(Callionymus Linnaeus, 1758) Род Каллионимусы, Пескарки, Дракон-рыбы, Genus Callionymus Linnaeus, 1758 (Dragonets, Stinkfishes) 98 видов
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 4. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1975 г.
2. Род CALLIONYMUS LINNE, 1758 — ДРАКОН-РЫБА
Callionymus Linnaeus, Syst. Nat., ed. X, pt. 1, 1758 : 249 (тип: Callionymus lyra Linne).
Calliurichthys Jordan et Fowler, Proc. U. S. Nat. Mus., 25, 1903 : 941 (тип: Calliurichthys japonicus Jordan et Fowler).
Два спинных плавника. В первом 3—4 слабых колючих луча, во втором 9—11 лучей. Тело и голова сильно приплюснуты. Нет оперкулярного клапана, шишкообразного выроста верхней челюсти и продольной кожистой складки ниже боковой линии. Жаберное отверстие расположено на спинной стороне головы. На нижней стороне костного выроста предкрышечной кости имеется шип, направленный острием вперед; на верхней стороне выроста имеются мелкая зазубренность или крупные изогнутые шипы, конец шипа заострен или крючкообразно загнут. Мягкие лучи второго спинного плавника простые, кроме самого последнего, ветвистого. Последний луч брюшного плавника соединен перепонкой с основанием грудного плавника.
Много видов. В водах Японии 16, из них в Японском море 10 видов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА CALLIONYMUS
(Ochiai et al., 1955, с дополнениями)
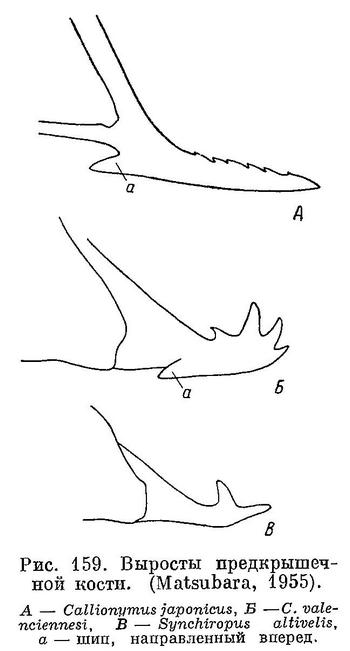
1 (4) . Предкрышечный костный вырост саблеобразной формы, прямой, с заостренной верхушкой; верхний край имеет мелкую зазубренность (рис. 159, А).

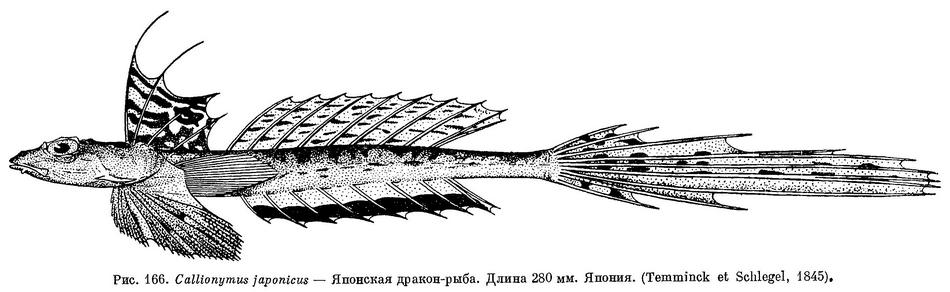
2 (3). На верхней поверхности головы (рис. 163) позади глаз 2 костные площадки с радиально расходящимися бугорками; промежуток между площадками покрыт гладкой кожей. (у Callionymus variegatus Temminck et Schlegel из Южной Японии эти площадки слиты в одну.) Во втором спинном плавнике 9, в анальном — 8 мягких лучей (последние лучи в обоих плавниках раздвоены). Первый спинной плавник с черным пятном между 3-м и 4-м лучами; у самцов первые 2 луча этого плавника удлинены в виде нити (см. рис. 166). На груди у самцов бывает темное пятно. 1. Callionymus japonicus Houttuyn.
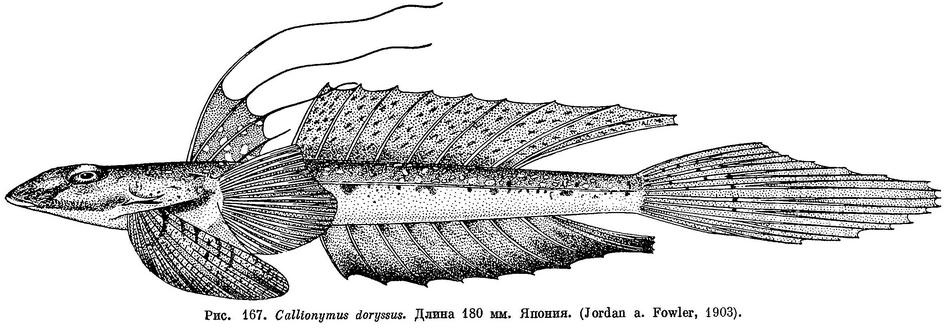
3 (2). Верхняя поверхность головы гладкая или с очень слабо выраженными двумя соприкасающимися площадками бугорков, различимыми у подсушенного экземпляра при значительном увеличении. Во втором спинном и в анальном плавниках по 9 лучей. Первый спинной плавник без черного пятна: лучи первого спинного плавника у взрослых рыб удлинены все (см. рис. 167). Нижняя сторона головы у самцов светлая. 2. Callionymus doryssus (Jordan et Fowler).
4 (1). Предкрышечный костный вырост на ( верхушке изогнут; верхний край вооружен 3 или более крупными изогнутыми шипами (см. рис. 159, Б).
