Trachurus indicus Nekrasov, 1966
Индийская ставрида, индийская ставрида побережья Омана, индийская ставрида Суэцкого залива, индийская ставрида Персидского залива, индийская ставрида отмели Сая-де-Малья (Rus),
Arabian scad
(Eng)
Синонимы:
Trachurus indicus sayademalha Nekrasov, 1966 Индийская ставрида отмели Сая-де-Малья (Rus),
Trachurus mediterraneus indicus Nekrasov, 1966
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus indicus Necrassov - индийская ставрида
Описание. D I, VIII, I 27-34, наиболее часто 31; А II, I 23-31, наичаще 27; L.l. 67-78, наичаще 73, из них колючих 33-42, наичаще 37. Боковая линия спереди расположена выше середины тела, с крутым изгибом под началом мягкого спинного плавника до его 4-11-го луча, далее прямая и расположена посредине тела; длина задней прямой части боковой линии больше хорды дуги передней части в 1,04 (0,95-1,30) раза. Высота наиболее высоких щитков кривой (передней) части боковой линии 7,9-18,5%, наичаще 13,23 % длины головы; высота щитков прямой (задней) части боковой линии 11,5-20,5%, наичаще 15,53% той же длины. Первый колючий щиток лежит под основанием 7-11-го луча второго спинного плавника. Спинные ветви боковой линии оканчиваются на вертикали основания VIIIDI (VILDI-IDII). Горизонтальный диаметр глаза 22,0-36,0%, наичаще 29,73%, длина верхней челюсти 34,0-45,0%, наичаще 39,73% длины головы. Тело толстое, округлое; наибольшая высота тела 21,7-27,1%, наичаще 23,14% его длины. Длина головы 27,0-34,0%, наичаще 29,3% длины тела, длина грудных плавников - 21,9-32,0%, наичаще 26,2%, расстояние от конца рыла до вертикали начала основания брюшных плавников 27,0-37,0%, наичаще 31,8%, расстояние от конца рыла до вертикали начала основания первого спинного плавника - 30,0-40,0%, наичаще 33,5%, расстояние от конца рыла до вертикали основания первого луча анального плавника - 47,4-59,8%, наичаще 53,1% длины тела. Последние лучи второго спинного и анального плавников по сравнению с предпоследними несколько увеличены, а расстояние между основаниями последнего и предпоследнего луча анального плавника в 1,10-1,95, наичаще в 1,40 раза превосходит расстояние между основаниями предпоследнего и предшествующего ему луча.
Спина темная, зеленовато-синяя, иногда с 8-11 широкими поперечными темными полосами; бока и брюхо перламутрово-серебристые; грудные и спинные плавники темно-серые, хвостовой - желтовато-серый, иногда серый, прочие плавники светлые; на жаберной крышке черное пятно.
Описание дано по 622 экз. длиной 9-29 см из Красного моря, Персидского и Аденского заливов, Аравийского моря, с банки Сая-де-Малья [Некрасов, 1966а, 1978].
Голотип - экземпляр длиной 267 мм добыт в районе с координатами 15°26' с.ш. 52°00' в.д. 4 мая 1963 г.; N 37519 в коллекции Зоологического института РАН (Санкт-Петербург).
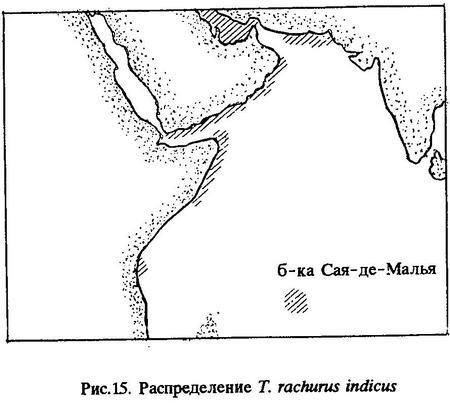
Распространение (рис.15). Наши исследования показали, что ареал Trachurus indicus значительно шире, чем мы считали ранее [Некрасов, 1964а, 1966а], и охватывает Суэцкий залив и прилегающие к нему воды Красного моря в районе Хургады, южную часть Красного моря, Аденский залив у побережий Омана и Маската, включая район о-ва Масира, Персидский залив у берегов Кувейта, Бахрейна и Катара, прибрежные воды Пакистана (у побережья Макран), Сомали (мыс Рас-Хафун) и далее на юг до залива Формоза, а также район банки Сая-де-Малья [Некрасов, 1977] (см.рис.15). Необходимо заметить, что в работе "Ставриды восточного побережья Африки" [Некрасов, 1970] я ошибочно указал среди районов обитания Trachurus delagoa банку Сая-де-Малья.
Сравнительные замечания. В 1966 г. мною была описана индийская ставрида, обитающая в Аравийском море, как подвид Trachurus mediterraneus indicus. Основой для выделения этой ставриды лишь на уровне подвида послужило отсутствие хиатуса в диагностических признаках Trachurus mediterraneus mediterraneus и Trachurus mediterraneus indicus. Позднее в литературе появились работы, справедливо подвергающие сомнению наше прежнее утверждение о том, что индийская ставрида является подвидом средиземноморского вида [Rass, 1973]. Дальнейшие исследования показали, что указанные филогенетически близкие формы различаются на уровне вида, хотя граница между ними не очень четкая. Ф.Г.Берри и Л.Коэн (1972) возвели описанную мною форму в ранг вида Trachurus indicus Necrasov. Мне остается только присоединиться к их мнению и добавить, что более детальное сравнительное изучение Trachurus indicus, добытых мною в Красном и Аравийском морях и Персидском заливе, и Trachurus mediterraneus mediterraneus, пойманных нами в Средиземном море у Александрии, выявило различия не только по высоте щитков в передней части боковой линии, по величине отношения хорды дуги кривой части боковой линии к длине тела и по длине грудных плавников, на что я указывал ранее [Некрасов, 1966а], но и по числу щитков в боковой линии, по отношению длины прямой части боковой линии к длине хорды дуги кривой, по длине спинной ветви боковой линии, по расстоянию от вершины рыла до вертикали начала основания анального плавника и некоторым другим признакам (табл.18).
