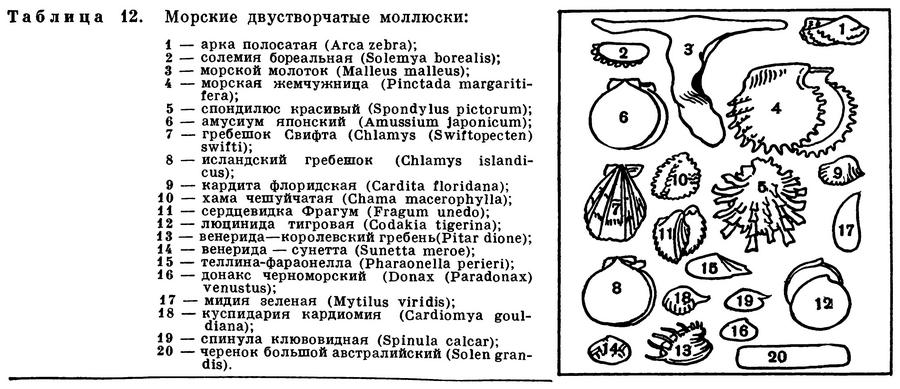
(Bivalvia Linnaeus, 1758) Класс Бивалвии, Двустворчатые, Двустворчатые моллюски, Безголовые, Безголовые моллюски, Пластинчатожаберные, Пластинчатожаберные моллюски, Топороногие, Топороногие моллюски, Class Bivalvia Linnaeus, 1758 (Bivalves) 11 отрядов
Синонимы:
Acephala Cuvier, 1798
Asiphonidae Choffat, 1886
Conchifera Lamarck, 1818
Lamellibranchia Blainville, 1834
Pelecypoda Goldfuss, 1820
Жизнь животных. Том 2. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
КЛАСС ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ (BIVALVIA)
Класс двустворчатых моллюсков имеет, как известно, четыре различных названия, каждое из которых в какой-то мере отражает главнейшие черты их строения. Название «двустворчатые» (Bivalvia) было впервые предложено Линнеем (1758) и является наиболее правильным, поскольку оно применимо ко всем представителям этого класса. Безголовыми (Acephala) они были названы Линком (1807), что отразило факт редукции у них головного отдела тела по мере развития у них в процессе эволюции двух створок раковины и смыкания этих створок вокруг тела моллюска. Третье название — «пластинчатожаберные» (Lamellibranchia), предложенное Блэнвилем в 1814 г., может быть полностью применимо лишь к одному отряду данного класса, поскольку остальные отряды имеют жабры иного строения; таким образом, это название неприменимо, как и четвертое — «топороногие» (Pelecypoda, Гольдфус, 1880), так как строение ноги у двустворчатых моллюсков весьма разнообразно. Таким образом, наиболее правильным и всеобъемлющим является первое, линнеевское наименование, которое и следует сохранить применительно к этому классу. (В разделе «Двустворчатые моллюски» читатель найдет после наименования рода некоторых моллюсков латинские названия, заключенные в скобки. Это названия подродов, которые приобрели широкое распространение в систематике моллюсков.)
Двустворчатые широко распространены в Мировом океане и его краевых морях, в реках и озерах и даже в прудах.
Общее количество видов двустворчатых составляет около пятнадцати тысяч, и большая их часть связана в своем обитании с солеными морскими водами, и лишь около одной пятой общего числа их видов населяет пресные воды. На суше двустворчатые моллюски не встречаются.
В морских водах они распространены чрезвычайно широко, встречаясь во всех климатических зонах, от теплых вод тропических морей до Арктики и Антарктики и до холодных глубин океанической абиссали. Они населяют практически все глубины Мирового океана — от приливо-отливной зоны (литорали) и прибрежных мелководий вплоть до огромных глубин впадин Мирового океана, где они были найдены на глубине почти 10,8 км.
Особенно изобилуют двустворчатые моллюски в прибрежных мелководных районах морей (до глубины 100—200 м), где они по своей биомассе (весу на 1 м2 площади дна) и по плотности поселений составляют часто большую часть всей обитающей здесь донной фауны.
В настоящее время количество видов глубоководных двустворчатых моллюсков, обитающих в абиссали Мирового океана (т. е. на глубине более 2000 м), судя по неполным еще данным, составляет около 400 видов, но и это число следует считать сильно преуменьшенным.
Велико разнообразие размеров, строения и окраски раковин двустворчатых моллюсков. Так, гигант среди моллюсков вообще, обитатель тропических морей тридакна может достигать 200 кг веса, а длина ее мощной раковины 1,4 м. Наряду с этим размеры ряда обычных глубоководных моллюсков не превышают 2—3 мм.
Пестрыми узорами и яркими красками блещут раковины и края мантии многих тропических мелководных моллюсков, незаметных среди зарослей водорослей, белых, розоватых, лиловых и желтоватых кораллов, ярких звезд и других беспозвоночных. Разнообразные выросты, шипы, чешуйки и ребра украшают створки этих моллюсков, что помогает им укрепляться в этих зарослях и противостоять действию морских волн и течений.
Более скромную окраску имеют раковины моллюсков, живущих на песчаных или илистых грунтах умеренной или арктической области.
Глубоководные формы имеют, как правило, бледно окрашенную раковину, часто очень тонкую и полупрозрачную.
Скромно окрашена в зеленоватые и коричневатые тона большая часть пресноводных форм.
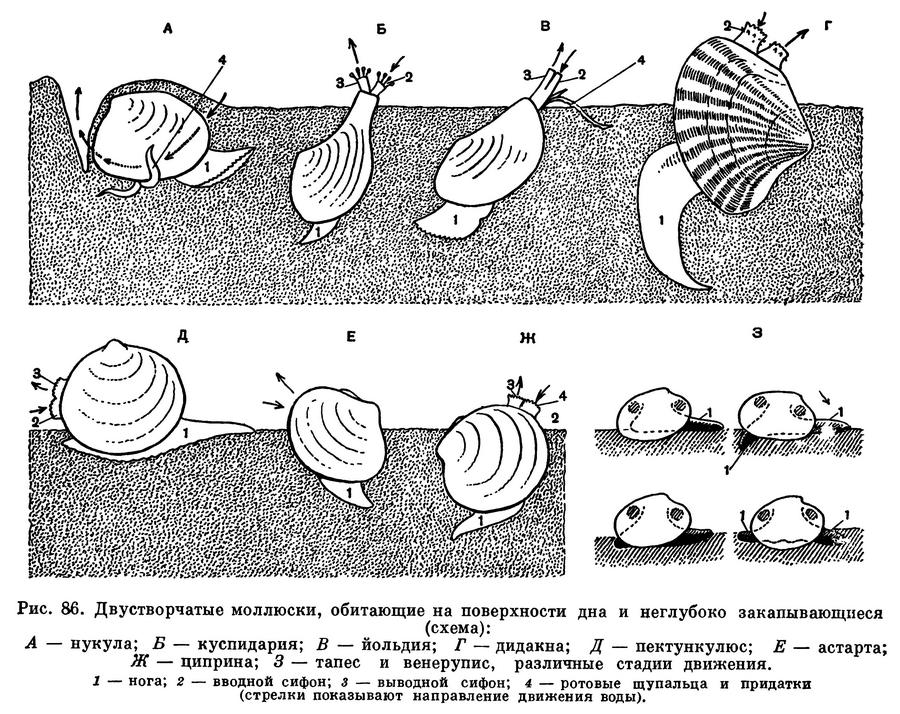
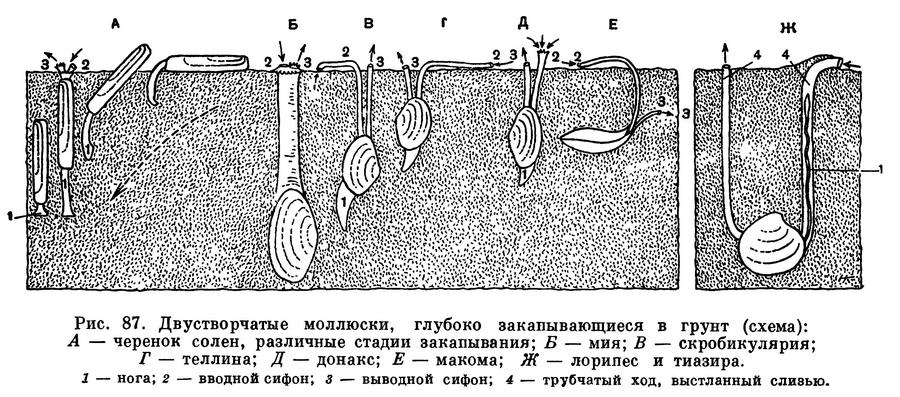
Большое разнообразие строения тела и раковины двустворчатых моллюсков тесно связано с образом их жизни, местом их обитания, с глубиной и с качеством грунта, на котором они живут, или прикрепляются, или в который закапываются. Это сказывается прежде всего на строении их раковины; на наличии на ней ребер с тем или иным вооружением ее так называемого замка, при помощи которого скрепляются створки; на наличии или отсутствии сифонов — особых выростов мантии (двух мягких лопастей, окружающих тело моллюска и выделяющих его раковину); на форме и размерах ноги и наличии в ней особой железы, выделяющей нити так называемого биссуса, при помощи которого они могут прикрепляться к грунту, а также и на многом другом. У форм, закапывающихся более или менее глубоко в мягкий грунт, сзади развиваются особые выросты мантии — сифоны, через которые засасывается и выводится вода, необходимая для дыхания и питания моллюска, погруженного в грунт. Таковы различные макомы, теллины, йольдии и др. Моллюски, живущие на поверхности грунта, ползающие или слегка закапывающиеся в него, имеют лишь зачаточные сифоны или вовсе лишены их (например, сердцевидки, венусы, астарты и др.). Моллюски, обитающие в прибрежных мелководных районах на более жестких песчаных грунтах с примесью камней, имеют более крепкие, толстые раковины (например, арки, скафарки, морские гребешки — пектены и хламисы), а различные обитатели мягких илистых грунтов имеют более тонкие раковины (батиарки, морские гребешки пропеамуссиумы и др.).
Многие формы, живущие на прибрежных мелководьях, прикрепляются нитями биссуса к камням, скалам, друг к другу, нередко образуя целые гроздья, сростки (многие мидии), или даже прирастают своими створками к камням, или срастаются друг с другом (устрицы).
Очень крепкой раковиной с острыми зубчиками на ребрах обладают многие моллюски-камнеточцы; некоторые из них выделяют особый кислый секрет, растворяющий известь прибрежных скал и камней, в которых они протачивают себе норки. Мягкое червеобразное тело древоточца тередо (Teredo) прикрыто лишь спереди маленькой сложной раковиной, служащей ему для сверления, а не для защиты тела; проводя всю жизнь в прогрызаемых в древесине ходах, эти моллюски не нуждаются в защите своего слабого длинного тела раковиной. Наконец, огромное разнообразие различных тропических моллюсков, постоянных обитателей коралловых рифов, тесно связано с их жизнью в мелководной, крайне разнообразной по характеру субстрата зоне.
Крепкие известковые раковины двустворчатых, как и других моллюсков, хорошо сохраняются в отложениях (глинах и песках) в течение целых геологических эпох. Остатки их поселений чрезвычайно ценны для геологов и палеонтологов. Эти остатки могут прекрасно характеризовать не только гидрологические и климатические условия, при которых эти отложения образовались (т. е. при которых жили найденные здесь виды моллюсков), но и возраст данной толщи отложений. Так, скопления ископаемых раковин ныне живущего в арктических морях холодноводного моллюска портландии арктической (Portlandia arctica) в отложениях севера Европы прекрасно указывают на то, что эти районы были ранее заняты холодными, слегка распресненными водами мелководного так называемого Йольдиева моря. Это море с холодноводной фауной, где руководящую роль играла арктическая портландия, было связано с периодом похолодания в послеледниковое время (примерно около 8—10 тыс. лет назад). И наоборот, отложения теплого Литторинового моря, образовавшегося позднее (3—5 тыс. лет назад), характеризуются присутствием остатков совсем других, тепловодных моллюсков, как например исландской циприны (Cyprina islandica), съедобной сердцевидки (Cerastoderma edulе), цирфеи гребенчатой (Zirfaea crispata) и др. Эти виды живут теперь лишь в Северной Атлантике, в наиболее теплых районах Баренцева и отчасти Белого морей, тогда как в эпоху Литторинового моря они продвигались дальше на север.
Представители класса двустворчатых моллюсков впервые появляются в отложениях в палеозое, т. е. в древнейших отложениях нашей планеты, а именно в верхнекембрийских слоях, образование которых насчитывает около 450—500 млн. лет. Найденные здесь первые двустворчатые моллюски относились к четырем родам, из которых такие, как Ctenodonta и Paleoneilo, имели гребенчатый замок и внешне напоминали современных ореховидок (Nuculidae) и маллеций (Malletiidae) из отряда гребенчатозубых (Taxodonta). Наибольшего видового разнообразия двустворчатые моллюски достигали в меловое время, т. е. за 100—130 млн. лет до нашего времени.
Таким образом, класс двустворчатых моллюсков является одной из самых древних групп донных беспозвоночных.
С давних времен многие двустворчатые моллюски используются человеком, они служили и служат предметом добычи. Их раковины постоянно встречаются в так называемых «кухонных кучах» доисторического человека, жившего у берегов морей, рек, озер. В раскопках стоянок человека палеолита в Крыму неизменно находят большое количество раковин устриц, мидий, гребешков и других моллюсков, которых промышляют и в настоящее время. Двустворчатых моллюсков добывают из-за их вкусного, очень полезного и легко усвояемого организмом человека мяса (как, например, устриц, мидий, морских гребешков, петушков-тапесов и венеруписов, мактр, песчаную ракушку, сердцевидок, арок, морских черенков и синовакул, пресноводных перловиц, лампсилин, беззубок, корбикул и др.).
По калорийности они могут даже превосходить мясо многих рыб, как морских, так и пресноводных. Пищевая ценность мяса моллюсков обусловливается также большим содержанием в них витаминов А, В, С, D и др., высоким содержанием таких редких в обычной пище человека минеральных веществ, как йод, железо, цинк, медь и др. Последние, как известно, входя в состав ряда ферментов, гормонов, играют исключительно большую роль в окислительном, углеводном и белковом обмене веществ, в регуляции гормональной деятельности. Мясо и раковины моллюсков широко используют для изготовления кормовой муки, идущей на откорм домашней птицы, а также для изготовления удобрительных туков.
В последние десятилетия, ввиду того что естественные природные запасы наиболее ценных съедобных моллюсков (даже в морях) истощены, а спрос на них продолжает увеличиваться, во многих странах их стали переселять в новые районы, акклиматизировать, а также и разводить искусственно как в морских, так и в пресных водах, на «фермах» — специально подготовленных отмелях и в охраняемых от хищников небольших бухтах, искусственных водоемах. Успешно разводят и культивируют не только морских моллюсков (устриц, мидий, морских петушков, тапесов), но и пресноводных (лампсилин).
В настоящее время значительно более половины добываемых двустворчатых моллюсков получают в результате их искусственного разведения. Вылов моллюсков в местах их естественного обитания в водоемах и их искусственное разведение стали теперь в ряде стран доходной и важной частью пищевой индустрии.
Двустворчатых моллюсков теперь нередко добывают на больших судах специально сконструированными орудиями лова; широко используется отлов моллюсков водолазами. Моллюски поступают на рынок не только в свежем и сушеном виде, но особенно в мороженом; сильно развилось также приготовление различных консервов из моллюсков.
Добыча двустворчатых моллюсков в последние десятилетия резко возросла. Если перед началом мировой войны ежегодная добыча их составляла около 5 млн. ц, то уже в 1962 г. она возросла примерно до 17 млн. ц и стала составлять около 50% мировой добычи всех морских беспозвоночных, или 4 % от общей мировой добычи (426 млн. ц) всех продуктов моря (рыбы, китов, беспозвоночных, водорослей).
Наибольшее количество (около 90%) двустворчатых моллюсков добывается в северном полушарии — в Тихом и в Атлантическом океанах. Промысел пресноводных двустворчатых моллюсков дает лишь несколько процентов от общей их мировой добычи. Особенно большое значение имеет промысел двустворчатых моллюсков в таких странах, как Япония, США, Корея, Китай, Индонезия, Филиппинские острова и другие острова Тихого океана. Так, в Японии добывается около 90 видов двустворчатых моллюсков, из которых около двух десятков видов имеют большое промысловое значение, а 10 видов разводятся искусственно. В европейских странах промысел и разведение двустворчатых моллюсков наиболее развиты во Франции, Италии и др.
В СССР промысловое значение имеют главным образом крупный приморский гребешок (Pecten (Patinopecten) yessoensis), а также различные мидии, белая ракушка (Spisula sachalinensis), песчаная ракушка (Mya (Arenomya) arenaria), петушки (Tapes, Venerupis) и некоторые другие.
Двустворчатые моллюски издавна добывались и ради их раковин, дающих не только прекрасное сырье для перламутровых изделий (многие пресноводные перловицы и жемчужницы, морские жемчужницы — пинктады, птерии и др.), но и ценнейший жемчуг. В начале текущего столетия были найдены промышленные способы более быстрым искусственным путем получать жемчуг (находки которого являются в природных условиях довольно редкой случайностью), не отличимый от жемчуга, образовавшегося естественным путем. Фермы для содержания морских жемчужниц и выращивания в них жемчуга особенно успешно работают в Японии. Так, уже в 1936 г. здесь было выращено 140 тыс. раковин морских жемчужниц и получено 26,5 тыс. жемчужин.
В некоторых странах, особенно в тропической части Тихого океана, добываемые раковины двустворчатых моллюсков широко используются для получения извести.
Почти все двустворчатые моллюски, за исключением крупных форм с крепкой, толстой раковиной, служат излюбленной пищей придонных рыб — бентофагов (т. е. питающихся донными животными), в том числе и многих промысловых рыб, как морских, так и пресноводных: камбал, некоторых тресковых (пикши), осетровых, многих карповых (лещ, сазан), зубаток, бычков и др. Некоторые рыбы благодаря преобладанию в их пище мелких моллюсков получили название «моллюскоедов», как, например, каспийская вобла. Районы, где наряду с другими донными животными (многощетинковыми червями, офиурами и др.) наблюдается массовое развитие мелких двустворчатых моллюсков, служат местами откорма различных придонных промысловых рыб.
Моллюсков охотно поедают многие крупные десятиногие раки (омары, раки-отшельники, крабы), морские звезды — исконные враги двустворчатых моллюсков. Промысловые устричные банки периодически очищают от морских звезд при помощи особых швабр, которые волокут по дну небольшие суда.
Немалую роль играют двустворчатые в питании промысловых камчатских «крабов» (Paralithodes kamtschatica).
