(Myctophidae Gill, 1893) Семейство Миктофиды, Миктофовые, Миктофидовые, Светящиеся анчоусы, Family Myctophidae Gill, 1893 (Lanternfishes) 33 рода 248 видов
Рыбы открытого океана. Н.В.Парин 1998 г.
Летучие рыбы питаются планктонными животными, обитающими в поверхностном слое, в основном мелкими ракообразными и крылоногими моллюсками, а также личинками рыб. Их основными пищевыми конкурентами в тропической зоне являются представители семейства светящихся анчоусов (Myctophidae) — группы, в целом характерной для глубоководной пелагиали, но включающей также виды, которые поднимаются в темное время суток к самой поверхности. Приповерхностные, или никтоэпипелагические (от греческого слова «никтиос» — ночь), миктофиды, принадлежащие к родам Myctophum, Symbolophorus и др., достигают высокой численности. Размах суточных вертикальных миграций этих небольших рыбок (обычная длина 6—10 см) достигает 500—900 м: за какой-нибудь час они преодолевают, таким образом, расстояния, в несколько тысяч раз превышающие длину их тела. Никтоэпипелагические светящиеся анчоусы достаточно многочисленны и в умеренных водах — там, где летучие рыбы, область распространения которых ограничена, грубо говоря, изотермой 20° на поверхности, не живут. В умеренно тепловодной зоне они уступают; роль массовых потребителей зоопланктона родственной группе — семейству макрелещуковых (Scomberesocidae).
Плодовитость эпипелагических рыб, как правило, очень велика. Только у миктофид (их длина, напомним, редко превышает 10 см), летучих рыб и макрелещуковых она исчисляется тысячами икринок, тогда как крупные рыбы поверхностного слоя продуцируют икру миллионами и десятками миллионов. Для преобладающего большинства эпипелагических рыб характерна порционность икрометания, приводящая к большой растянутости сроков нереста. В тропических водах в связи с относительным постоянством условий среды размножение может идти даже круглогодично (например, у летучих рыб и тунцов). Высокая плодовитость и ускоренное прохождение ранних стадий индивидуального развития направлены у костистых рыб на компенсацию огромной смертности икры и личинок при полном отсутствии родительской заботы о потомстве.
В трофическом отношении все эпипелагические рыбы разделяются на две большие группы — мирных планктофагов и хищников (преимущественно ихтиофагов). При этом рыб, питающихся исключительно мелким планктоном, в эпипелагиали открытого океана сравнительно немного. Наибольшей численности среди них достигают голоэпипелагические макрелещуковые и летучие рыбы, а также мероэпипелагические светящиеся анчоусы и выходящие в открытые воды рыбы прибрежного происхождения — лососи, сельдевые, ставриды, скумбрии, тресковые, возможно, нототениевые. К этой группе относятся, естественно, и все планктонные личинки и мальки рыб, которые могут использовать в пищу даже фитопланктон. Почти у всех эпипелагических планктофагов пищевые спектры очень широки, а локальные и сезонные изменения состава их пищи определяются в значительной степени только наличием доступных кормовых объектов.
Кончилась очередная океанологическая станция, опустела палуба. Судно на малом ходу неторопливо «переползает» с одной волны на другую, но забортные работы еще не завершены. С блока крамбола косо протянулся к корме и режет воду, уходя вглубь, напряженный, как струна, трос: идет траление разноглубинным тралом Айзекса-Кидда... Полчаса - час лова на выбранном горизонте (скажем, на глубине 1000 м), и вот трал поднят на борт грузовой стрелой - в свежую погоду эта операция не так уж проста, и требуется приложить немало мускульных усилий, чтобы усмирить летающий над палубой 100-килограммовый депрессор-заглубитель. Наконец, сетной мешок развязан и улов высыпан в таз. Чего там только нет. Ярко-красные креветки с длинными усами-антеннами, полупрозрачные миниатюрные кальмарчики, сальны, пиросомы, медузы... Но сейчас нас интересуют только рыбы. Их в трале больше, чем беспозвоночных, но какие же они странные, непохожие на тех, к которым мы привыкли с детских лет. Светящиеся анчоусы сверкают сине-зелеными или голубыми пуговичками фотофоров. У змеевидно удлиненного бархатно-черного идиаканта, кроме золотых фотофоров, светятся еще и фиолетовые пятна железистой ткани на теле. Еще более длинные нитехвостые угри удивляют своими клювовидными тонкими челюстями. Зубастый хаулиод, раскрыв пасть, демонстрирует устрашающие непропорционально большие клыки. Рыбы-топорики действительно очень напоминают хорошо известный плотницкий инструмент, в миниатюре отлитый из никеля или серебра да еще и украшенный изумрудами фотофоров.
Среди глубоководных пелагических рыб наибольшим разнообразием выделяется семейство светящихся анчоусов, или миктофовых (Myctophidae), детально изученное в последние годы трудами В. Э. Беккера (Институт океанологии АН СССР), Б. Нафпактитиса, Дж. Пэкстона, Б. Халли и других ихтиологов. Оно насчитывает более 210 видов, 92 из них входят в состав двух больших родов — Diaphus и Lampanyctus. Миктофиды — некрупные рыбки длиной от 2,5 до 25 см, большеглазые и большеротые (ротовая щель далеко заходит за вертикаль заднего края глаза), но в целом имеющие вполне стандартный «рыбообразный» облик (рис. 17). Их своеобразие состоит, однако, в необычайно сильном развитии светящихся органов — фотофоров и разнообразных желез на голове и туловище.

Особенно сложное строение имеют фотофоры — одинаковые по форме круглые бляшки, число которых у разных видов варьирует от 50 до 80 (их нет только у одного, самого глубоководного вида из рода Taaningichthys). Эти бляшки располагаются, как правило, в нижней части тела и сгруппированы в серии, генеральная топография которых характерна для всех представителей семейства (только у Scopelopsis multipunctatus мелкие фотофоры усеивают всю поверхность головы и туловища). В то же время число фотофоров в сериях и их взаимоположение специфичны для каждого вида. Основой отдельного фотофора является погруженная в кожу чешуя, которая видоизменена в чашечку, выстланную пигментным и отражающим слоями и содержащую внутри нежные светопродуцирующие клетки, окруженные студенистой тканью. Чешуйка, прикрывающая все эти образования снаружи, модифицирована в совершенно прозрачную двояковыпуклую линзу. Каждый фотофор имеет независимое кровоснабжение и иннервируется особым нервным ответвлением. Его голубоватый свет испускается в результате химической реакции, которая сводится к окислению специфического вещества — люциферина, выделяемого железистыми клетками, в присутствии особого фермента люциферазы.