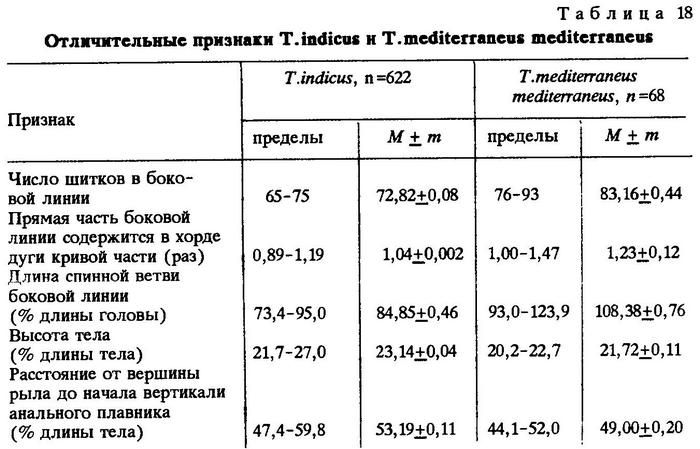
Нам представилась возможность собрать материал из разных районов обитания Trachurus indicus. Сравнивая географически разобщенные популяции, мы установили существенные различия по некоторым признакам. Приводим оценки тех морфологических признаков Trachurus indicus из различных районов Индийского океана, по которым установлены наиболее четкие различия между популяциями (табл.19-26).
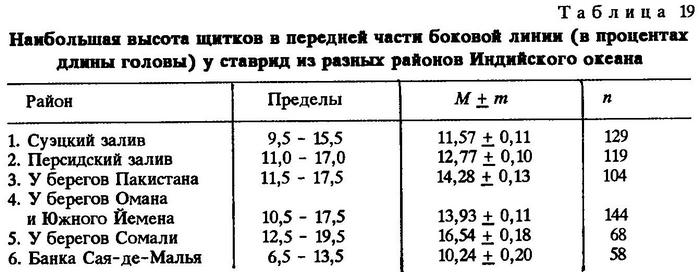
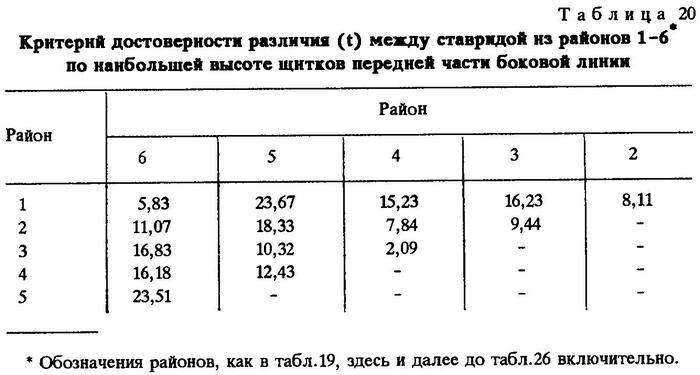

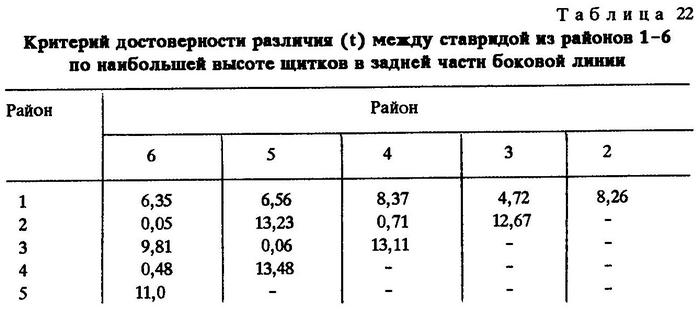
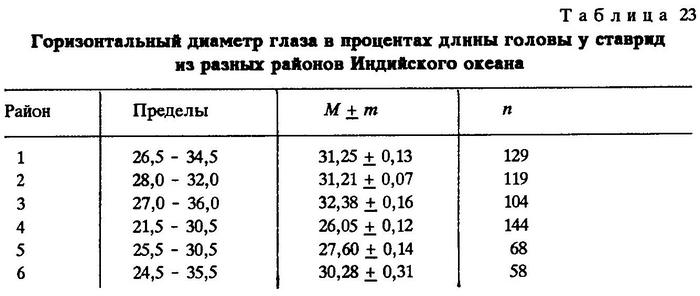
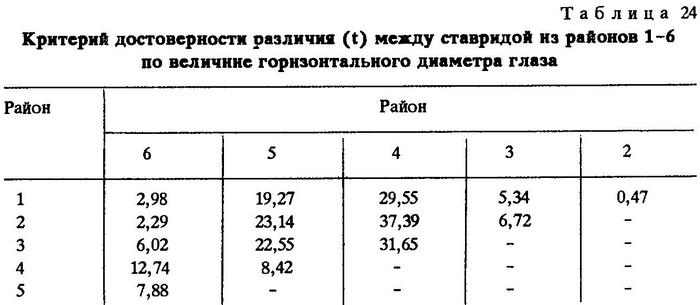
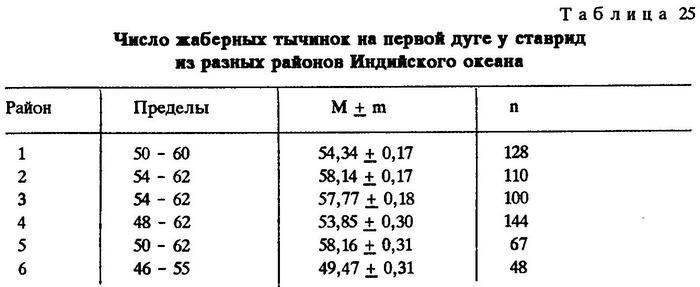
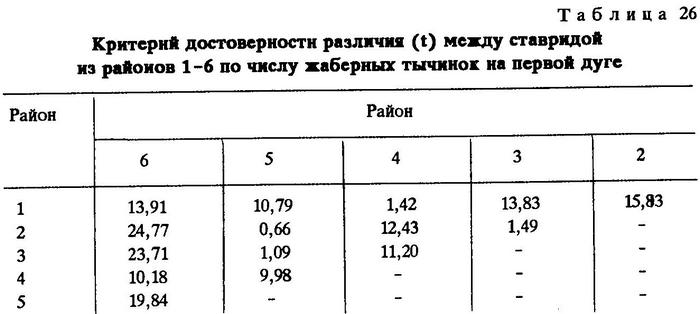
Из таблиц видно, что критерий достоверности различия (t) по указанным признакам почти везде значительно более 3,0. Это говорит о наличии локальных популяций индийской ставриды в обследованном регионе.
