Trachurus capensis Castelnau, 1861
Капская ставрида, южноафриканская ставрида (Rus),
Cape horse mackerel
(Eng)
Синонимы:
Selar tabulae Barnard, 1927
Trachurus capensis Castelnau, 1861
Trachurus trachurus capensis Aleev, 1957
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus capensis Castelnau - капская, или южноафриканская, ставрида
Описание. D I,VIII, I+28-36, наиболее часто 32-33; А II, I+25-31, наиболее часто 28; L.l. 68-80, наиболее часто 73. Спинные туловищные сейсмосенсорные каналы длинные: 18DII-27DII. Жаберных тычинок на первой дуге 62-76, на ее нижней части - 49-56. Щитки боковой линии крупные; колючих щитков 36-43. Первый колючий щиток лежит под основанием 4-7-го луча второго спинного плавника. Отношение задней части боковой линии к хорде дуги передней ее части 1,09. Отношение расстояний между основаниями последнего и предпоследнего лучей и между основаниями предпоследнего и предшествующего ему луча в А составляет 1,1-1,4. Высота наиболее высоких щитков в передней части боковой линии составляет 15,0-21,0 % длины головы. Высота наиболее высоких щитков в задней части боковой линии - 17,5-23,5% длины головы. Наибольшая высота тела 19,5-23,3% его длины. Длина головы 25,4-29,0%, наиболее часто 27,0-28,0% длины тела. Рыло коническое. Глаз снабжен жировым веком, его диаметр 20,2-28,2% длины головы. (Длина грудных плавников 23,1-29,1% длины тела. Рот косой, верхняя челюсть широкая. Ноздри смежные, заметно удалены от переднего края глаза. Расстояние от вершины рыла до вертикали начала основания брюшных плавников 28,9-32,5% длины тела, наиболее часто 31,0. Расстояние от вершины рыла до вертикали начала основания первого спинного плавника 30,9-34,9% длины тела, наиболее часто 32,9%. Расстояние от вершины рыла до вертикали основания первого луча анального плавника 50,6-58,6% длины тела, наиболее часто 54,7%.
Тело толстое, сжатое с боков. Длина головы больше, чем ее высота. Разрез рта на уровне центра глаза, косой. Нижняя челюсть выступает вперед; верхняя доходит до вертикали переднего края глаза. Зубы очень мелкие конические, на каждой челюсти, на языке, небе и сошнике. Щеки, затылочное пространство и часть жаберной крышки покрыты чешуей, остальные части головы голые. Мягкий спинной и анальный плавники с серповидными выемками в области 4-5-го и 3-4-го ветвистых лучей соответственно. Грудные плавники серповидные, оканчиваются на вертикали начала изгиба боковой линии. Брюшные плавники короче грудных, веерообразные.
Спина и верхняя часть головы темные, брюшко и бока серебристые, грудные и спинные плавники серые, хвостовой более светлый, желтоватый, брюшные и анальный плавники белые. Конец рыла и основания грудных плавников черные. На жаберной крышке черное пятно [Berry, Cohen, 1972; наши данные].
Распространение. Trachurus capensis в Атлантическом океане обитает у берегов Африки, от Анголы до мыса Доброй Надежды и на отмели Агульяс в Индийском океане [Алеев, 1957а; Световидов, 1964; Комаров, 1969]. По другим данным [Berry, Cohen, 1972], северной границей ее распространения служит 4° ю.ш.; известны также сообщения о том, что Trachurus capensis распространена вдоль побережья юго-восточной Африки до бухты Делагоа [Smith, 1961; Rass, 1973]. По нашим наблюдениям, от порта Дурбан до бухты Делагоа обитает только Trachurus delagoa [Некрасов, 1970].
Ю. Г.Алеев [1957а] рассматривал капскую ставриду в качестве подвида Trachurus trachurus capensis, который отличается от Trachurus trachurus trachurus несколько большим числом щитков в боковой линии (75) и их меньшей величиной.
Валидность вида Trachurus capensis восстановлена Ф.Берри и Л.Коеном [1972].
