Trachurus mediterraneus (Steindachner, 1868)
Средиземноморская ставрида, черноморская ставрида (Rus),
Mediterranean horse mackerel
(Eng)
Синонимы:
Caranx trachurus mediterraneus Steindachner, 1868
Suareus furnestini Dardignac & Vincent, 1958
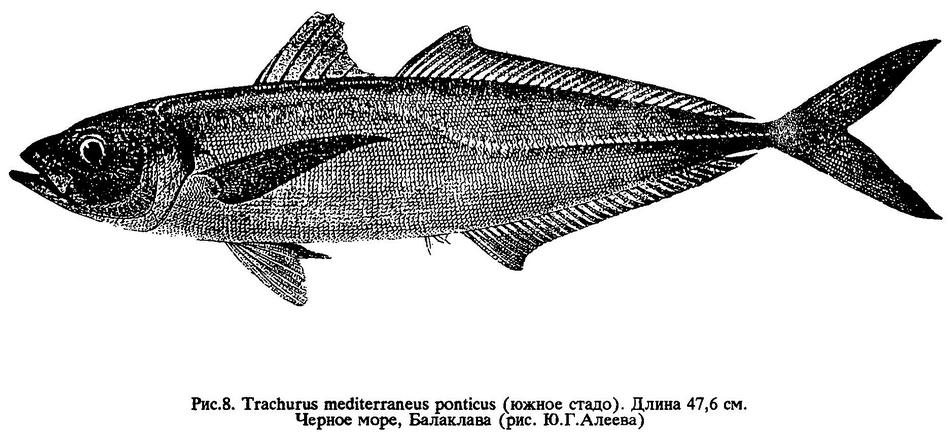
Trachurus mediterraneus ponticus Aleev, 1956 Черноморская ставрида (Rus),
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus mediterraneus (Steindachner) - средиземноморская ставрида
Описание. D I, VIII, I+26-34; А II, I+21-31; L.l. 75-95. Dll 5DI-6DII. Жаберных тычинок на первой дуге 45-61. Щитки боковой линии сравнительно мелкие; высота наиболее высоких щитков передней (кривой) части боковой линии 8,6-17,9% длины головы, высота наиболее высоких щитков задней (прямой) части боковой линии 11,9-19,2% той же длины. Кили на щитках развиты сравнительно слабо, шипы короткие. Прямая (задняя) часть боковой линии, несущая киль, сравнительно короткая; хорда дуги кривой части боковой линии 0,9-1,5 раза укладывается в длине ее прямой части. Тело сравнительно низкое, кругловатое; наибольшая его высота 16,8-24,0% длины тела до конца средних лучей хвостового плавника. Глаз небольшой, его диаметр 18,9-34,0% длины головы. Последние лучи второго спинного и анального плавников почти не отставлены от остальной части плавника; расстояние между основаниями последнего и предпоследнего лучей анального плавника в 1,1-1,3 раза превосходит расстояние между основаниями предпоследнего луча этого плавника и предшествующего ему (третьего сзади) луча. Грудные плавники сравнительно короткие, у взрослых не доходят до вертикали основания первого луча анального плавника или только слегка заходят за нее.
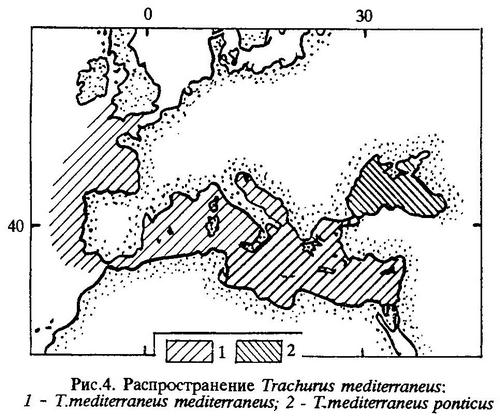
Распространен в Атлантическом океане у берегов Европы и в бассейне Средиземного моря (рис.4).
Вид представлен двумя подвидами: Trachurus mediterraneus mediterraneus (Steindachner) и Trachurus mediterraneus ponticus Aleev.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДВИДОВ Trachurus mediterraneus
1 (2). Высота наиболее высоких щитков передней (кривой) части боковой линии 12,4-17,9, в среднем 15,8% длины головы; высота наиболее высоких щитков задней (прямой) части боковой линии 14,7-19,2, в среднем 16,9% той же длины. Trachurus mediterraneus mediterraneus.
2 (1). Высота наиболее высоких щитков передней (кривой) части боковой линии 8,6-14,0, в среднем 11,0% длины головы; высота наиболее высоких щитков задней (прямой) части боковой линии 11,9-18,6, в среднем 14,8% той же длины. Trachurus mediterraneus ponticus.
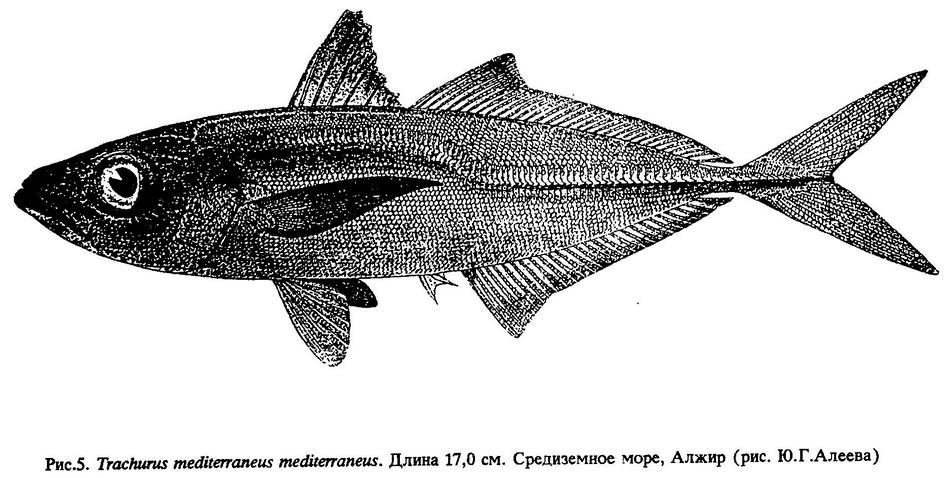
Trachurus mediterraneus mediterraneus - средиземноморская ставрида (рис.5.) в Атлантическом океане распространена у берегов Европы от Гибралтарского пролива до Бискайского залива включительно. В Средиземном море широко распространена и встречается как в его западных, так и в наиболее восточных районах, но в Мраморном, Черном и Азовском морях не отмечена. В пределах своего ареала образует несколько локальных стад, отличающихся не только некоторыми морфологическими признаками, но и по темпу роста. В Средиземном море является самой широко распространенной и многочисленной формой рода Trachurus.
Нерест средиземноморской ставриды наблюдается в мае-июле. Выметанные икринки имеют диаметр 1,00-1,04 мм. Молодь сопровождает медуз. Темп роста сравнительно высокий; годовики имеют длину около 11-13 см. В Адриатическом море двухгодовалые особи имеют длину около 15 см, трехгодовики - около 18 см, четырехгодовики - 20 см, восьмилетки достигают длины 27 см [Алеев, 1957а]. В южной части Бискайского залива были обнаружены экземпляры длиной до 46,4 см [Полонский, Байдалинов, 1964].
Пища состоит в основном из мелких рыб и ракообразных.
Средиземноморская ставрида имеет хозяйственное значение в странах Балканского полуострова, Алжире, Египте, Турции.
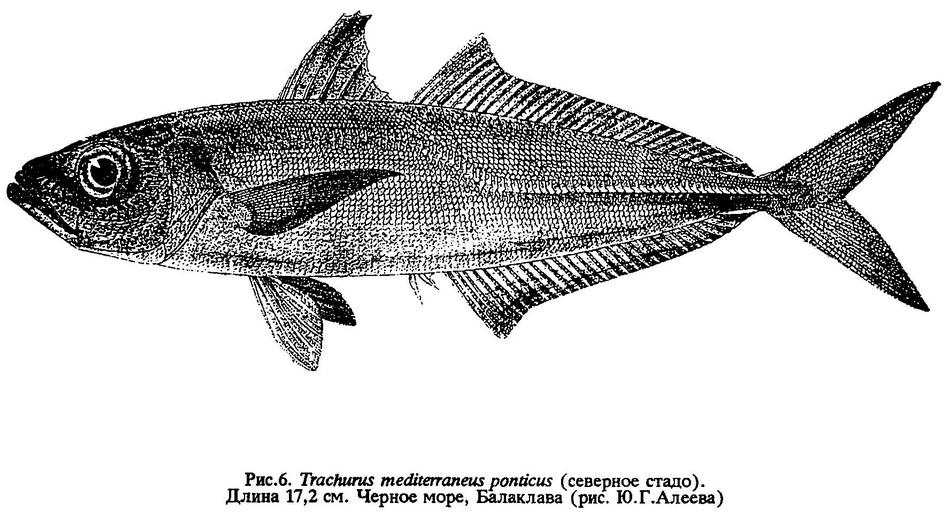
Trachurus mediterraneus ponticus - черноморская ставрида (рис.6) обитает в Мраморном, Черном морях, иногда заходит в южные и западные районы Азовского моря.
От типичной средиземноморской ставриды черноморская ставрида отличается меньшим числом ветвистых лучей в спинном и анальном плавниках, меньшей величиной щитков боковой линии и их несколько меньшей длиной в дополнительной спинной ветви боковой линии и некоторыми другими, менее существенными особенностями.