Анализируя данные таблиц, нетрудно заметить клинальную изменчивость некоторых признаков в направлении с севера на юг. Однако популяции, слагающие клины Trachurus indicus, достаточно хорошо изолированы. Считается, что популяции можно отнести к особому подвиду при значении CD равном 1,28 [Майр, 1971]. Судя по нашим данным, одна из популяций, обитающая на банке Сая-де-Малья, заслуживает подвидового ранга.
Наибольшие различия между ставридами банки Сая-де-Малья и ставридами, обитающими у мыса Рас-Хафун и у побережья Пакистана, наблюдаются по двум признакам - по числу жаберных тычинок на первой дуге и по наибольшей высоте щитков в передней части боковой линии.
По числу жаберных тычинок значение CD между популяциями ставрид из районов банки Сая-де-Малья и мыса Рас-Хафун CD равно 1,86, между популяциями из районов банки Сая-де-Малья и побережья Пакистана - 2,06. По второму признаку соответственно CD = 2,09 и 1,45.
Различия значений этих же признаков ставрид, обитающих на банке Сая-де-Малья и в более отдаленных районах, несколько сглаживаются. Так, сравнение популяции ставриды, обитающей на банке Сая-де-Малья, с популяцией, обитающей у берегов Омана и Адена, выявляет различие на уровне подвида лишь по одному признаку - по наибольшей высоте щитков в передней части боковой линии (CD = 1,32), а сравнение с популяцией, обитающей в Персидском заливе, дает различие на уровне подвида только по числу жаберных тычинок (CD = 2,20). Обращает на себя внимание тот факт, что популяция ставриды, обитающая на самом севере ареала вида - в Суэцком заливе, отличается от популяций, обитающих у берегов Омана и Южного Йемена, значительно сильнее, чем от популяций, обитающих в более южных широтах. Это вполне понятно, так как клинальная изменчивость при переходе из северного полушария в южное меняет направление градиента [Hoffmeister, Torre, 1961; цит. по Завадскому, 1968 и Некрасову, 1966б, 1966в].
Таким образом, мы установили, что в западной части Индийского океана обитает политипический вид Trachurus idicus с двумя подвидами: Trachurus indicus sayademalha Necrassov (голотип - экземпляр длиной 251 мм, 10°24' ю.ш., 61°50' в.д., 19 сентября 1964 г., N 42498 в коллекции Зоологического института РАН и 7 паратипов в той же коллекции) и Trachurus indicus indicus. Последний подвид включает в себя несколько локальных популяций, обитающих в Суэцком и Персидском заливах, у берегов Сомали, юга Аравии и Пакистана.
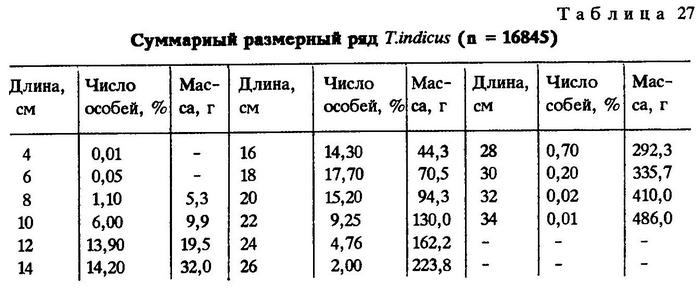
Биология. Суммарный размерный ряд Trachurus indicus, составленный на основании материалов из всех известных районов ее обитания, показывает, что основу уловов составляют особи средней длины 15,9 см и массой тела примерно 59 г, но отдельные экземпляры достигают длины 33 см и массы 486 г. (табл.27). Однако по отдельным районам наблюдаются значительные различия в длине тела Trachurus indicus.
Результаты распределения индийской ставриды позволили выделить четыре основных биологических группировки (стада), локализующихся в Суэцком заливе, у побережья Омана, в Персидском заливе и на отмели Сая-де-Малья. Первые три представляют собой локальные стада подвида Trachurus indicus indicus с сильной клинальной изменчивостью в широтном направлении. Четвертая - подвид Trachurus indicus sayademalha. В связи с этим нам представляется целесообразным излагать материалы по биологии и условиям обитания индийской ставриды отдельно по каждому стаду.
