Engraulis encrasicolus (Linnaeus, 1758)
Европейский анчоус, атлантическая анчоа, черноморский анчоус, азовский анчоус, черноморская хамса, азовская хамса, хамса (Rus),
European anchovy
(Eng)
Синонимы:
Anchoa argyrophanus Valenciennes, 1848 Атлантическая анчоа (Rus),
Anchovia argyrophana Valenciennes, 1848
Anchoviella guineensis Rossignol & Blache, 1961
Clupea encrasicolus Linnaeus, 1758
Engraulis amara Risso, 1827
Engraulis argyrophanus Valenciennes, 1848
Engraulis capensis Gilchrist, 1913
Engraulis encrasicholus maeoticus Pusanov, 1926 Азовская хамса (Rus),
Engraulis encrasicholus ponticus Alesandrov, 1927 Черноморский анчоус, черноморская хамса (Rus),
Engraulis encrasicolus russoi Dulzetto, 1947
Engraulis meletta Cuvier, 1829
Engraulis russoi Dulzetto, 1947
Engraulis vulgaris Nilsson, 1832
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Анчоус черноморский, или хамса — Engraulis encrasicholus ponticus Aleksandrov, 1927
У всех представителей рода тело вальковатое, округлое в поперечном сечении. Рот очень большой, нижний. Бока тела серебристые, спина серовато-коричневая. Длина тела примерно 12 см. Обитает в открытых частях Черного моря, зимой опускаясь на большие глубины. Играет огромную роль в промысле и в экосистеме моря, так как служит основной пищей крупных рыб и дельфинов.
Анчоус (хамса) азовская — Engraulis encrasicholus maeoticus Pusanov, 1926
Очень похож на предыдущий подвид, но спина более светлая, желтоватая. Длина тела, как правило, не более 8 см. Морской вид, зимует в Черном море, нерестится и нагуливается в Азовском море. Объект промысла.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
ЧЕРНОМОРСКАЯ ХАМСА — ENGRAULIS ENCRASICHOLUS PONTICUS ALEKSANDROV
Условия размножения. В пределах своего ареала анчоус Engraulis encrasicholus L. обитает при значительных колебаниях температуры и солености. По А. И. Александрову (1927), ареал анчоуса совпадает с распределением годовых изотерм от 8 до 22°, а в районах массового нахождения — от 10 до 20°.
Колебания солености в различных областях распространения этого вида также очень большие. В Атлантическом океане, где встречается указанный вид, соленость составляет 35—36‰; в Средиземном море он обитает при солености 38—39‰; в Черном море встречается повсеместно при солености 17—18‰; в северо-западной части Черного моря и в Азовском море обитает при солености 7—13‰. Наряду с большим разнообразием условий обитания прослеживаются общие для вида черты биологии размножения, которые являются, по-видимому, основным критерием пространственного распределения и численности стад в различных областях ареала.
Нерест анчоуса в Северном море происходит в мае — июле (Ehrenbaum, 1905—1909). Фаж (Fage, 1920), сопоставляя данные о размножении этого вида в северных и южных областях его распространения, указывает, что в Северном море и в Зюдерзее нерест начинается в последних числах мая и заканчивается в августе, с максимумом в июне и июле. В указанных районах нерест происходит при температуре воды 13,7—22° и солености 7—28‰. По данным Редеке (Redeke, 1916), анчоус нерестился на всем пространстве Зюдерзее (до его перекрытия), где соленость колебалась от 7 до 20‰. Наибольшее количество икринок и личинок встречалось в участках с соленостью 9—15‰.
После перекрытия Зюдерзее этот вид проник в Балтийское море, соленость поверхностных слоев которого не превышает 8‰.
У атлантических берегов Марокко анчоус также размножается в основном летом, иногда весной. В период нереста он является постоянным обитателем литоральной зоны (Furnestin J. et Furnestin M., 1959). По данным И. И. Казановой (1961), нерест Engraulis encrasicholus у западного побережья Африки происходит в течение почти всего года. В районах Дакара, Такоради и в Гвинейском заливе личинки этого вида встречались в планктоне в разные месяцы года.
Из наблюдений многих исследователей (Raffaele, 1888; Holt, 1899; Lo Bianco, 1908—1909; Fage, 1911, 1920) известно, что в Средиземном море нерест анчоуса продолжается от апреля до сентября с хорошо выраженным максимумом в июне и июле. Фаж (1920) указывает на нахождение очень мелких личинок в феврале и марте в районе Мессины и в ноябре у побережья Туниса, в Суэцком канале и прилежащих водах. Jlo Джудиче (Lo Giudice, 1922) отмечает, что в феврале и марте яйца и личинки анчоуса ловились в Генуэзском заливе.
Таким образом, при известных условиях нерест анчоуса может быть как очень ранним, так и очень поздним. В основном же, по заключению Фажа, это вид летнего размножения с четко выраженным тяготением в период нереста к повышенной температуре воды.
Растянутый период размножения Engraulis encrasicholus в Средиземном море Фаж (1937) объясняет неодновременным созреванием двух поколений анчоуса (годовиков и двухлеток), принимающих участие в нересте. В то же время Ло Джудиче (1922) связывает этот вопрос с существованием двух групп анчоуса — группы пелагической (открытого моря) и группы прибрежной. Первая группа размножается с марта по май в открытом море, вторая — с июня до сентября в прибрежных районах.
Из работ Фажа (1911, 1920) следует, что нерест анчоуса происходит только в прибрежных районах, преимущественно в районах промысла, который основан на концентрациях производителей в период размножения.
В Адриатическом море период размножения Engraulis encrasicholus также значительно растянут. В северных районах Вучетич (Vucetic, 1957) наблюдала нерест с апреля по октябрь. В средней Адриатике, по данным Гамулина (Gamulin, 1940), анчоус размножается с апреля по август с максимумом в мае. Как и в Средиземном море, в Адриатике он размножается в неглубоких местах вблизи берегов (Vucetic, 1957). Наши наблюдения (Дехник, 19636; Дехник, Синюкова, 1964) показали, что у албанского побережья икринки Engraulis encrasicholus встречались в заметном количестве в конце апреля и мае при температуре воды 15,0—19,5° и при солености 37,0 — 37,5‰. Сравнительно большая концентрация икринок анчоуса (свыше 1500 на десятиминутный лов ихтиопланктонной сети) наблюдалась в июле 1960 г. в северных районах Адриатического моря, особенно в Венецианском заливе над глубиной 30 м.