Светящиеся железы миктофовых рыб группируются в несколько типов. Так, у представителей большинства родов (за исключением Diaphus и Gynmoscopelus) развиты хвостовые железы, расположенные на верхнем и нижнем краях стебля. Они могут быть маленькими или большими, одиночными или составленными серией чешуевидных пластинок, простыми по строению или сложно организованными — с пигментной выстилкой, рефлектором и линзой. В некоторых родах (Lampanyctus и др.) верхне- и нижнехвостовые железы одинаково развиты у особей обоих полов, в других (Myctophum и др.) самцы имеют только верхние железы, а самки только нижние. Иногда такие железы присутствуют только у самцов или развиты у них гораздо лучше, чем у самок (род Electrona). Кроме хвостовых, у всех миктофид есть одна или несколько пар окологлазничных желез, которые лучше всего развиты у видов рода Diaphus. У некоторых из них эти железы очень велики и занимают почти всю предглазничную часть головы. В их строении и особенно размерах также нередко наблюдается половой диморфизм. Набор желез еще одного, последнего типа — чешуйковидных или аморфных скоплений светящейся ткани у оснований плавников, на верхней поверхности головы и в иных местах сильно варьирует у представителей разных родов.
Функция системы светящихся органов, по-видимому, многозначна, но главное ее назначение состоит, без сомнения, в обеспечении распознания особей своего вида, а в некоторых случаях и их пола. Сосредоточение светопродуцирующих источников преимущественно на нижней части тела позволяет предполагать, что здесь, имеет место и так называемый противотеневой эффект, способствующий лучшей маскировке рыбы на фоне рассеянного верхнего освещения. Есть также предположения, что вспышки хвостовых желез могут дезориентировать преследующего хищника.
Систематика миктофид, которую нередко вполне серьезно именуют «миктофидологией», - очень своеобразный раздел ихтиологической науки. Миктофидологи (в нашей стране эта специфическая профессия полноправно представлена В. Э. Веккером) определяют роды и виды, рассматривают их родственные отношения, строят систему семейств, основываясь главным образом на расположении фотофоров, каждую группу которых принято обозначать определенной буквенной аббревиатурой, а отдельные органы в ее составе — порядковыми номерами. Поэтому научные рисунки светящихся анчоусов часто напоминают технические чертежи, а видовые описания типа «SA01 под V04 или впереди его вертикали; SA02 примерно на вертикали AOa1; SA03 над АОа2 и т. д. понятны «без перевода» только тем, кто хорошо знаком с этой группой рыб.
Такая «геометричность» миктофид, казалось бы, очень облегчает их опознание: на схематических рисунках различия между видами очень наглядны и очевидны. Это, однако, вовсе не так. На самом деле в уловах глубоководных тралов крайне редко можно встретить идеально сохранившийся экземпляр с полным набором светящихся органов, - а установление видовой принадлежности по сохранившимся на своих местах остаткам полуободранных фотофоров — процедура, которая не терпит стандартного подхода. Определение поврежденных миктофид, на мой взгляд, имеет нечто общее с разгадыванием кроссворда, и поиск верного решения сам по себе очень увлекателен. Я люблю возиться с этими симпатичными рыбками, знаю многих из них «в лицо» и узнаю некоторых даже при полном отсутствии кожи, но тем не менее очень часто пасую перед непреодолимыми трудностями, доводя идентификацию только до родового уровня.
Светящиеся анчоусы очень широко распространены во всех широтах Мирового океана (их нет только в Центральном полярном бассейне), но достигают наибольшего видового разнообразия в тропических и субтропических водах. Все они принадлежат к числу океанических рыб, причем большинство видов постоянно обитают в толще воды и лишь немногие ведут бенто-пелагический образ жизни у материковых склонов и подводных поднятий. Среди пелагических миктофид резко преобладают среднеглубинные виды, которые держатся в светлое время суток на глубине порядка 300—1500 м. Для них очень характерны суточные вертикальные миграции с ночным подъемом на несколько сот метров в составе так называемых мигрирующих звукорассеивающих слоев (Звукорассеивающие слои - регистрируемые гидроакустическими приборами (эхолотами) слои, образованные скоплениями макропланктона.) (ЗРС) — комплекса макропланктонных и микронектонных животных, перемещающихся по вертикали в течение суток. В тропиках эти миграции, как правило, полностью укладываются в пределы основного термоклина (в высоких широтах верхняя граница сезонного слоя скачка также лимитирует выход миктофид к поверхности в летне-осеннее время). В то же время представители тепловодных родов Myctophum (кроме М. selenops), Symbolophorus, Gonichthys, Centrobranchus и Loweina (всего около 40 видов), относящихся к никтоэпипелагической группе, проникают ночью к самой поверхности воды. Батипелагические миктофиды (Taaningichthys, некоторые виды Lampanyctus и Lampadena) не меняют существенно горизонта своего обитания в течение суток.
Пелагические миктофиды — типичные макропланктонные рыбы, постоянно дрейфующие вместе с перемещающимися водами. Поэтому почти все видовые ареалы этих рыб находят объяснение в приуроченности их основ к тем или иным крупномасштабным круговоротам (из этого правила существуют, однако, отдельные не очень понятные исключения). Пассивным концентрированием особей определяются по-видимому, и скопления миктофид в некоторых районах схождения течений, например вдоль Южного полярного фронта.