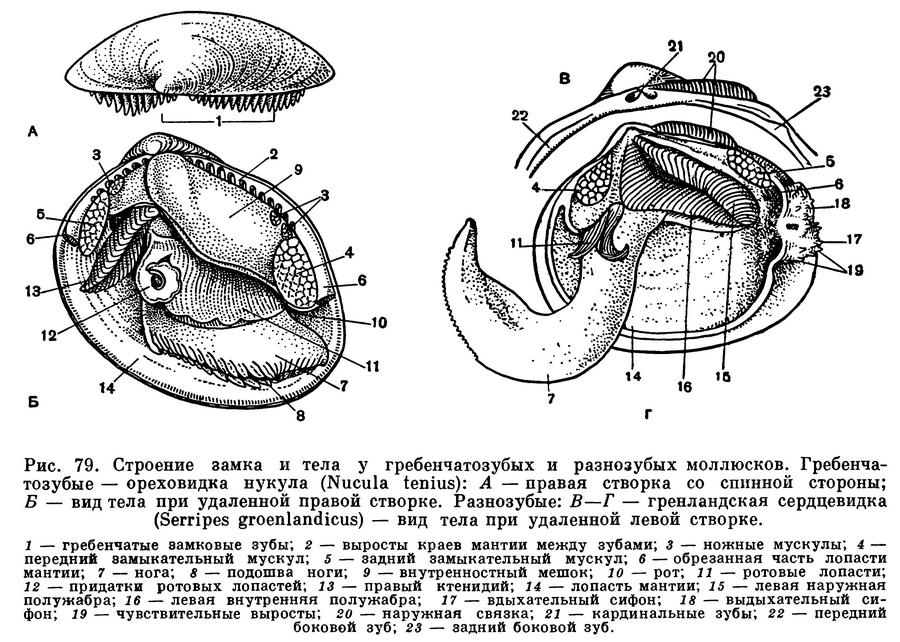
Каковы же основные черты строения двустворчатых моллюсков? Чтобы легче понять их устройство, представим себе книгу в переплете, поставленную корешком вверх. Обе половинки переплета будут соответствовать правой и левой створкам раковины, охватывающим тело моллюска с боков. Корешок книги будет аналогичен эластичной наружной связке (лигаменту), соединяющей обе створки на спинной стороне раковины и одновременно растягивающей их. Первый и последний листы книги соответствуют двум лопастям мантии, охватывающим тело с правой и левой сторон, а следующие два листа книги спереди и сзади будут аналогичны двум парам жабр с каждой стороны тела. И наконец, между обеими парами жабр внутри расположено само тело и нога — обычно довольно крупный мускулистый топоровидный или клиновидный орган, направленный вперед; у прикрепленных или малоподвижных форм нога может превращаться в небольшой вырост, и, наоборот, у активно двигающихся видов (например, у сердцевидок) нога становится сильной, слегка коленчатоизогнутой, приспособленной для продвижения в мягком песчанистом грунте.
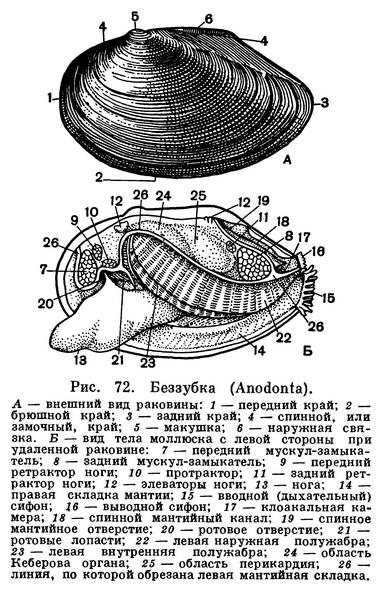
Расположение частей тела двустворчатого моллюска станет яснее, если рассмотреть вскрытого моллюска, например беззубку, обычную в наших пресноводных водоемах с илистым дном и медленно текущей или стоячей водой. Наиболее распространенной является обыкновенная беззубка (Anodonta cygnea) — довольно крупный моллюск из отряда настоящих пластинчатожаберных (Eulamellibranchia). При осмотре моллюска важно определить передний и задний концы раковины. Передний конец у беззубки легко узнать по более округлой форме раковины и по направленной вперед ноге; на заднем, несколько более суженном конце между створок видны короткие выросты мантии — сифоны. Вдоль верхнего спинного края, позади макушек, расположена довольно крупная наружная связка , или лигамент, — упругий эластичный тяж, при «сокращении» которого створки раскрываются. Он состоит из волокнистого рогового вещества, близкого к хитину, — конхиолина: образуется он из наружного покрова самой раковины (периострака). «Работа» лигамента определяется взаимодействием различно расположенных волокон конхиолина, из которых он состоит. Когда мускулы-замыкатели, сокращаясь, стягивают створки раковины, то волокна в нижней части лигамента сжимаются, а в верхней — растягиваются, а при расслаблении мускулов — наоборот; поэтому у мертвых моллюсков створки раковины всегда полуоткрыты. У двустворчатых моллюсков связка может быть наружной или внутренней или той и другой вместе.
У анодонты нет замковых зубов и спинной край гладкий, отсюда и ее название — беззубка (Anodonta). У большинства же двустворчатых моллюсков для более прочного соединения створок между собой под макушкой, изнутри, на спинном или замочном краю раковины, имеются разнообразные (по форме, числу и расположению) выросты, так называемые зубы, каждый из которых входит в соответствующее углубление на противоположной створке. Все это, вместе взятое, образует замок раковины. Устройство замка, характер, количество и расположение его зубов служит важным систематическим признаком у двустворчатых моллюсков и характерно для семейства, рода и вида. Лигамент также является частью замкового аппарата двустворчатых моллюсков, поскольку служит для соединения створок между собой.
Поверхность раковины у большинства двустворчатых моллюсков, в том числе и у беззубки, покрыта различно окрашенным наружным слоем, или периостраком. Он легко соскабливается ножом, и тогда под ним обнажается белый фарфоровидный, или призматический, известковый слой (ostracum). На нем хорошо видны концентрические линии — следы нарастания раковины, идущие параллельно ее краям. Внутренняя поверхность раковины у многих моллюсков, в том числе и у беззубки, выстлана перламутровым слоем (hypostracum).
Периострак, состоящий из конхиолина, устойчив к внешним воздействиям (как механическим, так и химическим) и служит, таким образом, хорошей защитой для внутреннего известкового слоя раковины. Особенно хорошо заметна стойкость периострака к действию растворенной в морской воде угольной кислоты. Она может накапливаться у дна, в самых придонных слоях, и в грунте, где живут моллюски (благодаря разложению органического вещества, отчасти и дыханию водных организмов), и возрастать по мере увеличения глубины и давления. Так, в Карском море часто встречаются совсем мягкие отмершие раковины астарт, йольдий или портландий с растворенной известковой частью раковины, от которой остался один неповрежденный мягкий роговой слой — периостракум.
Оба других слоя раковины состоят из известковых призмочек или пластинок, соединенных небольшим количеством конхиолина. В среднем (фарфоровом) слое они располагаются перпендикулярно к поверхности раковины, а во внутреннем (перламутровом) — параллельно ей; благодаря такому расположению получается интерференция света, которая и дает блеск и радужные переливы перламутра. Чем тоньше пластинки этого слоя, тем красивее и ярче этот блеск. Наиболее красивый перламутр бывает у тех моллюсков, у которых толщина пластинок перламутра в слое составляет 0,4—0,6 мк.
Раковина моллюска образуется в результате секреторной работы его мантии: вдоль ее края расположено большое количество железистых клеток, вырабатывающих различные слои раковины. Так, клетки особой мантийной бороздки, идущей вдоль всего края мантии, образуют наружный конхиолиновый слой, эпителиальные клетки так называемой краевой складки дают призматический слой раковины, а наружная поверхность мантии выделяет перламутровый слой.
Раковина двустворчатых моллюсков, состоящая более чем на 90% из СаCO3, содержит его в виде кальцита или арагонита, находящихся в различных соотношениях. У тропических моллюсков раковина содержит больше арагонита, а также довольно много стронция. Кристаллографическое изучение состава раковин ископаемых моллюсков позволяет теперь судить о температуре тех морей, в которых жили эти моллюски.
Кальций, откладывающийся в раковину мантией, поступает в нее не только через кровь, куда она попадает из пищи через кишечник, но, как показали последние эксперименты с радиоактивным кальцием, клетки мантии могут и сами извлекать кальций из воды.
Рост раковины происходит как путем общего утолщения створок в результате наслаивания все новых и новых известковых пластинок на внутренней поверхности створок, так и путем нарастания всей раковины по ее свободному краю. При наступлении неблагоприятных условий (в зимний период, при ухудшении питания и т. д.) рост раковины замедляется или даже приостанавливается, что хорошо заметно на поверхности раковины у многих моллюсков, где в это время образуются характерные сгущения линий, имеющих вид концентрических полос, идущих параллельно брюшному краю раковины. По зимним кольцам — сезонным остановкам роста — иногда можно определить приблизительно возраст моллюска. Однако у некоторых видов такие кольца неразличимы; у тропических форм, где нет сезонных явлений, такие кольца обычно вообще не образуются. У наших пресноводных перловиц и беззубок такие сезонные зимние перерывы роста имеются, поэтому годовые кольца выражены обычно хорошо.
Чтобы открыть створки раковины беззубки, надо прежде всего перерезать два имеющихся у нее внутри спереди и сзади довольно сильных мускула-замыкателя, стягивающих обе створки в поперечном направлении и закрывающих раковину. У живой беззубки легче сломать ее тонкую раковину, чем открыть ее, не перерезав этих мускулов.
Когда мускулы перерезаны, то створки сами свободно раскрываются, растягиваемые лигаментом, и можно видеть две мягкие полупрозрачные розоватые или желтоватые лопасти — мантию, охватывающую тело с боков. Края мантии слегка утолщены. Этим местом она прикрепляется к раковине, на внутренней поверхности створок которой образуется так называемая мантийная линия. Мантия беззубок, срастаясь сзади, образует два коротких сифона, опушенных короткими чувствительными выростами.
У моллюсков, закапывающихся в грунт, образуются длинные сократимые сифоны; места прикрепления втягивающих их мышц образуют на внутренней поверхности раковины отпечаток, так называемый синус. Чем глубже синус, тем длиннее сифоны у моллюсков, тем глубже они могут закапываться в грунт.
На брюшной стороне из-под края мантии вперед выдается довольно крупная клиновидная нога, направленная острым концом вперед. Нога у беззубки очень подвижна (как и у многих других моллюсков), и ее действие легко наблюдать в аквариуме. Как только анодонта успокоится, ее раковина приоткрывается, показываются розовато-желтые края мантии и наружу высовывается кончик ноги. Если все кругом спокойно, нога высовывается еще дальше (у крупных анодонт на 4—5 см), погружается в песок, и моллюск начинает двигаться вперед или закапываться в грунт передним концом, слегка приподнимаясь на своей ноге. На пройденном ею пути остается след в виде неглубокой бороздки.
Большая подвижность ноги беззубки обусловливается главным образом сокращением имеющихся в ней различных групп гладких мышц. Имеются парные передний и задний мускулы: ретракторы, втягивающие ногу косо вверх, протракторы, выдвигающие ногу вперед, и группа более мелких мускулов — поднимателей (элеваторов) ноги кверху. Все эти мышцы прикрепляются к внутренней поверхности створок раковины, где отпечатки мест их прикрепления довольно хорошо заметны (у ретракторов вдоль замочного края раковины). Кроме того, в ноге имеется еще целый ряд более мелких мышц, не прикрепляющихся к створкам и расположенных в ноге послойно и поперечно.
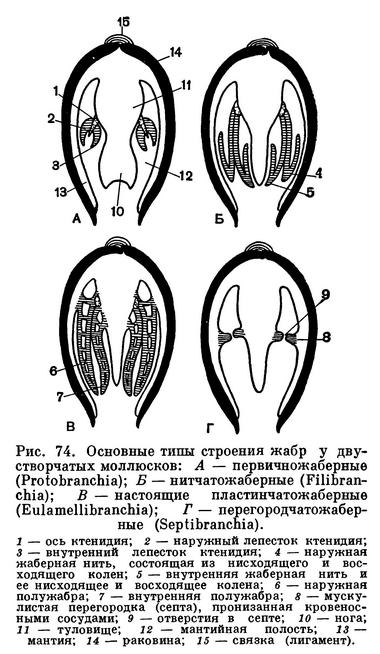
Если отвернуть мантийную лопасть кверху, то откроется мантийная полость беззубки, где расположены ее основные органы: ротовые лопасти, коричневатые жаберные листки (по два с каждой стороны тела), нога, основание которой находится между правой и левой жабрами. Спереди, в углублении между ногой и передним мускулом-замыкателем, помещается ротовое отверстие, окруженное двумя парами небольших треугольных сократимых околоротовых лопастей. Каждая жабра у анодонты состоит из двух листков полужабр, которые в свою очередь сложены двумя пластинками — восходящей и нисходящей.
Каждая жаберная пластинка состоит из рядов отдельных нитей (филаментов), а каждая нить соответственно образует восходящее и нисходящее колена. У анодонты между соседними нитями и между образуемыми ими коленами имеются сосудистые соединения (мостики), что характерно для всего отряда настоящих пластинчатожаберных. Каждая полужабра представляет собой, таким образом, решетчатую, сложно продырявленную двуслойную пластинку.
У представителей других отрядов двустворчатых моллюсков жабры имеют иное устройство (о чем будет сказано ниже).
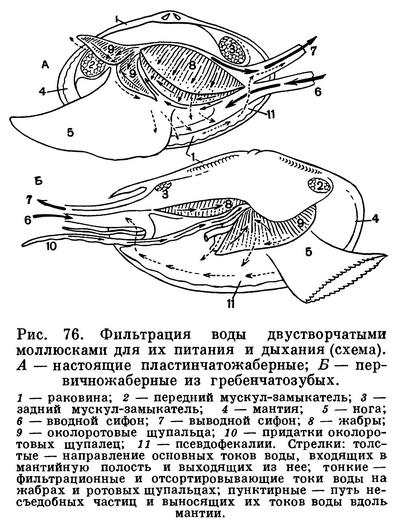
Мантийная полость и расположенные в ней жабры все время омываются током воды, который создается главным образом мерцанием мельчайших ресничек эпителия, покрывающего поверхность мантии, жабр, ротовых лопастей и стенок тела. Вода поступает в мантийную полость беззубки через нижний (дыхательный) сифон, попадая сначала в ее большую, нижнюю часть — дыхательную камеру, затем процеживается через щели в жабрах и уходит в верхнюю часть мантийной полости — выдыхательную камеру, откуда наконец и выходит наружу через верхний (выводной, или анальный) сифон. Засасывание воды через входной сифон происходит в результате разницы в гидростатическом давлении между поджаберным и наджаберным пространствами мантийной полости и между нею и окружающей моллюска водой; эта разница вызывается не только работой ресничного эпителия, но также и сокращением жаберных нитей и мускулатуры мантии и сифонов. Когда ток воды замедляется, входя в большую «вдыхательную камеру» мантии, грубые и крупные частицы выпадают из него и оседают на поверхности мантии, а затем удаляются из моллюска. Легко убедиться в наличии интенсивных токов воды, поступающей в мантийную полость, если положить беззубку в неглубокий сосуд с водой так, чтобы вода лишь слегка покрывала раковину. Дав ей успокоиться, надо подсыпать в воду вблизи заднего ее конца какой-нибудь порошок, остающийся взвешенным в воде (например, тушь, кармин, сухие тертые водоросли). Тогда можно увидеть, как крупинки порошка уходят через нижний (вводной) сифон внутрь раковины и через некоторое время с сильной струей воды выбрасываются наружу через верхний (выводной) сифон. Время от времени беззубка, часто без всякого внешнего раздражения, с силой захлопывает створки раковины и выбрасывает наружу струи воды, обновляя сразу всю воду, содержащуюся в мантийной полости. Вскоре створки раковины опять приоткрываются и нормальная медленная циркуляция воды возобновляется.
Чтобы убедиться в интенсивности работы ресничного эпителия, можно вырезать кусочек мантии беззубки и положить его внутренней поверхностью книзу на дно сосуда. Благодаря продолжающейся еще некоторое время работе ресничек этот кусочек будет слегка перемещаться и даже слегка ползти по наклонной плоскости.
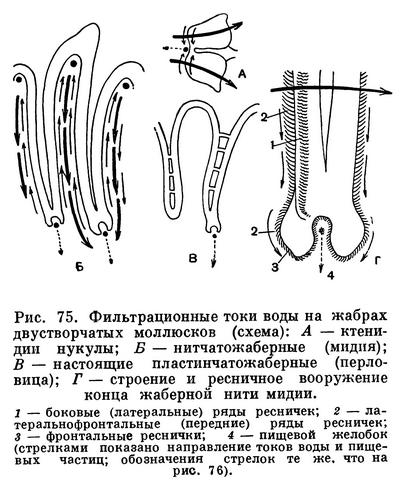
Засасывание воды и ее циркуляция внутри мантийной полости обеспечивают беззубку не только необходимым для ее дыхания кислородом, но и пищей. Как и все двустворчатые моллюски, беззубка лишена головы и целого ряда связанных с ней органов — обособленной глотки, слюнных желез, жестких образований для пережевывания пищи (как, например, хитиновых пластиночек — терки, имеющейся у брюхоногих моллюсков). Поэтому беззубка не может питаться крупными организмами. Она и большинство двустворчатых моллюсков (Eulamellibranchia и Filibranchia) являются активными фильтраторами. Такие моллюски питаются взвешенными в толще воды детритом (мельчайшими остатками отмерших растений и животных) и микропланктоном (одноклеточными водорослями, бактериями и очень мелкими животными). При помощи сложного ресничного механизма жабр и околоротовых лопастей моллюски отфильтровывают их из воды, отделяя несъедобную минеральную взвесь и крупные для них пищевые частицы.
Жаберные нити моллюсков имеют расположенные в определенных местах ряды различных по своим размерам ресничек, которые и могут отфильтровывать и отсортировывать пищевые частицы, обволакивать их слизью, а затем направлять в пищевые бороздки, расположенные вдоль брюшного края полужабр (в местах перехода нисходящих жаберных колен в восходящие) или у их основания. Ряды довольно крупных боковых, наиболее интенсивно работающих ресничек на жаберных нитях производят фильтрацию воды через узкие щели между жаберными нитями и обеспечивают прохождение ее из «вдыхательной» в «выдыхательную» камеру мантийной полости. Особенно крупные латеральнофронтальные реснички, сидящие по бокам жаберных нитей, отцеживают пищевые частички из воды или улавливают их обильно выделяемой слизью и проталкивают на наружную сторону жаберных нитей. Здесь расположены фронтальные реснички, которые собирают пищевые частички и направляют их вниз, к пищевому желобку. Пищевые частицы, собирающиеся в пищевых желобках, также обволакиваются слизью, образуют здесь комочки, уплотняются и благодаря работе ресничек желобка направляются к ротовым лопастям. Ротовые лопасти моллюсков — очень эффективный сортирующий аппарат, освобождающий пищу от несъедобных частиц. Они вооружены многими чувствительными элементами— хемо- и механорецепторами. Они имеют ряды поперечных бороздок, вооруженных особо длинными ресничками; самые мелкие частицы, пригодные для питания, по ряду таких бороздок направляются к ротовому желобку (расположенному у основания обеих лопастей), по которому они далее направляются к ротовому отверстию, где и заглатываются. По другим бороздкам (с ресничками, работающими в противоположном, чему предшествующих, направлении) более крупные частички и слизистые комочки, непригодные для питания, скатываются вниз и падают на мантию. Сильные реснички мантийных краев гонят эти частицы обратно к основанию вводного сифона; по мере продвижения туда эти частицы склеиваются, уплотняются и в виде так называемых псевдофекалий выбрасываются наружу.