В годичном жизненном цикле черноморской ставриды можно различать два хорошо разграниченных периода: летний и зимний. Первый период продолжается с апреля по ноябрь-декабрь, т.е. около восьми месяцев; второй - все остальное время. Для летнего периода характерно широкое распространение в пределах ареала, пребывание в верхних, хорошо прогретых слоях водной толщи (в Черном море - от поверхности до глубины 15-30 м, над слоем температурного скачка), подвижный образ жизни, интенсивное питание, быстрый рост и размножение. Для зимнего периода характерно, напротив, относительно малоактивное состояние, пребывание у дна, на глубинах до 80-100 м, концентрация в определенных, относительно очень небольших по площади районах у берегов, слабое питание, почти или совершенно прекращающееся в наиболее холодные месяцы года (январь-март), и приостановка в росте. В Черном море ставрида переносит зимой охлаждение до 7°С.
В центральных халистатических областях Черного моря ставрида встречается редко; это - как правило, молодь. В недавнем прошлом ставрида северного и восточного стад заходила в Азовское море, где регулярно встречалась с июня по сентябрь у берегов Керченского полуострова, в Утлюкском лимане и в северной части Сиваша, составляя там в некоторые годы предмет промысла. В последние годы этого не наблюдается. В зимнее время область распространения значительно сужается. В этот период ставрида уходит из северных районов и открытых частей Черного моря и концентрируется в наиболее теплых прибрежных районах в непосредственной близости от берегов, на сравнительно глубоких местах с глубинами от 30-40 до 100 м. У ставриды южного стада ("крупной") в те годы, когда она заходит в северную часть Черного моря, сезонные миграции имеют большую протяженность, а следовательно, и большую продолжительность, чем у ставрид других трех стад.
Следует отметить, что места зимовок черноморской ставриды представляют собой, как правило, глубокие заливы или бухты, иногда окруженные обрывистыми, почти вертикально опускающимися на десятки метров под воду берегами (например, в крымском районе), или же понижения морского дна, так называемые "ямы"( например, в юго- восточном районе Черного моря). Течения на местах зимовок значительно (иногда примерно вдвое) слабее, чем у незащищенных берегов или на возвышенностях морского дна (между "ямами"). Географическое расположение районов зимовок определяется особенностями температурного режима моря; все районы зимовок приурочены к наиболее теплым областям: к берегам южного Крыма, Грузии и Малой Азии. Расположение мест концентрации ставриды внутри названных районов определяется прежде всего особенностями подводного рельефа, береговой линии и системы течений. Зависимость между течениями и особенностями распределения ставриды внутри района зимовок проявляется тем сильнее, чем суровее зима, как это видно на примере крымского района зимовок. В наиболее холодные зимы ставрида находится в особенно депрессивном состоянии, менее подвижна и собирается в наиболее спокойных местах южнокрымской акватории (к западу от мыса Сарыч), что и создает в такие зимы предпосылки для удачного лова ее здесь.
В холодные зимы скопления ставриды у южных берегов Крыма (у мыса Айя) достигают большой величины. Стремление уклониться от движения в суровые зимы особенно сильно выражено: оно тем сильнее, чем более охлаждена вода. Наоборот, в теплые зимы ставрида держится не только в бухтах или заливах, но и у открытых берегов, перед мысами, на местах с более значительным течением. В такие зимы мы находим ее по всему прибрежью южного Крыма, от мыса Фиолент до мыса Аюдаг; держится она более разреженно и не представляет такого удобного объекта для лова, как в годы массовых концентраций у мыса Айя, что заметно отражается на величине зимне-весенних уловов у берегов южного Крыма. На местах зимовки у берегов южного Крыма ставрида начинает скапливаться в декабре. Это - жирная рыба с неразвитыми половыми продуктами (яичники и семенники на стадии II). В декабре, январе и феврале ставрида всех возрастов распределена в районе зимовки более или менее равномерно; в это время она держится в придонном слое. С марта или с первой половины апреля, а иногда и значительно раньше ( с января) ставрида начинает совершать периодические подъемы в толщу воды. Подъемы эти первое время происходят только в ночные часы, а затем (к весне) и днем, после чего через некоторое время (во второй половине апреля - мае) вертикальные миграции прекращаются, и ставрида постоянно остается в поверхностных, более прогретых слоях водной толщи. Крупная ставрида (длиннее 13 см), быстрее созревающая, весной первая подходит ближе к берегам, на более мелкие места (иногда почти к самому урезу воды), попадая в зону действия ставных орудий лова. Становясь к весне более подвижной, она чаще попадает в ставные невода и чаще облавливается кошельковыми неводами. Напротив, более мелкая и медленнее созревающая рыба дольше пребывает в относительно малоактивном состоянии, вследствие чего в меньшем числе попадает в ставные невода и реже облавливается кошельковыми неводами. В течение второй половины апреля и в мае ставрида северного стада покидает места зимовок.
Половой зрелости самцы достигают в годовалом возрасте при длине от 8,5 см и более; самки - в годовалом-двухгодовалом возрасте при длине от 9,6 см и более. По достижении половой зрелости ставрида нерестует ежегодно [Алеев, 1957а]. По наблюдениям Т.Е.Сафьяновой и Н.И.Ревиной [1960], "крупная" ставрида созревает в возрасте трех-четырех лет. Созревание половых продуктов начинается в марте. В этот период ставрида переходит к более подвижному образу жизни, начинает более интенсивно питаться, причем интенсивность ее питания увеличивается с повышением температуры воды.
Нерест ставриды в Черном море происходит с мая до конца августа, преимущественно в июне (у берегов Кавказа) и июле (у северных берегов моря). Нерест "крупной" ставриды зачастую происходит и на удалении до 100 миль от берегов, что связано с общим характером ее летнего распределения.
Икрометание порционное, в течение нерестового сезона самки выметывают две порции икры. Вымет икры приурочен к вечерним, ночным и утренним часам; однако в пасмурную погоду икрометание продолжается иногда на протяжении большей части суток.
Плодовитость мелкой ставриды - до 150-200 тыс. икринок. Максимальное количество икринок имела самка длиной 42 см - 2 055 тыс.шт. [Сафьянова, Ревина, 1960]. Икра пелагическая; икринка с узким перивителлиновым пространством и большой жировой каплей.
Вертикальное распределение икры ставриды в Черном море ограничивается преимущественно верхним 5-метровым слоем, хотя вообще икра встречается до глубин 20-25 м; личинки также встречаются в основном в самом поверхностном слое воды.
Во время нереста зрелые самцы и самки, по-видимому, группируются на определенное время в отдельные стаи по половому признаку. Об этом свидетельствуют наблюдающиеся в период нереста случаи чрезвычайно резкого преобладания в уловах различных орудий лова зрелых рыб какого-либо одного пола.
Нерестовый косяк имеет "послойное" строение: в верхней его части концентрируются самцы, в нижней - самки; выметываемая самками икра, всплывая, пронизывает слой молок, во время чего и происходит оплодотворение [Алеев, 1957а]. Развивающаяся икра ставриды встречается при температуре воды 15-26°С. Между температурой воды и количеством живой икры ставриды существует прямая зависимость. Наилучшая выживаемость икры отмечена при температуре воды от 19° до 23°С [Сафьянова, Ревина, 1960]. Икра развивается в воде соленостью от 13,8‰ и выше [Дехник и др., 1950]. Икрометание может происходить, однако, и при несколько более низкой солености (12-13‰), о чем свидетельствует установленный факт нереста этой формы в Арабатском заливе Азовского моря [Алеев, 1957а].
Эмбриональный период развития черноморской ставриды весьма непродолжительный. Икра на стадии дробления появляется в планктоне после 19 ч; в 23-24 ч преобладает уже икра на стадии морулы, между 1-4 ч ночи - на стадии гаструлы. При температуры воды 20-22°С эмбриональный период длится около полутора суток. Выметанная (набухшая) икринка имеет диаметр 0,76-1,2 мм, диаметр жировой капли составляет 0,21-0,29 мм. Только что вылупившийся из икринки эмбрион имеет длину около 1,75 мм, а вместе с выступающим вперед желточным мешком - около 2 мм. Вдоль спины предличинки располагаются два ряда меланофоров. Первые два дня предличинка плавает спиной вниз, на третьи сутки переворачивается спиной вверх и становится более активной; желточный мешок к этому времени значительно уменьшается. Для личинок длиной 3,5-9,5 мм характерно наличие двойного ряда шипов на предкрышке и пигментных полосок из одного ряда звездчатых меланофоров на средней линии тела и близ оснований спинного и анального плавников. Превращение личинки в малька заканчивается при длине около 10 мм; образование характерных для ставрид щитков на боковой линии начинается при длине 15-17 мм.
В местах нереста ставриды нагуливается и ее молодь. Мальки длиной 1-9 см сопровождают медузу Rhizostoma pulmo, скрываясь в момент опасности под ее колоколом. Сожительство мальков с медузами продолжается вплоть до декабря, т.е. до периода концентрации ставриды, включая и сеголетков, на местах зимовки. Благодаря значительной растянутости периода икрометания осенью (в ноябре-декабре) сеголетки имеют весьма различную длину - от 6 до 11 см.
