Trachurus murphyi Nichols, 1920
Перуанско-чилийская ставрида, перуанская ставрида, чилийская ставрида (Rus),
Chilean jack mackerel
(Eng)
Синонимы:
Caranx peruanus Tschudi, 1846
Caranx peruanus Tschudi, 1846
Trachurus murphyi Nichols, 1920
Trachurus symmetricus murphyi Nichols, 1920 Перуанско-чилийская ставрида (Rus),
Рыбы открытого океана. Н.В.Парин 1998 г.
Следует особо остановиться на периодически происходящих в разных районах и иногда продолжающихся несколько лет подряд массовых выходах в открытую пелагиаль стайных неритических рыб. Такие выходы отмечены как в тропической зоне (в тепловодной Атлантике, например, в последние годы резко увеличилась численность серого спинорога Balistes carolinensis, который стал вполне обычным и в открытом океане), так и в умеренных водах, для которых они особенно характерны. В качестве примеров, кроме уже упомянутых «океанической сельди» и путассу, нужно назвать японского анчоуса (Engraulis japonicus), сардинопсов (подвиды Sardinops sagax, включая иваси) и особенно скумбрию (Scomber japonicus) и ставрид из группы Trachurus picturatus. Все они принадлежат к числу видов, характеризующихся периодическими крупномасштабными изменениями численности (своего рода ритмическими «приливами» и «отливами» волн жизни), в основе которых, как считают В. П. Шунтов и другие исследователи, лежит ритмичность климатических процессов, в свою очередь определяемая космофизическими циклами (в частности, солнечной активностью). Их выходы в открытый океан всегда связаны с временным увеличением численности неритической популяции (или популяций) в области ее (их) нормального обитания.
Можно полагать, что выселению популяционного «избытка» в океан предшествует увеличение размаха двусторонних (нагульно-нерестовых) миграций, которые имеют некий предел дальности и становятся необратимыми при его превышении. Первоначально происходит, по-видимому, расширение нагульной части ареала во всех возможных направлениях — не только вдоль берегов (так обычно ориентированы нормальные сезонные миграции умеренноводных неритических рыб), но и от них — в открытые воды океана (увеличение площади нерестовой области в большей степени лимитируется консервативными требованиями вида в репродуктивный период). Пока эти миграции сохраняют двусторонний характер, часть популяции, находящаяся за пределами неритической зоны, должна считаться мероэпипелагической, однако у некоторых видов при большом росте численности стайные группы, достаточно далеко удалившиеся от традиционных миграционных путей, оказываются неспособными вернуться обратно. Эта часть популяций переходит тем самым в ксеноэпипелагическую категорию. Постоянно пополняясь за счет новых выселений из неритической зоны, она остается в океане вплоть до естественного вымирания (элиминации), если только не попадет в новые неритические районы или же в такие участки открытого океана, в которых условия среды окажутся благоприятными хотя бы для питания и роста, а в наиболее удачном варианте также для созревания, нереста и выживания на ранних стадиях жизненного цикла. Такие участки (я называю их «псевдонеритическими»), как правило, оказываются приуроченными к биологически продуктивным районам вблизи островов и поднятий или у зон конвергенции и дивергенций течений. В соответствии с возможными различиями условий среды в новых участках могут образовываться три типа популяционных группировок: 1) псевдопопуляции, не способные к нормальному воспроизводству и существующие только за счет пополнения извне; 2) временные зависимые популяции, которые могут существовать при отсутствии пополнения из исходной самовоспроизводящейся популяции лишь до тех пор, пока сохраняется благоприятная обстановка; 3) новые самостоятельные популяции.
Интересный материал для размышлений представляют сведения по биологии и распространению двух восточнотихоокеанских видов ставрид из группы Trachurus picturatus. Размножение калифорнийской ставриды (Trachurus symmetricus simmetricus) было отмечено по нахождениям икры и личинок в зоне 40-х параллелей северного полушария в 1955 г. в удалении до 1100 миль, а в 1972 г.— до 600 миль от берегов Северной Америки. Аналогичным образом нерест перуанско-чилийской ставриды (Trachurus symmetricus murphyi) в 40-х широтах южного полушария отмечен в 1985 г. С. А. Евсеенко на огромном расстоянии (около 3000 миль) от побережья Чили. Никаких свидетельств в пользу результативности такого нереста, впрочем, не существует, так как в этих океанических участках — у крайних границ своего распространения на запад — оба вида представлены, судя по имеющимся данным, только очень крупными особями длиной 45—60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб, а созревает ставрида в возрасте 2—3 лет). Таким образом, эти факты могут трактоваться лишь как доказательства существования временных псевдопопуляций (маловероятным представляется даже обособление зависимых популяций), которые не только существуют за счет постоянного пополнения из прибрежного запаса, но и являются фактически его излишком. Выселение перуанской ставриды в океан вполне объяснимо ростом ее численности в неритической зоне. Так, согласно рыбопромысловой статистике ФАО (Продовольственная и сельскохозяйственная организация ООН), уловы этого вида в прибрежных водах Перу и Чили (они, видимо, отражают и истинную численность ставриды) увеличились со 100 тыс. т в 1970 г. и 300—400 тыс. т в 1974—1976 гг. до 1100—1300 тыс. т в 1978—1980 гг., а в открытом океане эта рыба была обнаружена в 1978 г.
Цикличность процесса неизбежно предполагает грядущий спад численности и даже почти полное исчезновение этой рыбы из открытых для международного рыболовства вод. Правда, в те же сроки в каких-то иных участках Мирового океана могут появиться псевдопопуляции других видов. Прогнозировать ход этих пульсационных процессов — одна из важнейших, на мой взгляд, задач прикладной рыбохозяйственной науки.
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus murphyi Nichols - перуанская ставрида
Описание. D I, VIII+I, 29-38, в среднем 32; А II+I, 24-33, в среднем 27,7. Всех щитков в боковой линии 90-107, в среднем 98; колючих щитков - 44-56, в среднем - 47. Голова 25,1-31,5% длины тела, в среднем 28,2%. Верхняя (спинная) ветвь боковой линии оканчивается под основанием первого-седьмого луча второго спинного плавника, обычно под третьим лучом второго спинного плавника. Жаберных тычинок 51-66, в среднем 60,4; на нижней части жаберной дуги 37-48, в среднем 44,6. Щитки боковой линии средней величины, высоты наиболее высоких щитков передней (кривой) части боковой линии 16,5-20,0%, в среднем 16,9% длины головы; высота наиболее высоких щитков задней (прямой) части боковой линии 15,6-21,5%, в среднем 17,0% длины головы. Кили на щитках хорошо развиты, шипы на килевых щитках крепкие. Длина прямой части боковой линии, несущей киль, составляет 36,3-44,0%, в среднем 40,0% длины тела, а хорда дуги кривой (передней) части боковой линии - 41,0-48,0%, в среднем 45% длины тела. Грудные плавники 25,6-31,6%, в среднем 28,4% длины тела. Тело овальное, его высота 16,1-20,4%, в среднем 18,2% длины. Горизонтальный диаметр глаза 18,1-25,5%, в среднем 22,4% длины головы. Верхняя челюсть 33,1-38,9%, в среднем 36,2% длины головы. Изгиб боковой линии лежит под основанием 8-13-го, в среднем 10-11-го луча второго спинного плавника. Первый колючий щиток лежит под основанием 8-13-го, в среднем 11-го луча второго спинного плавника. Расстояние от вершины рыла до вертикали начала основания брюшных плавников 28,4-35,9%, в среднем 30,5% длины тела. Расстояние от вершины рыла до вертикали начала основания первого спинного плавника 29,5-37,6%, в среднем 33,1% длины тела. Расстояние от вершины рыла до вертикали начала основания анального плавника 50,0-61,1%, в среднем 57,2% длины тела. Отношение расстояния между последним и предпоследним лучами анального плавника к расстоянию между последним и предшествующим ему (третьим сзади) лучом анального плавника равно 1,3-2,4, в среднем 1,8 раза. Ответвления каналов боковой линии на верхней стороне головы и на спине хорошо развиты. Спина и верхняя часть головы темные, бока и брюхо серебристые, грудные, спинные и хвостовой плавники серые, брюшные и анальный плавники белые. Вершина рыла и основание грудных плавников черные. На жаберной крышке черное пятно.
Описание дано по 300 экз. длиной от 20,5 до 63,5 см, средней длиной 29,0 см, пойманных нами за пределами 200-мильной зоны Чили и Перу.
Распространение. Перуанская ставрида широко распространена как в прибрежных неритических районах, так и в открытом океане, от 3°ю.ш. (Галапагосские острова) до 57°ю.ш. (Магелланов пролив) и от берегов Эквадора, Перу и Чили далеко на запад в открытый океан, вплоть до района Новой Зеландии (177°з.д.) (рис.20).
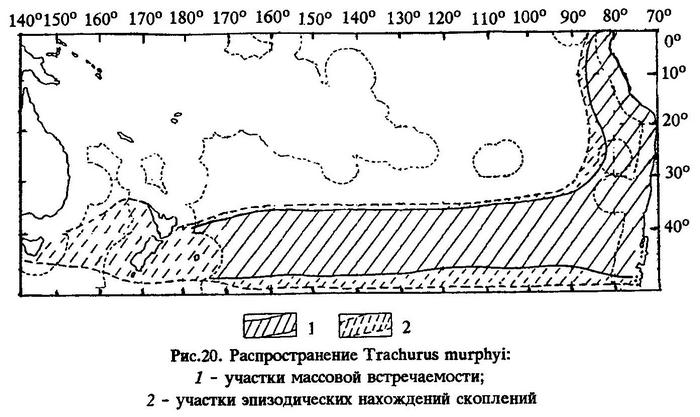
В южной части Тихого океана ареал перуанской ставриды вытянут с востока на запад. Северной его границей служит периферия субтропической фронтальной зоны, которая характеризуется повышенными меридиональными градиентами солености; в водах, имеющих соленость выше 35‰, перуанская ставрида практически не встречается.
Южная граница распространения этого вида определяется периферией антарктической полярной фронтальной зоны, или поверхностной изотермой 8°С.
Присутствие перуанской ставриды в водах Новозеландского плато в пределах 200-мильной экономической зоны японские исследователи [Kawachara et al., 1988] обнаружили во время совместной с новозеландцами съемки на НИС "Синкай Мару" в 1986 г. над подводными банками чатемского поднятия - Мерну и Верьян, около о-вов Чатем, а также с восточной стороны пролива Фово. Экспедиции ТИНРО в 1988 г. выполнялись с восточной и западной сторон экономической зоны Новой Зеландии. В обоих участках была обнаружена перуанская ставрида. Контрольное траление, выполненное в открытых водах Новозеландского плато дало 4.5 т перуанской ставриды. Две экспедиции ТИНРО в 1986 г. (НПС "Пулковский меридиан") и в 1987 г. (НИС "Пионер Николаева") в августе и ноябре имели в контрольных тралениях по нескольку экземпляров перуанской ставриды длиной 55-61 см. При работах в районе о-ва Снэрс (46-50°ю.ш., 166-179°в.д.) только за вторую половину марта 1990 г. ее присутствие в уловах отмечалось 33 раза.