У двустворчатых моллюсков из группы Protobranchia (ореховидок, лед, йольдий, портландий и т. п.), имеющих наиболее просто устроенные лепестковидные жабры — ктенидии, ротовые щупальца очень большие, сократимые и снабжены длинным желобчатым выростом. При помощи его они собирают с поверхности грунта мелкие пищевые частички — детрит, которые затем переносятся ресничками по желобку на пластинки ротовых щупалец, где и подвергаются сортировке; жабры-ктенидии служат главным образом для создания токов воды. Работа отфильтровывающего и сортирующего аппарата двустворчатых моллюсков довольно совершенна. Так, мидии могут отфильтровывать частицы размером от 40 до 1,5—2 мк (лучше всего — в 7—8 мк), полностью извлекая их из воды. Они задерживают одноклеточные водоросли и жгутиковых; более тяжелые частицы минеральных взвесей, даже размером в 4—5 мк, не задерживаются мидиями. Из смеси водорослей и пурпурных бактерий устрицы извлекают только водоросли; они обычно задерживают жгутиковых, водоросли и органические частицы размером более 2—3 мк и пропускают все частицы размером в 1 мк и меньше.
Двустворчатые моллюски профильтровывают очень большие объемы воды. Так, устрица может профильтровать за час около 10 л воды; мидия — до 2—5 л (при более высокой температуре воды больше, при более низкой — меньше); съедобная сердцевидка при температуре воды в 17—19,5°С — от 0,2 до 2,5 л, в среднем 0,5 л воды за час; мелкие морские гребешки фильтруют со скоростью 1 л в час на 1 г их веса, а старые — только 0,7 л.
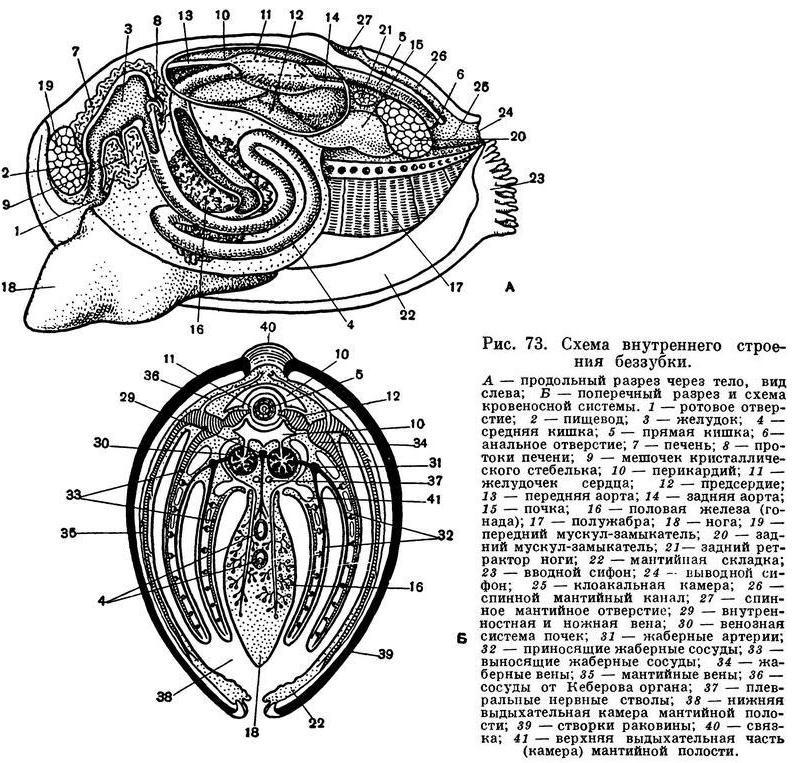
Пищеварительная система беззубки, как и у всех двустворчатых моллюсков, состоит из короткого пищевода, более или менее округлого желудка, средней и задней кишки; в желудок открываются протоки парной пищеварительной железы — печени, а на брюшной стороне выступает конец так называемого кристаллического стебелька. Отходящий от желудка кишечник (средняя кишка) в основании ноги, в массе половых желез образует 1—2 оборота, затем переходит на спинную сторону и, пронизывая нижнюю стенку перикардиальной сумки, проходит через желудочек сердца, выходит за пределы перикардия через его спинную часть, проходит над задним замыкательным мускулом и оканчивается анальным отверстием, открывающимся в клоакальную камеру мантийной полости с ее выводным сифоном. Часть кишечника, идущая от перикардия до анального отверстия, обычно называется прямой или задней кишкой. Кишечный тракт двустворчатых моллюсков не имеет мышечных волокон, и передвижение пищи в нем происходит благодаря работе выстилающего его ресничного эпителия. Удалению непереваренных остатков способствует мышечная связка, окружающая анальное отверстие.
Попав через короткий пищевод в желудок, пищевые частицы благодаря деятельности ресничного тока и желудочного желобка отсортировываются на мелкие и крупные. Крупные пищевые частицы попадают в кишечник, а более мелкие разносятся по складкам желудка и собираются у выступающего конца кристаллического стебелька. Его выступающий конец все время вращается, что способствует перемешиванию пищевых частиц и их сортировке. Кристаллический стебелек образуется в особом мешковидном органе и представляет собой стекловидную палочку студенистого вещества, состоящего из белка типа глобулина с адсорбированными в нем ферментами (амилазой и др.), способными переваривать углеводы (крахмал, гликоген). Попадая в слабокислую среду кишечника, он начинает растворяться и выделять адсорбированные в нем ферменты — единственные выделяемые двустворчатыми моллюсками в кишечный тракт для внеклеточного переваривания пищи. Мелкие пищевые частицы, обработанные энзимами кристаллического стебелька, поступают из желудка в выросты печени. Она состоит из очень большого числа удлиненных слепых трубочек — дивертикул и не является пищеварительной железой в обычном смысле; в ней не вырабатываются и не выделяются в кишечный тракт какие-либо пищеварительные ферменты, и она является органом внутриклеточного (а не внеклеточного) переваривания пищи и всасывания. Внутриклеточное переваривание у двустворчатых моллюсков производится главным образом фагоцитирующими блуждающими клетками — амебоцитами. Они во множестве встречаются не только в дивертикулах печени, но и в желудке и в средней кишке. Амебоциты обладают различными ферментами и способны переваривать не только углеводы, но и белки и жиры и т. д. Блуждающие клетки могут проходить через эпителий кишечного тракта в его просвет и обратно возвращаться в ткани. Заглатывают и переваривают пищевые частицы также и клетки печени; они также могут блуждать по просвету дивертикул и обратно возвращаться в стенки печени. Блуждающим клеткам и принадлежит основная роль в переваривании пищи у двустворчатых моллюсков.
При отмирании амебоцитов и клеток печени их пищеварительные ферменты могут поступать в просвет кишечного тракта. Поэтому следы различных ферментов (протеаз, липаз) находят в вытяжках из печени и желудка двустворчатых моллюсков.
Далеко не все попавшие в кишечный тракт организмы перевариваются двустворчатыми моллюсками. Нередко, особенно при большом количестве пищи, в фекальных массах моллюсков находят живых диатомей (одноклеточные водоросли с кремниевым скелетом), мелких веслоногих рачков и т. п. Поэтому при искусственном разведении двустворчатых моллюсков (устриц) в выростных бассейнах стараются поддерживать определенную (не избыточную) концентрацию планктонных водорослей, которыми они питаются.
Из сказанного видно, что пищеварение у двустворчатых моллюсков очень своеобразно. Они могут переваривать внеклеточно лишь углеводы, а белковые и жировые компоненты их пищи перевариваются фагоцитирующими блуждающими амебоцитами и клетками их «печени». Таким образом, двустворчатые моллюски являются очень специализированной группой животных, приспособленной к питанию детритом, одноклеточными водорослями и бактериями.
Кровеносная система у беззубок, как у всех двустворчатых моллюсков, незамкнутая, и кровь — гемолимфа — циркулирует не только по кровеносным сосудам — артериям и венам, но и в пространствах между органами, и в соединительной ткани по целой системе лакун и синусов, не имеющих собственных стенок. Артериальная кровь течет главным образом по сосудам, а венозная система имеет преимущественно лакунарный характер. Кровь прогоняется через всю систему благодаря сокращению сердца, а также мускулатуры тела. Сердце у двустворчатых моллюсков (у анодонты) состоит из желудочка и двух предсердий и лежит в перикардиальной полости, или околосердечной сумке, расположенной на спинной стороне тела. Перикардий представляет собой вытянутый тонкостенный мешочек, наполненный гемолимфой, и является у двустворчатых моллюсков частью сильно уменьшенной в объеме их вторичной полости тела. Желудочек обладает мощными мышечными стенками и имеет вид мешка грушевидной формы, своим широким концом обращенного назад. Предсердия очень тонкостенны, полупрозрачны и чаще всего имеют вид вытянутых треугольников, вершины которых открываются в желудочек; при входе в последний они снабжены небольшими полулунными складками — клапанами, пропускающими кровь только из предсердия в желудочек.
У беззубки, как у большинства двустворчатых, желудочек пронизан проходящей через него задней кишкой, но его полость при этом вполне замкнута и отделена от кишки своей стенкой. Из желудочка кровь расходится по телу: к заднему концу — через заднюю аорту, которая разделяется на две артерии, питающие сосуды задней части мантии и заднего замыкательного мускула; к переднему концу — через переднюю аорту и отходящие от нее артерии к ноге, к внутренностям и к передней части мантии. Из артериальных сосудов кровь изливается в не заполненные тканями промежутки, и через систему лакун кровь, ставшая венозной, собирается по синусам и венам в большой продольный венозный синус, лежащий между выделительными органами. Пройдя отсюда через венозную систему почек, она вливается в приносящие парные жаберные артерии, проходящие у основания каждой жабры. Из них венозная кровь по приносящим жаберным сосудам нисходящих жаберных пластинок растекается по жаберным нитям и их сосудистым перемычкам. Окисленная в жабрах артериальная кровь, насыщенная кислородом, по выносящим сосудам восходящих жаберных пластинок вливается в парные (с каждой стороны моллюска) жаберные вены, откуда поступает в предсердия. В предсердия поступает и та часть крови, которая, минуя жабры и почки, окислилась в сосудах мантийных складок и по мантийным венам поступила в наружные жаберные вены. Мантия с ее сильно разветвленными кровеносными сосудами играет у двустворчатых моллюсков очень большую роль в дыхании и обогащении крови кислородом.
То, что у большинства двустворчатых моллюсков желудочек сердца оказывается пронизанным прямой кишкой, находит свое объяснение в особенностях их эмбрионального развития и в эволюции всей этой группы. У ряда низших представителей двустворчатых имеется не только два предсердия, но и два раздельных желудочка, лежащих по бокам кишки (у арок); у других — непарный желудочек лежит над кишкой (ореховидки, аномии, лимы), у третьих — он лежит книзу от кишки (устрицы, жемчужницы и др.). Все это указывает на то, что расположение кишки и сердца по отношению друг к другу претерпевало большие изменения в процессе эволюции двустворчатых моллюсков, и на то, что первоначально у них было два раздельных желудочка, которые затем слились вместе. Частота сокращения сердца у двустворчатых моллюсков, являющихся в общем малоподвижными организмами, невелика, обычно не более 15—30 раз в минуту, в то время как у таких подвижных и активных моллюсков, как головоногие, сердце сокращается 40—80 раз в минуту. Все отделы сердца двустворчатых моллюсков могут сокращаться автономно.
У двустворчатых моллюсков, как и вообще у беспозвоночных с незамкнутой кровеносной системой, кровяное давление очень низкое и сильно изменчиво.
Кровь — гемолимфа — двустворчатых моллюсков играет огромную роль в их жизни, обмене веществ. Она выполняет целый ряд функций: обеспечивает внутренние органы и ткани кислородом и питательными веществами, уносит продукты их жизнедеятельности (углекислоту, продукты азотистого обмена и т. п.), создает и поддерживает постоянство их внутренней среды (ионного состава, осмотического давления). Наконец, играет очень большую роль в создании гидравлического механизма давления, необходимого тургора (напряжения), а также при движении моллюсков. Изучение циркуляции крови в теле двустворчатых моллюсков объяснило явление разбухания ноги, что наблюдается при движении и закапывании животного. Оно происходит благодаря наполнению ее кровью, которая и придает ноге необходимую упругость, создает необходимый тургор. Когда нога вытягивается и ножные мышцы расслабляются, кровь поступает по артерии в ногу, а при ее сокращении уходит обратно в тело. Так, у морского черенка, который может очень быстро закапываться, нога сначала погружается в грунт и кровь быстро втекает в нее, расширяя конец ноги в виде диска; последний служит якорем, когда ножные мышцы, сокращаясь, тянут моллюска вниз. При подъеме моллюска из грунта на поверхность ножные мышцы расслабляются, а конец ноги снова расширяется, заполняясь кровью; держась на таком «якоре», нога вытягивается, так как часть крови поступает в верхнюю часть ноги и подталкивает моллюска вверх. Приток, нагнетание необходимого для набухания ноги количества крови и ее отток регулируются так называемым кеберовым органом, играющим при этом как бы роль клапана.
В противоположность животным с замкнутой кровеносной системой двустворчатые моллюски, как и все беспозвоночные, обладающие незамкнутой кровеносной системой, имеют довольно значительный объем крови — гемолимфы. У моллюсков (кроме головоногих) он составляет 40—60% (объемных процентов) к весу их тела без раковины. У пресноводной жемчужницы (Margaritifera) и у мидии (Mytilus californianus) на 100 г веса тела объем крови составляет около 50 мл.
В крови двустворчатых моллюсков имеется много форменных элементов, главным образом различных форм амебоцитов (лейкоцитов). Количество их колеблется у разных видов от 6 000 до 40 000 в 1 мм3. У двустворчатых моллюсков имеются и эритроциты; иногда их даже может быть больше, чем некоторых форм лейкоцитов. Гемоглобин обнаружен у довольно многих видов (арки, морские черенки, теллины, пектункулюсы, астарты и др.).
Важная для газообмена (для снабжения органов и тканей кислородом) способность крови насыщаться кислородом у двустворчатых моллюсков очень мала и составляет 1—5% к объему их крови. Так, 100 мл крови беззубки способны поглотить всего лишь 0,7 мл кислорода, а у мидий — 0,3 мл. Беззубка потребляет 0,002 мл O2 на 1 г своего веса за один час (при 10°С); устрицы соответственно — 0,006 мл O2 (при 20°С); мидии — 0,055 мл O2. Несколько больше обычно потребляют виды более подвижные, как, например, морские гребешки Pecten grandis, потребляющие 0,07 мл O2 на 1 г своего веса за 1 час (или 70 см3 O2 на 1 кг веса). Мелкие формы также нередко потребляют больше кислорода, чем крупные. Например, шаровка роговая при оптимальной для нее температуре воды (18°С) потребляет 0,05 мг O2 на 1 г веса за час, но при снижении температуры воды до 0,5°С потребление кислорода у нее почти прекращается. В потреблении кислорода, т. е. в интенсивности обмена, у многих двустворчатых моллюсков наблюдаются сезонные колебания; так, мидии в летнее, наиболее активное время их жизни потребляют примерно в два раза больше кислорода, чем в зимнее, холодное время года.
Многие двустворчатые моллюски могут довольно долгое время жить при очень малом содержании кислорода в воде или даже при его отсутствии. Так, песчаная ракушка (Муа arenaria) может жить в бескислородных условиях при 14°С до 8 дней, а при 0° даже несколько недель; виргинская устрица также выносит такие условия в течение недели и более. Обмен веществ в такие периоды анаэробиоза резко снижается, но моллюски могут при этом получать необходимый для их жизни кислород путем интрамолекулярного дыхания — гликолитического расщепления их запасных веществ (углеводов и жиров) по типу брожения. Такая способность ко временному (факультативному) анаэробиозу (аноксибиозу) особенно характерна и необходима для видов, живущих на литорали (как, например, для песчаной ракушки, мидий, балтийской макомы, съедобной сердцевидки). Во время отлива они закрывают свои раковины, небольшое количество кислорода в их мантийной полости довольно быстро исчезает, и они начинают жить за счет процессов аноксибиоза. Во время прилива они раскрывают свои раковины, непрерывно фильтруют воду и дышат кислородом, растворенным в воде; вначале у них резко (в несколько раз) повышается интенсивность фильтрации и потребления кислорода, а затем через некоторое время приходит в норму, характерную для их жизни в воде.
Органами выделения у двустворчатых моллюсков являются почки, а также, но в меньшей степени так называемый кеберов орган; последний представляет собой железистое утолщение передней части и передне-боковых стенок перикардиальной сумки.
Почки, или боянусовы органы, открываются своими внутренними концами в перикардий, а наружными — в мантийную полость. Почки у беззубки имеют вид двух темно-зеленых изогнутых трубчатых мешочков; один конец имеет железистые стенки и представляет собственно выделительную часть почки, другой имеет вид пузырька, где накапливаются продукты обмена веществ, подлежащие удалению из тела.
У двустворчатых моллюсков нет такой концентрации частей центральной нервной системы (нервных узелков, или ганглиев), как у брюхоногих моллюсков. У беззубки, например, имеется одна пара ганглиев над ртом, немного позади него, другая пара — глубоко в ноге и третья — позади заднего замыкательного мускула. Между первой и второй парой ганглиев, а также между первой и третьей проходит по паре нервных стволов, а каждая пара узелков соединена между собой поперечными перемычками (комиссурами).
Органы чувств у двустворчатых по сравнению с другими классами моллюсков развиты довольно слабо. Однако эти органы у них довольно разнообразны по своему строению и рассеяны в различных частях тела: вдоль наружных краев мантии, на концах сифонов, на первых жаберных нитях, около ротового отверстия на околоротовых щупальцах, на краях заднего замыкательного мускула, в выдыхательной камере, около задней кишки и т. д. Эти органы чувств представляют собой как довольно сложные образования — глаза, или фоторецепторы, органы равновесия — статоцисты, или статорецепторы, так и более простые — осфрадии, различные чувствительные выросты, а иногда и просто скопления пигментированных чувствительных клеток.
Фоторецепторы у двустворчатых моллюсков могут быть устроены очень различно: от простых эпителиальных пигментных (оптических органелл) до довольно сложных глаз с хрусталиком и сетчаткой. Такие глаза могут быть очень многочисленными, особенно у свободноживущих форм, как например мантийные глаза у гребешков, у которых их насчитывается иногда до 100 на обоих краях мантии.
Различно устроенные глаза и глазки могут располагаться у двустворчатых также на первых жаберных нитях (жаберные глаза у арок, аномий), на коротких выростах вокруг отверстий сифонов (у некоторых сердцевидок и др.).
У многих двустворчатых имеются так называемые оптические органеллы, сферические или удлиненные, концентрирующие свет на особом внутриклеточном иннервированном образовании (ретинелле). Такие фоторецепторы разбросаны на концах сифонов и в других частях тела моллюсков.