Индийская ставрида Суэцкого залива. Суэцкий залив представляет собой относительно мелководный участок северной части Красного моря. Здесь температура воды у дна (50-80 м) зимой составляет 17,1-17,3°С , а летом достигает 26,0°С. Кислородный режим в Суэцком заливе удовлетворительный. Насыщенность воды кислородом претерпевает лишь небольшие изменения и в течение всего года близка к 100%. Так, даже в придонном слое насыщение воды кислородом составляет в декабре 98-102%, в марте - 98-104%, в апреле - 98-103% и лишь в сентябре падает до 71%. Анализ видового состава уловов показал, что в Суэцком заливе и сопредельном с ним участке Красного моря (район Хургады) ставрида зимой составляет около 15,0%, а летом 32,3% общего вылова рыбы. Южнее Хургады взрослые особи не отмечены, а молодь лишь в небольшом количестве встречалась в уловах бортовых ловушек.
В Суэцком заливе, в районе архипелага Хургада и в бухте Сафага индийская ставрида - самая мелкая, ее средняя длина составляет всего 11,2 см , а средняя масса - 22 г. Однако в пределах отдельных районов размерный состав ставрид значительно варьируется и по участкам распределения и по сезонам. Так, в северной части Красного моря в районе Хургады пробы, взятые в марте-апреле 1965 г., состояли из наиболее крупных рыб. Более мелкие рыбы встречались в центральной части Суэцкого залива как в зимние, так и в весенние месяцы. В северной части залива во все периоды наблюдений (осенью, зимой, весной) вылавливались самые мелкие рыбы. Во всех участках, охваченных наблюдениями, можно было заметить увеличение средних размеров ставриды от осени к весне. Возможно, что эти изменения в размерном составе уловов ставриды в какой-то мере отражают отсутствие материалов за полный годовой цикл. Но несомненно, что в северной части залива обитают самые мелкие рыбы с большей долей неполовозрелых особей.
Trachurus indicus Суэцкого залива - рыба с коротким жизненным циклом и малой длиной тела. Основу уловов составляют особи в возрасте от сеголетков до трехгодовиков. Более старые рыбы (до пяти годовиков) встречаются в небольшом количестве. Длина тела ставриды из уловов тралами и бортовой ловушкой колеблется от 3 до 17 см.
Половая зрелость у индийской ставриды наступает в основном в возрасте годовика при достижении размеров 10-11 см.
Нам представилась возможность проследить за созреванием гонад индийской ставриды в Суэцком заливе по сезонам года. Было установлено, что у Trachurus indicus в течение почти всего периода исследований наблюдались гонады на различных стадиях развития: от II стадии до преднерестовой, нерестовой и посленерестовой. Лишь в пробе, взятой в летний сезон 1966 г. в северной части залива, все рыбы, даже наиболее крупные, имели гонады в состоянии покоя. В зимне-весенний период наблюдалось резкое уменьшение количества рыб с половыми железами на стадии II, а также ювенильных особей (размерами до 7-8 см), и происходило активное созревание большинства рыб. Если в ноябрьских пробах количество рыб с признаками прошедшего нереста составляло 5%, то в декабре оно увеличилось до 40-50%, а в январе - до 70-100%. Преобладающими на протяжении этого периода стадиями зрелости половых продуктов у ставриды были III-IV и IV для впервые созревающих рыб и VI-III и VI-IV для рыб, созревающих повторно. Разгар нереста происходит, по-видимому, с января по апрель, когда количество особей с текучими и посленерестовыми гонадами достигает максимума. В этот период наблюдается наиболее низкая температура воды в заливе. Все эти наблюдения дают основание полагать, что активный нерест ставриды в Суэцком заливе начинается в ноябре и продолжается до апреля включительно, т.е. в холодный период года. Следует отметить, что в течение пяти-шести зимне-весенних месяцев происходит последовательное вступление в нерестовую часть стада все новых и новых групп вновь созревающих рыб, которые в силу многопорционности икрометания ставриды обеспечивают непрерывность нереста на протяжении длительного периода. Абсолютная плодовитость
Trachurus indicus при длине 20 см близка к 60-90 тыс. икринок.
Анализ питания показал, что основу пищи Trachurus indicus в Суэцком заливе составляют веслоногие рачки. В светлое время суток ставрида питается менее интенсивно, чем ночью. Так с 8 ч. утра до 17 ч. средний балл наполнения желудков рыб не превышал 0,6 и редко достигал 1. В ночное время наполнение желудков достигало в среднем 2,3-2,7 балла.
Такой физиологический показатель, как жирность, обнаруживает у индийской ставриды популяции Суэцкого зклива определенную тенденцию к уменьшению от осени к весне. Средняя жирность ставриды в ноябре была равна 3,65%, в марте - 0,77% и в апреле - 0,37%. Снижение жирности, очевидно, связано с активно проходящим в этот период нерестом ставриды.
Больших перемещений в Красном море Trachurus indicus, по-видимому, не совершает. Установлена вертикальная миграция рыб в зависимости от времени суток. В дневное время ставрида обитает у дна и облавливается тралами. Ночью она поднимается в толщу воды, где может вылавливаться бортовыми ловушками и кошельковыми неводами, составляя до 42-46% общего улова вместе с другим видом ставриды рода Decapterus.