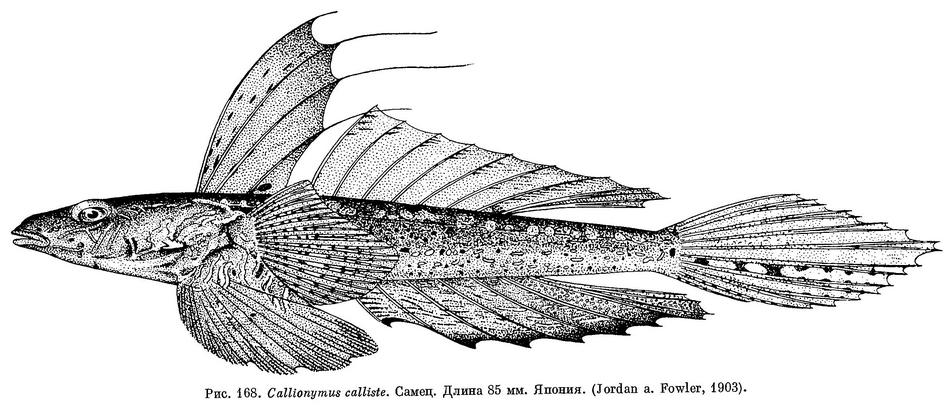
5 (6). Во втором спинном плавнике 8, в анальном — 7 мягких лучей (последний луч у обоих плавников раздвоен). Нет дополнительной веточки на спинной стороне хвостового стебля, связывающей боковые линии противоположных сторон тела. Глаз относительно умеренный, около 25% длины головы. (Близкий вид Callionymus phasis Günther, известный у берегов преф. Эхиме, отличается более крупным глазом — 33% длины головы.) Самцы в отличие от самок имеют нитевидно удлиненные лучи в первом спинном плавнике (см. рис. 168). 3. Callionymus calliste Jordan et Fowler.
6 (5). Во втором спинном и анальном плавниках по 9 мягких лучей (Последний луч у обоих плавников раздвоен). Имеется дополнительная веточка на спинной стороне хвостового стебля, связывающая боковые линии противоположных сторон тела (рис. 164). Этот признак не изображен на большинстве рисунков.
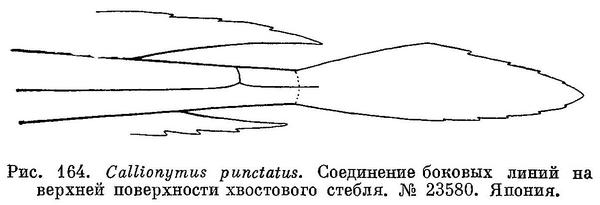
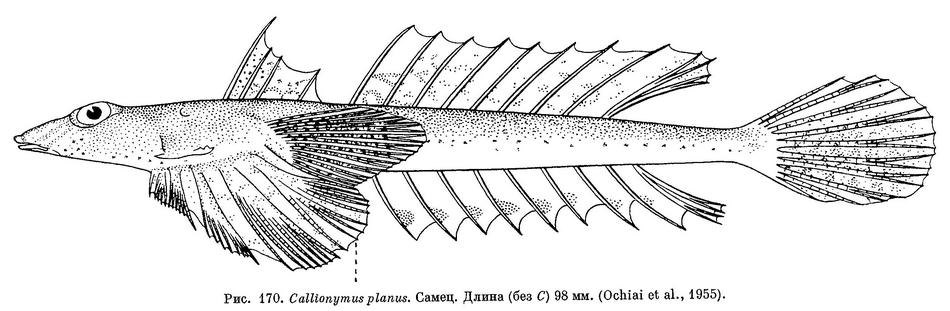
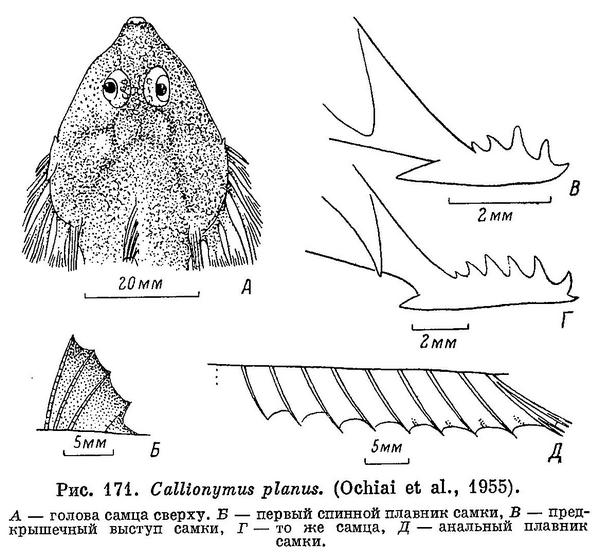
7 (8). Предкрышечный костный вырост довольно толстый, с 5—6 острыми шипами по верхнему краю. Голова уплощена, ее ширина больше длины. Первый спинной плавник у самцов с темным рисунком из многочисленных, густо расположенных извитых линий и точек (см. рис. 170), а у самок с мелкими темными пятнышками (см. рис. 171, Б). 4. Callionymus planus Ochiai.
8 (7). Предкрышечный костный вырост довольно тонкий, с 3—4 острыми шипами по верхнему краю. Голова умеренно уплощена, такой же ширины и даже меньше ее длины.
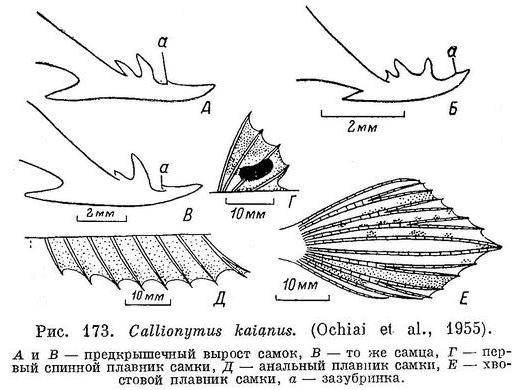
9 (10). Самый задний шип предкрышечного костного выроста с зазубринкой (см. рис. 173, а). У самца 2 центральных луча хвостового плавника заметно удлинены. 5. Callionymus kaianus Günther.
10 (9). Самый задний шип предкрышечного костного выроста без зазубринки. Лучи хвостового плавника не удлинены, а если удлинены, то почти все, а не только 2 центральных луча.
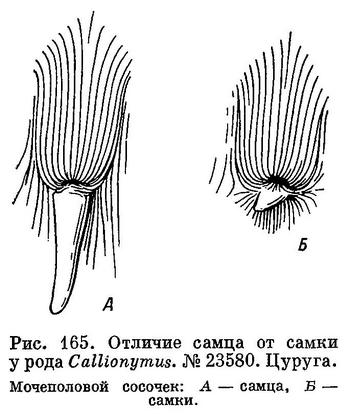
11 (22). Мочеполовой сосочек удлинен (самцы) (рис. 165, А).