Биология. По-видимому, в пределах ареала капской ставриды обитают два ее стада: северное - в водах Атлантики от Анголы до ЮАР и южное - на отмели Агульяс.
Капская ставрида в водах Атлантики достигает длины 75 см. Половозрелой она становится при длине 22-26 см в возрасте 2-3 лет. У юго-западного побережья Африки она встречается на шельфе до глубины 300 м вдоль всего побережья от Анголы до мыса Доброй Надежды.
Промысловые концентрации ставриды обнаружены на глубинах от 60 до 270 м. Чаще всего наиболее мощные скопления встречаются в придонном слое между изобатами 120 и 180 м на стыке вод холодного Бенгельского течения и встречного течения южного направления [Комаров, 1962]. В зоне перемешивания создаются условия, наиболее благоприятные для нагула и нереста ставриды. Эта зона служит постоянным местом обитания рыб различных возрастов.
Промысловые скопления ставриды в районе 17-25° ю.ш. в конце 1960 - начале 1961 гг. состояли из рыб различных возрастных групп размером от 14 до 45 см. Молодые особи придерживались более теплых вод, поэтому они чаще встречались на северной кромке скоплений и на малых глубинах. Рыбы старших возрастов тяготеют к большим глубинам с более низкой температурой воды. В южной части ареала (в районе Уолфиш-Бей) встречаются лишь половозрелые рыбы.
В южной Атлантике отмечена зависимость поведения ставриды от режима ветров. Сильные южные ветры усиливают Бенгельское течение, вследствие чего воды, обедненные кислородом, вторгаются в зону смешанных вод, захватывая ее южную и прибрежную части. При таких условиях ставрида в придонные слои не опускается и до ослабления ветров держится в толще воды или смещается с глубин 120-170 м на глубины 170-210 м, постепенно продвигаясь к северу.
Миграции капской ставриды к настоящему времени практически не изучены, однако, по некоторым наблюдениям, имеют место ее горизонтальные кормовые миграции вдоль юго-западного побережья Африки.
Вертикальные перемещения капской ставриды известны в двух формах. Одна из них - обычные суточные вертикальные миграции, свойственные многим видам рыб: в светлое время суток рыбы держатся в нижних слоях воды, а в темное - поднимаются в верхние; как исключение в зоне заморных придонных вод капская ставрида к грунту не опускается. Другая форма суточных вертикальных перемещений отличается от обычной тем, что иногда в местах опускания вод ставрида мигрирует в придонные слои и ночью.
В придонном слое ставрида может образовывать плотные скопления как днем, так и ночью, причем в лунные ночи скопления у дна более плотные, чем в темные.
В разные годы темп роста капской ставриды меняется незначительно. У берегов Анголы и Намибии встречается ставрида длиной от 11 до 45 см в возрасте от 1 до 9 лет.
Время закладки годовых колец на отолитах капской ставриды в Атлантическом океане приходится на весенний период (октябрь-ноябрь), совпадающий с наибольшим похолоданием вод и с началом наиболее интенсивного нереста ставриды.
К концу первого года жизни южноафриканская ставрида
северного стада имеет длину в среднем 13 см, к концу второго - 18,5 см.
Абсолютная плодовитость капской ставриды в Атлантике колеблется от 12,7 до 740,0 тыс. икринок, составляя в среднем 175,0 тыс.
Нерест ставриды продолжается с различной интенсивностью почти круглый год. Различные группы созревают в разное время. Период наиболее массового икрометания приходится на весну и начало лета (ноябрь-апрель). Икрометание порционное. На нерест ставрида мигрирует в участки с более теплой водой, т.е. в северном направлении; после окончания икрометания откочевывает к югу, образуя скопления вдоль свала глубин.
В зоне бенгельского апвеллинга происходит подъем вод, затрудняющий поступление кислорода из верхних слоев в нижние, поэтому в некоторых местах в придонном слое образуется его дефицит. Так, в декабре 1960 г. в районе 20-24° с.ш. в придонном слое насыщение кислородом составляло всего 3-5% (абсолютное содержание 0,2-0,3 мл/л). Ставрида избегает таких участков и придерживается районов, где насыщение воды кислородом в придонных слоях не опускается ниже 6%.