В декабре 1987 г. на РТМС "Звезда Севастополя" в Тасмановом море в точке 41°50'ю.ш. и 164°20'в.д. в ночное время при температуре воды у поверхности 14,8°С в слое 10-80 м было поймано 10 экз. самцов перуанской ставриды длиной 55-61 см и массой 2,0-2,4 кг. Их желудки были растянуты от большого количества съеденных эуфаузиид. Позднее в Тасмановом море были получены уловы перуанской ставриды другими судами.
Распространение перуанской ставриды непосредственно связано с системой Перуанского течения, Антарктическим циркумполярным течением и мезомасштабными динамическими образованиями (вихри, меандры, ринги, апвеллинговые зоны).
Считается, что с 70-х годов ареал перуанской ставриды расширился, особенно после Эль-Ниньо 1976 г., когда ее значительные скопления появились вдоль всего перуанского побережья [Zuta, 1983].
Причины появления большого количества перуанской ставриды и некоторых других видов рыб многие авторы связывают с катастрофическим Эль-Ниньо 1972-1973 гг., когда ухудшились условия нереста анчоуса, погибла вся молодь, а половозрелые особи были в чрезмерном количестве выловлены, что резко сократило среднее число нерестующихся особей, необходимое для ежегодного пополнения.
В результате произошло снижение биомассы анчоуса с 20-30 млн.т до 2-3 млн.т за 10-летний период. Снижение потребления планктона и ослабление конкуренции между видами, питающимися планктоном, позволило шире распространиться ставриде, сардине, скумбрии и другим видам рыб [Jordan, 1983].
Важно отметить, что эти изменения в видовом составе в ЮВТО развивались с различными характеристиками у берегов Перу и Чили. В то время когда анчоус традиционно удерживал высокий уровень биомассы и свои нерестилища в Перу, в Чили величина его биомассы составляла примерно 1/10 перуанской. Это подтверждается тем, что в Перу максимальный вылов рыбы достигал 12 млн.т (1970 г.), а в Чили - порядка 1 млн.т (1971 г.). Последующее снижение биомассы анчоуса в водах Перу в 10 раз означало высвобождение неиспользуемой органической материи, что создало благоприятные условия для распространения других видов рыб. Можно предположить, что у чилийского побережья этот эффект был значительно слабее из-за меньшей потерянной биомассы анчоуса. Следовательно, можно говорить о происшедшей замене анчоуса другими видами рыб, в частности ставридой, сардиной, скумбрией и т.д., в водах Перу; что же касается вод Чили, то здесь, по-видимому, произошел процесс активизации экосистемы - увеличения продукции [Jordan, 1983].
Подводя итог вышеизложенному, можно сказать, что пока неизвестно, обитали ли скопления перуанской ставриды далеко за пределами 200-мильной экономической зоны Чили постоянно и не были обнаружены ранее лишь из-за отсутствия современных технических средств (мощных судов, орудий лова, гидроакустической аппаратуры и т.п.) или это временное явление, вызванное влиянием катастрофического Эль-Ниньо 70-х годов [Павлов и др.,1984], или в силу каких-либо других причин в последние годы происходит проникновение перуанской ставриды из восточных районов, где находится основа ее ареала, все далее на запад. Возможно, это следствие многолетней флюктуации численности популяции в восточной части океана, в результате чего произошел выход в периферические районы ареала излишка особей. Не исключено, что увеличение численности ставриды в открытом океане связано с освоением ею новой экологической ниши, освободившейся в результате сокращения численности китов, использовавших те же пищевые компоненты. Дальнейшие исследования покажут, какая из гипотез верна.
Это и определит стратегические перспективы промысла перуанской ставриды в открытом океане.
Биология. На шельфе Перу в уловах доминирует ставрида длиной от 27 до 47 см. Наиболее крупная ставрида длиной 64-70 см и массой до 2900 г, отмечена южнее 16°ю.ш. Подавляющую часть уловов составляют особи длиной 30-38 см и массой 300-800 г. Южнее 16-17°ю.ш. в уловах преобладают ставриды длиной 34-45 см. Зависимость масса - длина выражается уравнением:
W = 0,05068 • ls2,56298,
где W - масса целой рыбы, г; ls - максимальная длина рыбы, см [Абрамов, Котляр, 1980].
У берегов Чили основу промысла составляют особи длиной от 26 до 34 см. Максимальная длина ставриды в промысловых уловах Чили - 40 см, хотя по литературе известны экземпляры и до 65 см [Serra, 1983]. Однако в рейсе НИС "Академик Книпович" у берегов Чили в 1973 г. на шельфе ловили ставриду длиной до 73 см (табл. 40).
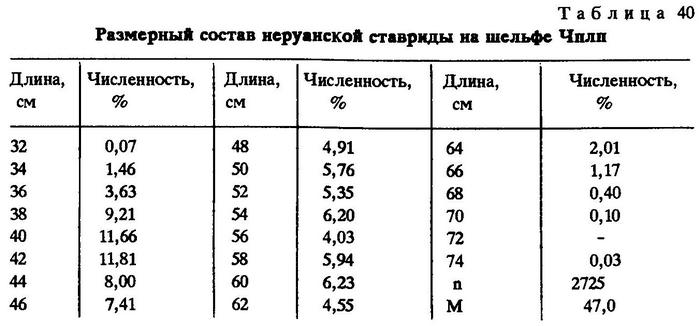
За все годы промысла перуанской ставриды в открытых водах юго-восточной части Тихого океана (за 200-мильной экономической зоной) в траловых уловах рыба была представлена размерами от самых минимальных - до 66 см и до 2900 г массой. Тем не менее все годы в открытом океане ловили мелкую и среднеразмерную ставриду. Лишь в 1990 г. средний размер ставриды в уловах несколько увеличился - до 34,8 см, чего не наблюдалось никогда ранее. Изменился и возрастной состав ставриды в уловах (табл.41).
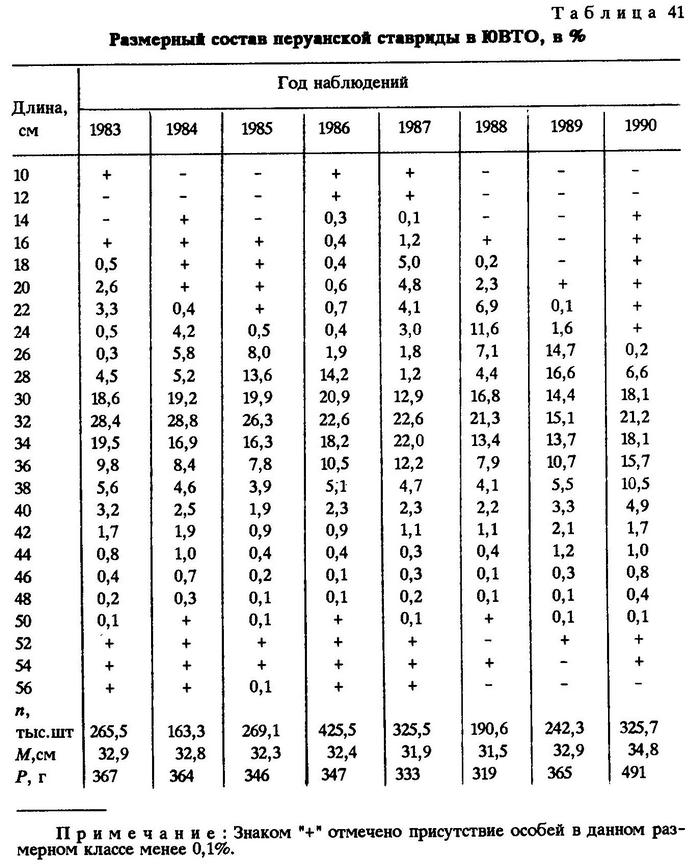
Промысловые скопления перуанской ставриды в разных районах и в разное время были образованы различными размерными группами.
Наряду с сезонной изменчивостью в распространении ставриды существуют различия в пространственном распределении различных размерных групп рыб. Кроме наличия большого числа элементарных популяций перуанской ставриды, отличающихся друг от друга средними размерами и обитающих по соседству друг с другом, установлено также и то, что более мелкая рыба преобладает на северных, северовосточных участках, а в южных участках обитает более крупная рыба. Это было показано нами еще в 1980 г., когда средние размеры рыб прямо пропорционально увеличивались в южном направлении через каждые два градуса широты.
Так, например, в 1990 г. в ЮВТО на участке между 12 и 25°ю.ш. вдоль 200-мильной экономической зоны, как и во все предшествующие годы, объектом промысла была мелкоразмерная ставрида (М = 25,3 см; n = 3375):
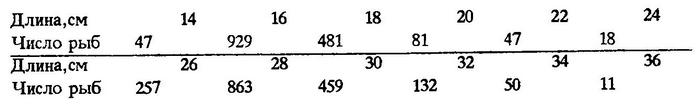
Оказалось, что чем ближе к западной периферии ареала поймана перуанская ставрида, тем она крупнее [Некрасов, 1992] (рис.21).
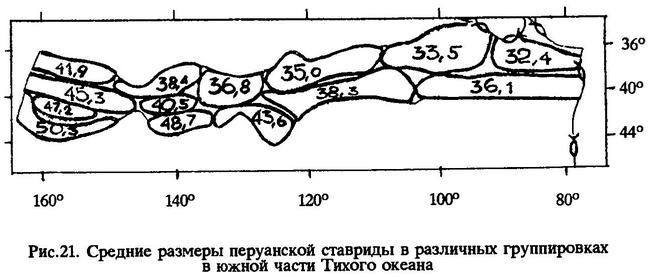
Закономерное увеличение средних размеров ставриды подтверждают материалы по ее размерному составу за 1980-1984 гг., сгруппированные через каждые 10° по долготе, Видно, что южнее 40°ю.ш. ставрида крупнее, чем к северу от этой параллели на тех же меридианах; сходная закономерность распределения разноразмерных групп рыб выявилась и в 1987 г. (табл. 42).
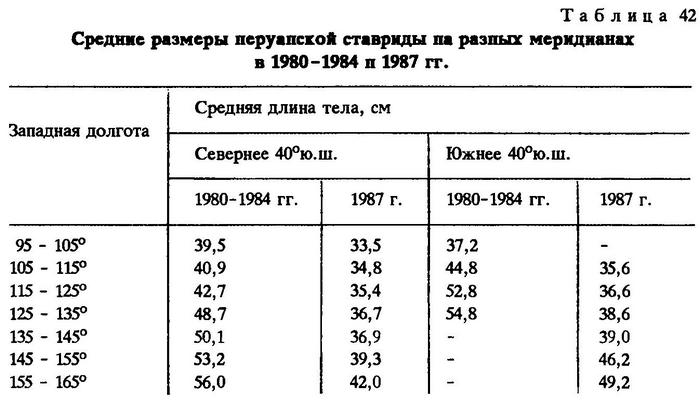
Возрастание средних размеров рыб свидетельствует о том, что ставрида мигрирует в западном направлении. Причем многолетние наблюдения за состоянием запасов перуанской ставриды и ее распределением в южной части Тихого океана показывают, что это невозвратные миграции.