Пища взрослых особей состоит, главным образом, из мелкой рыбы и некоторых ракообразных. Из рыб наиболее существенное значение в питании имеют черноморская хамса Engraulis encrasicolus ponticus, шпрот Sprattus sprattus phalericus, тюлька Clupeonella delicatula delicatula некоторые бычки {Pomatoschistus, Knipovitschia, Aphia), атерина Atherina mochon pontica, песчанка Ammodytes cicerelius, из ракообразных - мизиды (Mesomysis sp., Macromysis sp. и др.) и креветки (Leander sp., Crangon sp.). Кроме того, в пище присутствуют Amphipoda (Gammaridae), Isopoda (Idotea sp.), Polichaeta и другие беспозвоночные, а также пелагические мальки разных видов рыб, в частности, барабули Mullus barbatus ponticus, кефалей (Mugil cephalus, Liza spp.), ошибня Ophidion barbatum, различных сельдевых (Clupeidae) и др. Удельный вес этих компонентов в пище ставриды в различных районах моря в разные месяцы года бывает неодинаковым, в одних случаях преобладают ракообразные, в других - рыба. Ракообразные в питании мелкой ставриды играют более значительную роль, чем в питании крупной ставриды южного стада; последняя во взрослом состоянии питается почти исключительно рыбой: хамсой, шпротом, атериной, пелагической молодью барабули и др. В пище ставриды северного и восточного стад рыба в особенно больших количествах присутствует во второй половине лета и осенью, когда у берегов появляется много молоди хамсы, шпрота, тюльки и т.д. Мальки длиной до 2-4 см питаются планктоном (Sagitta, Acartia, Centropages и др.).
В течение суток наблюдаются два периода наиболее интенсивного питания: утром, после восхода солнца, и вечером, перед его заходом; с наступлением темноты ставрида перестает захватывать пищу. С наибольшей интенсивностью питание происходит в июле и августе, когда температура воды в море достигает максимума, т.е. 22-25°С; этот же период является и периодом наиболее быстрого роста. Менее интенсивны питание и рост весной и осенью. Зимой, при температуре воды ниже 10°С, питание почти полностью прекращается. В северных районах Черного моря, где температура воды особенно низка, ставрида в наиболее холодные месяцы года (январь-март) практически перестает питаться. Сезонные изменения интенсивности питания определяют в основном сезонные изменения жирности. Так, если в сентябре у ставриды северного стада содержание жира в теле составляет 12-13%, то к маю оно уменьшается до 6-7%; в июне-июле, несмотря на увеличение интенсивности питания, жирность остается на низком уровне - около 2-3%, что связано с расходованием большого количества энергии на построение половых продуктов. Увеличение жирности начинается после окончания массового нереста, с конца июля - начала августа. Более крупные рыбы всегда жирнее мелких [Алеев, 1957а]. Наиболее высокой жирностью отличается крупная ставрида южного стада.
В пищевых цепях Черного моря ставрида может занимать разные места. Взрослая ставрида, особенно "крупная" южная, во многих случаях является окончательным звеном пищевой цепи. С другой стороны, мелкая ставрида представляет собой связующее звено между беспозвоночными и мелкими рыбами, которыми она питается, и более крупными пелагическими хищниками, для которых она служит добычей: пеламидой Sarda sarda, крупным луфарем Pomatomus saltatrix, тунцом Thunnus thynnus и дельфинами Delphinus delphis. В то же время ставрида является конкурентом этих крупных хищников, так как все они питаются главным образом не ставридой, а именно той мелкой рыбой, которая составляет пищу ставриды: хамсой и шпротом. Конкурентом ставриды в питании в Черном море, преимущественно в его западной половине, является также скумбрия Scomber scombrus.
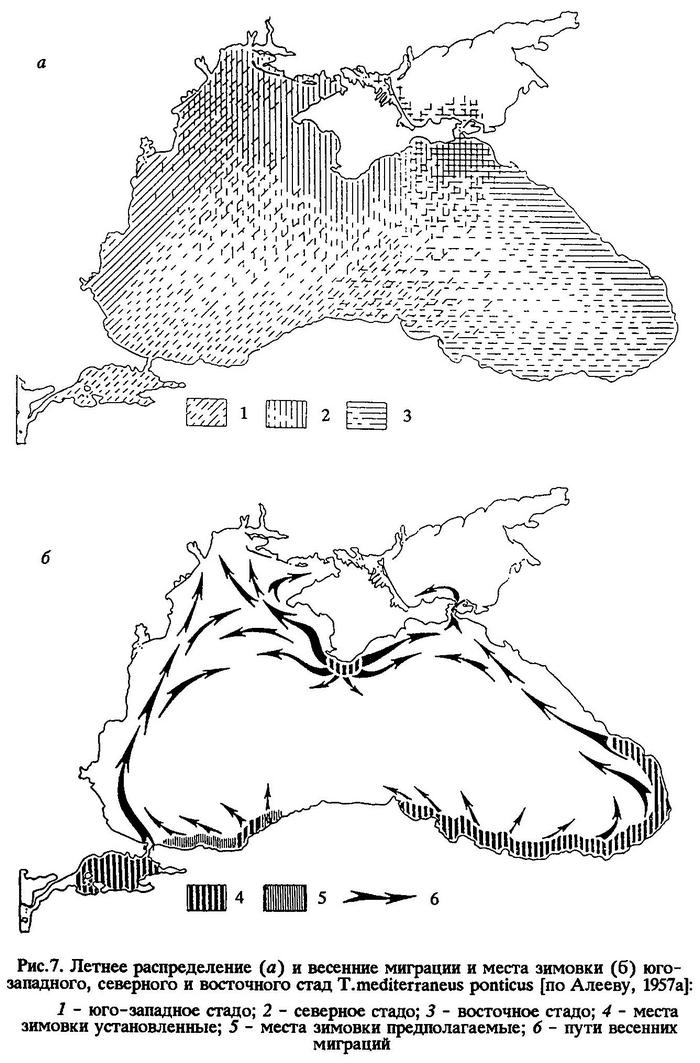
В пределах своего ареала черноморская ставрида представлена четырьмя локальными стадами: юго-западным (босфорским), северным (крымским), восточным (кавказским) и южным (анатолийским) (рис.7 и 9).
Стада эти, кроме географической локализации мест зимовки, нагула и нереста, существенно отличаются одно от другого возрастным составом; ставрида разных стад имеет различный темп роста, разный характер питания, разные сроки нереста и т.д.
Юго-западное стадо распределяется в Мраморном море, Босфоре, а также в юго-западном и западном районах Черного моря. Ставрида этого стада летом встречается в Мраморном море и Босфоре, у северо-западных берегов Малой Азии, у берегов Болгарии и Румынии. Нерест происходит на всей этой площади, однако в Мраморном море и в прибосфорской области летом остается только незначительная часть стада, основная же его масса отходит севернее, к берегам Болгарии и Румынии. В придунайском районе ставрида юго-западного стада летом частью смешивается со ставридой крымского стада. В румынских водах зимой она отсутствует с декабря по апрель, в болгарских - с января по март.
Места зимовки расположены в основном в Мраморном море, в связи с чем ставрида этого стада дважды в году мигрирует через Босфор, осенью (в сентябре-ноябре) уходя из Черного моря в Мраморное и возвращаясь в него весной (в мае-июне). Частью места зимовки этого стада располагаются и в прибосфорском районе Черного моря у северозападных берегов Малой Азии, в частности в районе Зон- гулдак и Эрегли [Алеев, 1957а].
Ставрида юго-западного стада достигает длины 30 см в восьмилетнем возрасте. Рост этой ставриды представлен в табл.3. В последние годы в промысловых скоплениях этого стада обычно наблюдаются лишь первые три возрастные группы.

Северное (крымское) стадо занимает летом северозападную часть Черного моря и воды Крыма вплоть до Керченского предпроливного пространства. Нерест наблюдается в пределах всего района. Когда Азовское море было экологически чистым, ставрида этого стада заходила в него и встречалась, главным образом, у северных берегов Керченского полуострова.
Места зимовки ставриды крымского стада располагаются у берегов южного Крыма, от мыса Фиолент до мыса Аюдаг. Эта ставрида отличается наименьшей продолжительностью жизни и самым медленным ростом; максимальный установленный возраст - 5 лет. Основную массу стада обычно составляют три первые возрастные группы. Длина не более 20 см, очень редко - до 25 см, масса 95-100 г (см.рис.6). Рост ставриды северного стада показан в табл.4.

В придунайском районе Черного моря летом происходит частичное смешение северного стада с юго-западным, а в районе Керченского предпроливья - с восточным.
Восточное (кавказское) стадо распределено в восточной части Черного моря - у берегов Кавказа и северо-восточных берегов Малой Азии. Летом основная масса ставриды этого стада держится у берегов Кавказа, где и происходит нерест. Места зимовки располагаются у юго-восточных берегов Черного моря, к югу от мыса Пицунда; на запад район зимовок простирается до Синопа.
По темпу роста ставрида восточного стада почти не отличается от ставриды северного стада. Попадаются экземпляры в возрасте до 6 лет при длине до 26 см, обычно наиболее многочисленны особи в возрасте до 4 лет. Темп роста ставриды восточного стада показан в табл.5.

Ставрида восточного стада нерестится в массе в июне, тогда как ставрида северного стада (крымского) - в июле. Разница в сроках нереста объясняется тем, что ставрида восточного стада, зимующая в юго-восточном районе Черного моря при несколько более высоких температурах воды, находится зимой в менее депрессивном состоянии, чем та, которая проводит зиму у крымских берегов. Благодаря этому ставрида восточного стада по сравнению с крымской весной созревает в массе быстрее.
Летнее распределение юго-западного, северного и восточного стад устанавливается с июня и сохраняется до конца сентября (см.рис.7), зимнее - с конца декабря до апреля; промежуточные периоды можно назвать периодами осенних (с октября по декабрь) и весенних (с апреля по конец мая) миграций.
Нерестилища юго-западного, северного и восточного стад располагаются в основном в области общего кольцевого течения моря и в прибрежных районах.