Питание капской ставриды смешанное, она потребляет в пищу как планктон, так и мелкую рыбу.
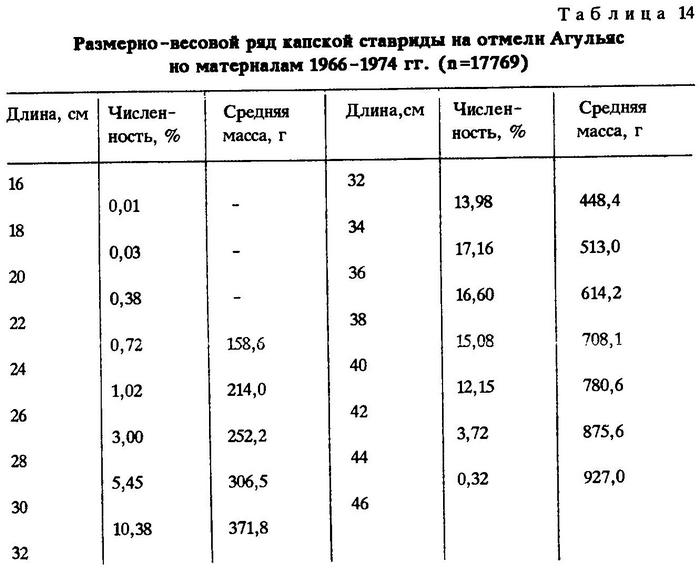
Анализ возрастного состава капской ставриды на отмели Агульяс свидетельствует о том, что здесь в промысловых уловах встречаются особи в возрасте от 1 до 11 лет средней длиной от 18,8 до 40,5 см (n=790).
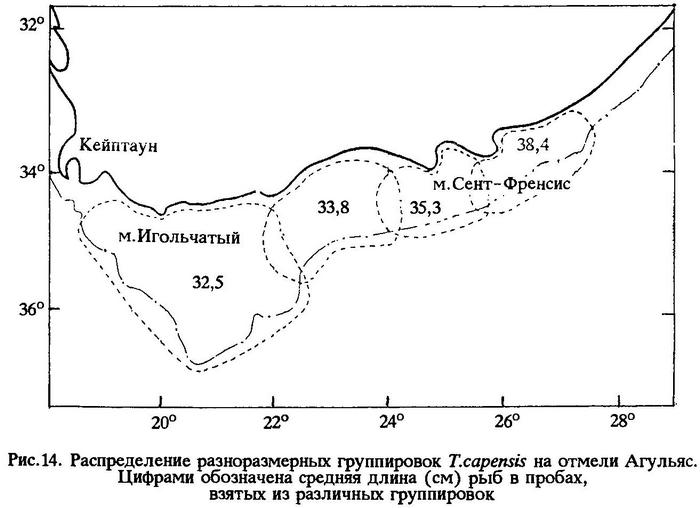
Как было указано выше, капская ставрида обитает не только в Атлантике, но и в Индийском океане, от мыса Игольного до порта Дурбан. По нашим наблюдениям, на отмели Агульяс уловы состояли из рыб длиной от 16 до 45 см (в среднем 35,5 см) и средней массой 539,5 г (табл.14; рис.14).
Суточные вертикальные миграции капской ставриды наблюдаются и на отмели Агульяс. В дневное время ставрида находится в придонном слое, а с наступлением сумерек поднимается в толщу воды, в результате чего резко снижаются ее уловы.
Капская ставрида достигает половой зрелости на 2-3-м году жизни при размерах 22-26 см. Наши материалы показывают, что ставрида, обитающая на отмели Агульяс, нерестует в течение всего года и на всех ее участках. Всюду и всегда встречаются рыбы с гонадами на различных стадиях развития: и незрелые, и готовые к нересту, и уже отметавшие ту или иную порцию икры.
Основываясь на данных анализа состояния гонад, мы пришли к выводу, что капской ставриде свойственно порционное икрометание [Некрасов, 1968,1970], однако число порций не было установлено с полной достоверностью.