Температура воды в местах концентрации икринок достигала 23—25°, т. е. почти, максимальных значений для этих районов. Наоборот, соленость была минимальной (34‰) из известных значений для поверхностных вод Средиземного моря.
Период нереста Engraulis encrasicholus в Черном море совпадает с указанным периодом для Адриатического и в целом для Средиземного морей. Он начинается в последних числах апреля — начале мая и заканчивается в сентябре — первых числах октября. Черноморская хамса размножается при температуре воды 13—26° и солености 9,7—19,3‰ (Дехник и Павловская, 1950; Дехник, 1954; Зайцев, 1959а; Einarsson a. Gürtürk, 1960; Георгиев и др., 1961, 1962).
Вначале при относительно низкой температуре воды нерестится небольшое количество производителей. Массовое икрометание начинается при температуре воды 17—18°. Многими исследователями установлено, что нерест Engraulis encrasicholus ponticus в Черном море происходит на всей его площади. Как в прибрежных, так и в открытых районах встречаются значительные концентрации икринок и личинок (Книпович, 1926; Никитин, 1929; Водяницкий, 1930а; Пузанов, 1936; Малятский, 1940а; Майорова, 1941а; Казанова, 1947; Дехник и Павловская, 1950; Дехник, 1954; Einarsson a. Gürtürk, 1960; Дехник и др., 1970).
В Азовском море период нереста менее продолжителен. Икрометание начинается в мае или июне (в зависимости от весеннего прогрева воды) и заканчивается к августу или сентябрю (Дементьева, 1958). Развитие икринок и личинок Engraulis encrasicholus maeoticus происходит при температуре 18— 26° (Дементьева, 1958) и солености 10—17‰ (Морозова и Каракаш, 1939). Оптимальными условиями развития Т. Ф. Дементьева считает температуру воды 22,5—24° и соленость — 11,7—11,9‰. Нерест хамсы в Азовском море происходит на всем пространстве, за исключением Таганрогского залива (Книпович, 1926; Пузанов, 1936). В годы с небольшим паводком Дона, когда соленость Таганрогского залива повышается, нерест отмечается также в западной части залива (Корнилова, 1960). Наиболее интенсивное икрометание, по наблюдениям В. П. Корниловой (1960), происходит в прибрежных районах моря. По нашим наблюдениям, проведенным в течение двух нерестовых сезонов (1962—1963), большая концентрация икринок отмечена на всем пространстве Азовского моря как в прибрежных, так и в открытых районах (Дехник, 1967).
Интенсивность размножения рассматриваемого вида в значительной степени определяется соленостью воды. Из данных, приведенных Фажем (1920), следует, что распределение икринок и личинок Engraulis encrasicholus в северных областях распространения в основном ограничено изогалиной в 30‰, а максимальные концентрации приурочены к районам с соленостью не более 15‰. В этой же работе Фаж подчеркивает хорошо выраженное тяготение анчоуса в период размножения к воде пониженной солености в пределах распределения солености в Средиземном море. Ж. Фюрнестен и М. Фюрнестен (1959) также отмечают стремление анчоуса в период размножения к солоноватым водам. Данные многих упомянутых выше исследователей свидетельствую о том, что только в районах с низкой соленостью (в пределах ареала) наблюдается значительная концентрация икринок и личинок этого вида в планктоне.
Решающее влияние на размножение анчоуса оказывает температура. Нерест Engraulis encrasicholus в северных широтах повсеместно происходит в наиболее теплое время года. Нормальное развитие икринок и личинок ограничено диапазоном температуры воды 13—26° (с незначительными возможными отклонениями в ту и другую сторону). Оптимальные условия для развития создаются при прогреве воды от 18—20 до 25—26°. Ж. Фюрнестен и М. Фюрнестен (1959) приходят к заключению, что анчоус — эвритермная и эвригалинная рыба в период нереста становится стенотермной.
Суточный ритм размножения. В размножении хамсы и развитии выметанных икринок четко выражен суточный ритм (Дехник, 1959). Вымет и оплодотворение икры происходят закономерно в ночные часы. Только что выметанные икринки с обозначающимся протоплазматическим куполом на анимальном полюсе встречаются в планктоне от 20 до 24 ч. В каждый период времени в планктоне встречаются развивающиеся икринки хамсы двух этапов, один из которых относится к вымету в ближайшие ночные часы, другой — к вымету в предшествующую ночь. В отдельных случаях вылавливаются икринки трех этапов, два из которых смежные. Это определяется неодновременным выметом икринок в пределах каждого ночного отрезка времени, в течение которого происходит размножение хамсы. В определенные часы в планктоне встречаются икринки только одного этапа развития — обычно IV или V. Этот период времени находится в соответствии с длительностью периода развития каждого суточного вымета и зависит от температуры воды. Так, при температуре 20—24° выклев личинок происходит обычно в промежуток времени между 14 и 18 ч. До появления нового вымета икры (22—24 ч) в планктоне находятся икринки только одного суточного вымета ближайших ночных часов. Наличие в планктоне в каждый отрезок времени икринок хамсы только двух (или одного) этапов развития было отмечено Е. Г. Косякиной (1938).
Суточное распределение икринок по этапам развития может быть прослежено по их суммарному количеству из горизонтальных уловов ихтиопланктонной сетью в разгар нереста (июль — август 1961, 1962 гг.). Из приведенных данных (табл. 4) следует, что икринки хамсы на I этапе развития встречаются в планктоне между 20 и 08 ч следующего дня. В остальные часы суток икра хамсы на I этапе развития в планктоне не встречается. Нахождение икринок на последующих этапах развития соответственно сдвигается во времени (табл. 4).
