(Maxillopoda Dahl, 1956) Класс Максиллоподы, Челюстеногие, Челюстеногие ракообразные, Class Maxillopoda Dahl, 1956 (Maxillopods) 22 отряда
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ПОДКЛАСС МАКСИЛЛОПОДЫ (MAXILLOPODA)
Грудной отдел у Maxillopoda состоит из определенного числа сегментов, обычно из 6 или реже оно сокращено до 5 или 4. Грудные ножки служат только для передвижения или для возбуждения токов воды, но не несут дыхательной функции и лишены внутренних отростков, подающих пищу ко рту. Ротовые придатки хорошо развиты, часто служат для фильтрации. Сложные глаза и жабры обычно отсутствуют, кровеносная система сильно упрощена или отсутствует. Развитие со сложным метаморфозом.
В состав этого подкласса наряду со свободноживущими рачками, сохраняющими типичное для Maxillopoda строение, входят ракообразные, ведущие сидяче-прикрепленный и паразитический образ жизни, сильно изменивший их организацию.
Maxillopoda распространены чрезвычайно широко. Они населяют как море, так и пресные воды, живут как на дне, так и в толще воды, обитают в подземных и в капиллярных водах, пропитывающих песок, опускаются на огромные океанические глубины, паразитируют на других животных, в том числе и на других ракообразных, — словом, они эволюционировали в различных направлениях и приспособились к существованию в самых разнообразных условиях.
В этот подкласс включают 5 отрядов — веслоногих, карпоедов, мистакокарид, аскоторацид и усоногих.
ОТРЯД ВЕСЛОНОГИЕ РАКООБРАЗНЫЕ (COPEPODA)
При изучении жизни каждого водоема — от маленькой лужи до океана — исследователь встречается с этими рачками. Планктонная сеть, протянутая на любой глубине океана, в жарких тропиках или в холодных приполярных водах, приносит улов, состоящий преимущественно из веслоногих рачков. В ручьях и озерах глубоких темных пещер также обитают эти ракообразные. Они массами плавают во всех обычных пресноводных водоемах, часто не уступая по численности ветвистоусым. Некоторые из них паразитируют на рыбах и других водных животных, причем нередко под влиянием паразитизма меняются настолько, что не сразу можно угадать их истинную природу. Веслоногие массами поедаются рыбами. Сельдь, иваси и многие другие рыбы питаются почти исключительно веслоногими. Большинство этих ракообразных — животные микроскопические, длина их тела от 1 мм и меньше до 5 мм; лишь отдельные глубоководные и паразитические виды достигают в длину 1—3 см.
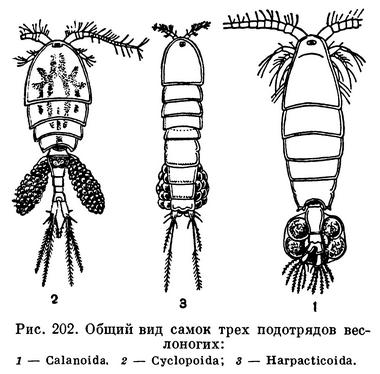
Тело свободноживущих веслоногих разделяется на головогрудь, грудь и брюшко (рис. 202). Голова слитная, без всяких следов сегментации, срастается с первым грудным сегментом, образуя головогрудь. Передний конец головы часто вытянут в загнутый вниз клюв, или рострум. Очень характерно отсутствие парных фасеточных глаз; на лобной части головы расположен только науплиальный глазок. Именно это обстоятельство позволило датскому натуралисту Мюллеру в свое время назвать обычных пресноводных веслоногих «циклопами» в честь одноглазого гиганта греческой мифологии.
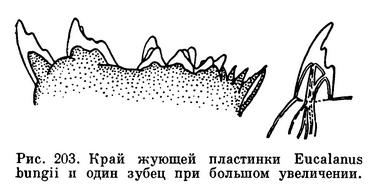
Голова снабжена 5 парами придатков. Передние антенны часто очень длинные, иногда длиннее тела, и участвуют в плавании и парении рачков. Кроме того, они выполняют и функции органов чувств: на них сидят чувствительные щетинки и цилиндрические чувствительные придатки. Задние антенны короткие, обычно двуветвистые. Жвалы мощные и имеют двуветвистый щупик. Их жевательная сильно хитинизированная часть обладает острыми зубцами, помогающими раздроблять пищу. При внимательном рассмотрении зубцов жвал некоторых морских веслоногих выяснилось, что эти зубцы покрыты кремневыми коронками, увеличивающими их прочность (рис. 203). Открытие кремневых коронок интересно в двух отношениях. Во-первых, оно указывает на способность веслоногих ракообразных усваивать и концентрировать кремний; такой способности лишены почти все высшие беспозвоночные — черви, моллюски, а также другие членистоногие. Во-вторых, можно надеяться найти в геологических отложениях кремневые коронки древних веслоногих, почти совсем не сохранившихся в ископаемом состоянии.
Передние челюсти веслоногих устроены очень сложно, так как снабжены внутренними и наружными лопастями и многочисленными перистыми щетинками. Задние челюсти имеют только внутренние лопасти и также многочисленные щетинки. К головным придаткам присоединяется пара одноветвистых ногочелюстей, принадлежащих переднему слившемуся с головой грудному сегменту.
Задние антенны, щупики жвал и передние челюсти фильтрующих веслоногих совершают частые и беспрерывные взмахи, создающие круговороты воды, которые приносят взвешенные пищевые частицы. Эти частицы отфильтровываются главным образом щетинками задних челюстей.
Грудной отдел состоит из 5 сегментов с ясно видимыми границами между ними. Все 5 пар грудных ножек у примитивных веслоногих устроены одинаково. Каждая ножка состоит из 2-члениковой основной части и двух обычно 3-члениковых ветвей, вооруженных шипами и щетинками. Эти ножки совершают одновременные взмахи, действуя как весла и отталкивая тело рачка от воды. У многих более специализированных видов пятая пара ножек самца преобразована в аппарат, приспособленный для удерживания самки во время спаривания и прикрепления сперматофоров к ее половым отверстиям. Нередко пятая пара ног редуцирована.
Брюшной отдел состоит из 4 сегментов, но у самок часто их число меньше, так как некоторые из них сливаются между собой. На переднем брюшном сегменте открывается парное или непарное половое отверстие, и у самки этот сегмент часто крупнее остальных. Брюшко заканчивается тельсоном, с которым сочленены фуркальные ветви. Каждая из них вооружена несколькими очень длинными, иногда перистыми щетинками. Эти щетинки особенно сильно развиты у планктонных видов, у которых они приспособлены для парения в воде, так как препятствуют погружению рачка.
Строение паразитических видов сильно отклоняется от описанного. Их головные придатки превращаются в органы прикрепления к хозяину и в связи с этим часто имеют вид хитиновых крючков, погруженных в кожу рыб или других водных животных. Плавательные ножки иногда исчезают совсем, или число их сокращается. В большей или меньшей степени исчезает сегментация, покровы становятся мягкими. Специализированные паразиты приобретают червеобразную форму тела и теряют всякое сходство с ракообразными. Однако можно подобрать полную серию переходов между такими предельно упрощенными видами и нормальными свободноживущими. Кроме того, яйца прикрепляются к телу паразитических видов совершенно так же, как к телу свободноживущих, и из яиц выходят такие же личинки.
Дыхание веслоногих осуществляется всей поверхностью тела, жабры отсутствуют. С этим, возможно, связано и слабое развитие или даже отсутствие кровеносной системы. Сердце есть только у представителей подотряда Calanoida, да и у них оно невелико, хотя и бьется очень часто: например, у морского рачка Labidocera оно совершает более 150 ударов в минуту. У остальных веслоногих полостная жидкость приводится в движение сокращениями кишечника.
Все веслоногие раздельнополы. Самцы обычно меньше самок и у свободноживущих видов отличаются от самок искривленными (одной или обеими) передними антеннами, а также иным строением пятой пары грудных ножек. Особенно резко различны самцы и самки многих паразитических видов. У некоторых из них прикрепленные к хозяину самки полностью теряют сегментацию и конечности, а маленькие самцы сохраняют облик поздней личинки и свободно плавают. У других (семейство Lernaeopodidae) маленькие самцы, также сходные с поздней личинкой, своими клешневидными челюстями и ногочелюстями прикрепляются к крупным червеобразным паразитическим самкам.
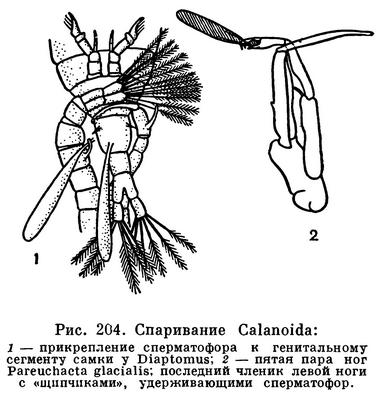
При спаривании самец удерживает самку пятой парой грудных ножек и первыми антеннами и при помощи той же пятой пары ног приклеивает колбасовидный сперматофор близ ее половых отверстий, т. е. к нижней стороне первого брюшного сегмента. У некоторых видов одна из ветвей пятой пары ног самца снабжена на конце щипчиками, захватывающими сперматофор и переносящими его на нужное место (рис. 204). Из сперматофора сперма попадает в семеприемник самки. При выметывании яиц они оплодотворяются.
Большинство морских планктонных видов откладывает яйца прямо в воду, но у всех пресноводных и у всех паразитических, а также у морских донных и прибрежных видов яйца склеиваются между собой специальным секретом и прикрепляются близ половых отверстий самки. У одних веслоногих таким образом формируется один, у других — два яйцевых мешка, которые самка носит на себе до выхода из яиц личинок.
Из яйца выходит личинка науплиус. Личинка многократно линяет и постепенно приближается по своим признакам к взрослому рачку. Различают 12 личиночных стадий веслоногих. Первые две стадии — ортонауплиуса — характеризуются присутствием только обеих пар антенн и пары жвал, у следующих четырех стадий — метанауплиуса — закладываются и развиваются остальные ротовые придатки, но тело остается несегментированным. Последние 6 стадий называются копеподитными и отличаются сегментацией заднего конца тела и постепенным развитием грудных ножек. Для завершения метаморфоза разным веслоногим требуется различное время, да и биология личинок далеко не у всех видов одинакова.
Образ жизни, способ питания и среда обитания веслоногих рачков настолько разнообразны, что лучше рассмотреть этот отряд не целиком, а каждый из входящих в него подотрядов в отдельности.
В пределах отряда веслоногих принято различать 5 подотрядов, из которых 2 объединяют исключительно паразитические виды. (Некоторые исследователи разделяют паразитических веслоногих на 5 или 6 подотрядов.) Свободноживущие веслоногие относятся к подотрядам Calanoida, Cyclopoida и Harpacticoida (рис. 202).
Calanoida — исключительно планктонные животные. Их голова и грудь значительно длиннее узкого брюшка, передние антенны очень длинные, превосходят голову и грудь, а нередко и все тело рачка, если есть яйцевой мешок, то один.
Cyclopoida обитают преимущественно на дне или в придонных слоях воды. Среди них есть и паразиты. Их голова и грудь относительно всего тела гораздо короче, чем у Calanoida, передние антенны короткие, не достигают и середины головогрудного отдела, брюшко уже груди. Самки всегда носят два яйцевых мешка.
Harpacticoida, за единичными исключениями, живут на дне и больше ползают, чем плавают. Тело их червеобразно благодаря тому, что брюшной отдел почти не отличается по ширине от грудного. Передние антенны очень короткие, самки большинства видов образуют один яйцевой мешок.
Представители всех трех подотрядов населяют как моря, так и пресные воды.
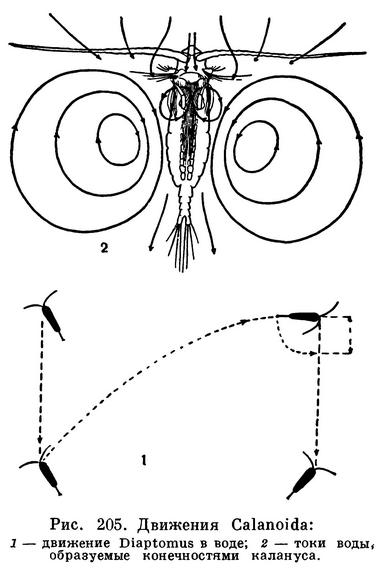
ПОДОТРЯД КАЛАНИДЫ (CALANOIDA)
Вся организация Calanoida превосходно приспособлена к жизни в толще воды. Длинные антенны и перистые щетинки фуркальных ветвей позволяют морскому Calanus или пресноводному Diaptomus неподвижно парить в воде, лишь очень медленно погружаясь. Этому способствуют находящиеся в полости тела рачков капли жира, уменьшающие их удельный вес. Во время парения тело рачка располагается вертикально или наклонно, причем передний конец тела расположен выше заднего. Опустившись на несколько сантиметров вниз, рачок делает резкий взмах всеми грудными ножками и брюшком и возвращается на прежний уровень, после чего все повторяется сначала. Таким образом, путь рачка в воде рисуется зигзагообразной линией (рис. 205, 1). Некоторые морские Calanoida, как например приповерхностный вид ярко-синего цвета Pontellina mediterranea, совершают настолько резкие скачки, что выпрыгивают из воды и пролетают какое-то пространство по воздуху наподобие летучих рыб.