Экологически выделяется ставрида южного стада, которое по сравнению с прочими тремя занимает явно обособленное положение. Рыбы этого стада резко отличаются от рыб других стад значительно более высоким темпом роста и более крупными размерами, за что получили название "крупная" ставрида (рис.8). Им также свойственна большая продолжительность жизненного цикла.
Ставрида южного стада совершает более протяженные миграции и значительно меньше привязана к шельфу. "Крупная" ставрида представляет форму, менее привязанную к шельфу, чем рыбы остальных трех стад, которые больше тяготеют к прибрежью и редко встречаются за пределами 50-мильной прибрежной полосы. Весной и летом она встречается как у берегов, так и в удалении от них до 70-100 миль и более. Это - хищник, тогда как ставриды других стад имеют в основном смешанное питание. "Крупная" ставрида наиболее теплолюбивая, не зимует у северных берегов Черного моря, а в юго-восточном районе держится зимой в самых теплых местах. Ареал ее в значительной степени наложен на ареалы других трех стад, чего нельзя сказать об ареалах последних, которые в целом не перекрываются. Все это дает основание полагать, что ставрида южного стада представляет собой хорошо выраженную экологическую расу. Некоторые исследователи [Водяницкий, 1974] считают, что "крупная" черноморская ставрида в годы второй мировой войны в большом количестве проникла из Средиземного моря в Черное. Здесь она нашла благоприятные условия, в частности, обильную пищу в виде мелких пелагических рыб, в результате чего произошла вспышка ее численности и ускорился рост особей. В качестве подтверждения этой гипотезы приводится пример кратковременной вспышки развития в Черном море дальневосточного вселенца-моллюска рапаны, который был завезен в Новороссийскую бухту в предвоенные годы. В дальнейшем этот моллюск размножился и распространился по Черному морю. Размеры его достигали 15 см, но постепенно он стал мельчать, и теперь такие экземпляры очень редки. Так и "крупная" черноморская ставрида, по мнению этих авторов, в дальнейшем постепенно мельчала. Часть этого стада попала в сети, часть вымерла, а потомство превратилось в знакомую нам черноморскую ставриду [Водяницкий, 1974].
Большинство авторов, однако, считает, что "крупная" черноморская ставрида всегда обитала в Черном море. В больших количествах она была обнаружена сразу после войны, когда рыбная разведка в Керчи получила самолеты для поиска рыбы и наводки на нее промысловых судов. С воздуха были обнаружены плотные гигантские косяки рыбы на большом расстоянии от кавказского побережья. Это и послужило основанием для организации промысла "крупной" ставриды. Позднее, в 1966 г., в Стамбуле на сессии Международного совета по изучению Средиземного моря наши ученые убедились в том, что "крупная" ставрида никогда не проходила через Босфор в Черное море, более того, она вообще неизвестна в Средиземном море [Шульман, 1979].
Наши исследования [Некрасов, 19666] показали, что "крупная" черноморская ставрида отличается от средиземноморской на уровне подвида. В то же время как по внешнему виду, так и по внутреннему строению "крупная" ставрида - точная увеличенная копия мелкой ставриды.
Южное стадо обитает в южной части ареала Trachurus mediterranreus ponticus - в анатолийских водах Черного моря и прибосфорском районе. Здесь южная ставрида была известна издавна - ее давно знали у берегов Болгарии [Максимов, 1914] и у берегов Кавказа [Зернов, 1913]. "Крупная" ставрида хорошо знакома аджарским рыбакам.
Зимовка "крупной" ставриды происходит в самом теплом районе Черного моря, у его юго-восточных берегов: по восточному берегу - к югу от мыса Кодор (хотя севернее Батуми зимует редко), по анатолийскому берегу - на запад, видимо, не далее мыса Киремит. В некоторые годы часть южного стада ставриды уходит на зимовку в Мраморное море и распространяется там вплоть до Дарданелл. Этот район зимовки не является для южного стада обычным, и здесь встречаются только отдельные экземпляры и далеко не ежегодно.
Весной, в мае, "крупная" ставрида распространяется из юго-восточного района моря (с мест зимовки) на запад, до прибосфорского района и берегов Болгарии. В это же время другая часть южного стада ставриды мигрирует на северо-восток, появляясь у аджарских берегов (рис.9).
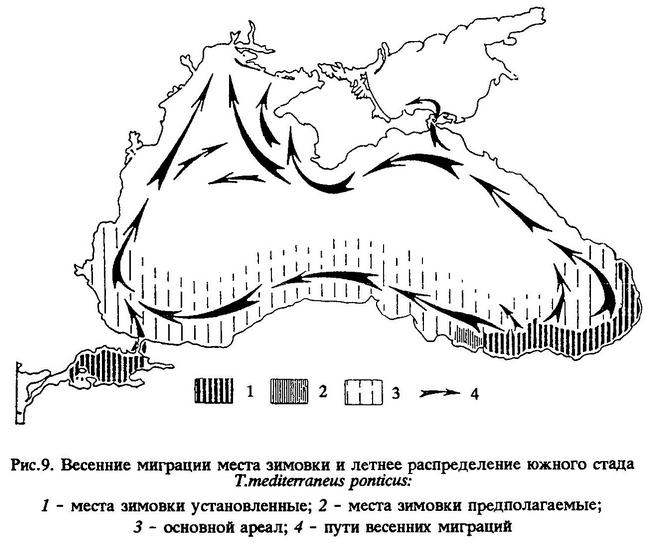
В годы значительного увеличения численности "крупная" ставрида распространялась летом значительно шире и держалась тогда не только в южных районах Черного моря, но проникала и к его северным берегам (как это наблюдалось, например, в 1950-1964 гг.). В такие годы из юго-восточного района моря она шла весной вдоль анатолийских берегов не только на запад, до прибосфорского района, откуда распространялась далее на север к берегам Болгарии и Румынии, в некоторых случаях доходя до Одессы, но вначале и на северо-восток, а затем вдоль берегов Кавказа на север до Керченского пролива и Крыма, а иногда еще дальше, вплоть до Азовского моря (у северного берега Керченского полуострова, к западу до Арабатского залива), Каркинитского залива и Тендры. В годы такого широкого распространения "крупной" ставриды нерест ее в июне-июле наблюдался как у анатолийских берегов, берегов Болгарии, Румынии, Крыма и Кавказа, так и далеко от берегов в открытом море.
Весной "крупная" ставрида появляется раньше всего у берегов Грузии - в апреле или первой половине мая, когда температура поверхностного слоя воды повышается до 9°С. В конце мая-начале июня она наблюдается иногда против Сочи и Туапсе, в удалении от берегов на 50-100 миль. Присутствие ее в этих удаленных от берегов районах связано, скорее всего, с наличием в них в указанный период года мелких пелагических рыб - хамсы и шпрота, которые составляют ее основную пищу. В течение июня наблюдается продвижение на север, в район мыса Меганом-Новороссийск; в июле "крупная" ставрида появляется у южных берегов Крыма. В северных районах Черного моря и в Азовском море (куда заходит редко) она остается в разные годы до сентября-октября включительно, затем исчезает до следующей весны; в водах южного Кавказа в октябре-ноябре хорошо прослеживается осенняя миграция. У берегов Болгарии "крупная" ставрида ловится в мае и июне, во время весенней миграции на север, и осенью, во время обратной миграции на юг.
"Крупная" ставрида (южного стада) является рыбой с длительным жизненным циклом и достигает возраста 14 лет при длине до 55 см [Ревина и др., 1960]. Темп ее роста выше, чем ставрид всех других стад Trachurus mediterraneus ponticus (табл.6).

По данным Т.Е.Сафьяновой и Н.И.Ревиной [1960] темп роста "крупной" ставриды несколько выше (табл.7).
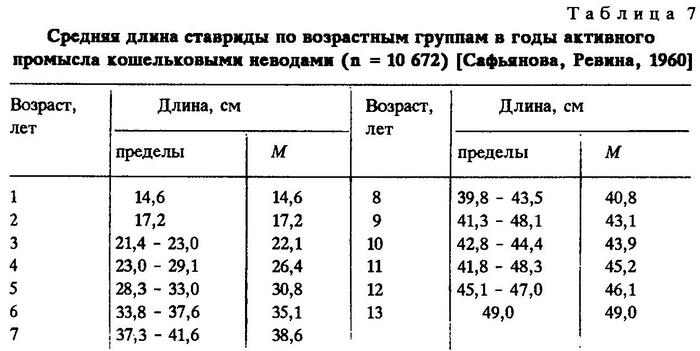
Наибольший прирост длины у "крупной" ставриды, в среднем 10,1 см, наблюдается в первый год жизни. В 2-7-летнем возрасте годовые приросты составляют около 4 см. На 8-9-м году жизни резко снижается линейный, а в возрасте 10 лет - и весовой прирост. В дальнейшем длина и масса "крупной " ставриды увеличивается незначительно, что свидетельствует о наступлении предельного возраста этой рыбы [Сафьянова, Ревина, 1960].
С начала развития активного промысла "крупной" ставриды был собран материал по ее размерному составу в промысловых уловах (табл.8).

В первые два года интенсивного промысла средний размер ставриды был наименьшим. В дальнейшем количество мелкой рыбы в уловах резко уменьшилось, и средняя длина рыб значительно увеличилась. В 1958 г. рыбы длиной до 30 см в среднем составляли всего лишь 0,9% улова, в 1957 г. - 1%, тогда как в 1956 г. их доля в годовом улове достигала 21,7%, в 1955 г.- 47,5% и в 1954 г.- 46,7%.
