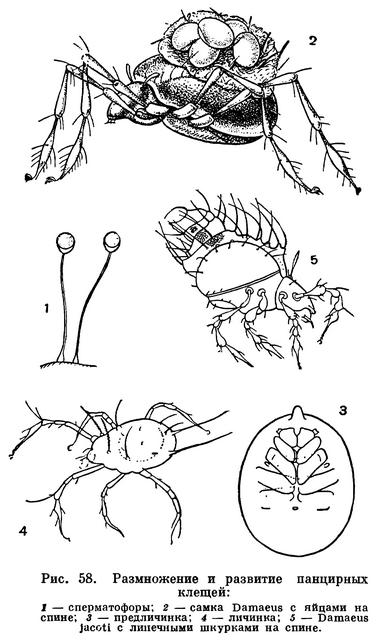
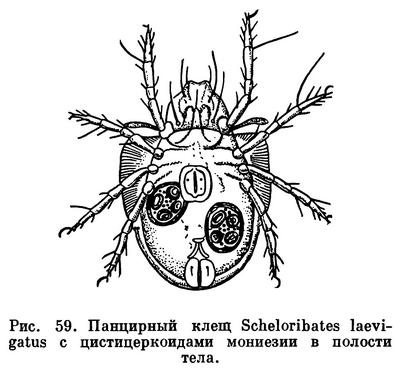
Тироглифоидние клещи (надсем. Tyroglyphoidea) называются также амбарными, так как среди них немало форм, повреждающих зерно в хранилищах, муку и другие пищевые продукты. Тироглифиды по количеству видов уступают орибатидам, но численность их в природе и в запасах продуктов бывает огромна. Заслуга всестороннего изучения тироглифид и разработки мер защиты пищевых запасов принадлежит коллективу советских ученых во главе с профессорами Московского университета З. С. Родионовым и А. А. Захваткиным. Последним написана монография «Тироглифоидные клещи» («Фауна СССР», 1941).
Места обитания тироглифид очень разнообразны: почва, лесная подстилка, скопления всевозможных растительных остатков, гниющая древесина, вытекающий сок деревьев, грибы, лишайники, мхи, корни, клубни и зеленые части высших растений, гнезда млекопитающих, птиц, насекомых и т. п. Характерный набор видов заселяет зерно в элеваторах и складах, куда клещи заносятся с полей при уборке урожая. При влажности зерна около 17% клещи выедают зародыш, к которому проникают через повреждения оболочки, а при более высокой влажности начинается их массовое размножение в зерне, причем они питаются и эндоспермом. Помимо непосредственного повреждения, клещи снижают всхожесть зерна, засоряют его экскрементами и шкурками, приводят к склеиванию в комки и самосогреванию, заражают гнилостными микроорганизмами.
Наиболее серьезные вредители зерна — мучной клещ (Tyroglyphus farinae) и удлиненные клещи (Tyrophagus noxius, Tyrophagus perniciosus). Муке сильно вредит Aleuroglyphus ovatus.
Защита зерна от амбарных клещей включает ряд мероприятий. Расселительные способности тироглифид не позволяют полностью предотвратить их попадание в склады. Поэтому особое значение приобретает режим хранения, исключающий размножение клещей: охлаждение зерна и поддержание низкой его влажности, устранение факторов, вызывающих самоувлажнение и самосогревание, доступ сырого воздуха и т. п. Из химических мер борьбы применяют обработку зерна газообразным хлорпикрином и порошкообразными ядами (акарицидами), но необходимость этих «пожарных» мер обычно бывает результатом плохих условий хранения запасов.
Тироглифиды вредят не только зерну, и мучным продуктам. Сырный клещ (Tyrolichus casei) размножается на сырах; клещ Carpoglyphus lactis живет на веществах, содержащих молочную, уксусную и янтарную кислоты, — сухих фруктах, кислом молоке, старом сыре, пиве, гнилых овощах и т. п. Интересен винный клещ (Histiogaster bacchus), который иногда в массе размножается на поверхности вина в чанах разливочных цехов винных заводов и портит вино. Виды этого рода живут в природе на вытекающем бродящем соке деревьев, особенно дуба. Как выяснилось, эти клещи попадают на винные заводы с дубовыми бочками, в которых транспортируется вино. Луковичный клещ (Rhizoglyphus echinopus) живет на корнях растений и сильно повреждает луковицы, корне- и клубнеплоды в хранилищах.
Паразитические акаридии представлены несколькими надсемействами: перьевые клещи птиц (надсем. Analgesoidea), паразиты млекопитающих — волосяные клещи (надсем. Listrophoroidea) и чесоточные клещи (надсем. Sarcoptoidea), некоторые паразиты насекомых и др. Все они по происхождению связаны с тироглифоидными клещами и перешли к паразитизму через гнездовое сожительство с животными-хозяевами. Это постоянные наружные или внутрикожные паразиты с грызущими ротовыми органами, питающиеся не кровью, а пером, волосом, кожей, выделениями кожных желез и т. п. Переход к постоянному паразитизму сопровождался выработкой ряда приспособлений. Расселительная дейтонимфа (гипопус) исчезла за ненадобностью. У наружных паразитов (эктопаразитов), подвергающихся механическому воздействию хозяина — счесыванию и раздавливанию, возникли защитные и прицепные приспособления: уплощенное тело, покрытое щитками, крепкие ноги с крупными присосками на лапках, выросты и разнообразные прицепные щетинки, прикрепительный аппарат для захвата волоса у волосяных клещей и т. п.
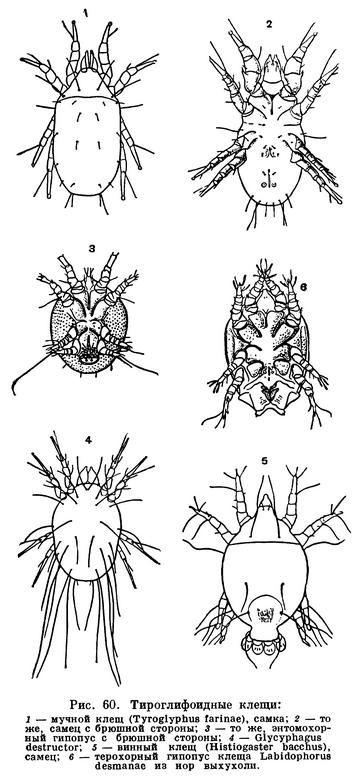

Наиболее разнообразны перьевые клещи (надсем. Analgesoidea), которых только на птицах Палеарктики обнаружено около 1000 видов. Эти высокоспециализированные паразиты живут на бородках перьев и на коже птиц. На одном виде птиц обычно встречается несколько специфических видов перьевых клещей, причем каждый обитает на разных участках оперения, нередко на определенных перьях, с чем в свою очередь связана различная форма тела клещей и строение прицепных приспособлений. Перьевые клещи питаются отмершими частичками эпидермиса кожи и перьев, но главным образом жировой смазкой оперения птиц, выделяемой копчиковой железой. Самец спаривается с телеонимфой, у которой имеется копулятивный канал и которая после этого линяет, превращаясь в оплодотворенную самку. Яйца приклеиваются к бородкам пера. В большинстве случаев, кроме шестиногой личинки, имеется две нимфальные фазы — прото- и телеонимфа, но у некоторых видов бывает еще покоящаяся дейтонимфа, которая соответствует гипопусу и появляется при неблагоприятных условиях, например в период линьки птицы-хозяина. Течение жизненного цикла и все поведение перьевых клещей подчинены жизненному ритму птиц-хозяев и закономерно изменяются в зависимости от возраста птицы, наступления и характера линьки оперения, осенних перелетов и т. д.
Большинство перьевых клещей не причиняет птицам заметного вреда, но есть виды, вызывающие тяжелые поражения кожи. Таков ножной зудень (Knemidocoptes mutans), прежде причислявшийся к чесоточным клещам, который живет под чешуями неоперенной части ног кур и других домашних птиц и вызывает тяжелое заболевание «известковые ноги». При этом заболевании чешуи оттопыриваются, ноги покрываются белесыми бугристыми корками, под которыми происходит отмирание (некроз) тканей, и птица гибнет.
Чесоточные клещи (надсем. Sarcoptoidea) представлены немногими видами, паразитами млекопитающих животных и человека, возбудителями различных форм зудневой чесотки. У человека чесотку вызывает чесоточный зудень (Acarus siro). Именно этот паразит в трудах Аристотеля обозначается словом «клещ» (греческое ахара, латинское Acarus) и указывается в качестве причины чесотки.