Биология светящихся анчоусов изучена явно недостаточно. Известно, однако, что они почти безвыборочно питаются разнообразным мезопланктоном и принадлежат к числу консументов второго — третьего уровня. Продолжительность их жизни невелика: большинство тропических и мелкие умеренноводные виды живут всего 1—2 года, а более крупные умеренноводные — 4—5 (по другим данным — 7—8) лет. В тропической зоне миктофиды размножаются круглогодично, а за ее пределами преимущественно в теплое время года, причем их личинки доминируют в эпипелагическом ихтиопланктоне всех районов открытого океана (это является косвенным доказательством их высокой численности). Некоторые тропические формы (в частности, никтоэпипелагические виды из родов Myctophum и Symbolophorus, а также Notolychnus valdiviae и др.) относятся к моноцикличным рыбам: они созревают уже в возрасте нескольких месяцев и после довольно продолжительного многопорционного нереста погибают примерно в годовалом возрасте. Индивидуальная плодовитость разных видов сильно варьирует, достигая нескольких тысяч икринок, и находится в прямой зависимости от размеров производителей.
Как правило, миктофиды держатся разреженными стайками, хотя в некоторых участках океана (например, в зоне антарктической конвергенции) они образуют более или менее значительные скопления. Такие скопления более характерны, однако, для отдельных участков над материковым склоном, и образуют их в основном специфические «присклоновые» виды. Для одного такого вида — Ceratoscopelus maderensis — у восточного побережья Северной Америки отмечены на глубине 600 м плотные линзовидные стаи диаметром до 100 м и толщиной до 10 м, в которых плотность рыб достигает 10—15 экз./м3 (в одной стае, следовательно, может насчитываться 300—400 тыс. особей средней длиной около 6 см). В целом светящиеся анчоусы, несомненно, являются наиболее многочисленной группой мезопелагических рыб, играющей огромную роль в пищевых цепях открытого океана.
У многих рыб вертикальные миграции начинают формироваться еще на личиночной стадии развития. Известно, например, что личинки многих массовых миктофид встречаются у поверхности воды только днем (это пример так называемой инвертированной, или обращенной, миграции), что прямо связывается с существованием у них суточного хода двигательной активности, обусловленной ритмом питания.
Размах вертикальных перемещений может сильно варьировать. Наибольшей протяженности — с глубины 400— 800 (1000) м до поверхности — они достигают у никтоэпипелагических рыб, уже упоминавшихся выше. К ним относятся около 40 видов светящихся анчоусов из 5 родов, акула-карлик, змеиная макрель, которые, по-видимому, поднимаются к поверхности каждую ночь, а также отдельные виды винцигуерий и астронестов (например, Astronesthes aiger), у которых такая миграция не имеет массового характера, и значительная часть популяции остается ночью в подповерхностных слоях.
У преобладающего большинства тропических мезопелагических рыб вертикальные миграции полностью укладываются в пределы своего биотопа — основного термоклина (т. е. между нижней границей поверхностного изотермического слоя и верхней границей глубинных однородных вод). При этом положение верхней границы ночного подъема разных видов довольно сильно варьирует: по-видимому, оно контролируется в основном температурой воды (может быть, также положением слоя концентрации пищевых объектов). Известную роль в размахе вертикальной миграции играет и освещенность (многие миктофиды, например, поднимаются в новолуние на 30—50 м выше, чем в полнолуние). Имеются различия и в батиметрическом распределении разных размерно-возрастных групп одного вида. У некоторых видов (из родов Diaphus, Ceratoscopelus и др.) в нижние слои эпипелагиали проникают, как правило, только неполовозрелые особи.
Мигрирующие по вертикали рыбы перемещаются в толще воды в многовидовых ассоциациях — звукорассеивающих слоях, образующих особые сообщества неполного состава (без автотрофного звена), которые можно называть схизоценами. Эти слои объединяют рыб, креветок, эвфаузиид, кальмаров, киленогих и крылоногих моллюсков, сифонофор, сальп и других животных. Суточные миграции ЗРС (как и другие виды вертикальных перемещений) связывают мезопелагических консументов с пищевыми ресурсами вышележащих горизонтов и имеют существенное значение в перераспределении органического вещества в океане. Эти слои имеют очень широкое распространение и наблюдаются почти повсеместно. В продуктивных районах часто формируются сразу несколько звукорассеивающих слоев, перемещающихся по-разному, хотя иногда и сливающихся вместе ночью или днем. Основную роль в мигрирующих ЗРС играют макропланктонные рыбы-планктофаги, среди которых резко преобладают миктофиды из родов Diaphus, Ceratoscopelus, Lampanyctus, Hygophum и др., составляющие до 80—90% общей численности рыб в схизоцене. В некоторых участках океана к доминирующим формам относятся также винцигуерии и батилагиды. Хищные рыбы представлены в сообществах ЗРС хаулиодами, стомиями, гоностомами, астронестами, идиакантами и другими относительно крупными видами отряда Stomiiformes, питающимися только в пределах самого слоя. Глубины дневного и ночного положения звукорассеивающих слоев колеблются в довольно значительных пределах — у светящихся анчоусов, например, амплитуда суточных перемещений варьирует от 200 м (Diaphus regani) до 700—800 м (Ceratoscopelus warmingii), — но почти всегда остаются в границах мезопелагиали, т. е. зоны, занятой промежуточными водами. В светлое время суток мигрирующие рыбы живут на глубине 300—1000 м (обычно глубже 400—500 м), а ночью концентрируются в слое от 50-100 до 300 м.
Икра и ранние личинки самых разных рыб глубоководной пелагиали — светящихся анчоусов, циклотон, удильщиков — проходят развитие в поверхностных горизонтах, но размножающихся особей в эпипелагиали никогда не наблюдали. Логично заключить, что нерест происходит на глубине обычного обитания взрослых рыб, откуда оплодотворенные икринки, обладающие положительной плавучестью, быстро всплывают. Вылупившиеся личинки по мере развития и роста постепенно опускаются все глубже и глубже.