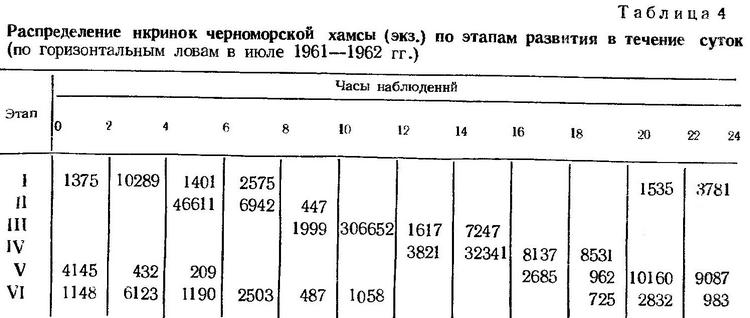
Чередование этапов эмбрионального периода развития черноморской хамсы (табл. 4.) находится в соответствии с четко выраженным ритмом ее размножения. Закономерность ежесуточного поступления икринок хамсы в планктон определяется неодновременным созреванием разных возрастных групп нерестовой популяции и растянутым многопорционным нерестом отдельных особей. Это важное приспособление направлено к сохранению численности стада. Спорадически проявляющиеся неблагоприятные для развития гидрологические условия (резкие колебания температуры воды, шторма) приводят к элиминации отдельных суточных выметов и при длительном периоде размножения не могут отразиться на общей эффективности нереста.
Строение икринок. Икринки черноморской хамсы имею эллипсоидальную форму. Перивителлиновое пространство очень узкое, желток почти вплотную примыкает к наружной оболочке икринки. По мере развития эмбриона перивителлиновое пространство увеличивается главным образом у анимального и вегетативного полюсов. Оболочка икринок тонкая, прозрачная. Желток, как у всех сельдевых, разделен на крупные дольки. Жировой капли нет (рис. 4) В желтке содержится много воды (больше 90%), этим в основном определяется хорошая плавучесть икринок хамсы (Крыжановский. 1956).
Данные о величине икринок черноморской хамсы содержатся в работах многих исследователей. В. А. Водяницкий (1930а, 1936), С. М. Малятский (1940а), В. А. Водяницкий и И. И. Казанова (1954) указывают, что продольный диаметр икринок хамсы колеблется от 1,5 до 1,9 мм, поперечный — от 0,8 до 1,2 мм. По наблюдениям Ю. П. Зайцева (1958а, 1959а), в северо-западной части Черного моря икринки хамсы указанного размера не встречаются. На основании большого количества измерений живых икринок, выловленных в приднепровском районе в течение всего периода размножения, автор отмечает, что длина продольного диаметра изменяется от 1,01 до 1,60 мм и поперечного — от 0,72 до 1,03 мм. Соответствующие размеры икринок черноморской хамсы приводят Р. М. Павловская (1955) и Т. В. Луговая (19636) для районов Крыма и Кавказа. По нашим данным, продольный диаметр живых икринок изменяется от 1,00 до 1,90 мм, поперечный — от 0,66 до 1,20 мм.
Величина икринок черноморской хамсы закономерно уменьшается от начала к концу нерестового сезона (Зайцев, 1958а, 1959а; Луговая, 19636) (табл. 5).
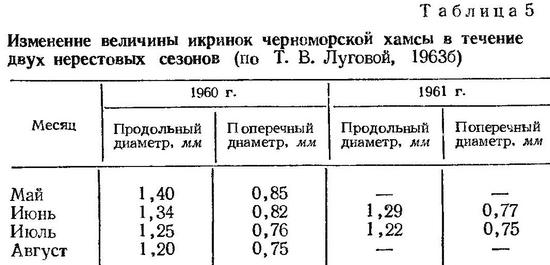
Икринки Engraulis encrasicholus из разных районов ареала существенно отличаются по размерам и форме. Демир (Demir, 1959) приводит данные о размерах и форме икринок анчоуса в Черном, Мраморном, Эгейском и Средиземном морях и приходит к выводу, что с повышением солености размер икринок уменьшается и они приобретают более вытянутую форму. Обстоя тельный анализ этих изменений дан в работе Т. В. Луговой (1963а). Сопостав ление размеров икринок из разных районов ареала, проведенное автором, свидетельствует об увеличении продольного и уменьшении поперечного диаметров икринок в направлении от районов с низкой соленостью к районам с высокой соленостью. От Зюдерзее с соленостью 10—15‰ к восточным районам Средиземного моря с соленостью 39—40‰ соотношение большого и малого диаметра икринок повышается от 1,29 до 2,55. В морях с пониженной соленостью воды (Азовское, Черное, Зюдерзее до перекрытия) форма и размеры икринок существенно изменяются даже при небольшом повышении солености. Так, от Азовского к Черному морю при повышении солености от 12 до 18‰ соотношение диаметров икринок изменяется от 1,29 до 1,66 (Луговая, 1963а). Соответственно четко прослеживается уменьшение объема икринок анчоуса при повышении солености. В морях с океанической соленостью объем икринок в 3,5—4 раза меньше, чем в морях с пониженной соленостью (Луговая, 1963а). Уменьшение размеров пелагических икринок с повышением солености воды четко показано в работах Ю. П. Зайцева (1955а, 1958а).