Самки чесоточного зудня 0,3 мм длиной, тело округлое с короткими ногами; самец вдвое мельче. Покровы кожистые, бороздчатые. Самка питается кожей, прогрызая в ее роговом слое извитые ходы до 15 мм длиной, которые различимы через поверхность кожи в виде сероватых линий. Яйца откладываются в ходах, над ними самка обычно выгрызает вентиляционные отверстия. Личинки и протонимфы живут в этих ходах, где питаются остатками изгрызенной самкой кожи и тканевой жидкостью. Сами они ходов не прогрызают. Протонимфы превращаются в телеонимф, которые выходят на поверхность кожи обычно ночью, когда больной спит. Здесь часть их превращается в самцов, которые спариваются с женскими телеонимфами — будущими самками. Самцы выгрызают в коже небольшие ходы, в которые прячутся. Оплодотворенные телеонимфы вгрызаются в кожу и превращаются в самок. Заражение происходит при соприкосновении с больным чесоткой, в редких случаях через платье больного. Человека могут заражать и другие чесоточные клещи, которые паразитируют на собаках, овцах, козах, свиньях, лошадях и других животных. Чесоточные зудни родов Acarus, Psoroptes, Chorioptes сильно вредят в животноводстве.
Лечат чесотку химическими препаратами в виде мазей, растворов; животных купают в ваннах или обрабатывают газообразными препаратами в специальных камерах.
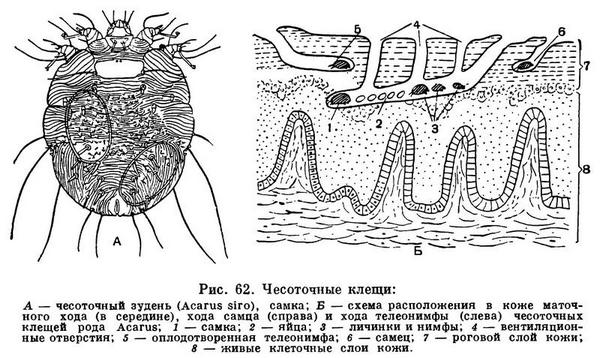
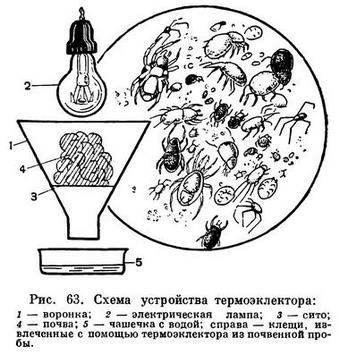
ПОДОТРЯД ТРОМБИДИФОРМНЫЕ КЛЕЩИ (TROMBIDIFORMES)
Представителей этого подотряда, в отличие от предыдущего, можно назвать сосущими акариформными клещами, так как большинство их питается жидкой животной или растительной пищей и хелицеры приспособлены для прокалывания. Характерно развитие пары трахей, открывающихся стигмами близ хелицер. Классификация подотряда сложна, насчитывается до 20 надсемейств и 70 семейств. Многочисленные ряды форм ведут от почвенных клещей к разнообразным обитателям растений, паразитам животных, водяным клещам и др. Судя по характеру жизненных циклов, все это на первый взгляд необозримое многообразие представляет два главных эволюционных направления, ведущих начало от примитивных Endeostigmata. Одно из них характеризуется сокращением жизненного цикла, поздние фазы развития уподобляются ранним (педоморфоз), число линек сокращается, и размножение происходит во все более раннем возрасте (неотения), что обусловлено монотонностью условий и скрытым образом жизни этих форм на протяжении цикла развития. Таковы многие семейства почвенных Trombidiformes, обычно объединяемые в группу Prostigmata, у которых взрослые особи отвечают тритонимфам исходного цикла, и связанные с ними по происхождению постоянные обитатели растений — тетраниховые и галлообразующие клещи, а также ряд постоянных паразитов животных. Другое направление представляют краснотелки и пресноводные клещи, у которых возник личиночный паразитизм, резкая смена образа жизни в течение цикла и связанный с этим сложный метаморфоз.
Почвенные Trombidiformes (табл. 8, 1—6) чрезвычайно разнообразны. Располагая микроскопом или лучше бинокуляром и несложным прибором для добывания клещей, можно увидеть эти формы. Клещей «выгоняют» из почвы с помощью термоэклектора. Это воронка с вложенным в нее ситом, на которое помещают почву или иной субстрат, подлежащий обработке. Над воронкой горит электрическая лампа, а под горлышко ставят чашечку с водой. По мере просушивания почвы лампой клещи и другие подвижные беспозвоночные уходят вниз, проваливаются сквозь сито и падают в чашечку. Часть их тонет, но через несколько часов поверхность воды покрывается бактериальной пленкой, по которой клещи свободно ходят. Если теперь поместить чашечку под бинокуляр, то глазам предстанет удивительный мир. Тут много уже знакомых нам орибатид и тироглифид, а также гамазоидных клещей, о которых речь ниже, но наиболее разнообразны тромбидиформные клещи. Здесь множество различных Prostigmata. По поверхности бегают крошечные беловатые клещи семейства Tydeidae, шустрые, подергивающие ногами клещи семейства Eupodidae. Вдруг среди этой мелюзги в поле зрения появляется крупный (в действительности всего 0,3 мм) лиловый клещ с острым «носом» — вытянутыми сомкнутыми хелицерами. Это бделла (сем. Bdellidae), хищница, пронзающая хелицерами, как копьем, ногохвосток и других клещей. Она очень прожорлива, ее нередко видишь с насаженной на хелицеры ногохвосткой, которую она высасывает на ходу. Вот другая хищница — рагидия (сем. Rhagidiidae), похожая на сольпугу в миниатюре, очень подвижная, нежная, но со здоровенными клешнями хелицер, жительница глубоких почвенных микропещер. Вот шествует бархатисто-красная краснотелка (сем. Trombidiidae), кажущаяся гигантом в сравнении с остальными. А вот удивительное нежнейшее существо с непомерно длинными ногами, подобие миниатюрного сенокосца — клещ Linopodes, живущий на грибах. Поверхность воды усеяна клещами-тарсонемидами (Tarsonemini), похожими на мельчайших черепашек, странными клещами-криптогнатидами (сем. Cryptognathidae) с капюшоном над ротовыми органами, которые выбрасываются на длинной «шейке», и множеством других форм. Здесь столько всего, что один перечень видов занял бы несколько страниц.
Почвенные Trombidiformes дали начало множеству групп клещей, больших и малых по числу видов, из которых мы упомянем только некоторые. Своеобразны клещи-тарсонемиды (надсем. Tarsonemini), очень различные по образу жизни (рис. 64). Эти мельчайшие формы с сосущими ротовыми органами в виде «головки» и черепитчатыми щитками частью живут в почве, частью перешли на растения, насекомых и позвоночных в качестве паразитов. Размножение на ранних фазах (неотения) сочетается у них с утробным развитием молоди, вплоть до рождения взрослых клещей, иногда личинкоподобных. Злаковый клещ (Siteroptes graminum) повреждает злаки и переносит споры ржавчины. Пчелиный клещ (Acarapis woodi) паразитирует в трахеях пчел и сильно вредит в пчеловодстве. Клещи рода Pygmephorus живут в шерсти грызунов. Пузатый клещ (Руemotes ventricosus) паразитирует на насекомых и может нападать на человека, вызывая уколами сильное раздражение кожи. Самка рождает взрослых клещей. В период размножения ее брюшко сильно вздуто и на нем держатся крошечные самцы, которые сосут гемолимфу матери и оплодотворяют рождаемых ею самок.