Насколько сейчас известно, почти все глубоководные пелагические рыбы относятся к порционно нерестующим. Число икринок в порции подвержено довольно сильной изменчивости. В районе Гавайских островов оно варьирует, по данным Т. Кларка, у миктофид от 30—150 (крохотный Notolychnus valdiviae длиной не более 2,5 см) до 2,6—9,5 тыс., а у других мезопелагических рыб от 70—250 до 27—61 тыс. ооцитов (относительно крупная Gonostoma elongatum длиной до 27,5 см), причем общее число порций составляет 10—20. Антарктический батилаг (Bathylagus autarcticus) выметывает от 250 до 1300 икринок в 3—4 порциях, а североатлантический светящийся анчоус (Benthosema glaciale) — 190—470 икринок, возможно, при разовом нересте. В яичниках «светлых» циклотон насчитывали 0,1—0,9 тыс. ооцитов, «черных» циклотон — около 2—10 тыс., угря Derichthys serpentinus — 4,1 тыс., рыб-удильщиков, как правило, 7,5—10,8 тыс., но у очень крупной (65 см) самки цератии (Ceratias holboelli) — почти 5 млн. ооцитов. Суммарная абсолютная плодовитость у рыб мезо- и батипелагиали в общем невелика и заметно уступает плодовитости эпипелагических рыб, которые, впрочем, имеют гораздо более крупные средние размеры. Относительная плодовитость (число икринок на 1 г массы тела) у глубоководных видов также достоверно меньше, чем у приповерхностных.
Размножение глубоководных пелагических рыб в тропических водах, по всей вероятности, продолжается в течение всего года, но у краев тепловодной зоны либо имеет пик в летне-осеннее время, либо вообще ограничено этим сезоном. В то же время в субполярных водах существуют как зимненерестующие, так и летненерестующие виды (в Северной Пацифике, например, к числу первых принадлежит миктофида Stenobrachius leucopsarus).
Некоторые из упоминавшихся специфических особенностей батипелагических рыб, связанные с их размножением, в частности, половой диморфизм по размерам (и крайнее его проявление — самцовый паразитизм) и по строению органов обоняния, а также синхронный гермафродитизм, обеспечивающий возможность нереста любой пары случайно встретившихся производителей, несомненно, объясняются трудностями отыскания партнеров в условиях разреженного существования.
Данных по возрасту глубоководных рыб пока совсем немного, и почти все цифры, приводимые здесь, получены Дж. Фитчем и Р. Лавенбергом при исследовании отолитов отдельных экземпляров из вод, прилегающих к берегам Калифорнии. Продолжительность жизни этих рыб в общем невелика. Скорее всего созревают (еще до наступления годовалого возраста) и меньше всего живут (1,5—2 года) мигрирующие в эпипелагиаль тропические миктофиды и винцигуерии: до наступления половой зрелости они быстро растут, а затем почти беспрерывно нерестятся вплоть до самой смерти. Так, Benthosema pterotum, наиболее массовый вид миктофид Аравийского моря, становится половозрелой к 6—7 месяцам жизни при длине порядка 40 мм (максимальная длина этого вида 50 мм) и гибнет после окончания нерестового периода в возрасте чуть больше 1 года. Созревание слабо мигрирующих и постоянно остающихся на глубинах, а также обитающих в высоких и умеренных широтах рыб наступает позже (у батипелагического угря Serrivomer sector, например, в возрасте около 4, у северного светящегося анчоуса Benthosema glaciale — в 2—3 года). Продолжительность их жизни составляет от 3—5 (циклотоны, рыбы-топорики, батилаги, умеренно-холодноводные и глубинные миктофиды, веретенники рода Lestidium и др.) до 6—8 лет (хаулиоды, идиаканты, кинжалозуб, китовидки, живоглоты) и даже 10—18 лет (представители родов Poromitra, Nemichthys, Gigantactis). Это свидетельствует о более высоком уровне энергетических процессов у мигрирующих по вертикали видов по сравнению с ограниченно мигрирующими и немигрирующими.
В каждом биотопе глубоководной пелагиали есть и планктоядные и хищные рыбы. Среди мезопелагических планктофагов доминируют, как уже отмечалось, мигрирующие по вертикали миктофиды и винцигуерии. Исследования, выполненные в последние годы Т. А. Гореловой, Т. Кларком и другими специалистами, показывают, что те из них, которые достигают ночью поверхности, питаются в основном приповерхностными беспозвоночными (копеподы понтеллиды и др.), а перемещающиеся в составе ЗРС без выхода в верхний однородный слой — мигрирующим вместе с ними интерзональным мезопланктоном (главным образом также ракообразными — копеподами, остракодами, амфиподами, эвфаузиидами) и являются основными потребителями этого планктона. Рыбы-мигранты питаются по преимуществу ночью или в вечерние и утренние часы (т. е. на закате и восходе солнца) и, подобно всем прочим зоофагам океанской толщи, захватывают обнаруженную ими (как правило, визуально) добычу, какой бы размер она ни имела, поодиночке (единственным фильтратором среди глубоководных рыб является, насколько сейчас известно, большеротая акула мегахазма). Об этом свидетельствует, в частности, явное преобладание в их желудках непрозрачных, более или менее контрастных по окраске объектов. Несмотря на очень широкие пищевые спектры и их большое сходство у разных видов тропических миктофид, между ними, по меньшей мере в некоторых районах, существуют определенные различия в составе пищи, которые, по всей вероятности, связаны с различиями в длине рыб (корреляция между размерами потребителя и жертвы часто выражена довольно явно) и в видовом составе планктона на горизонтах их ночного подъема. Это, по-видимому, способствует снижению межвидовой конкуренции за пищу в условиях ограниченных трофических ресурсов, поскольку у миктофид, живущих в более продуктивных по планктону умеренных и холодных водах, различий в составе пищи фактически не обнаружено. В олиготрофных участках океана миктофиды демонстрируют исключительно высокую кормовую пластичность: Ceratoscopelus warmingi, например, может использовать в пищу не только излюбленных копепод, но также хетогнат, оболочников, сифонофор, личинок рыб и даже фитопланктон.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Семейство Миктофовые, или Светящиеся анчоусы (Myctophidae)
Семейство светящихся анчоусов, или
миктофовых, содержит 30 родов и около
200 видов и является, таким образом,
наиболее крупным в подотряде
миктофовидных (Myctophoidei) и одним из самых
больших семейств глубоководных
костистых рыб вообще. Светящиеся анчоусы
необычайно широко распространены в
Мировом океане — разные виды этой группы
известны от Шпицбергена и северной части
Берингова моря до шельфовых ледников
Антарктиды. Наконец, многие виды
семейства достигают огромной численности
и представляют определенный
практический интерес. Все это делает
светящихся анчоусов чрезвычайно перспективным
и многообещающим объектом
исследования, так как на их примере могут быть
выяснены общие черты биологии и
закономерности распространения, присущие
многочисленным обитателям средних
слоев пелагиали Мирового океана.