Данные вышеприведенных таблиц свидетельствуют об увеличении средних размеров ставрид, мигрирующих в западном направлении, но в 1987 г. было отмечено общее значительное снижение размеров рыб. Сходная картина наблюдалась нами в ЮВТО и в 1989 г., что свидетельствует о подходе молодых многочисленных и высокоурожайных поколений перуанской ставриды. Преобладание более молодых рыб, очевидно, говорит о хорошем состоянии популяции.
Одной из причин миграции перуанской ставриды в западном направлении к Новой Зеландии можно считать отсутствие для рыб старшего возраста (длиной 32 см и более и в возрасте 4 года и старше) энергетически выгодного корма на востоке Тихого океана, вблизи рыболовных зон Перу и Чили. По-видимому, эпипелагиаль за пределами шельфовых зон Перу и Чили служит частью нагульного ареала только для неполовозрелых особей перуанской ставриды (длиной менее 30 см и моложе 3 лет), рыбы старшего возраста не находят здесь энергетически выгодного корма. Отсутствие такого корма обусловливает миграции ставриды старших возрастных групп на запад, где в летне-осенний период на отдельных участках нотальной зоны южной Пацифики концентрации макропланктона, включая основной корм перуанской ставриды в этом районе - сальп и эвфаузиид, достаточно велики [Елизаров и др., 1992].
Исследования, польских ученых в районе между 20 и 48°ю.ш., вне 200-мильной экономической зоны прибрежных государств, до 90°з.д. [Wyniki zwiadu...,1980; Majewski, Kurrowicki, 1983] показали, что в уловах встречалась ставрида длиной от 17 до 73 см (модальные классы 33-47 см (87%), средняя длина 41,3 см). Зависимость масса-длина выражается уравнением:
W = 0,0091354 • L2,958.
Между 17°20'ю.ш. и 20°26'ю.ш. в уловах встречалась ставрида длиной 6-12 см [Wyniki zwiadu..., 1980]. Средняя длина рыб в районе 37-48°ю.ш. и 78-86°з.д. равнялась 38,6 см (от 32 до 50 см) [Majewski, Kurrowicki, 1983].
Исследования немецких ученых, проведенные в районе между 39-46°ю.ш. и от границ рыболовной зоны Чили до 83°з.д., показали, что средняя длина перуанской ставриды ЮВТО оказалась в пределах от 32,2 до 35,9 см. Минимальная длина в уловах составляла 25 см, максимальная - 60 см. Обнаружено, что по мере продвижения с севера на юг средняя длина ставриды в уловах увеличивается. Так, например, в северной части района основной размерный ряд ставриды был 29-33 см (пределы - 26-46 см, средняя длина 32,2 см), в западной части района соответственно 30-34 см (25-44 см, средняя длина 33,5 см), в южной части - 33-37 см (26-49 см, средняя длина 35,9 см). Даже в районе 46°50'ю.ш. средняя длина ставриды равнялась 41,8 см, а в районе 50°22'ю.ш. - 42,2 см. Каких-либо изменений размерных рядов в течение пяти недель работы не отмечалось [Lambert, Sievert, 1982].
Польским исследователям в районе 37-48°ю.ш. и 78-86°з.д. удалось установить аналогичную картину, причем средняя длина рыб возрастает от 38,6 до 43,0 см [Majewski, Kurrowicki, 1983].
Возраст перуанской ставриды немецкие исследователи определяли по отолитам. Однако использование этой регистрирующей структуры вызвало у них затруднение в чтении годовых колец. Лишь возраст ставриды, в котором она впервые встречается за 200-мильной экономической зоной, полагают они, можно считать точным: он равен трем годам. Минимальная длина в этом возрасте 24 см, максимальная - 34 см [Lambert, Sievert, 1982].
Известно, что у ставрид нет половых различий по длине тела [Алеев, 1957а; Некрасов, 1979]. В частности, нет их и у перуанской ставриды, о чем говорят материалы А.А.Абрамова и А.Н.Котляра (1980).
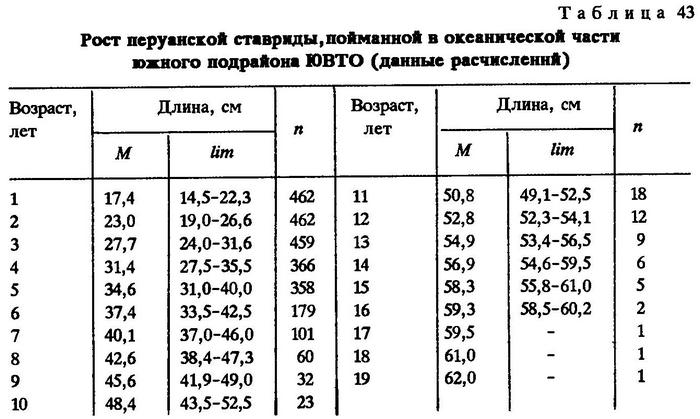
Возраст перуанской ставриды в наших исследованиях определялся как по отолитам, так и по чешуе. На отолитах рыб в возрасте старше пяти лет годовые кольца трудно различимы, а на чешуе рыб всех размеров кольца видны так же, как и у всех других видов ставрид, поэтому по возможности возраст определяли по чешуе. Первое кольцо считали мальковым, поскольку обратное расчисление роста показывает, что первое кольцо закладывается у перуанской ставриды длиной 7,5-14,1 см. В июле в океанической части ареала встречается молодь ставриды длиной 11-15 см. Нерест ставриды здесь длится с ноября по март, т.е. с начала нереста могло пройти 3-7 мес. Но в июле в Южном полушарии разгар зимы - время закладки зимнего кольца, что и отражается на регистрирующих структурах рыб при данной длине. Первые два годовые кольца видны хуже последующих. Они более расплывчаты, так как первые два года жизни фиксируются на регистрирующих структурах зимой, а все последующие являются нерестовыми [Некрасов, 1979]. Наши исследования показали, что на первом году жизни перуанская ставрида достигает длины от 14,5 до 22,3 см, в среднем 17,4 см; на втором - 19,0-26,6 см, в среднем 23,0 см; на третьем - 24,0- 31,6 см, в среднем 27,7 см и т.д. Максимальный экземпляр в наших исследованиях длиной 62 см имел возраст 19 лет (табл.43).
По данному ключу нами была рассчитана возрастная структура перуанской ставриды в уловах промыслового флота СССР (табл.44).
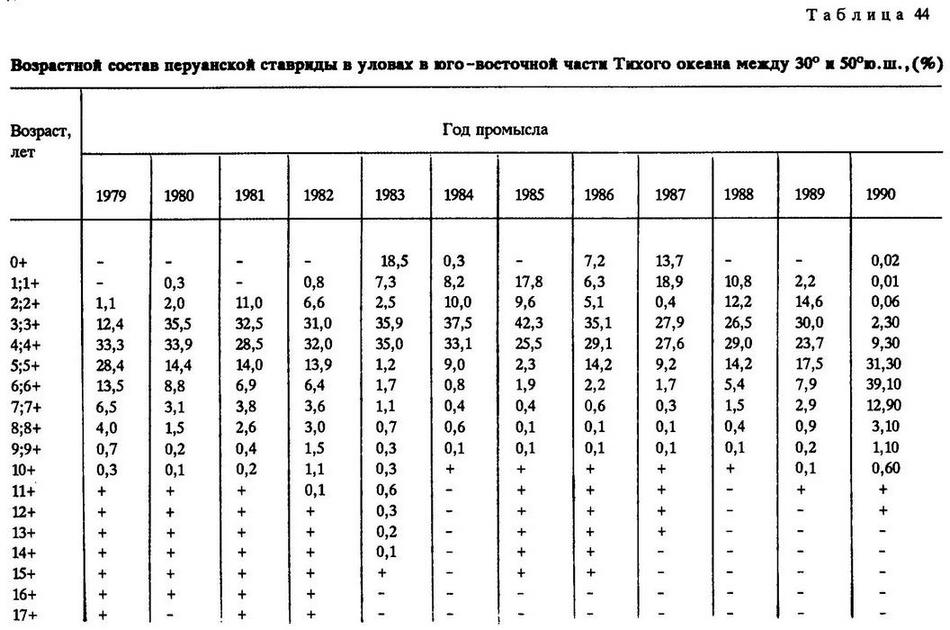
Из года в год в уловах в северном подрайоне постоянно и значительно преобладают по численности четырехлетки. Причем на следующий год вылов данного поколения резко сокращается.
Анализ размерной и возрастной структуры облавливаемой части скоплений перуанской ставриды свидетельствует о том, что в ЮВТО на участке 12-25°ю.ш. основу промысла в 1990 г. составили годовики с двухлетками (41,7%) и трехлетки (39,7%), на участке 30-43°ю.ш. - шестилетки (30,9%) и семилетки (32,8%).
Обращает на себя внимание, что на участке ЮВТО между 30 и 50°ю.ш. во все предшествующие годы преобладали в уловах пятилетки, а в 1990 г. даже рыбы более старшего возраста. В период Эль-Ниньо из островной зоны вод Чили, что расположена севернее 30°ю.ш., выходят на юг в большом количестве особи в возрасте 2+, и тем не менее на следующий год эта возрастная группа не становится преобладающей в уловах. Например, после Эль-Ниньо 1987 г. наибольшую по численности часть уловов ставриды в 1988 г. составили особи в возрасте 4+, а не 3+, как это можно было бы предположить. Это лишний раз подтверждает, что основное пополнение промысловой части популяции ставриды в ЮВТО, обитающей южнее 30°ю.ш., происходит, по-видимому, за счет эмиграции из вод Чили. На акватории же, прилегающей к водам Перу, ставрида в определенный период года в отсутствие Эль-Ниньо выходит далеко в открытый океан.
Таким образом, во все годы промысла всегда основу вылова по численности составляли четырехлетки независимо от того, более урожайным было это поколение по сравнению с другими или нет.
Из этого следует, во-первых, то, что такая ситуация (при постоянной интенсивности промысла) скорее всего возможна лишь в том случае, если в район промысла мигрирует из 200-мильной экономической зоны Перу значительная доля четырехлеток, поколение которых в предыдущие годы не выходило из зоны. Во-вторых, на следующий год четырехлетки, став пятилетками, а также и более взрослые особи в основной своей массе не выходят вновь за 200-мильную зону Перу.