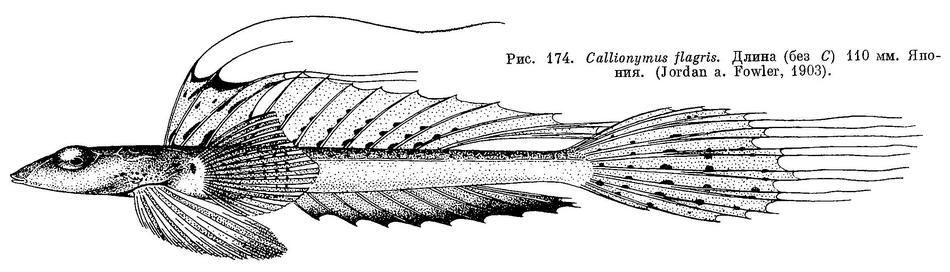
12 (13). Лучи хвостового плавника (5 средних) нитевидно удлинены. Лучи первого спинного плавника (кроме второго луча) сильно нитевидно удлинены (см. рис. 174). 6. Callionymus flagris Jordan et Fowler.
13 (12). Лучи хвостового плавника нитевидно не удлинены или только слегка удлинены.
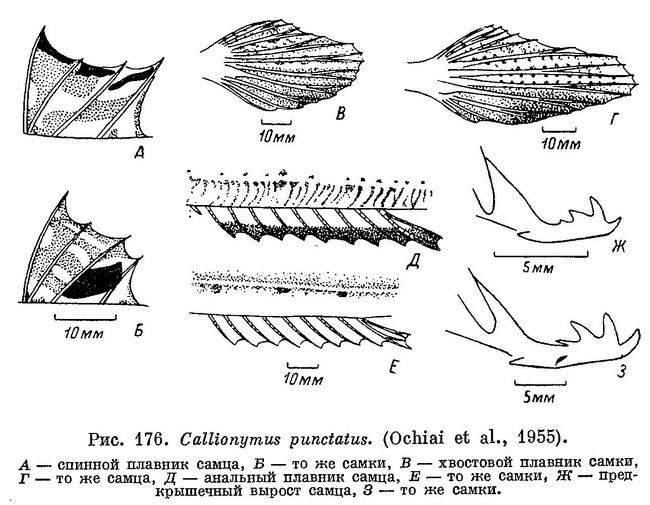
14 (15). Лучи первого спинного плавника не удлинены. У самцов верхний край перепонок первого спинного плавника с характерной окраской в виде узкой черной полоски из полулунных пятен (см. рис. 176, А), а у самок с овальным черным пятном на перепонке этого плавника
между III и IV лучами (см. рис. 176, Б). 7. Callionymus punctatus Richardson.
15 (14). Все или некоторые лучи первого спинного плавника нитевидно удлинены.
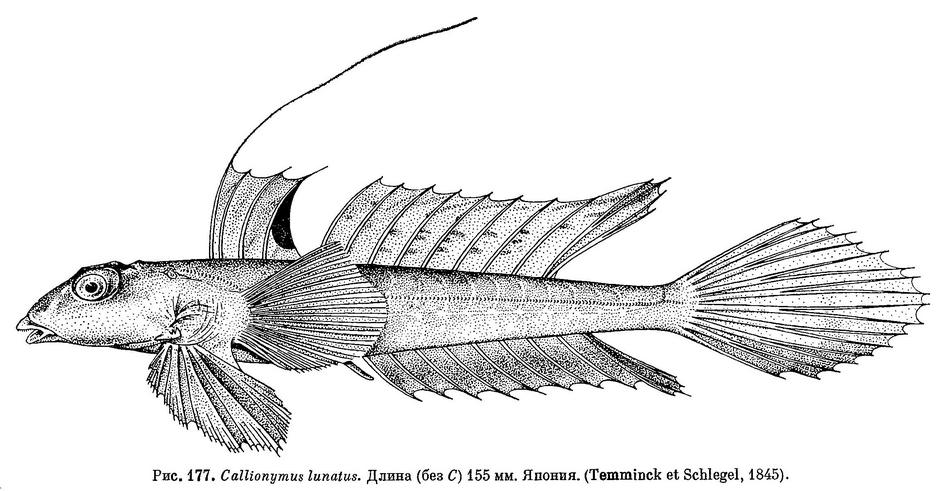
16 (17). Только первый луч первого спинного плавника нитевидно удлинен; остальные лучи не имеют резко выраженного удлинения (см. рис. 177 и 178, А); имеется черноватое пятно на перепонке позади последнего луча. 8. Callionymus lunatus Temminck et Schlegel.
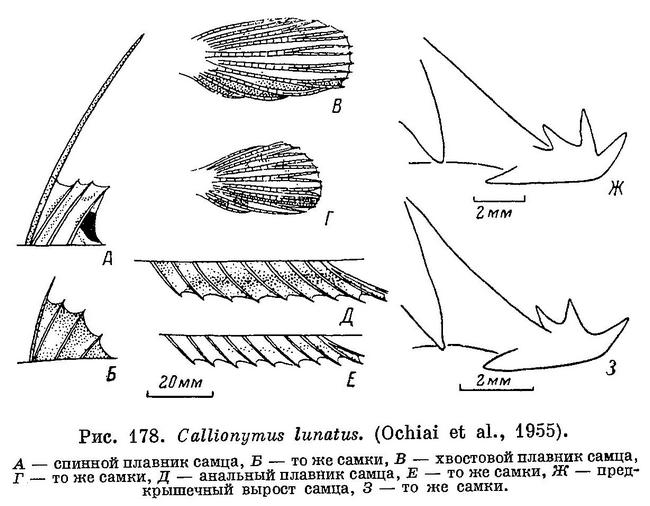
17 (16). Все лучи первого спинного плавника более или менее нитевидно удлинены.
18 (21). Передние 2 луча первого спинного плавника длиннее остальных. Перепонка между лучами первого спинного плавника такой же высоты или ниже перепонки между лучами второго спинного. Диаметр глаза равен или немного меньше длины рыла.