Органы равновесия у двустворчатых представляют собой пузыревидное впячивание эпителия, хорошо иннервируемое, выстланное изнутри ресничным эпителием, замкнутое (статоцист) или открытое (статокрипт). Они содержат внутри твердое минеральное зерно (статолит) или мелкие песчинки (статоконии). Обычно, например у беззубки, статоцисты расположены недалеко от ножного ганглия или на спинной стороне моллюска. Органы равновесия хорошо развиты у свободноживущих форм, например у морских гребешков.
Осфрадии представляют собой обычно очень мелкие парные пигментированные хорошо иннервируемые группы чувствительных клеток. Они могут располагаться в различных местах — на ноге, в области жабр, задней кишки и т. д. Их роль еще недостаточно ясна: являются ли они хеморецепторами или органами осязания.
Анодонты, как и большинство двустворчатых, раздельнополы, но в условиях водоемов со стоячей водой могут встречаться отдельные гермафродитные особи или даже целые их колонии. При этом во избежание самооплодотворения сначала продуцируются мужские половые продукты — сперматозоиды, а затем женские — яйца. Парные, сильно расчлененные половые железы (гонады) у двустворчатых (в том числе и у анодонты) залегают в спинной части ноги, где их окружает петля кишечника и выросты печени; выводные протоки гонад открываются в мантийную полость рядом с отверстиями выделительной системы. Только у некоторых самых примитивных двустворчатых моллюсков протоки половых желез открываются общим отверстием с выделительной порой. У некоторых пресноводных двустворчатых половой диморфизм выражен настолько резко, что самцов и самок одного и того же вида иногда относят к разным видам.
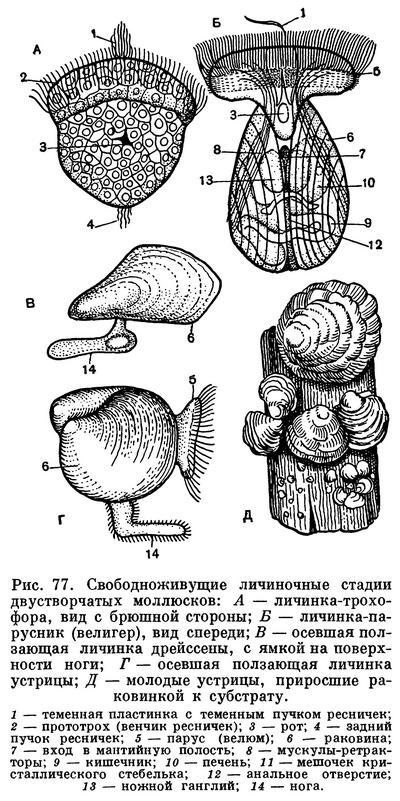
Развитие молоди у двустворчатых моллюсков происходит разнообразно. Почти все морские формы, обитающие на мелководьях, откладывают яйца прямо в воду, где и происходит оплодотворение, или же оно происходит в мантийной полости материнской особи. Яйца свободно плавают в воде, реже склеиваются или прикрепляются к раковинам, водорослям. Исключение представляют живородящие (точнее, личинкородящие) формы, как некоторые устрицы, арки и др.
Оплодотворенные яйца двустворчатых моллюсков, пройдя стадию дробления спирального типа, образуют трохофорообразную личинку, сходную с личинкой многощетинковых червей (полихет). Однако в процессе эмбрионального развития двустворчатых моллюсков почти отсутствует процесс сегментации, столь характерный для развития личиночных стадий кольчатых червей. У личинки двустворчатых моллюсков имеется зачаток ноги и первичной раковины (продиссоконха), которая первоначально закладывается в виде одной пластинки, расположенной на спинной стороне личинки. После ряда изменений трохофора, у которой возникает парус (velum) — ресничный теменной диск, двустворчатая раковинка и зачатки других органов, превращаются в велигер. Наличие свободноплавающей личинки (велигер) представляет собой очень важную жизненную стадию моллюсков, так как обеспечивает им возможности широкого расселения, поскольку взрослые двустворчатые моллюски ведут обычно малоподвижный или даже прикрепленный образ жизни. Вместе с тем эта личиночная стадия жизни, как и у других беспозвоночных, является самой чувствительной к неблагоприятным внешним условиям, и лишь высокая плодовитость двустворчатых моллюсков обеспечивает сохранение вида и его расселение.
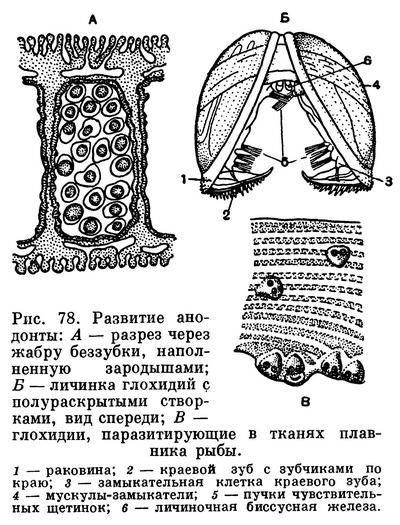
У некоторых видов двустворчатых эта пелагическая личиночная стадия утрачена и яйца развиваются внутри мантийной полости материнского организма (как у многих пресноводных форм), так что из материнской раковины выходят уже почти готовые молодые особи. У некоторых пресноводных же форм цикл развития осложняется своеобразным паразитизмом личинок на теле рыб. Этот период следует за периодом первоначального развития на жабрах материнской особи, куда и откладываются их яйца, при этом у некоторых видов на жабрах образуются специальные выводковые камеры (марзупиум). Примером может служить развитие личинки анодонты — глохидия, протекающее, как и у всех Unionidae, в наружных жабрах материнской особи. В дальнейшем он становится паразитом рыб, прикрепляясь к их коже или к жабрам биссусом или зубцами раковинки. Погружаясь в ткани рыбы, глохидий окружается цистой, питается за счет соков тела хозяина. Этот паразитический период продолжается 1—2 месяца, после чего моллюски выходят в воду и, опускаясь на дно, переходят к свободному образу жизни.
У ряда морских холодноводных и, видимо, у многих глубоководных океанических видов двустворчатых моллюсков развитие может происходить с очень укороченной стадией плавающей личинки или совсем без нее. В последнем случае образуются немногочисленные крупные яйца, снабженные большим количеством питательного вещества. Это позволяет им развиваться вне зависимости от наличия пищи в окружающей воде, что особенно важно для глубоководных форм, где количество пищи для молоди у дна очень ограниченно.
Основываясь на строении раковины, замка, жабр, на расположении и количестве мускулов-замыкателей, связки и пр., различают отряды: Гребенчатозубые, Связочнозубые, Настоящие пластинчатожаберные и Перегородчатожаберные.
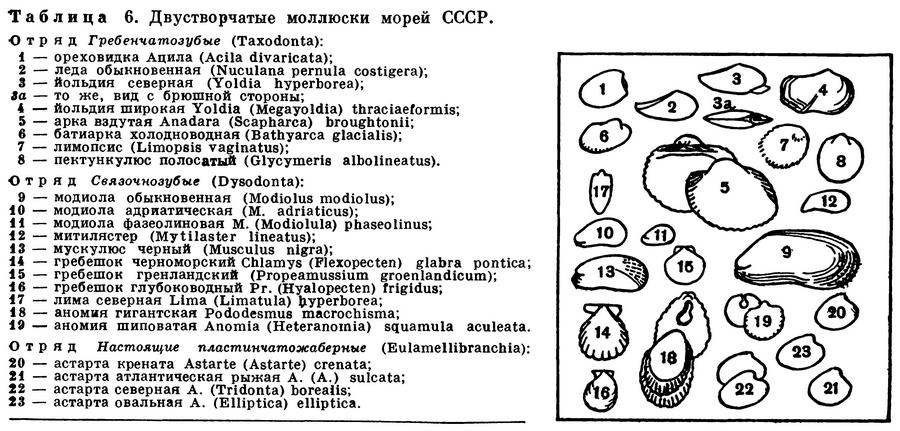
ОТРЯД ГРЕБЕНЧАТОЗУБЫЕ (TAXODONTA)
К этому отряду относятся двустворчатые моллюски, имеющие ряд примитивных черт в своем строении, например в первую очередь замок «таксодонтного типа», т. е. состоящий из ряда мелких зубчиков, расположенных вдоль спинного (верхнего) края раковины. Особенно ясно выражены такие признаки у видов из надсемейства нукулид (Nuculacea), являющихся наиболее древними среди прочих гребенчатозубых. У нукулид имеются наиболее просто устроенные жабры типа ктенидиев, состоящие из ряда мелких округлых листочков, по одному ряду с каждой стороны тела, сидящие на общей оси (Protobranchia); нога у них со складывающейся подошвой, а полость гонад соединяется с почками.
Виды второго большого надсемейства арок (Arcacea), хотя и имеют существенные отличия от нукулид, но также сохраняют ряд примитивных черт в строении гребенчатого замка и жабр. Их жабры состоят из рядов тонких, свободных, не сросшихся между собой нитей (Filibranchia). Нога без подошвы, маленькая и нередко образует биссусные нити.
Моллюски, относящиеся к отряду гребенчатозубых, очень широко распространены в Мировом океане и его окраинных морях; они встречаются во всех наших морях, кроме Азовского и Каспийского озера-моря, обитают на самых разнообразных глубинах, почти от уреза воды вплоть до ультраабиссальных глубин океанических впадин (более 6000 м). Рассмотрим наиболее важные из них.
К надсемейству Nuculacea относятся семейства: ореховидки (Nuculidae), ледиды (Nuculanidae, или Ledidae) и маллеции (Malletiidae).
Представители первых двух широко распространены в Мировом океане, особенно в мелководных шельфовых морях, где многие массовые виды входят в состав биоценозов донной фауны. В семействе же Malletiidae преобладают более глубоководные виды, которые встречаются обычно на больших глубинах, особенно в абиссали — на ложе океана, до 4—6 км.
Из первого семейства наиболее известна тонкая ореховидка-нукула (Nucula tenuis) — одна из обычнейших форм морей северного полушария, где обитает на илах до глубин не более 100—150 м, а в северных частях Атлантического и Тихого океанов изредка встречается и на глубинах около 1000 м. На мелководьях Баренцева моря и у берегов Мурмана эта ореховидка, как и другие двустворчатые, является излюбленной пищей ряда донных промысловых рыб (пикши, камбал, молоди тресковых), в желудках которых встречается десятками.
Из семейства Nuculanidae наиболее известны различные йольдии, портландии, леди и др.
Виды рода Йольдия (Yoldia) очень обычны и многочисленны в морях северного полушария. У настоящих йольдий поверхность раковины гладкая, у видов подродa Cnesterium она украшена тонкими косыми линиями; у видов подрода Megayoldia раковина самая крупная среди всех йольдий (до 6—7 см), с широким, как бы обрубленным сзади концом. Для йольдий характерно наличие пары длинных сросшихся сифонов, широкие, сократимые ротовые лопасти и сильная мускулистая нога со складывающейся подошвой. При таком «вооружении» они легко могут закапываться в мягкий илистый грунт прибрежных мелководий, выставляя над дном лишь задний конец раковины, а также сифоны и ротовые щупальца. При этом сифоны и околоротовые лопасти могут вытягиваться далеко за пределы раковины. Лопасти «ощупывают» поверхность дна вокруг моллюска, собирая детрит с его поверхности, а через сифоны идет фильтрация воды из придонного слоя; йольдии наиболее обычны на глубине до 100 м, редко глубже.
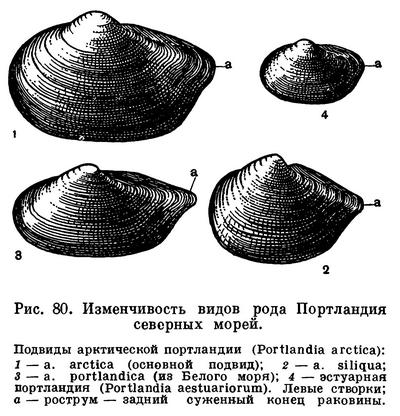
Особый интерес представляют широко распространенные на мелководьях арктических морей небольшие холодноводные моллюски портландии, раньше также называвшиеся йольдиями.
Арктическая портландия играла большую роль в северных морях прошлых геологических эпох. Она была руководящей формой (характернейшей и массовой) холодного «Йольдиевого моря» (названного так по ее имени), которое около 8—10 тыс. лет назад покрывало в межледниковый период весь север Европы вплоть до Балтийского моря. Остатки массовых поселений арктической портландии указывают на пределы распространения этого моря, образовавшегося вслед за отступлением края ледника, и на условия, при которых существовала населявшая его в то время донная фауна.
В составе настоящей морской фауны моллюсков большую роль играют леды — представители рода Nuculana, или Leda, куда входит много видов. Для них характерна удлиненная раковина с сильно оттянутым задним ее концом, вследствие чего ее длинные несократимые сифоны лежат в раковине, как в футляре. У типичного и наиболее обычного для этого рода вида обыкновенная леда (Nuculana pernula) поверхность зеленовато-желтой раковины покрыта тонкой концентрической исчерченностью. Обыкновенная леда обитает почти во всех морях северного полушария, где образует несколько подвидов.
В третье семейство отряда гребенчатозубых — Malletiidae — входит несколько типично глубоководных океанических родов, среди которых имеются очень древние примитивные формы: Malletia, Tindaria, Neilonella, Spinula и др.
К Nuculacea (Protobranchia) также относят очень интересное семейство солемий (Solemyidae), представители которого известны еще из отложений девона.
Как и нукулы, они обладают жабрами- ктенидиями и ногой с подошвой; их удлиненно-овальная раковина совсем лишена зубов и покрыта тонким периостракумом, выступающим в виде радиальных выростов за брюшной край их раковины. Хотя они и не имеют сифонов, но могут закапываться в грунт. Многие солемии глубоководные, имеются и литоральные виды.
Ко второму надсемейству Arcacea из отряда гребенчатозубых относятся семейства арок (Arcidae), лимопсисов (Limopsidae), пектункулюсов (Glycymeridae), большая часть видов которых обитает в теплых морях и на тропических мелководьях.
У Arcacea удлиненно-трапециевидная или округлая раковина, с прямым или слегка изогнутым замочным краем, с многочисленными вертикальными или скошенными зубами. Раковина с радиальными ребрами. Нередко она покрыта снаружи тонкими волосовидными выростами периострака или гладкая. Между спинным краем раковин и макушками имеется ромбовидная площадка; нога слабая, имеется биссус. Жабры нитевидные, несросшиеся, сифонов нет.
В наиболее богатое семейство арок (Arcidae) входит около 200 видов, из которых в наших северных морях широкое распространение имеют только три североатлантических вида из рода батиарок (Bathyarca) с сероватой бархатистой опушкой поверхности раковины. В Белом море батиарок нет. В Черном море встречается лишь одна мелкая (около 1 см) арка — Arca (Arcopsis) lactea — средиземноморско-атлантический вид.
ОТРЯД СВЯЗОЧНОЗУБЫЕ (DYSODONTA)
В этот отряд входит 13 семейств моллюсков, весьма различных по устройству своей раковины и тела, а также по распространению и образу жизни. Вместе с тем их объединяет большая или меньшая редукция переднего мускула-замыкателя, который либо может быть сильно уменьшенным в размере (Неравномускульные — Anisomyaria), или имеется лишь один крупный мускул-замыкатель, обычна расположенный посредине раковины (Одномускульные — Monomyaria).
Нередко весь этот отряд связочнозубых называют неравномускульными. Общим для всего отряда является отсутствие настоящего замка, однако у некоторых (Mytilidae) под макушкой имеются по краю раковины зубовидные выросты. Соединение створок между собой происходит главным образом при помощи лигамента (связки). Жабры нитчатые (Filibranchia), состоят из двух складчатых (наружных и внутренних) листков (полужабр) с каждой стороны тела. Они образованы длинными опушенными ресничками, тонкими согнутыми жаберными нитями, образующими сперва нисходящее, а затем восходящее колена. Отдельные жаберные нити не срастаются, а лишь сцепляются между собой при помощи жестких ресничек, сидящих на особых дисках. Восходящее и нисходящее колена жаберных нитей могут срастаться соединительнотканными перемычками лишь в местах их сгиба или у концов их восходящих колен.
Почти все связочнозубые являются формами эпифауны, т. е. живут на поверхности грунта, прикрепляясь к нему своим биссусом или даже прирастая створкой к камням и скалам, или свободно лежат на поверхности грунта и даже могут плавать; некоторые из них живут частично «воткнувшись» в грунт. Молодые формы обычно свободно передвигаются при помощи своей ноги, которая у взрослых обычно редуцируется. Все они активные фильтраторы.
Из этого отряда рассмотрим лишь некоторых его представителей, интересных с точки зрения их строения, образа жизни или хозяйственного значения.

К семейству Mytilidae относится много массовых видов: мидий, модиол, мускулюсов, митилястеров и др., широко распространенных в морях земного шара, преимущественно в мелководных их районах. Все они имеют клиновидную «митилидного» типа раковину; передний мускул-замыкатель маленький и узкий, а задний — большой, округлый.
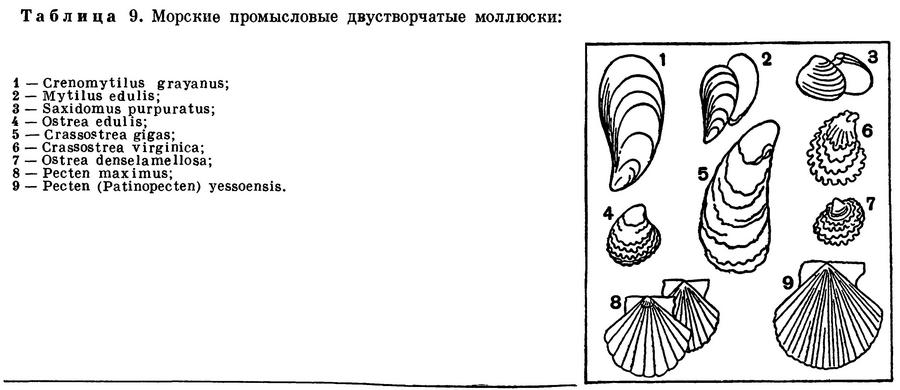
Наиболее широко известны мидии, многие виды которых являются съедобными и промысловыми: обыкновенная съедобная мидия (Mytilus edulis), дальневосточная гигантская мидия, или черная ракушка (Crenomytilus grayanus), средиземноморско-черноморская мидия (Mytilus galloprovincialis), калифорнийская (Mytilus californianus), мидия Магеллана (Mytilus magellanicus) и др.
Мидии — обычные обитатели прибрежной зоны, где они часто образовывают массовые поселения. Срастаясь биссусами друг с другом, они образуют на берегах так называемые щетки; их большие скопления на литорали и на подводных мелководьях в более открытых частях моря носят название мидиевых банок.
У мидий удлиненная клиновидная раковина, суженная спереди, расширенная сзади. В связи с их прикрепленным образом жизни макушка сдвинута на передний (приостренный) конец раковины. Окраска раковины темная, часто иссиня-черная, внутренняя поверхность имеет тонкий перламутровый слой. Имеется несколько мелких замковых зубчиков; биссус хорошо развит.