Индийская ставрида оманского побережья. Район оманского побережья расположен в северной части Аравийского моря над материковым склоном вдоль юго-восточного берега Аравийского полуострова между мысом Рас-Фартак и 60° в.д. Гидрологический режим этого района, как и всего Аравийского моря, определяется муссонным характером атмосферной циркуляции. Примерно с октября по май в этом районе господствуют устойчивые северо-восточные ветры (зимний муссон), в летние месяцы - сильные юго-западные ветры (летний муссон). В соответствии с муссонным характером ветровой деятельности движение вод в районе побережья Омана подвержено значительным сезонным колебаниям. В целом поверхностный слой воды этого района благоприятен для обитания морских животных. Для него характерны высокая температура (21-27,5°С), соленость 36,5-36,8‰, высокое насыщение кислородом (105-110%) и достаточно высокое содержание биогенных элементов. Все это обеспечивает активное продуцирование органического вещества, в результате чего этот район является одним из наиболее продуктивных в западной части Индийского океана. Но на глубине 150-200 м и ниже залегает мощный слой подповерхностных вод Аравийского моря, имеющих несколько пониженную соленость (35,5-36,0‰) и температуру (15-20°С), крайне низкое содержание кислорода (до 0,5 мл/л), а в некоторых участках - отчетливые признаки сероводородного заражения. При сгонных ветрах (период летнего муссона) эти воды поднимаются на шельфовые участки, что пагубно сказывается на местной фауне. В этот сезон основная масса рыбы мигрирует на мелководные участки (с глубиной 30-40 м) прибрежной зоны, которые вследствие ветрового перемешивания хорошо аэрированы. В этом районе семейство Carangidae представлено большим числом видов, из которых наиболее многочислен Trachurus indicus. Здесь индийская ставрида крупнее, чем в Суэцком заливе, имеет большую ценность как сырье и по праву считается одной из основных промысловых рыб района. В уловах эта ставрида представлена рыбами длиной от 10 до 30 см, при среднем размере 19 см и средней массе около 100 г. В пробах отмечены рыбы в возрасте от 2 до 6 лет с преобладанием четырех-пятигодовиков. Половой зрелости ставрида достигает на 1-2-м году жизни при размерах 11-12 см. В Аравийском море у побережья Омана половозрелость индийской ставриды наступает при длине тела 11,5-13,0 см, т.е. при несколько большей, чем у ставрид, обитающих в Суэцком и Персидском заливах.
Анализ половых желез показал, что в течение всего периода наблюдений, который охватывал и зимние, и летние месяцы, в уловах встречаются особи с гонадами на различных стадиях зрелости - от незрелых до текучих, посленерестовых и с повторно созревающими очередными порциями половых продуктов. Это свидетельствует о чрезвычайно растянутом периоде нереста, в результате чего едва ли можно ожидать образования стабильных нерестовых концентраций. Однако в течение всего года в районе оманского побережья обнаруживаются плотные скопления индийской ставриды, состоящие из рыб различных размеров, возрастов и зрелости. Устойчивость этих скоплений определяется гидрометеорологическими условиями.
Обращает на себя внимание тот факт, что летом в Аравийском море в уловах нет молоди. Видимо, пик нереста все-таки приурочен к определенному периоду.
Собранный нами материал в годичной экспедиции в северо-восточную часть Аравийского моря, обработанный Т.И.Фурсой (1969, 1973), дал возможность установить, что во все сезоны года в данном районе часто встречались икра и личинки ставрид, составляя до 8,8% улова. Не менее богат личинками рода Trachurus и Аденский залив [Кракатица, Фурса, 1968]. Но наиболее многочисленны личинки Trachurus indicus в двух участках у побережья Омана - в районе бухты Саукара и в заливе Масира [Цокур, 1974]. У западных берегов Индии личинок Trachurus indicus не обнаружено [Фурса, 1969].
На примере популяции индийской ставриды, обитающей у берегов Омана, А.Г.Цокур (1974) убедительно показал, что основная масса личинок (97,4%) распространена на шельфе или в непосредственной близости от него. Кроме того, им было показано, что даже в таком теплом водоеме, как Аравийское море, личинки Trachurus indicus придерживаются участков с наиболее низкой температурой воды - до 16,7°С в слое 0-100 м. Наши наблюдения показали, что в Персидском заливе нерест этой ставриды происходит при еще более низкой температуре - до 15,4°С.
Средняя масса индийской ставриды у оманского побережья значительно выше, чем у ставриды, обитающей в Суэцком заливе, что связано, очевидно, с более благоприятной кормовой базой. В этом районе ставрида интенсивно питается как днем, так и ночью. Так, например, между 12 и 14 ч. средний балл наполнения желудков был равен 2,0, а ночью, между 0 и 2 ч. - 1,8.
Одной из особенностей индийской ставриды района оманского побережья является высокая степень ее географической и экологической локализации. Весь жизненный цикл этой рыбы протекает в пределах данного района, где в зависимости от океанографических условий она совершает лишь местные перемещения, но не столько вдоль берега, сколько между относительно глубоководными и прибрежными участками. Оптимальная глубина обитания ставриды в период зимнего муссона (т.е. в период наиболее благоприятных условий среды) - 90-140 м. Однако при выходе на шельф холодных и обедненных кислородом вод подповерхностного слоя индийская ставрида, как отмечалось выше, покидает эти глубины, перемещаясь в прибрежные мелководные участки. Индикатором появления на шельфе подповерхностных вод и начала миграции ставриды является температура воды. Нами ни разу не были отмечены скопления ставриды в местах с температурой воды ниже 20°С. Но главной причиной ее отхода на мелководье, конечно, является дефицит кислорода. Индийская ставрида очень чувствительна к этому фактору и при насыщении кислородом воды, не превышающем 30%, в уловах отсутствует, хотя некоторые другие виды рыб (заурида, розовый карась и др.) продолжают оставаться в этом районе [Некрасов, 1964а, 1973].