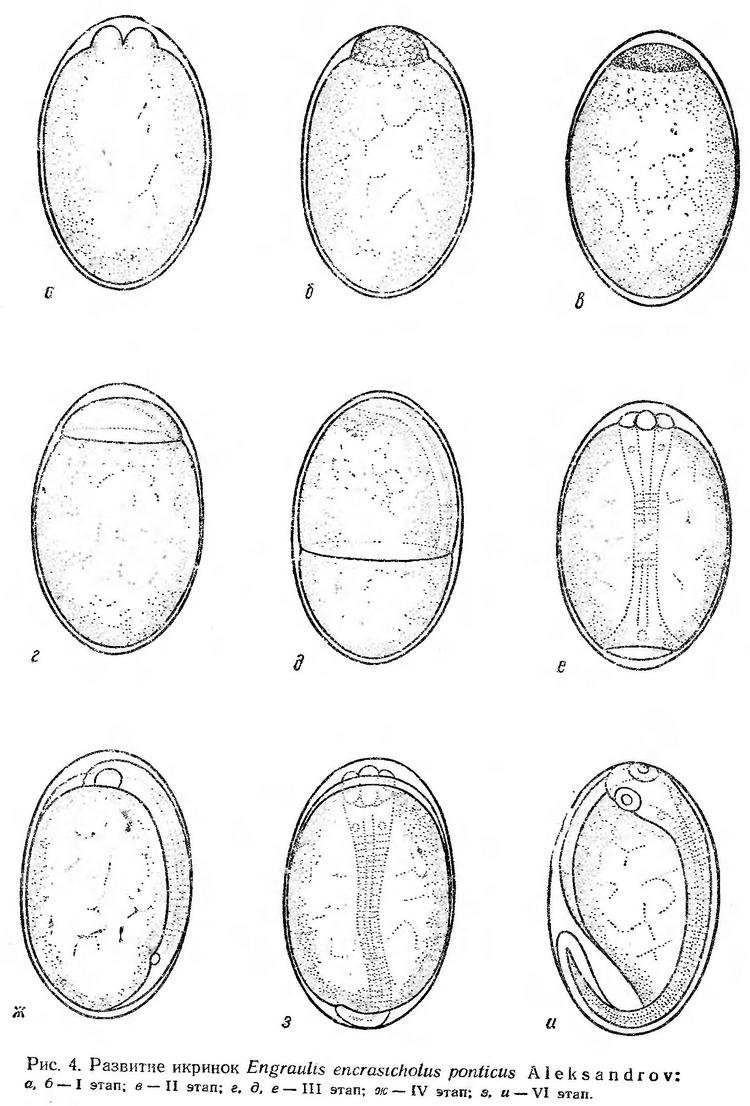
Эмбриональное развитие черноморской хамсы прослежено на икринках, собранных в море между 22 и 24 ч. Температура воды в опытах в период эмбрионального и постэмбрионального развития изменялась от 22,0 до 23,4°. Наблюдения за развитием икринок хамсы были начаты в 23 ч. В протоплазматическом бугорке, занимающем очень небольшую часть поверхности желтка, наметилась первая борозда дробления (рис. 4, а). В 2 ч 50 мин следующих суток икринки были на стадии мелкоклеточной морулы (рис. 4, б). Таким образом, процесс дробления продолжался около 4 ч (табл. 6).
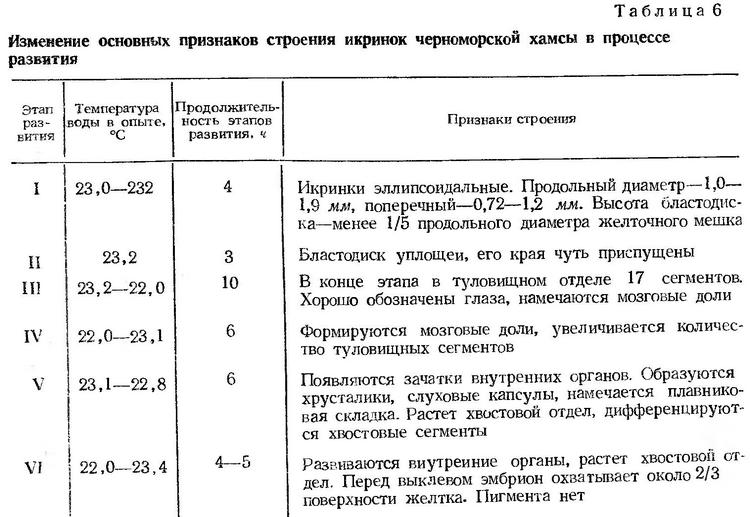
Начало смещения бластомеров к поверхностному слою (образование эпителиальной бластулы) сопровождается появлением в перибласте многочисленных пузыревидных образований, которые по мере развития перемещаются по всей поверхности желтка к вегетативному полюсу. Значение этих образований не вполне ясно и возникают они, как будет показано в последующих описаниях эмбрионального развития черноморских рыб, не у всех видов.Образование эпителиальной бластулы (II этап) при температуре воды 23,2° происходило в течение 3 ч. Бластодиск уплощен, края его чуть приспущены. При определенных поворотах икринки под многослойной пластинкой просматривается бластоцель (рис. 4, в). Через 7 ч после появления первой борозды дробления начинается процесс обрастания желтка бластодиском. Почти одновременно происходит подворачивание краев бластодиска и образование краевого (зародышевого) утолщения (рис. 4, г). По мере наползания бластодиска на желток зачаток зародыша удлиняется и утолщается. Когда края бластодиска заходят за экватор, появляются первые туловищные сегменты, намечаются зачатки глаз (рис. 4, д). Через 14 ч после начала дробления бластодерма охватывает около 2/3 поверхности желтка. В туловищном отделе насчитывается 9 сегментов, появляется купферов пузырек. Головной конец значительно расширен. Перед закрытием бластопора в туловищном отделе насчитывается 12—15 сегментов, четко обозначены глаза, намечаются мозговые доли. Купферов пузырек перемещается к заднему концу тела (рис. 4, е). Процесс гаструляции и образования зародышевой полоски (III этап) при температуре воды 23,2—22,0° продолжается около 10 ч.
Ко времени замыкания бластопора (рис. 4, ж) количество туловищных сегментов увеличивается до 20. Сохраняется купферов пузырек, обозначаются мозговые доли. IV этап, заканчивающийся образованием хвостовой почки при температуре воды 22,0—23,1°, продолжался около 6 ч.
На V этапе развития наряду с ростом хвостового отдела дифференцируются хвостовые миотомы, четко выявляются мозговые доли, большие с хрусталиками глаза. Формируются слуховые капсулы, сердце, печень, кишечник, появляется плавниковая складка. V этап развития продолжался в опыте при температуре воды 23,1 — 22,8° около 6 ч.
К началу пульсации сердца (VI этап) эмбрион хамсы охватывает чуть больше половины поверхности желтка (рис. 4, з). Сердце пульсирует 91—93 раза в минуту. Одновременно с пульсацией сердца начинаются слабые подергивания эмбриона. В течение VI этапа происходит формирование заложенных зачатков личиночных органов. Тело эмбриона совершенно лишено пигмента. По мере развития движения эмбриона становятся более энергичными. Перед выклевом голова эмбриона прижата к оболочке, которая постепенно утончается под воздействием ферментов вылупления и разрывается. Ко времени вылупления эмбрион охватывает около 2/3 поверхности желтка (рис. 4, и). Выклев происходит с головного конца и начинается через 4—5 ч после появления пульсации сердца. Перед выклевом сердце пульсирует 110—115 раз в минуту.