Мидий употребляют в пищу в жареном, вареном и в консервированном виде (сырые мидии быстро портятся и становятся ядовитыми). Пищевая ценность мидий высока: мясо свежих мидий содержит около 10% белков, 1% жиров, 0,5% углеводов (гликогена), а также витамины В и С и много разных полезных для организма человека солей.
Из мелких мидий делают также очень питательную кормовую муку, которую используют для корма домашних животных.
Любопытны по образу жизни морские, или каменные, финики из родов Литофага (Lithophaga) и Ботула (Botula). Они имеют низкую удлиненную раковину с гладкими или исчерченными створками, покрытыми толстым периостраком. Они ведут «скрытый» образ жизни, протачивая в известковых камнях и скалах ходы и норки, где и живут, прикрепляясь к их стенкам биссусом и выставляя свои длинные сифоны. Поэтому находить и собирать их очень трудно. Часто они живут вместе с другими моллюсками, например камнеточцами фоладами. Хорошо известны такие виды, как средиземноморская литофага (Lithophaga lithophaga), 8 см длиной, калифорнийская ботула Botula (Adula) californiensis — до 4 см длиной, и др. Морские финики протачивают свои ходы, выделяя особый кислый секрет (сок), растворяющий известковую породу прибрежных скал. Железа, выделяющая этот секрет, находится в передней части края мантии; поэтому они имеют тонкую раковину, которая не участвует в сверлении. На берегах Средиземного моря морские финики ценятся как лакомое блюдо.
Очень интересны моллюски из тропического надсемейства птерий (Pteriacea), раковины у которых обладают хорошо развитым перламутровым слоем, передний мускул-замыкатель очень мал или редуцирован, а нога обычно обладает сильно развитым биссусом. Сюда относятся различные пинны и атрины, или раковины-перья (сем. Pinnidae), морские молоточки (Malleus, сем. Isognomonidae) и настоящие морские жемчужницы, или крыловидки (сем. Pteriidae). Пинны и атрины имеют очень характерную клиновидную, нередко очень крупную раковину без замковых зубов и с очень сильно развитым биссусом. Поверхность раковины обычно украшена радиальными ребрами или множеством чешуек.
К родам Pinna и Atrina относится около 20 видов, живущих в Средиземном море, в Атлантическом, Тихом и Индийском океанах. Благородная пинна (Pinna nobilis), живущая в Средиземном море и в Атлантике, достигает 30 см длины, являясь самой крупной среди двустворчатых моллюсков европейских морей. Внутри раковины пинн красивого красноватого цвета нередко встречаются красноватые же или темные жемчужинки.
Еще более интересны виды из тропического индо-тихоокеанского рода Malleus. За причудливую форму раковины они получили название «морской молоточек». Замковый край у раковин маллеуса сильно удлинен, створки же толстой известковой раковины сужены и вытянуты наподобие ручки молотка. Внутри раковин Malleus malleus, Malleus albus и других видов этого рода иногда также встречается жемчуг, но реже, чем у обычных жемчужниц.
Морские жемчужницы издавна славятся высоким качеством своего перламутра и способностью образовывать самый лучший и ценный жемчуг. Для них характерна крупная различной формы раковина, с прямым замочным краем, обычно вытянутым сзади в уховидный или клювовидный выступ; замковый край с одним-двумя зубовидными утолщениями и передний мускул-замыкатель редуцированы, биссус сильно развит. Раковина птерий состоит из двух слоев: наружного (призматического) и мощного внутреннего (перламутрового) слоя. Наиболее крупная из жемчужниц Pinctada margaritifera может достигать 30 см в поперечнике и веса 10 кг (обычно они мельче).
К морским жемчужницам-крыловидкам относится много тропических видов, распадающихся на два рода: птерии (Pteria, или Avicula) и пинктады (Pinctada). Наибольшее промысловое значение имеют: Pinctada margaritifera, мелководный индо-тихоокеанский вид, живущий также у берегов Центральной Америки и Австралии, Pinctada martensii — у берегов Японии; цейлонская жемчужница (Pinctada vulgaris) и др. После постройки Суэцкого канала цейлонская жемчужница проникла и в Средиземное море.
Пинктады и птерии ведут сидячий образ жизни, прикрепляясь своим густым и прочным биссусом к подводным скалам и камням. Часто они образуют банки — колонии и грозди на глубине от 5—6 до 60 м (обычно на глубине 10—15 м), обитая здесь вместе с другими формами эпифауны, т. е. животными, сидящими на поверхности жесткого грунта, — кораллами, губками, гидроидами и др. Жемчужницы не выносят опреснения, являясь типичными обитателями чисто морских районов. Они растут очень медленно: в возрасте трех лет они достигают размеров всего 5—6 см.
К группе одномускульных (Monomyaria) из отряда Dysodonta относятся три богатых видами семейства моллюсков. Таково в первую очередь семейство Морские гребешки (Pectinidae), насчитывающее много родов и видов, широко распространенных почти во всех морях и океанах, на самых разнообразных глубинах вплоть до ультраабиссальных; так, в Курило-Камчатской впадине Тихого океана на глубине 8100 м «Витязем» был найден крупный полупрозрачный Delectopecten, и это пока наибольшая глубина встречаемости морских гребешков.
Особенно богат и разнообразен мир морских гребешков в водах прибрежных мелководий субтропической и умеренной зон Мирового океана.
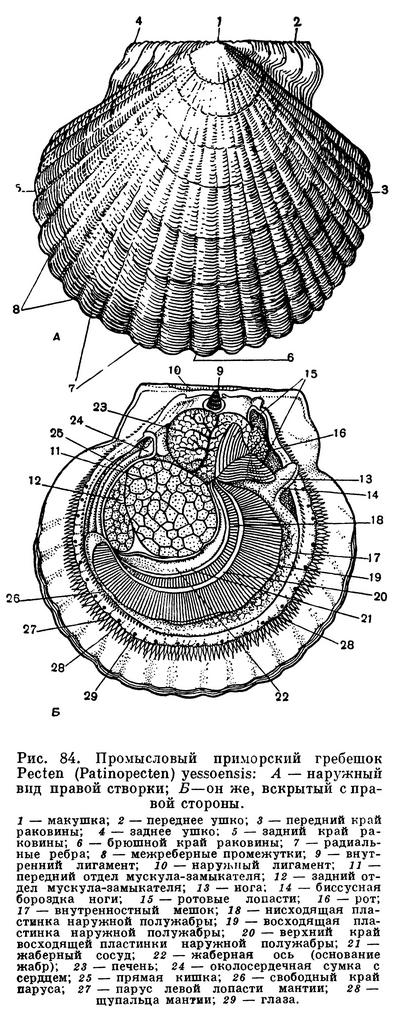
В морях Советского Союза встречается сравнительно немного видов морских гребешков, наибольшее их количество обитает в морях Дальнего Востока. На прибрежных мелководьях Японского моря, до глубины около 50 м, от Кореи до Сахалина и Южно-Курильских островов обитают хорошо известные дальневосточные гребешки: крупный (до 20 см в поперечнике, реже — больше) приморский промысловый гребешок Pecten (Patinopecten) yessoensis, с белой радиальноребристой раковиной, и очень красивый гребешок Свифта — Chlamys (Swiftopecten) swifti. В южной части Японского моря встречается также и японский гребешок Фаррера (Chlamys farreri nipponensis).
В Беринговом, Охотском морях и в южной части Чукотского моря обитает беринговоморский гребешок (Chlamys beringianus), а также ряд других видов из рода Chlamys. Беринговоморский гребешок наиболее обычен на глубинах от 50 до 100 м, встречается также у Тихоокеанского побережья Америки до Калифорнии.
В дальневосточных морях и особенно в Баренцевом и Белом морях и в юго-западной части Карского моря (куда проникают с запада более теплые воды) обитает довольно крупный (до 8 см в поперечнике) красивый исландский гребешок (Chlamys islandicus). Он обычен также и у берегов Исландии, Норвегии, Южной Гренландии, у Атлантического побережья Северной Америки. Встречается на глубинах до 100 м и входит в состав некоторых биоценозов донной фауны. Мясо исландского гребешка очень вкусно, но промысла на него в наших морях нет.
Близки к рассмотренному выше семейству морских гребешков моллюски из семейств спондилюсов (Spondylidae) и лим (Limidae).
Лимы имеют изящную овально-ребристую раковину; замковые зубы не развиты, мускул-замыкатель один. Мантийных глаз нет, но имеются чувствительные выросты по краю мантии, часто очень длинные и ярко окрашенные. Имеется также мантийный парус, поэтому лимы могут плавать скачками, как и гребешки. Иногда лимы образуют биссус, с помощью которого некоторые виды, например зияющая лима (Lima hians), могут строить «гнезда», скрепляя биссусом куски ракушки, мелкие камешки, обломки кораллов и пр. После того как этот материал скреплен сетями из биссусных нитей, лима выстилает свой «дом» изнутри нежной пряжей из этих нитей, и он становится похожим на уютное птичье гнездо. По-видимому, лима, живущая в таком гнезде, редко его покидает, выпуская наружу лишь густую подвижную бахрому краев мантии, которые способствуют созданию токов воды для ее питания и дыхания.
Однако многие лимы ведут и свободный образ жизни, как, например, небольшие белые северные лимы — Lima (Limatula) hyperborea, обитающие в северных морях, или многие более глубоководные виды, встречающиеся в открытых частях морей и реже на ложе океанов.
Моллюски из семейства Аномии (Anomiidae) ведут прикрепленный образ жизни. Только что осевшие на скалы и камни молодые аномии прочно прикрепляются к ним биссусом, который позже пропитывается известью, превращаясь в твердый стебелек, выходящий в отверстие в нижней створке раковины. Зубов нет; форма раковины неправильно округлая, так как повторяет обычно неровности камней, к которым плотно прилегает. Обычна для морей теплой и умеренной зон Атлантики Anomia ephippium, а в дальневосточных морях гигантская аномия — Pododesmus (Monia) macrochisma, или Monia macrochisma. Размеры первой — около 1—2 см, а второй — до 9—10 см. Обе обитают на камнях и скалах прибрежной зоны и небольших глубин. Anomia ephippium встречается также в Средиземном море, но в Черное не проникает.
К аномиям близка крупная тропическая плакуна (Placuna placenta), имеющая почти плоскую полупрозрачную раковину 13—14 см в поперечнике. Она живет на скалистых грунтах в Индийском и в западной части Тихого океана. В Индии, Китае, на о. Целебес и на Филиппинах ее створки издавна употребляют вместо стекол в окнах домов. Даже теперь в одной только Маниле для этих и других целей ежегодно добывается около 5 млн. штук плакун; в настоящее время ее здесь даже разводят. Раковины плакун размалывают также в серебристый порошок, из которого приготовляют краску. Мясо плакун нередко употребляют в пищу.
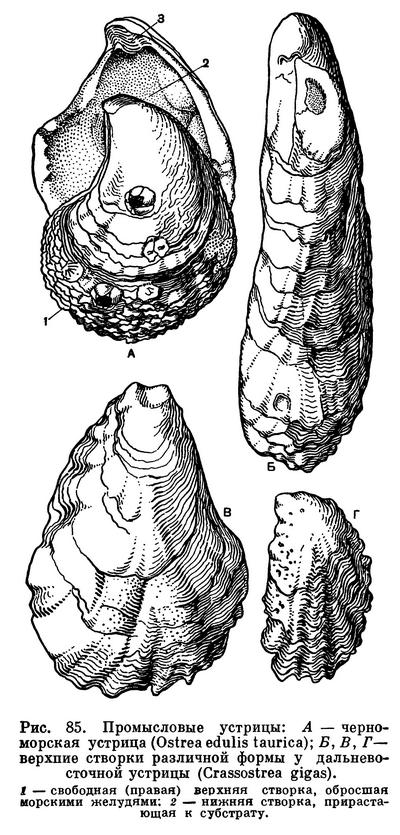
Устрицы (сем. Ostreidae) — одна из самых популярных промысловых групп двустворчатых моллюсков, которые употребляются человеком в пищу с незапамятных времен. Об этом свидетельствует нахождение их раковин в так называемых «кухонных кучах» — отбросах, оставшихся от древних поселений людей, живших еще в каменном веке на берегах морей (Черного, западной части Балтийского моря и др.). Упоминание о промысле устриц находят еще у Плиния (т. е. за 150 лет до начала нашего летосчисления). Все это говорит о том, что высокие вкусовые и питательные качества устриц были известны людям уже очень давно. В результате долгого хищнического истребления устричных банок уже в середине XIX в. встал вопрос о необходимости строгого регулирования промысла устриц и об искусственном их разведении. В Японии культура устриц возникла еще в XVII в.
Устрицы имеют несимметричные грубо-чешуйчатые створки раковин изменчивой формы; при этом левая имеет блюдцеобразную форму, а правая (более плоская) прикрывает ее, как крышечкой. Левой (нижней) створкой устрица прирастает к камням или скалам, а часто и к створкам других устриц или иных сидячих моллюсков. Нога и биссус у взрослых устриц полностью редуцированы.
Известно около 50 видов устриц. Все они тепловодны и не проникают на север далее 66° с. ш. Основным промысловым видом в разных морях служит обыкновенная, или съедобная, устрица (Ostrea edulis), обычная у берегов Европы — от Норвегии до Алжира и в Средиземном море, а в Балтийском море она отсутствует из-за пониженной его солености. Издавна промышляется и разводится: так, во Франции устриц начали разводить в семидесятых годах прошлого столетия.
Настоящая съедобная устрица — очень изменчивый вид (как и вообще большинство видов устриц) и образует различные местные расы и морфы, нередко считающиеся даже отдельными видами, как например адриатическая устрица (Ostrea edulis adriatica), богатые устричники которой были известны издавна, скаловая устрица (Ostrea edulis sublamellosa) и наша черноморская грядовая устрица (Ostrea edulis taurica). У атлантических берегов Франции промышляют также португальскую устрицу (Crassostrea angulata).
В Черном море устрицы встречаются как в защищенных бухтах и на прибрежных скалах, так и на мелководьях более открытых частей моря, образуя в зоне ракушечника биоценозы устричных гряд и банок. Большие устричные банки у Крымского и Кавказского берегов (например, Гудаутская банка) почти не используются промыслом. Кроме того, они сильно пострадали от выедания устриц и мидий хищным брюхоногим моллюском рапаной (Rapana).
В США промышляют и разводят: по Атлантическому побережью виргинскую устрицу (Crassostrea virginica), а по Тихоокеанскому — калифорнийскую устрицу (Ostrea lurida).
У берегов Японии встречается несколько видов устриц, из которых промышляют и разводят гигантскую устрицу (Crassostrea gigas, или Ostrea laperousi), японскую устрицу (Crassostrea nippona), листоватую устрицу (Ostrea denselamellosa) и др. Обычная в Японском море гигантская устрица встречается у берегов Приморья и Сахалина, образуя богатые скопления — устричники, которые у нас еще не используются. В начале текущего столетия эту устрицу перевезли на Тихоокеанское побережье США (штат Вашингтон), где ее широко промышляют.
Гигантская устрица очень изменчива по форме — от длинной продолговатой до овально-укороченной. Много других видов устриц промышляется в Австралии, Новой Зеландии, в Индии и многих других странах.
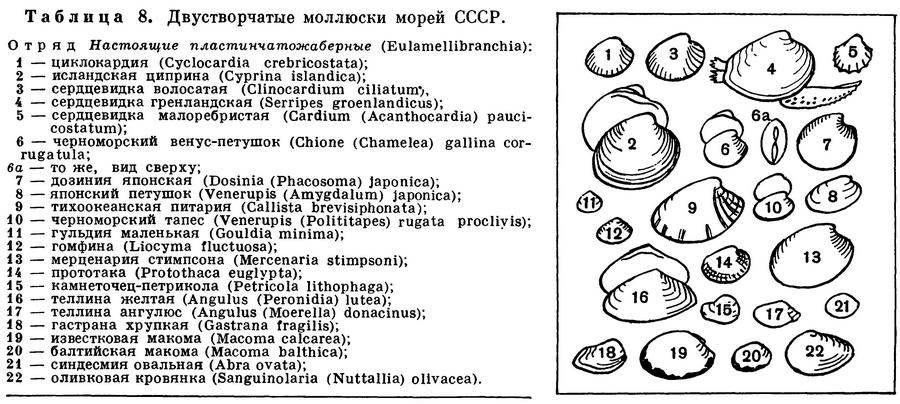
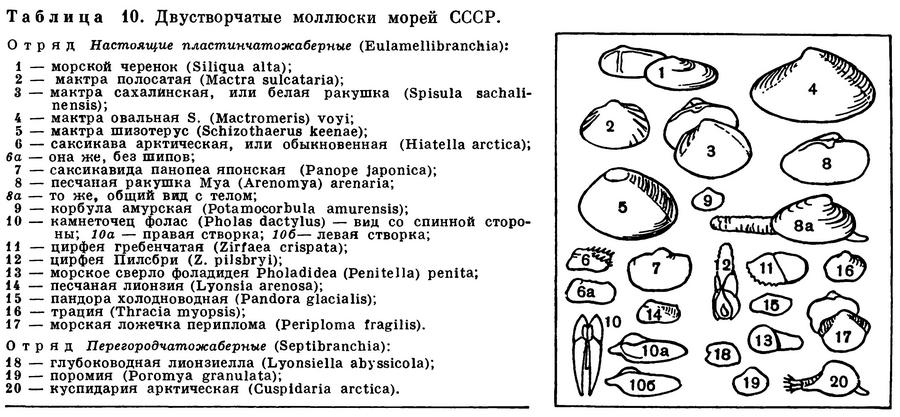
ОТРЯД НАСТОЯЩИЕ ПЛАСТИНЧАТОЖАБЕРНЫЕ (EULAMELLIBRANCHIA)
К отряду настоящих пластинчатожаберных моллюсков относится наибольшее число видов, которые обитают как в морских, так и в пресных и солоноватых водах. Встречаются во всех морях и в океанах на самых различных глубинах — от приливо-отливной зоны и почти до самых больших глубин Мирового океана (10,8 км), в одной из глубоководных впадин в южной части Тихого океана.
Основным признаком отряда является строение их жабр, которые представляют собой две двухслойные решетчатые, сложнопродырявленные полужабры с каждой стороны тела. Каждая полужабра состоит из рядов коленчатоизогнутых жаберных нитей, соединенных между собой соединительноткаными или сосудистыми спайками и образующих внутреннюю и наружную жаберные пластинки с полостью между ними. Спайки имеются и между восходящими и нисходящими коленами самих жаберных нитей.
Замок устроен различно, но у большинства видов он так называемого гетеродонтного типа и состоит из центральных (кардинальных) и боковых зубов; характер, количество и расположение зубов могут варьировать, отдельные части замка или весь замок в целом могут быть редуцированы. Передний и задний замыкательные мускулы одинаково развиты.
Многие настоящие пластинчатожаберные моллюски являются формами инфауны, т. е. обитают в толще различных грунтов, в большей или меньшей степени закапываясь в них. Для глубоко закапывающихся форм характерны уплощенные, гладкие или очень слабо скульптированные раковины, сросшаяся мантия с отверстием для выхода ноги, длинные разделенные или сросшиеся сифоны.