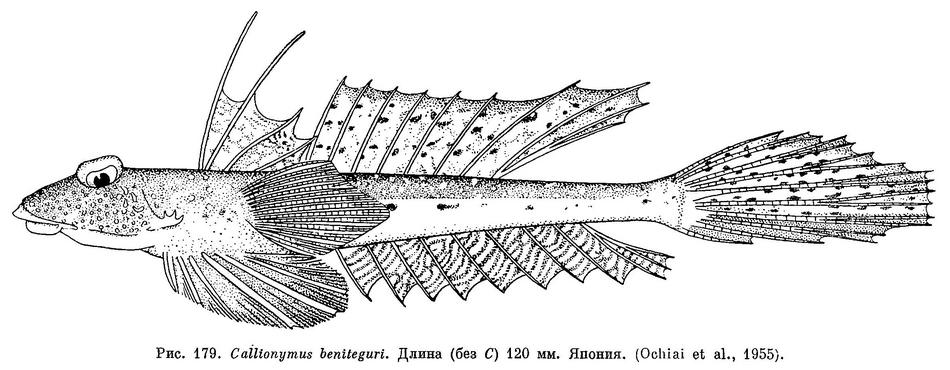
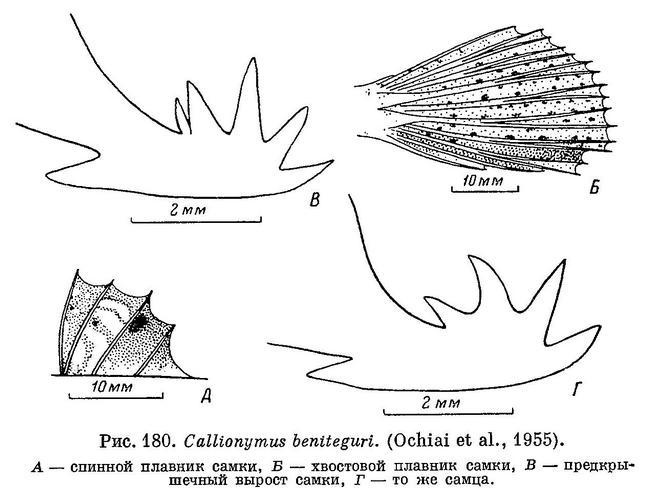
19 (20). Три средних луча хвостового плавника длиннее других (см. рис. 179). Задние лучи спинного и анального плавников простираются до основания хвостового и даже слегка дальше. Нет большого черного пятна на первом спинном плавнике; анальный плавник тусклый или черный с многочисленными косыми волнистыми белыми полосками. 9. Callionymus beniteguri Jordan et Snyder.
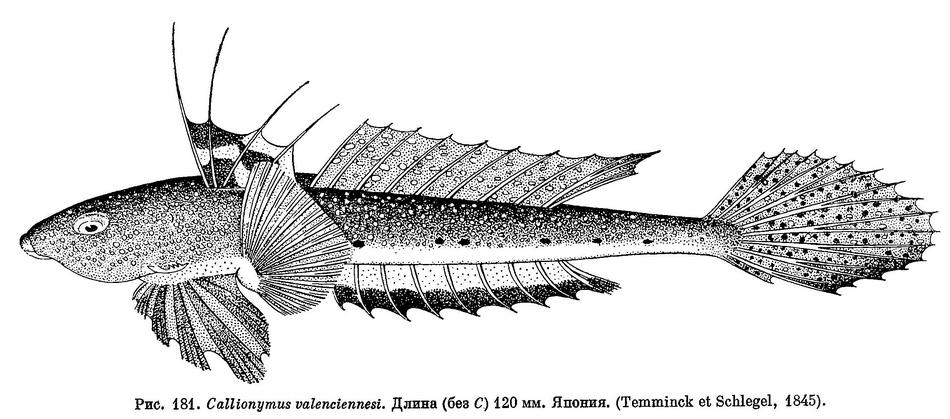
20 (19). Хвостовой плавник равномерно закругленный (см. рис. 181). Задние лучи спинного и анального плавников не достигают основания хвостового. Имеется несколько черноватых пятен на первом спинном плавнике; анальный плавник светлый, но наружный край черный. 10. Callionymus valenciennesi Temminck el Sclilegel.
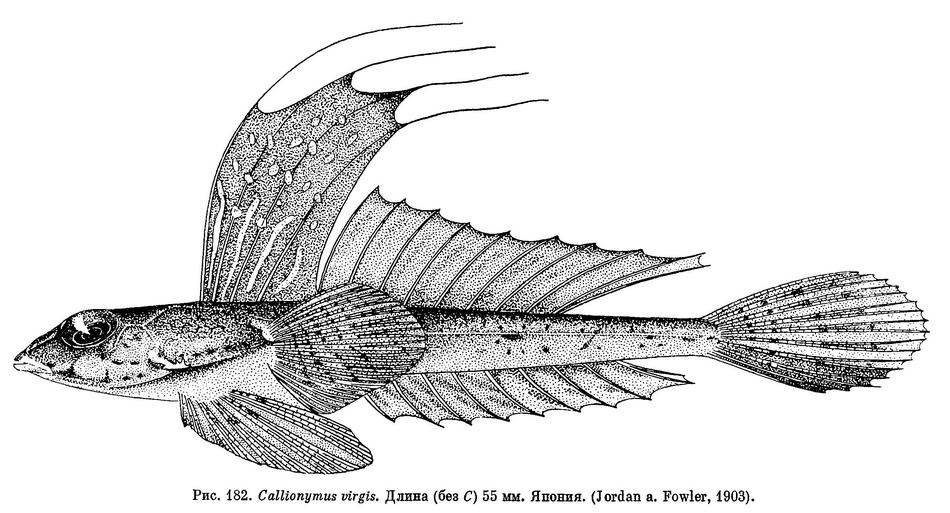
21 (18). Все лучи первого спинного плавника очень сильно удлинены (см. рис. 182); перепонка между лучами почти вдвое выше перепонки между лучами второго спинного плавника; перепонка последнего луча слегка прикреплена сзади к основанию первого луча второго
спинного плавника. Диаметр глаза больше длины рыла. 11. Callionymus virgis Jordan et Fowler.
22 (11). Мочеполовой сосочек очень короткий или рудиментарен (самки) (см. рис. 165, Б).
23 (30). Последние лучи спинного и анального плавников, прижатые к телу, не достигают основания хвостового плавника.
24 (25). Лучи первого спинного плавника более или менее нитевидно удлинены, причем первый луч, самый длинный и прижатый к телу, достигает основания второго спинного (см. рис. 175). Анальный плавник
бледный. 6. Callionymus flagris Jordan et Fowler.