Если грудные ножки действуют время от времени, то задние антенны, щупики жвал и передние челюсти вибрируют беспрерывно с очень большой частотой, совершая до 600—1000 ударов ежеминутно. Их взмахи вызывают мощные круговороты воды с каждой стороны тела рачка (рис. 205, 2). Эти токи проходят через образованный щетинками челюстей фильтрационный аппарат, и отфильтрованные взвешенные частицы проталкиваются вперед к жвалам. Жвалы размельчают пищу, после чего она поступает в кишечник.
Для определения интенсивности фильтрации Calanus применялись водоросли, меченные радиоактивными изотопами углерода и фосфора. Оказалось, что один рачок за сутки пропускает через свой фильтрационный аппарат до 40 и даже до 70 см3 воды, причем питается он преимущественно ночью.
Питание водорослями необходимо для многих Calanoida. Так, например, половые продукты Calanus finmarchicus созревают только при достаточном потреблении рачком диатомовых водорослей.
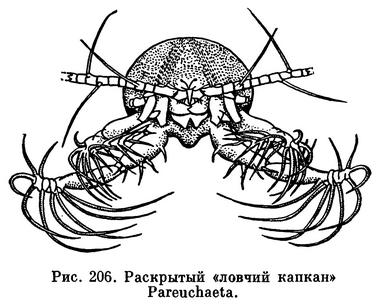
Помимо фильтраторов, среди Calanoida есть и хищные виды, большинство которых обитает на значительных или больших океанических глубинах, где планктонные водоросли не могут существовать из-за отсутствия света. Задние челюсти и ногочелюсти таких видов снабжены крепкими острыми шипами и приспособлены для схватывания жертв. Особенно интересны приспособления для добывания пищи у некоторых глубоководных видов. Винкстед наблюдал, как глубоководная Pareuchaeta неподвижно висит в воде, расставив в стороны свои удлиненные ногочелюсти, образующие нечто вроде капкана (рис. 206). Как только между ними оказывается жертва, ногочелюсти смыкаются, капкан захлопывается. При крайней разреженности организмов на больших океанических глубинах такой способ охоты оказывается наиболее целесообразным, так как затрата энергии на активные поиски жертв не окупается их поеданием.
С особенностями движения и питания Calanoida связана сложная проблема их суточных вертикальных миграций. Скорость передвижений рачков при их вертикальных миграциях измеряется величинами порядка 10—30 см в минуту. Если принять во внимание длину их тела (для Calanus finmarchicus, например, около 2 мм), то такую скорость приходится считать значительной. При этом не только подъем кверху, но и опускание вниз осуществляется за счет активных движений рачков, а не за счет их пассивного погружения.
Не следует думать, что, совершая вертикальные миграции, все рачки одновременно двигаются в каком-нибудь определенном направлении. Английский ученый Бэйнбридж опускался под воду и проводил наблюдения над мигрирующими веслоногими. Он видел, как в одном и том же слое воды часть рачков движется вверх, а другая — вниз. В зависимости от того, какое движение преобладает, перемещается вверх или вниз вся масса животных.
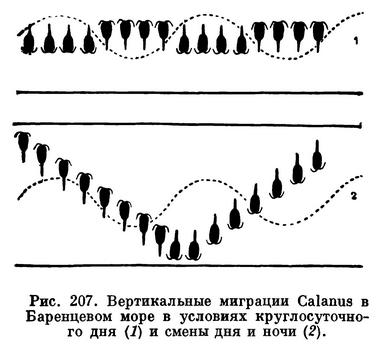
Вопрос о причинах вертикальных миграций до сих пор окончательно не выяснен. Совершенно очевидно, что стремление рачков подняться в поверхностные слои объясняется изобилием там планктонных водорослей, которыми питаются веслоногие-фильтраторы. Менее понятны причины, заставляющие рачков покидать эти богатые пищей слои. Многие исследователи считают, что свет вредно влияет на рачков и, избегая его, они утром начинают уходить вниз. Важное значение света подтверждается наблюдениями В. Г. Богорова над вертикальным распределением веслоногих в Баренцевом море летом, т. е. в условиях круглосуточного освещения. Оказалось, что в это время Calanus finmarchicus неизменно находится на одной глубине, там, где условия освещения для него наиболее благоприятны. В этом районе моря в толще воды наблюдаются внутренние волны, которые должны то несколько приподнимать, то несколько опускать рачков. Очевидно, рачки активно передвигаются в противоположном направлении, так как они в течение круглых суток не выходят за пределы определенного горизонта. Осенью, когда восстанавливается смена дня и ночи, возобновляются обычные вертикальные миграции (рис. 207). Не только солнечный, но и лунный свет заставляет рачков уходить из поверхностных слоев воды в более глубокие.
Протяженность сезонных миграций обычно бывает больше, чем суточных. Первые иногда захватывают 3—4 тысячи метров, а вторые — самое большее несколько сотен метров.
Представители подотряда Calanoida преимущественно морские животные. В настоящее время известно около 1200 морских видов этих рачков, принадлежащих к 150 родам и 26 семействам. В пресных водах обитает всего около 420 видов, распределяющихся между 12 родами и 4 семействами.
Проведенные в последнее время подробные исследования фауны морских каланид показали, что прежние представления о широком распространении многих видов этих ракообразных неправильны. В каждой части океана обитают в основном присущие только ей виды. Каждый вид морских каланид расселяется благодаря несущим рачков течениям. Так, например, поступающие в Полярный бассейн ответвления Гольфстрима заносят туда каланид из Атлантического океана. В северо-западной части Тихого океана в водах теплого течения Куросио обитают одни виды, а в водах холодного течения Оясио — другие. Нередко по фауне каланид удается определить происхождение тех или иных вод в определенных частях океана. Особенно резко различаются по своему составу фауны вод умеренных широт и вод тропиков, причем тропическая фауна богаче видами.
Каланиды обитают на всех океанических глубинах. Среди них ясно различаются поверхностные виды и виды глубоководные, никогда не поднимающиеся в поверхностные слои воды. Как уже указывалось, на больших глубинах преобладают хищники, а на малых — фильтраторы. Наконец, существуют виды, совершающие вертикальные миграции огромного диапазона, то поднимающиеся к поверхности, то опускающиеся на глубину до 2—3 км.
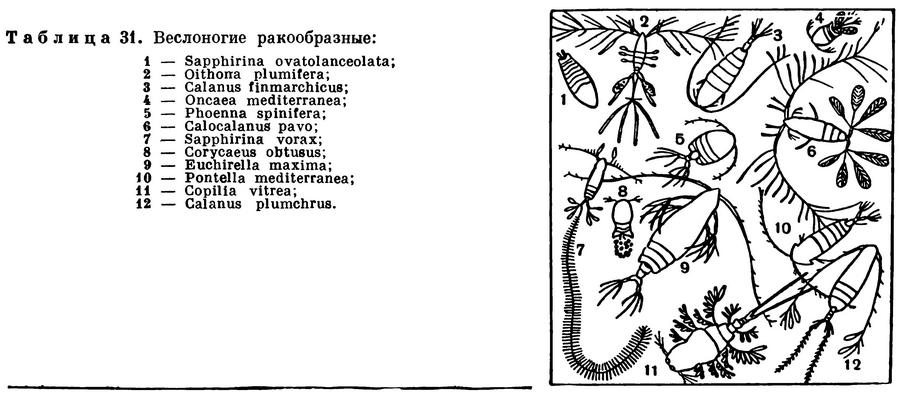
Некоторые мелководные виды каланид в умеренных водах развиваются в огромном количестве и по весу составляют преобладающую часть планктона. Так, например, планктон Баренцева моря приблизительно на 90% состоит из Calanus finmarchicus (табл. 31, 5). Эти рачки характеризуются высокой питательностью: в их теле содержится 59% белков, 20% углеводов и часто более 10—15% жиров.
Многие рыбы, а также усатые киты питаются главным образом каланидами. Таковы, например, сельдь, сардина, скумбрия, анчоус, шпрот и многие другие. В желудке одной сельди было обнаружено 60 000 проглоченных ею веслоногих рачков. Из китов активно потребляют огромные массы каланид финвал, сейвал, синий кит и горбач.
Некоторые пресноводные каланиды появляются в водоемах только в определенное время года, например весной. В весенних лужах часто попадается сравнительно крупный (около 5 мм) Diaptomus amblyodon, окрашенный в яркий красный или синий цвет. Этот вид и некоторые другие широко распространенные пресноводные каланиды способны к образованию покоящихся яиц, стойко переносящих высыхание и вымерзание и легко разносящихся ветром на далекие расстояния.
ПОДОТРЯД ЦИКЛОПЫ (CYCLOPOIDA)
Другой подотряд веслоногих ракообразных — Cyclopoida — наибольшим количеством видов представлен в пресных водах.
Пресноводные циклопы живут во всевозможных водоемах, от мелких луж до крупных озер, причем нередко встречаются в очень большом количестве экземпляров. Основной зоной их обитания служит прибрежная полоса с зарослями водных растений. При этом во многих озерах к зарослям определенных растений приурочены определенные виды циклопов. Так, например, для Валдайского озера в Ивановской области описано 6 группировок растений с соответствующими им группировками видов циклопов.
Сравнительно немногие виды могут считаться настоящими планктонными животными. Некоторые из них, принадлежащие преимущественно к роду Mesocyclops, постоянно обитают в поверхностных слоях воды, другие (Cyclops strenuus и другие виды того же рода) совершают регулярные суточные миграции, опускаясь днем на значительную глубину.
Плавают циклопы несколько иначе, чем каланиды. Одновременно взмахивая четырьмя парами грудных ножек (пятая пара редуцирована), рачок делает резкий прыжок вперед, вверх или вбок, а затем при помощи передних антенн может некоторое время парить в воде. Поскольку центр тяжести его тела смещен вперед, во время парения его передний конец наклоняется и тело может принять вертикальное положение, причем погружение замедляется. Новый взмах ножками позволяет циклопу подняться. Эти взмахи молниеносны — они занимают 1/60 секунды.
Пресноводные циклопы распространены очень широко. Некоторые виды встречаются почти повсеместно. Этому способствуют прежде всего приспособления к перенесению неблагоприятных условий, в частности способность рачков переносить высыхание водоемов и в виде цист пассивно расселяться по воздуху. Кожные железы многих циклопов выделяют секрет, обволакивающий тело рачка, часто вместе с яйцевыми мешками, и образующий нечто вроде кокона. В таком виде рачки могут подвергаться высыханию и вмерзанию в лед, не теряя жизнеспособности. В опытах Камерера циклопы быстро выводились при размачивании сухого ила, сохранявшегося около 3 лет. Поэтому нет ничего удивительного в появлении циклопов в весенних лужах, возникающих при таянии снега, в только что залитых рыбоводных прудах и т. д.
Некоторые циклопы приспособились к жизни в солоноватых и даже в очень соленых водоемах. Род Halicyclops, например, весьма обычен в Каспийском море и не встречается в пресной воде. Microcyclops dengizicus широко распространен только в солоноватых и соленых водоемах зоны пустынь (Ирак, Индия, о. Гаити, Египет, Калифорния, в СССР — в Карагандинской области, в Муганской степи) и прекрасно живет даже при солености, превосходящей морскую (до 41‰). Многие обычные пресноводные виды могут существовать и в солоноватой воде, как например Mesocyclops leuckarti в Финском и Ботническом заливах.