О том, что ставрида относится к рыбам с порционным икрометанием, говорит и следующий факт. При измерении ооцитов на срезе яичника ставриды, находящегося на VI- IVn стадии зрелости, выяснилось, что икринки имеют самые различные размеры. Основная масса ооцитов диаметром до 150 мкм находилась в периодах ядерных преобразований и протоплазматического роста, т.е. они являлись резервным фондом для нереста будущего года. Ооциты диаметром от 150 до 550 мкм находились в периоде трофоплазматического роста и образовывали генерации текущего года.
Для более показательной картины асинхронности было просчитано количество ооцитов в поле зрения микроскопа на срезах яичников различных стадий зрелости. Оказалось, что в созревающих гонадах ооциты находятся в разных фазах развития, но преобладают ооциты в фазе интенсивного накопления желтка (16,4%), в то время как в гонадах VI-II стадии зрелости ооциты в этой фазе развития отсутствуют совершенно.
Гистологический анализ гонад ставриды с отмели Агульяс на различных стадиях зрелости в мае-июле показал, что ооциты созревают неодновременно. Асинхронность в развитии ооцитов проявляется уже на ранних стадиях и особенно четко выражена в фазах вителлогенеза. Поэтому при переходе яичника из III в IV стадию зрелости не все ооциты успевают закончить трофоплазматический рост. В результате этого после вымета первой порции икры яичник переходит не на VI, как у единовременно нерестящихся рыб, а на VI-IVn стадию зрелости. Яичники на VI-IVn стадии как по своему внешнему виду, так и по гистологическому строению отличаются от яичников IV стадии лишь большим количеством лопнувших фолликулов.
Яичники ставриды на II стадии зрелости характеризуются хорошо сформированными яйценосными пластинками, несущими комплекс оогоний и ооцитов периода ядерных преобразований и протоплазматического роста (диаметр их достигает 150 мкм). Самки с яичниками на II стадии зрелости имеют коэффициент зрелости 0,92 и не будут участвовать в нересте текущего нерестового сезона.
Яичники ставриды на VI-II стадии зрелости имеют комплекс оогоний, ооцитов периодов ядерных преобразований и протоплазматического роста и, кроме того, резорбирующиеся остаточные дефинитивные ооциты и резорбирующиеся фолликулы. Самки с гонадами на VI-II стадии зрелости имеют коэффициент зрелости от 0,54 до 1,20, в среднем 0,95; они тоже не будут участвовать в нересте текущего года, так как готовая икра уже выметана, а для развития генерации следующего года требуется длительное время.
Яичники ставриды IV стадии зрелости имеют комплекс оогоний и ооцитов (диаметром до 550 мкм) периодов ядерных преобразований, протоплазматического и трофоплазматического роста. Таким образом, в яичниках IV стадии зрелости наряду с резервными, составляющими запас для будущего нереста, имеются ооциты еще в трех фазах развития, среди которых преобладающими по массе являются ооциты фазы интенсивного накопления желтка.
Яичники на VI-IVn стадии зрелости имеют комплекс ооцитов и оогоний периодов ядерных преобразований, протоплазматического и трофоплазматического роста и, кроме того, резорбирующиеся остаточные дефинитивные ооциты. Самки с яичниками на VI-IVn стадии зрелости имели коэффициент зрелости от 0,94 до 2,96, в среднем 1,63. В текущем нерестовом сезоне такие самки будут участвовать в нересте не менее трех раз.
Таким образом, анализ гистологических препаратов яичников ставриды с отмели Агульяс показывает, что у этой ставриды имеет место асинхронность оогенеза, которая обусловливает развитие и вымет не менее четырех порций икры в течение одного нерестового сезона [Шиленкова, 1970].
