(Sardinops Hubbs, 1929) Род Сардинопсы, Сардины-сардинопсы, Genus Sardinops Hubbs, 1929 (Indo-Pacific sardines, Indo-Pacific pilchards) 1 вид
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Сардины рода Сардинопс (Sardinops)
достигают длины 30 см и веса 150 г и
выше. Тело толстое, брюхо не сжато с боков.
Спина сине-зеленая, бока и брюхо
серебристо-белые, вдоль каждого бока тянется
ряд темных пятен, числом до 15. На
поверхности жаберной крышки есть радиально расходящиеся борозды. Сардинопсы
очень похожи на настоящую сардину-пильчарда, отличаясь от нее
укороченными жаберными тычинками у угла сгиба
первой жаберной дуги, несколько
большим ртом (задний край верхней челюсти
заходит за вертикаль середины глаза) и
характером чешуйного покрова: у сардинопсов все чешуи одинаковые, средней
величины (50—57 поперечных рядов чешуи),
а у пильчардов под крупными чешуями
скрыты более мелкие. Количество
позвонков от 47 до 53.
В роде сардинопс имеется только один
вид — сардина-сардинопс (Sardinops
sagax), состоящий из пяти подвидов.
Сардины-сардинопсы — стайные планктоноядные рыбы, живущие
преимущественно в пределах температуры воды от 10
до 20°С и совершающие кормовые и
нерестовые миграции. Откармливаются у
берегов, для нереста обычно отходят в море.
Множество хищных рыб и птиц кормятся
сардиной. Сардины-сардинопсы наряду
с сельдью и треской — важнейшие
промысловые рыбы мира. Дальневосточная
сардина (японское название ма-иваси)
достигала в 1936—1939 гг. огромной
численности, заходя на север до Камчатки
и давая в то время уловы до 2,4—2,8 млн. т. Особенно много ловили ее
у берегов Японии и Кореи; в Советском
Союзе ловили до 100—140 тыс. т.
Молодая сардина, в возрасте от 2 до 6 лет (17—23 см длины), начинала путь на север
из вод Южной Японии с марта, проходя
до 23—33 км в сутки и появляясь в водах
Приморья обычно с июня до сентября.
Здесь она откармливалась планктоном,
главным образом рачками, а с конца
сентября отходила обратно на юг, имея в
марте наименьший район распространения.
Таким образом, область ее
распространения в связи с сезонными изменениями
температуры воды расширяется летом
(в теплые годы до Камчатки) и
сокращается зимой. Нерест сардины происходит
у берегов Южной Японии с декабря по
март, главным образом при температуре
воды 13—18°С, у Северной Японии позже,
до июня.
Но с 1940 г. численность стада
дальневосточной сардины начала быстро
уменьшаться, по-видимому, в связи с
похолоданием воды, сильно сократившим
воспроизводство. Область распространения
сардины сократилась за счет северных районов,
куда она перестала заходить.
Уловы калифорнийской сардины с 1945 г.
тоже начали сокращаться.
С 1948 г. начал расти промысел
южноафриканской сардины, понемногу
увеличиваются уловы перуанской сардины.
Большие стаи австрало-новозеландской
сардины еще ожидают развития их
промысла.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
6. Род SARDINOPS HUBBS, 1929
Sardinops Hubbs, Proc. Calif. Acad. Sci., 18, 11, 1929 : 264 (тин: Meletta caerulea Girard). — Световидов, Фауна СССР, Рыбы, 2, 1, 1952 : 175 (описание).