В южном подрайоне ЮВТО во все годы промысла основу уловов составляют пятилетки. Однако здесь нет такого резкого уменьшения доли более старших поколений в общем вылове. Кроме того, на следующий год промысла здесь нет резкого уменьшения доли вылова бывших пятилеток, а теперь шестилеток, как это было с лидирующим поколением в северном подрайоне.
Обращает на себя внимание и то, что в южном подрайоне ежегодно доля четырехлеток в общем вылове по численности меньше доли пятилеток. Селективностью орудий лова этот факт объяснить нельзя, так как в северном подрайоне, где четырехлетки составляют основу вылова, ловят теми же орудиями. Видимо, в южном подрайоне пополнение промысловых скоплений происходит из 200-мильной зоны.
Возможно, что скопления перуанской ставриды, обитающие в открытой части океана, являются самостоятельной популяцией, обособленной от популяций, обитающих на шельфе и в прилегающих к нему водах. По крайней мере к таким выводам пришли исследователи на основании обнаруженных ими различий по морфологическим признакам [Константинов и др., 1979]. Кроме того, генетический анализ показал различия между группиррвками ставриды Перуанской и Чилийской котловин по соотношению фенотипов эстераз [Алексеев, 1984; Батальянц,1984; Коваль, 1984]. Однако ряд авторов оспаривает правомочность выделения двух и более популяций на основании биологических параметров, поскольку, в частности, не обнаружено достоверных различий в размерно-возрастном составе, в темпе роста и пополнения, показателях смертности и др. По-видимому, эти авторы упускают из виду то, что дать перечень всех признаков, в которых могут проявляться обязательные свойства популяции у рыб, невозможно. Понятно, что две популяции не могут быть идентичными. Две неразличимые по любым признакам, но репродуктивно изолированные внутривидовые группировки будут полноценными популяциями. Но этот признак - репродуктивная изолированность популяции, - проявляющийся в разобщенности районов или сроков нереста или в том и другом, не абсолютен, так как в любом из перечисленных вариантов возможно смешение икры и молоди без последующей дифференцировки по местам или срокам нереста [Алексеев; 1984], что, впрочем, принципиально отличает популяцию от биологического вида, поскольку в этом случае изоляция не стопроцентная.
При современном популяционном подходе к существованию вида, трудно найти биолога, который был бы действительно убежден в том, что перуанская ставрида, с ее обширным ареалом от Перу и Чили до Новой Зеландии и Тасманова моря, была бы представлена лишь единой популяцией.
Для перуанской ставриды характерны сезонные миграции. В северном подрайоне за 200-мильной экономической зоной Перу и Чили ставрида появляется лишь в определенные сезоны, где ее активный промысел ведут и российские суда. С наступлением лета она мигрирует в зону. Обычно мощные промысловые скопления перуанской ставриды в северном подрайоне наблюдаются с июня по декабрь. Эти сроки могут изменяться, поскольку они обусловлены образованием меандров перуанского океанического течения. В годы Эль Ниньо ставрида в северном подрайоне за 200-мильной зоной Перу не отмечена. По-видимому, она вытесняется теплым течением в островную экономическую зону Чили (острова Хуан-де-Фернандес), а частично даже в южный подрайон. В южном подрайоне ставрида обитает круглый год, совершая сезонные миграции с севера на юг на расстояние до 600 миль.
Вертикальное распределение перуанской ставриды очень широко. В северной части ареала, например, она наблюдается от поверхности до глубины 200 м, а в южноцентральных участках ареала - на глубинах до 300 м. Вне 200-мильных рыболовных зон установлены суточные вертикальные миграции ставриды. Днем она образует скопления различной плотности на глубинах порядка 40-250 м, ночью - ее плотные малоактивные скопления концентрируются на глубинах до 50-60 м, главным образом в слое 20-40 м [ Serra, 1983; Павлов, Каратаева, 1984].
В результате анализа распределения и поведения перуанской ставриды установлено, что ей присущ циклический ритм активности: максимальная активность в светлое время суток и снижение активности в темное время суток.
В дневное время перуанская ставрида образует подвижные скопления, состоящие из отдельных косяков различного размера и плотности, обычно на глубинах от 100 до 250 м. С заходом солнца с глубин 200-250 м начинается подъем ставриды в верхние слои. Сравнительно крупные косяки перуанской ставриды с вертикальным развитием от 5 до 20 м (мода 6-12 м), поднимаясь в верхние слои воды, распадаются на более мелкие с вертикальным развитием 2-4 м, образуя менее активные скопления на глубинах от 10 до 80 м в виде дорожек, лент, пятен. В это время ставрида активно питается и успешно облавливается тралом.
Перед восходом солнца вновь происходит перераспределение косяков ставриды. Из мелких косяков и разреженных скоплений образуются более крупные и подвижные косяки, которые очень трудно облавливаются тралом [Короткое и др., 1990].
Половые продукты впервые у перуанской ставриды начинают созревать при длине 25 см на третьем году жизни [Абрамов, Котляр, 1980]. Самцы созревают при несколько меньших размерах, чем самки. Около 30% рыб созревает при длине 24-27 см [Андрианов, 1985]. Дж.Серра [Serr, 1983] считает, что длина перуанской ставриды, при которой она достигает половой зрелости, равна 31 см. По К.Кайзеру [Kaiser, 1973], у берегов Чили ставрида также начинает созревать на третьем году жизни, но при этом имеет длину 36 см и массу 600 г. Интересно отметить, что ближайший родственник перуанской ставриды - калифорнийская ставрида впервые начинает созревать при длине тела 25 см, а при длине 35 см все рыбы уже половозрелые [MacGregor, 1966].
Изучение динамики зрелости половых продуктов перуанской ставриды показало, что в северном и южном подрайонах гонады у рыб созревают в разное время года.
В северном подрайоне ЮВТО гонады достигают IV стадии зрелости обычно в июне, и активный нерест наступает в августе, т.е. в середине зимы южного полушария. В южном подрайоне нерест начинается лишь поздней весной (в ноябре) и затухает к концу лета - началу осени, т.е. сроки нереста в северном и южном подрайонах четко соответствуют правилу Ортона, согласно которому популяции, близкие к теплым границам своего распространения, размножаются в холодное время, а популяции, близкие к холодным границам своего распространения - в теплое. Это обеспечивает появление и рост молоди в оптимальных для нее температурных условиях, так как молодь менее терпима к крайним условиям среды, чем взрослые особи.
Изучение хода развития гонад половозрелой (средняя длина 35 см) ставриды в течение годового репродуктивного цикла позволило установить, что в зимне-весенний нагульный период гонады ставриды находятся на VI-II, II-III и III стадиях зрелости, а значение гонадосоматического индекса колеблется от 0,35 до 1,30 у самок и несколько ниже у самцов. С интенсификацией генеративного обмена к началу весны на III стадии зрелости гонадосоматический индекс равен 1,30 у самок и 1,00 у самцов. Перед нерестовым сезоном, когда гонады находятся на IV стадии зрелости, гонадосоматический индекс у самок достигает 2,56 и 2,30 у самцов.
После первого периода нереста гонады ставриды переходят в VI-III стадию зрелости (гонадосоматический индекс 1,60 у самок и 1,46 у самцов). После непродолжительного нагула, за время которого младшие размерные группы ооцитов достигают дефинитивного состояния, ставрида вновь вступает в нерест, и по окончании его гонады переходят в стадию выбоя VI-II, завершающую годовой репродуктивный цикл [Воронина, Елисеева, 1985].
Асинхронность в созревании одноразмерных группировок ставрид в северном и южном подрайонах указывает на нарушение панмиксии, а это обстоятельство свидетельствует о возможном существовании двух популяций перуанской ставриды в открытой части океана.
Наши наблюдения показали, что зимой (июль - август) независимо от размерного состава перуанская ставрида в южном подрайоне распределяется преимущественно между 42° и 45°ю.ш. С наступлением весны она постепенно смещается в северном направлении к местам нереста (34-39°ю.ш.). Первыми на нерест подходят крупные особи, которые, следовательно, и нерестятся при более низких температурах, чем особи более мелкие. Так, например, особи со средними размерами 52-54 см нерест начинают при температуре воды на поверхности 10,2-12,3°С, а с размерами 46-50 см - при 12,5-15,5°С. К концу сентября в уловах доминируют нерестующие особи еще более мелкие, а крупные составляют незначительную часть улова.
В юго-западной части Тихого океана нерестующиеся ставриды впервые были отмечены на 135°в.д., при этом центр нереста смещается на восток. По-видимому, мелкие особи нерестятся позже крупных, распространенных далее к западу. Так, например, если в районе 132°з.д. в середине января нерест в основном закончился, и ставрида рассредоточилась на большой акватории, то восточнее в феврале она в массе еще находилась в нерестовом состоянии, причем нерест ее проходил при более высокой температуре воды на поверхности - 15,5-18,5°С, и только в конце февраля наметилась тенденция к его затуханию.
В общем, чем ближе конец нерестового периода, продолжающегося с весны до середины лета, тем меньше средние размеры нерестующих особей.
По нашим наблюдениям, в течение нереста ставрида отметала четыре порции икры и промежуток между ними ориентировочно составил 25-40 суток. Вымет икры длится не более 2-3 суток, остальное время ставрида находится на VI-IV и VI-IV-V стадиях зрелости половых продуктов. В декабре в западной части района в уловах встречалось до 5-10% отнерестившихся особей (на стадии VI-II), причем значительную их часть составляли крупные рыбы. В районе 130-135 з.д. в начале февраля крупных нерестующих особей было уже не более 5-7%, в то время как на востоке юго-западной части Тихого океана они составляли до 40-60%. И только когда их осталось не более 15%, ставрида стала смещаться в южном направлении к местам нагула (февраль).
На западе ареала (45°15' - 45°25' ю.ш. и 155°20' - 152°54' з.д) уже в начале января перуанская ставрида полностью отнерестилась, и 10-15% ее находилось на стадии VI-II и 85-90% - на стадии II-III. Впервые нагульная перуанская ставрида на 47° ю.ш. и 135° з.д. была отмечена в конце января.
Для перуанской ставриды, по-видимому, характерны сезонные миграции к местам нагула и нереста. С окончанием нереста в северной части ареала между 34° и 39° ю.ш. ставрида мигрирует в южном направлении к субантарктическому фронту, располагающемуся между 46° и 50° ю.ш. Следует заметить, что хотя наиболее благоприятной для нереста считается температура воды 14,0-16,5°С, незначительная часть крупной ставриды нерестится и при более низкой температуре. В дальнейшем, по-видимому, икра и личинки ставриды дрейфуют с течением в восточном направлении, причем наибольшая часть их попадает в южный подрайон. Подрастая, молодь ставриды начинает более активно мигрировать в западном направлении. Таким образом, океаническая группировка перуанской ставриды постоянно пополняется рыбой из южного подрайона; при этом с продвижением на запад количество ее заметно убывает. Обратно половозрелая ставрида из западной океанической части в восточный океанический подрайон, и тем более на шельф, по-видимому, не возвращается.