Светящиеся анчоусы — небольшие
рыбки, обычной «рыбообразной» формы.
Взрослые особи самых мелких видов —
нотолихна (Notolychnus valdiviae), диогенихта (Diogenichthys atlanticus) имеют
длину всего 2,5—3 см, а длина наиболее
крупных экземпляров некоторых видов
рода Лампаникты (Lampanyctus) и рода
Нотоскопелы (Notoscopelum) не превышает
20—30 см. Все тело этих рыб покрыто
довольно крупной чешуей, как правило,
циклоидной, и только у нескольких видов
имеется ктеноидная чешуя с более или менее
развитыми шипиками по заднему краю.
Боковая линия обычно хорошо развита, но у
видов примитивного рода Протомиктофы
(Protomyctophum) и видов рода Центробранх (Centrobranchus) она отсутствует.
Плавники довольно большие, и только у
некоторых видов рода Лампаникт
(Lampanyctus) грудные плавники очень малы,
а у одного из них (Lampanyctus achirus) вовсе
отсутствуют. Анальный плавник начинается
под концом основания спинного или сразу
за ним. На верхней стороне хвостового
стебля, над концом анального плавника,
всегда имеется хорошо развитый жировой
плавничок.
Голова у светящихся анчоусов довольно
большая, ее длина составляет от одной
четверти до одной трети длины тела (без
хвостового плавника). Размер глаз у этих
рыб значительно варьирует, от средней
величины до очень больших, почти до
трети длины головы. Несколько видов рода
Протомиктоф (Protomyctophum)
отличаются так называемым
«полутелескопическим» строением глаз (у них зрачок
смещен вверх). Вооруженный
многочисленными мелкими зубами рот светящихся
анчоусов простирается за задний край
глаза. Именно большой рот этих рыб,
похожий на рот настоящих анчоусов
(семейство Engraulidae), и послужил
поводом для их названия, хотя в
действительности эти две группы довольно далеки
друг от друга и принадлежат к разным
отрядам.
Наиболее характерной чертой внешнего
облика светящихся анчоусов являются
различного рода светящиеся органы. По
местоположению и строению их можно
разделить на четыре типа: фотофоры и
хвостовые, окологлазничные и туловищные
железы. Наиболее интересно и сложно
устроены фотофоры. В отличие от таких
же органов неоскопелов (семейство Neoscopelidae) каждый фотофор светящихся
анчоусов помещается в чашеобразной
впадине мышц, резко отграничен от
окружающих тканей, имеет самостоятельное
кровоснабжение и обслуживается
специальным нервом. Со стороны тела фотофор
подстилается слоем черного пигмента,
который немного выступает за края органа
и на поверхности тела окружает его
сплошным черным кольцом. На пигментной
выстилке лежит блестящий рефлектирующий
слой. Затем следуют студенистая ткань
и продуцирующие свет железистые клетки. Снаружи фотофор прикрыт
измененной в виде линзы чешуйкой. Под линзой
находится полулунная шторка — септа
(особенно хорошо заметная у диафов —
видов рода Diaphus), с помощью
которой рыба, по-видимому, может
произвольно изменять силу и направление света.
Располагаются фотофоры почти у всех
видов семейства только на нижней
половине тела, ниже боковой линии, и образуют
вполне определенные группы, каждая из
которых имеет свое название. Число фотофоров в группах и их взаимоположение
специфичны для вида и, вероятно, играют
определенную роль в поведении этих рыб.
Кроме только что описанных «основных»
фотофоров, у нескольких видов имеются
и так называемые «вторичные» фотофоры,
располагающиеся на каждой чешуйке или
только на некоторых из них. Размеры
«вторичных» фотофоров, как правило,
меньше, чем «основных», и лишь у скопелопсиса (Scopelopsis multipunctatus)
величина тех и других органов примерно
одинакова.
Хвостовые светящиеся железы
встречаются у многих видов семейства и могут
иметь различное строение. Иногда это
сложные органы, имеющие, как и
фотофоры, черную пигментную выстилку,
рефлектор и линзоподобные структуры. В
таких случаях хвостовые железы играют
роль вторичных половых признаков,
причем у самцов они обычно устроены
сложнее и помещаются на верхней стороне
хвостового стебля, а у самок их строение
проще и располагаются они на нижней
стороне тела, между анальным и хвостовым
плавниками. Такие железы отсутствуют
у мальков и развиваются только ко
времени наступления половой зрелости. У
других видов (рода Lampanyctus и
близких к нему) хвостовые железы
представляют собой простые белые или
желтоватые чешуевидные пластинки, лежащие на
поверхности тела. В таких случаях они
одинаковы у рыб разных полов и
появляются во время развития очень рано,
одновременно с появлением фотофоров.
Окологлазничные светящиеся железы
особенно характерны для диафов
(видов рода Diaphus), хотя имеются и у
многих других представителей семейства.
Строение этих желез у разных видов диафусов различно и часто служит хорошим
признаком для их определения. Величина
их также значительно варьирует: от
маленьких, округлых, не более фотофора,
железок до огромных и сложных органов,
занимающих всю предглазничную область
головы. Интересно, что у диафов,
которые не имеют хвостовых светящихся
желез, половой диморфизм выражен в
строении и величине окологлазничных
светящихся органов (у самок многих видов
они меньше, чем у самцов).
Туловищные светящиеся железы
представляют собой участки светящейся ткани
без каких-либо дополнительных устройств
вроде рефлектора или линзы.
Располагаются такие железы на поверхности
тела (и очень легко утрачиваются, не
оставляя, в отличие от фотофоров, никаких
следов), обычно вдоль оснований
плавников и реже между ними, вдоль спины или
брюшка. Туловищные железы особенно
характерны для высокоорганизованных
форм семейства, таких, как лампаникты
(Lampanyctus) и близкие к ним.
Продолжительность жизни светящихся
анчоусов невелика. Особи сравнительно
крупного светлого лампаникта (Stenobrachius leucopsarus), достигающего
длины около 10 см, живут всего 3, редко 4
года. Срок жизни более мелких форм (Notolychnus, Diogenichthys,
Centrobranchus и т. д.), вероятно, еще меньше.