Резкое уменьшение в уловах кошельковых неводов мелкой рыбы объясняется отсутствием должного пополнения промыслового стада ставриды молодью. Все эти годы промысел в основном базировался на вылове рыб урожайных поколений 1946-1949 гг.
Максимальный вылов "крупной" ставриды кошельковыми неводами был в 1956 г. В 1957 и 1958 гг. годовой улов ставриды и средний улов ее на судно заметно снизился.
Анализ данных по возрасту и размерному составу уловов, а также показатели промысла свидетельствовали о постоянном уменьшении запаса "крупной" ставриды. В последующие годы численность старших поколений еще более сократилась. После 1967 г. промысел "крупной" черноморской ставриды утратил свое значение. В настоящее время "крупная" ставрида встречается единично.
Хозяйственное значение. В последние годы в Азовском и Черном морях резко ухудшилась экологическая обстановка. Это связано не только с последствиями предшествующего зарегулирования стока рек, безвозвратного забора воды, с изменениями солевого состава речных вод под влиянием водохранилищ, но и с резким возрастанием загрязнения от сточных вод, в которых в большом количестве содержатся органические формы фосфора, азота, пестициды, тяжелые металлы, нефтепродукты и т.д. Уровень загрязнения в этих морях во многих местах сейчас превышает ПДК в 2-5, а иногда и в десятки раз. В результате произошло эвтрофирование прибрежных вод, существенно увеличилась их токсичность. Так, на Азовском море в последние 4 года летом регулярно наблюдается массовая смертность рыб. К бедам от загрязнения добавились экологические последствия появления в Черном и Азовском морях гребневика - мнемиопсиса, завезенного зерновозами из прибрежных вод США. В Азовском море им с 1989 г. уничтожается большая часть кормовых ресурсов, что обрекает рыб на полуголодное существование [Зиланов и др., 1992]. Гребневик летом почти полностью выедает корм пелагических промысловых рыб, а также их икру и личинок. Так, например, по данным ЮгНИРО, в 70-х гг. в Черном море летом средняя биомасса кормового зоопланктона составляла 113 мг/м3, а в 1992 г. - 8-10 мг/м3. В последние три-четыре года в связи со вспышкой численности гребневика снизились уловы черноморской хамсы в 30 раз, почти на нет свелся промысел азовской хамсы и тюльки. Подобная катастрофа коснулась и Турции.
Короче говоря, произошел естественный процесс. Сработал общебиологический закон: адаптация вселенца всегда сопровождается вспышкой его численности в первые годы освоения нового ареала. Но согласно этому же закону численность вселенца в последующие годы непременно должна значительно снижаться, что и наблюдается в настоящее время, благодаря чему заметно увеличилась биомасса черноморской хамсы.
В последние годы резко падает и численность ставриды на зимовальных скоплениях. Последние мощные скопления на зимовках наблюдались в наших водах в 1984-1985 гг. В настоящее время в традиционных местах зимовок ставрида встречается единично. Причем вначале пропали промысловые скопления ставриды в наших водах, а в 1991 г. - и в водах Турции.
О депрессивном состоянии черноморской ставриды говорят и ее биологические показатели. По устному сообщению заведующего лабораторией ЮгНИРО А.Н.Михайлюка, наблюдается значительное снижение среднего возраста, отмечается начало нереста в возрасте 1 года.
Следует особо остановиться на проблеме "крупной" черноморской ставриды.
В природе хорошо известны массовые вспышки численности видов растений и животных. Они связаны с колебаниями климатических факторов, создающих очень благоприятные условия размножения, роста и нагула. Эти условия обычно повторяются с определенными интервалами времени. В последние десятилетия у биологов сложилось мнение, что чаще всего эти колебания связаны с 11-летней периодичностью солнечной. активности. Примером таких вспышек являются массовые нашествия саранчи, распространение грызунов и т.д. "Крупная" черноморская ставрида дала вспышку численности, известную человечеству только один раз. Весь промысел "крупной" ставриды в 50-х - начале 60-х гг. базировался на одном высокоурожайном поколении конца 40-х гг. [Шульман, 1979]. К сожалению, 11-летней цикличности вспышки численности "крупной" ставриды не наблюдается. Сейчас лишь изредка попадаются в уловах отдельные ее экземпляры. Так, в 1986-1987 гг. "крупную" черноморскую ставриду обнаруживали у берегов Крыма. В 1981 г. у берегов Грузии судно ЮгНИРО обловило тралом косячок "крупной" ставриды средней длиной тела 30 см.
В 1992 г. В.В.Кузнецов сделал доклад на симпозиуме в Канаде, в котором высказал гипотезу о цикличности колебаний численности животных не только с периодичностью 11 лет, но и 95, 115 лет. Меридиональные переносы тепла при повышении солнечной активности примерно с вековой периодичностью снижают даже численность людей. С такой периодичностью, в частности, изменяется численность сардины-иваси. Сейчас численность сардины-иваси резко сокращается. Но интересно отметить, что в противофазе с численностью сардины-иваси находится численность японской ставриды, численность которой в последние годы заметно увеличивается.
Почему бы нам не предположить, что колебание численности "крупной" черноморской ставриды также подчиняется закономерности с периодичностью в 95 или 115 лет?
В связи с изложенным о хозяйственном значении черноморской ставриды сегодня приходится говорить, к сожалению, в основном в прошедшем времени.
Численность черноморской ставриды подвержена значительным межгодовым колебаниям (флюктуациям). Еще раньше по различным оценкам, в разные годы величина ее запаса изменялась от 75 до 800 тыс. т.
Основная масса улова Украины и России обеспечивалась промыслом северного стада черноморской ставриды. Меньший улов обеспечивался за счет восточного стада. Юго-западное стадо играло значительную роль в промысле Румынии и Болгарии.
Промысел ставриды северного и восточного стад в основном велся в зимне-весеннее время на местах зимовок.
У берегов Крыма плотные зимовальные скопления начинали образовываться во второй половине, реже - в середине ноября. Однако массовыми они становились лишь в конце ноября или даже в декабре при температуре воды около 12°С, и в это время создавалась стабильная обстановка промысла. У берегов Кавказа стабилизация скоплений происходила при такой же температуре несколько позже, как правило, в конце декабря. Промысел черноморской ставриды зимой велся конусными сетями с привлечением рыб на электросвет. Вылов ее кошельковыми неводами в этот период был крайне незначителен.
Наилучшими условиями для промысла черноморской ставриды являются температура воды ниже 10°С и маловетреная погода. Замечено, что с понижением температуры воды реакция на свет у ставриды, а следовательно, и ее вылова конусной сетью увеличивается. Есть основания полагать, что некоторое влияние на эту реакцию оказывает чередование фаз луны, вызывая ее ухудшение при полнолунии, особенно если оно сопровождается безоблачной погодой.
В марте, с распадом скоплений и увеличением подвижности черноморской ставриды, вылов конусными сетями снижался. Тем не менее вылов ставриды в этом месяце мог быть даже выше,чем в январе-феврале. Объясняется это увеличением числа добывающих судов, которые обычно переводились с добычи хамсы при ухудшении ее промысла в марте. Лов черноморской ставриды конусными сетями прекращался в марте-апреле. С прогревом воды и выходом рыбы на меньшие глубины становился возможным ее лов кошельковыми неводами. Лов велся, как правило, у побережья Грузии в апреле-мае. Осенью тоже можно было ловить мигрирующую черноморскую ставриду кошельковыми неводами. Лов велся в октябре-декабре у побережья Грузии и в значительно меньшей степени у берегов Северного Кавказа и Крыма. При высокой величине запаса отдельные уловы кошельковыми неводами у берегов Грузии бывали в конце августа и в сентябре.
С апреля по октябрь черноморская ставрида добывалась также ставными неводами. Летний лов ставными неводами был приурочен в основном к северному району Черного моря; более низкие уловы были обычно в летнее время у крымских и кавказских берегов и еще более низкие - в Керченском проливе и в южной части Азовского моря. Доля вылова ставными неводами в целом была невелика.
В 1936-1939 гг. уловы ставриды в Румынии составили около 0,6-2,0 тыс.ц, в Болгарии - 1,1-2,8 тыс.ц, в Турции - 3,9-5,9 тыс.ц; уловы СССР в тот же период составляли 0,6-7,0 тыс. ц [Алеев, 1957а].
С 1947-1948 гг. в Черном море появилась в промысловых количествах "крупная" ставрида.
В 1947-1953 гг. наиболее обычные уловы ставриды в советских водах Черного моря составляли около 16-20 тыс.ц, уловы отдельных лет (1950) доходили до 60 тыс.ц. В 1954 г. благодаря частичному освоению запасов южного стада улов черноморской ставриды резко увеличился и составил около 110 тыс.ц (табл.9).

В первые годы массового появления "крупной" ставриды в наших водах не было организовано специального лова ее. Ставриду в небольшом количестве вылавливали в прибрежной зоне ставными неводами, поэтому уловы ее в то время составляли несколько сот центнеров в год.
В 1953 г. на промысле "крупной" ставриды начали применять кошельковые невода, с помощью которых производили облов косяков как в поверхностных слоях, так и в толще воды, и не только в прибрежной зоне, но и в открытом море. К концу 1953 г. промыслом "крупной" ставриды занимались 25 судов, вооруженных капроновыми кошельковыми неводами. Лучшие результаты были получены при облове косяков кошельковыми неводами больших размеров (600 м и более) с механизированной выборкой.
Быстрому развитию активного промысла ставриды способствовали организация разведки ее скоплений с помощью авианаблюдения и проведения эхолотной съемки с поисковых судов. Судовая и авиационная разведка не только обнаруживала косяки ставриды, но и наводила на них промысловые бригады.