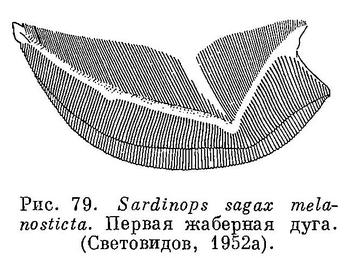
Близок к Sardinella. Жировые веки обычно одинаково хорошо развиты сзади и спереди глаз. На вертикальной части ключицы кожистого выроста нет. На крышечной кости есть радиальные бороздки. Брюшные килевые чешуи сравнительно хорошо развитые, образуют явственный киль, брюхо слегка закругленное. Рот конечный, нижняя челюсть не выдается вперед, сравнительно большой: сочленение нижней челюсти с черепом за вертикалью середины глаза, у крупных — под вертикалью заднего края его, задний конец верхнечелюстной кости достигает или почти достигает до вертикали середины глаза, у крупных простирается несколько далее. Сошник без зубов, челюстные и нёбные кости также без зубов, лишь в задней части по нижнему краю верхнечелюстной кости есть мелкие зубчики. Жаберные тычинки нижней части жаберной дуги укорачиваются к углу дуги (рис. 79), тычинки верхней части дуги сильно налегают снаружи на тычинки нижней части дуги, тычинки тонкие и длинные, до 100 на нижней части дуги. Брюшные плавники позади начала спинного. Основание спинного плавника в бороздке из чешуй, более крупных и удлиненных в задней части плавника и прикрывающих последние лучи его до вершины их. Чешуи на боках тела одинакового размера. Поперечные бороздки на чешуях с боков тела немногочисленные, неясно выраженные, прерванные посредине. На боках тела за жаберной крышкой сверху по темному пятну и по ряду пятен позади этого пятна, ниже которого у некоторых бывает еще 1—2 ряда из меньшего числа пятен. Икринки пелагические, с жировой каплей и с большим околожелтковым пространством. Морские рыбы средних размеров, до 300 мм, иногда немного больше. (Световидов, 1952а : 175).
1 вид, в Японском море подвид (Sardinops sagax melanosticta (Schlegel, 1846)).
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
6. Род SARDINOPS HUBBS
Sardinops Hubbs, Proc. Calif. Acad. Sci., XVIII, 11, 1929 : 264 (тип: Meletta caerulea Girard = Sardinops sagax caerulea).
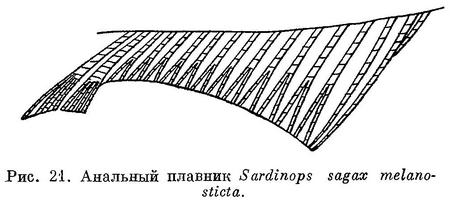
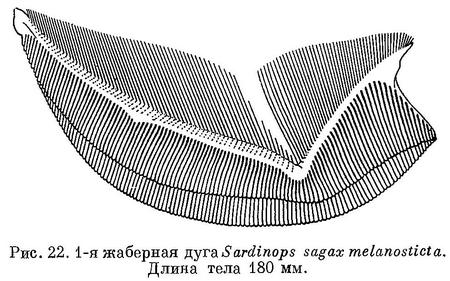
По отсутствию заметной медиальной вырезки в верхней челюсти и по наличию удлиненных чешуй (ala) при основании хвостового плавника и удлиненных последних лучей анального плавника (рис. 21) близок, к Sardinella. Жировые веки обычно одинаково хорошо развиты сзади, и спереди глаз. На вертикальной части ключицы кожистого выроста нет. На крышечной кости есть радиальные бороздки. Брюшные килевые чешуи сравнительно хорошо развитые, образуют явственный киль, брюхо слегка закругленное. Рот конечный, нижняя челюсть не выдается вперед, сравнительно большой: сочленение нижней челюсти с черепом за вертикалью середины глаза, у крупных — под вертикалью заднего края его, задний конец верхнечелюстной кости достигает или почти достигает до вертикали середины глаза, у крупных простирается несколько далее. Сошник без зубов, челюстные и нёбные кости также без зубов, лишь в задней части по нижнему краю верхнечелюстной кости есть мелкие зубчики. Жаберные тычинки нижней части жаберной дуги укорачиваются к углу дуги (рис. 22), тычинки верхней части дуги сильно налегают снаружи на тычинки нижней части дуги, тычинки тонкие и длинные, до 100 на нижней части дуги. Отростки плавательного пузыря помещаются в prooticum и pteroticum. Брюшные плавники позади начала спинного. Основание спинного плавника в бороздке из чешуй, более крупных и удлиненных в задней части плавника и прикрывающих последние лучи его до вершины их. Чешуи на боках тела одинакового размера. Поперечные бороздки на чешуях с боков тела немногочисленные, неясно выраженные, прерванные посредине. На боках тела за жаберной крышкой сверху по темному пятну и по ряду пятен позади него, ниже которого у некоторых бывает еще 1—2 ряда из меньшего числа пятен. Икринки пелагические, с жировой каплей и с большим околожелтковым пространством. Рыбы средних размеров, до 300 мм, иногда немного более.