Как и все другие виды ставрид, индийская ставрида района оманского побережья совершает суточные вертикальные миграции: днем она держится у дна, а ночью поднимается в поверхностные слои воды и рассеивается, поэтому максимальные уловы тралом наблюдаются в дневное время.
Индийская ставрида Персидского залива имеет среднюю длину 16,1 см и среднюю массу 61 г.
В Персидском заливе, особенно в его северной части у берегов Кувейта, в массе ставрида появляется начиная с ноября, когда температура воды у дна снижается до 23,5°С. В течение всего зимнего периода в траловых уловах встречаются в основном половозрелые особи относительно крупных размеров - от 10 до 24 см. В мае, при значительном потеплении воды, крупные особи в уловах исчезают, но у южной границы Кувейта и у берегов Саудовской Аравии в массе появляются молодые рыбы длиной 8-12 см. В июне, июле и августе ставрида нами отмечена лишь единично в центральной части Персидского залива и близ о-ва Бахрейн на глубине 72 м и более.
Половой зрелости в Персидском заливе ставрида достигает в возрасте годовика при длине 10-11 см. Здесь наши наблюдения показали, что нерест Trachurus indicus начинается в ноябре, хотя в то же время еще у 60% особей гонады находятся на II стадии зрелости. В декабре-феврале разгар нереста. В марте нерест заканчивается. В мае все особи имеют половые продукты в состоянии покоя. В июле у некоторых крупных особей гонады находятся на III стадии зрелости, а у одного экземпляра были даже на стадии III-IV.
Таким образом, нами установлено, что и в Персидском заливе интенсивный нерест индийской ставриды наблюдается в наиболее холодное время года. Для рода Trachurus этот факт отмечен и другими исследователями [Оверко, 1969; Седлецкая, 1973 и др.].
Выделение таксономических единиц низших рангов, обоснование норм вылова, составление рыбохозяйственных прогнозов и т.п. - все эти важные вопросы в большей мере проясняются в связи с установлением возраста и темпа роста рыб. Определение возраста рыб имеет важное практическое значение. В основе его лежит подсчет регулярно формирующихся слоев в минерализованных структурах организма [Чугунова,1959; Брюзгин,1969; Мина, Клевезаль, 1976]. Однако до настоящего времени не существует единого мнения относительно того, что определяет формирование годовых отметок на регистрирующих структурах. Если для рыб умеренных и высоких широт известно, что закладка колец так или иначе связана со сменой сезонов, и очевидно, что годовое кольцо закладывается один раз в году, то для тропических видов, обитающих в условиях отсутствия резких колебаний температуры воды, продолжительности светового дня, кормовой базы и пр., причины закладки колец до настоящего времени не выяснены.
Долгое время считалось, что ритмичность роста рыб обусловлена лишь воздействием температуры воды. Существовал даже термин "зимнее кольцо", т. е. полагали, что кольцо образуется зимой при самой низкой температуре воды [Морозов, 1934 и др.]. В дальнейшем было показано, что у рыб умеренной зоны и субтропиков кольцо закладывается весной [Чугунова, 1940а; Ел-Зарка, 1960, Аl-Hamed, 1966 и др.], летом [Соловьева, 1938; Суворов, 1948; Сафьянова, Ревина, 1967] или осенью [Меnon, 1953; Буй Дин Чунг, Дружинин, 1965 и др.]. К тому же было экспериментально доказано образование "зимнего кольца" у мерланга, которого выдерживали в течение зимы в танке с теплой водой [Меnon, 1953]. Эти факты свидетельствуют о явной переоценке роли температурного фактора. Тем не менее и в настоящее время иногда утверждают, что разница в 4-5°С достаточна, чтобы вызвать замедление роста рыб и появление кольца на регистрирующих структурах [Скорняков, 1964 и др.]. В связи с этим обращает на себя внимание тот факт, что у рыб тропического и особенно экваториального климатического пояса, где нет резкой смены сезонов, скорость роста также неравномерна в течение года [Mohr, 1921; Chidambaram, Krishamurthy, 1951; Jhingran, 1957; Seshappa, 1958; Balan, 1959; Липская, 1966a; Решетников, Кларо, 1976 и др]. Еще в начале нашего века высказывалось сомнение в том, что причиной этого является чередование климатических условий [Томпсон, 1926].
Утверждение, что ритмичность роста рыб определяется их питанием [Чугунов, 1926; Соловьева, 1938; Суворов, 1948; Nair, 1949; Radhakrishnan, 1954], также вызывало возражения. Обращалось внимание на значительные изменения в кальциевом обмене у рыб в период созревания гонад. Большое количество кальция мобилизуется на построение половых продуктов, и, по-видимому, в результате этого на чешуе и костях появляются зоны с малым его количеством [Липская, 1967, 1969; Решетников, Кларо, 1976 и др.]. Уместно отметить, что изменения количества растворенного в морской воде кальция не влияют на образование микрокристаллических зерен СаСO3 на отолитах, - скорость, роста отолита при этом не изменяется [Takaaki, 1960].
Н.Я.Липская (1967,1969) показала, что у тропических рыб, несмотря на интенсивное питание в период нереста (а это характерно для многих морских рыб с длительным порционным нерестом [Асланова, 1954; Павловская, 1963]), большие приросты на чешуе отмечены у особей, потреблявших меньше корма и не участвовавших в нересте, тогда как закладка кольца происходит у рыб, нерестящихся и потребляющих много корма. Это обстоятельство Н.Я.Липская объясняет тем, что в нерестовый период на формирование половых продуктов затрачивается в 40-60 раз больше энергии, чем на рост. А так как у тропических рыб время нереста очень растянуто, то закладка кольца у них происходит по завершении периода наиболее интенсивного нереста в любое время года [Липская, 1966а,б, 1969; Липская, Овен, 1970; Решетников, Кларо, 1976].