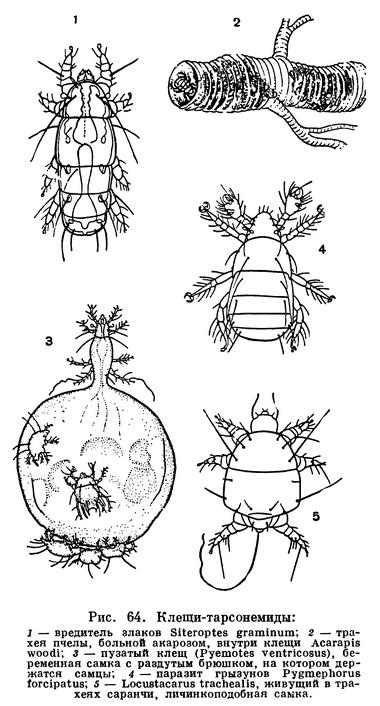
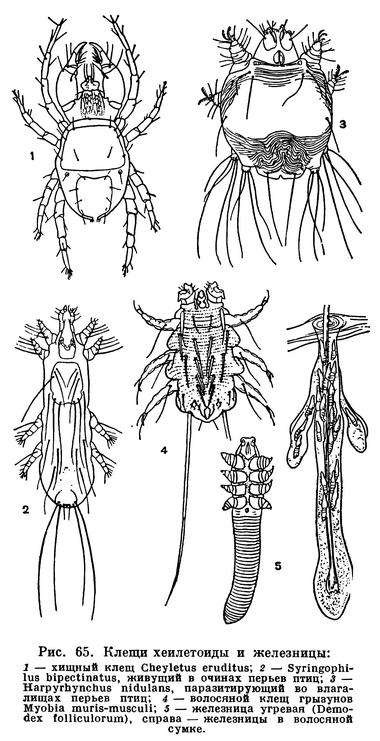
Другой пример перехода к паразитизму представляют простигматические клещи надсемейства Cheyletoidea (рис. 65). Хищные клещи (сем. Cheyletidae), с крупными хватательными педипальпами, живут в почве, растительных остатках, гнездах насекомых и позвоночных, нередко переходя на животных в погоне за клещами-паразитами. С этим семейством связано по происхождению несколько семейств своеобразных паразитов позвоночных. Клещи семейств Syringophilidae и Harpyrhynchidae живут в полостях очинов перьев, на коже и в подкожной клетчатке птиц. Волосяные клещи (сем. Myobiidae) паразитируют на мелких млекопитающих. Они держатся на волосе и высасывают содержимое волосяного фолликула, лимфу и плазму крови. От миобиид ведут начало железницы (сем. Demodicidae), которые живут в сальных железах и волосяных сумках млекопитающих и благодаря эндопаразитизму изменились до неузнаваемости. Они так малы, что их можно рассмотреть только при сильном увеличении микроскопа. Тело червеобразное, кутикула кольчатая, ноги короткие, как бы обрубленные, ротовые органы сосущие; они кладут яйца; личинки, нимфы и взрослые по облику сходны. Виды рода Demodex — паразиты разных млекопитающих, диких и домашних, некоторые сильно вредят в животноводстве, вызывая железничную чесотку (демодикоз). У человека это заболевание вызывает железница угревая (Demodex folliculorum), паразитирующая в сальных железах.
Сосущие растительноядные клещи также связаны по происхождению с почвенными Prostigmata. Уже среди последних есть формы, высасывающие почвенные водоросли, соки мхов, но наиболее разнообразны заселившие высшие растения тетраниховые и галлообразующие клещи.
Тетраниховые клещи (надсем. Tetranychoidea) представлены несколькими семействами, среди которых наиболее распространены паутинные клещи (сем. Tetranychidae) и плоскотелки (сем. Tenuipalpidae, рис. 66, 67, табл. 9). Тело тетранихид овальное, цельное, у плоскотелок уплощено и разделено на протеро- и гистеросому. Это мелкие клещи 0,2—1 мм длиной. Кутикула тонкая, у плоскотелок уплотненная, зернистая, сетчатая. Характерна окраска различных оттенков зеленого, желтого, красного цвета. Щетинки обычно игольчатые, но бывают листовидные, перистые и др. Ротовые органы приспособлены для прокола эпидермиса и сосания сока. Основные членики хелицер сросшиеся, в них помещаются два выдвижных длинных стилета — измененные концевые членики хелицер. Внутри коротких, сросшихся в основании педипальп у паутинных клещей помещаются паутинные железы, протоки которых открываются на концевом членике. Близ хелицер открываются стигмы трахей. Ноги тонкие, на лапках имеется сложный коготковый аппарат, с помощью которого клещи удерживаются на листьях: средний коготок (эмподий), иногда с придатками, и пара боковых, с двумя булавчатыми волосками каждый. Наружный половой аппарат упрощен, нет ни клапанов, ни половых присосок. У самок половое отверстие прикрыто кожной складкой, у самцов имеется копулятивный орган на заднем конце тела.
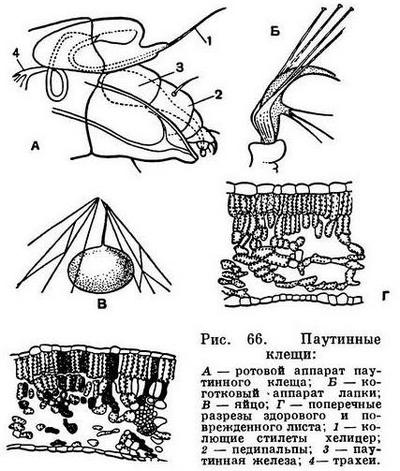
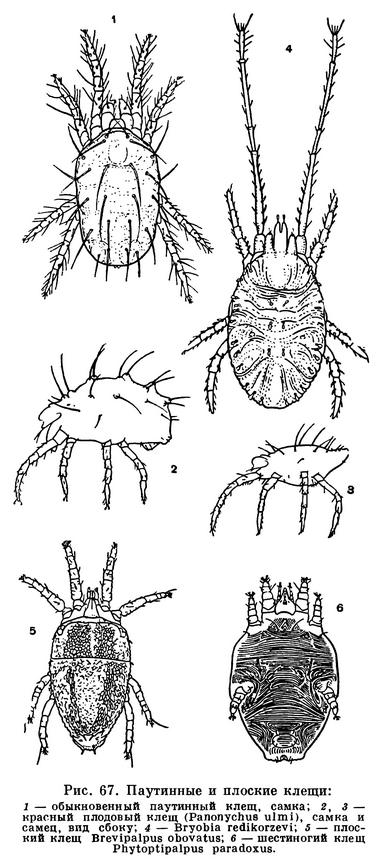
Круг кормовых растений тетраниховых клещей в целом широк. Паутинные клещи живут главным образом на лиственных деревьях, но немало видов обитает на травянистых растениях, в частности клещи-бриобии (подсем. Bryobiinae) в основном живут на травах. На хвойных встречаются немногие виды. Клещи-плоскотелки также в основном обитатели листопадных древесных растений, но немало обитателей хвойных, а также злаков. Большинство видов имеет более или менее ограниченный набор кормовых растений (олигофаги), некоторые живут только на одном виде растений (монофаги), но есть и такие, которые могут жить и размножаться на сотнях видов как древесных, так и травянистых растений (полифаги).
Большинство видов семейства Tetranychidae в том или ином количестве выделяет паутину, которой покрываются листья и другие части растений. Паутинный покров служит защитой клещам и особенно характерен для видов, живущих большими колониями. На паутинках клещи также расселяются, подхваченные токами воздуха.
Наиболее вредоносен обыкновенный паутинный клещ (табл. 9), которому разные специалисты до сих пор придают разные латинские названия из ранее употреблявшихся: Tetranychus telarius, Tetranychus urticae, Tetranychus turkestani и др. Этот вид очень широко распространен, в СССР наиболее обилен в южных районах, обычен в средней полосе, где в годы с жарким летом нередко размножается в массе, в оранжереях и на комнатных растениях встречается вплоть до Заполярья. Крайне многояден, известно несколько сотен видов кормовых растений, травянистых и древесных, кроме хвойных. Питающиеся клещи зеленые и желтоватые, живут колониями на нижней поверхности листьев, покрывая ее слоем паутины. Самка в течение двух-трех недель жизни при благоприятных условиях откладывает свыше 100 яиц. Длительность генерации — две-три недели, поколения накладываются во времени. В размножающейся колонии клещей можно найти все фазы — яйца, личинок, нимф, самок и самцов. Зимует оплодотворенная самка в состоянии диапаузы, которая наступает под влиянием сокращения длины светового дня. Во второй половине лета, когда условия еще подходящие для размножения клещей, в колониях начинают появляться готовящиеся к зимовке самки. Они перестают питаться, приобретают оранжево-красный цвет и уходят с растения, выделяя по пути много паутины. Зимуют они в растительных остатках, под комочками почвы, собираясь в плотные скопления. Нередко самки еще на растении собираются в клубочки, которые свешиваются на паутинных тяжах. В южных районах и в оранжереях клещи размножаются непрерывно, давая в год до 20 поколений. Потенциальная размножаемость колоссальна. Приводят несложный подсчет: если бы в потомстве каждой самки выживало 50 особей, то при 15 поколениях в год от одной исходной самки получилось 6*1035 особей, которые покрыли бы земной шар сплошным слоем в 2,5 м. Поэтому вспышки массового размножения возникают внезапно при очень низкой первоначальной численности клещей.
Обыкновенный паутинный клещ на юге сильно вредит хлопчатнику, плодовым, бахчевым культурам, в более северных районах — оранжерейным и парниковым растениям, в особенности огурцам. Значителен экономический ущерб в хлопководстве. Листья хлопчатника покрываются багрово-красными сливающимися пятнами и опадают. Отмирают бутоны, урожай коробочек снижается на 30—50%. Характерную картину повреждения хлопчатника в Средней Азии называют «ургамчак», «канасы», в Закавказье — «чор».
Яблоням, сливам и другим плодовым вредит красный плодовый клещ (Panonychus ulmi), до 0,45 мм, темно-красный, спинные щетинки крупные, сидят на бугорках. Этот вид распространен в южной половине Европейской части СССР, в Крыму, на Кавказе. Зимуют яйца в трещинах коры, личинки появляются с распусканием листьев, на юге бывает несколько поколений в сезон. Помимо плодовых, этот клещ живет на различных лиственных породах, в особенности из розоцветных.