С 1954 до 1967 г. весной, летом и осенью у восточных и северо-восточных берегов моря существовал интенсивный кошельковый промысел.
Ныне ставрида южного стада ("крупная" ставрида) утратила промысловое значение.
Сравнительные замечания. В 1966 г. мною была описана индийская ставрида, обитающая в Аравийском море, как подвид Trachurus mediterraneus indicus. Основой для выделения этой ставриды лишь на уровне подвида послужило отсутствие хиатуса в диагностических признаках Trachurus mediterraneus mediterraneus и Trachurus mediterraneus indicus. Позднее в литературе появились работы, справедливо подвергающие сомнению наше прежнее утверждение о том, что индийская ставрида является подвидом средиземноморского вида [Rass, 1973]. Дальнейшие исследования показали, что указанные филогенетически близкие формы различаются на уровне вида, хотя граница между ними не очень четкая. Ф.Г.Берри и Л.Коэн (1972) возвели описанную мною форму в ранг вида Trachurus indicus Necrasov. Мне остается только присоединиться к их мнению и добавить, что более детальное сравнительное изучение Trachurus indicus, добытых мною в Красном и Аравийском морях и Персидском заливе, и Trachurus mediterraneus mediterraneus, пойманных нами в Средиземном море у Александрии, выявило различия не только по высоте щитков в передней части боковой линии, по величине отношения хорды дуги кривой части боковой линии к длине тела и по длине грудных плавников, на что я указывал ранее [Некрасов, 1966а], но и по числу щитков в боковой линии, по отношению длины прямой части боковой линии к длине хорды дуги кривой, по длине спинной ветви боковой линии, по расстоянию от вершины рыла до вертикали начала основания анального плавника и некоторым другим признакам (табл.18).
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Ставрида черноморская — Trachurus mediterraneus ponticus (Aleev, 1956)
Боковая линия на всем протяжении прикрыта костными щитками. Рот конечный, большой. D1 VII—IX. D2 I 26—34. А II, I 21—31. Хищная пелагическая рыба. Длина тела не превышает 40 см (обычно мельче). Обитает по всей акватории Черного моря. Промысловый объект.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
ЧЕРНОМОРСКАЯ СТАВРИДА — TRACHURUS MEDITERRANEUS PONTICUS ALЕЕV
Условия размножения. Нерест этого вида в разных районах ареала происходит в основном в наиболее теплое время года. В Черном море ставрида нерестится с июня по август (Дехник и Павловская, 1950; Павловская, 1954; Ткачева, 1957; Ревина, 1958; Георгиев и др., 1960, 1961, 1962), разгар икрометания приходится на июль (Дехник и Павловская, 1950).
По данным Арима (Arim, 1957), икринки и личинки этого вида встречаются в районе Тробзона со второй половины июня до конца июля; в августе ловится молодь под куполами медуз.
Период размножения Trachurus mediterraneus в Средиземном море точно не установлен. У Санцо (1932 г) есть единственное указание на нахождение икринок этого вида в районе Мессины в июне. По нашим наблюдениям, икринки Trachurus mediterraneus (?) встречались только в летнее время — с июня по август, наиболее часто в июле (Дехник, Синюкова, 1964).
В период размножения ставрида обитает преимущественно в прибрежных районах. В Черном море икринки и личинки встречаются вдоль всего побережья, в основном в пределах 10—25-мильной зоны (Дехник и Павловская, 1950). В годы резкого увеличения численности крупной формы ставриды она нерестилась на большой площади Черного моря — до 80 и более миль от берега (Ревина, 1958).
Проведенные нами наблюдения свидетельствуют о том, что численность икринок и личинок ставриды уменьшается от берегов к открытому морю. Заметные различия выявляются из сопоставления прибрежного 10-мильного района (1960—1962 гг.) с открытым морем (1963, 1965 гг.) (табл. 21). Данные Р. М. Павловской (1954) также свидетельствуют о тяготении этого вида в период размножения к прибрежным районам.
В Мраморном море икринки и личинки ставриды встречаются над глубинами 20—350 м (Demir, 1958а, 19586), как и в Черном море в основном в прибрежной области. В то же время наблюдения Н. И. Ревиной (1958) показали, что в отдельные годы значительные концентрации икринок и личинок могут быть обнаружены в открытом море.
В пределах своего ареала ставрида нерестится при значительных колебаниях температуры и солености. Нормально развивающиеся икринки ставриды были найдены нами в Азовском море (в районе Казантипского маяка) при температуре 24,4° и солености 11,55‰. В Черном море развивающиеся икринки и личинки ставриды встречаются при температуре воды 15,1—25,9°. Ю. П. Зайцев (1959а) указывает на нахождение икринок ставриды при температуре 11,5°. Соленость в местах нахождения икринок и личинок в Черном море колеблется от 13,8 до 19,3‰. В Средиземном море они развиваются при солености 37,5—39,1‰ и температуре 15—26°.
Приспособление этого вида в период раннего онтогенеза к значительным колебаниям температуры и солености — одна из важнейших причин, определивших широкое распространение и высокую численность локальных стад в условиях Черного моря.
Суточный ритм размножения. Свежевыметанные икринки ставриды встречаются в планктоне между 16 и 24 ч. Как установлено для многих других видов, в планктоне одновременно развиваются икринки двух суточных порций — ближайшего суточного вымета и вымета предшествующих суток. В определенный отрезок времени, когда инкубационный период икринок вымета предшествующих суток закончился и произошел выклев эмбрионов, в планктоне находятся икринки одной суточной порции — вымета ближайших суток.
Распределение икринок ставриды по этапам развития в течение суток может быть отчетливо прослежено по поверхностным горизонтальным уловам ихтиопланктонной сетки в разгар нереста (табл. 18).

Из приведенных данных видно, что икринки на I этапе развития встречаются в период времени от 16 до 02 ч следующих суток с максимальной численностью между 20 и 24 ч. В сумеречное время (16—20 ч) икринки на начальных стадиях развития встречались единично. Следовательно, нерест ставриды происходит в основном между 20—24 ч.
Икринки на II этапе развития, соответственно малой его длительности (см. ниже), встречались на протяжении небольшого периода времени — от 22 до 04 ч следующих суток с максимумом между 24 и 02 ч. Встречаемость последующих этапов развития соответственно сдвигается во времени: икринки на III этапе встречаются между 24 и 12 ч следующих суток с максимумом в период от 02 до 08 ч, на IV этапе — от 04 до 16 ч с максимумом распределения между 10 и 12 ч; на V этапе — между 12 и 22 ч и на VI этапе — между 18 и 02 ч.
В летнее время (с июня по август) в планктон ежесуточно поступают икринки ставриды. В любом районе моря в ночное время можно встретить свежевыметанные икринки (на этапе дробления). Ежесуточное поступление икринок в планктон определяется растянутым многопорционным нерестом ставриды, который свойствен многим другим рыбам Черного моря (Овен и др., 1970).
Строение икринок. Икринки ставриды пелагические, сферической формы, с одной жировой каплей. Оболочка очень тонкая, прозрачная, желток сегментированный (рис. 22). Диаметр икринок черноморской формы Trachurus mediterraneus колеблется от 0,73 до 1,00 мм, жировой капли — от 0,19 до 0,29 мм (Дехник и Павловская, 1950; Водяницкий и Казакова, 1954; Зайцев, 1959а; Георгиев и др., 1960). Средний размер живых развивающихся икринок по нашим измерениям в разные месяцы нерестового сезона составляет 0,84 мм, жировой капли — 0,26 мм.
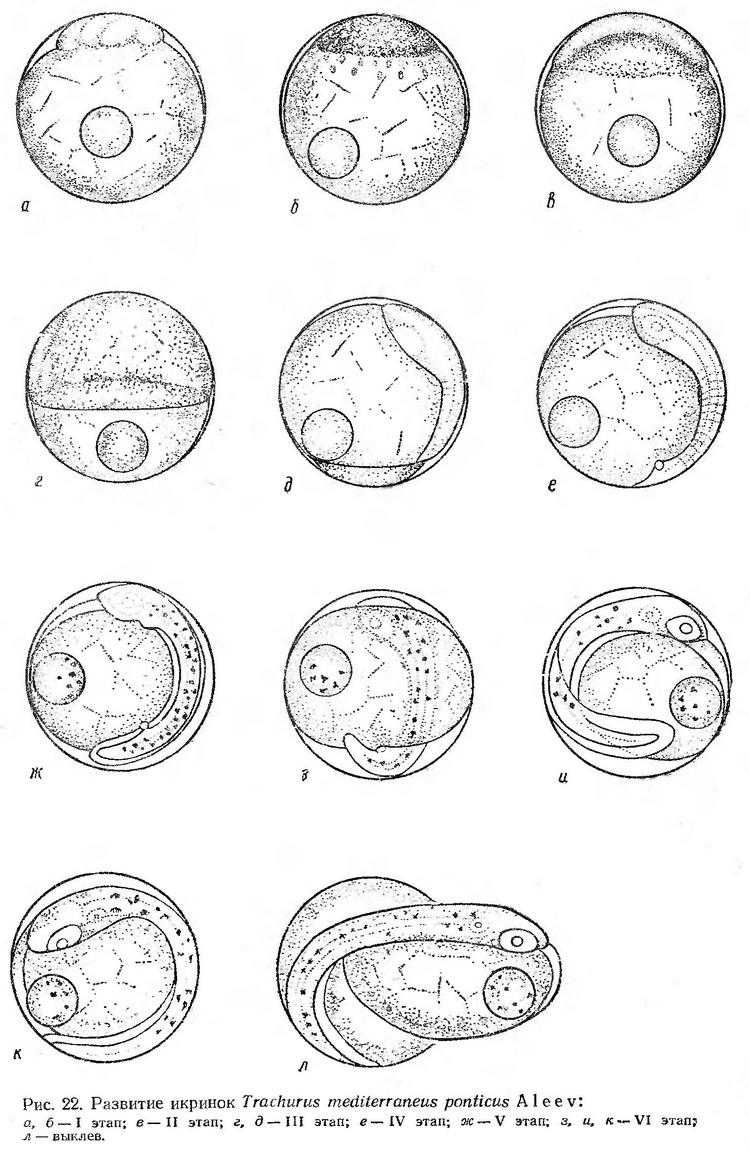
По размерам и величине жировой капли икринки ставриды близки к икринкам другого массового вида — султанки. Икринки обоих видов встречаются в планктоне в летний период в основном в одних и тех же районах. Некоторые отличительные признаки икринок обоих видов указаны А. И. Смирновым (1949, 1953). Икринки ставриды в массе заметно крупнее, чем икринки султанки, на что указывают их средние размеры в разные месяцы нерестового сезона. Жировая капля у первого вида также более крупная, ее поверхность несколько неровная, как бы шероховатая, в отличие от султанки, жировая капля которой имеет четкие очертания. По мере развития эмбрионов различия этих двух видов становятся более отчетливыми (рис. 22 и 30).