Установлено, что нерест ставриды в западных участках юго-западной части Тихого океана начинается примерно на 1,5 мес раньше и отнерестившиеся особи уходят на нагул раньше, чем в восточных участках.
Зона нереста перуанской ставриды более размыта, чем нерестилища сардины и анчоуса. Ее икра рассеивается на большой площади. Было установлено, что ставрида нерестится в зонах с сильными турбулентными течениями [Bakun, Parrish, 1982]. Известно, что многие пелагические виды рыб, например, сельдевые и скумбрии, выбирают для своего нереста наиболее стабильные зоны. Ставрида же выискивает на первый взгляд наиболее неблагоприятные зоны для нереста. Максимальное число икринок на станцию составило 2400, что является относительно небольшой величиной по сравнению с 40000 икринок сардины [Serra, 1983].
Перуанская ставрида размножается в водах Перу и Чили до 41°ю.ш. [Абрамов, Котляр, 1980; Serra, 1983]. Максимальные концентрации икры и мальков наблюдаются в прибрежных водах у Антофагасты и далее к югу. В ходе исследований вдоль чилийского побережья, проведенных на расстоянии до 200 миль, отмечена значительная плотность распределения икры и мальков без заметного уменьшения ее в западном направлении [Serra, 1983].
Репродуктивный ареал перуанской ставриды вне 200-мильной зоны простирается в юго-восточных водах Тихого океана от 3° до 43°ю.ш. [Горбунова и др., 1985]. В сороковых широтах почти на всех ихтиопланктонных станциях в период нереста обнаруживаются личинки ставриды, в марте 1982 г. основу ихтиопланктона эпипелагиали района между 40-43°ю.ш. составляли ее икринки и личинки. Они обнаруживались на каждой станции в верхнем 50-метровом слое над термоклином. Численность икринок ставриды была довольно высокой (от 50 до 279 экз/м2). В субантарктической фронтальной зоне интенсивный нерест перуанской ставриды проходил как в 300 милях, так и в 600 милях от побережья. Высокая численность икры ставриды в мористых участках позволяет предполагать наличие нереста этого вида и в более западных районах сороковых широт. О.Рохас [Rojas, 1980] при анализе проб ихтиопланктона обнаружил высокую концентрацию личинок перуанской ставриды на траверсе Талькахуано на расстоянии до 900 миль от берега. Средняя плотность распределения личинок составляла 23 экз. на станцию, что является довольно значительной величиной для данных мест. Наши исследования показали, что икра перуанской ставриды встречается в большом количестве на 160° з.д., примерно в 3600 милях от берегов Чили и в 600 милях от Новой Зеландии [Некрасов, 1992б].
Исследовано питание личинок перуанской ставриды длиной от 2,2 до 9,0 мм и массой 0,04-5,6 мг. Установлено, что у личинок лишь при длине 3,0 мм, как правило, полностью резорбируется желточный мешок и имеется хорошо развитый кишечник. С этого момента они переходят на активное питание. Возраст таких личинок составляет 3-4 суток. Скорость роста личинок в условиях эксперимента до перехода на активное питание - примерно 0,3 мм в сутки [Липская, 1985].
Постоянными компонентами в пище личинок перуанской ставриды являются циклопоиды, составляющие в июле - августе 54%, а в ноябре - декабре 57% всей потребляемой пищи. В основном это различные виды Oncaea. Из копепод 24,4% составляют Vicrosetella rosea. Отмечена четкая тенденция повышения размеров потребляемых пищевых организмов с увеличением длины личинок. Личинки длиной 8-9 мм питаются в основном взрослыми формами копепод размерами до 1,1 мм, тогда как личинки длиной 3,5 мм потребляют пищевые организмы размерами 0,08-0,15 мм. Следовательно, увеличение размера личинки примерно втрое (от 3,5 до 9 мм) обеспечивает ей возможность использовать пищевые организмы размерами 0,08-1,12 мм, т.е. пищевые возможности их примерно в семь-восемь раз выше, чем у личинок длиной 3,5 мм, что в конечном счете обусловливает лучшее их выживание на ранних стадиях развития [Липская, 1985].
Взрослые особи перуанской ставриды питаются преимущественно эуфаузиидами, креветками, гипериидами, рыбами, крылоногими моллюсками, кальмарами, сальпами, гребневиками и медузами. В дневных уловах желудки перуанской ставриды более наполнены [Lambert, Sievert, 1982]. Следует отметить, что ставрида продолжает питаться и в период нереста, правда, интенсивность питания снижается. Если в нерестовый период средний балл наполнения желудков ставрид составляет 0,24, то в начале нагула - 1,6.
Исследования немецких ученых показали, что содержание жира в перуанской ставриде колеблется от 0,5 до 7,0%. Примечательно, что выловленная в южных районах ставрида обладает довольно высокой жирностью. Во время нереста жирность ставриды падает до минимума [Lambert, Sievert, 1982]. Жиронакопление на внутренностях ставриды хорошо согласуется с сезонами. Во всех подрайонах наибольшая степень жирности приходится на осенне-зимний период и минимальная жирность - на весну. Обращает на себя внимание также и то, что у группировок ставрид, обитающих в более холодных водах, степень жиронакопления выше.
На соотношении полов у перуанской ставриды необходимо остановиться особо. В большинстве научных отчетов о работе в рейсах научно-поисковых и научно-исследовательских судов в ЮВТО, посвященных изучению биологии перуанской ставриды, и о многочисленных наших наблюдениях отмечено, что в океанической части ареала преобладают самцы. Это интересно хотя бы потому, что известно огромное количество противоположных примеров.
У многих видов рыб соотношение полов в нерестовой части популяции близко к равному [Никольский, 1961; Правдин, 1966]. И вообще в популяциях многих двуполых организмов соотношение самцов и самок примерно равное [Пианка, 1981]. Для полной точности различают соотношение полов: первичное (пренатальное, или эмбриональное), вторичное - соотношение полов у потомства при рождении (для рыб - постэмбриональная и мальковая фазы) и третичное - соотношение полов у взрослых, размножающихся особей [Геодакян и др., 1967]. Обычно, когда специально не оговаривают, под соотношением полов в популяции имеют в виду третичное соотношение полов. Ч.Дарвин полагал, что соотношение полов, равное 1:1, выгодно группе, так как в пределах каждого пола оно до минимума сокращает конкуренцию за полового партнера [цит. по: Пианка, 1981]. Р.А.Фишер [Fisher, 1930] показал, что для видов, выращивающих свое потомство, наилучшие результаты получаются, если в возрасте половой зрелости достигается соотношение самцов и самок 1:1. Другими авторами было высказано мнение, что соотношение полов 0,5 (доля самцов в популяции) в репродуктивном возрасте максимально облегчает встречу особей противоположного пола и способствует понижению степени инбридинга [Геодакян и др.,1967].
И тем не менее, несмотря на все перечисленные преимущества равного соотношения полов, мы редко встречаем популяции, в которых соотношение полов равно 1:1. Например, по данным Е.К.Суворова (1948), это соотношение следующее. Волжский лещ: самцов 47,0% - самок 53,0%; треска Охотского моря: самцов 41,4-50,5 - самок 58,6-49,5%.
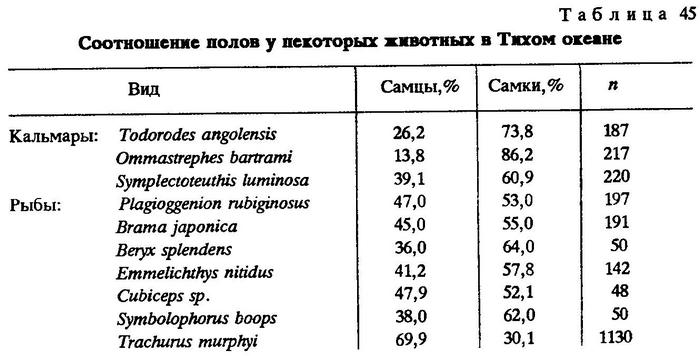
По нашим наблюдениям в 1982 г. в 4-м рейсе НПС "Возрождение", в Тихом океане за пределами 200-мильных экономических зон соотношение полов у большинства различных животных было таким, что, как и в примере Е.К.Суворова (1948), в популяциях преобладали самки. Лишь в популяции перуанской ставриды значительно преобладали самцы (табл.45).
С точки зрения нового подхода соотношение полов в популяции - переменная величина, тесно связанная с условиями среды и определяющая эволюционную пластичность вида. Очевидно, соотношение полов в популяции отражает специфику взаимосвязей популяции данного вида со средой. Оно не остается постоянным и в различных частях ареала вида. Половая структура популяции вида - такое же его приспособительное свойство, как и все другие признаки и свойства вида [Никольский, 1965]. Именно поэтому в стабильной эволюционной ситуации "вековой" оптимум соотношения полов устанавливается на относительно низком уровне (меньше самцов), а некоторые виды или популяции вида даже полностью утрачивают самцов.
В связи с этим большой интерес вызывает обнаруженное нами значительное преобладание по численности самцов у перуанской ставриды в юго-западной части Тихого океана (см. табл.45).
Известно, что распространение большинства видов рода Trachurus ограничено неритическими зонами, пределы которых они не покидают на протяжении всего жизненного цикла. Лишь несколько видов этого рода, среди них и перуанская ставрида, встречаются не только на шельфе, но и на расстоянии сотен и даже тысяч миль от побережья.
Таким образом, если концепция В.А.Геодакяна верна и в стабильной эволюционной ситуации "вековой" оптимум соотношения полов представлен преобладанием самок, то мы должны сделать вывод, что популяция перуанской ставриды в юго-западной части Тихого океана обитает в лабильных условиях. Однако, согласно концепции В.А.Геодакяна, при ухудшении условий среды в популяции должны гибнуть в большем количестве самцы, и в связи с этим возникает вопрос: почему самцов в популяции, несмотря на это, больше?
Чтобы ответить на этот вопрос, важно знать механизм изменения соотношения полов, в какую сторону и при каких условиях происходит изменение половой структуры.
Прежде всего следует обратить внимание на преобладание самцов в популяциях перуанской ставриды, обитающих не только в океанической эпипелагиали юго-западной части Тихого океана, но и у обитающих на шельфе. Наши наблюдения показали преобладание самцов в океанических популяциях перуанской ставриды, что имеет место в течение нескольких поколений и в различных районах.
Конечно, преобладание самцов в скоплении перуанской ставриды в юго-западной части Тихого океана, на периферии ареала вида, можно объяснить, по-видимому, следствием миграции от Американского континента в сторону Новой Зеландии. Известно, что в период миграций иногда замечается переформирование соотношения полов в скоплениях рыб, так как самцы активнее и подвижнее самок [Шмидт, 1936; Асланова, 1952]. Однако такого объяснения явно недостаточно, когда мы знакомимся с соотношением полов скоплений ставриды в южном подрайоне юго-восточной части Тихого океана, где в течение всего года последовательно встречаются нерестующиеся, нагульные и преднерестовые особи ставриды. К тому же мы располагаем данными из шельфовых участков ареала этого вида, полученными в 1973 г. в 12-м рейсе НПС "Академик Книпович" в водах Чили. Эти данные говорят о том, что в то время на шельфе в популяции перуанской ставриды тоже преобладали самцы. Соотношение полов составляло 51,7% : 48,3% (n = 1716).