Галлообразующие, или четырехногие, клещи (сем. Eriophyidae, надсем. Eriophyoidea, или Tetrapodili) — крайне специализированные паразиты растений (рис. 68). Эти мельчайшие существа (0,1—0,2 мм) высасывают содержимое растительных клеток и вызывают у растений появление тех или иных уродливых образований, наростов, уродливости роста побегов, цветков и плодов, ржавых пятен, волосистых подушек (эринеумов), разнообразных галлов (табл. 9). Часть видов живет на поверхности растений (подсем. Phyllocoptinae), большинство же — в галлах (подсем. Eriophyinae и Phytoptinae).
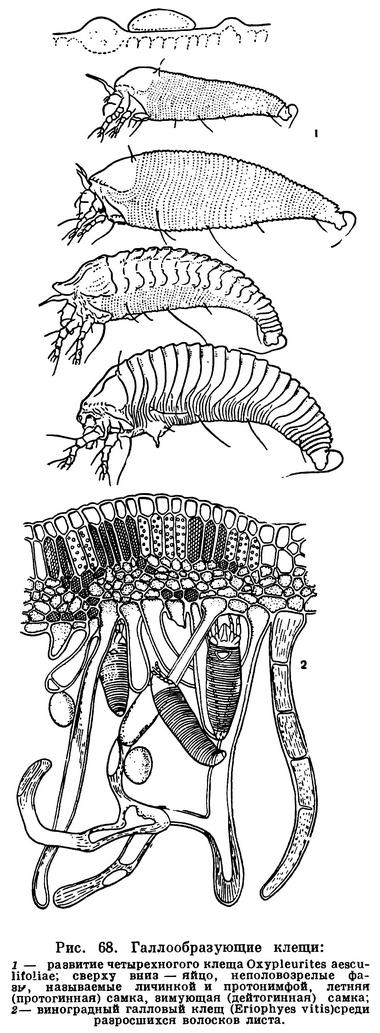
По происхождению эриофииды связаны с тетраниховыми клещами. Связующим звеном служат похожие на клещей-плоскотелок шестиногие клещи (сем. Phytoptipalpidae), живущие на растениях и размножающиеся в личиночной фазе. Эволюционный ряд от почвенных Prostigmata к тетранихидам, шестиногим и галловым клещам иллюстрируют рисунки 67, 68. Обращает на себя внимание внешнее сходство галловых клещей с железницами. Обе группы — конечные этапы паразитической специализации разных ветвей Prostigmata, одни эндопаразиты животных, другие — растений.
Жизненные циклы эриофиид строго согласованы с сезонными явлениями в жизни растений и в деталях разнообразны. Для ряда видов характерно образование двух типов самок — летних (протогинных), которые живут и размножаются в галлах, и зимних (дейтогинных), которые появляются к осени, уходят из галлов и зимуют в почках, трещинах коры, стерне и т. п. Весной они переходят на зеленые части растений и служат основательницами колоний клещей во вновь образуемых галлах.
Многие виды галловых клещей вредят культурным растениям. Виноградный войлочный клещ (Eriophyes vitis) вызывает на нижней стороне листьев винограда волосистые пятна, сначала белые, затем буреющие. Вред ощутим в засушливые периоды. Грушевый клещ (Eriophyes piri) вызывает образование бляшковидных выпуклостей на листьях груши, яблони и других плодовых. Смородинный почковый клещ Eriophyes ribis) вызывает вздутие почек смородины. Клещ Aceria tulipae живет под чешуйками луковиц лука, чеснока, тюльпана, вызывая сморщивание и высыхание луковиц. Галловые клещи переносят вирусы растений; сейчас их интенсивно изучают в этом отношении.
Клещи-краснотелки (надсем. Trombea) — формы со сложным метаморфозом, который возник в результате того, что одни фазы развития взяли на себя функцию питания, а другие за этот счет перестали питаться и превратились в формообразовательные покоящиеся фазы, подобные куколке насекомых (рис. 69). Но в отличие от последних, у которых куколка одна, у краснотелок фазы питания и формообразования чередуются (рис. 56, 10). Дейтонимфы и взрослые краснотелки — хищники, высасывают мелких членистоногих, нередко их яйца, и также весьма прожорливы. Избыточное питание активных фаз, в первую очередь личиночный паразитизм, было той основой, на которой возник метаморфоз краснотелок. Личинки перешли к паразитизму от питания мелкими членистоногими (энтомофагии), начав присасываться к более крупным насекомым (клещи сем. Trombidiidae) и к позвоночным (сем. Trombiculidae).
Ярко-красные крупные виды семейства Trombidiidae (род Trombidium и многие другие) нередко попадаются на влажной почве, а их мелкие личинки — присосавшимися к насекомым. Краснотелки семейства Trombiculidae мельче, их личинки часто в большом количестве встречаются на мелких млекопитающих и птицах, особенно на ушах грызунов. Личинки тромбикулид нападают на человека, вызывая раздражение кожи (тромбидиоз), особенно в теплых влажных районах.
Тромбикулиды передают человеку японскую речную лихорадку «цуцугамуси» — опасное заболевание, распространенное в Юго-Восточной Азии, на островах Тихого океана и недавно обнаруженное у нас на Дальнем Востоке. Болезнь вызывается особыми микроорганизмами — риккетсиями, которыми личинки заражаются от грызунов и насекомоядных. Личинка питается только раз, но риккетсии сохраняются в организме клеща в течение метаморфоза и передаются через яйцо (трансовариально) голодным личинкам следующего поколения, которые и заражают человека.
Водяные клещи представлены двумя неродственными группами: пресноводные клещи — гидрахнеллы (группа Hydrachnellae) и морские клещи — галакариды (Halacarae). Гидрахнеллы (рис. 70) обычны в пресных водоемах, особенно стоячих; их известно более 2000 видов, группируемых в 9 семейств и 3 надсемейства (по новейшей классификации Б. А. Вайнштейна) (Прежде насчитывали до 20 семейств.). Это в большинстве хорошо плавающие формы, часть видов постоянно держится на водной растительности. Взрослые обычно 1—2 мм длиной, но немало и более крупных, до 3—5 мм и даже 8 мм. Это самые крупные представители отряда. Тело цельное, обычно округлое, покровы кожистые, плотные, нередко покрыты шипиками, сетчатой скульптурой или щитками различной величины и формы.
Окраска разнообразна — ярко-красная, оранжевая, желтая, бурая, зеленая, синеватая, иногда пестрая (табл. 9). Обычно развита пара двойных глаз. Ноги снабжены плавательными волосками. Тазики ног крупные, сращены с телом в виде коксальных щитков. Ротовые органы объединены в «головку», хелицеры с когтевидным концевым члеником, когтевидный вырост на педипальпах иногда образует с их последним члеником род хватательной клешни. Близ ротовых органов открываются дыхальца хорошо развитых ветвящихся трахей. Половой конус прикрыт половыми створками, у самок выполняет роль яйцеклада, у самцов — органа, выделяющего сперматофоры; половые присоски варьируют в числе.
Гидрахнеллы родственны краснотелкам, о чем свидетельствует их строение и особенно метаморфоз. Сперматофорное оплодотворение осложнено различными способами спаривания, при котором самец переносит слизистый сперматофор с помощью третьей пары ног, подтягивает самку к сперматофору, прикрепленному к субстрату, и т. п. Гидрахнеллы кладут яйца на поверхность подводных предметов. Плодовитость самки может достигать нескольких сотен яиц. Яйца окружены набухающим секретом, нередко тысячи кладок сливаются в сплошной ослизненный слой. Предличиночная фаза проходит в яйце. Личинки гидрахнелл — паразиты, главным образом водных насекомых, по облику напоминают личинок краснотелок. Они либо отыскивают насекомых, плавая в толще воды, либо всплывают и подстерегают насекомых, плавающих по поверхности и вылетающих из воды. Насекомые- хозяева разнообразны: водные жуки, клопы, стрекозы, поденки, веснянки, ручейники, различные двукрылые. Клещи рода Unionicola не только в личиночной фазе, но и во всех последующих паразитируют в мантийной полости и на жабрах пластинчатожаберных моллюсков — беззубок и др. Личинки присасываются к насекомым надолго, как бы срастаясь ротовыми органами с покровом, сосут гемолимфу и постепенно очень сильно увеличиваются, по весу в сотни раз. Паразитизм, помимо питания, имеет расселительное значение, личинки заносятся насекомыми в новые водоемы и отпадают в воду. Дальнейшее развитие представляет гистолитический метаморфоз, как у краснотелок. Однако покоящиеся фазы — прото- и тритонимфа протекают здесь под покровом предшествующих. Линечные шкурки сбрасываются по две. Под покровом личинки образуется кутикула протонимфы (аподерма), под ней кутикула дейтонимфы, и уже затем шкурки личинки и протонимфы сразу сбрасываются (рис. 56, 11). То же происходит при превращении дейтонимфы во взрослого клеща. Поэтому внешне линек только две. В периоды метаморфоза клещ прицепляется к подводному предмету и переходит в покоящееся состояние — окукливается.