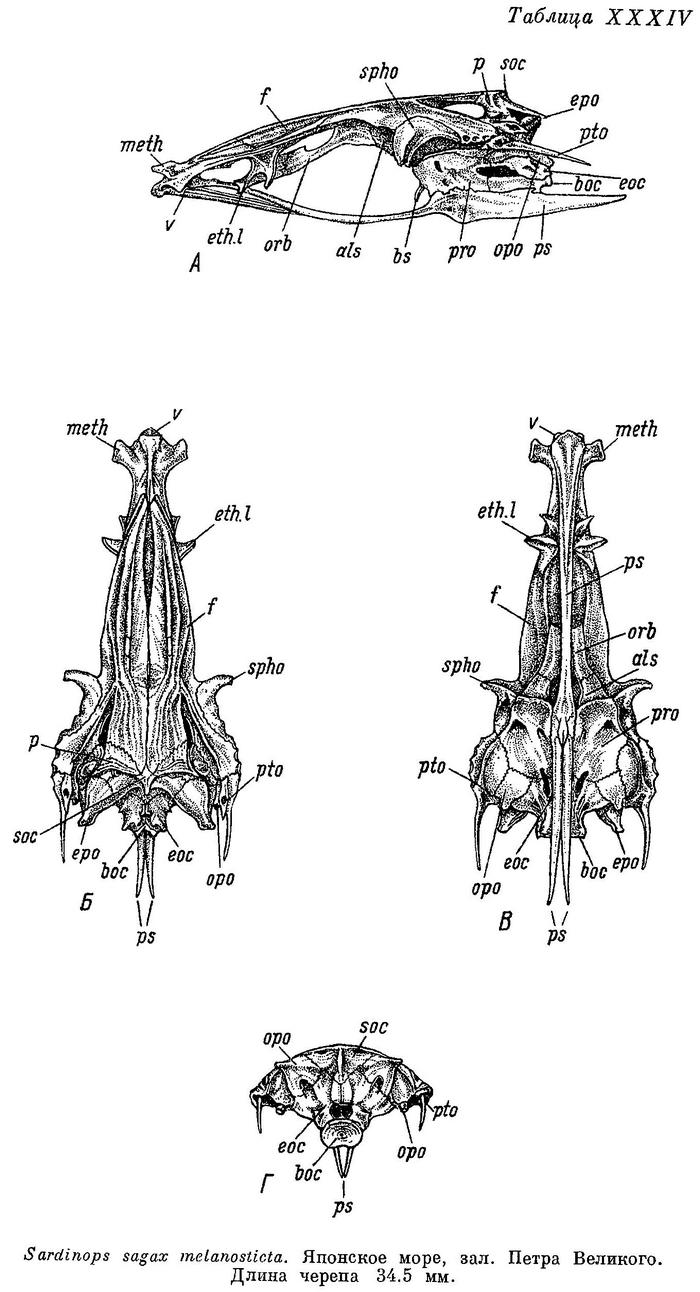
Скелет. (Описание и рисунки скелета головы см. у Филипса (Phillips, 1942 : 463—500, f. 1—23).) Bulla prootica меньше, чем bulla pterotica. Bulla prootica снаружи на черепе сбоку (табл. XXXIV, А) образует слабую, едва выдающуюся над поверхностью выпуклость; bulla pterotica имеет заметное вздутие лишь сверху под передней половиной fossa praeepiotica, сбоку на этой кости вздутие едва заметно, иногда отсутствует. Оба отростка плавательного пузыря с каждой стороны расположены в продольном углублении вдоль переднего края occipitale laterale и отчасти прикрыты сверху гребнем, отделяющим это углубление от отверстия n. vagus; выше каждый отросток помещается в костном канале, отверстие в который лежит впереди отверстия n. vagus значительно ниже opisthoticum. Fenestra auditiva значительных размеров, узкая и длинная, спереди близко подходит к bulla prootica, сзади к каналу, в котором расположен отросток плавательного пузыря. Fossa praeepiotica небольшая, низкая. Foramen temporale большое, более высокое и длинное, чем fossa praeepiotica, наиболее высокое сзади. Поверхность для причленения hyomandibulare сплошная, не разделена на две части на sphenoticum и pteroticum. Нижний край сошника и передней части парасфеноида почти прямые, слегка изогнутый лишь под ethmoidalia lateralia, передняя часть парасфеноида сверху закругленная, крылья парасфеноида выдаются вниз под миодомом, прямые снизу, суженные сзади черепа и заостренные на конце. Нисходящий отросток базисфеноида тонкий и длинный, снизу почти достигает парасфеноида. Задний край supraoccipitale и каждого epioticum на черепе сверху (Б) образует тупой угол, близкий к прямому. От заднего конца pteroticum отходит длинный и тонкий, слегка изогнутый шип, длина которого значительно варьирует; сверху под ним второй шип, более короткий, с отверстием у основания сверху. Передний конец каждого sphenoticum в виде отростка, почти равного по ширине на всем протяжении и тупого на конце, направленного в стороны и немного вперед. Передние и задние крылья ethmoidalia lateralia слегка выдаются в стороны из-под переднего