Весь процесс эмбрионального развития хамсы при температуре воды 22,0—23,4° длился около 1,5 суток (табл. 6).
Постэмбриональное развитие. Только что выклюнувшиеся личинки хамсы имеют длину, по нашим данным, 2,28—2,52 мм. Меньшие размеры выклюнувшихся личинок (1,58—1,60 мм) указывают В. А. Водяницкий и И. И. Казанова (1954). По данным Р. М. Павловской (1955), выклюнувшиеся личинки имеют длину 1,75—1,80 мм. Приведенные разными авторами различные размеры личинок хамсы объясняются значительными колебаниями размеров икринок как в течение нерестового сезона, так и по районам Черного моря. Указанные различия также определяются температурой воды, при которой происходит развитие икринок. При более низкой температуре эмбрионы выклевываются более развитыми и больших размеров. В. А. Водяницкий и И. И. Казанова приводят размеры личинок, по-видимому, по фиксированным материалам.
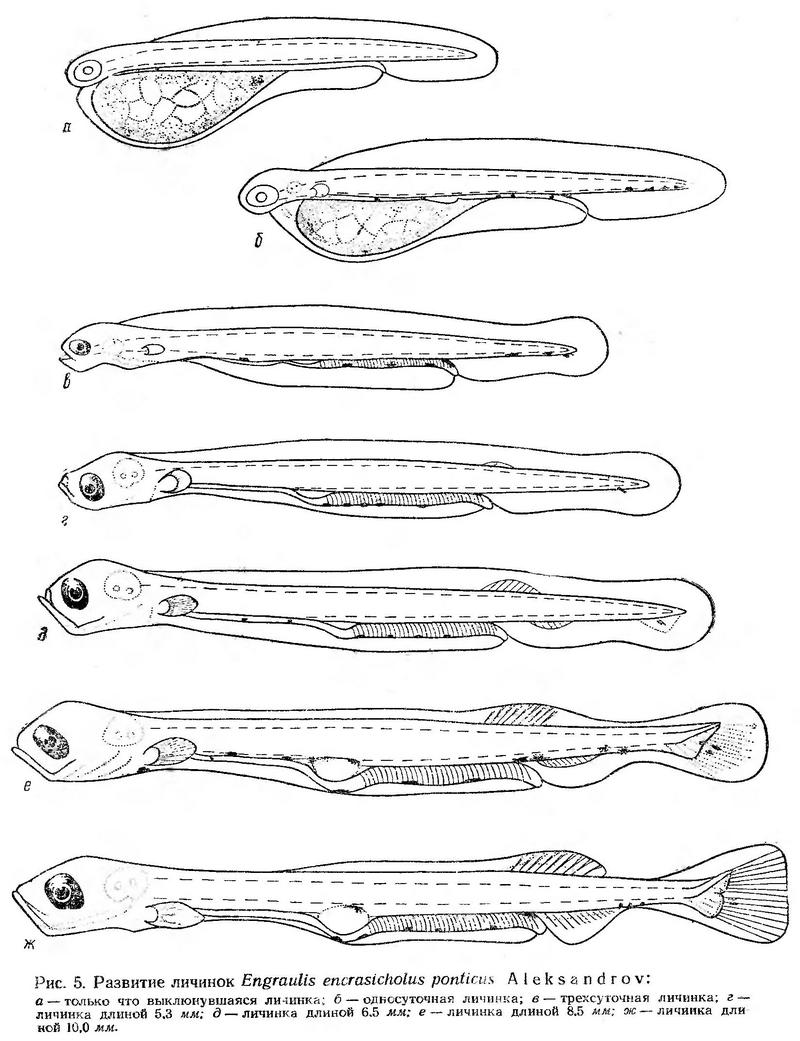
Тело только что выклюнувшихся личинок хамсы тонкое, длинное, сплющенное с боков. Голова плотно прижата к желтку и слегка загнута вниз. Анус отодвинут далеко назад (рис. 5, а); антеанальное расстояние составляет около 70% длины тела. Кишечник длинный и прямой, без петель. Личинки лишены пигмента, тело их прозрачное. В круглых аквариумах объемом 15 л они распределяются по всей толще воды. В покое держатся почти в строго вертикальном положении головой вниз. Затем быстро переворачиваются головой кверху и начинают подниматься по изогнутой линии при помощи волнообразных изгибов длинного тонкого тела. Периоды движения кратковременные, обычно не более 2—3 сек; периоды покоя длятся от 2—3 до 25—60 сек. Время от времени личинки делают стремительные броски (1,5—2 см/сек).
Односуточные личинки хамсы достигают в среднем 3,5 мм длины. Желточный мешок сокращается в размере (рис. 5, б). Формируются грудные плавники, намечается ротовая ямка. Вдоль вентральной стороны тела над кишечником появляются мелкие точечные меланофоры. Тело личинок совершенно прозрачное, и они с трудом просматриваются в падающем и отраженном свете. Периоды покоя, когда личинки медленно пассивно погружаются головой вниз, длятся от нескольких секунд до минуты. Периоды движения увеличиваются до 4—10 сек. Иногда личинки проплывают за это время расстояние в 15—17 см. Обычно разовые перемещения составляют не более 2—3 см. Изредка наблюдаются стремительные броски.
В течение вторых суток развития при температуре 21,5° почти полностью резорбируется желточный мешок, прорезается рот, формируются жаберные крышки, увеличиваются грудные плавники, пигментируются глаза. Скорость движения двухсуточных личинок хамсы составляет в среднем 6,9 см/мин (Синюкова, 1969).
Трехсуточные личинки достигают длины 3,88 мм. Рот большой, открытый, глаза интенсивно пигментированы (рис. 5, в). В состоянии покоя личинки занимают косое положение; головной конец по-прежнему обращен вниз. При движении они принимают горизонтальное положение. Периоды покоя сокращаются, обычно они не превышают 10 сек. Моменты движения также непродолжительные, но отдельные броски стремительные — за 1 сек личинки могут проплыть 2—3 см.