Некоторые исследователи полагают, что у многих видов рыб умеренной зоны [Чугунова, 1940а,б; Замахаев, 1948], а также у тропических (Naseef,1961; Довгопол, 1969; Белан, 1971] нерестовые отметки являются годовыми кольцами. Наряду с этим в многочисленных работах последних лет рассматривается влияние на закладку колец у тропических рыб муссонов, сезонов дождей и других факторов внешней среды.
Для проверки предположения о связи нереста с закладкой колец на регистрирующих структурах тропических рыб были проведены исследования в Аравийском море.
Анализ материала показал наличие колец на чешуе и отолитах собранных экземпляров. Во все сезоны у донных и пелагических рыб на отолитах и чешуе отмечались большие и малые приросты, только что сформированное кольцо. Так, по данным за 1961-1973 гг. в популяциях индийской ставриды зимой с законченным последним кольцом было 77,0% особей, с приростом - 23,0%, весной - 59,8 и 40,2%, летом - 45,1 и 54,9%, осенью - 52,0 и 48,0%. Как видим, наибольшее число рыб с законченным кольцом приходится на зиму. Тогда же отмечено и наибольшее число отнерестившихся рыб (табл.28, 29).
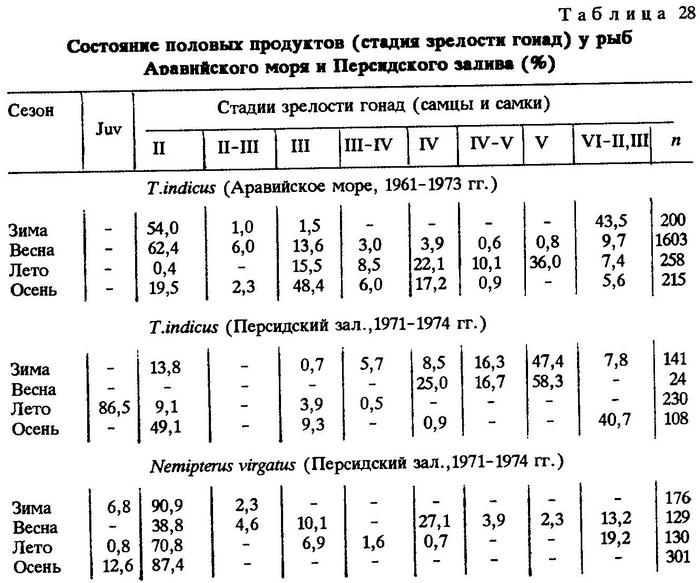
Следует заметить, что последнее кольцо по краю чешуи или отолита трудно различимо. Наиболее четко оно проявляется при приросте, пусть даже незначительном. Поэтому, хотя сама закладка кольца относится к периоду наиболее интенсивного нереста, кольцо обнаруживается несколько позже, что совпадает с периодом затухания нереста. Итак, можно сделать вывод, полностью совпадающий с утверждениями других авторов о том, что кольца на регистрирующих структурах тропических рыб, в частности индийской ставриды Аравийского моря, закладываются во время нереста.
Поскольку из литературы известно, что у некоторых тропических рыб, возможно, образуется два кольца в год [Hattori, 1953; Seshappa, Bhimachar, 1954; Скорняков, 1964; Poinsard, Тrоаес,1966; Решетников, Кларо, 1976 и др.], нет оснований исключать возможность двукратного нереста, а стало быть, и вероятности образования двух колец за год жизни и у индийской ставриды.
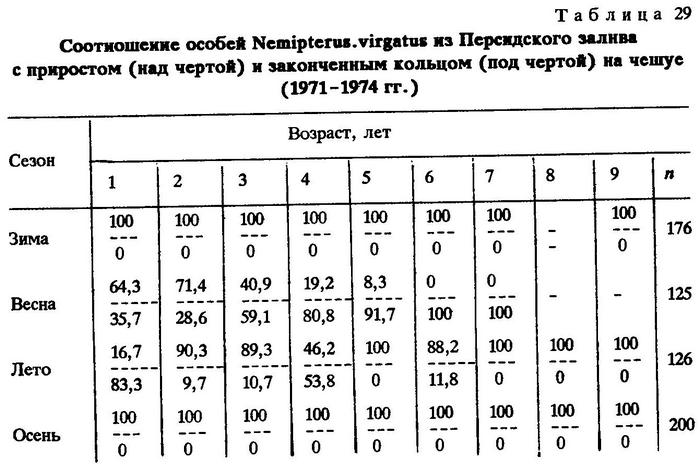
Чтобы убедиться в обратном или подтвердить это предположение, мною был привлечен материал, собранный в 1971-1974 гг. в Персидском заливе у берегов Кувейта. Регион интересен большой сезонной изменчивостью температуры (среднеянварская температура в поверхностном слое воды 15,9°С, среднеавгустовская - 32,9°С), а также четкой приуроченностью сроков нереста рыб к сезонам, что полностью соответствует правилу Ортона: виды, близкие к теплым границам своего распространения, размножаются в холодное время; виды, близкие к холодным границам своего распространения, размножаются в теплое время. Это обеспечивает появление и рост молоди в оптимальных для нее температурных условиях, так как молодь менее терпима к крайним условиям, чем взрослые особи [Овен, 1976]. Одни виды, например, индийская ставрида, нерестятся в холодное время года, другие, например, нитепер Nemipterus virgatus, - в теплое (см.табл.29). Эти виды являются удобными объектами нашего исследования и по той причине, что в своей массе они впервые нерестятся в конце первого года жизни, как было непосредственно установлено при длительных (2,5 года) наблюдениях в Персидском заливе (пробы отбирали ежемесячно).