Дейтонимфы и взрослые клещи — хищники, питаются мелкими рачками, личинками водных насекомых. Взрослую фазу гидрахнелл нередко называют имагинальной. Действительно, плотный покров, развитые трахеи, укрупнение размеров, сложные формы спаривания, долговечность (многие виды живут более года), способность активно расселяться — все это признаки имаго, сходные с теми, которые мы видели у орибатид, с тем различием, что здесь имаго водное и способ его формирования иной. Имагинальность гидрахнелл выступает особенно отчетливо, если сравнить взрослую фазу этих клещей с тем, что мы видели у примитивных Acariformes и в особенности у неотенических представителей отряда.
В водоемах разных типов видовой состав гидрахнелл различен. Наиболее обильны и разнообразны они среди зарослей водной растительности в прудах и озерах, где обычны многие виды родов Eylais, Hydrachna, Piona, Arrhenurus и др. Беднее фауна рек, ручьев, временных водоемов. Специфичные виды найдены в подземных водах и горячих источниках.
Галакариды (рис. 71) в большинстве морские формы, но есть и пресноводные. По происхождению они связаны с Prostigmata. Это мелкие клещи (0,2—1,7 мм). На спине обычно четыре щитка, на брюшной стороне — два и по бокам сросшиеся пластинчатые тазики ног. Ноги не приспособлены для плавания, плоско расставлены, клещи держатся на поверхности подводных предметов. Ротовые органы в виде «головки», хелицеры колющие. Половой конус самки превращен в яйцеклад, у самца — в копулятивный орган. Яйца откладывают на подводные предметы, иногда носят на задних ногах. Развитие — как у большинства Prostigmata: яйцо, личинка, прото-, дейтонимфа, размножающаяся фаза. Галакариды высасывают сок водорослей, разлагающиеся остатки животных, мелких беспозвоночных и их яйца. Паразитизм редок. Известны паразиты морских ежей, хитонов, речного рака. Галакариды широко распространены в морях, до полярных широт, живут главным образом в прибрежной зоне до 30—40 м, особенно на участках, богатых водорослями. Известно около 200 видов семейства Halacaridae.
Данные о водяных клещах имеются в двух выпусках «Фауны СССР» (1940 и 1952), написанных И. И. Соколовым.
ОТРЯД ПАРАЗИТИФОРМНЫЕ КЛЕЩИ (PARASITIFORMES)
Этот отряд по числу видов меньше предыдущего, насчитывает около 4000, но цифра эта также далека от действительной, так как изучение ряда семейств только начато. Сюда относятся три основные группы, рассматриваемые обычно как надсемейства: гамазоидные клещи (надсем. Gamasoidea), уроподы (надсем. Uropodoidea) и иксодовые клещи (надсем. Ixodoidea). Первые два надсемейства велики и разнообразны, наряду со свободноживущими формами здесь немало сожителей (комменсалов) насекомых и позвоночных, а среди гамазоидных клещей и паразитов последних. Иксодовые клещи представлены целиком кровососущими паразитами наземных позвоночных.
Выяснение происхождения этого отряда и его положения среди арахнид затрудняется тем, что даже наименее специализированные его представители значительно видоизменены, а палеонтологических материалов почти нет. Своеобразный тип строения и индивидуального развития этого отряда был выяснен автором в 1947 году при изучении наименее изменившихся почвенных гамазоидных клещей семейства Rhodacaridae. Так как среди паразитиформных клещей сегментированные формы не сохранились, пришлось прибегнуть к косвенным методам выяснения сегментального состава тела и его изменений в индивидуальном развитии. На теле этих клещей имеется сложный и постоянный набор щетинок и лировидных органов чувств. Изучение численного состава, топографии этих органов, а также порядка их появления по фазам развития показало, что они образуют поперечные ряды, соответствующие слившимся сегментам. В результате такого, как его теперь называют, хетологического анализа была получена теоретическая картина сегментации тела (в форме карты хетомерии, т. е. предполагаемых посегментных наборов щетинок), которая совпала с другими анатомическими данными (расположение мышц и др.) и, что особенно важно, с сегментацией зародыша. В дальнейшем среди низших уропод удалось найти формы, у которых частично сохранилась сегментация брюшка, также подтвердившая теоретическую картину. В результате оказалось возможным описать строение и развитие паразитиформных клещей в форме, сравнимой с другими арахнидами, причем выявилось большое своеобразие этого отряда и особенно резкие его отличия от акариформных клещей.
Туловище исходных форм (рис. 72) разделено на головогрудь и брюшко, причем граница между ними делит предполовой сегмент на часть, которая примыкает к головогруди, и часть, примыкающую к брюшку. В составе головогруди слились шесть сегментов, несущих конечности, и боковые участки головной лопасти, передняя часть которой отделена и входит в состав сложно устроенного ротового аппарата — гнатемы. В брюшке объединены шесть сегментов и два-три недоразвитых концевых.
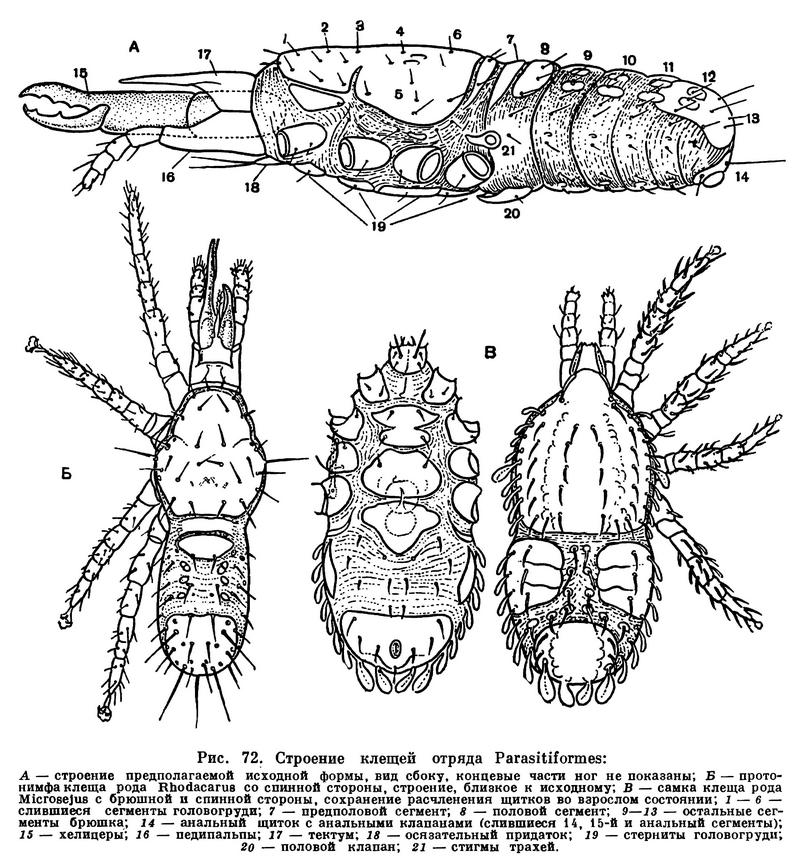
Для отряда в целом характерно образование сложного имагинального панциря, причем изучение индивидуального развития и эволюционных рядов форм позволяет понять, как он образуется. В исходном состоянии наружный скелет представлен многими посегментными склеритами. Головогрудь покрыта щитом, передний отдел которого соответствует трем передним сегментам и отделен швом. Снизу между подвижно причлененными тазиками ног расположены стерниты, сливающиеся в дальнейшем в цельную грудину. Судя по всему, ближайшие предки Parasitiformes имели членистое брюшко с небольшими парными тергитами и стернитами — местами прикрепления посегментных спинно-брюшных мышц. Слияние сегментов начинается с заднего конца брюшка, сливающиеся тергиты образуют здесь сначала небольшой пигидиальный щиток, который затем разрастается вперед, превращаясь в сплошной верхний брюшной щит (нотогастер). У многих форм последний сливается далее с головогрудным щитом в общий спинной щит и следы разделения головогруди и брюшка исчезают.
На нижней стороне брюшка у исходных форм имеется небольшой анальный щиток с анальным отверстием, прикрытым клапанами, а на восьмом сегменте — половой щиток, прикрывающий половое отверстие. В дальнейшем нижняя поверхность брюшка уплотняется двумя способами: либо анальный щиток разрастается вперед, образуя так называемый вентро-анальный щит, либо половой щиток разрастается назад, образуя генито-вентральный щит. У самцов в связи с выработкой своеобразного способа копуляции половое отверстие смещается вперед, к основанию гнатемы, и принимает вид небольшой вороночки. У самок ряда гамазоидных клещей в связи с относительно крупными размерами яиц и особенно при живорождении половой щиток увеличивается, занимает пространство между третьей и четвертой парами ног, а под ним вокруг крупной поперечной половой щели образуется сложный внутренний скелет, широко раздвигающийся при выходе молоди. У иксодид и уропод, откладывающих яйца, более мелкие относительно самки, половое отверстие невелико.