Наконец, чтобы убедиться в том, не является ли преобладание самцов в популяциях во всем роде Trachurus обычным, мы привлекли для сравнения наши материалы по другим видам этого рода, собранные в разные годы в Индийском и Атлантическом океанах (табл. 47).
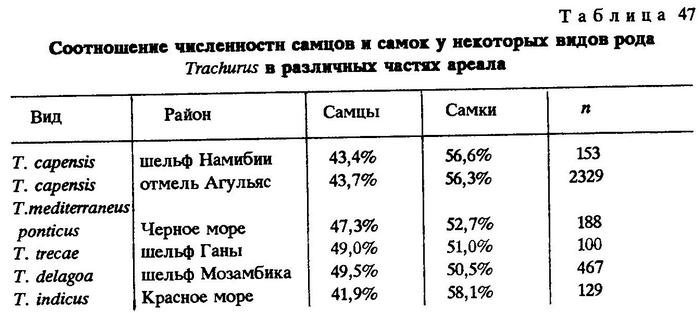
Как видно, у большинства видов рода Trachurus в популяциях преобладают самки. Итак, объяснение высокого уровня соотношения полов у океанических популяций перуанской ставриды только миграциями представляется недостаточным и неубедительным. Возможно, это является следствием пока нам неизвестных суровых условий среды обитания вида. Преобладание самцов в популяции перуанской ставриды, обитающей на шельфе, очевидно, свидетельствует об изменчивой среде ее обитания, что, по-видимому, вынуждает ставрид осваивать новые биотопы в открытом океане. Известно, что перуанская ставрида, как и калифорнийская, выходит далеко за границу континентального шельфа, на расстояние многих сотен миль от него [Roedel, Fitch, 1952; Ahlstrom, 1956; Алеев, 1957а]. Н.В.Парин (1968), рассматривая в сравнительном аспекте состав ихтиофауны открытой и прибрежной эпипелагиали, пришел к выводу, что основная масса рыб, слагающих ихтиофауну открытой эпипелагиали, принадлежит к филогенетическим молодым группам. Очевидно, это справедливо и для перуанской ставриды.
Скопления перуанской ставриды, относительно недавно вышедшие в открытую эпипелагиаль, обладают высокой пластичностью, что дает возможность освоить новые биотопы, отвечая еще более высоким уровням соотношения полов, т.е. большим количеством самцов в популяции.
Те же тенденции наблюдаются и при более кратковременных изменениях среды в пределах одного поколения. В экстремальных условиях среды третичное соотношение полов падает, т.е. снижается количество самцов в популяции.
По-видимому, для объяснения описанной ситуации нужен иной подход.
Мы предприняли попытку на примере перуанской ставриды выяснить природу данного явления.
Нам представилась возможность изучить размерно-половое соотношение в океанических популяциях перуанской ставриды, обитающих в северном, южном и западном районах ареала. Скопления перуанской ставриды были образованы различными размерными группами рыб и в траловых уловах представлены особями длиной от 21 до 65 см [Некрасов, Тимохина, 1987].
Размерно-половое соотношение в популяции перуанской ставриды, обитавшей в 1980 году в южной океанической части ареала, показано на рис.22а. Видно, что среди мелкоразмерных особей длиной 26-28 см в возрасте 3 лет, самцов было 33,3%, а самок 66,7%. С увеличением длины рыб это соотношение изменялось в сторону увеличения численности самцов, и среди рыб длиной 44-50 см в возрасте 8-11 лет самцы составляли уже 79,6-72,7%.
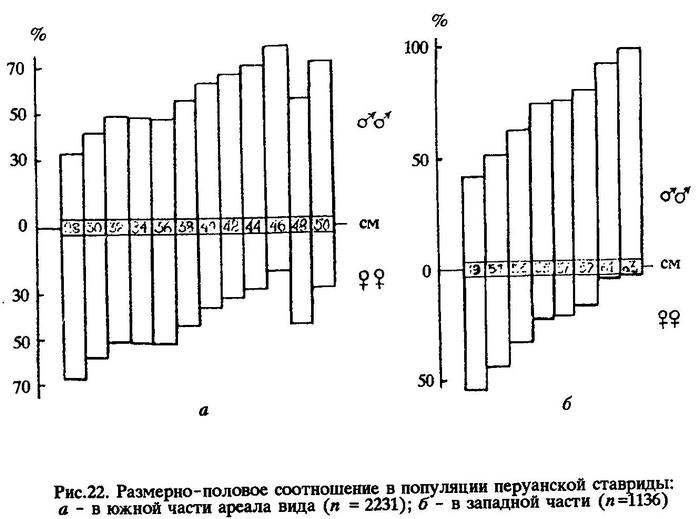
Аналогичное размерно-половое соотношение наблюдалось в 1980 г. в северной океанической популяции (табл. 48) с той лишь разницей, что здесь тенденция к увеличению относительной численности самцов среди особей мелких размеров обнаруживалась уже у рыб длиной 25-27 см (59,2%), а среди рыб длиной 36-38 см в возрасте 5-6 лет самцов было 64,3%.
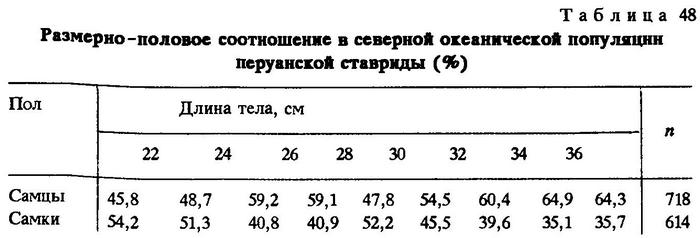
Следовательно, южная и северная океанические популяции перуанской ставриды имеют характер размерно-половых соотношений третьего типа. Но наиболее четко закономерность увеличения числа самцов к концу жизни выражена в океанической популяции перуанской ставриды, обитающей в западной части ареала. На рис.22б видно, что среди рыб длиной 60-62 см в возрасте 16-18 лет самцы составляют 93,9%, а в размерной группе 62-64 см (19 лет и старше) вообще нет самок.
В 1981 г. в юго-восточной части Тихого океана за пределами 200-мильной экономической зоны Перу и Чили характер размерно-половых соотношений у популяций перуанской ставриды также был третьего типа (табл.49).
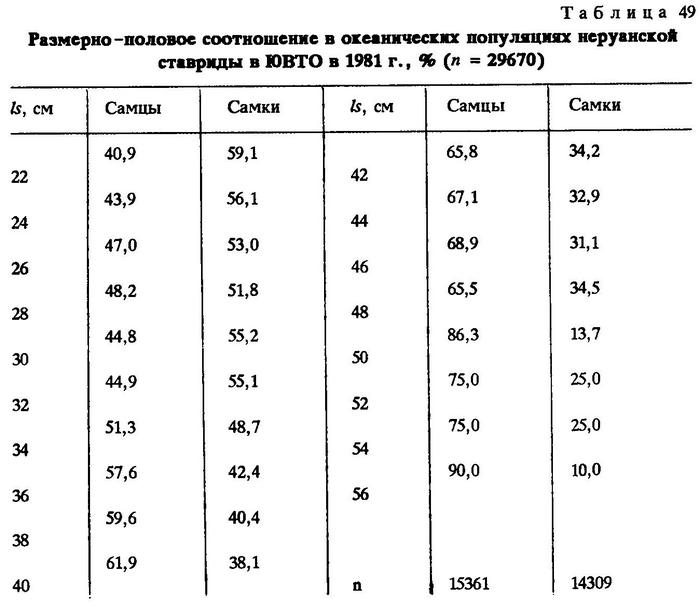
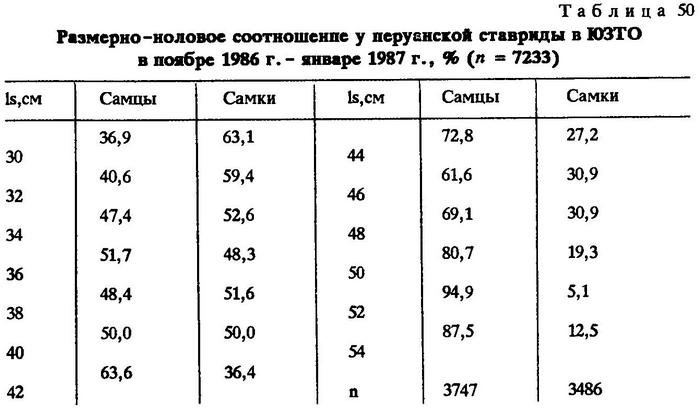
Аналогичная картина наблюдалась нами и в 1986-1987 гг. (табл.50).
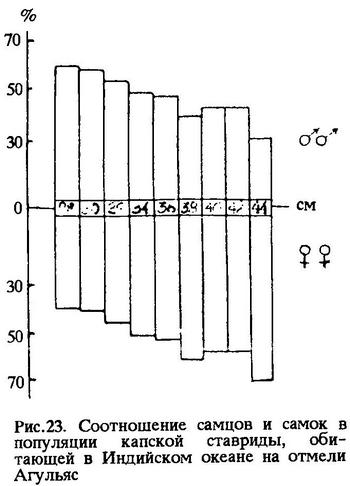
Интересно отметить, что у других видов рода Trachurus, обитающих на шельфе, например, у Trachurus mediterraneus ponticus, Trachurus trecae, Trachurus delagoa, Trachurus capensis размерно-половое соотношение второго типа. Наиболее наглядно это видно на примере популяции капской ставриды Trachurus capensis, обитающей на отмели Агульяс (рис.23) (n = 2329 экз.). Если при длине тела капской ставриды 38-42 см в возрасте 7-10 лет самцы составляли лишь 41,6-41,4%, а среди более крупных рыб длиной 42-44 см в возрасте 12 лет и более лет самцов было всего 28,6%, то среди рыб длиной 26-30 см в возрасте 3-4 года доля самцов составила 59,2-58,0%.
He только у ставрид, включая перуанскую, к концу жизни особей наблюдается увеличение количества самок. Так, например, соотношение полов у морской камбалы Pleuronectes platessa в Северном море следующее: среди молоди длиной 6-7 см на 100 самок приходится 150 самцов; чем крупнее камбала, тем меньше попадается самцов, а среди рыб длиной 67-70 см встречаются только самки. То же наблюдается и у других камбал [Суворов, 1948; Шатуновский, 1963].