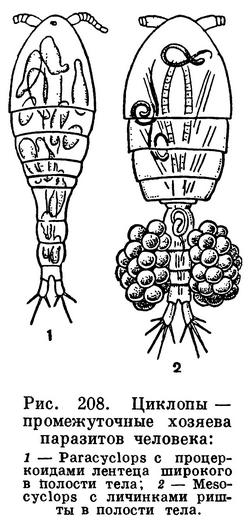
Значение циклопов в жизни наших внутренних водоемов очень велико. С одной стороны, они служат пищей многим рыбам и их молоди, но, с другой стороны, выступают в качестве конкурентов тех же самых рыб, поедая множество мелких животных, как планктонных, так и донных. Не следует забывать и о том, что циклопы способствуют заражению человека некоторыми паразитическими червями, в первую очередь широким лентецом (Diphyllobothrium latum) и риштой (Dracunculus medinensis). Циклопы заглатывают плавающих в воде личинок этих червей. Корацидий широкого лентеца, попав в циклопа, сбрасывает свой мерцательный покров, проникает из кишечника в полость тела рачка и превращается там в стадию процеркоида. Если такой циклоп будет проглочен рыбой, процеркоид перейдет в стадию плевроцеркоида, а затем, при поедании рыбы человеком, кошкой и другими млекопитающими, из плевроцеркоида возникнет взрослый червь, часто вызывающий тяжелое малокровие. Личинки ришты, проглоченные циклопами, также проникают в полость их тела и остаются там в течение 1—1,5 месяцев. При питье сырой воды человек проглатывает циклопов и заражается риштой.
Морские представители подотряда Cyclopoida менее разнообразны, чем пресноводные. Среди них обычны и нередко многочисленны в морском планктоне виды рода Oithona. Очень характерны также крупные (до 8 мм) уплощенные виды рода Sapphirina, поверхность тела которых отливает яркими синими, золотистыми или темно-красными тонами (табл. 31, 1). Другой близкий морской род — Oncaea (табл. 31, 4) — обладает железами, выделяющими светящийся секрет, и нередко, совместно с другими организмами, вызывает свечение моря.
ПОДОТРЯД ГАРПАКТИЦИДЫ (HARPACTICOIDA)
Об образе жизни представителей третьего подотряда — Harpacticoida — известно гораздо меньше. Эти червеобразные, большей частью очень мелкие рачки, чрезвычайно разнообразны как в морских, так и в пресных водах, но никогда не встречаются в массовом количестве. Описано более 30 семейств и несколько сотен видов Harpacticoida.
Большинство гарпактицид ползает по дну и донным растениям. Лишь единичные виды способны продолжительное время плавать и входят в состав морского планктона (Microsetella). Гораздо более характерны целые группы родов и видов гарпактицид, приспособившиеся к обитанию в особых, необычных условиях, в частности в капиллярных ходах между песчинками морских пляжей и в подземных пресных водах.
Всего несколько лет тому назад зоологи применили очень простой прием, позволяющий изучать население капиллярных ходов морского песка. На пляже, выше уровня моря, выкапывается яма. В ней постепенно накапливается вода, заполнявшая капилляры песка. Эту воду фильтруют через планктонную сеть и таким образом добывают представителей своеобразной фауны, получившей название интерстициальной.
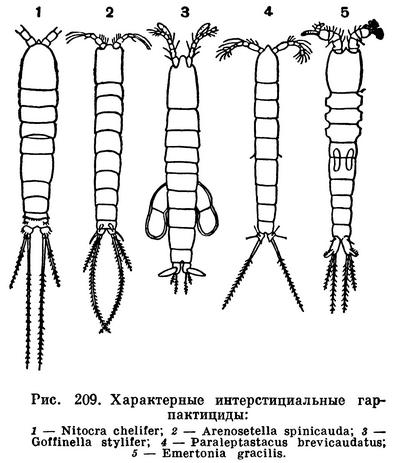
Гарпактициды составляют заметную часть интерстициальной фауны. Их обнаружили всюду, где проводились соответствующие исследования, — на пляжах Англии, вдоль европейского и американского берегов Атлантического океана, на Средиземном и Черном морях, у берегов Африки и Индии, на островах Мадагаскар, Реюньон и Багамских. Большинство интерстициальных гарпактицид принадлежит к особым, обитающим только в таких условиях родам, отличающимся необыкновенно тонким и длинным телом (рис. 209), позволяющим рачкам двигаться в узких капиллярных ходах. Замечательно, что некоторые из этих специализированных видов были найдены в очень отдаленных друг от друга местах. Так, например, на Багамских островах оказались Arenosetella palpilabra, ранее известная только из Шотландии, и Horsiella trisaetosa, ранее известная только из Кильской бухты. Трудно объяснить подобное распространение, поскольку покоящихся яиц у интерстициальных гарпактицид нет.
Чрезвычайно богата видами гарпактицид своеобразная фауна Байкала. Она состоит из 43 видов, из которых 38 эндемичны для этого озера. Особенно много этих рачков в прибрежной части Байкала, на камнях и водных растениях, а также на растущих здесь губках. По-видимому, они питаются за счет губок и в свою очередь становятся жертвами также ползающего по губкам бокоплава Brandtia parasitica.
Некоторые виды гарпактицид приурочены только к водоемам, очень бедным солями, характеризующимся повышенной кислотностью, т. е. связанным с верховыми, сфагновыми, болотами. Таков, например, Arcticocamptus arcticus, биология которого подробно изучена Е. В. Боруцким.
Arcticocamptus arcticus широко распространен на севере Европы, от Большеземельской тундры до Скандинавии, на западном побережье Гренландии, на Новой Земле. Кроме того, он найден в Альпах и в нескольких пунктах средней полосы Европейской части СССР, в том числе в Косине под Москвой, в Звенигороде, под Ярославлем и т. д. Повсюду он обитает в водоемах, связанных со сфагновыми болотами.
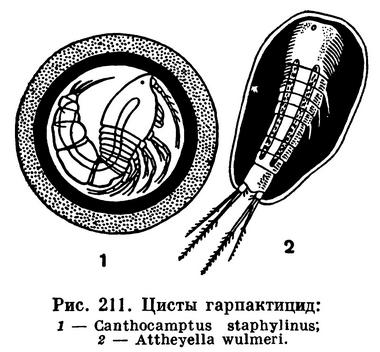
Если Arcticocamptus arcticus в активном состоянии существует весной, то некоторые другие виды пресноводных гарпактицид встречаются только зимой или только летом. При этом известны виды, проводящие период покоя не в стадии покоящегося яйца, как Arcticocamptus arcticus, а в стадии цист, несколько напоминающих описанные выше цисты циклопов. У Canthocamptus staphylinus такие цисты округлые, у Attheyella wulmeri и Attheyella northumbrica они овальные, причем фуркальные щетинки рачка торчат из оболочки (рис. 211).
Практическое значение гарпактицид несравненно меньше, чем каланид и циклопов. В некоторых водоемах они составляют заметную часть пищи рыб, в особенности их молоди.
ПАРАЗИТИЧЕСКИЕ ВЕСЛОНОГИЕ
Паразитические веслоногие ракообразные прикрепляются к самым различным рыбам, как морским, так и пресноводным, а также ко многим морским беспозвоночным — сифонофорам, кораллам, моллюскам, полихетам, эхиуридам, высшим ракообразным, иглокожим и к некоторым другим. Как правило, каждый вид паразита паразитирует на определенном виде хозяина, однако есть и менее разборчивые, способные использовать разные виды рыб.
Большинство паразитических веслоногих представляет собой эктопаразитов. Они поселяются на наружных покровах, на жабрах, на глазах, реже в ротовой полости рыб. Среди паразитов морских беспозвоночных много эндопаразитов. Так, например, Monstrillidae живут в кровеносных сосудах полихет, Mytilicola — в кишечнике моллюсков, несколько родов — в кишечнике иглокожих.
Паразитические веслоногие принадлежат к трем подотрядам — Cyclopoida, включающему наряду с паразитическими и только что рассмотренные свободноживущие виды, Caligoida и Lernaeoida, в состав которых входят исключительно паразитические виды.
Паразитические Cyclopoida отличаются от свободноживущих прежде всего строением своих головных придатков. Задние антенны крючковидные, приспособленные для прикрепления к хозяину. Жвалы, если есть, не бывают жующего типа, другие ротовые части также видоизменены или отсутствуют. Тело некоторых видов сегментировано так же, как у свободноживущих, но у других сегментация в той или иной степени исчезает. У таких специализированных видов редуцируются и грудные ножки. К этому подотряду относится большое количество паразитов пресноводных и морских рыб, а также морских беспозвоночных.
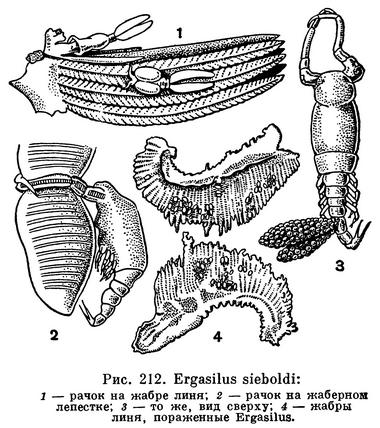
Одними из самых обычных паразитов как пресноводных, так и морских рыб можно считать представителей рода Ergasilus, прикрепляющихся к жабрам линей, щук и многих других рыб. Нередко к одной рыбе прикрепляется по нескольку десятков, а иногда даже до 3000 рачков, что приводит ее к гибели. Замечено, что, чем старше рыба, тем больше рачков на ней паразитирует. Рачки охватывают жаберные стебельки своими крючковидными задними антеннами, при смыкании образующими кольцо, на котором паразит, по выражению В. А. Догеля, болтается, как замок на скобе (рис. 212, 3). Постепенно эпителий жабр разрушается.
Вышедшие из яиц личинки Ergasilus ведут свободный образ жизни. Через 2—2,5 месяца рачки достигают половозрелости и спариваются. Оплодотворенные самки активно двигаются против течения. Это помогает им оседать на жабры рыб, поскольку из-под жаберной крышки направляется ток воды.
Таким же путем жабры рыб поражаются глохидиями перловиц. Интересно, что между эргазилидами и глохидиями существует антагонизм: одни вытесняют других и совместно на жабрах одной рыбы не встречаются.
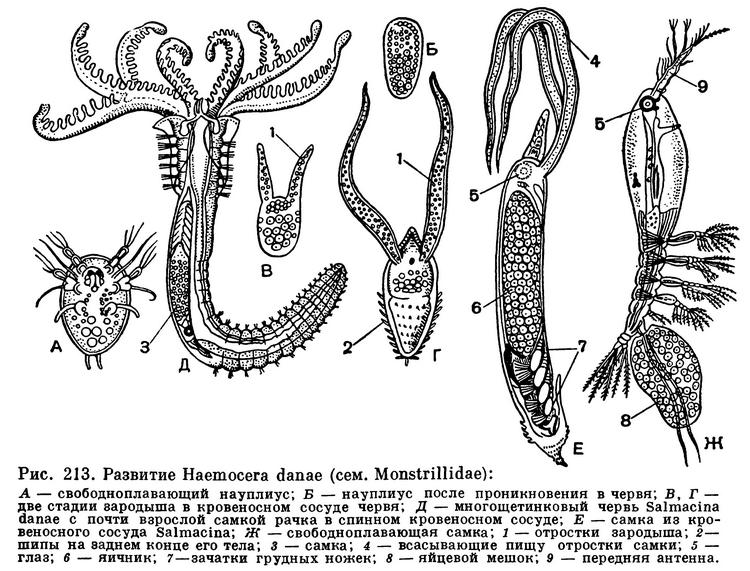
Если эргазилиды паразитируют во взрослом состоянии и ведут свободный образ жизни на ранних стадиях своего развития, то представители семейства Monstrillidae, паразитирующие в морских многощетинковых червях, имеют совсем другой жизненный цикл. Взрослые рачки оживленно плавают подобно обычным свободноживущим веслоногим. Замечательно, однако, что они полностью лишены кишечника и совсем не питаются. Из яиц этих самок выходят также свободноплавающие науплиусы, как и взрослые, лишенные кишечника. Науплиусы отыскивают определенные виды полихет (из родов Polydora, Salmacina и др.) и внедряются в них, проникая в спинной кровеносный сосуд. Здесь науплиус линяет, превращаясь в многоклеточное овальное тельце. Впоследствии у этого зародыша на переднем конце вырастают два длинных придатка, служащих для всасывания пищи. Зародыш линяет еще раз и преобразуется в длинное колбасовидное тело, внутри которого формируется взрослый рачок с хорошо развитыми половыми органами. Он прорывает стенку кровеносного сосуда и покровы хозяина и приступает к активному существованию (рис. 213).
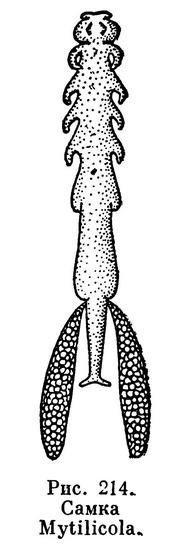
Из паразитов беспозвоночных наибольшее хозяйственное значение имеет Mytilicola (рис. 214). Взрослые самки рачка достигают 8 мм длины и живут в кишечнике мидий и устриц. Тело паразита вытянутое, червеобразное, но следы сегментации сохраняются; грудные ножки редуцированы. Науплиусы и метанауплиусы живут свободно, первая копеподитная личинка внедряется в кишечник моллюсков и превращается там во взрослого рачка. Заражение Mytilicola вызывает массовую гибель мидий. Запасы этих промысловых моллюсков у берегов Голландии в результате заражения рачками в 1949 и 1950 годах уменьшились в 10 раз.