По бокам тела образуются лентовидные или округлые так называемые перитремальные склериты, на которых открываются дыхальца трахей, а нередко еще набор боковых (плейральных) склеритов.
У многих свободноживущих форм во взрослом состоянии части скелета сливаются в сплошной твердый панцирь, что характерно для ряда гамазид и особенно уропод. У паразитических клещей, насасывающих много крови, напротив, наблюдается редукция скелета и освобождение тела от щитков.
Хелицеры и педипальпы входят в состав гнатемы, или «головки», строение которой у Parasitiformes иное, чем у акариформных клещей. Тазики педипальп срастаются не только друг с другом, но и с передней частью головной лопасти (так называемым тектумом), образуя склеритное кольцо (основание гнатемы), из которого выдвигаются хелицеры и в котором помещается сосательный желобчатый конус (эпистом), ведущий в глотку. По бокам основания ротового аппарата причленяются щупальца педипальп, а снизу видоизмененные жевательные лопасти. Гнатема причленяется перепончатой шейкой и весьма подвижна как целое. Хелицеры удлиненные, 3-члениковые, втягиваются в кожистые влагалища. У многоядных и хищных форм они клешневидные, нередко очень мощные (табл. 8, 12), у кровососущих паразитов они превращаются в колющие стилеты или более сложные образования, служащие для фиксации в коже хозяина. У ряда уропод, высасывающих свою добычу или соки растений, хелицеры также длинные, стилетовидные. У самцов хелицеры обычно служат копулятивными органами и в той или иной мере видоизменены. На пальцах клешни имеются щели или жгутовидные придатки — сперматодактили, служащие для переноса сперматофора в половое отверстие самки. У некоторых гамазид эти придатки пронизаны каналами, по-видимому, наполняемыми спермой, а у самок имеются семеприемники. Способ копуляции в этих случаях точно не выяснен, но не исключено, что он сходен с тем, что мы видели у пауков, только совокупительными органами служат не педипальпы, а хелицеры.
Ноги несут на конце лапок эластичный придаток с парой коготков и присоской между ними. У гамазид ноги обычно длинные, свободноживущие формы в большинстве быстро бегающие хищники. Передние ноги тоньше других и выполняют роль усиков, на конце лапок имеется сложно устроенный тарзальный орган, в составе которого, помимо длинных осязательных щетинок, имеются органы химического чувства. Ноги уропод короткие, вкладываются в специальные ямки при замирании. У постоянных паразитов ноги нередко толстые, с мощными коготками и шиловидными щетинками, служат для прикрепления к хозяину.
Дышат паразитиформные клещи с помощью трахей, которые открываются парой стигм по бокам тела над основаниями третьей-четвертой пар ног или позади последней. Трахеи хорошо развиты, ветвятся и имеют спиральные опорные утолщения. Стигмы открываются на перитремальных щитках, чаще лентовидных. По этим щиткам вперед от стигм обычно тянется трубка — перитрема, пронизанная порами, через которую, по-видимому, происходит дыхание при закрывании стигм. У иксодовых клещей этих трубок нет и стигмы открываются на округлых или запятовидных щитках, причем их запирательный аппарат также хорошо развит.
По внутреннему строению Parasitiformes напоминают сенокосцев. Характерны несколько пар крупных слепых выростов средней кишки, мальпигиевы сосуды, сокращение коксальных желез до одной не всегда функционирующей пары, небольшое сердце, крайняя концентрация нервной системы и т. п.
Биология размножения своеобразна. О способе копуляции мы говорили. Обычно особи соприкасаются брюшными сторонами и самец с помощью хелицер вводит сперматофор в половое отверстие самки или приклеивает близ него. Большинство уропод и иксодовые клещи кладут яйца, среди гамазид наблюдаются все переходы от яйцекладущих к живородящим, причем есть формы, рождающие личинок и
нимф. В большинстве случаев яйца малы абсолютно и велики относительно самки, в связи с чем они вызревают и откладываются по одному. Но у иксодовых клещей, крупных кровососущих паразитов, все созревшие яйца находятся в половых путях самки и откладываются затем одной кладкой, обычно по нескольку тысяч.
Индивидуальное развитие чрезвычайно своеобразно (рис. 73). Выяснение сегментального состава тела и его изменений в послезародышевом развитии в сочетании с эмбриологическими данными позволяет понять наблюдаемые здесь явления. Эмбриональное развитие в этом отряде довольно однотипно, причем нет и следов тех примитивных черт, которые мы видели у акариформных клещей. Несмотря на малые размеры яиц, дробление поверхностное, и порядок сегментации зародыша вторично изменен, как у большинства арахнид. Наиболее характерно, что к концу эмбрионального развития зародыш приобретает строение, вполне типичное для арахнид вообще. Имеется не шесть, а восемь пар развитых расчлененных ног и полный состав сегментов брюшка, свойственный взрослым клещам этого отряда. Но далее у вполне развитого зародыша происходят необычайные явления. В течение нескольких часов (что удается наблюдать прижизненно, например, в прозрачных яйцах гамазоидных клещей) происходит сокращение нескольких сегментов в области соединения головогруди и брюшка. По мере роста зародыш как бы сжимается в средней части. При этом ноги четвертой пары съеживаются до небольших клеточных скоплений и уходят под покровы. В результате получается шестиногая личинка со «спрятанными» до времени ногами четвертой пары и укороченным брюшком. В таком виде она покрывается кутикулой и вскоре выходит наружу. Предличиночной фазы здесь вообще нет.
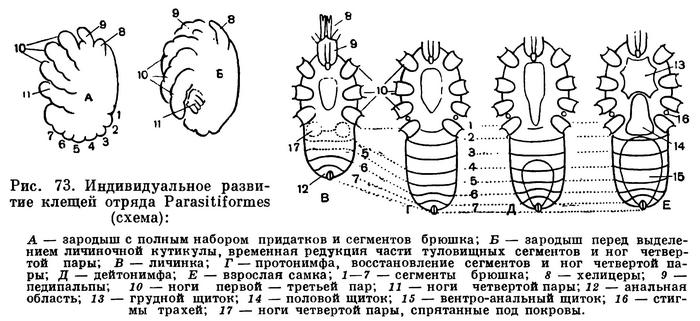
Послезародышевое развитие, помимо шестиногой личинки, включает две нимфальные фазы и взрослую. Нимф здесь называют прото- и дейтонимфой, но они ни в какой мере не соответствуют таковым акариформных клещей, и в их названии отражена лишь последовательность линек. При превращении личинки в протонимфу временно «спрятанные» сегменты и ноги четвертой пары как бы развертываются и нормальная организация клеща сразу восстанавливается. Протонимфа замечательна тем, что наиболее отчетливо олицетворяет предковое состояние этого отряда. Все, что временно извращено у личинки, у протонимфы восстановлено, но в то же время у нее еще минимальны вторичные изменения, наступающие на более поздних фазах по мере усложнения имагинального панциря. Поэтому протонимфа служит ключом, позволяющим понять организацию Parasitiformes. Расчлененный скелет, небольшой головогрудной щит и грудина, маленький пигидий на конце брюшка и посегментные склериты перед ним, стигмы еще без перитрем и т. д. — все это прототипические черты отряда (рис. 72, Б). При превращении в дейтонимфу в основных чертах оформляется вторичный наружный скелет, развиваются перитремы, посегментные наборы щетинок дополняются до окончательного состава. При превращении во взрослого клеща образуется половое отверстие, у самок — половой щиток, а у самцов — копулятивные органы хелицер. Происходит окончательное, нередко очень сильное уплотнение всего панциря.
Таковы в общих чертах особенности строения и развития этого отряда клещей. Сравнивая этот отряд с предыдущим (Acariformes), легко видеть, насколько существенны различия между ними. Их не только невозможно объединить, как это делается по традиции, но они оказываются стоящими на совсем разных эволюционных уровнях и в противоположных концах общей системы паукообразных. Сравнение Parasitiformes с другими арахнидами приводит к выводу, что они произошли от каких-то сенокосцеподобных форм, близких к некоторым ископаемым отрядам арахнид каменноугольного периода (Architarbi и др.), которые дышали легкими. Прежде Parasitiformes были более сходны с родственными им отрядами, но в дальнейшем благодаря измельчанию их строение изменилось (слияние сегментов, образование «головки» и др.), а индивидуальное развитие глубоко извратилось, внешне уподобившись развитию других клещей. Рассмотрение причин последнего явления увело бы нас в область специальных вопросов сравнительной эмбриологии. Заметим только, что, судя по картинам, наблюдаемым в эмбриональном развитии, ближайшие предки Parasitiformes не имели шестиногой личинки, из яйца выходила восьминогая форма, как у большинства арахнид (рис. 74, 1).