Плавая с открытым ртом, личинки наталкиваются на пищевые организмы и могут их схватывать. Поскольку в этом возрасте остаток желтка очень небольшой, а активность личинок значительно возрастает, расход энергии должен пополняться за счет внешней пищи. В это время личинки питаются, по-видимому, в значительной степени пассивно, захватывая вместе с водой при плавании фитопланктонные организмы и малоподвижные формы зоопланктона. По наблюдениям Л. А. Дука (1961), у таких личинок в кишечниках встречаются иногда яйца и мелкие науплиусы Copepoda.
Личинки в возрасте четырех суток плавают в нормальном горизонтальном положении и только в покое принимают вертикальное или косое положение, как бы подвешиваясь хвостом к поверхностной пленке. Грудные плавники становятся подвижными, в связи с чем личинки приобретают способность быстро поворачиваться и плавать в разных направлениях. Скорость их движения увеличивается до 16 см/мин (Синюкова, 1969).
Пяти- и шестисуточные личинки в условиях опыта находятся в угнетенном состоянии. Движения их становятся вялыми, периоды покоя увеличиваются. На шестые-седьмые сутки личинки погибают. Последующее описание морфологических признаков личинок дано по фиксированному материалу.
При длине личинок 5,3—5,5 мм закладываются лучи спинного плавника, намечаются лучи в хвостовом плавнике (рис. 5, г). Желточного мешка нет, питание внешнее. В пищевом комке личинок встречаются малоподвижные формы зоопланктона, в основном яйца, науплиусы и метанауплиусы Сореpoda, реже копеподитные и взрослые формы Oithona minuta (Павловская, 1955, 1958а, 19586; Дука, 1961, 1964а, 19646). Фитопланктонные организмы в пище личинок хамсы встречаются редко (Дука, 1961).
У личинок длиной 6,5—7,0 мм лучи в спинном плавнике достигают половины высоты плавниковой складки, формируются лучи в анальном плавнике. Сохраняются пигментные клетки вдоль нижнего края кишечника (рис. 5, д). Основную роль в питании этих личинок играют более подвижные формы зоопланктона, как Oithona minuta и копеподиты Acartia clausi (Дука, 1961, 1964а). Это свидетельствует об увеличении скорости движения личинок, о возросшей их активности.
У личинок длиной 8,5 мм продолжается дифференцировка непарных плавников. Лучи спинного плавника раздваиваются. По верхнему краю плавательного пузыря и на вентральной стороне хвостового отдела появляются меланофоры. Плавательный пузырь наполнен воздухом (рис. 5, е). При длине личинок 10,0—10,5 мм плавниковая кайма редуцируется, четко дифференцируются непарные плавники, заканчивается формирование лучей (рис. 5, ж). В пищевом комке личинок наряду с излюбленными формами (науплиусы и метанауплиусы Copepoda, взрослые формы Oithona minuta) встречаются крупные подвижные организмы, такие как Acartia clausi (Дука, 1961, 1964а).
Распределение и численность икринок и личинок. Сопоставление видового состава икринок и личинок летнеразмножающихся рыб Черного моря (Дехник и др., 1970) свидетельствует о значительном преобладании икринок и личинок хамсы над другими видами. Как отмечалось, хамса интенсивно размножается на всем пространстве моря. Как в прибрежных, так и в открытых районах моря икринки и личинки этого вида встречаются в большом количестве (Никитин, 1929; Водяницкий, 1930а; Пузанов, 1936; Малятский, 1840а, 19406; Дехник и Павловская, 1950; Дехник 1954; Einarsson a. Gürtürk, 1960; Георгиев и др., 1961, 1962; Дехник и др., 1970).
Икринки хамсы обладают высокой плавучестью (Зайцев, 1953, 19556). По данным многих исследователей (Никитин, 1929; Водяницкий, 1930а; Елизарова, 1936; Косякина, 1938; Малятский, 1940а, 1940б), их распределение ограничено преимущественно поверхностным однометровым слоем воды. Наблюдениями Ю. П. Зайцева (1961а, 1964а) установлено, что икринки и личинки хамсы входят в состав гипонейстона. Наряду с этим дальнейшее изучение вертикального распределения ихтиопланктона методом серийных горизонтальных ловов (Дехник, 1954, Дехник и др., 1970) показало, что икринки хамсы, как и других теплолюбивых видов, распространены во всем верхнем теплом слое моря (до глубины 20—25 м) и образуют иногда более высокие концентрации в нижележащих слоях (табл. 7). Широкое вертикальное распределение пелагических икринок определяется в значительной степени перемешиванием водных слоев, а также колебаниями плотности морской воды, вызываемыми изменением температуры и солености. Быстрые вертикальные перемещения пелагических икринок можно хорошо наблюдать в высоких аквариумах, лишь слабое покачивание которых приводит к размещению икринок во всем слое воды от дна до поверхности.
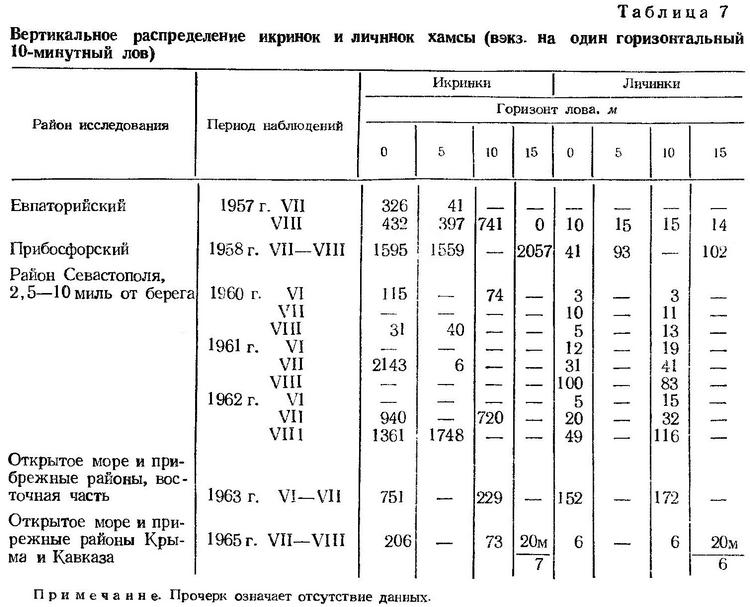
В вертикальном распределении личинок хамсы закономерно прослеживается увеличение их численности с глубиной (в слое 0—15 л). Приведенные в табл. 7 данные получены в основном по многосуточным наблюдениям с примерно одинаковым числом дневных и ночных ловов. Суточные миграции в пределах теплого слоя моря, которым ограничено распределение личинок хамсы, не выражены.