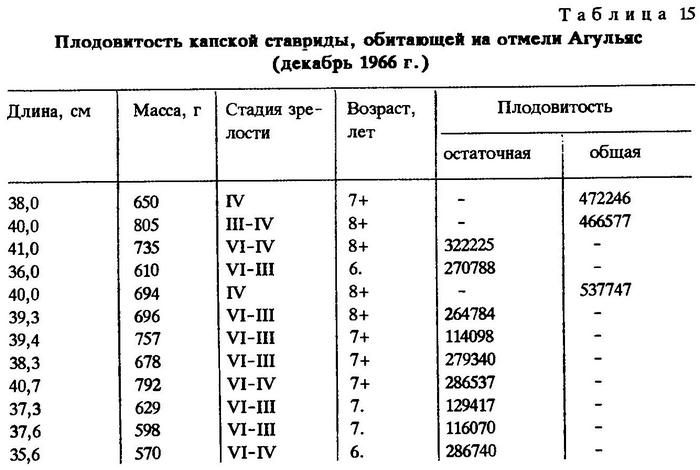
При определении плодовитости порционно нерестующих рыб приходится различать общую (индивидуальную) и остаточную плодовитость. Наши наблюдения показали,что у капской ставриды длиной 38-40 см индивидуальная плодовитость близка к 0,5 млн. икринок (табл.15).
Соотношение полов у капской ставриды близко 1:1, но в разные периоды это обычное соотношение несколько меняется.
Основу питания ставриды на отмели Агульяс составляют ракообразные семейств Euphausiidae и Hypereidae, а также виды Eucalanus, Calanus и Centropagus, Gymnoplea и Amphipoda [Гробов, 1969]. В дневное время питание ее несколько активнее, причем в ритме питания можно подменить примерно четырехчасовую периодичность, которая сглаживается в сумерки. В период нереста питание капской ставриды не прекращается.
Во время интенсивного откорма ставрида придерживается районов опускания поверхностных вод на шельф, где наблюдается максимальное количество зоопланктона (более 1 г/м3).
Хозяйственное значение. Еще в 50-х гг. промыслом ставриды у юго-западного побережья Африки занимались только Ангола и Южно-Африканская Республика. Ниже приведены уловы для того периода по годам в тыс.т [по данным ФАО]:

Южно-Африканская Республика добывает ставриду в основном вместе с сардинопсом кошельковыми неводами. Уловы ставриды Южно-Африканской Республики колеблются по годам и периодам, максимум вылова приходится на май. В Южно-Африканской Республике ставриду в основном используют для изготовления кормовой муки. Часть сырца идет на производство консервов.
По данным Международной комиссии по рыболовству (ИКСЕАФ), сегодня в юго-восточной Атлантике более 65% общего улова рыб и нерыбных объектов приходится на район Намибии и прилегающие воды Анголы, находящиеся под воздействием Бенгельского течения и связанных с ним апвеллингов.
Основу международного промысла морских рыб у намибийского побережья (между устьями рек Кунене и Оранжевой в районе 17°17' - 28° 38' ю.ш.) составляют ставрида и мерлуза (свыше 80% уловов). По данным за 1986 г., общий улов ставриды в этом районе достиг 555 тыс.т [Золотова, 1990]. В число стран, ведущих здесь наиболее интенсивный промысел, входят Россия, ЮАР, Испания, Румыния и Германия (табл.16).

Вопрос о современном состоянии запасов ставриды у побережья Намибии остается дискуссионным. Многие эксперты ИКСЕАФ придерживаются оптимистической точки зрения, согласно которой существующий уровень промыслового изъятия вида не превышает допустимых величин, что позволило сохранить возможный вылов ставриды в этом районе на уровне 440 тыс.т в 1987 г. и 472 тыс.т в 1988 г. Несколько ранее на совещании в Гаване (1986) запасы ставриды были оценены здесь в 2500-3000 тыс.т, а величина допустимого вылова на 1987 г. - в 400-500 тыс.т, что практически соответствует рекомендациям ИКСЕАФ [Экспресс-информация..., 1990].
Запасы капской ставриды на 1987 г., по оценке ВНИРО, выполненной согласно рекомендациям ИКСЕАФ с использованием средних значений пополнения за последние 7 лет, составляли 3,09 млн.т, ОДУ - 500 тыс.т. Таким образом, очевидно, что сегодня запасы капской ставриды находятся на относительно высоком устойчивом уровне. Это определяет выбор режима промысла для прогноза на будущие годы. С учетом фактической интенсивности промысла других стран возможный вылов России может составить 300 тыс.т.