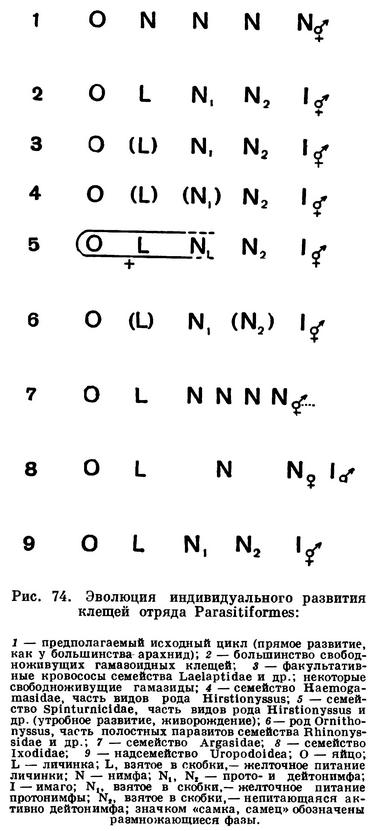
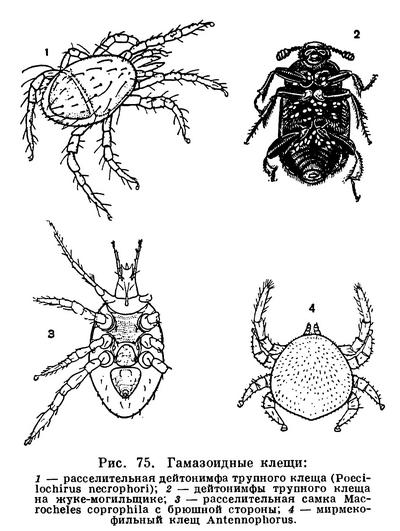
Малые размеры также открыли перед этим отрядом большие эволюционные возможности: заселение скважин почвы и всевозможных органических субстратов, переход к паразитизму. Однако характер эволюции Parasitiformes иной, чем у предыдущего отряда (Acariformes). Среди непаразитических форм господствуют хищники, разрывающие добычу хелицерами, причем хищничество сочетается с потреблением животных остатков и растительной пищи подобно тому, что мы видели у сенокосцев. Иной и характер паразитизма — переход от хищничества к сосанию крови наземных позвоночных. При этом разнообразие паразитов здесь создается не в силу разных источников питания, а благодаря различным взаимоотношениям с животными-хозяевами. В этом отряде можно различить несколько типов кровососущих паразитов, происходящих один от другого: подстерегающие временные паразиты — убежищные, т. е. нападающие на хозяина в гнезде, норе и иных укрытиях, и внеубежищные, или пастбищные, подстерегающие животных в открытой природе; постоянные паразиты — наружные, которые живут и размножаются на теле хозяина, и полостные паразиты, перешедшие к эндопаразитизму в дыхательных органах животных, и др. Как мы увидим, каждый из этих типов характеризуется целым комплексом приспособительных изменений строения, биологии и индивидуального развития, т. е. тем, что В. Н. Беклемишев в своих трудах по сравнительной паразитологии называет жизненной схемой вида. Специализация индивидуального развития идет при этом либо по линии живорождения, либо по линии увеличения или уменьшения числа нимфальных линек (рис. 74).
Питаясь кровью позвоночных, паразитиформные клещи вступили в тесные взаимоотношения с возбудителями многих болезней животных и человека — фильтрующимися вирусами, риккетсиями, бактериями, простейшими-кровепаразитами и др., и стали их специфическими переносчиками и хранителями. Наиболее важны в этом отношении иксодовые клещи, по числу переносимых болезней занимающие одно из первых мест среди кровососущих членистоногих. Они передают такие опасные заболевания, как энцефалиты, клещевые тифы, туляремию, чуму, бруцеллез, гемоспоридиальные болезни домашних животных и др. Эти клещи составляют важнейший объект медицинской и ветеринарной акарологии и паразитологии вообще. Их изучением и разработкой мер защиты человека и животных занимаются многие паразитологи, эпидемиологи и врачи во всех странах мира. Специальная литература по клещам — переносчикам болезней огромна. Советская наука по праву занимает в этой области ведущее место. Теория природной очаговости трансмиссивных болезней, т. е. передаваемых клещами и насекомыми, разработанная Е. Н. Павловским, и учение о жизненных схемах переносчиков болезней, созданное В.Н. Беклемишевым, служат основой исследовательской и практической работы в этой области и широко признаны у нас и во всем мире. Данные о кровососущих клещах изложены в целом ряде научных и учебных руководств: Е. Н. Павловский, Курс паразитологии человека, изд. АН СССР, т. I, 1945, т. II, 1949; «Учебник медицинской энтомологии», под ред. В. Н. Беклемишева, Медгиз, 1949; «Определитель членистоногих, вредящих здоровью человека», под ред. В. Н. Беклемишева, Медгиз, 1958, и др. Современный уровень знаний о кровососущих клещах хорошо отражают соответствующие главы в книге «Переносчики возбудителей природноочаговых болезней», составленной коллективом специалистов под руководством и при участии П. А. Петрищевой (Медгиз, 1962).
Гамазоидные клещи (надсем. Gamasoidea) представлены свободноживущими и паразитическими формами и составляют большую часть отряда, до 30 семейств. Непаразитические гамазиды — хищники или многоядные формы, населяют почву, лесную подстилку, скопления всевозможных гниющих остатков. Многие живут в гнездах общественных насекомых, птиц и млекопитающих. Широко распространен паразитизм на наземных позвоночных, причем наблюдаются все стадии перехода от гнездового сожительства и хищничества к питанию кровью животных и различным способам паразитирования: подстерегающие гнездовые и открытоживутцие паразиты, живущие постоянно на теле хозяев, в дыхательных путях и т. п.
Среди непаразитических гамазид (рис. 75) распространено пассивное расселение (форезия) на насекомых и других животных, что особенно характерно для видов, размножающихся в недолго существующих, эфемерных, субстратах — помете, трупах и т. п. Расселяются чаще взрослые самки, но иногда дейтонимфы.
Клещи-уроподы (надсем. Uropodoidea) — мелкие или более крупные формы (до 2 мм) с очень твердым имагинальным панцирем. Тело обычно щитообразное, выпуклое сверху и плоское снизу, овальное или грушевидное, с выступающим передним краем. Иногда панцирь покрыт сложной скульптурой (табл. 8, 14). Есть круглые в очертании совсем плоские формы, у которых край панциря образован слившимися уплощенными щетинками. Ротовые органы спрятаны в углублении панциря (камеростоме) и прикрываются снизу плоскими тазиками передних ног. Хелицеры обычно длинные, с небольшой колющей клешней. Ноги короткие, могут плотно прижиматься к телу и вкладываются в углубления на нижней стороне, что наблюдается при замирании (танатозе), очень характерном для этих медлительных клещей. Стигмы с небольшими искривленными перитремами. Половое отверстие расположено в средней части тела, у самца небольшое, зияющее, у самки прикрыто твердым щитком. Уроподы кладут яйца. У личинки и протонимфы на теле несколько щитков, которые у дейтонимфы сливаются; при превращении во взрослого клеща панцирь сильно уплотняется.
Уроподы живут в почве, лесной подстилке, навозе, много среди них мирмекофильных видов — специфических обитателей муравейников. Питание разнообразно: хищники, высасывающие почвенных нематод, личинок насекомых; формы, питающиеся органическими остатками; есть виды, сосущие сок растений и повреждающие рассаду в парниках. Некоторые имеют расселительных дейтонимф. Последние прикрепляются к насекомым, чаще жукам, с помощью секрета, выделяемого из анального отверстия и застывающего в виде гибкого стебелька (рис. 78, 2).
Уроподы изучены недостаточно. Насчитывается до 15 семейств этих клещей. Среди них наименее специализированные формы позволяют установить общность происхождения уропод и гамазоидных клещей. У представителей некоторых близких к уроподам семейств частично еще сохранилась членистость брюшка (сем. Microsejidae и др.).
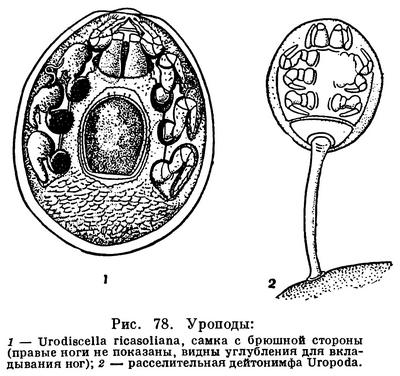

Иксодовые клещи (надсем. Ixodoidea) представлены двумя целиком паразитическими семействами: аргасовые клещи, или аргазиды (сем. Argasidae), и собственно иксодовые клещи, или иксодиды (сем. Ixodidae). Ixodoidea — высокоспециализированные подстерегающие паразиты, питающиеся кровью наземных позвоночных: аргазиды — убежищные формы, иксодиды — пастбищные. Это самые крупные клещи, сильно увеличивающиеся в размерах при насасывании крови; голодные клещи обычно 3—5 мм длиной, сытые — 10—15 мм, некоторые до 25 мм. Многие виды могут сосать кровь человека. Сюда относится большинство клещей — переносчиков возбудителей инфекционных болезней человека и животных.