Представители подотряда Caligoida характеризуются расширенным, сплющенным в спинно-брюшном направлении телом, сегментация грудного отдела в той или иной степени утеряна, самки имеют очень крупный передний брюшной (генитальный) сегмент, к которому прикрепляются два яйцевых мешка, ротовые придатки образуют хоботок, позволяющий высасывать кровь хозяина. Самки и самцы мало различаются по величине и по строению.
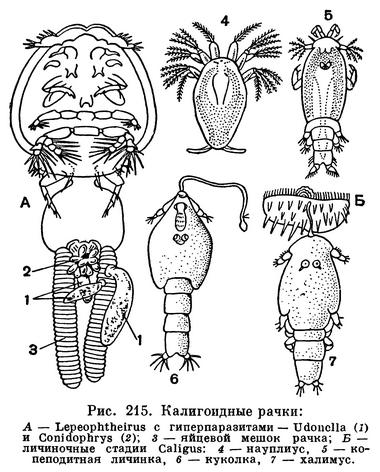
Эти рачки паразитируют главным образом на рыбах, как морских, так и пресноводных, но род Archicaligus живет на замечательном головоногом моллюске Nautilus. Взрослые особи могут на некоторое время покидать хозяина, плавать, а затем садиться на другого. Науплиусы ведут свободное существование. Личинка на копеподитной стадии снабжена специальной лобной нитью, при помощи которой она прикрепляется к хозяину. После этого она переходит в стадию куколки, помещающейся в кутикулярной оболочке предыдущей стадии, неподвижной и непитающейся. Куколка линяет и превращается в также прикрепленную последнюю личиночную стадию — халимус, которая преобразуется, наконец, во взрослого рачка (рис. 215, Б).
Некоторые калигиды, будучи сами паразитами рыб, подвергаются нападению других паразитов. Так, например, на Lepeophtheirus, живущем на многих морских рыбах, паразитируют плоские черви Udonella и инфузории Conidophrys, выступающие в качестве «сверхпаразитов», т. е. паразитов паразита (рис. 215, А).
Последний подотряд паразитических веслоногих — Lernaeoida — объединяет наиболее сильно видоизмененных под влиянием паразитизма рачков. Взрослые половозрелые самки обычно имеют червеобразное тело без всяких следов сегментации или даже разделения на отделы тела, с выростами на переднем конце, погруженными в кожу хозяина, и с двумя яйцевыми мешками на заднем конце. Самцы гораздо мельче самок и по своему строению напоминают копеподитные стадии. Рачки паразитируют на рыбах, реже на амфибиях и китах. Некоторые из них достигают очень крупных размеров: паразит китов Penella balaenoptera длиною 32 см может считаться крупнейшим представителем веслоногих. Он погружен в наружные покровы китов на 5—7 см.
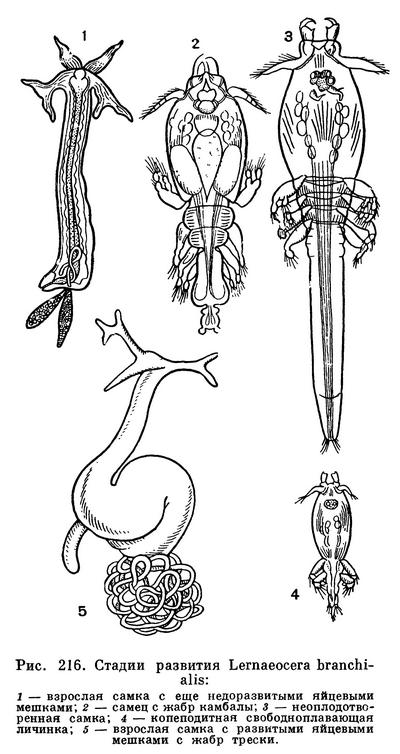
Развитие этих рачков обычно укорочено. Стадии науплиуса и метанауплиуса они проходят в яйце. Из него вылупляется свободная копеподитная личинка, достигающая половозрелости. Вскоре происходит копуляция, после которой самцы умирают. Молодые самки семейства Lernaeidae сначала паразитируют на одних рыбах, например на камбаловых, а затем переходят на тресковых, теряя следы сегментации, конечности и превращаясь в червеобразные существа (рис. 216). Первым хозяином гигантской Penella служат головоногие моллюски, а вторым — киты. У представителей другого обширного семейства этого подотряда — Lernaeopodidae — смены хозяев не происходит и копеподитная личинка совершает свой метаморфоз на одном хозяине.
Лернеиды нередко вызывают гибель пораженных ими рыб, как морских, так и пресноводных. Они вредят лососевым рыбам, разрушая их жабры, описаны случаи смерти морских рыб, вызванной этими рачками. Глубоко внедряясь в тело рыб, некоторые из лернеид достигают сердца и печени своих хозяев, другие вызывают образование кровоточащих ран на поверхности тела хозяина. Кроме того, питаясь кровью рыб, эти паразиты сильно их истощают.
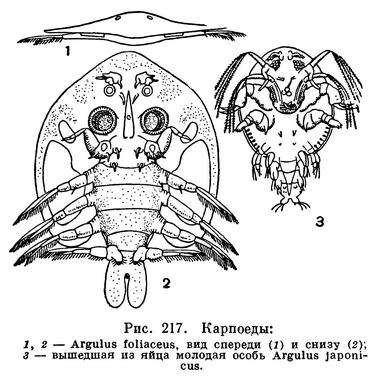
ОТРЯД КАРПОЕДЫ, ИЛИ КАРПОВЫЕ ВШИ (BRANCHIURA)
На пресноводных рыбах паразитируют не только веслоногие ракообразные. Нередко на поверхности их тела, особенно там, где кожа тоньше и не так сильны токи омывающей рыбу воды, т. е. за жаберными крышками и за грудными плавниками, можно найти сравнительно крупных (3—8 мм) очень плоских рачков, прикрепленных к коже рыбы присосками и жадно сосущих их кровь.
В отличие от веслоногих их нельзя считать постоянными паразитами: насытившись, карпоеды оставляют рыбу и с огромной скоростью уплывают прочь. Карпоеды живут не только в пресных водах, но и в море. Всего их известно около 60 видов.
По своему строению эти рачки довольно резко отличаются от веслоногих (рис. 217). Их плоское тело явственно подразделяется на два отдела — передний и задний. Передний, головогрудной, отдел сверху покрыт широким головогрудным щитом — карапаксом, на спинной поверхности которого расположена пара крупных фасеточных глаз, а между ними три простых науплиальных глазка. Частично этот щит покрывает и передний из четырех относящихся к заднему отделу тела свободных грудных сегментов, снабженных каждый парой двуветвистых плавательных ножек.
Тело заканчивается укороченным брюшным отделом, слившимся с широкими фуркальными ветвями.
Придатки переднего отдела приспособлены к паразитическому существованию, а придатки заднего отдела — к плаванию. Обе пары антенн очень короткие. Жвалы образуют направленный назад хоботок, вводимый в покровы рыб и служащий для сосания их крови. По направлению вперед от него отходит острый шип, также помогающий рачку удерживаться на коже рыб. По бокам от хоботка расположены мощные присоски, представляющие собой очень сильно измененные передние челюсти. Задние челюсти одноветвистые и служат для прикрепления к хозяину. Четыре пары грудных ножек несут многочисленные перистые щетинки и совершают сильные взмахи, благодаря которым рачок может быстро плавать спинной стороной вверх. Кишечник карпоедов так же приспособлен к паразитическому существованию, как и их ротовые придатки. Средний его отдел снабжен разветвленными слепыми выростами, служащими резервуарами для высосанной крови. Благодаря способности запасать пищу впрок карпоеды могут до 3 недель не питаться, постепенно расходуя свои запасы.
При нападении рачка на рыбу зрение не играет никакой роли. Рачки руководствуются главным образом движением воды и отчасти химическим чувством. Они воспринимают соответствующие раздражения многочисленными чувствительными щетинками, расположенными на разных частях тела. Если в аквариум с карпоедами опустить фильтровальную бумажку и подвигать ею в воде, рачки бросаются на бумажку, но, прикоснувшись к ней, отплывают прочь. Если бумажкой предварительно обтерта рыба, карпоеды задерживаются на ней несколько дольше. Карпоеды преследуют рыбу, быстро двигаясь в том же направлении параллельно ей, а затем садятся на голову жертвы. Впоследствии они переползают на те части тела рыбы, которые наименее сильно омываются водою и где покровы сравнительно тонкие, т. е. прикрепляются преимущественно позади жаберных крышек и грудных плавников. Присосавшись к рыбе, рачки продолжают действовать своими плавательными ножками, возбуждая ими ток воды, необходимый для дыхания. Оно осуществляется через тонкостенные участки карапакса. Сердца у карпоедов нет; кровь в полости тела двигается благодаря сокращениям кишечника и мускулатуры брюшного отдела.
Никакого предпочтения определенным видам рыб карпоеды не оказывают. Больше того, они могут нападать и на других водных позвоночных — на тритонов и на головастиков лягушек. Некоторые морские виды карпоедов паразитируют на головоногих моллюсках, например Argulus arcussonensis на каракатицах.
Самец оплодотворяет присосавшуюся к рыбе самку, удерживая ее задние грудные ножки своими и подсовывая под нее свой брюшной отдел так, чтобы отверстия его семепроводов совпали с отверстиями ее яйцеводов. Сперма попадает непосредственно в половые пути самки. Перед откладкой яиц самка покидает рыбу и плавает в поисках подводных растений, камней или другого подходящего субстрата. Она откладывает двойной ряд яиц (обычный Argulus foliaceus от 20 до 250 яиц) на этот субстрат, прикрепляя их специальным секретом. Обычно яйца выметываются не сразу, а с перерывами, во время которых самка снова нападает на рыб и сосет их кровь. В этом отношении карпоеды отличаются от большинства других ракообразных, самки которых носят яйца на себе в яйцевых мешках или в специальной выводковой сумке.
Через 3—5 недель из яиц Argulus foliaceus выходят молодые, но еще не вполне сформировавшиеся рачки (рис. 217, 3). Они имеют длинные задние антенны, с помощью которых плавают, их жвалы снабжены щупиками, передние челюсти еще не превращены в присоски, плавательные ножки недоразвиты, а карапакс укорочен. Личинки прикрепляются к рыбам, используя для этого передние антенны и концевые шипы передних челюстей. В течение первых 8 дней пребывания на рыбе рачок дважды линяет, причем его задние антенны и мандибулы укорачиваются, но зато полностью развиваются плавательные ножки. После третьей линьки за счет передних челюстей начинают образовываться присоски; этот процесс заканчивается только после пятой линьки. Так, постепенно, в результате линек, происходящих каждые 3—4 дня, формируется взрослый рачок. У некоторых других видов карпоедов развитие сокращено еще больше: из яйца выходит вполне сформировавшаяся, но еще неполовозрелая особь.
Карпоеды довольно широко распространены как в пресных водах, так и в морях и могут считаться весьма обычными паразитами рыб, в том числе и хозяйственно ценных. Однако вред, приносимый ими рыбам, невелик. В очень редких случаях они вызывают гибель рыб, да и то только молодых.
ОТРЯД МИСТАКОКАРИДЫ (MYSTACOCARIDA)
Песчаные морские пляжи южных морей привлекают массу отдыхающих. Искупавшись, любители загорать растягиваются на песке и подставляют свое тело под жаркие лучи солнца. Но вряд ли кто-нибудь из них знает, что в толще песка, на котором он расположился, идет интенсивная жизнь мелких своеобразных существ.
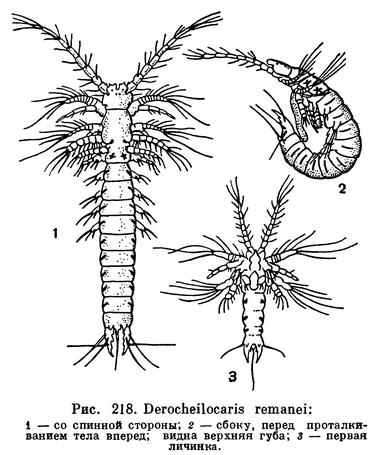
Только поверхностные слои песка, да и то на некотором расстоянии от уреза воды, лишены влаги. На той или иной глубине песок пропитан морской водой. Она заполняет узкие капиллярные пространства между песчинками, и в этих извилистых ходах, диаметр которых обычно меньше 1 мм, обитает особая фауна, получившая название интерстициальной. В нее входят представители разных групп животных — простейшие, кишечнополостные, различные черви, водяные клещи и многие другие, но особенно многочисленны приуроченные к таким своеобразным условиям ракообразные. Выше уже упоминались интерстициальные гарпактициды, однако представители этого подотряда веслоногих встречаются не только в пропитывающей песок воде, но и в других весьма различных условиях. Совместно с гарпактицидами в капиллярных ходах пляжей Средиземного моря и некоторых участков Атлантического и Индийского океанов живут мелкие (до 0,5 мм) рачки, принадлежащие к особому отряду мистакокарид, который приспособлен к существованию исключительно в интерстициальных водах.