Скопление капской ставриды, обитающее на отмели Агульяс, до настоящего времени не используется отечественным промыслом. Следует отметить, однако, что ставрида с отмели Агульяс значительно отличается от атлантической ставриды по вкусовым качествам. Мясо ее содержит белка - 19,6%, жира - 5,83%, золы - 1,16%, влаги - 73,80%; оно очень вкусное, нежное и высококалорийное (135 кал. на 100 г). Рекомендуется использовать для приготовления копченостей и консервов в масле [Ковальчук, 1965, 1968].
Интересно отметить, что у других видов рода Trachurus, обитающих на шельфе, например, у Trachurus mediterraneus ponticus, Trachurus trecae, Trachurus delagoa, Trachurus capensis размерно-половое соотношение второго типа. Наиболее наглядно это видно на примере популяции капской ставриды Trachurus capensis, обитающей на отмели Агульяс (рис.23) (n = 2329 экз.). Если при длине тела капской ставриды 38-42 см в возрасте 7-10 лет самцы составляли лишь 41,6-41,4%, а среди более крупных рыб длиной 42-44 см в возрасте 12 лет и более лет самцов было всего 28,6%, то среди рыб длиной 26-30 см в возрасте 3-4 года доля самцов составила 59,2-58,0%.
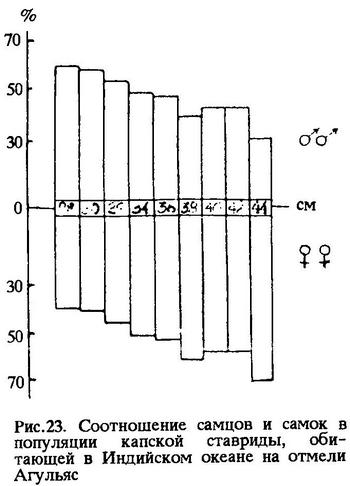
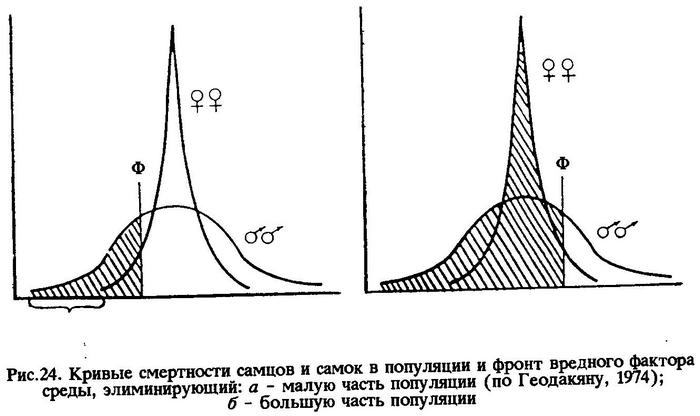
Значит, более тесная связь особей мужского пола со средой обеспечивается большей величиной их фенотипической дисперсии по сравнению с особями женского пола. Типичной иллюстрацией к этому служит размерно-половое соотношение в популяции капской ставриды, обитающей на отмели Агульяс (рис.23). В этом случае кривые смертности имеют вид, изображенный на рис.24. По оси абсцисс - устойчивость к вредному фактору среды; по оси ординат - количество особей с данной устойчивостью. На рисунке видно, что кривая смертности женских особей имеет положительный эксцесс, а мужских - отрицательный. И это понятно, потому что у мужских особей разнообразных отклонений от моды популяции больше, чем у женских, а за счет этого образуется зона дискомфорта (Зд). Зона дискомфорта тем шире, чем популяция пластичнее. На рис.24а показано, что вредный фактор среды (Ф - фронт вредного фактора) элиминирует малую часть популяции (заштриховано на рисунке), состоящую преимущественно из самцов. Именно поэтому к концу жизни поколения в популяции капской ставриды остается больше самок. Такая форма связи со средой ценой высокой смертности самцов, как уже отмечалось выше, выгодна для популяции в целом.