Благодаря таким сифонам, находясь «в укрытии» в толще грунта (спасаясь от врагов всех двустворчатых моллюсков — морских звезд или брюхоногих моллюсков — букцинид, натицид и др.), они могут засасывать необходимую им для дыхания и питания воду из самых придонных слоев, собирать детрит с поверхности грунта.
Формы, ползающие по поверхности грунта или лишь частично и неглубоко закапывающиеся в него, относятся к так называемой онфауне. Они обычно обладают крепкой, вздутой ребристой раковиной с хорошо развитым замком; у них несросшаяся мантия, сильная нога, небольшие или даже зачаточные сифоны. Ребра укрепляют створки раковины, и различные выросты на них препятствуют «проваливанию» животных в грунт и помогают им удерживаться в самых поверхностных слоях грунта.
У форм, закапывающихся, вбуравливающихся в грунт, наблюдается ослабление и даже полная редукция замковых зубов и усиление роли внутренней связки, а сверлящие формы лишены и замка и связки.
Отряд Настоящие пластинчатожаберные включает более ста семейств и разделяется на четыре подотряда: Расщепленнозубые (Schizodonta), Разнозубые (Heterodonta) — самый большой по количеству семейств, родов и видов, Адапедонты (Adapedonta) и Аномальносвязочные (Anomalodesmata).
Обширную группу настоящих пластинчатожаберных моллюсков, целиком приспособившихся к жизни в пресной воде, составляют прежде всего моллюски из большого подотряда расщепленнозубых; из подотряда разнозубых к пресным водам приурочены моллюски из надсемейства Sphaeriacea, а также почти все виды из семейства Dreissenidae.
Все эти указанные группы настоящих пластинчатожаберных широко распространены в пресных водах всех континентов, особенно в водах Америки, где их видовое разнообразие особенно велико.
Большинство видов расщепленнозубых относится к надсемейству Unionacea, или Najadacea, в которое входит 2 семейства: пресноводные жемчужницы (Margaritiferidae) и униониды (Unionidae). У всех у них раковина имеет хорошо развитый перламутровый слой, нога большая, топоровидная, связка наружная, позади макушек; мантия несросшаяся, сифоны зачаточные. Замок, если он имеется, очень изменчивого строения и относится к особому унионидному («псевдогетеродонтному») типу; зубы могут быть более или менее расщепленными, расходящимися и несут функции центральных или боковых зубов.
Очень характерно развитие у Unionacea: молодь развивается у материнских особей внутри жаберных полостей, играющих роль выводковой сумки — марзупиума, где и формируется особая личинка — глохидий, которая затем выходит в воду и закрепляется на жабрах или на коже различных рыб (окунь, чехонь, язь, ерш и др.). Затем глохидии инкапсулируются, паразитируя в течение одного-двух месяцев на рыбах, где и происходит их превращение в молодых моллюсков. На одной рыбе может «выкормиться» огромное количество глохидиев: так, на жабрах одного небольшого окуня длиной около 8 см было найдено около 400 глохидиев. Выйдя из мешочков (капсул), молодь моллюсков оседает на дно.
Унионид употребляют в пищу, но используют главным образом на откорм домашней птицы и скота (свиней); раковины многих их видов, а также и жемчужниц используют для различных изделий из перламутра. В 1963 г. мировая добыча пресноводных двустворчатых моллюсков, главным образом Unionidae (и отчасти Corbiculidae), составляла 350 тыс. ц.
В семейство пресноводных жемчужниц входит всего один род — Margaritifera, или Margaritana, виды которого живут в Северной Америке, Европе и на севере Восточной Азии, в том числе на Сахалине и в Японии.
Это наиболее примитивные формы из Unionacea, имеющие редуцированные боковые зубы; жабры их не срастаются сзади с мантией. У нас известна главным образом обыкновенная жемчужница (Margaritifera margaritifera) длиной до 12 см. Обитает в небольших реках севера нашей страны, а также на севере Европы, Америки и в Японии. Добывается ради жемчуга давно. В реках Дальнего Востока живет несколько видов жемчужниц, например: даурская жемчужница (Margaritifera dahurica) длиной до 18 см, обитающая в бассейне р. Амура и в Приморье (ее издавна промышляли ради перламутра и жемчуга); камчатская жемчужница (Margaritifera middendorffi) длиной 9 см; овальная раковина этой жемчужницы отличается от беззубок значительно большей толщиной створок, мощным перламутровым слоем, наличием замка с одним-двумя центральными зубами.
Оплодотворение, выход яиц и заражение ими жабр у жемчужниц наблюдается в июле — августе. В отличие от перловиц и беззубок их яйца развиваются до глохидиев во всех четырех (наружных и внутренних) полужабрах материнского организма. Глохидии очень мелкие и лишены клювовидного острого зубца; уже в сентябре они встречаются на коже и жабрах рыб (хариуса, горчака и др.).
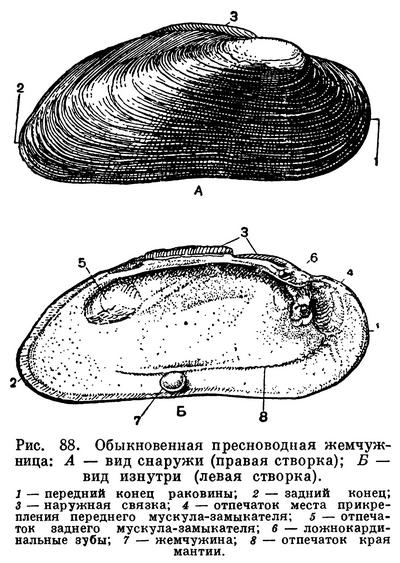
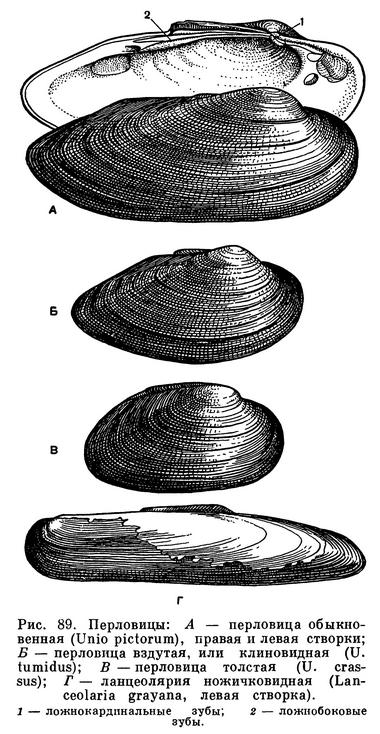
В семейство Unionidae входит около 80 родов, объединяемых в 6 подсемейств, из которых наиболее важны перловицы (Unioninae), беззубки (Anodontinae), а также самое обширное из них подсемейство лампсилин (Lampsilinae), куда входит более 20 родов и около 600 видов, обитающих исключительно в Северной и Центральной Америке, преимущественно в реках, впадающих в Тихий океан.
Перловицы широко распространены в Европе, Азии, Африке, Северной и Центральной Америке, Японии, на Малайском архипелаге.
Наибольшее разнообразие перловиц наблюдается в Восточной Азии, где известно около тридцати их родов и большое число видов; в Северной Америке известно около 12 родов, в Африке 3 рода, а в Европе 1 род — настоящие перловицы (Unio, или Nodularia).
Настоящие перловицы (Unio) обладают крепкой раковиной, хорошо развитым замком с одним-двумя центральными и хорошо развитыми пластинчатыми боковыми зубами. Наружные полужабры сзади до конца срастаются с мантией, а края мантии срастаются только над верхним (выводным) сифоном; верхний и нижний (вводной) сифоны разделены перегородкой.
В Европейской части СССР хорошо известны и широко распространены три очень сильно изменчивых вида. Перловица обыкновенная (Unio pictorum), длиной до 14,5 см. Длинная узкая раковина ее с почти параллельными спинным и брюшным краями. Обитает в реках и озерах всей Европейской части нашей страны, в Средней и Северной Европе.
Перловица клиновидная, или вздутая (Unio tumidus), с выпуклой, суженной на заднем конце раковиной, длиной до 11 см. Ее распространение сходно с предыдущим видом.
Перловица толстая (Unio crassus), с овальной, довольно вздутой раковиной, нередко с розовым перламутровым слоем внутри. Обитает в реках Европы и Европейской части СССР, в озерах обычно не встречается.
В реках Дальнего Востока, в озере Ханка, в Приморье наиболее хорошо известна: дальневосточная перловица (Unio douglasiae), а также ножичковидная ланцеолярия (Lanceolaria grayana), с сильно вытянутой почти цилиндрической раковиной, закругленной спереди и суженной сзади. Оба эти вида обитают также и в Китае, а дальневосточная перловица — также и в Японии.
В Западной и Центральной Сибири в настоящее время перловиц нет; богатая их фауна, бывшая здесь в плиоцене, вымерла во время ледникового периода, а физические преграды (горные цепи и большие сибирские реки), видимо, препятствовали проникновению в Сибирь амурских видов перловиц. Отсутствие их в среднеазиатских реках (Аму-Дарья и Сыр-Дарья) объясняется, видимо, большой мутностью воды этих рек.
Обнаружить перловиц в реке или в озере нетрудно по их следам — бороздкам, которые они оставляют на мягком грунте дна. Наиболее благоприятен для них илисто-песчаный грунт; вязкого ила они избегают.
Подсемейство беззубок (Anodontinae) охватывает значительное разнообразие форм пресноводных двустворчатых, большинство родов и видов которых живет в Северной и Центральной Америке, но где они все же значительно уступают по количеству видов лампсилинам (особенно) и перловицам.
Беззубки лишены замковых зубов, их раковины большей частью тонкостенные, с довольно слабым перламутровым слоем; края мантии образуют лишь зачаточные сифоны; их молодь — эмбрионы — вынашивается с осени в течение всей зимы (в период так называемой жаберной беременности) в наружных полужабрах. Перед этим периодом у них развиваются особые вентиляционные трубочки, проводящие воду и значительно улучшающие снабжение кислородом развивающихся в жабрах эмбрионов. Выбрасывание обычно довольно крупных глохидиев, вооруженных двумя (по одному на каждой створке) крючками, происходит весной.
У нас хорошо известна беззубка обыкновенная европейская (Anodonta cygnea), достигающая 20 см длины. Живет она в Средней и Северной Европе, в Европейской части СССР и в Западной Сибири, обитая в прудах, озерах и речных заводях и затонах. Это очень изменчивый вид, образующий (чуть ли не в каждом большом водоеме, районе и т. п.) различные морфы, экологические расы, разновидности. Некоторые из них считаются нередко даже отдельными видами, как, например: живущая в озерах и речных старицах вытянутая беззубка (Anodonta cygnea cellensis), обитающие в озерах и реках рыбья беззубка (Anodonta cygnea piscinalis) и утиная беззубка (Anodonta cygnea anatina).
В реках Европейской части СССР обитает также и узкая беззубка (Pseudanodonta complanata) с очень сжатой, удлиненно-овальной раковиной; проникает в сильно опресненные участки наших внутренних морей. На Кавказе в реках, старицах, озерах с прозрачной водой обитает кавказская беззубка (Anodonta cyrea).
На Дальнем Востоке, в Приморском крае, а также в Китае в реках и озерах обитают наиболее крупные виды из рода анодонт с сильно вздутой раковиной — беззубка Вуда — Anodonta (Pteranodon) woodiana, до 15 см длиной, и сводчатая беззубка — Anodonta (Haasiella) arcaeformis, до 10 см. В бассейнах рек Амура и Уссури, в озере Ханка, озерах Монголии и реках Китая, в Японии обитает наиболее крупный вид пресноводных двустворчатых моллюсков гребенчатка (Cristaria plicata), одна из разновидностей которой (Cristaria plicata v. herculea) достигает 34 см длины и веса 1 кг. Встречается она на глубине 1,5—2 м, на мягких илистых грунтах, в хорошо прогреваемых и защищенных участках озер или в реках со слабым течением. В Китае гребенчатку используют для добычи перламутра. У гребенчатки широкая, довольно толстостенная раковина, у которой спинные края образуют высокий плоский гребень. Замковые зубы рудиментарны. В озере Ханка гребенчатка может встречаться в количестве до 10 экземпляров на 1 м2 дна.
Лампсилин (Lampsilinae) в США не только вылавливают, но довольно давно искусственно разводят, особенно некоторые быстро растущие виды из родов Lampsilis, Obovaria и др. При этом рыб искусственно заражают их глохидиями. У многих лампсилин наблюдается половой диморфизм, выраженный даже в форме их раковины. Выводковая камера образуется в задней части наружных полужабр.
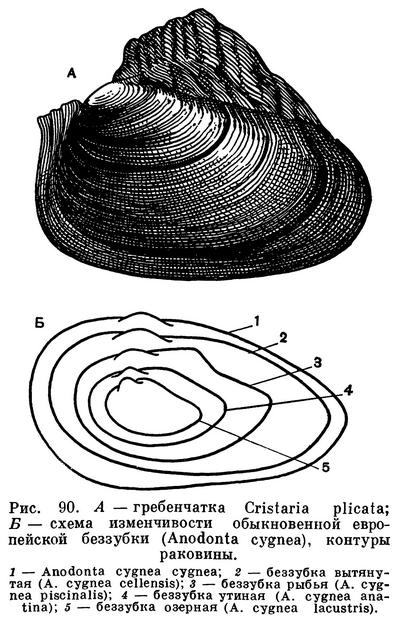
Кроме перловиц и беззубок, постоянными обитателями наших пресноводных водоемов являются мелкие моллюски из подотряда разнозубых, из надсемействa Sphaeriacea. Это прежде всего такие широко известные и обычные обитатели прудов, рек, озер и луж, как шаровки (Sphaerium) и горошинки (Pisidium) из семейства Sphaeriidae.
Шаровки — это мелкие двустворчатые с округлой раковинкой, которая у самых крупных видов — у речной шаровки (Sphaerium rivicola) — достигает лишь 2,5 см длины. Шаровки «живородящи»: яйца развиваются у них в особых выводковых камерах, которые образуются на внутренних жабрах, так что из материнской раковины выходят уже готовые молодые ракушки. Интересно отметить, что количество этой молоди обычно не превышает десятка, в то время как глохидии у перловиц выбрасываются сотнями. У шаровок забота о потомстве оказывается весьма эффективной, так как обеспечивает им выживание и расселение вида. Гермафродиты.
Шаровки встречаются в Европе, Азии, Америке, Африке. Из обитающих в речных водах в пределах СССР наиболее обычны речная шаровка (Sphaerium rivicola), прочная шаровка (Sphaerium solidum), некоторые разновидности роговой шаровки — Sphaerium corneum; остальные разновидности этой шаровки обитают в стоячих водоемах.
Самые мелкие представители семейства горошинки (Pisidium) населяют наши пресные водоемы, как стоячие, так и проточные, почти повсеместно, от рек, озер до прудов, болот и пещер.
Горошинки чрезвычайно разнообразны и изменчивы и образуют большое количество видов и подвидов, чему, видимо, способствует изолированное их местообитание в отдельных водоемах. Наиболее обычны речная горошинка (Pisidium amnicum), блестящая горошинка (Pisidium nitidum), болотная горошинка (Pisidium casertanum) и др.
Кроме шаровок и горошинок, к Sphaeriacea также относится семейство Corbiculidae, виды которого обитают в пресных и солоноватых водах субтропиков и тропиков. Крупные виды корбикул промышляют. У нас в Закавказье и в Средней Азии в озерах, реках, оросительных каналах, а также в заливах Южного Каспия обитает западная корбикула (Corbicula fluminalis), а в Приморье — восточная корбикула (Corbicula fluminea).
Своеобразную группу представляют собой небольшие пресноводные и солоноватоводные моллюски дрейссены (Dreissena, или Dreissensia) и конгерии (Congeria) из семейства Dreissenidae. Хотя дрейссены по форме раковины напоминают мелких митилид, однако они им не родственны, а относятся к отряду настоящих пластинчатожаберных (Eulamellibranchia). Дрейссены имеют заостренный передний конец раковины и расширенный задний. Замковых зубов нет. Внутри, в самой передней части раковины образуется перемычка (септа), к которой прикрепляется передний небольшой мускул-замыкатель. Мантия имеет отверстие для выхода ноги и образует два коротких сифона; нога с биссусом. Развитие со свободно плавающей личинкой.
Дрейссены раздельнополы. Яйца выметываются прямо в воду в небольших слизистых комочках, в которых содержится много мелких яиц. После их оплодотворения развивается личинка велигер, которая плавает короткое время, а потом оседает на дно, после чего прикрепляется биссусом к субстрату. Наличие плавающей личинки — черта, свойственная чисто морским моллюскам, и это указывает на морское происхождение дрейссен.
Речная дрейссена (Dreissena polymorpha) имеет зеленовато-желтую очень изменчивой формы раковину до 3—5 см длиной, нередко с рисунком из поперечных или зигзагообразных коричневых полосок; образует много подвидов.
Дрейссена живет до 7—8 лет. Половые продукты у нее созревают в апреле, после чего начинается порционное икрометание, которое продолжается все лето, благодаря чему ее личинки (велигер) наблюдаются в планктоне водохранилищ с конца мая (когда температура воды повышается до 15—20°) до глубокой осени.
Кроме речной дрейссены, имеется еще ряд видов дрейссен, свойственных только Каспийскому или Аральскому морю (так называемые эндемичные формы, т. е. нигде больше не встреченные).
В Каспийском и Аральском морях все дрейссены поедаются донными рыбами.
В Каспийском море обитают: каспийская дрейссена (Dreissena caspia), с более мелкой, узкой, удлиненной раковиной, живущая при солености 5—11‰, и морская клювовидная дрейссена (Dreissena rostriformis, или Dreissena distincta), более крупная, овально-каплевидной формы, с маленькими острыми отвернутыми в разные стороны носиками, живущая при солености до 12,7‰, и др. Считают, что в настоящее время каспийская дрейссена вымерла. Другие представители Dreissenidae живут в пресных или солоноватых водах Америки, Африки, Индии.
Возвращаясь к рассмотрению чисто морских моллюсков из отряда настоящих пластинчатожаберных, начнем с наиболее массовых форм подотряда разнозубых (Heterodonta).
Одной из обычных и массовых групп двустворчатых моллюсков морей северного полушария являются различные астарты (сем. Astartidae). Для них характерна большая внутривидовая изменчивость, как и для других моллюсков, которые образовывают массовые поселения. Астарты в целом связаны с холодными или умеренными водами северного полушария, где они живут главным образом на шельфе (до глубин 200 м). Несколько видов известно также в Антарктике.
Замок у всех астарт имеет довольно сходное устройство: 2 центральных (кардинальных) зуба и по одному слабому боковому в каждой створке.
Астарты не имеют сифонов, поэтому не могут глубоко закапываться в мягкий грунт, на поверхности которого они обычно живут. Молодь всех астарт служит пищей многим донным рыбам (камбалам, треске, пикше). Астарты играют большую роль в сообществах донной фауны Северной Атлантики, наших северных и дальневосточных морей.