Мистакокариды были открыты только в 1943 году на американском побережье Атлантического океана. Впоследствии они были обнаружены на Атлантическом побережье Европы и Африки и вдоль почти всех берегов западной половины Средиземного моря, далее, в Южной Африке и, наконец, в Чили. При предварительных исследованиях интерстициальной фауны Черного и Каспийского морей найти этих рачков пока не удалось, так что для СССР они еще не указаны. Все они принадлежат к одному роду Derocheilocaris, включающему всего 3 вида — американский Derocheilocaris typicus, европейско-африканский Derocheilocaris remanei и чилийский Derocheilocaris galvarini.
Подобно другим представителям интерстициальной фауны мистакокариды имеют длинное и тонкое червеобразное тело, позволяющее им ползать по узким ходам между песчинками. Голова удлиненная и устроена своеобразно. Ее передняя часть — акрон, несущая передние антенны, отделена швом от задней части, на которой расположены задние антенны и хорошо развитые ротовые придатки. С задней частью головы частично сливается первый грудной сегмент, конечности которого преобразованы в ногочелюсти. Далее следуют 4 свободных грудных сегмента. Каждый из них снабжен парой коротких нерасчлененных придатков с щетинками на концах, представляющих собой редуцированные грудные ножки. Брюшной отдел состоит из 6 сегментов, подобных грудным, но лишенных конечностей. Он заканчивается длинными когтевидными фуркальными ветвями. На всех грудных и брюшных сегментах, за исключением последнего, с боков расположены щелевидные углубления. Назначение этих органов неизвестно (рис. 218, 1).
Мистакокариды раздельнополы, но самцы и самки внешне совершенно сходны. Процессы спаривания и откладки яиц этих ракообразных наблюдать не удалось. Изучение их личинок показало, что из яйца выходит метанауплиус длиною 0,15 мм. Личинка линяет 9 раз, причем при каждой линьке добавляется по одному сегменту и постепенно развиваются недостающие конечности. Такое равномерное и постепенное превращение личинки во взрослое животное и значительное количество личиночных стадий говорит о примитивном характере постэмбрионального развития мистакокарид.
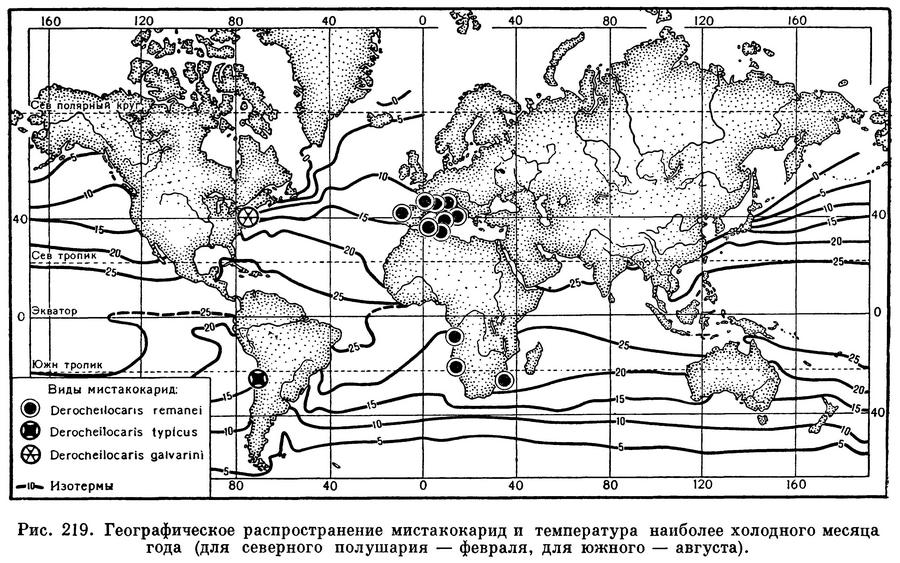
Далеко не каждый песчаный пляж населен мистакокаридами. Они могут жить только в очень мелком песке, частицы которого имеют диаметр около 0,2 мм. Как в более, так и в менее крупнозернистом грунте они не встречаются. Гораздо менее чувствительны эти рачки к солености и температуре. Наблюдения на пляжах Средиземного моря показали, что мистакокариды без всякого вреда для себя переносят колебания солености от 10 до 32‰ и температуры от 10 до 26°С. Все известные их местонахождения как в северном, так и в южном полушарии приурочены к зонам, температура которых даже в самое холодное время года не опускается ниже 10° С (рис. 219).
Может быть, именно этим объясняются неудачи поисков мистакокарид в наших морях.
Распространение мистакокарид ставит перед исследователями некоторые трудные и еще не решенные вопросы. На рисунке 219 видно, что Derocheilocaris remanei образует отдельные колонии, удаленные друг от друга на сотни и тысячи километров. Никаких покоящихся стадий у мистакокарид нет, возможности их расселения крайне ограничены, и, следовательно, эти колонии полностью изолированы одна от другой. В таких случаях обычно представители различных изолированных колоний эволюционируют в разных направлениях, что приводит в конце концов к возникновению разных видов. Между тем мистакокариды Средиземного моря и атлантического побережья Южной Европы совершенно идентичны рачкам с атлантического побережья Африки. Приходится признать, что эволюция обитателей интерстициальных вод идет очень медленно; поэтому, несмотря на давнюю изоляцию отдельных колоний мистакокарид, строение составляющих их рачков осталось неизменным.
Если предположение о замедленном темпе эволюции обитателей интерстициальных вод правильно, то можно рассчитывать встретить среди них древние, мало изменившиеся виды. К таким видам следует причислить и мистакокарид, которым свойственны многие чрезвычайно примитивные признаки (например, строение нервной системы и ротовых придатков) и примитивный характер развития.
ОТРЯД МЕШКОГРУДЫЕ РАКООБРАЗНЫЕ (ASCOTHORACIDA)
В 1887 году на организованной незадолго до этого на Соловецких островах биологической станции проводились исследования фауны Белого моря. Известный русский зоолог Н. Вагнер вскрывал морских звезд. В полости тела одной из них он обнаружил разветвленное мягкое тело какого-то паразита. Вскоре будущий академик В. М. Шимкевич нашел такое же существо в полости тела многолучевой морской звезды Solaster. В 1889 году он писал про свою находку: «Настолько загадочная форма, что почти ничего невозможно сказать относительно ее природы». Этой «загадочной формой» заинтересовался другой крупный зоолог и будущий океанограф — Н. М. Книпович, детально ее изучивший. В результате кропотливых многолетних исследований он установил, что это существо принадлежит к группе паразитических ракообразных, которую в то время относили к отряду усоногих, а теперь считают самостоятельным отрядом мешкогрудых ракообразных. Н. М. Книпович назвал изученное им существо Dendrogaster, что значит «разветвленный живот».
Организация Dendrogaster очень сильно изменена под воздействием паразитизма. Тело самки состоит главным образом из двух боковых многократно ветвящихся отростков. Эти два ответвления отходят от срединной непарной части, на самом переднем конце которой находятся передние антенны, а также сложенные в виде пирамиды жвалы и челюсти, очень сильно видоизмененные. Никаких других признаков ракообразных (сегментации, туловищных конечностей и т. д.) животное не сохранило (рис. 220, А). В ответвлениях тела самки помещаются карликовые паразитические самцы, более сходные с обычными ракообразными. Тело их сегментировано, голова снабжена такой же ротовой пирамидой, как у самок, но грудной отдел несет 5 пар двуветвистых ножек. Необычный вид самцу придает пара очень длинных задних выростов, в которые заходят ветви кишечника и семенники (рис. 220, Б).
Не все мешкогрудые видоизменены в такой степени, как самки Dendrogaster. Среди них есть и виды, сохранившие облик нормальных ракообразных, есть и менее измененные, чем Dendrogaster. Представителей этого отряда по степени упрощения организации можно расположить в определенный ряд, по которому можно судить о том, как шла эволюция мешкогрудых ракообразных в связи с приспособлением к паразитическому существованию и с переходом с одних хозяев на других.
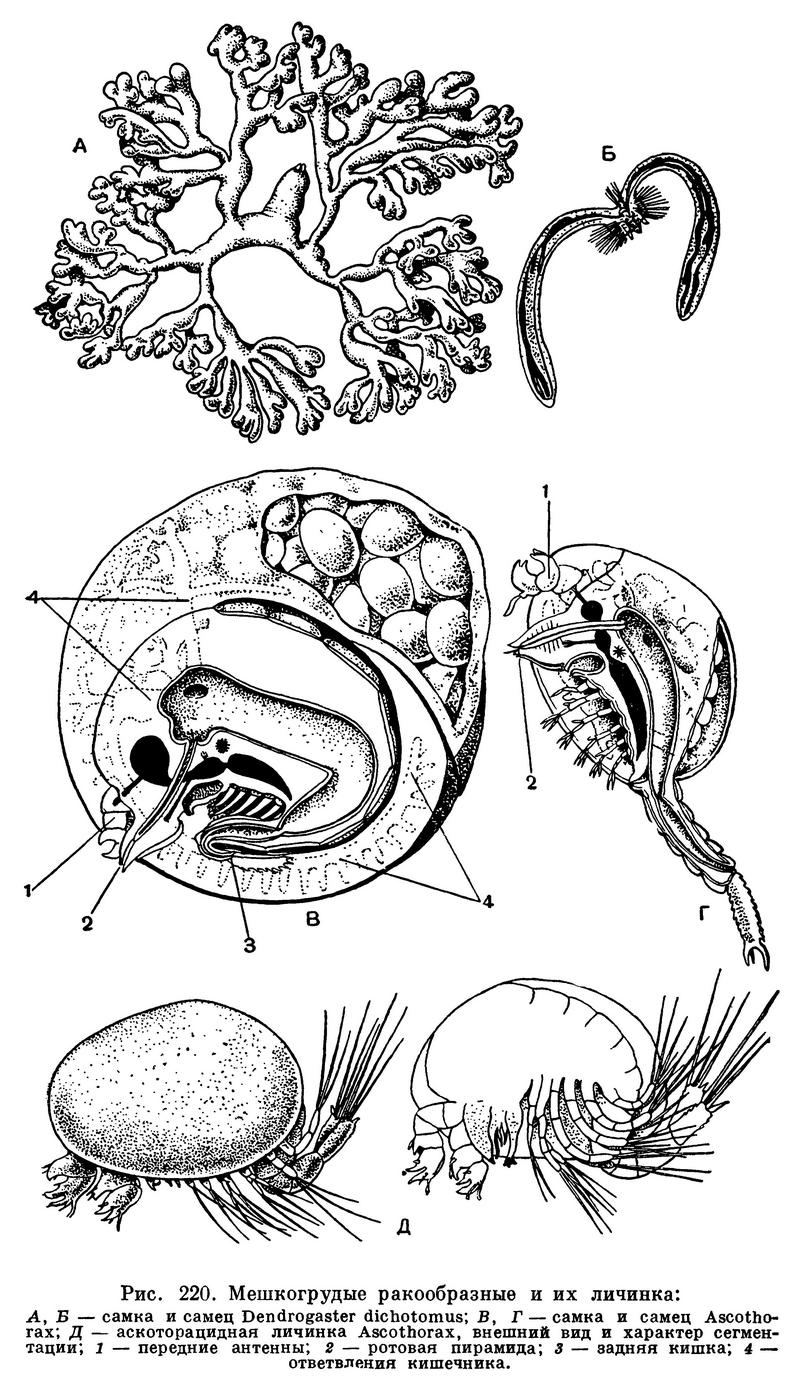
Рассмотренное раньше семейство Dendrogastridae, представители которого паразитируют в полости тела морских звезд и морских ежей, отличается от семейства Sinagogidae тем, что мантия самок чрезвычайно сильно разрастается с боков, принимая разветвленную древовидную форму или имея вид мешка, иногда раздвоенного. Собственно тело рачка помещается в самой передней части непарного срединного ствола мантии; оно не сегментировано, лишено грудных ножек или они не членистые. Маленькие самцы паразитируют в мантийной полости самок.