конца лобных костей, задние крылья суживаются к концу, направлены в стороны, передние крылья обращены концами в стороны. Поперечные отростки мезэтмоида направлены немного вперед, не заостренные на концах. Череп довольно широкий, наибольшая ширина его немного более двух раз, ширина лобных костей впереди sphenoticum около 3.5—4 раз в его длине. Сошник и передняя часть парасфеноида на черепе снизу (В) плоские, сошник с коротким и небольшим гребнем на головке, закругленным сверху, парасфеноид сзади от сошника суживается, крылья парасфеноида разделяются под передней третью prooticum, тесно сближены впереди. Задний конец opisthoticum слегка удлинен в виде короткого, тупого на конце отростка. С каждой стороны на черепе сзади (Г) в нижней части epioticum и верхней части occipitale laterale располагаются fossae temporales posteriores, между основанием отростка каждого opisthoticum и шипом каждого pteroticum располагаются fossae subtemporales. Сочленовная головка на hyomandibulare сверху одна, не обособлена на две части, расширенная спереди. Quadratum с небольшой выемкой в верхней части, передний угол вытянут. Нижнечелюстная кость невысокая, сочленовная кость удлиненная и низкая, немного короче нижнечелюстной. Предкрышечная и крышечная кости широкие, последняя шире первой. Описание сделано по нескольким скелетам Sardinops sagax melanosticta.
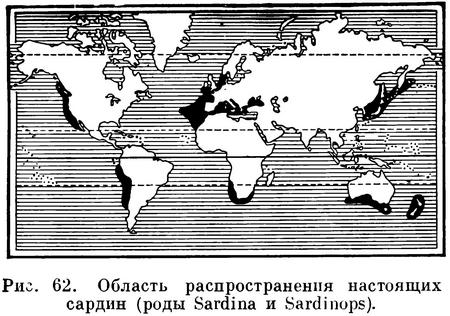
Морские рыбы. Один вид в субтропических частях Тихого, Индийского и отчасти Атлантического океанов у берегов Сев. и Южн. Америки, восточной Азии, Новой Зеландии, южных берегов Австралии и южной оконечности Африки (рис. 23). Sardinops dakini Whitley (Mem. Queensl. Mus., XI, 1937 : 114; известно 3 экз. от берегов северной части Квинслэнда, Австралия), выделяемая позднее в подрод (Fusciclupea Whitley, Austr. Zool., v. 9, 4, 1940 : 401, f. 7), судя по всему, относится к роду Sardinella, так как жаберные тычинки нижней части дуги у этого вида не налегают на тычинки нижней части дуги, радиальные бороздки на крышечной кости отсутствуют (они, как полагает автор описания, атрофированы или редуцированы до одной).