И вот на фоне изложенного неожиданным противоречием открывается перед нами картина размерно-полового соотношения в океанических популяциях перуанской ставриды к концу жизни в сторону увеличения доли самцов. Чем объяснить это нетипичное явление, когда при третичном соотношении полов с возрастом увеличивается относительная численность самцов? Почему самок с возрастом гибнет больше, чем самцов, и как это увязывается с изложенной выше концепцией В.А.Геодакяна, согласно которой все должно быть наоборот?
Значит, более тесная связь особей мужского пола со средой обеспечивается большей величиной их фенотипической дисперсии по сравнению с особями женского пола. Типичной иллюстрацией к этому служит размерно-половое соотношение в популяции капской ставриды, обитающей на отмели Агульяс (рис.23). В этом случае кривые смертности имеют вид, изображенный на рис.24. По оси абсцисс - устойчивость к вредному фактору среды; по оси ординат - количество особей с данной устойчивостью. На рисунке видно, что кривая смертности женских особей имеет положительный эксцесс, а мужских - отрицательный. И это понятно, потому что у мужских особей разнообразных отклонений от моды популяции больше, чем у женских, а за счет этого образуется зона дискомфорта (Зд). Зона дискомфорта тем шире, чем популяция пластичнее. На рис.24а показано, что вредный фактор среды (Ф - фронт вредного фактора) элиминирует малую часть популяции (заштриховано на рисунке), состоящую преимущественно из самцов. Именно поэтому к концу жизни поколения в популяции капской ставриды остается больше самок. Такая форма связи со средой ценой высокой смертности самцов, как уже отмечалось выше, выгодна для популяции в целом.
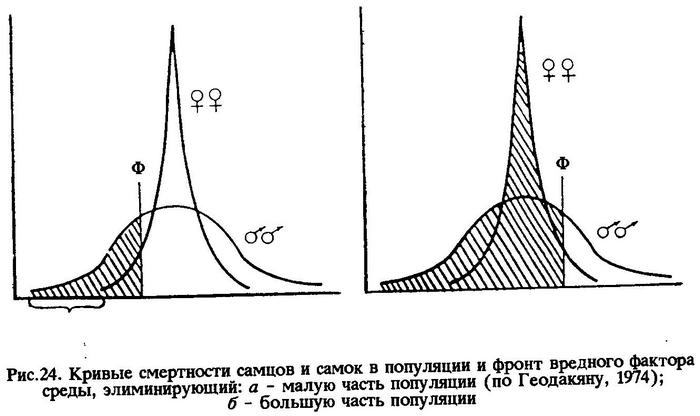
В случае с океаническими популяциями перуанской ставриды, в которых к концу жизненного цикла поколения остается больше самцов, мы обязаны были бы предположить, что, при прочих равных условиях, фенотипическая дисперсия больше у особей женского пола. И это явилось бы редчайшим исключением в природе. Но исключения здесь нет.
Принимая во внимание то, что популяция на изменение условий среды закономерно отвечает изменением половой структуры, мы вправе считать, что условия обитания популяций перуанской ставриды нестабильные. Нестабильные условия среды и обусловливают численное преобладание самцов. На океанические популяции перуанской ставриды вредный фактор (или факторы) среды, вероятно, действует постоянно и элиминирует большую часть популяции (см.рис.24б). Но так как фенотипическая дисперсия у самцов выше, чем у самок, то имеется широкая зона дискомфорта не только в левой части вариационной кривой, но и в правой. И, стало быть, при более мощном воздействии вредного фактора в правой части остается больше самцов. Именно поэтому к концу жизненного цикла поколения перуанской ставриды, обитающей в океанической части ареала, остается больше самцов.
Изложенная трактовка образования трех типов размерно-половых соотношений у рыб объясняет, почему большинство видов рыб имеет соотношение полов второго типа. Виды рыб с первым и третьим типами соотношения полов испытывают сильный пресс вредных факторов среды, поэтому, чтобы противостоять этому, они должны были либо выработать в процессе эволюции чрезвычайно высокую плодовитость, либо приспособиться к охране своего потомства. Эволюция перуанской ставриды пошла по первому пути.
Хозяйственное значение. Промысел перуанской ставриды ведется довольно интенсивно как на шельфе, так и в открытой части океана. Только вылов Чили и Перу превышает 2 млн.т. Особенно оживился промысел после катастрофического падения уловов анчоуса в Перу и Чили в начале 70-х годов. В последнее время уловы перуанской ставриды имеют тенденцию к увеличению. Установлено, что в период Эль-Ниньо 1982-1983 гг. произошли изменения в распределении ставриды. Впервые за многие годы исследований ее обнаружили у берегов Эквадора.
В последние годы вылов перуанской ставриды у берегов Перу достиг 200 тыс.т, а у берегов Чили - приблизился к 2,5 млн.т [Yearbook..., 1992].
По данным Перу, возможный ежегодный вылов перуанской ставриды и скумбрии в 50-мильной прибрежной зоне оценивается в 2,5 млн.т, а общие запасы ставриды в 1984 г. составляли 8,5 млн.т против 4,5 млн.т в 1981 г. [Another fisheries..., 1984].
Промысел перуанской ставриды в Чили начался относительно недавно. В 1977 г. от моновидового промысла анчоуса был совершен переход к поливидовому, что вызвано в основном увеличением численности популяций сардины, ставриды и скумбрии. С 1978 г. перуанская ставрида занимает в Чили второе место в общем вылове рыбы, и ее добыча в этом году составила 586,7 тыс.т, в 1979 г. - 597,5 тыс.т, в 1984 г. - 1,4 млн.т, в 1990 г. - 2,5 млн.т [Yearbook..., 1992].
Кроме местного промысла, в водах ЮВТО ведется и экспедиционный лов ставриды другими странами в основном за пределами 200-мильных экономических зон прибрежных государств (табл.51) [FAO, 1985, 1989, 1992].
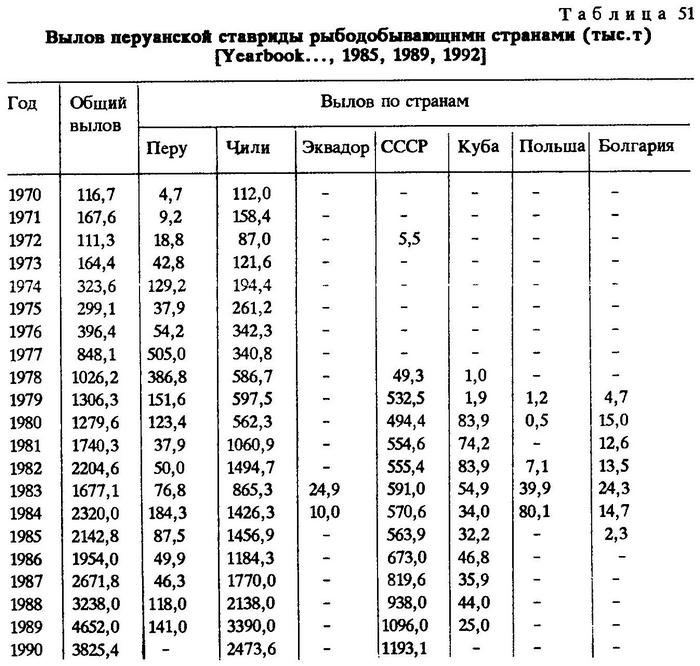
Динамика развития советского промысла перуанской ставриды протекала следующим образом. В 1978 году ВРПО "Запрыба" была организована поисково-промысловая экспедиция в ЮВТО на РТМС "Звезда". В результате успешных поисковых работ за пределами 200-мильной экономической зоны Перу были обнаружены большие концентрации перуанской ставриды. Судно имело промысловые уловы. Это послужило поводом для формирования нового для отечественного океанического рыболовства промыслового района.
В сентябре 1978 г. в ЮВТО были размещены первые промысловые суда, и к концу года было добыто 49,3 тыс.т ставриды, а в конце января 1979 г. вылов ее уже составил 532 тыс.т. С тех пор промысел перуанской ставриды в ЮВТО стал круглогодичным.
В 1985 году существенно возрастает значение промысла перуанской ставриды в новом районе - в юго-западной части Тихого океана.
Сырьевая база в ЮВТО в районе 3-30°ю.ш. отличается неустойчивостью. Промысел сезонный (с июня по декабрь), а в период явления Эль-Ниньо он полностью отсутствует.
На участке ЮВТО между 30° и 50°ю.ш. промысловый запас перуанской ставриды позволяет вести крупномасштабный промысел, но условия здесь более сложные по сравнению с северным участком - большая частота штормовых дней, значительная удаленность от базового порта Кальяо (Перу). Поэтому в те годы, когда возможен промысел между 3° и 30°ю.ш., в районе 30-50°ю.ш. вылов сокращается из-за перевода части промысловых судов на северный участок ЮВТО.
Еще более сложными являются условия промысла в ЮЗТО, западнее 105°з.д. Рыба здесь ловится более крупная, и для ее облова требуется большая скорость траления, что не всем судам доступно. Поэтому в ЮЗТО промысловый период продолжается всего 2-3 месяца, в то время, когда в ЮВТО промысловая обстановка ухудшается.
Вылов перуанской ставриды отечественными судами отличался высокой стабильностью, но с 1992 г. он полностью прекратился.
Промысел ставриды в ЮВТО велся на сильно разреженных скоплениях на большой акватории. Разноглубинный траловый лов является единственным видом лова, обеспечивающим эффективный промысел [Дондуа и др., 1984]. При выборе типа судна для успешного ведения промысла в ЮВТО, очевидно, следует учитывать, что в осенне-зимний период в районе южнее 35°ю.ш. наблюдаются частые штормы и крупная зыбь. В таких погодных условиях вести промысел могут только суда, обладающие достаточной мощностью силовой установки. Наиболее результативными являются суда типов РТМС, БАТМ, БАТГ, БАТСП, БМРТИБ. Эти суда имеют возможность маневрировать с тралом и делать сравнительно быстрые переходы из одного района в другой. Скорость передвижения перуанской ставриды достаточно высокая; для облова ее все суда работают по следующей схеме. Вначале идут курсом на ветер с поиском скоплений, пригодных для облова. Затем разворачиваются на обратный курс, отдают трал и буксируют его по ветру с полным использованием мощности силовой установки. Для проведения очередного траления вновь заходят на ветер, причем по причине подвижности рыбы часто судам приходится вновь искать скопление, которое только что облавливалось. В этом случае скорость хода судна играет существенную роль.
Ставрида облавливается в основном с 18 до 6 ч, поэтому за ночь удается сделать не более двух-трех продуктивных тралений. При усилении ветра суда типа БМРТ и РТМА часто не в состоянии сделать очередной забег и оказываются в пролове [Дондуа и др., 1984].