Светящиеся анчоусы всю свою жизнь
проводят в толще вод открытого океана.
В тропической области их размножение
происходит в течение круглого года. В
субтропических и умеренных водах имеет
место, по крайней мере для некоторых
видов, сезонность размножения. Мелкие
личинки светящихся анчоусов держатся
у поверхности моря и входят в состав
поверхностного планктона. По мере роста,
с переходом к мальковой фазе развития,
молодь опускается в более глубокие слои
воды и постепенно переходит к образу
жизни взрослых рыб. Плодовитость
светящихся анчоусов довольно велика, и самки
длиной около 7 см выметывают 7—8 тыс.
икринок.
Пищу светящихся анчоусов составляют
преимущественно планктонные
ракообразные, но иногда мальки и личинки других
рыб. Хотя светящиеся анчоусы
приспособлены к питанию мелкими организмами,
иногда они заглатывают и довольно
крупную добычу.
Массовые виды светящихся анчоусов
в свою очередь служат важными
объектами питания более крупных рыб
(лососей, морских окуней, дальневосточной
трески, отчасти некоторых тунцов и марлинов), а также морских котиков и
китов. Один из видов рода нотоскопелов
(Notoscopelus) составляет в пище котиков
на местах их зимовки в водах Японии
около 60% всего их рациона. Примерно
столько же составляют они и в питании
дальневосточных лососей в районах их
откорма.
Светящиеся анчоусы населяют
преимущественно верхний тысячеметровый слой
вод океана и на глубинах более 1000 м
уступают свое место наиболее
многочисленной группе — циклотонам (Cyclothone,
семейство Gonostomidae). Среди них есть,
однако, виды, которые, насколько
сейчас известно, не встречаются выше 700—800 м (Taaningichthys bathyphilus). По
наблюдениям с французского батискафа
FNRS-3 в Средиземном море, светящиеся
анчоусы встречаются до глубины 2300 м
(до максимальной глубины этого
погружения).
По-видимому, большинство светящихся
анчоусов совершают суточные
вертикальные миграции и в темное время суток
поднимаются в верхние слои воды, а с
рассветом покидают их и уходят на глубину.
В этом отношении семейство может быть
разделено на две группы. Одна из них
включает 30—40 видов так называемых
«приповерхностных» светящихся
анчоусов (роды Myctophum, Symbolophorus,
Gonichthys и др.). Характерной
особенностью этих видов является то, что они
в темное время суток поднимаются к
самой поверхности воды и, таким образом,
ведут в это время отнюдь не
глубоководный образ жизни. На этом основании их
следует рассматривать в качестве
временного компонента приповерхностного
комплекса рыб (постоянными членами
которого являются летучие рыбы, корифены,
тунцы и т. д.). Все приповерхностные
виды обладают положительной реакцией
на свет и привлекаются его
искусственными источниками. В свете лампы,
опущенной тропической ночью с борта судна,
почти всегда можно наблюдать этих
стремительных, сверкающих серебром рыбок,
преследующих добычу или преследуемых
кальмарами или гемпилами (змеиными
макрелями).
Вторая группа светящихся анчоусов
объединяет остальные виды семейства.
Они в ночное время также поднимаются
на меньшие глубины, но, как правило, не
достигают поверхности моря. Известно,
что многие из этих видов входят в состав
населения звукорассеивающих слоев и
мигрируют вместе с ними.
В связи с различиями в глубине
обитания светящихся анчоусов изменяются
и некоторые черты их строения.
Особенно интересны в этом отношении
изменения размеров испускающих и
воспринимающих свет органов, т. е. фотофоров
и глаз. Оказывается, что самые крупные
глаза и фотофоры присущи не наиболее
глубоководным, а, напротив,
поднимающимся к поверхности видам. С
увеличением глубины обитания и те и другие
органы закономерно уменьшаются. То же
явление известно и для других батипелагических рыб. Исчерпывающего
объяснения ему пока не найдено. По-видимому,
прямой связи между изменениями
величины фотофоров и глаз не существует.
Если размеры фотофоров
пропорциональны яркости свечения и если верно,
что они создают «противотеневой эффект»
(об этом подробнее говорилось в очерке
о семействе гоностомовых), тогда можно
полагать, что уменьшение их величины
у более глубоководных форм является
прямым следствием уменьшения яркости
верхнего фона в результате поглощения
дневного света в толще воды. Понятно,
что для создания «противотеневого
эффекта» на менее ярком фоне нужны менее
яркие источники света.
Как уже говорилось, светящиеся
анчоусы представляют собой одну из наиболее
широкораспространенных групп
глубоководных пелагических рыб. Наибольшего
разнообразия они, однако, достигают
в субтропических и тропических водах.
Огромное большинство видов семейства
встречается между 40—45-й параллелями
северного и южного полушарий. К югу
от 45° ю. ш. постоянно обитает около 20
видов и примерно столько же — к северу
от 45° с. ш. В Северной Атлантике,
однако, с мощным и теплым потоком Северо-Атлантического течения многие тепловодные виды проникают и дальше на север,
почти до 60° с. ш. Вообще же
распространение отдельных видов светящихся
анчоусов (как и других мелких
глубоководных пелагических рыб) связано с водами
определенных структур. Так, например,
в Атлантическом океане распространение
нескольких видов этого семейства очень
четко ограничено водами северного
центрального круговорота, образуемого
Гольфстримом, Канарским, Северным
пассатным и Антильским течениями. Та же
картина наблюдается и в северной части
Тихого океана, где целый ряд видов
обитает в круговороте, образуемом Куро-Сиво, его продолжением и Северным
пассатным течением. В средних и высоких
широтах, где основные течения имеют
широтное направление, ареалы светящихся
анчоусов также вытянуты в широтном
направлении, и их распространение в этих
районах носит ярко выраженный
зональный характер. Границы распространения
некоторых видов светящихся анчоусов
оказываются настолько четкими, что по
ним могут быть обозначены
биогеографические зоны. Так, например, в южной
части Тихого океана антарктическую зону
населяет один вид рода — антарктическая
электрона (Electrona antarctica), а
следующую к северу соседнюю нотальную
зону — другой вид (Electrona subaspera).