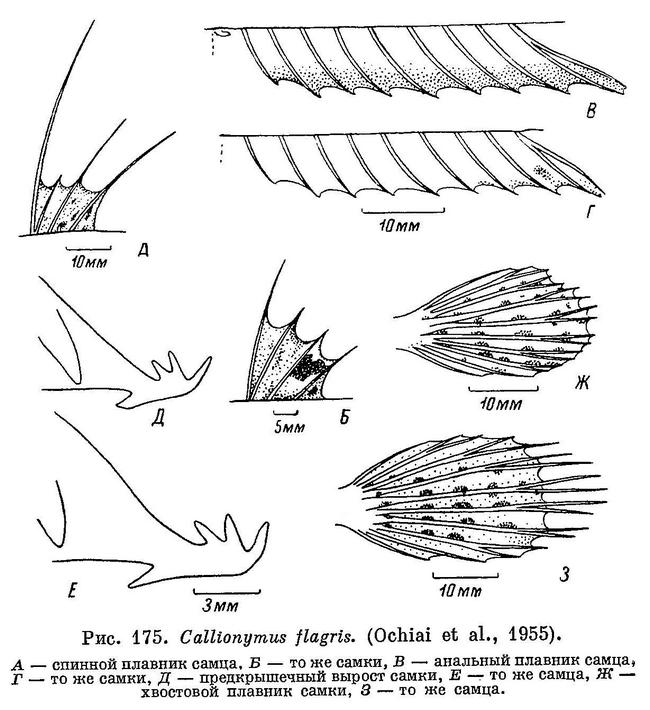
25 (24). Лучи первого спинного плавника не удлинены, за исключением первого луча, который не достигает или достигает с большим трудом основания первого луча второго спинного плавника.
26 (29). Первый спинной плавник однообразно темный (см. рис. 178, Б).
27 (28). Глаз умеренной величины; его диаметр примерно равен длине рыла. Вдоль середины анального плавника проходит слабая темная полоска (см. рис. 178, Е). 8. Callionymus lunatus Temminck et Schlegel.
28 (27). Глаз довольно большой; его диаметр больше длины рыла. Анальный плавник без темной полоски. 11. Callionymus virgis Jordan et Fowler.
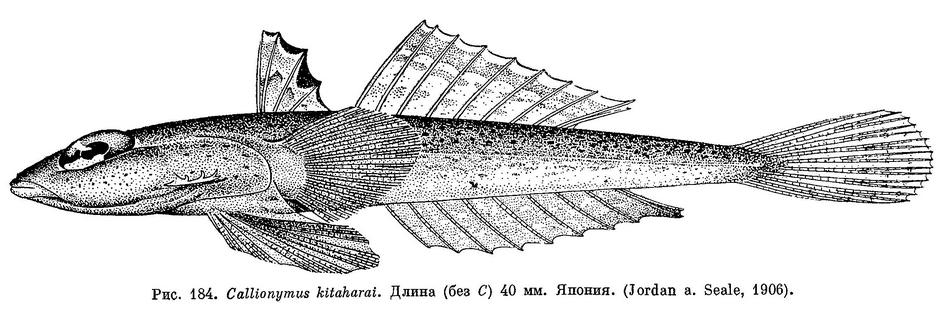
29 (26). Первый спинной плавник с темным пятном полулунной формы на перепонке между вторым и третьим лучом; другое — сходное, но более крупное пятно, на перепонке между третьим и четвертым лучом (см. рис. 184). 12. [Callionymus kitaharai Jordan et Seale].
30 (23). Последние лучи спинного и анального плавников у взрослых, прижатые к телу, простираются до основания хвостового плавника. Анальный плавник тусклый. У просмотренных нами самок длиной 37 и 74 мм лучи спинного, а тем более анального плавников не достигали основания хвостового, хотя по характерному пятну на первом спинном плавнике и другими признакам эти экземпляры несомненно принадлежат к Callionymus punctatus.
31 (32). Первый спинной плавник с многочисленными, очень мелкими темными крапинками и крупными белыми пятнами на перепонках между первым и третьим лучами (см. рис. 180, А), остальная часть плавника однообразно темная. 9. Callionymus beniteguri Jordan et Snyder.
32 (31). Первый спинной плавник с большим черноватым пятном на перепонке между третьим и четвертым лучами (см. рис. 176, Б). 7. Callionymus punctatus Richardson.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
МАЛАЯ МОРСКАЯ МЫШЬ — CALLIONYMUS BELENUS RISSO
МОРСКАЯ МЫШЬ — CALLIONYMUS FESTIVUS PALLAS
Условия размножения. Нерест обоих видов в Черном море происходит летом. Впервые В. А. Водяницкий (1930в) отметил нахождение икринок Callionymus festivus в Новороссийской бухте в середине мая. Дальнейшими наблюдениями Е. Г. Косякиной (1938) было показано, что икринки Callionymus встречаются в этом районе с середины мая до середины сентября. В районе Карадага икринки Callionymus festivus ловились с июня по сентябрь (Овен, 1959). У побережья Болгарии нерест этого вида происходит с мая по август; икринки встречаются в планктоне с начала июня до конца августа при температуре воды 16—25°.
По наблюдениям Ю. П. Зайцева (1959а), нерест Callionymus belenus происходит с конца мая до середины сентября. Икра встречается в толще воды при температуре 10,0—24,5° и солености 9,51—18,30‰. Наиболее интенсивный нерест наблюдается в июле. У побережья Болгарии Callionymus belenus размножается с июня до начала сентября. Икринки встречаются в планктоне при температуре воды 20—25°.
По нашим наблюдениям, икринки Callionymus sp. встречаются преимущественно в узкой прибрежной зоне до 1—1,5 миль от берега с июня по август при температуре воды 19,2—23,6° и солености 17—18‰. Личинки рассеиваются на более широком пространстве моря и в значительном количестве встречались в уловах до 7,5—10 миль от берега.
В Средиземном море Callionymus festivus размножается с мая по август (Raffaele, 1888; Lo Bianco, 1908—1909). Икринки Callionymus belenus ловились в районе Марселя в апреле (Holt, 1899). Личинки и мальки этого вида вылавливались с июня по сентябрь (Fage, 1918). На основании этих данных Падоа (1956д) считает, что этот вид размножается в Средиземном море весной и летом. По наблюдениям Фажа (1918), личинки Callionymus belenus обитают в поверхностных слоях у берегов: ни одной личинки не было поймано при длине ваера более 95 м. Об условиях нахождения личинок Callionymus festivus в Средиземном море данных нет.
Строение икринок. Икринкн Callionymus festivus и Callionymus belenus сферические, очень мелкие, без жировой капли. Оболочка икринок тонкая, совершенно прозрачная. Поверхностный слой желтка разделен на мелкие дольки. Перивителлиновое пространство очень узкое (рис. 47). Диаметр икринок Callionymus festivus, по данным В. А. Водяницкого и И. И. Казановой (1954), 0,62—0,75 мм. Длина только что выклюнувшихся личинок около 1,5 мм. Соответствующие размеры икринок этого вида приводят Ж. М. Георгиев и др. (1960).