Проведенные многосуточные наблюдения на небольшом пространстве моря (район Севастополя) позволили выявить дисперсность (пятнистость) в распределении личинок хамсы (табл. 8). Численность личинок в очень короткие периоды времени на небольшой площади моря колеблется от единиц до нескольких десятков и даже сотен экземпляров.

Колебания численности в уловах нельзя отнести за счет времени суток, так как высокие или низкие уловы приходятся на разные часы наблюдений (табл. 8). Мы полагаем, что приведенные данные достаточно хорошо иллюстрируют пятнистость распределения личинок или, иными словами, их стайное поведение, что, как показано многими исследователями, имеет многообразное приспособительное значение (Мантейфель, 1955; Никольский, 1955; Keenleyside, 1955; Радаков, 1956, 1958; Мантейфель и Радаков, 1960; Орфеев, 1963; Радаков, 1965).
Численность икринок и личинок хамсы в планктоне, как и других видов рыб, значительно колеблется в течение нерестового сезона. Первые икринки хамсы появляются в планктоне в конце апреля — начале мая. В это время нерест хамсы, по-видимому, оказывается безрезультатным в отношении воспроизводства. Личинки ее начинают встречаться в уловах в июне, а у северного побережья Черного моря обычно во второй половине июня при темпетуре воды 17—18° (Малятский, 1940а, 1940б; Виноградов, 1948, 1949; Дехник, 1954). Насколько известно, ни в одной из работ по ихтиопланктону Черного моря нет указания на нахождение личинок хамсы в планктоне в апреле или мае.
Однако в июне эффективность нереста хамсы, по нашим данным, еще низкая. Так, в Евпаторийском районе с 3 по 6 июня 1958 г. при значительной численности икринок хамсы (до 250— 300 шт. в одном 10-минутном лове) личинок в планктоне не было. Температура воды у поверхности колебалась от 18,1 до 19,1°. С 1 по 19 июня 1959 г. проводились наблюдения в халистатической области западной половины Черного моря. За первые 10 суток численность икринок в планктоне составляла в среднем 60 экз/м2 в слое 25—0 м. За это же время было обнаружено только 3 предличиики хамсы в горизонтальных ловах. В последующие 9 суток численность икринок увеличилась до 84 экз/м2, а численность личинок составляла в среднем только 0,1 экз/м2.
В течение трехлетних наблюдений в районе Севастополя при среднем количестве икринок в июне, достигающем 70 экз/м2, численность личинок составляла в среднем лишь 2,2 экз/м2. Отношение численности личинок к количеству икринок в июне оставалось очень низким, в среднем 3,2%.
В более южных районах, в частности у побережья Болгарии, количество личинок хамсы в июне бывает значительным. К сожалению, в работе Ж. М. Георгиева, К. Александровой и Д. Хр. Николова (1961) данные о численности личинок рассчитывались по поверхностным горизонтальным ловам и выражались средней величиной на одну станцию. Эти результаты не могут быть сопоставлены с нашими. Однако более высокие концентрации личинок в июне (в отдельные годы) по сравнению с июлем, по данным указанных авторов, свидетельствуют о высокой эффективности нереста в данный период.
Максимальная концентрация икринок и, следовательно, наиболее интенсивный нерест хамсы происходит в июле и августе. Численность икринок хамсы в июле в разных районах Черного моря колеблется в среднем, по нашим данным, от 23 до 868 экз/м2 (табл. 9) с индивидуальными колебаниями от единиц до 5 тыс. экз/м2. Численность личинок в это время также значительно возрастает и соответственно этому повышается процентное соотношение личинок и икринок.
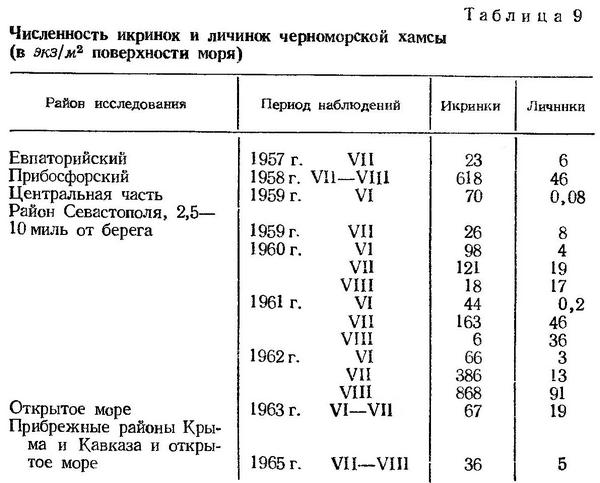
В августе интенсивность нереста хамсы обычно несколько снижается, а количество личинок в отдельных случаях превышает количество икринок за счет расширения размерного ряда и преобладания в планктоне крупных личинок (Дехник и др., 1970). В отдельные годы нерест хамсы в августе может быть очень интенсивным. Так, в 1962 г. в районе Севастополя численность икринок хамсы в планктоне была необычно высокой и соответственно наблюдалась большая концентрация личинок (табл. 9). В отдельных случаях численность личинок достигала 400—500 экз/м2. Соотношение между численностью личинок и икринок в августе обычно наибольшее.
В сентябре икринки хамсы немногочисленны в планктоне, количество личинок не превышает, по нашим данным, 20 экз/м2 и составляет в среднем в прибрежном районе 9 экз/м2. О низкой интенсивности нереста хамсы в сентябре свидетельствуют данные и других исследователей (Дехник и Павловская, 1950; Овен, 1959; Георгиев и др., 1961). В октябре личинки хамсы встречаются очень редко, нахождение икринок не отмечено (Дехник, 1954).