Диаметр икринок Trachurus mediterraneus из Средиземного моря, по данным Падоа (Padoa 1956в), 1,00—1,04 мм, жировой капли — 0,24 мм. По нашим измерениям (Дехник, Синюкова, 1964), диаметр икринок этого вида (?) колеблется от 0,74 до 0,94 мм, жировой капли — от 0,19 до 0,24 мм. Преобладали икринки размером 0,84—0,85 мм с жировой каплей 0,22—0,24 мм. Принимая во внимание размер икринок, жировых капель и пигментацию личинок рода Trachurus, преобладающих в наших материалах из Средиземного моря, мы предположительно отнесли их к Trachurus mediterraneus. По размерам и величине жировой капли эти икринки близки к икринкам этого вида из Черного моря и заметно отличаются от описанных Падоа. Можно предположить, что описанные Падоа под видом Trachurus mediterraneus икринки в действительности относятся к Trachurus trachurus.
Эмбриональное развитие ставриды изучалось на икринках, собранных в море в июне и июле между 22 и 23 ч. В это время в планктоне икринки ставриды находятся на начальных стадиях развития (8—16 бластомеров) (рис. 22, а). Первый этап развития — дробление при средней температуре 19,6° — продолжался около 4 ч. Зародышевый диск составляет небольшую часть поверхности желтка. Его высота около 0,20 мм, длина основания 0,63 мм (рис. 22, б, табл. 19).
Образование эпителиальной бластулы происходит очень быстро. При температуре воды 20,6° уже через 2 ч после окончания дробления четко просматривается бластоцель. Зародышевый диск чуть уплощен, начинается обрастание желтка с одновременным подворачиванием краев бластодиска. Через 1,5—2 ч после начала обрастания при температуре 20,6° бластодерма охватывает около 1/3 поверхности желтка (рис. 22, в). Одновременное подворачиванием краев бластодиска продолжается его рост. Когда его края заходят за среднюю линию желтка, намечается зародышевая полоска (рис. 22, г). Через 8,5 ч после начала обрастания на вегетативном полюсе остается небольшая желточная пробка (рис. 22, д). Головной конец зародыша сильно расширен. Появляется нервный тяж, мозговые пузыри, намечаются глазные бокалы и первые туловищные сегменты. Этап обрастания и гаструляции при температуре 20,6—20,0° продолжается около 9 ч.
После замыкания бластопора происходит быстрая дифференцировка зародышевых органов. Головной конец четко обособляется и достигает анимального полюса. В глазах появляются хрусталики, намечаются зачатки слуховых капсул, происходит быстрая сегментация туловищного отдела (рис. 22, е). IV этап при температуре 20,0—19,4° продолжается около 6 ч. Ко времени образования хвостовой почки эмбрион охватывает немногим более половины поверхности желтка. Наряду с ростом хвостового отдела происходит дифференцировка хвостовых миотомов; появляются зачатки кровеносных сосудов, сердца, кишечника, печени, образуется плавниковая кайма. На теле зародыша и на жировой капле появляются крупные меланофоры (рис. 22, ж).
При температуре 19,4—20,4° V этап продолжается около 7 ч. Когда эмбрион охватывает около 2/3 поверхности желтка, начинается пульсация сердца и слабые подергивания эмбриона (рис. 22, з). На теле появляется диффузный каротиноидный пигмент. По мере роста зародыша и развития зачатков органов движения эмбриона становятся энергичнее. Перед выклевом хвост зародыша почти смыкается с головой, остается свободной примерно 1/10 часть поверхности желтка. Эмбрион вместе с желтком свободно поворачивается внутри яйцевой оболочки, сердце интенсивно пульсирует — 120—150 раз в минуту. В заглазничной области и антеанальной части тела образуются сгущения коричневого пигмента (рис. 22, и, к). Процесс выклева длится несколько минут. Головной конец выпячивается через разрыв оболочки и затем последовательными толчками освобождается туловище и хвост (рис. 22, л). Этап подвижного эмбриона длится около 8 ч (рис. 22, з—л). Весь процесс эмбрионального развития ставриды при температуре воды 19,5—20,6° продолжается около 36 ч (табл. 19).

Постэмбриональное развитие. Личинки ставриды выклевываются длиной 1,63—2,00 мм. Большой яйцевидный мешок выдается за голову. У переднего края желтка расположена сравнительно крупная жировая капля. Анальное отверстие открывается в край плавниковой каймы за серединой тела. Имеются небольшие зачатки грудных плавников. Голова, передняя часть тела и жировая капля окрашены коричневатым пигментом. На спинной стороне тела, на голове и на жировой капле располагаются крупные точечные меланофоры (рис. 23, а). Выклюнувшиеся личинки занимают вертикальное положение. Они как бы подвешиваются к поверхностной пленке головой или правильнее вытянутой частью желтка и свободно парят в воде. Жировая капля, расположенная у самого переднего края желточного мешка, способствует сохранению такого положения. Личинки длительное время находятся в покое. Паузы 5—10 — 30 сек сменяются короткими перемещениями, продолжительностью не более 2 — 3 сек. Движения скачкообразные. В покое личинки медленно погружаются, не меняя положения, затем скачками достигают приповерхностного слоя, на некоторое время замирают и снова медленно опускаются. Парящие в вертикальном положении личинки с гонким почти прозрачным телом, по-видимому, едва заметны в воде.
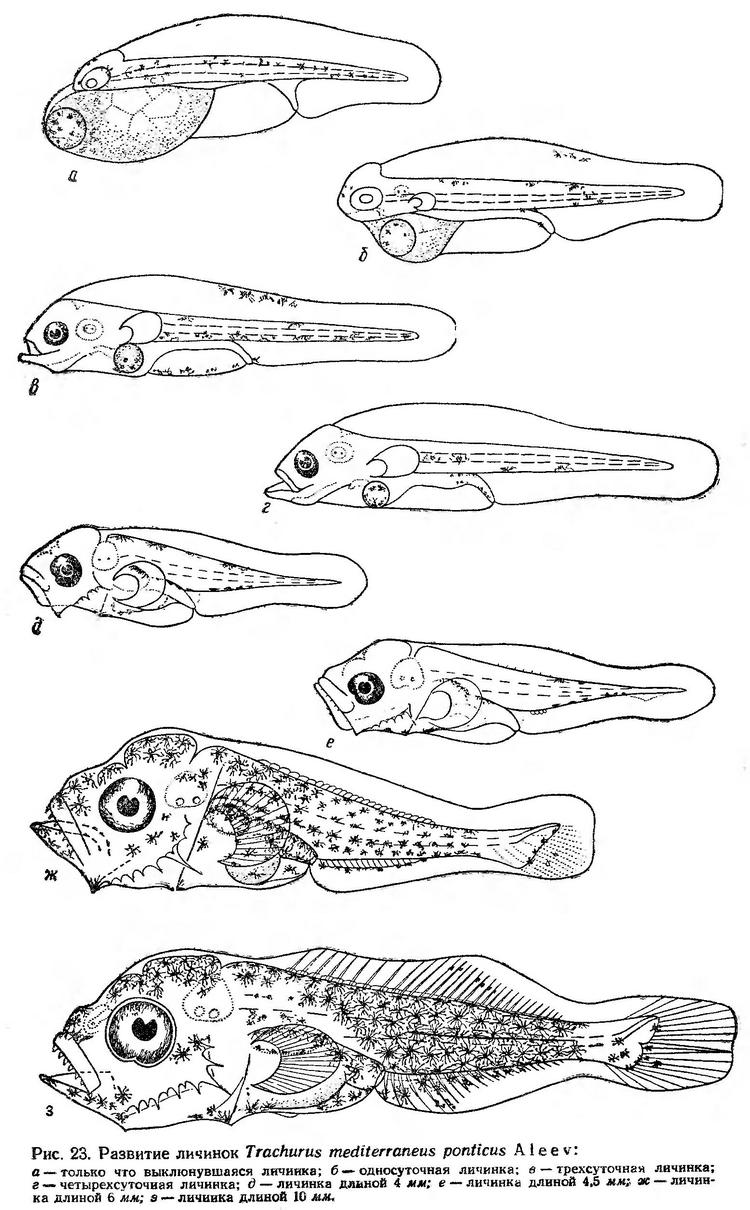
В течение первых суток развития значительная часть желточного мешка резорбируется. Намечается ротовая ямка, увеличиваются зачатки грудных плавников. Фоновая коричневая окраска делается более интенсивной (рис. 23, б). Активность личинок заметно возрастает. Периоды покоя продолжаются не более 10 сек. Личинки «отрываются» от поверхностной пленки, принимают горизонтапьное положение (брюшной стороной кверху). Время от времени делают стремительные короткие броски, почти не меняя положения или чуть переворачиваясь на бок.