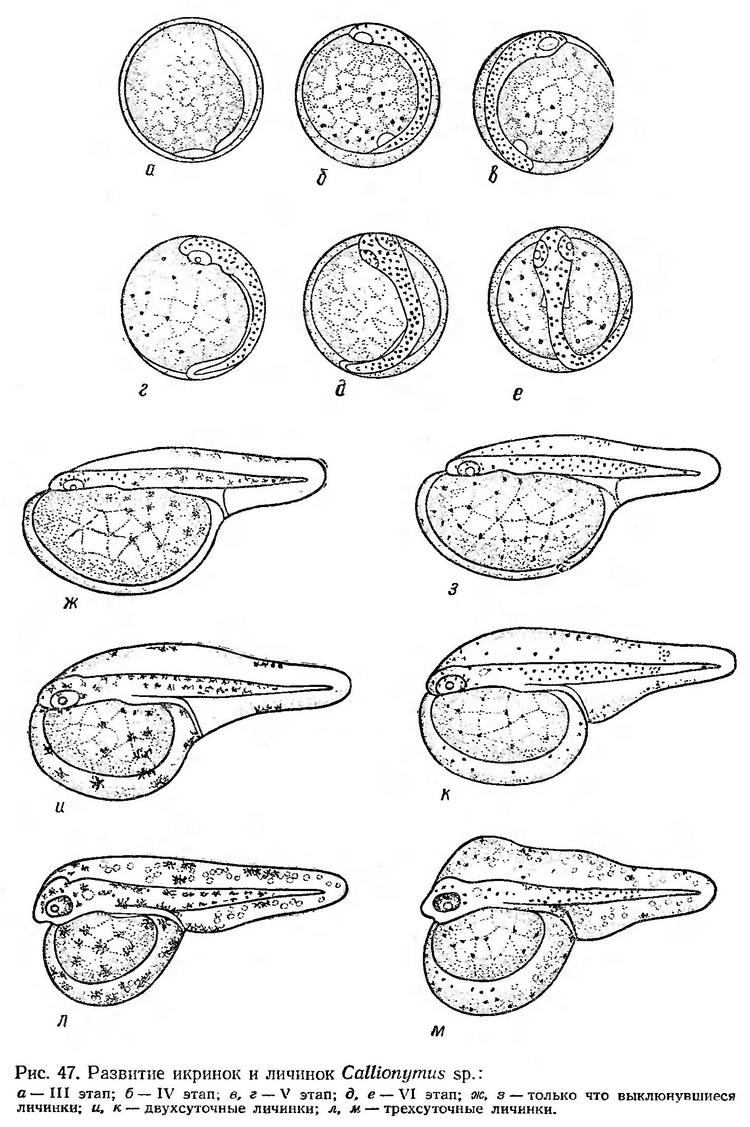
Диаметр живых икринок Callionymus belenus, по данным Ю. П. Зайцева (1959а), 0,65—0,81 мм. Средний размер уменьшается от июня к августу от 0,72 до 0,70 мм. Зародыш и желточный мешок пигментированы звездчатыми клетками, которые в проходящем свете имеют темно-коричневую окраску, а в отраженном — лимонно-желтую. Выклюнувшиеся личинки очень мелкие. Их длина 1,02—1,32 мм (Зайцев, 1959а). Икринки Callionymus belenus у болгарского побережья несколько мельче — 0,61—0,70 мм (Георгиев и др., 1960).
В Средиземном море икринки этих видов значительно мельче. Так, Холт (1899) для икринок Callionymus belenus указывает размеры 0,58—0,64 мм и Рафаеле (1888) для икринок Callionymus festivus — 0,56—0,60 мм.
Эмбриональное развитие Callionymus sp. прослежено с конца III этапа на икринках, выловленных в море в ранние утренние часы (5—6 ч). Количество икринок в наших сборах, как будет показано ниже, невелико. На основании имеющихся материалов было трудно проследить суточный ритм размножения и развития икринок. Недавно выметанные икринки в наших сборах были отмечены в промежуток времени между 17 и 22 ч.
Ко времени замыкания бластопора зародышевая полоска имеет четкие очертания. Головной конец заметно расширен, намечаются глазные пузыри. В туловищном отделе появляются 1—2 первых сегмента (рис. 47, а). В момент замыкания бластопора появляется купферов пузырек, обособляются глаза, формируются мозговые доли, число туловищных сегментов увеличивается до 6—7. На теле эмбриона и на желтке появляются мелкие точечные хроматофоры, которые в проходящем свете имеют темно-коричневую окраску, в отраженном — желтую (рис. 47, б). IV этап при температуре воды 21,4° продолжается 2—2,5 ч.
Ко времени образования хвостовой почки в туловищном отделе насчитывается около 20 сегментов. На V этапе развития в глазах появляются хрусталики, образуются слуховые капсулы, намечается плавниковая складка. Пигментация тела усиливается (рис. 47, в, г). Когда хвост эмбриона несколько заходит за середину желтка, начинаются слабые подергивания эмбриона. Зачаток сердца очень маленький, и пульсация его в это время не прослеживается. V этап, который заканчивается с появлением подвижности эмбриона, продолжается при температуре воды 25,0° около 2 ч. При дальнейшем развитии эмбриона уменьшается желток и увеличивается соответственно перивителлиновое пространство. На VI этапе появляются зачатки грудных плавников, дифференцируются мозговые доли, наряду с увеличением хвостового отдела образуются хвостовые миотомы. Значительно усиливается пигментация тела и желточного мешка (рис. 47, д, е). Перед выклевом эмбрион охватывает около 2/3 поверхности желтка. Прослеживается очень слабая пульсация сердца. Эмбрион часто и энергично подергивается. Голова прижимается к оболочке, выпячивает ее, и в этом месте оболочка разрывается. Выклев происходит в течение нескольких минут — через 3 ч с момента обнаружения подвижности эмбриона.
Весь процесс эмбрионального развития, исходя из времени вымета икринок (17—22 ч), при температуре воды 21,4—25,0° продолжается около суток.