В Адриатическом море наибольшая численность икринок хамсы, по нашим данным, в разгар нереста (конец июня — июль) составляла 186 экз/м2, численность личинок колебалась от 2 до 44 экз/м2.
Фаж (1920) приводит данные о количественном распределении икринок анчоуса в Зюдерзее (до его перекрытия). По наблюдениям в течение ряда лет, в 10 см3 воды насчитывалось 204—3780 икринок. Эти данные свидетельствуют об очень высокой концентрации икринок анчоуса в планктоне Зюдерзее, значительно превышающей концентрацию их в Черном и Азовском морях, где хамса одна из самых многочисленных рыб.
В Азовском море в разгар нереста (вторая половина июня — начало июля) максимальная численность икринок хамсы, по нашим данным, в 1962 и 1963 гг. достигала 2170—2180 экз/м2, что в пересчете на объем составляет более 300 экз/м3. Численность личинок в эти же сезоны достигала 180 экз/м2 или 26 экз/м3 (Дехник, 1967).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Наиболее широкое распространение
имеет европейский анчоус, обитающий в
Атлантическом океане от Канарских
островов до Бискайского залива, во всех
районах Средиземного и Черного морей, а в
летнее время заходящий также в Северное
(до берегов Южной Норвегии),
Балтийское и Азовское моря. В пределах своего
ареала этот вид, переносящий большие
колебания солености и температуры,
образует несколько обособленных форм —
атлантическую, средиземноморскую,
черноморскую и азовскую.
Черноморский анчоус, или хамса (Е.
encrasicholus ponticus), постоянно живет
в Черном море, будучи вполне обычным
у всех берегов. Летом хамса широко
рассеивается на всем пространстве моря и
придерживается верхних слоев воды,
расположенных над слоем
температурного скачка. Особенно много этой рыбы
бывает летом в хорошо прогреваемой и
богатой кормовым планктоном северо-западной
части Черного моря. Зимой, когда
поверхностная вода сильно охлаждается,
а штормы достигают большой силы, хамса
концентрируется в ограниченных
прибрежных районах, ведет малоподвижный
образ жизни, слабо питаясь и опускаясь
на глубину 70—80 м. Здесь она держится
главным образом в придонных водах
и лишь в теплые, тихие дни поднимается
к поверхности. Впрочем, в мягкие зимы
хамса может и вообще не опускаться на
глубину. В наших водах основные районы
зимовки располагаются у южного
побережья Крыма и у берегов Грузии.
Весной, обычно в начале апреля, хамса
поднимается с глубин и начинает активно
питаться планктоном (в основном
мелкими ракообразными). Сначала она
появляется у берегов близ районов зимовки, но
вскоре уходит в море и быстро
рассредоточивается по его площади. Размножение
хамсы происходит в Черном море
повсеместно и продолжается в течение всего
теплого времени года — с мая по сентябрь,
причем, судя по нахождениям
икринок, наиболее интенсивный нерест идет
в местах массового развития планктона.
Плодовитость хамсы составляет около
20—25 тыс. икринок, выметываемых двумя-
тремя (иногда даже четырьмя) порциями.
Хамса имеет очень короткий
жизненный цикл — ее предельный возраст
составляет всего 3—4 года. В первые два года
жизни эта рыба довольно быстро растет,
достигая к концу этого срока 10—11 см
в длину, но в дальнейшем скорость ее
роста замедляется, и предельные размеры
не превышают 13, редко 15 см. В общем
составе нерестующей популяции хамсы
преобладают двух- и трехлетние рыбы, а на
четырехлеток приходится лишь около 1 %.
Черноморская хамса — самая
многочисленная рыба моря. Она служит
основной пищей большинства хищников этого
водоема — пеламиды, скумбрии, белуги
и других рыб, а также дельфинов, чаек
и буревестников.
Азовская хамса (Е. encrasicholus maeoticus) отличается от черноморской
светлой окраской и меньшими размерами —
ее обычная длина составляет 8—9 см,
а предельная — не более 10—11 см. Эта
рыба проводит в Азовском море только
лето. Там она активно питается, там же
происходит икрометание (в июне — июле)
и нагул мальков. Осенью азовская хамса
всех возрастов уходит через Керченский
пролив в Черном море и, продвигаясь
вдоль берегов Кавказа и Крыма, подобно
черноморской хамсе залегает в
зимовальные ямы. Зимовка азовской хамсы
в разные годы происходит в различных
районах, но большая часть ее
обыкновенно остается на зиму в районе
Новороссийска или несколько южнее.
Во время зимовальной миграции (как
и при обратном перемещении) хамса
движется огромными косяками,
сопровождаемыми массой кружащихся над ними
чаек и буревестников, а также
дельфинами. Иногда мигрирующая хамса в
большом количестве заходит в бухточки и
заливы; особенно много этой рыбы
наблюдалось в те времена, когда промысел ее
был еще слабо развит. Вот как описывает
подобный заход хамсы в Балаклавскую
бухту один из первых исследователей
черноморского рыболовства Н. Я.
Данилевский: «Самый замечательный пример этого
рода был в 1859 г., когда изобилие хамсы,
зашедшей в бухту, вместо благодеяния
природы обратилось в настоящее
народное бедствие. От натиска сзади зашедшая
в бухту хамса не могла возвратиться
назад. Здесь залив, имеющий более версты
в длину и около 100 сажен в ширину, до
того переполнился рыбой, что не было
видно воды. От такого скучения она вся
задохлась и задушила всю прочую рыбу,
которая выскакивала на берег. Морские
раки тоже выползли вон из воды. От
гниения этой рыбы распространилось такое
зловоние, что серебро в шкафах и
масляные картины с белилами совершенно
почернели. Из угла бухты, где она мелка,
заставляли выгребать хамсу и уносить
в мешках. Ее зарывали в землю,
некоторые удобряли поля, но все это не могло
уменьшить зловония... Невыносимый
запах продолжался с год в окрестностях
Балаклавы, и рыба совершенно исчезла из
бухты. На другой год можно было видеть
с набережной... в тихую погоду целые
кучи дохлой рыбы на дне, лежавшей
точно скирды. По словам жителей, тут
погибли миллионы пудов хамсы. В малых
размерах повторилось это в 1867 г.».