Из приведенных данных (см.табл. 29) хорошо видно, что индийская ставрида нерестится один раз в год; нерест продолжается с осени до весны с паузой летом. Осенью, зимой и весной темп роста рыб замедляется (наибольшие приросты отмечены к концу лета), причем закладка кольца приходится чаще на конец зимы - начало весны, т. е. происходит один раз в год - во время нереста. Эту нерестовую отметку можно считать годовым кольцом. Аналогичные данные получены и в отношении нитепера, однако нерест и закладка кольца у него приходится на теплое время года (см.табл. 28 и 29).
Определение на этом основании возраста индийской ставриды и нитепера, а также средней длины тела рыб разного возраста в Персидском заливе и, далее, сравнение между собой рыб с одинаковым количеством колец на чешуе и отолитах из Персидского залива и Аравийского моря показало, что средние значения длины тела у них очень близки (табл.30). Этот факт позволяет заключить, что и в Аравийском море они нерестятся раз в год и нерестовая отметка соответствует событию "год жизни".

Понятно, что констатация этого факта недостаточна для решения всех вопросов, связанных с определением возраста и роста тропических рыб. Немаловажно установить причины образования годовых колец у неполовозрелых особей тех видов, которые нерестятся не на первом году жизни. Тем не менее есть надежда, что наши выводы будут полезны в практике рыбохозяйственных исследований тропических рыб, приступающих к нересту в конце первого года жизни.
В траловых уловах у побережья Омана за весь период работ преобладали четырех-пятигодовики, составляя 32,8% и 32,5% соответственно. В Персидском заливе основу уловов составляли трех- и четырехгодовики (34,2 и 40,7%). В Суэцком заливе в районе Хургады до 71,2% уловов составляли двухлетки.
Хозяйственное значение. Вылов индийской ставриды в Индийском океане в период ее активного промысла достигал 33 тыс.т. Промысловое освоение районов западной части океана рыбодобывающими судами СССР было начато в 1963 г. В Индийский океан вышли первые траулеры - РТМ "Евпатория", "Алушта", "Руставели", "Кореиз", которые производили опытно-промысловый лов донных рыб главным образом в районе мыса Рас-Фартак и продолжали его с перерывами и в 1964 г. Более полное развитие промысла в Индийском океане началось с 1965 г., когда траулеры типа РТМ и СРТМ стали работать здесь в течение круглого года, причем в среднем на лову единовременно находилось пять-шесть судов типа РТМ. Основными районами промысла в 1965 г. были прибрежные воды Пакистана, дававшие 44,3% общегодовой добычи в Индийском океане; значительную роль играли районы мыса Рас-Фартак (28,1%) и мыса Рас-Хафун (13,6%). В 1966 г. промысловое использование сырьевых ресурсов районов Индийского океана продолжало усиливаться, и единовременно на промысле находились в среднем около 10 судов типа РТМ. В этом же году был освоен новый район промысла - прибрежье Омана, - который дал наибольшую долю годовой добычи в Индийском океане (44,6%) и по своему значению опередил район Пакистана (42,9%) [Травин, 1968]. При этом доля ставриды в очень разнообразных уловах достигала 16,6%. С середины 1967 г. из-за закрытия Суэцкого канала советский траловый промысел в Индийском океане прекратился и возобновился только с середины 1969 г. В Аравийское море было направлено несколько траулеров "Дальрыбы", а к концу 1970г. их число возросло до десяти [Соловьев, Травин, 1970]. В течение этих лет практически использовались лишь районы Аравийского моря.
Промысловый район оманского прибрежья расположен в северной части Аравийского моря на шельфе юго-восточного берега Аравийского полуострова, между мысом Рас-Фартак и 60° в.д. Для этого района, как и для большинства районов Индийского океана, характерно большое разнообразие видового состава ихтиофауны, причем около 50 видов имеют промысловое значение, но ни один из них не является постоянно доминирующим. Как правило, уловы состоят из трех-четырех и более промысловых видов рыб. Ставриды здесь представлены несколькими видами, из которых наибольшее значение имеет Trachurus indicus.
Оманское стадо индийской ставриды более многочисленно, чем стада Персидского и Суэцкого заливов. Ставрида, обитающая в Суэцком заливе, не может иметь большого промыслового значения из-за мелких размеров и невысокой пищевой ценности. В Персидском заливе ставрида не образует мощных скоплений. Ставрида оманского стада значительно крупнее ставрид указанных выше двух стад, имеет большую ценность как сырье и по праву считается одной из основных промысловых рыб района оманского прибрежья, в особенности у о-ва Масира и в бухте Саукара.
В уловах советских промысловых флотилий, работавших у побережья Омана в 1966 г. и первом квартале 1967 г., на долю ставрид приходилось до 16,6% общей добычи рыбы, а в декабре - феврале в уловах некоторых траулеров они составляли от 60 до 90% уловов.
Учитывая большое промысловое значение Trachurus indicus, приводим некоторые техно-химические данные. По материалам лаборатории технологии ЮгНИРО, мясо этой рыбы содержит белка - 20% (19,1%-20,9%), жира - 6,0% (4,8%-7,1%), золы - 1,4%. Вареное мясо этой ставриды обладает нежной консистенцией, приятным вкусом с легкой кислинкой. Использовать индийскую ставриду из оманского района рекомендуется для приготовления консервов в масле и для копчения [Ковальчук, 1965, 1968; Яковлева, Ярославцева, 1967].