Постэмбриональное развитие. Только что выклюнувшиеся личинки Callionymus sp., по нашим данным, имеют длину 1,25—1,30 мм. Они очень слабо развиты. Голова плотно прижата к большому яйцевидному желточному мешку. Грудные плавники зачаточные. Кишечник не сформирован. Сердце пульсирует очень слабо. Широкая плавниковая оторочка окаймляет тело. Хвостовой отдел короткий. Антеанальное расстояние составляет 67—68% длины тела. По характеру пигментации выклюнувшихся личинок можно разделить на две группы. У личинок первой группы (рис. 47, ж) на спинной стороне и на боковых поверхностях тела расположены ветвистые темно-коричневые (в проходящем свете) хроматофоры. В средней части хвостового отдела тела и по краям плавниковой каймы в этой же области находятся скопления крупных хроматофоров. Такое же скопление расположено по краю спинной плавниковой каймы над задней частью туловищного отдела. На хвостовом стебле пигмента нет. Крупные ветвистые хроматофоры расположены отдельными небольшими скоплениями на желточном мешке. Тело личинок окрашено в бледно-желтый цвет.
Личинки второй группы (рис. 47, з) имеют такую же бледно-желтую фоновую окраску. На теле, на плавниковой кайме и на желточном мешке более или менее равномерно распределены темно-коричневые компактные (слабо разветвленные) хроматофоры. Отдельных скоплений пигмента, как у личинок первой группы, нет. Одна или две пигментные клеточки имеются в области хвостового стебля.
Личинки парят в воде у самой поверхности в перевернутом горизонтальном положении. Большой желточный мешок при очень малых размерах тела сильно затрудняет движение. Личинки в основном вращаются вокруг оси тела при помощи вибрирующих изгибов хвоста, время от времени штопорообразно поднимаясь вверх, и быстро переходят в состояние покоя.
Уже к концу первых суток развития личинки приобретают способность к горизонтальным перемещениям; при этом они поворачиваются на бок и в таком положении проплывают расстояние в 2—3 мм и затем снова переходят в состояние покоя. В минуту насчитывается 8—11 таких перемещений.
К двухсуточному возрасту длина личинок увеличивается до 1,4—1,6 мм. Желточный мешок заметно уменьшается, голова частично освобождается от него и сразу не намечается ротовая ямка. Хвостовой отдел удлиняется; антеанальное расстояние уменьшается до 55—57% длины тела. Зачатки грудных плавников увеличиваются; в глазах появляется пигмент. Формируется кишечник. Сердце пульсирует четко, ритмично. В спинной плавниковой кайме образуется полость, которая по мере рассасывания желтка заполняется водой. Различия в характере пигментации сохраняются (рис. 47, и, к). Личинки опускаются в придонный слой аквариума. В периоды покоя они держатся в горизонтальном положении брюшной стороной кверху. При движении принимают боковое положение. Время от времени личинки поднимаются вертикально вверх и тотчас же снова опускаются на дно.
На третьи сутки после выклева грудные плавники приобретают подвижность, благодаря чему личинки при плавании делают повороты, меняют направление движения. Но движения личинок по-прежнему ограниченные, кратковременные. К этому времени передняя часть головы освобождается от желточного мешка, намечается ротовая ямка, глаза интенсивно пигментируются, усиливается пигментация тела (рис. 47, л, м). Желточный мешок несколько уменьшается, но он еще достаточно большой, в связи с чем личинки по-прежнему плавают сравнительно медленно, в основном по кругу, К концу третьих суток личинки в наших опытах погибли. Дальнейшее описание дано по фиксированному материалу.
У личинок длиной 1,8 мм (рис. 48, а) сохраняется небольшой остаток желточного мешка. Рот прорезался, но челюсти еще не оформлены. Усиливается пигментация глаз; грудные плавники увеличиваются. Кишечник изгибается и утолщается. Плавательный пузырь заполняется воздухом. Питание эндогенное.
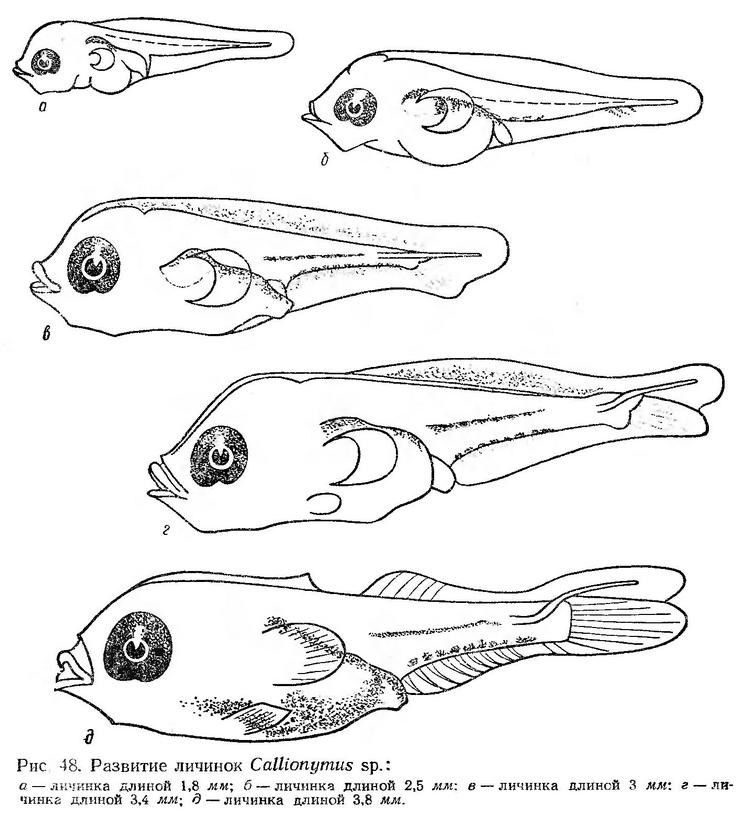
По мере дальнейшего развития голова и туловище личинок становятся массивными. Личинки длиной 2,5 мм имеют широкий петлеобразно изогнутый кишечник, большую массивную голову, широкое туловище и сравнительно короткий постанальный отдел (около 45% длины тела). У личинок оформляются челюсти, жаберный аппарат. Большие веерообразные грудные плавники хорошо развиты. В хвостовом плавнике намечаются лучи (рис. 48, б). Личинки наряду с использованием имеющегося еще желтка переходят на экзогенное питание. В кишечниках встречается растительная и животная пища: жгутиковые, диатомовые водоросли, инфузории, науплиусы и метанауплиусы Copepoda, личинки пластинчатожаберных моллюсков (Дука, 1966). Личинки длиной 3 мм еще имеют небольшой остаток желточного мешка, но в основном уже перешли на внешнее питание. В кишечниках встречаются науплиусы и метанауплиусы Copepoda, взрослые формы Oithona minuta, Podon polyphemoides, Tintinnidae. В хвостовом плавнике начинают формироваться лучи (рис. 48, в).