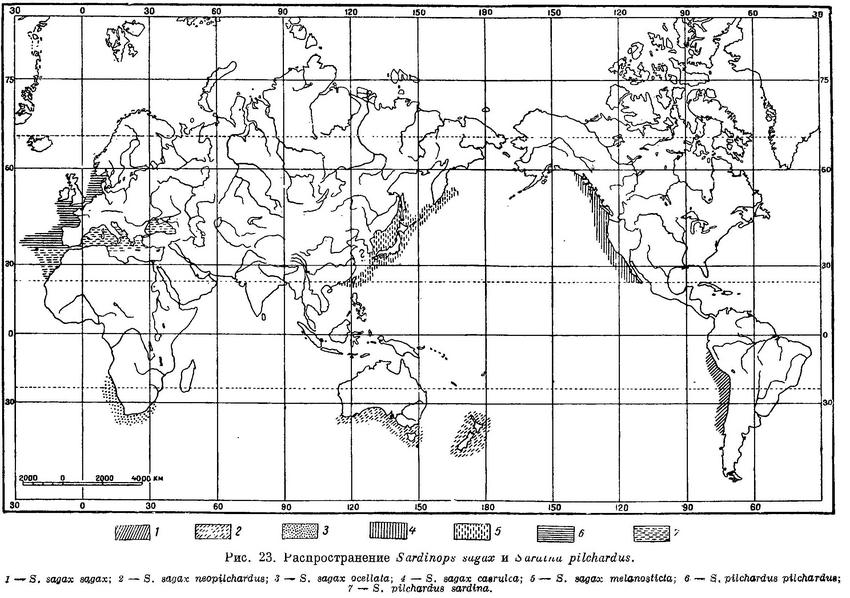
Субтропический представитель сельдевых северной части Тихого океана (Sardinops sagax), судя по всему, проник сюда из южного полушария. Близкие между собой субтропические роды Sardinops и Sardina наиболее близки в свою очередь к тропическому роду Sardinella и, повидимому, имеют общее происхождение. Род Sardina обособился в субтропических морях северного полушария, Sardinops, судя по его распространению (Sardinops sagax sagax — тихоокеанские берега Южн. Америки, Sardinops sagax neopilchardus — у южных берегов Австралии и вокруг Новой Зеландии, Sardinops sagax ocellata — берега южной Африки, Sardinops sagax melanosticta — берега юго-восточной Азии, Sardinops sagax caerulea — тихоокеанские берега Сев. Америки) — в субтропических морях южного полушария. Современное биполярное распространение Sardinops sagax сложилось в ледниковое время, когда вследствие похолодания климата имел место обмен субтропических элементов фауны обоих полушарий (Берг, 1920 : 273; 1947 : 128; Berg, 1933 : 449). Род Sardina в южное полушарие распространиться не мог, так как наиболее близкое место, куда бы он мог в первую очередь попасть (берега южной Африки), оказалось занятым совершенно сходным биологически родом Sardinops (Sardinops sagax ocellata). В свою очередь те же причины препятствовали распространению и Sardinops sagax ocellata в северную часть Атлантического океана к берегам Европы. Род Sardinops имел возможность беспрепятственно расселиться лишь в северную часть Тихого океана, куда он и проник как вдоль американских (Sardinops sagax caerulea), так и азиатских берегов (Sardinops sagax melanosticta), дав таким образом биполярное и амфипацифическое распространение (Андрияшев, 1939). Южное полушарие было, повидимому, центром происхождения не только рода Sardinops, но и рода Sprattus, большинство видов которого распространено также здесь.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Sardinops Hubbs 1929, тип: Meletta coerulea Girard 1854, Калифорния. Несколько видов — на Дальнем Востоке, в Японии, у берегов сев.-зап. Америки, Чили. Нов. Зеландии, Австралии, южн. Африки. В Японии и в Приморье на север до Командорских островов Sardinops sagax coerulea n. melanosticta (Temminck et Schlegel) 1846, «иваси», дальневосточная сардина. Описание см.: С. М. Кагановская, Рыбн. хоз. Дальн. Востока, XIII, № 1, 1935, стр. 38—40. — А. Г. Кагановский (устное сообщение) наблюдал дальневосточную сардину в совершенно пресной воде — в одном ключе, впадающем в бухту Джигит (бухта Рында) в сев. Приморье. Этот вид иногда заходит в опресненную часть Амурского Лимана.