Обитатели бездны. Уильям Кроми 1971 г.
Второе место по численности в списке глубоководных обитателей по праву занимают светящиеся анчоусы — маленькое, большеглазое создание, очень напоминающее распространенную мелководную разновидность рыб. Большинство их живет на глубине от 100 до 500 метров, но многие ночью поднимаются на поверхность в поисках пищи. Только тут и можно представить, насколько они многочисленны. Однажды ночью английский корабль погоды, находясь в Северной Атлантике, в течение пяти часов плыл по морю мерцающих, прыгающих светящихся анчоусов. Должно быть, то было сказочное зрелище, потому что серебристое брюхо этих рыб, бока, голова и хвост усеяны напоминающими пуговицы огоньками. Иногда огоньки на хвосте настолько ярки, что при их свете можно читать; огоньки на теле сверкают, словно зеленые драгоценные камни, иногда они бывают окрашены в желтый или красный цвет. Хотя существа эти так крохотны, что на фунт пойдут сотни этих рыб, на теле у них не меньше ста огоньков. У каждой из 170 разновидностей свой, характерный рисунок огней. Биб утверждал, что он «с первого взгляда мог определить, какие именно виды и сколько их находится в каждом новом улове, руководствуясь одними лишь светящимися иероглифами».
Зачастую дневная окраска светящегося анчоуса столь же поразительна, как и его ночные огни. У некоторых видов радужная, с медным отливом спина с темно-синими пятнами, разбросанными кое-где, серебристое брюхо и бока с мерцающим розовым или зеленым оттенком.
Расположение и цвет огней может служить признаком пола их обладателя, подобно тому как по оперению можно определить пол птицы. Самцы светящегося анчоуса имеют сверху на хвосте крупные, мощные огни, в то время как у самок на нижней части хвоста находятся более тусклые «лампы». Вероятно, светящийся анчоус узнает своих сородичей по расположению огней на брюхе, а по хвостовым огням определяет их пол.
Живой свет в значительной мере выполняет функции, которые в освещенных водах выполняет цвет. Взять, к примеру, образование косяков. Узнав «родственные» огни, некоторые виды могут формировать косяки; во всяком случае, ориентируясь по однородным огням, находящиеся в одном косяке рыбы держатся вместе.
Биб наблюдал, как «довольно крупные копеподы и другие организмы» устремились в полосу света, отбрасываемого вниз огнями на брюхе светящегося анчоуса. «Вслед за тем рыба, изогнувшись, схватила несколько этих животных».
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
XLIX. Сем. MYCTOPHIDAE — МИКТОФИДОВЫЕ, СВЕТЯЩИЕСЯ АНЧОУСЫ
Myctophidae, Regan, Ann. Mag. Nat. Hist., (8), 7, 1911 : 128. — Parr, Bull. Bingham Oceanogr. Coll., 3, 3, 1928 : 47. — Bolin, Bull. Stanf. Ichth., 1939 : 89. — Fraser-Brunner, Proc. Zool. Soc., London, 118, 4, 1949 : 1063. — Bolin, Rep. Sci. Res. «Mich. Sars» Exped., 4, 2, 7, 1959 : 3.
Scopelidae, Берг, Система рыб, 1940 : 257.
Близко к сем. Sudidae, отличаясь наличием срединного киля на этмоиде и тем, что парасфеноид простирается вверх к лобным между боковыми этмоидальными, тогда как у Sudidae эти кости соприкасаются по средней линии. Зубы на сошнике, если имеются, так же как у Sudidae, представлены 2 хорошо отделенными друг от друга участками. Тело обычно удлиненное, сжатое с боков, голова также сжата с боков, глаза боковые по положению, рот конечный (Regan, 1911а : 128). Парр (Parr, 1928) принимает 3 подсемейства. В Японском море известны представители только подсемейства Myctophinae, которое отличается от других тем, что фотофоры расположены в строгом порядке в виде отдельных рядов, а не в беспорядке, или вообще отсутствуют. Так как при определении родов и видов этого подсемейства основное значение имеет характер расположения светящихся органов, приводим схему их расположения (рис. 149) и пояснительный к ней текст, как они даны в работе Парра (Parr, 1928 : 50, 51, fig. 6) и Болина (Bolin, 1939).
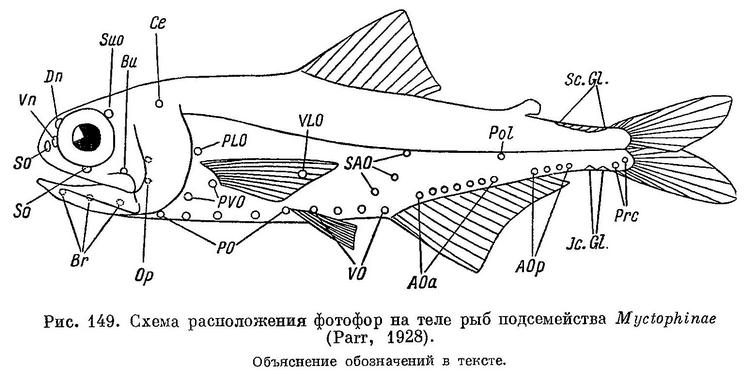
Под термином «фотофор» Парр (Parr, 1928) понимает только мелкие, правильно округлые или почковидные светящиеся органы, входящие в состав указанных на схеме рядов на теле и голове, но не другой тип светящихся органов в форме чешуй или пластинок. Бесцветная центральная часть фотофора на теле окружена отчетливо выраженным черным кольцом; фотофоры на голове могут быть несколько отличными; их расположение на схеме и их название даны по Болину (Bolin, 1939) из работы Е. Б. Куликовой (1961 : 9, рис. 2). В схеме (рис. 149) ряды фотофор носят следующие названия. Эта терминология приведена в работе Брауэра (Brauer, 1906).

Буквенные обозначения отдельных групп фотофор у рыб подсемейства Myctophinae принимаются без изменения всеми последующими исследователями для того, чтобы обеспечить сравнение с прежними описаниями, но, как указывает Фрезер-Бруннер (Fraser-Brunner, 1949), эти буквенные обозначения следует рассматривать только как простые значки, а не как сокращение названий этих групп, так как в ряде случаев эти названия не соответствуют действительному расположению фотофор.