У личинок длиной 3,4—3,5 мм голова и туловище становятся еще более массивными (рис. 48, г). Туловищный отдел удлиняется; антеанальное расстояние увеличивается до 60—62% длины тела. Высота тела за анусом составляет 15—16% длины тела. Челюсти и жаберный аппарат оформлены. Глаза большие, интенсивно пигментированы. Формируется хвостовой плавник; длинный, тонкий уростиль загибается вверх. Закладываются лучи в анальном плавнике. Личинки переходят на питание крупными планктонными организмами. Наряду с Oithona minuta в кишечниках встречаются Acartia clausi, Pseudocalanus elongatus, Centropages kröyeri, а также личинки моллюсков и науплиусы баллянусов (Дука, 1966).
При длине 3,8—4,0 мм закладывается колючий спинной плавник, формируются мягкий спинной и анальный плавники. Рот становится выдвижным (рис. 48, д). Антеанальное расстояние увеличивается до 65—66% длины тела.
По наблюдениям Л. А. Дука (1966), личинки Callionymus sp. размером 3,5—6,0 мм большую часть времени держатся у дна аквариума, облавливая в этом слое воды планктон. Изредка личинки поднимаются в верхние слои, где обычно концентрируются кормовые организмы, и активно охотятся за ними. Автором были выявлены интересные особенности в питании личинок, которые позволяют отнести их по типу питания к «засадчикам». Увидев движущийся организм, личинка настораживается и, поворачивая глаза или голову, следит за движением жертвы. Когда организм приближается на расстояние примерно 1 см, она производит сильный бросок и схватывает добычу. По данным Л. А. Дука, личинки Callionymus sp. — эврифаги: в пищевом комке личинок насчитывается до 16 кормовых организмов. Излюбленные организмы — яйценосные самки Oithona minuta — составляют 35—77% по весу и 82—91 % по количеству от состава пищевого комка. О большом значении Oithona minuta в питании Callionymus sp. свидетельствуют также высокие индексы избирательной способности по этой форме. Большое значение в питании наряду с Oithona minuta имеют Podon polyphemoides, Acartia clausi и Paracalanus parvus.
Количество этих организмов в пищевом комке личинок в разные годы различно, что в большой степени зависит от количественных соотношений этих форм в планктоне (Дука, 1966).
Распределение и численность икринок и личинок. Икра Callionymus sp. встречается в планктоне летом у берегов Крыма, Кавказа, в северо-западной части моря, в частности в Одесском и Каркинитском заливах, у побережья Болгарии (Водяницкий, 1930 в; Косякина, 1938; Дехник и Павловская, 1950; Павловская, 1950; Дука, 1959; Зайцев, 1959а; Овен, 1959; Георгиев и др., 1960). В уловах обычно отмечены единичные икринки. В опресненном Одесском заливе икринки Callionymus belenus благодаря высокой плавучести занимают второе место по численности после икринок хамсы. В отдельные периоды наблюдений численность икринок достигала 6—7 экз. под 10 м2 поверхности моря. В июне 1952 г. было отмечено максимальное количество — 107 икринок под 10 м2 (Зайцев, 1959а).
В наших материалах из района Севастополя во все годы наблюдений отмечены единичные икринки Callionymus sp. Они вылавливались в основном из поверхностного слоя в количестве не более 11 экз. в одном 10-минутном горизонтальном лове; обычно в улове насчитывалось 1—2 икринки.
В сборах ихтиопланктона вблизи берегов в районе Карадага и Батуми в конце июня — начале июля количество икринок Callionymus sp. было несколько больше, в среднем 5—8 экз. в одном горизонтальном лове.
Пелагические личинки и мальки Callionymus sp. длиной 2,7—6,4 мм, по данным З. М. Пчелиной (1940), ловились в Новороссийской бухте и перед входом в нее с июня по сентябрь. Автор отмечает, что «днем они чаще встречались в промежуточном слое, а ночью нередко также у поверхности и у берегов» (Пчелина, 1940, стр. 64). Наблюдения Ю. П. Зайцева (1959а) показали, что личинки Callionymus belenus в природе и в эксперименте обитают преимущественно близ самой поверхности воды. Наряду с этим Л. А. Дука (1966) приводит данные, свидетельствующие о том, что личинки Callionymus sp. в светлое время суток держатся в слое 5—15 м, а ночью поднимаются к поверхности.
Наши экспериментальные наблюдения показали, что у поверхности держатся личинки недавно выклюнувшиеся; достигнув возраста 2—3 суток, они опускаются на дно аквариума.
На основании круглосуточных наблюдений, проведенных в море, также выявляется, что личинки Callionymus sp. обитают в промежуточном слое, а не у поверхности. По горизонтам лова они распределяются следующим образом. На 0 м выловлено 46 личинок (22 лова), на 10 м — 229 (38 ловов), в слое 25—0 м — 141 (49 ловов).
В уловах встречались личинки размером от 1,4 до 5,5 мм, преобладали личинки размером 2—3 мм.
Интересно отметить, что в наших сборах ихтиопланктона в разные годы наблюдений личинки Callionymus sp. встречались значительно чаще и в большем количестве, чем икринки.
В отдельных случаях количество личинок достигало 50—70 экз. под 1 м2; икринки встречались обычно единичными экземплярами.
Значительные различия в численности икры и личинок Callionymus sp. в планктоне определяются следующими причинами:
1. Вымет и развитие икринок происходит в узкой прибрежной зоне не далее, по-видимому, 0,5—1 мили от берега, где сборы ихтиопланктона производились очень редко.
2. Эмбриональный период развития Callionymus sp. продолжается около суток. В течение этого времени икринки не могут быть вынесены далеко от берега. Вымет икринок происходит в определенное время суток — в сумеречные часы; в другие часы суток икринки в планктон не поступают. Планктонный период жизни личинок Cailionymus sp. заканчивается при длине 6—7 мм в возрасте около двух недель. За это время личинки могут быть вынесены течениями в открытое море или активно расселиться в поисках скоплений пищевых организмов на значительной площади в верхних теплых слоях моря.