Аргазиды характерны для стран с жарким сухим климатом, у нас они распространены в южных районах, главным образом в Средней Азии и Закавказье, где встречается до 17 видов трех родов. Клещи рода Ornithodoros заражают человека спирохетами клещевого возвратного тифа (рекурренса) и поэтому наиболее изучены. Полные данные о них имеются в книге М. В. Поспеловой-Штром «Орнитодорины фауны СССР» (изд. АМН СССР, 1953) и в монографии Н. А. Филипповой «Клещи-аргазиды» («Фауна СССР», 1966).
Иксодиды (сем. Ixodidae) встречаются в различных ландшафтно-климатических зонах. Это в большинстве пастбищные паразиты, подстерегающие животных-хозяев в открытой природе. При таком образе жизни вероятность встречи паразита с хозяином невелика и поддержание численности паразита обеспечивается целым рядом специальных приспособлений. Круг животных-прокормителей, как правило, довольно широк. Взрослые клещи большинства видов питаются на крупных животных — копытных, хищниках, а личинки и нимфы — на грызунах, насекомоядных, мелких хищниках, птицах, ящерицах. Взрослые клещи многих видов могут сосать кровь человека. Иксодиды способны долго голодать, обычно по нескольку месяцев, некоторые виды до двух лет. Клещи всползают на хозяина с земли или растительности. Они подстерегают животных на низкорослых растениях, сидя в характерной позе с выставленными вперед передними ногами, на лапках которых имеются обонятельные органы (органы Галлера). Прицепившись к животному, клещ отыскивает подходящее место и присасывается.
Иксодиды на каждой фазе развития питаются один раз. Они присасываются надолго, обычно на несколько дней, и высасывают за это время очень много крови, увеличиваясь по весу в сотни раз. Все их строение приспособлено к такому способу питания. Эти крупные клещи имеют мешковидное тело, одетое эластичной кутикулой, которая сильно растягивается по мере сосания крови. У личинок, нимф и самок, выпивающих много крови, имеется только небольшой спинной щиток и пара маленьких перитремальных щитков по бокам тела, на которых открываются дыхальца хорошо развитых трахей (у личинки перитрем и дыхалец еще нет). У самцов, насасывающих меньше крови или не питающихся, щиток покрывает всю спину, щитки имеются также на брюшной стороне.
По данным Б. И. Померанцева (Иксодовые клещи, «Фауна СССР», 1950), в СССР встречается 54 вида иксодид. В тундре их почти нет. В таежных лесах, северных и горных на юге, преобладает род Ixodes, в широколиственных, особенно на Дальнем Востоке, — Haemaphysalis. Для степей характерен род Dermacentor, хотя немало видов и других родов. Для полупустыни особенно характерны виды рода Rhipicephalus, для пустыни — Hyalomma.
Медицинское и ветеринарное значение иксодид очень велико. Только на территории СССР они причастны к распространению более 20 заболеваний, вызываемых вирусами, риккетсиями, бактериями, кровепаразитарными простейшими и др. Среди иксодид нашей фауны до 30 видов сохраняют возбудителей болезней, есть виды, способные хранить около десятка различных инфекций. В ряде случаев возбудитель размножается в клещах и передается потомству через яйцо, причем сам клещ нисколько не страдает от болезнетворного начала. Специфическая связь иксодид с большим числом инфекций не случайна и, очевидно, объясняется типом паразитизма этих клещей, прежде всего особенностями питания, в недавнее время детально описанного в превосходных работах Ю. С. Балашова. Медленное насасывание огромного количества крови, длительное пребывание паразита в состоянии как бы сращенном с хозяином, рост и развитие органов клеща в процессе кровососания, строгая согласованность питания и созревания яиц, доведенная до единственного в жизни гонотрофического цикла, наконец, разнообразие и смена используемых хозяев — все это создает неповторимо благоприятные условия для заражения клеща и взаимоприспособления возбудителя болезни и клеща-переносчика.

Защита человека и животных от кровососущих клещей составляет неотъемлемую часть общей системы борьбы с паразитами и болезнями, проводимой в нашей стране. Конкретные меры основываются на глубоком изучении болезни и ее возбудителя, клещей-переносчиков и животных-хозяев, путей циркуляции инфекции в очаге, условий заражения человека и т. д. Истребительные меры сочетаются с профилактическими. Клещей уничтожают химическими препаратами на скоте и в открытой природе, что в ряде случаев сочетают с уничтожением грызунов. Химический метод широко применяется в очагах энцефалита: леса вокруг населенных пунктов, участки лесоразработок обрабатывают препаратами ДДТ и гексахлорана опыливанием с самолетов, задымлением с помощью специальных шашек и т. п. Важное значение имеет личная профилактика: предохранительная вакцинация, периодический взаимный осмотр и удаление заползших клещей при передвижении по эпидемиологически опасной местности, защитный противоклещевой костюм, пропитывание одежды отпугивающими клещей препаратами (репеллентами). В борьбе с гемоспоридиозами домашних животных, помимо истребительных мер, используется рациональная система выпаса с учетом зараженных пастбищ и другие меры.
Как говорилось, иксодовые клещи присасываются не сразу, а довольно долго ползают, отыскивая на коже подходящее место. Поэтому тщательный взаимный осмотр одежды и поверхности тела считают наиболее простым и надежным приемом предохранения человека от нападения клещей.
Если клещ присосался, его следует немедленно удалить. Это лучше делать острым пинцетом, крепко удерживая основание «головки» клеща, чтобы последняя не осталась в коже. Обнаруженных клещей сжигают или погружают в дезинфицирующий раствор; место присасывания смазывают иодом.
Помимо трех выше указанных основных группировок — гамазоидных клещей, клещей-уропод и иксодовых клещей, — в состав отряда Parasitiformes входят некоторые другие, небольшие и пока малоизученные группы клещей. Таковы, в частности, своеобразные тропические клещи-голотиры (надсем. Holothyroidea), которые прежде занимали положение обособленного подотряда в системе Acarina, а при разделении клещей на отряды были отнесены А. А. Захваткиным к отряду Parasitiformes.
Голотиры — крупные клещи, до 7 мм длиной, с округлым, выпуклым сверху и плоским снизу туловищем и очень твердыми темноокрашенными покровами. Ноги относительно длинные, хелицеры клешневидные, в основании ротового аппарата имеется теркоподобный орган; округлое половое отверстие расположено между тазиками четвертой пары ног и прикрыто щитками.
Виды рода Holothyrus найдены в Новой Гвинее, на Цейлоне, в Австралии, на островах Индийского океана. Голотиры ядовиты, известны случаи гибели домашней птицы при заглатывании этих клещей.
ОТРЯД КЛЕЩИ-СЕНОКОСЦЫ (OPILIOACARINA, ИЛИ NOTOSTIGMATA)
Клещи-сенокосцы, или опилиакарины, — небольшой отряд клещеобразных арахнид, по некоторым признакам напоминающий примитивных сенокосцев, по другим — паразитиформных клещей и в то же время очень своеобразный. Это мелкие формы, около 1 мм длиной, с продолговато-овальным туловищем и довольно длинными ногами. Покровы кожистые, бороздчатые. Два задних сегмента головогруди и восемь сегментов брюшка разделены спинными бороздами. Имеются двойные боковые глаза. Дыхание трахейное, четыре пары маленьких стигм открываются на спинной стороне четырех передних сегментов брюшка. Хелицеры короткие, клешневидные, жевательные отростки тазиков педипальп с придатками сложной формы, ротовое отверстие ограничено с боков ребристыми лопастями, которые называют «теркой». Половое отверстие расположено между тазиками ног третьей пары, самки имеют яйцеклад. Из яйца вылупляется шестиногая личинка, которая превращается в восьминогую нимфу; общее число линек не установлено. В кишечнике опилиакарин найдены остатки мелких членистоногих, но как они поедаются — неизвестно.
Известно всего несколько видов клещей-сенокосцев, относящихся к роду Opilioacarus единственного семейства Opilioacaridae, найденных в жарких и теплых странах. Все они описаны по немногим экземплярам, обнаруженным в почве, лесной подстилке, под камнями и т. д. Они известны из Южной Европы, Аравии, Патагонии, Техаса. У нас клещи-сенокосцы найдены в Средней Азии: Opilioacarus hexophthalmus в Семиречье и Opilioacarus italicus в отрогах Угамского хребта к северо-востоку от Ташкента.