Близки к астартам кардиты, венерикардии и другие кардитиды (Carditidae), но они обладают более мощным замком, обычно с удлиненными задними кардинальными зубами. Живут главным образом в теплых и тропических водах. Очень красивые и ярко окрашенные раковины у представителей родов Cardita и Glans. На мелководьях Берингова и Охотского морей обитает несколько видов циклокардий: Cyclocardia borealis, Cyclocardia crebricostata, Cyclocardia paucicostata и др.
В Средиземном море и в Атлантическом океане живет интересный моллюск из семейства Glossidae — морское сердце (Isocardia cor, или Glossus rubicundus), по форме раковины действительно напоминающий человеческое сердце. Раковина тонкая, вздутая, размером до 5 см, имеет выпуклые сильно закрученные внутрь макушки.
Исландская циприна (Cyprina islandica, или Arctica islandica) из семейства Arcticidae представляет интерес как единственный современный представитель рода; тепловодный североатлантический европейский вид, обитающий в наших северных морях — в западной прибрежной части Баренцева моря, а также и в наиболее теплых районах Белого моря. Циприна имеет крупную (до 12 см) выпуклую округлую раковину, покрытую коричневым лоснящимся периостраком. Замок сложный, с хорошо развитыми кардинальными и боковыми зубами.
Мелкие циприны служат пищей ряду промысловых донных рыб; крупными питаются многие морские птицы, например серебристая чайка. Птица, нырнув, достает со дна крупную циприну, а затем, взлетев в воздух, роняет ее на прибрежные камни, о которые разбивается раковина даже крупных циприн. После этого мясо моллюсков поедается птицами.
Богатое видами (около 200) семейство Люциниды (Lucinidae) распространено главным образом в тропических морях. Они играют здесь большую роль в питании рыб, а некоторые виды используются в пищу и человеком. Много крупных и красивых видов с нежно окрашенными раковинами, как например Codakia tigerina, достигающая размера 12 см, обитает в Атлантике, в Тихом и Индийском океанах.
В Черном море встречается лишь два мелких вида — Divaricella (Lucinella) divaricata и Loripes lacteus.
В питании многих донных рыб заметное место занимают мелкие моллюски из близкородственного люцинам семейства Thyasiridae — тиазиры (Thyasira, или Axinus) и аксинопсис (Axinopsis). Они обладают хрупкой округлой раковиной, нередко с радиальной складкой, лишенной зубов, и не имеют сифонов; нога червеобразная с расширением на конце, способная очень сильно вытягиваться на расстояние в 4—5 раз большее, чем сам моллюск. При помощи такой ноги они закапываются в мягкий грунт приблизительно на глубину, равную длине своей вытянутой ноги. При этом они строят два трубчатых хода до поверхности грунта и выстилают их слизью, выделяемой ногой. Эти ходы выполняют для них роль вдыхательного и выдыхательного сифонов и прочищаются ногой.
Примером моллюсков-комменсалистов могут служить многие мелкие моллюски из Leptonidae и Montacutidae, живущие на жабрах и ножках некоторых десятиногих раков, на иглокожих, в трубках червей и т. п.
Очень красивы хамы (Chama) из семейства Chamidae, которые живут в тропических морях, прирастая одной из створок раковины к скалам и камням.
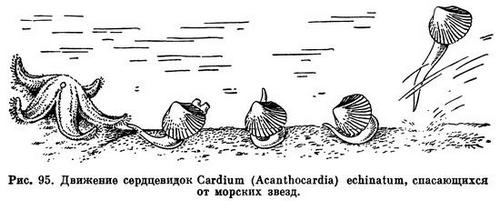
Сердцевидки, или кардиумы (сем. Cardiidae), — очень большая группа мелководных, преимущественно тропических и субтропических, форм, распадающихся на 5 подсемейств, куда относится более 20 родов и много видов. Для них характерна сердцевидная раковина с радиальными ребрами, часто украшенными шипами или чешуйками, или имеющая острое ребро («киль») на задней стороне раковины. Замок хорошо развит, обычно с двумя кардинальными и с передним и задними боковыми зубами в каждой створке, или зубы могут быть частично редуцированы, как у группы солоноватоводных Limnocardiinae и др.
Сердцевидки обладают сильной клиновидной ногой и с ее помощью не только передвигаются и быстро закапываются в грунт, но и подпрыгивают на несколько сантиметров над дном, когда им грозит опасность со стороны их врагов, в первую очередь со стороны морских звезд. Из-за своих коротких сифонов они не могут закапываться глубоко, нередко выставляя над грунтом заднюю половину раковины с опущенными сифонами.
Сердцевидки, особенно мелкие или молодые формы, охотно целиком поедаются рыбами, а у крупных форм рыбы нередко откусывают лишь их ноги.
Сердцевидки съедобны, и ряд их видов промышляют. Так, в 1962 г. в Испании их было добыто 99 тыс. ц, а в Голландии — 24 тыс. ц.
Особенно красивы и разнообразны по форме, окраске и «вооружению» ребер раковины сердцевидок, живущих в мелководных тропических районах, на коралловых рифах и у островов Тихого, Индийского и Атлантического океанов.
Сердцевидка Ламарка (Cerastoderma edule lamarcki) обладает более косо-овальной или даже округло-ромбической раковиной с килевым перегибом в задней части раковины, с менее уплощенными, а в задней части даже угловатыми ребрами; обитает как у берегов Европы, так и в Средиземном, Адриатическом, Черном, Азовском, Каспийском морях, часто в эстуариях, лиманах, лагунах, при солености от 2—4 до 40‰, на песчанисто-илистых и даже смешанных грунтах, до глубины 10—15 м, реже — глубже. По форме раковины и по размерам сердцевидка Ламарка очень изменчива: в Азовском, Каспийском и Аральском морях она сильно мельчает. В Балтийском море Cerastoderma edule заметно мельчает при понижении солености.
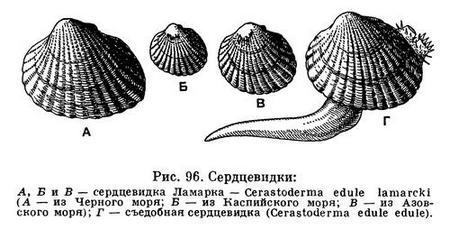
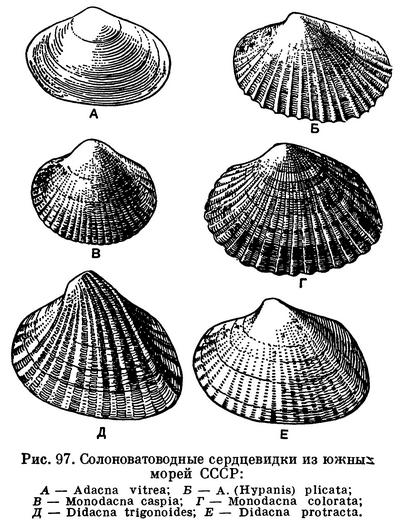
Особого упоминания заслуживает крупная (до 8—10 см длиной) холодноводная гренландская сердцевидка (Serripes groenlandicus), с гладкой округлой раковиной и с малозаметными ребрами на переднем и заднем ее концах. Окраска взрослых темная, сероватая, а молодых более светлая, с зигзагообразным рыжеватым рисунком; такие формы ранее считали отдельным видом и называли гренландской афродитой.
Большой интерес представляет группа сильно изменчивых видов сердцевидок, обитающих в Каспийском, Аральском и опресненных участках и лиманах Азовского и Черного морей из родов Дидакна (Didacna), Монодакна (Monodacna) и Адакна (Adacna). Их нередко выделяют даже в подсемейство Limnocardiinae, поскольку они обитают в солоноватых водах; у них нет боковых зубов и наблюдается редукция кардинальных зубов. Это реликты более богатой сарматской морской фауны, обитавшей в районе наших южных морей в прежние геологические эпохи. Уже в плиоцене Понтическое озеро-море имело богатую дидакнами и монодакнами фауну.
Все виды лимнокардиид, кроме крупных дидакн, служат пищей многим каспийским донным рыбам, в том числе осетровым.
Виды рода Didacna — ныне эндемики Каспийского моря (т. е. обитают только в нем). У них довольно крупная (до 5—6 см) треугольно-овальная или овальная раковина, с ясным килеватым перегибом перед задней частью раковины, с плоскими радиальными ребрами, с двумя, реже одним, кардинальными зубами; сифоны зачаточные, синуса нет. Из нескольких ныне живущих в Каспийском море (кроме сильно опресненных районов Северного Каспия) дидакн наиболее обычны и многочисленны Didacna trigonoides, Didacna barbot de marnyi, Didacna crassa и Didacna protracta. Первая встречается преимущественно в мелководной прибрежной зоне, на песчаных и смешанных твердых грунтах, а последняя из указанных видов — на мягких заиленных грунтах, на глубине около 50 м и более.
Монодакны обладают округло-овальной, зияющей сзади раковиной, обычно с плоскими радиальными ребрами, с небольшими сифонами и неглубоким синусом; у монодакн обычно один конусовидный кардинальный зуб, а иногда и рудимент второго (на правой створке). В Каспийском море широко распространена Monodacna caspia, особенно изобилующая в северной его части. Обитает при солености от 0,1 до 12—14‰. Встречается также в лиманах Черного моря. В Таганрогском заливе Азовского моря, при солености не более 4‰, живет более крупная (до 4 см) Monodacna colorata, которую недавно нашли и в Северном Каспии. Она акклиматизировалась в Веселовском водохранилище, и ее переселили в Цимлянское водохранилище для улучшения его кормности для рыб.
Настоящие адакны (Adacna) имеют также не килеватую, зияющую спереди и сзади, почти гладкую, с очень низкими плоскими ребрами раковину, обычно лишенную зубов или с одним рудиментарным зубом; сифоны длинные, синус глубокий.
В Каспийском море на илистых грунтах, на глубине до 40 м и более, обитают адакны с овально-треугольной, хрупкой, нежной раковиной — Adacna vitrea, а также Adacna minima, живущая и в Аральском море.
Наиболее крупная из адакн — Adacna laeviuscula (до 4 см), с хрупкой, косо-овальной раковиной, живет главным образом в Северном Каспии при солености до 10‰. В Каспийском море и в лиманах Черного моря при солености до 8—14‰ живет небольшая, до 3—4 см, адакна с многочисленными раздвинутыми резкими острыми ребрами — Adacna (Hypanis) plicata.
Особый интерес представляют близкородственные сердцевидкам виды тридакн (сем. Tridacnidae). Это типичнейшие обитатели тропических морей, особенно характерные для зоны распространения коралловых рифов в Индо-Тихоокеанской и Австралийской областях. К этому семейству относится два рода: собственно тридакна (Tridacna) и гиппопус (Hippopus), охватывающие около двух десятков видов. Поистине «царь-ракушкой» среди всех современных двустворчатых моллюсков по своим размерам и мощности раковины является гигантская тридакна (Tridacna gigas), достигающая иногда 1,4 м длины и общего веса 200 кг, причем вес только ее тела достигает 30 кг. Однако обычные размеры тридакн меньше — около 1 м, чаще 30—50 см.
Тридакны интересны по своему внутреннему строению и по симбиозу с одноклеточными водорослями зооксантеллами, что встречается у моллюсков крайне редко.
По своему образу жизни среди тридакн различают свободно лежащих на дне, куда относятся более крупные формы (Tridacna derasa, Tridacna gigas, Hippopus hippopus и др.); все они могут сильно обрастать другими животными и водорослями. Другие, более мелкие, как, например, Tridacna crocea, имеют сильный биссус и широкое отверстие для его выхода между створками раковины на спинной стороне. Погружаясь в расселины между кораллами, механически «вбуравливаясь» в щели, эти тридакны образуют углубления, где и сидят макушкой вниз, время от времени механически расширяя их и прикрепляясь внутри этих углублений своим биссусом.
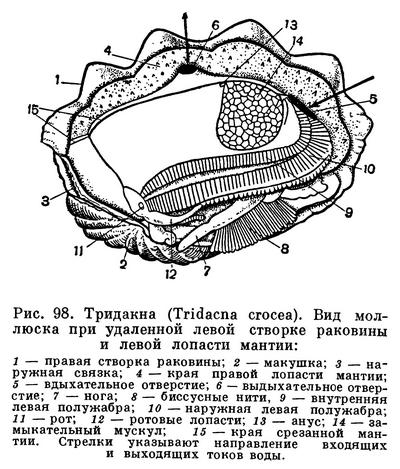
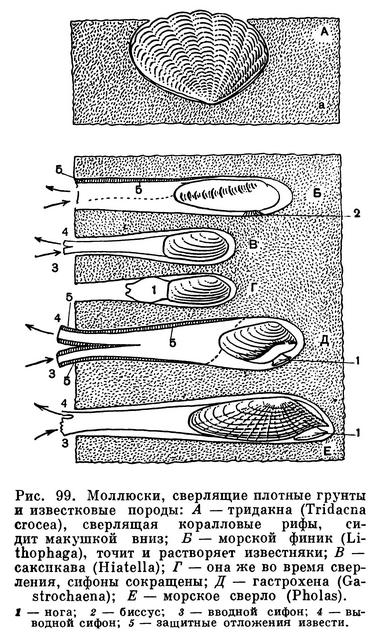
Наш обзор будет неполным, если не рассмотреть хотя бы вкратце такие богатые видами, преимущественно тропические и южнобореальные, группы разнозубых моллюсков, как семейства Венериды (Veneridae), включающее более 400 видов, и Теллиниды (Tellinidae) — более 200 видов.
Венериды (различные венусы, тапесы, венеруписы, дозинии, прототаки, мерценарии и т. д.) широко распространены в умеренно тепловодных и тропических морях, обитая на мелководьях прибрежной зоны, преимущественно на песчаных грунтах. Не обладая длинными сифонами или даже совсем лишенные их, венериды не могут глубоко закапываться и живут лишь в самом поверхностном слое грунта, активно ползая при помощи своей довольно сжатой клиновидно-топоровидной ноги. Раковины венерид очень красивы, различной величины и формы. Ребра раковин иногда могут быть пластинчатыми с бугорками и даже с причудливыми выростами, как, например, у королевского гребня — Pitar dione. Замковая пластинка крепкая, с тремя кардинальными зубами, удлиненными или в той или иной мере расщепленными, могут быть и передние боковые зубы.
Большинство венерид съедобно, и многие их виды являются промысловыми, а в последнее время венерид начали также разводить искусственно и акклиматизировать в новых для них районах.
В Черном море живет несколько атлантическо-средиземноморских видов венерид. Они обитают у берегов, главным образом в зонах биоценоза песка, илистых прибрежий ракушечника (особенно) и мидиевого ила, на глубинах до 50—70 м (реже несколько глубже).
Одним из самых обычнейших, известных и массовых видов моллюсков является черноморский венус-петушок — Chione (Chamelea) gallina corrugatula.
He менее известны и обычны черноморские венеруписы — Venerupis (Polititapes) rugata proclivis и Venerupis (Polititapes) discrepans.
Очень близко к венеридам семейство петрикол (Petricolidae), к которому относится самый обычный камнеточец Черного моря, средиземноморско- атлантический вид Petricola lithophaga, сверлящий здесь известковые породы, прибрежные скалы, а также плотные глины. Благодаря такому образу жизни раковина петриколы может быть сильно деформирована. Мантия у петрикол сросшаяся, с небольшим отверстием для выхода небольшой ноги; сифоны длинные, также частично сросшиеся. Местами петриколы встречаются на берегах в огромном количестве, и поэтому берега здесь разрушаются волнами гораздо быстрее. Для сверления моллюску служит очень крепкая овально-треугольная раковина, с тонкими многочисленными мелкими радиальными гранулированными ребрами; такая структура раковины напоминает поверхность рашпиля; закрепившись ногой в какой-нибудь точке и делая вращательные движения, петрикола медленно соскабливает слой за слоем породу этими зубчиками, постепенно погружается в протачиваемый ею ход и остается в нем.
Обширная группа умеренно теплолюбивых и тропических форм, объединяемых в надсемейство Tellinacea, распадается на четыре семейства: собственно Теллины (Tellinidae), Скробикулярии (Scrobiculariidae), Донаксы (Donacidae) и Псаммобии (Sanguinolariidae). Все они имеют относительно тонкую и более легкую, чем у венерид, овальную или удлиненную раковину. Замок состоит в основном из двух центральных кардинальных зубов в каждой створке, из которых один нередко раздвоен сверху, часто есть и боковые зубы. Нога языковидная, подвижная, без биссуса. Синус глубокий, сифоны длинные, разделенные и очень подвижные. Последние очень помогают им в собирании пищи — детрита — с поверхности мягкого илисто-песчаного или песчаного грунта, когда моллюск глубоко закапывается в него. Раковина теллинид часто бывает красиво окрашена и снаружи и изнутри, как, например, у ярко-розовой тропической Pharaonella perieri или как у атлантическо-средиземноморских и живущих также и в Черном море видов: желтая — у хрупкой гастраны (Gastrana fragilis), розовые — у мелких ангулюсов — Angulus (Fabulina) fabula и Angulus (Moerella) donacinus.
Благодаря небольшим размерам и тонкой раковине почти все Tellinacea — излюбленная пища придонных рыб (камбал и др.); многие Tellinacea съедобны.
Как указывалось выше, в наших северных морях из семейства Tellinidae обитают только виды рода Macoma; у видов этого рода отсутствуют боковые зубы, и раковины их обычно белые, покрытые очень тонким серым периостраком. В морях северного полушария, в том числе и в наших северных и дальневосточных морях, наиболее широко распространена умеренно холодноводная известковая макома (Macoma calcarea). Она нередко образует здесь массовые скопления на мягких заиленных песках, на глубине до 100—120 м; ее количество может достигать нескольких сот экземпляров на 1 м2, а биомасса — нескольких сот граммов на 1 м2. Macoma calcarea входит в состав многих сообществ донной фауны, являясь их руководящей, массовой или одной из самых характерных форм.
Едва ли не самой известной формой среди маком является балтийская макома (Macoma balthica).
Одной из самых крупных теллинид, обитающих во всех наших дальневосточных морях, а также в Чукотском море и вдоль западных берегов Северной Америки, является Angulus (Peromelia) lutea (до 10 см длиной).
Очень красивы маленькие (не более 2—3 см) округло-треугольные, с макушкой, сильно сдвинутой к заднему краю, блестящие раковинки донаксов (семейство Donacidae). Они бывают окрашены в самые различные цвета, нередко с лучистыми радиальными полосками, благодаря чему их называют морскими бабочками. Около 50 видов этого семейства — обитатели песчаных мелководий умеренной и тропической зон. У нас в Черном море живут два вида донаксов, из которых Donax (Paradonax) venustus наиболее обычен.
Значительно более крупны (до 8—10 см длиной) различные псаммобии (семейство Sanguinolariidae, или Psammobiidae, или Gariidae, или Asaphidae).
Около 100 видов этого семейства обитает главным образом на небольших глубинах и илистых грунтах, преимущественно в тропиках. Их удлиненно-округлые раковины обычно зияют спереди и сзади и окрашены в коричневые и фиолетовые тона. У берегов в южной части Охотского моря и в Японском довольно обычна оливковая кровянка (Sanguinolaria (Nuttallia) olivacea).