К трехсуточному возраст, личинки достигают 2,63—2,72 мм длины. У них сохраняется еще небольшой остаток желтка; жировая капля заметно уменьшается в размере. Прорезается рот и приобретает подвижность. Формируются челюсти (рис. 23, в). Большие веерообразные грудные плавники при движении личинок вибрируют, благодаря чему улучшается плавучесть личинок. Они начинают двигаться в разных плоскостях, меняя направление движения. Глаза интенсивно пигментированы, и, по-видимому, в этом возрасте личинки уже начинают различать плавающие предметы и выбирать среди них необходимые. Об этом свидетельствуют, как это видно из изложенного ниже, наблюдения В. И. Синюковой (1963а, 1964, 1969) за питанием личинок в море.
Личинки в состоянии покоя занимают горизонтальное или косое положение, ориентируясь брюшной стороной кверху. При движении быстро переворачиваются спинной стороной кверху, иногда делают стремительные броски на короткие расстояния.
По наблюдениям Н. И. Ревиной (1958, 1964) и В. И. Синюковой (1963а, 1964, 1969), у личинок такого размера, выловленных в море, в кишечниках уже встречается пища. В пищевом комке насчитывается 6—8 видов фито- и зоопланктонных организмов. Основные объекты питания таких личинок — науплиусы Copepoda размером 0,088 мм и инфузории, преимущественно Metacylis mediterranea размером 0,060 мм. Даже в те периоды наблюдений, когда в планктоне науплиусов Copepoda и инфузорий было мало, в пищевом комке личинок они составляли основную долю (Синюкова, 1964). Следовательно, эти организмы активно выбирались личинками ставриды из общей массы зоопланктона, что свидетельствует о способности личинок видеть и различать плавающие в воде предметы. Таким образом, уже в возрасте трех суток, имея еще запас питательного материала в виде желтка и жира, личинки ставриды переходят на внешнее питание.
К четырехсуточному возрасту средняя длина личинок не изменяется. У них еще сохраняется незначительный остаток желтка и жировая капля (рис. 23, г). Плавательный пузырь наполняется воздухом. Рот ритмично подвижен. Личинки начинают плавать спиной кверху. В связи с использованием желтка они переходят к активному образу жизни, что связано с потребностью добывания пищи. В этом возрасте они уже способны к длительным целенаправленным движениям. При помощи больших подвижных грудных плавников и сильных изгибов хвостового отдела личинки могут достаточно быстро и точно делать повороты. Периоды покоя значительно сокращаются; они в несколько раз короче периодов движения. В пищевом комке таких личинок также преобладают науплиусы Copepoda и инфузории (Синюкова, 1963а, 1964, 1969).
Личинки более позднего возраста в экспериментальных условиях не живут. Личинки длиной 4,0 и 4,5 мм (рис. 23, д, е) зарисованы по фиксированному материалу, остальные — по живому (из планктона).
При достижении личинками длины 4,0 мм у них заметно изменяются пропорции тела. Голова делается массивной, значительно увеличивается высота туловища по отношению к его длине и укорачивается антеанальное расстояние. Челюсти снабжены зубами, на предкрышках развиваются сильные колючки. Вдоль спины располагаются крупные звездчатые меланофоры. Черные пигментные клетки имеются также на нижней стороне хвостового отдела и над кишечником. Плавниковая кайма несколько уменьшается по высоте (рис. 23, д). Личинки интенсивно питаются. В светлое время суток они почти никогда не встречаются с пустыми кишечниками (Синюкова, 1963а, 1964,1969). В пищевом комке этих личинок, как и у более мелких, встречается до 12 видов планктонных организмов, но качественный состав их значительно отличается. Так, в пище личинок отсутствуют инфузории и появляются взрослые формы Oithona minuta, Paracalanus parvus и Podon polyphemoides. Основное значение в питании этих личинок имеет Oithona minuta.
У личинок длиной 4,5 мм начинается дифференцировка непарных плавников, намечаются основания лучей в спинном, анальном и хвостовом плавниках (рис. 23, е).
Личинки длиной 6 мм имеют большие хорошо развитые грудные плавники со сформированными лучами. Закладываются лучи в хвостовом плавнике, продолжается формирование спинного и анального плавников. Появляются зачатки брюшных плавников. Голова и тело личинок интенсивно пигментированы меланином (рис. 23, ж).
У личинок длиной 8—10 мм заканчивается формирование непарных плавников (рис. 23, з). Они способны к очень быстрым движениям, мгновенно изменяют направление, делают резкие повороты. Питаются личинки главным образом взрослыми формами Oithona minuta размером 0,25—0,28 мм, Paracalanus parvus и метанауплиусами Copepoda (Синюкова, 1963а, 1964). У личинок в этом возрасте развивается инстинкт защиты от врагов. Они собираются стайками и часто находят убежище под куполами медуз, куда они прячутся при приближении опасности.
Распределение и численность икринок и личинок. Данные различных исследователей, как и наши наблюдения, показывают, что икринки и личинки ставриды в основном входят в состав прибрежного ихтиопланктона, характеризующегося разнообразным видовым составом и значительно более высокой общей численностью в сопоставлении с открытым морем.
Икринки ставриды, как и икринки многих других теплолюбивых рыб, распределяются в верхних теплых горизонтах — до слоя температурного скачка (Косякина, 1938; Павловская, 1954; Ревина, 1958). Ниже слоя скачка встречаются мертвые, выпадающие из планктона икринки.
Личинки ставриды распределяются в пределах этого же слоя, однако, как и личинки хамсы, они концентрируются преимущественно в слое 5—15 м (Павловская, 1954; Ревина, 1958; Синюкова, 1964). Наши наблюдения за распределением личинок на 0 и 10 м показали закономерное увеличение их численности с глубиной (табл. 20).

В табл. 21 приведены данные по изменению численности икринок и личинок ставриды в течение нерестового сезона и в разные годы наблюдений.

Первые икринки ставриды появляются в планктоне с конца мая (Дехник, Павловская, 1950). У побережья Болгарии наиболее ранний нерест отмечен в первых числах июня (Георгиев, Александрова, Николов, 1961).
В начале нерестового сезона численность икринок ставриды в планктоне незначительна. Личинки появляются единичными экземплярами в первых числах июня. К середине июня численность икринок и личинок в планктоне значительно возрастает. В июле наблюдаются максимальные концентрации икринок и личинок этого вида. В августе интенсивность нереста обычно снижается и численность икринок и личинок в планктоне уменьшается (табл. 21).
В сентябре икринки ставриды ловятся единичными экземплярами, личинки в планктоне не встречаются, что, по-видимому, как и у хамсы, свидетельствует о безрезультатности нереста в это время.
Численность икринок и личинок ставриды в планктоне значительно колеблется по годам (табл. 21). В одном и том же районе (район Севастополя, 2,5—10 миль от берега) пределы колебаний численности икринок за три года наблюдений (1960—1962) выражаются соотношением 1 : 3. Численность личинок в эти годы наблюдений колебалась в среднем от 0,2 до 8 экз. под 1 м2. Наиболее высокая концентрация икринок и личинок ставриды отмечена для прибрежного района (Камышовая бухта, 1—1,5 мили от берега, наблюдения 1959 г). Наоборот, в отрытом море (наблюдения 1963, 1965 г.) численность икринок и личинок этого вида во время разгара нереста (конец июня — начало августа) была низкой (табл. 21).
Данных о численности икринок и личинок Trachurus mediterraneus в Средиземном море в литературе нет. Наши наблюдения (Дехник, Синюкова, 1964) показали, что численность икринок и личинок этого вида (?) в Средиземном море чрезвычайно низкая. На один горизонтальный лов приходилось не более 25 икринок (обычно до 10) и не более 17 личинок (чаще 2—4). Однако необходимо отметить, что наши работы проводились в основном в значительном удалении от берегов, тогда как нерест этого вида, как показано выше, происходит преимущественно в прибрежных районах. Следовательно, приведенные данные не могут характеризовать интенсивность нереста этого вида в Средиземном море.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
У восточных берегов Атлантического
океана, от Бискайского залива до
Зеленого мыса, в Средиземном и Черном
морях обитает средиземноморская ставрида
(Trachurus mediterraneus). В пределах
своего ареала средиземноморская
ставрида образует ряд резко локализованных
стад, различающихся размерами
входящих в них особей. Биология ее очень
сходна с биологией обыкновенной
ставриды. Она имеет также большое промысловое
значение и добывается теми же орудиями
лова, что и предыдущий вид.
Черноморскую ставриду считают особым подвидом
(Т. mediterraneus ponticus), причем
различают две ее формы: обычную — мелкую,
до 20 см длины, и южную — крупную, до
55 см. Нерест ее происходит вдоль всех
берегов Черного моря с мая по август,
в основном в июне, при температуре
воды 17—23° С. Питается ставрида главным
образом мелкой рыбой и ракообразными.
В зимнее время опускается на склоны
ям, на глубину от 30 до 80—100 м, почти
или совсем не питаясь. Ставрида — одна
из основных промысловых рыб Черного
моря.