В несколько ином направлении эволюционировало семейство Lauriidae. Относящиеся к нему рачки паразитируют на кораллах, причем самки — эндопаразиты, сидящие в цистах под сарколеммой, а самцы — эктопаразиты тех же хозяев, внутри которых живут самки. У самок мантия разрастается вверх и вниз и превращается в толстую пластинку, внутри которой разветвляются яичники и печеночные выросты; грудные ножки в той или иной степени недоразвиты, сегментация сохранилась. Самцы приблизительно такие же, как у других мешкогрудых, значительно мельче самок.
Таким образом, при переходе мешкогрудых от эктопаразитизма к эндопаразитизму отличия между самцами и самками становились все более резкими, самки теряли сегментацию, их грудные ножки редуцировались, а мантия разрасталась. Самцы изменялись гораздо меньше самок.
Все мешкогрудые ракообразные прикрепляются к хозяину при помощи передних антенн, концевые членики которых образуют подобие клешни. Задние антенны отсутствуют. Жвалы и челюсти сложены вместе и составляют колющий хоботок. Этим хоботком эктопаразитические Sinagoga прокалывают ткани своих хозяев и сосут их соки. Ascothorax питается разрушенными им клетками стенок бурсы офиур, а паразит неправильных морских ежей — Ulophysema — поедает половые железы хозяина. Разветвленные самки Dendrogaster заглатывают клетки полости тела морской звезды, одевающие паразита сплошным слоем.
Обычно присутствие паразита вызывает кастрацию хозяина. Если Ulophysema непосредственно разрушает половые железы морских ежей, то Ascothorax и Dendrogaster подавляют действие половых желез офиур и морских звезд механически.
Специальных органов дыхания у мешкогрудых ракообразных нет. Кислород усваивается стенкой мантии. Весьма вероятно, что разветвленная мантия Dendrogaster представляет собой приспособление, позволяющее использовать наибольшее количество кислорода, растворенного в полостной жидкости морской звезды, благодаря значительному увеличению усваивающей поверхности. Молодые самки, двигая ветвями мантии, могут медленно перемещаться в полости тела хозяина.
Все мешкогрудые ракообразные раздельнополы, за исключением паразита одиночных кораллов Petrarca bathyactidis, которому свойствен гермафродитизм. Чем более специализированы роды мешкогрудых, тем сильнее выражены различия между самцами и самками. У всех внутренних паразитов самки гораздо крупнее самцов и гораздо сильнее видоизменены под воздействием паразитизма. Обычно совместно с одной самкой живет по нескольку самцов, а у Dendrogaster они паразитируют в выростах мантии самки.
Оплодотворенные самки откладывают яйца в выводковые камеры, расположенные с внутренней стороны мантии. У Dendrogaster в этих же камерах паразитируют самцы. Число откладываемых самкой яиц различно у разных родов, причем, чем глубже приспособления к паразитизму, тем плодовитость выше. Малоизмененная Sinagoga откладывает всего 50—60 яиц, Ascothorax — 250—300, а Dendrogaster — около 20 000.
Аскоторацидные личинки проникают в своих хозяев и там развиваются во взрослых рачков. Если такая личинка окажется в одиночестве, она становится самкой. Если же она прикрепилась около вполне сформировавшейся самки или на поверхности ее тела, то развивается самец.
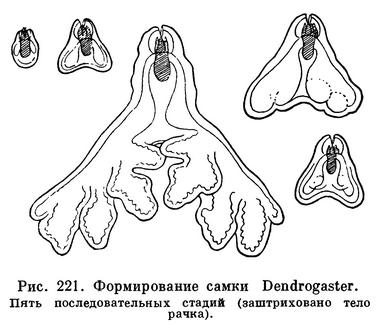
Процесс превращения аскоторацидной личинки во взрослую самку Dendrogaster весьма своеобразен. Личинка садится на поверхность тела морской звезды, линяет, теряет сегментацию и грудные ножки и приобретает грушевидную форму. После этого она проникает через стенку тела хозяина в его половые железы. Последующий рост происходит исключительно за счет разрастания мантии; собственно тело зародыша почти не увеличивается в размерах (рис. 221).
Мешкогрудые ракообразные, в особенности Sinagogidae, широко распространены в Мировом океане. Dendrogastridae обитают в умеренных и высоких широтах северного полушария, Lauriidae — преимущественно в тропиках. Они поражают не только мелководных, но и глубоководных животных. Так, например, в Беринговом море в морских звездах Psilaster pectinatus и Eremicaster tenebrarius с глубин 2300 и 3940 м найдены Dendrogaster astropectinis и Dendrogaster beringensis. Всего известно около 40 видов мешкогрудых, половина которых принадлежит к описанному впервые Н. М. Книповичем роду Dendrogaster.
ОТРЯД УСОНОГИЕ РАКООБРАЗНЫЕ (CIRRIPEDIA)
Чарлз Дарвин в своей автобиографии писал: «В октябре 1846 г. я начал работать над «Усоногими ракообразными»... я занялся изучением и анатомированием ряда обычных форм, и это постепенно привело меня к исследованию всей группы. В течение восьми лет я непрерывно работал над этим предметом и в конце концов издал два толстых тома, содержащих описание всех известных современных видов, и два тонких in quarto о вымерших видах. Не сомневаюсь, что сэр Э. Литтон-Булвер, выведя в одном из своих романов профессора Лонга, который написал два увесистых тома о ракушках, имел в виду меня».
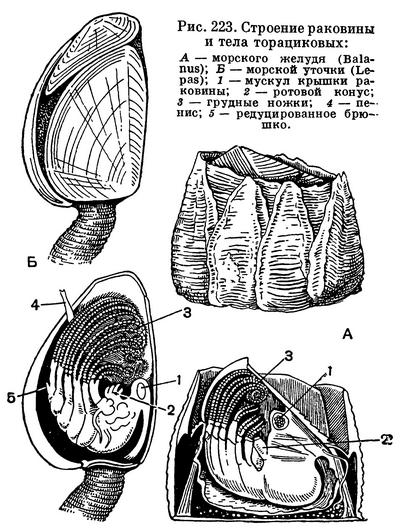
Усоногие ракообразные, которым отдал восемь лет своей жизни великий натуралист, во многих отношениях замечательны, и интерес к ним Ч. Дарвина вполне понятен. Внешне они даже непохожи на ракообразных. Во взрослом состоянии они ведут сидячий образ жизни, прикрепляясь к всевозможным подводным предметам — скалам, камням, сваям, днищам судов, а также к самым разнообразным морским животным. Тело большинства этих рачков заключено в твердую известковую раковину, состоящую из различного количества отдельных пластинок. Часть этих пластинок подвижно сочленена между собой. Рачок может их раздвигать, и в образующуюся щель просовываются время от времени грудные ножки, совершающие характерные взмахи. При этом вода с находящимися в ней организмами и их остатками загоняется внутрь раковинки; так осуществляется питание и дыхание.
Твердая известковая раковина, постоянное прикрепление к субстрату и отсутствие внешних признаков ракообразных долго мешали выяснению истинной природы усоногих. Даже такой зоолог, как Ж. Кювье, по примеру предыдущих исследователей относил усоногих к моллюскам. Ж. Б. Ламарк рассматривал усоногих как переходную группу от червей к моллюскам. Только в 1830 году, после того как стали известны личинки усоногих, сходные с личинками других ракообразных, была доказана принадлежность усоногих к этому классу.
В настоящее время в этом никто не сомневается. Больше того, нетрудно подметить многие признаки, сближающие усоногих с внешне совершенно несходными с ними веслоногими. Однако усоногие эволюционировали в ином направлении, чем подавляющее большинство остальных ракообразных. Среди непаразитических представителей этого класса только они, вместо того чтобы совершенствовать свою двигательную систему, перешли к прикрепленному существованию и развили мощное средство пассивной защиты — толстую известковую раковину, несколько напоминающую раковину моллюсков. В связи с прикрепленным образом жизни организация усоногих очень сильно изменилась: исчезли наружная сегментация и хитиновый панцирь, бесполезный при наличии защитной известковой раковины, перестроилась голова, которой рачки прикрепляются к субстрату, укоротился и почти исчез брюшной отдел. Изменилась и функция конечностей. Они перестали служить для передвижения и приспособились к тому, чтобы загонять воду в мантийную полость.
Часть усоногих ракообразных перешла от сидяче-прикрепленного образа жизни на поверхности тела различных морских животных к паразитическому существованию, сменив питание планктоном на использование соков своих хозяев. Можно наметить полную серию переходов от свободного существования этих ракообразных к паразитизму. Как и в других подобных случаях, паразитизм вызвал ряд изменений организации усоногих и эти изменения отразились на приспособлениях к прикрепленному образу жизни. Среди паразитических усоногих известны предельно упрощенные формы, утерявшие все признаки не только отряда, но даже класса ракообразных и типа членистоногих. Только изучение строения личинок позволяет понять их истинную природу.
Все усоногие — морские обитатели, причем некоторые из них могут выносить значительное опреснение. Тело их окружено двойной мантийной складкой, соответствующей карапаксу других ракообразных. Мантия непаразитических форм выделяет различное количество известковых пластинок, образующих защитную раковину. Мягкое тело подразделено на головной и грудной отделы. Передние антенны служат для прикрепления к субстрату, снабжены цементными железами и очень сильно укорочены. Задние антенны отсутствуют, ротовые придатки упрощенного строения. На грудном отделе расположено 6 пар двуветвистых ножек. У сверлящих усоногих число грудных ножек сокращено до 4 пар и они одноветвисты. У подотряда корнеголовых ракообразных, все представители которого — паразиты, ротовых придатков и грудных ножек нет совсем.
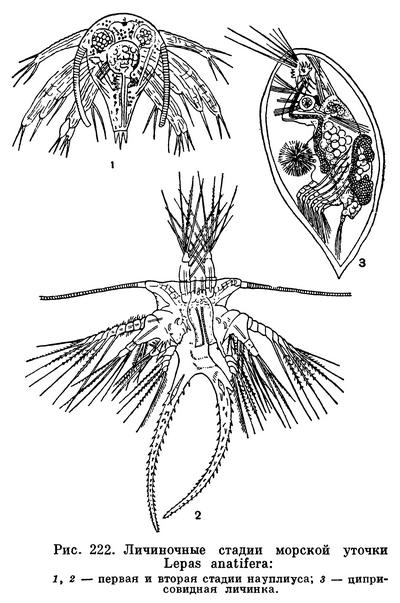
Из яиц усоногих выходит типичный науплиус, отличающийся от науплиусов других ракообразных присутствием боковых выростов на переднем конце тела, так называемых «рожек». Перелиняв несколько раз, науплиус переходит в стадию метанауплиуса, для которой характерно появление челюстей и зачатков других конечностей и начало развития фасеточных глаз по бокам от науплиального. «Рожки» у метанауплиуса обычно сохраняются, задняя часть тела часто снабжена выростами. За метанауплиальной стадией следует свойственная только усоногим стадия циприсовидной личинки, иногда называемая также «куколкой». Она обладает двустворчатой хитиновой раковиной, покрывающей все ее тело, 4-члениковыми передними антеннами с присасывательными дисками на предпоследнем членике и с цементными железами при основании, упрощенными жвалами и челюстями и 6 парами грудных двуветвистых плавательных ножек. На переднем конце личинки расположены науплиальный и пара фасеточных глаз. Присутствие двустворчатой раковины придает этой личинке внешнее сходство с представителями отряда ракушковых ракообразных, в частности с относящимся к нему родом Cypris, почему личинка и получила название циприсовидной. Циприсовидная личинка при помощи своих грудных ног свободно плавает в воде, а затем прикрепляется передними антеннами к подходящему субстрату. Дальнейшее ее превращение во взрослого рачка у разных подотрядов происходит по-разному.
В отряд усоногих включают три подотряда, резко различающихся как по строению, так и по образу жизни.
ПОДОТРЯД ТОРАЦИКОВЫЕ РАКООБРАЗНЫЕ (THORACICA)
Основу обрастаний судов составляют усоногие ракообразные — морские желуди. Они нередко покрывают днище корабля сплошным слоем. С китобойной матки длиной 140 м, плававшей в течение 7 месяцев к югу от экватора, сняли 40 т морских желудей. На наших дальневосточных судах за год плавания нарастает 10—12 кг морских желудей на 1 м2 поверхности днища.