Из семейства Scrobiculariidae (или Semelidae) наиболее известны различные синдесмии (Abra, или Syndesmya). Из двух атлантическо-средиземноморских видов синдесмий, обитающих в Черном море, овальная синдесмия (Abra ovata) обитает также и в Азовском море. Это мелкий моллюск (до 2,5 см), с треугольно-овальной тонкой, слегка приоткрытой спереди и сзади раковиной; живет на илистых грунтах и небольших глубинах (не более 20—40 м); может выносить значительное понижение солености (до 2‰), предпочитает жить даже в более опресненных участках. Может также выносить и недостаток кислорода в воде, перенося даже временные заморы, наблюдающиеся иногда в центральной части Азовского моря (в районе, занятом биоценозом этой синдесмии).
Для улучшения кормовой базы осетровых рыб синдесмия была пересажена из Азовского моря в Каспийское. Она здесь хорошо прижилась, расселилась и теперь вошла в пищевой рацион многих каспийских донных рыб. Наибольшая плотность поселений синдесмий в Каспии достигает 7000 экземпляров на 1 м2, а биомасса — 300 г на 1 м2.
К подотряду Adapedonta относится пять надсемейств: Черенки (Solenacea) — с одним семейством, Мактры (Mactracea) — с четырьмя семействами, Мии (Myacea) — с тремя семействами, Саксикавы (Saxicavacea, или Hiatellacea) — с одним семейством и Бессвязочные (Adesmacea) — с двумя семействами. Их объединяет большая или меньшая редукция замка вплоть до потери всех зубов и усиление роли внутренней связки. Многие виды из основных семейств, относящихся к первым трем группам, являются объектами промысла, а также играют большую роль в питании рыб.
Большинство видов из этих групп — закапывающиеся формы, а бессвязочные представляют собой специализированные группы моллюсков — камнеточцев и древоточцев.
Черенки (сем. Solenidae) — типичные обитатели песчаных грунтов, в которые они закапываются при помощи своей сильной расширенной на конце ноги. Их раковины очень характерной формы: тонкостенные, вытянутые, узкие, с параллельными спинным и брюшным краями, почти прямоугольные (реже удлиненно-овальные), зияющие с обоих концов, с сильно сдвинутой на передний край макушкой.
В Черном море на глубине до 10 м живет атлантическо-средиземноморский вид обыкновенный черенок (Solen vagina, или Solen marginatus), с очень удлиненной (до 12 см) узкой прямоугольной раковиной; макушка на самом переднем конце спинного края.
Настоящие мактры (Mactridae) широко распространены на песчаных мелководьях тепловодных морей бореальной зоны, а также и в субтропической зоне; в арктических морях их нет. Замковая площадка довольно широкая, крепкая; особенно характерен замок: с одним раздвоенным лямбдовидным кардинальным зубом в левой створке, а в правой — с двумя сходящимися к макушке тонкими кардинальными зубами; позади заднего центрального зуба правой створки располагается довольно большая треугольная площадка для прикрепления внутренней связки. Обычно имеются пластинчатые (приближенные к центральным) боковые зубы.
Мактры съедобны и многие служат предметом промысла в США, Японии, Китае и т. д. Их продают в свежем, мороженом, сушеном и особенно в консервированном виде.
Так, у Атлантического побережья США в 1963 г. было добыто 860 тыс. ц мактр, главным образом Spisula solidissima.
Эта крупная (до 12—15 см) мактра обильно заселяет прибрежные песчаные пляжи, начиная почти от уреза воды. После сильных зимних штормов ее миллионами находят в выбросах на берегу, за что и называют прибойной мактрой.
В Японии очень интенсивно добывают несколько видов крупных мактр, больше всего овальную мактру — Spisula (Mactromeris) voyi, затем сахалинскую мактру, или белую ракушку — Spisula sachalinensis, полосатую мактру (Mactra sulcataria) и др.
В Приморском крае промысловые запасы сахалинской мактры весьма значительны, и при ловле драгой один промысловый бот может добыть в среднем 2—3 тыс. штук в день. Эта крупная (до 12 см) белая ракушка здесь типичный обитатель мелкозернистых песков в прибойных бухтах, где живет на глубине до 5 м или несколько глубже. Размножается в июле — августе и быстро растет, достигая к пяти годам среднего размера около 8 см; затем ее рост замедляется; живет 10—13 лет. Ее молодь развивается на больших глубинах, а затем откочевывает к берегам.
Овальная мактра обитает у берегов всех наших дальневосточных морей до глубины около 75 м, а также у Тихоокеанского побережья Канады; полосатая мактра, как и сахалинская, обитает в южной части Охотского и Японского морей, а последняя также и у берегов Китая.
Почти всесветное распространение имеет обыкновенная, или арктическая, саксикава (Hiatella arctica). Это небольшой моллюск с грубой, толстой, неправильной, более или менее удлиненно-прямоугольной, зияющей с обоих концов раковиной, лишенной зубов (может быть лишь рудимент одного зуба). Мантия сросшаяся, сифоны довольно длинные, сросшиеся. Живет свободно на дне или в норках, которые может высверливать в различных грунтах. При сверлении саксикава закрывает отверстия своих сифонов и опирается сократившимися, сжавшимися сифонами в стенки норки.
К надсемейству Myacea относится довольно большое число видов из трех семейств: Мии (Myidae) — 8 родов, Корбулы (Corbulidae, или Aloididae) — 4 рода и Gastrochaenidae — 3 рода.
У всех Myidae замок слабо развит, раковина без зубов или с одним зубовидным выростом в каждой створке; помимо наружной, сильно развита внутренняя хрящеватая связка, которая прикрепляется одним концом к ложкообразному выросту замочного края под макушкой в левой створке, а другим концом — соответственно к полукруглой выемке в правой створке. Мантия сросшаяся, с отверстием для выхода ноги и с длинными покрытыми кутикулой сифонами.
К глубоко зарывающимся в мягкий илисто-песчаный грунт прибрежных мелководий формам относятся хорошо известные в северных и дальневосточных морях крупные моллюски мии: песчаная ракушка — Mya (Arenomya) arenaria с овально-яйцевидной раковиной (длиной до 10 см) и Mya truncata с округло-угловатой, сзади вертикально обрубленной раковиной (до 5—6 см длиной).
Песчаная ракушка — более тепловодный амфибореальный вид, обитатель приливо-отливной зоны и самых мелководных районов прибрежной зоны (не более 10 м глубиной); в арктических морях не встречается. Другой вид — Mya truncata — живет несколько глубже первого и распространен очень широко почти во всех морях северного полушария.
Mya (Arenomya) arenaria легко переносит опреснение воды; Mya truncata придерживается более постоянной и более высокой морской солености.
Песчаная ракушка поступает на рынок в свежем или мороженом виде, реже — сушеная и копченая, но большей частью в консервированном виде; особенно много из нее приготовляют бульона-экстракта. Песчаную ракушку используют также для наживки рыболовной снасти.
Большой интерес представляют камнеточцы и древоточцы из группы лишенных связок моллюсков (Adesmacea) из семейств морских сверлильщиков — фоладид (Pholadidae) и древоточцев (Teredinidae). Это формы, приспособившиеся к обитанию в ходах, проточенных ими в скалах, известняках, плотных породах или в дереве. Изменения в строении тела и раковины, которые претерпела эта группа моллюсков, оказались настолько глубокими, что многие из них утратили свой исходный внешний вид двустворчатых моллюсков, став неузнаваемыми, как, например, корабельный червь (Teredo).
Его длинное голое тело напоминает червя, а не моллюска; маленькая своеобразная раковина, играющая роль сверла, располагается на самом переднем конце тела; поверхность высверливаемого в дереве хода покрыта известковыми выделениями мантии моллюска и образует как бы трубку, защищающую нежное тело.
В связи с приспособлением к механическому сверлению твердых или плотных пород грунта раковины фоладид (Pholadidae) очень крепкие, сложной формы, со своеобразной скульптурой поверхности, неравносторонние (макушка сдвинута к переднему, реже — к заднему концу), со значительным переднебрюшным и задним зиянием. Части створок, граничащие с верхним (замочным) краем, в большей или меньшей степени отогнуты наружу, образуя так называемую макушечную складку. Передняя большая часть раковины вооружена резкими шиповатыми ребрами.
Сифоны у фоладид мощные, сросшиеся, нередко покрытые хитинизированным периостраком. Нога довольно хорошо развитая, короткая, на конце усеченная, с подошвой, служит для прикрепления к стенкам норки во время сверления. Некоторые фоладиды светятся, как, например, встречающийся в Черном море камнеточец фолас — Pholas dactylus. У него светятся пять хорошо ограниченных участков, включая и оба сифона; светящаяся слизь выделяется особыми железами с длинными протоками и при раздражении выпускается через сифон.
Фоладиды довольно крупны, некоторые достигают 10—12 см длины. Живут главным образом в прибрежной зоне, на малых глубинах. Фоладиды могут повреждать не только известковые породы, песчаник, мергель, но и дерево и даже бетон, что способствует более быстрому разрушению последнего в морской воде.
Некоторые виды фоладид проделывают свои ходы также и в плотном иле или в глине: атлантические и средиземноморские Pholas dactylus и Barnea candida, или Barnea spinosa; обитающая по европейскому и американскому побережьям цирфея гребенчатая (Zirfaea crispata); по Тихоокеанскому побережью Северной Америки — Zirfaea pilsbryi и в Японском море — морское сверло — Pholadidea (Penitella) penita.
Не менее известны и обычны сверлящие дерево фоладиды из рода Xylophaga — ксилофаги, как, например, Xylophaga dorsalis, живущая в Атлантическом океане и Средиземном море. Они часто встречаются в кусках плавающей и затонувшей древесины, в верхней оболочке кокосовых орехов и пр. и могут сверлить различные предметы вплоть до изоляции подводных кабелей. Много новых видов ксилофаг было найдено датской экспедицией на «Галатее» в глубоководных впадинах Тихого океана, например Xylophaga grevei, найденная на глубине более 7 тыс. м, что является пока наибольшей глубиной нахождения морских сверлильщиков.
Хотя ксилофаги точат свои ходы в древесине и пропускают большое количество древесных опилок через свой приспособленный к этому кишечник, они не могут переваривать древесину. Таким образом, проточенные ходы в древесине служат ксилофагам лишь для укрытия, как и всем другим сверлящим норки фоладидам.
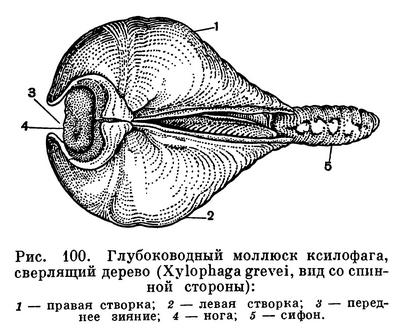
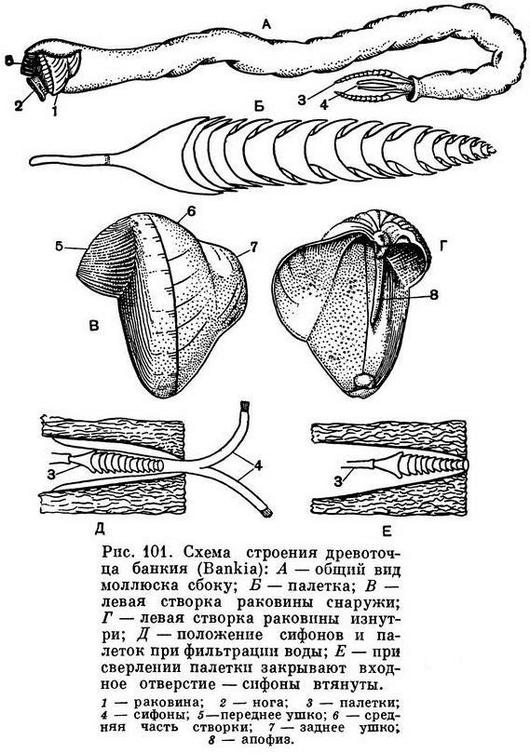
Наиболее совершенные приспособления к протачиванию ходов в древесине наблюдаются у морских древоточцев — весьма специализированных моллюсков из семейства Teredinidae — опасных вредителей деревянных свай, пристаней, подводных частей деревянных судов. Эти моллюски наносят огромный материальный ущерб в теплых и особенно в тропических водах. Морские древоточцы очень широко распространены и встречаются почти во всех морях обоих полушарий, отсутствуя лишь в холодных арктических водах.
Большинство известных в настоящее время видов древоточцев (около 70) относится к двум основным родам — Тередо (Teredo) и Банкия (Bankia).
Обычным местом их обитания служат куски дерева, сваи пристаней, деревянные суда, корневища морских трав, а в тропиках также заросли мангровых деревьев, затонувшие в море кокосовые орехи и др.
Лишь единичные виды древоточцев безвредны, как, например, живущий на песчаных мелководьях и прибрежных пляжах Kuphus polythalamia; он обитает в Тихом океане у Филиппин, на Малайском архипелаге, у Северной Австралии; строит в песке довольно толстые известковые трубки более метра длиной. Безвредны и древоточцы Zachsia, обитающие среди корневищ морской травы зостеры и филлоспадикс.
Створки раковины служат прекрасным инструментом для сверления древесины. Ее передняя часть небольшая, треугольная, выступающая в виде ушка, несет горизонтальные тонко зазубренные гребни; средняя, основная часть вооружена мощными грубозубчатыми гребнями, идущими под прямым углом к гребням передней части. Задняя часть раковины гладкая. На створках изнутри имеется длинный вырост — апофиз. У терединид, как и у фоладид, сокращение заднего и переднего мускулов-замыкателей происходит поочередно. При этом передние концы створок раздвигаются с такой силой, что острые зубчики раковины сдирают слой древесины, как рашпилем. Нога при этом играет роль присоски, которая удерживает моллюска в протачиваемом им ходе; сверлящие движения происходят ритмически — 8—12 раз в минуту. Ходы древоточцев выстланы внутри тонким слоем известковых отложений; поперечный разрез куска дерева, источенного ими, имеет вид пчелиных сот.
Интересно отметить, что у древоточцев их соседние ходы в древесине никогда не пересекаются, как бы густо они ни заселяли зараженную ими древесину. Повороты и изгибы ходов, которые приходится делать моллюскам, чтобы избежать друг друга, несомненно, приводят к еще большему «изрешечиванию» древесины.
Все кусочки древесины, соскобленные раковиной, заглатываются через рот, проходят через пищеварительный тракт, а непереваренные остатки древесины выводятся через выдыхательный сифон наружу.
В средних широтах древоточцы размножаются только летом, а в тропиках круглый год.
В морях СССР обитает несколько видов древоточцев, из которых наиболее известны: в Черном и Балтийском морях — Teredo navalis (до 25 см длиной), распространенныи также в водах Северной Атлантики и северной части Тихого океана; в юго-западной части Баренцева моря — Nototeredo norvagica (до 50 см длиной) и еще более крупный Teredo megotara. Последний также обитает в Северной Атлантике, Средиземном море и в Тихом океане. В Японском море у наших берегов обитает крупная Bankia setacea (длиной до 60 см) и несколько более мелких видов Teredo, в том числе обитающая в зарослях морских трав Zachsia zenkewitchi.
Меры борьбы с морскими древоточцами очень разнообразны, но сводятся в основном к покрытию подводных сооружений слоем специальных красок и к пропитыванию древесины токсическими для моллюсков веществами, в первую очередь креозотом, каменноугольным лаком, карболовой кислотой и др. По отношению к деревянным судам давно практикуется регулярный заход на 3—4 дня в пресную воду в период размножения тередо и оседания его личинок, которые очень чувствительны к сильному опреснению, осушка судов сроком от 5 до 30 дней и некоторые другие методы борьбы.
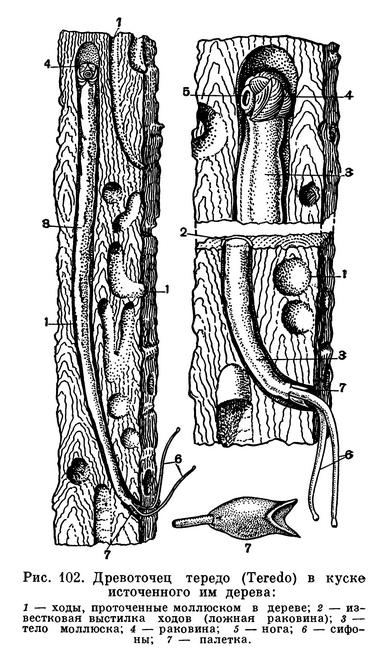
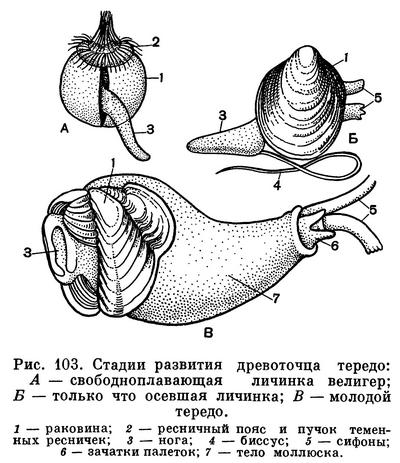
К подотряду аномальносвязочных (Anomalodesmata) относится небольшое число видов, обитающих в различных морях; раковины со слабым замком, часто даже лишенные зубов, с более или менее обызвествленной внутренней связкой; наружная полужабра нередко загнута вверх или редуцирована.
В морях СССР обитают виды только из четырех семейств этого подотряда: Лионзии (Lyonsiidae), Пандоры (Pandoridae), Трации (Thraciidae) и Морские ложечки (Periplomatidae). К отряду аномальносвязочных относят также очень интересных и необычных по своему внешнему виду тропических моллюсков, известных под названием «морских леечек», из семейства Clavagellidae. Их маленькие тонкостенные раковинки срастаются одной створкой (Clavagella) или обеими (Brechites, или Penicillus) с длинной известковой трубкой — ложной раковиной, выделяемой сифонами. Края мантии сросшиеся, с отверстием для ноги; сифоны длинные, сросшиеся. Нижний конец такой трубки расширен в виде диска, вооруженного по краям мелкими трубочками, при помощи которых они закрепляются в грунте, а сама трубка торчит вверх. Верхний конец трубки открытый и несет по краям листовидные выросты.
ОТРЯД ПЕРЕГОРОДЧАТОЖАБЕРНЫЕ (SEPTIBRANCHIA)
К этой интересной, но еще мало изученной группе моллюсков Septibranchia относятся небольшие, преимущественно глубоководные формы. У многих из них жабры маленькие, редуцированные или преобразованные в мускулистую поперечную перегородку — жаберную септу с несколькими отверстиями в ней. Нога без биссуса; внутренний лигамент имеет известковое ядро (литодесму). Наиболее распространенными из этого отряда являются роды и виды семейства куспидарий (Cuspidariidae). Их раковина оттянута сзади в виде более или менее длинного рострума (выроста), где лежат сифоны.
К отряду перегородчашожаберных относятся также еще два семейства: Поромии (Poromyidae) и Вертикордии (Verticordiidae), среди которых имеется много глубоководных видов, обитающих в открытых частях многих морей при высокой (морской) солености и в океанах.
Виды из отряда перегородчатожаберных не встречаются во внутренних морях СССР и при пониженной солености.
Перегородчатожаберные моллюски по способу питания, вероятно, мелкие хищники.