Рыбы этого семейства признавались глубоководными (Gilbert, 1913 : 68), но, как указывает Фрезер-Бруннер (Fraser-Brunner, 1949 : 1020), они обычны не только на глубинах, но также в верхних слоях океанов и даже встречаются на поверхности, а поэтому указания на нахождение представителей этого семейства в Японском море не противоречат представлению о молодости больших глубин этого моря.
Из большого числа родов этого семейства (Fraser-Brunner, 1949; Parr, 1929, 1931, 1934) в Японском море обнаружено 2 рода, кроме того, еще 1 род в Желтом море у берегов п-ова Корея и 1 род в южной глубоководной части Охотского моря.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. MYCTOPHIDAE
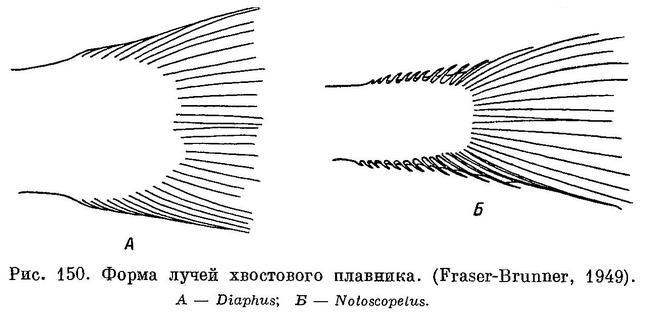
1 (4). Укороченные лучи хвостового плавника мягкие (рис. 150, А).
2 (3). Основание анального плавника много длиннее основания спинного. 1. [Myctophum Rafinesque].
3 (2). Основание анального плавника не длиннее или очень немного длиннее основания спинного, обычно несколько короче. 2. Diaphus Eigenmann et Eigenmann.
4 (1). Укороченные лучи хвостового плавника жесткие, в виде колючек (рис. 150, В).
5 (6). Основание спинного плавника не длиннее анального. 3. [Lampanyctus Bonaparte].
6 (5). Основание спинного плавника длиннее анального. 4. Notoscopelus Günther
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
XX. Сем. MYCTOPHIDAE — СВЕТЯЩИЕСЯ АНЧОУСЫ
Myctophidae Jordan a. Evermann, Fish. N. America, I, 1896 : 550. — Regan, Ann. Mag. Nat. Hist., (8), VII, 1911 : 128. — Parr, Bull. Bingh. Oceanogr. Coll., III, 3, 1928 : 47—156 (монографич. обзор подсемейства). — Taning, Vidensk. Meddel. Dan. Naturh. Foren. Københ., 86, 1928 : 49—69 (обзор североатлантических видов). — Воlin, Stanf. Ichth. Bull., I, 4, 1939 : 89—156 (обзор тихоокеанско-американских видов). — Fraser-Brunner, Proc. Zool. Soc. London, 118, pt. II, 1949 : 1019—1106 (всесветная ревизия семейства).
Scopelidae Günther (1864) et auctorum.
Тело умеренно удлиненное, сжатое с боков; покрыто крупной легко опадающей чешуей. Рот большой, конечный. Весь край верхней челюсти образован межчелюстной. Мезокоракоида нет. Этмоид со срединным гребнем. Парасфеноид впереди, между ethmoidalia lateralia, доходит до лобных костей. Жаберные отверстия продолжены вперед, перепонки не приращены к межжаберному промежутку. Жаберных лучей 8—10. Interoperculum не редуцировано. Боковая линия имеется. На боках тела обычно правильно расположенные светящиеся органы — фотофоры (нет у Scopelengys Alcock). Кроме фотофор могут быть еще светящиеся железы на голове и хвостовом стебле. Жировой плавник имеется. Брюшные плавники позади грудных, из 6—11 лучей. Известны по крайней мере с миоцена.
В диагностике многочисленных видов сем. Myctophidae большое значение имеет число и расположение фотофор, для условного обозначения которых обычно принимается следующая схема (рис. 61):
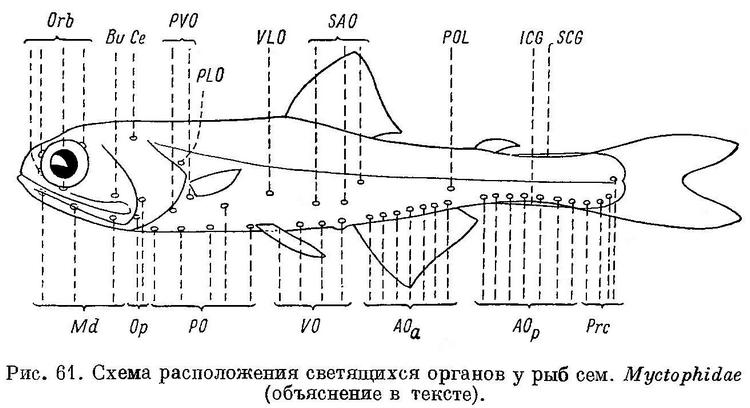
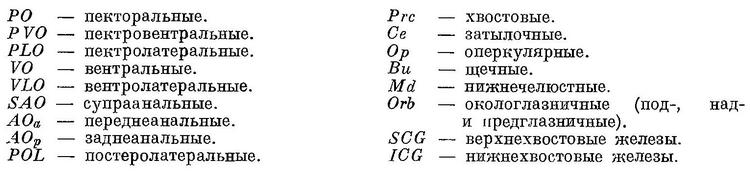
Мелкие батипелагические рыбки, широко распространенные во всех океанах. Хозяйственного значения не имеют; некоторые виды служат пищей пелагическим хищным рыбам. Из 24 родов семейства в северных морях могут встретиться 2 вида, относящиеся к 2 близким родам; их можно различать по следующим признакам:
1 (2). Основание спинного плавника заметно короче основания анального. Грудной плавник длиннее брюшного. Хвостовых фотофор 2.
Диаметр глаза более 30% длины головы. 1. Myctophum glaciale (Reinh.).
2 (1). Основание спинного плавника заметно длиннее основания анального. Грудной плавник короче брюшного. Хвостовых фотофор 3—4.
Диаметр глаза менее 30% длины головы. 2. Notoscopelus elongatus (Costa).