Морские желуди и относящиеся к тому же подотряду морские уточки (табл. 32), отличающиеся присутствием стебелька, на котором расположено защищенное известковыми пластинками тело рачка, поселяются не только на кораблях. Прибрежные скалы и камни усыпаны ими, они прикрепляются к раковинам моллюсков, к панцирям ракообразных, к раковинам себе подобных, иногда к рыбам. Некоторые виды приспособились к существованию на коже китов и акул. Есть морские желуди, сидящие на скалах значительно выше уровня моря и довольствующиеся брызгами, которые долетают до них только при сильных штормах. С другой стороны, известны усоногие, обитающие на глубине до 6860 м. Всего описано около 700 видов этих ракообразных.
Весьма своеобразны способ питания и функции грудных ножек морской уточки Lepas fascicularis (табл. 32, 2), широко распространенной в тропической и субтропической частях всех океанов. Эти рачки прикрепляются к каким-нибудь плавающим предметам (кусочки дерева, пемза, водоросли) или к выделяемому ими пенистому поплавку и целыми группами плавают на самой поверхности моря. Они могут грести своими ножками и активно подплывать к сифонофорам (Velella, Porpita), которых обгрызают жвалами.
Есть морские уточки, добывающие себе пищу не прибегая к помощи грудных ножек, которые в таких случаях укорачиваются и упрощаются. Malacolepas conchicola живет в мантийной полости двустворчатых моллюсков и использует пищевые частицы, поступающие через вводной сифон хозяина. Pagurolepas conchicola селится в раковинах брюхоногих моллюсков, занятых раками-отшельниками, и питается экскрементами последних. Некоторые морские желуди прикрепляются к коже китов и в той или иной степени питаются за счет своих хозяев. Н. И. Тарасов и Г. Б. Зевина пишут: «Род Coronula не полностью перешел к питанию за счет тканей и крови китов, причем молодые особи, по-видимому, в большей степени питаются биосестоном [взвешенными в воде частицами], нежели взрослые, сросшиеся с кожей хозяина, проникшей в полости стенки их домика. Tubicinella глубоко погружена в кожу кита, а взрослых Cryptolepas невозможно механически отделить от плотно и полностью облекающей их кожи единственного хозяина этого вида — серого кита Eschrichtius gibbosus». У таких специализированных паразитов грудные ножки укорочены, а известковые пластинки домика уменьшены и частично исчезли совсем (табл. 32, 12).
Весьма своеобразны половая система и особенности размножения Thoracica. В отличие от подавляющего большинства остальных членистоногих эти рачки, за немногими исключениями, гермафродиты. Каждая особь обладает и мужскими и женскими половыми железами. Яйцеводы открываются при основании передней пары грудных ножек. Выходящие из них яйца откладываются в мантийную полость. Семепроводы впадают в длинный трубкообразный мужской совокупительный орган — пенис, представляющий собой вырост редуцированного брюшного отдела и помещающийся непосредственно позади задней пары грудных ножек. При спаривании пенис распрямляется, высовывается из раковины наружу, проникает в мантийную полость соседней особи и выделяет туда сперму, оплодотворяющую яйца. Поскольку обычно морские желуди и морские уточки сидят в непосредственной близости друг к другу, происходящий, таким образом, процесс перекрестного оплодотворения осуществляется без всяких затруднений. В некоторых случаях может происходить самооплодотворение, как это следует из опытов по изоляции отдельных особей, которые и в одиночестве продолжали размножаться.
Некоторые исследователи предполагают, что некогда все усоногие были раздельнополыми. Те виды, которые имеют мало редуцированных самцов, должны считаться наиболее древними, а те, которые совсем их лишены и представлены только гермафродитными особями,— наиболее молодыми.
Весьма интересен, хотя и до настоящего времени совершенно неясен, вопрос об условиях, определяющих развитие личинки в самца, в самку или в гермафродитную особь. По некоторым наблюдениям, в случае попадания личинки в мантийную полость гермафродитной или женской особи из нее развивается самец, а в случае ее прикрепления к неорганическому субстрату — гермафродитная или женская особь. Эта гипотеза нуждается в дополнительной проверке.
Морские желуди растут довольно быстро. В тропиках некоторые виды достигают половозрелости через 8—16 дней после оседания. Обычному в Балтийском, Черном и Азовском морях Balanus improvisus, а также дальневосточному Balanus cariosus для этого требуется 3 месяца. Продолжительность жизни разных видов колеблется от 1—2 до 5—7 и более лет.
Как уже указывалось, морские желуди и морские уточки населяют самые различные глубины. Некоторые виды хорошо приспособлены к существованию в приливо-отливной зоне. Они активны только во время прилива, а когда вода отступает, они плотно закрывают крышки домика и погружаются в состояние, сходное с анабиозом. При переносе в аквариум рачки в течение нескольких дней сохраняли тот же ритм, прекращая биение ножек в часы отлива. Однако, даже находясь вне воды, животные продолжают дышать, хотя и менее интенсивно, чем под водой. Только при наступлении резко неблагоприятных условий они впадают в состояние «скрытой жизни» и в течение нескольких месяцев сохраняют жизнеспособность в сухом состоянии, без вреда для себя подвергаются длительному действию пресной воды, а в условиях опыта переносят заливание вазелиновым маслом, содержание в азоте и водороде, т. е. продолжают существовать при полном отсутствии кислорода.
Некоторые морские уточки прикрепляются не к дну, а к плавающим в воде предметам — кускам дерева, судам, а также к различным животным. Таковы виды родов Lepas и Conchoderma. Обычный широко распространенный вид Conchoderma auritum встречается и на кусках дерева, и на днищах судов, и на китах, морских черепахах и медленно плавающих рыбах. При этом Conchoderma auritum всегда садится только на твердую поверхность. На китах эта морская уточка находится только на прикрепившихся к их коже морских желудях Coronula (табл. 32, 5) и на китовом усе, а у кашалотов — на зубах, образуя целые гроздья. Уховидные придатки рачка, служащие, вероятно, для отвода профильтрованной воды, сильнее развиты у особей, сидящих на быстро движущихся животных.
Практическое значение Thoracica определяется прежде всего тем, что морские желуди составляют основу обрастаний судов. Как уже указывалось, обросшее морскими желудями (и другими животными) судно теряет исходную скорость. Соответственно увеличиваются время рейса, затраты на топливо и все остальные эксплуатационные расходы. Корабли периодически приходится очищать от обрастаний, а это обходится очень дорого.
Подсчитано, что торговый флот США ежегодно терпит убытки от обрастаний, превосходящие 100 миллионов долларов.
Во многих странах усиленно работают над проблемой защиты судов от обрастаний, в частности над изобретением таких красок для днищ кораблей, которые были бы ядовиты для оседающих на суда личинок морских желудей. Некоторые успехи в этом направлении достигнуты, но «необрастающие» краски очень дороги.
Морские желуди вредят не только судам. Проникнув в Каспийское море, они стали оседать на ставных сетях, неводах и других рыболовных орудиях и принесли немало неприятностей местным рыбакам, которые раньше с ними не сталкивались.
С другой стороны, несомненно и положительное значение морских желудей и морских уточек. Их личинки нередко составляют существенную часть прибрежного морского планктона и в большом количестве поедаются некоторыми планктоноядными рыбами. Так, например, у берегов Норвегии весной сайда питается преимущественно личинками усоногих, поедает их и беломорская сельдь, а также некоторые другие рыбы. Взрослые усоногие в малой степени используются рыбами, которые часто не могут оторвать их от субстрата и раскусить. Тем не менее морские желуди отмечены в качестве объектов питания трески, пикши, зубатки, осетра и др.
Человек также не брезгает усоногими. Из крупного морского желудя Balanus psittaceus в Чили делают консервы. В Испании и Португалии добывают морскую уточку Mitella pollicipes, которая используется и в пищу, и для приготовления тука. Многие другие усоногие также, несомненно, съедобны. Тур Xейердал, описывая свое замечательное путешествие на «Кон-Тики», рассказывает, между прочим, о том, как он и его спутники варили суп из морских желудей, которыми быстро обрастал их плот, и высоко оценивает вкусовые качества этих рачков. Весьма возможно, что в будущем человек станет использовать усоногих в большей степени, чем ныне.
ПОДОТРЯД АКРОТОРАЦИКОВЫЕ РАКООБРАЗНЫЕ (ACROTHORACICA)
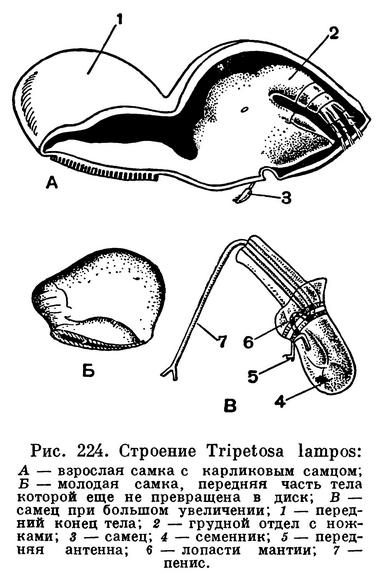
Занимаясь усоногими ракообразными, Ч. Дарвин открыл один сильно отличающийся по своему строению и образу жизни от морских желудей и морских уточек вид, сверлящий раковины моллюсков, который он считал необходимым выделить в особый подотряд. В настоящее время таких видов известно более десятка. Они точат ходы в раковинах брюхоногих и панцирных моллюсков, а также в стволах кораллов (рис. 224).
Будучи хорошо защищены известковой толщей раковины моллюска или ствола коралла, они, естественно, лишены собственных известковых пластинок, свойственных их только что рассмотренным родичам. Только на передней части головы самки есть хитиновая пластинка, принимающая, вероятно, участие в процессе сверления. Эта передняя часть головы перетяжкой отделена от задней части и от грудного отдела. Вокруг ротового отверстия расположены пара жвал и две пары челюстей, а позади рта пара передних грудных ножек. На значительном расстоянии от них находятся собранные в одну группу остальные три пары одноветвистых грудных ножек. Грудные ножки совершают около 60 взмахов в минуту, но не высовываются при этом за края мантии, как это бывает у морских желудей и морских уточек. Тем не менее благодаря взмахам грудных ножек вода внутри мантийной полости рачка все время сменяется и приносит достаточное количество мелких пищевых частиц. Они поступают в кишечник, лишенный заднего отдела и анального отверстия, но зато снабженный многочисленными ветвящимися слепыми выростами, заходящими в голову, в грудные ножки и в другие части тела.
Все Acrothoracica, за исключением живущего в кораллах рода Lithoglyptes, раздельнополы. Карликовые (0,4—2 мм) самцы живут поодиночке, прицепившись к мантии самки, длина которой достигает 2 см. Они обычно сохраняют облик циприсовидной личинки или имеют очень упрощенное строение. Из оплодотворенного яйца выходит науплиус, такой же, как у Thoracica. Перелиняв три раза, он превращается в метанауплиуса, который дает начало типичной циприсовидной личинке. Циприсовидная личинка садится на подходящий субстрат, сбрасывает свою раковину и превращается в сформировавшегося рачка. Рачок приступает к сверлению известняка, пользуясь хитиновой пластинкой и выделяя вещество, растворяющее соли кальция.
Обыкновенно Acrothoracica селятся в пустых раковинах уже погибших брюхоногих моллюсков (Buccinum, Fusus и др.) или в раковинах, занятых раками-отшельниками.
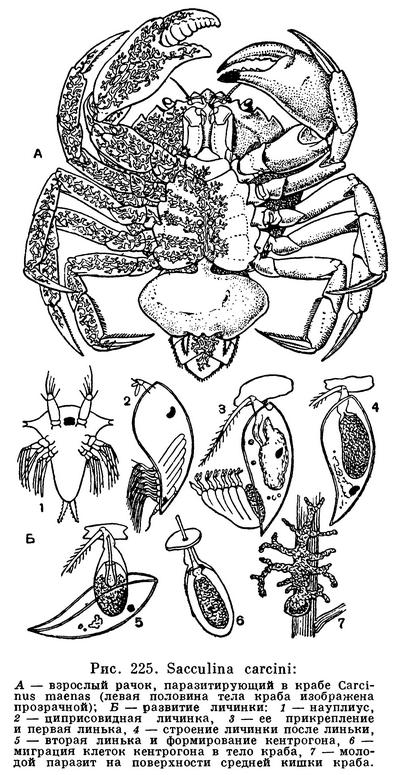
ПОДОТРЯД КОРНЕГОЛОВЫЕ РАКООБРАЗНЫЕ (RHIZOCEPHALA)
Еще больше отличаются от обычных усоногих ракообразных паразитические корнеголовые, организация которых предельно упрощена под воздействием паразитизма. Взрослые представители этого подотряда не сохраняют совершенно никаких признаков членистоногих животных. Они живут в десятиногих ракообразных — в крабах, раках-отшельниках и некоторых креветках.
Все тело краба, зараженного корнеголовым рачком Sacculina, пронизано ветвящимися, как корни растений, отростками паразита. Они оплетают все внутренние органы хозяина — кишечник, половые железы, нервную систему, внедряются в мускулатуру. Этими отростками рачки высасывают соки краба. Отростки впадают в обширный мешок, расположенный под брюхом хозяина на поверхности его тела. В мешке находятся сильно развитые яичники и маленькие семенники. Все остальные органы — кишечник, органы чувств и органы выделения — полностью отсутствуют.
Систематическое положение этого существа, так же как и в других подобных случаях (например, описанного выше дендрогастера), выясняется при изучении его развития (рис. 225, Б). Из яиц, откладываемых в полость мешка, выходит типичный для усоногих науплиус, выбирающийся наружу и плавающий в воде. Через 8 дней он превращается в метанауплиуса, который после 4—5 линек переходит в столь же типичную стадию циприсовидной личинки. Все эти личиночные стадии корнеголовых отличаются от соответствующих им стадий других усоногих главным образом отсутствием кишечника. Они питаются за счет содержащихся в их теле запасов желтка. Проплавав 3—4 дня, циприсовидная личинка прикрепляется передними антеннами к основанию щетинок конечностей или брюшной стороны тела молодых крабов. После этого она испытывает удивительные превращения. При очередной линьке она теряет большую часть своего тела со всеми конечностями, но сохраняет мантию, раковину и глаз, а затем линяет еще раз и отбрасывает и их. От личинки остается маленькое овальное мешковидное тельце, внутри которого располагаются недифференцированные клетки, имевшиеся еще у науплиуса. Эта стадия получила название стадии кентрогона. Покровы и передние антенны кентрогона образуют пронизанный каналом вырост, пробуравливающий покровы краба. Через этот канал кучка недифференцированных клеток проникает в тело краба и, вероятно, кровью хозяина переносится на поверхность его средней кишки.
Здесь начинается рост паразита. Отростки его тела удлиняются, разветвляются и начинают оплетать внутренние органы краба. Они не разрушают ни кишечника, ни половых желез хозяина, всасывая питательные вещества из омывающей их крови и выделяя в организм хозяина отбросы своей жизнедеятельности. Через 7—8 месяцев паразит образует мешковидный вырост. Когда он достигнет 2 мм в поперечнике, он прорывает покровы хозяина и выпячивается наружу, а затем продолжает расти в течение 6 недель, увеличиваясь до 12 мм в диаметре. К этому времени в мешковидном выросте заканчивается созревание половых желез, которые начинают функционировать. Кроме того, здесь же формируется очень слабо развитая нервная система. Всего для развития взрослой саккулины требуется, таким образом, 9 месяцев с момента прикрепления к хозяину циприсовидной личинки. Паразит живет около 3 лет, и хозяин обычно переживает его и продолжает существовать после смерти саккулины.
Присутствие саккулины оказывает мощное воздействие на организм краба. Заражение саккулиной молодого самца подавляет развитие его половых желез. С каждой последующей линькой он становится все более похожим на самку: брюшной отдел его расширяется, исчезают превращенные в совокупительный орган передние брюшные конечности, меняется характер щетинок брюшка. Японские зоологи Ишикава и Янагимачи описали зараженного саккулиной самца краба Eriocheir japonicus, семенники которого под влиянием паразита переродились в яичники, продуцировавшие нормальные яйца. Если саккулина заразила взрослого крупного самца или самку, это не приводит к изменению вторичнополовых признаков хозяина, но нарушает деятельность его половых желез — происходит паразитарная кастрация.
Нет сомнения в том, что причиной таких изменений в организме краба служит воздействие саккулины на его органы внутренней секреции. Непосредственно или выделяя специфические вещества, паразит подавляет выделение гормонов, действующих на формирование вторичнополовых признаков самца и контролирующих деятельность половых органов самца и самки.
Паразиты раков-отшельников Peltogaster и Peltogastrella также вызывают паразитарную кастрацию своих хозяев, но заражение ими самцов не приводит к развитию у них вторичнополовых признаков самок. В отличие от саккулины у этих паразитов есть дополнительные самцы, сохраняющие облик циприсовидных личинок. Семенники гермафродитных особей очень малы и не функционируют. Самцы прикрепляются передними антеннами к мешковидной части гермафродитных особей, и масса заполняющих их недифференцированных клеток переселяется в недоразвитый семенник гермафродита. Часть этих клеток преобразуется в сперматозоиды.
При линьке раков-отшельников наружные мешковидные части их паразитов отваливаются вместе с покровами хозяина. Однако это не приводит к гибели паразита: из корневидной, находящейся внутри рака-отшельника части паразита вновь отрастает его мешковидная часть. У Chologaster наблюдалось четырехкратное отпадение мешков и замена их новыми.
Способность к почкованию особенно ярко проявляется у Thompsonia — паразита креветок из семейства раков-щелкунов. На теле зараженной креветки всегда находится большое количество мешковидных паразитов, объединенных общей корневой системой. Она возникает из одной личинки, а затем образует много мешковидных особей.
Корнеголовые ракообразные широко распространены во всех морях и океанах, за исключением холодных южных и северных морей. Есть они и в Черном море, и в наших дальневосточных морях. В Северной Атлантике, Средиземном и Черном морях около 6 % представителей массовых видов крабов заражено саккулиной. В тропиках некоторые корнеголовые вместе со своими хозяевами обитают и в пресных водах. Всего известно около 120 видов этих удивительных ракообразных.
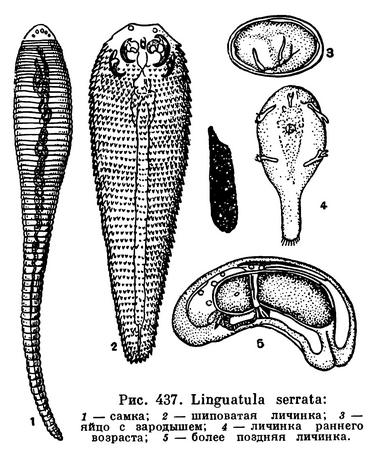
КЛАСС ЯЗЫЧКОВЫЕ, ИЛИ ПЯТИУСТКИ (LINGUATULIDA, или PENTASTOMIDA)
Язычковые очень своеобразная группа паразитических животных, неясная по систематическому положению. Во взрослом состоянии язычковые паразитируют в дыхательных путях и в легких млекопитающих и рептилий, реже птиц, причем, как правило, сменяют животных-хозяев в течение жизненного цикла. Наиболее известна носовая пятиустка (Linguatula serrata, синоним Linguatula rhinaria), во взрослом состоянии паразитирующая в носовой полости и лобных пазухах некоторых хищных и травоядных животных.
Тело взрослой пятиустки вытянутое, червеобразное, суженное в задней половине. Самка крупная, 8—13 см длиной, самец много мельче, всего до 2 мм. Тело кольчатое, но лишь внешне, во внутреннем строении сегментации не обнаруживается. Тело покрыто тонкой кутикулой. У переднего конца вентрально расположено ротовое отверстие, на заднем конце — анальное. Близ рта имеются две пары придатков, обычно трактуемые как видоизмененные конечности. В большинстве случаев это две пары 2-члениковых хитинизированных крючков. У некоторых тропических видов эти крючки сидят на вершине мускулистых бугорков, причем у личинок этих видов пара таких же бугорков с крючками имеется еще и на заднем конце тела. Первоначально околоротовые крючья принимали за ротовые отверстия, откуда и пошло неправильное, но укоренившееся в литературе название «пятиустки».
Внутреннее строение, как и внешнее, упрощено в связи с паразитизмом. Кишечник прямой, трубчатый. Дыхательная, кровеносная и выделительная системы отсутствуют. Нервная система сильно концентрирована, надглоточный ганглий слабо развит, все узлы нервной цепочки слиты в подглоточное нервное скопление. Глаз нет. Мускулатура поперечнополосатая. Язычковые раздельнополы. Половое отверстие у самки открывается в задней части тела, у самца — близ ротового отверстия. У самок имеется длинный трубчатый яичник, два яйцевода и длинное влагалище с парой семеприемников; у самца — семенники, семенной резервуар, два семепровода и парный копулятивный орган.
Взрослая носовая пятиустка паразитирует в носовой полости и лобных пазухах собак, волков, лисиц, реже встречается у лошадей, рогатого скота, овец и других животных, в организм которых попадает, по-видимому, случайно. Мелкие яйца пятиустки (0,09 мм) выходят из основного хозяина с носовой слизью, рассеиваются повсюду и попадают на растительность. Яйца проглатываются с растениями травоядными животными — промежуточными хозяевами паразита. Последними чаще всего служат зайцы и кролики. В желудке этих животных из яйца выходит крошечная личинка. Ее короткое тело сзади сужено, по бокам имеются две пары мускулистых придатков с крючковидными коготками, на переднем конце расположены три хитиновых стилета, которые считают сверлящим аппаратом. Личинка проходит сквозь стенку кишечника, странствует в теле хозяина и проникает в печень, брыжейку и иные органы; здесь она инкапсулируется и становится неподвижной. В таком состоянии личинка находится несколько месяцев, растет, линяет и достигает 4—6 мм длины. Теперь она имеет две пары околоротовых крючьев, кутикула несет поперечные ряды мелких шипиков. Личинка становится похожа на взрослую пятиустку (самку), но мельче и еще лишена органов размножения. Такая шиповатая личинка была первоначально описана как особый вид пятиустки (Pentastomum denticulatum). Шиповатая личинка странствует в теле животного и может снова инкапсулироваться в различных органах. Иногда она проникает в носовую полость и может выпасть со слизью наружу или, пробравшись в кишечник, выводится наружу с испражнениями. Обычно такие личинки гибнут, но иногда, по-видимому, могут попадать в окончательного хозяина — хищника, будучи втянуты в носовую полость, например, при обнюхивании следа. В некоторых случаях шиповатая личинка, проникнув в дыхательные пути промежуточного хозяина, не выпадает наружу, а развивается во взрослую пятиустку. Обычно же превращение личинки во взрослую пятиустку происходит в окончательном хозяине — хищнике, который заражается, съедая промежуточного хозяина. Собака, волк или лисица, в носовой полости которых живут пятиустки, рассеивают их яйца, зайцы, кролики заражаются личинками, хищник, съедая зайца или кролика, получает пятиустку. Такой же «обмен» другими, специфическими, видами язычковых происходит между некоторыми змеями и грызунами, которыми питаются змеи. Известно несколько случаев заражения человека носовой пятиусткой, по-видимому, от собак.
Описано более 70 видов язычковых, объединяемых в 15 родов и 2 большие группы — Cephalobaenida и Porocephalida, которые обычно считают отрядами. Большинство видов имеет основными хозяевами крупных рептилий — змей, крокодилов и др. и распространено в тропических странах. Личинки встречаются у различных млекопитающих, а также у рыб, служащих пищей основным хозяевам. Некоторые виды паразитируют у птиц, например Reighardia sternae у чаек в Арктике. В Африке, Южной Америке, Индии у крокодилов паразитируют язычковые родов Sebekia, Leiperia, Diesingia и др., а их личинки — в рыбах. В трахеях и легких питонов и других больших тропических змей поселяются язычковые родов Porocephalus, Kiricephalus, Armillifer и др. Промежуточными хозяевами для личинок служат мелкие млекопитающие, из более крупных — антилопы, жирафы, обезьяны, а также человек. Если человек проглатывает яйца этих паразитов, то их личинки, видимо, не причиняют ему какого-либо заметного вреда. Но при съедании сырого мяса с инкапсулированными личинками более позднего возраста последние проникают сквозь стенку кишечника, внедряются в печень, легкие и другие органы, в результате чего развиваются перитониты, пневмония с тяжелым, даже смертельным исходом.
Виды родов Linguatula — паразиты млекопитающих, распространены они в Евразии, Африке, Америке, Австралии. Упоминавшаяся носовая пятиустка (Linguatula serrata), имея хозяином собаку, всюду следует за человеком. Интересно, что у собаки динго, индийской по происхождению, паразитирует особый вид пятиустки (Linguatula dingophila), причем предполагают, что этот вид дифференцировался как самостоятельный после проникновения собаки в Австралию вместе с человеком четвертичного периода.
Происхождение язычковых не выяснено. Иногда их исключают из типа членистоногих и рассматривают как крайне изменившихся в связи с паразитизмом кольчатых червей. Однако ряд признаков говорит о принадлежности язычковых к Arthropoda: хитинизированная кутикула, отсутствие ресничек, поперечнополосатая мускулатура и др. Правда, положение язычковых среди членистоногих неясно. Попытки сблизить их с хелицеровыми лишены достаточных оснований. Сходство язычковых с некоторыми эндопаразитическими клещами (железницы и др.), на которое иногда указывают, носит чисто внешний характер.