Наиболее результативными бывают траления с 20 до 4 ч. Во время первого траления рыба держится, как правило, в слое 25-50 м, и верхнюю подбору трала ведут выше косяка на глубине 5-15 м. При втором тралении, особенно под утро, трал опускают ниже косяка. Перуанская ставрида очень пуглива и подвижна, особенно в дневное время. От шума судового винта она быстро уходит на глубину 70-250 м. Поэтому дневные уловы не дают желательных результатов и хорошие дневные подъемы редки. Днем трал следует вести примерно на 130 м ниже косяка [Кавкайкин, 1981].
Учитывая время суточных вертикальных миграций перуанской ставриды и ее активность, более благоприятным периодом тралового лова следует считать ночное время с 20 до 24 ч, когда рыба, поднявшись в верхние горизонты, начинает активно питаться и слабее реагирует на судно и трал (Коротков и др., 1990).
Кроме того, перуанская ставрида, как и другие рыбы, контактируя с орудиями лова, адаптируются к ним, т.е. постоянно происходит процесс ее "обучения", вырабатываются новые формы оборонительных реакций. У перуанской ставриды очень быстро выработалась оборонительная реакция на систему судно - трал. При подходе трала рыбы стали заныривать на глубину 100 м и более, хотя в начале освоения этого района промысла подобных реакций у рыб не наблюдалось [Коротков и др., 1990].
Учитывая высокую подвижность перуанской ставриды, ее повышенную реакцию на внешние раздражители и сложные гидрометеорологические условия в ЮВТО, к тралам и условиям лова должны предъявляться следующие требования: вертикальное раскрытие не менее 50 м, скорость траления не менее 5 узлов, длина вытравленных ваеров при облове рыбы в верхних слоях не менее 400 м и достаточная их прочность для ведения промысла при волнении моря 6-7 баллов и крупной зыби.
Наиболее полно удовлетворяют этим требованиям конструкции тралов, приведенные в табл.52.

Для успешного облова поверхностных (ночных) или более глубоководных (дневных) скоплений ставриды следует менять вооружение и оснастку тралов. На ночное время необходимо настраивать доски "на всплытие", уменьшать загрузку нижней подборы, массу грузов-углубителей и длину регулировочных цепей. Для дневной работы оснастка тралов приводится в прежнее (стандартное) состояние.
Как уже отмечалось, для облова быстроподвижных скоплений ставриды необходимо иметь достаточно высокие параметры устья тралов. При этом большое значение имеет вооружение верхней подборы. Для увеличения вертикального раскрытия тралов верхняя подбора оснащается гидродинамическими щитками либо устройствами "Гиплан-3" [Дондуа и др., 1984].
Перспективы промысла. На сегодня мы имеем много данных по промысловому запасу ставриды в ЮВТО, полученных различными методами. По данным АтлантНИРО, с 1982 по 1985 г. в ЮВТО и ЮЗТО он колебался от 9,8 до 11,7 млн.т. Т.В.Дехник и М.В.Назарова (1988) с помощью ихтиопланктонного метода оценили запас 1983 г. в 10,0 млн.т и 1984 г.- в 13 млн.т. По нашим данным, в ЮВТО и ЮЗТО промысловый запас перуанской ставриды составляет 13-15 млн.т.
По данным А.А.Елизарова с соавторами (1992), последние 5-7 лет биомасса перуанской ставриды находится на относительно стабильном уровне и колеблется в пределах 17-22 млн.т (в северном подрайоне ЮВТО - 1,3-2,4 млн.т, в южном подрайоне ЮВТО и восточной части ЮЗТО - 10-14 млн.т, в центральной и западной частях ЮЗТО - 6-8 млн.т). Для района ЮЗТО (105-165°з.д.) близкие оценки биомассы перуанской ставриды получены по результатам анализа данных тралово-акустических съемок в 1987 г. (около 8 млн.т). Наблюдения показывают, что величина промыслового запаса перуанской ставриды в ЮВТО южнее 30°ю.ш. за последние десять лет не подвергалась резким колебаниям.
Можно легко себе представить, какие потенциальные возможности таит в себе этот промысловый район. Тем не менее, несмотря на столь оптимистичные данные о запасе ставриды в Тихом океане, есть мнение, что перуанская ставрида в открытых водах, возможно, в ближайшем будущем исчезнет. Н.В.Парин (1988) считает, что ставрида в открытых водах ЮВТО представляет собой временную псевдопопуляцию, которая не только существует за счет постоянного пополнения из прибрежного запаса, но и является фактически его излишком. "Никаких свидетельств в пользу результативности нереста не существует, - пишет Н.В.Парин, - так как в этих океанических участках ставрида представлена только очень крупными особями длиной 45-60 см (там нет ни мальков, ни сеголетков, ни созревающих рыб...)". И далее: "Выселение перуанской ставриды в океан объяснимо ростом ее численности в неритической зоне, и рассчитывать на неограниченно долгое существование перуанской ставриды в открытом океане не приходится. Цикличность процесса неизбежно предполагает грядущий спад численности и даже почти полное исчезновение этой рыбы из открытых для международного рыболовства вод" [Парин, 1988, с. 100-101].
Вывод из изложенного однозначный: действительно зачем вести расчеты возможного оптимального, рационального вылова от запаса, если все равно выход ставриды в открытые воды - это биологический тупик? Стало быть, ее можно вылавливать всю - все равно она погибнет, не дав потомства.
Наши данные также подтверждают тот факт, что пополнение промысловой части стада происходит, по-видимому, из вод Чили. Но есть некоторые нюансы.
Прежде всего неизвестен период цикличности. Суждения некоторых ученых о том, что это был выход одного высокоурожайного поколения перуанской ставриды, оказались несостоятельными. В течение двенадцати лет промысла основу его каждый год составляет новое поколение. Но как долго будут выходить в открытый океан новые поколения ставриды? Конечно, можно проводить аналогии с таким видом, как сардина-иваси, появившимся в больших количествах у наших дальневосточных берегов в 30-х годах, затем на многие годы исчезнувшим и вновь появившимся там же в значительном количестве, а теперь вновь исчезающим. Но ведь можно провести и аналогию с распределением ближайшего родственника перуанской ставриды - калифорнийской ставридой (Trachurus symmetricus). Примерно 2 млн.т калифорнийской ставриды с момента ее обнаружения в открытом океане в начале 50-х годов до наших дней ежегодно обитает в океанической зоне вод США. Нередки случаи ее появления на расстоянии 1500 миль от континента [Некрасов, Каратаева, 1987]. По-видимому, можно предположить, что она существовала там и раньше многие годы, но технические возможности не позволяли ее обнаружить. Почему бы не предположить, что такое явление имеет место и у перуанской ставриды в ЮВТО?
Немаловажным фактом является и то, что нерест перуанской ставриды в открытом океане все-таки результативен. Еще в 1980 г. в ЮВТО во время экспедиции на НИС "Профессор Месяцев" обнаруживали мальков длиной 5-13 см. Мальков таких же размеров ловили и другие поисковые суда (НПС "Сократ", 1980). На 117°з.д. и 38-40°ю.ш. в экспедициях на РТМС "Полтава" и НПС "Плунге" обнаруживали молодь ставриды уже в 1979 г. Ежегодно облавливаются особи ставриды размерами 11-17 см на участке между 10° и 30°ю.ш. Нами в экспедиции на РТМС "Возрождение" еще в 1982 г. была обнаружена в ЮЗТО развивающаяся икра перуанской ставриды, описанная С.А.Евсеенко; в 1987 г. нами в большом количестве облавливались икра и личинки перуанской ставриды на акватории от 160°з.д. и восточнее, вплоть до вод Чили. В 11-м рейсе РТМС "Возрождение" 17 апреля 1987 г. в координатах 35° 06'ю.ш. и 100° 05'з.д. улов молоди ставриды со средними размерами 10,1 см составил 100 кг. Минимальная длина рыб была 9 см, максимальная - 12 см.
Факт увеличения средних размеров перуанской ставриды, мигрирующей в западном направлении в ЮВТО и ЮЗТО, свидетельствует, по-видимому, о том, что ставрида не возвращается на восток. Но из этого все-таки отнюдь не следует, что вся популяция ставриды существует в открытом океане только благодаря тем особям, которые размножаются в прибрежных водах Южной Америки.
Нам представляется вполне допустимым, что особи перуанской ставриды, родившиеся в открытых частях ЮВТО и ЮЗТО и ежегодно обнаруживаемые в каждой экспедиции на различных стадиях развития, переносятся периферийными струями Антарктического циркумполярного течения (АЦТ) в экономические зоны Чили и Перу.
Известно, что общий перенос водных масс АЦТ имеет устойчивое восточное направление со средней скоростью 20- 40 см/с [Зарипов, Кизнер, 1983], а по другим данным, на 40°ю.ш. и 125°з.д. - со средней скоростью 30 см/с [Саруханян, 1980]. В связи с этим можно предположить, что мальки перуанской ставриды со средними размерами 3,5 см, пойманные в 10-м рейсе РТМС "Возрождение" на 137°з.д., развились из икры, выметанной в еще более западном участке. Зная скорость течения, можно рассчитать скорость переноса ихтиопланктона от наиболее западного места обнаружения нами икры ставриды на 160°з.д. до 137°з.д., т.е. до места первого обнаружения мальков. Это расстояние, равное примерно 1080 милям, ихтиопланктон с течением АЦТ преодолеет за 77,3 сут, или за 2,5 мес. Аналогичные расчеты показывают, что от 160°з.д. ихтиопланктон и мальки будут перенесены к 115°з.д. за 5 мес, а до вод Чили - за 9 мес. Также известно, что к концу первого года жизни перуанская ставрида достигает длины примерно 18 см. Если даже допустить, что рост рыбы на первом году жизни равномерный, то средний прирост за месяц составит 1,5 см. Следовательно, за 2,5 мес переноса ихтиопланктона от 160°з.д. до 137°з.д. ставрида может достичь уже длины в среднем 3,7 см. Этот размер близок к тем размерам, которые наблюдаются у мальков на этой долготе. На 115°з.д. ставрида, перенесенная АЦТ от 160°з.д., через 5 мес достигает средней длины 7,5 см, а за 9 мес - уже у вод Чили - 13,8 см. Именно таких размеров рыб мы и ловили на 115°з.д. и восточнее.
Следует отметить, что в ЮВТО между 30° и 50°ю.ш. двухлетки в промысловых уловах наблюдаются очень редко, да и то лишь на участках, прилегающих к водам Чили. По-видимому, они в основной своей массе, так же как и сеголетки, заносятся АЦТ в воды Чили. Но уже трех- и четырехлетки, способные энергично противостоять потоку АЦТ, в открытых водах ЮВТО составляют основу промысла.
Следовательно, признавая, что хотя промысловый запас в ЮВТО и ЮЗТО и формируется, по-видимому, за счет эмиграции перуанской ставриды из вод Южной Америки, мы, однако, не можем исключить и возможности значительного пополнения запаса за счет особей, родившихся в открытом океане.
Даже если допустить, что далеко не весь промысловый запас в ЮВТО и ЮЗТО пополняется рекрутами, родившимися в открытом океане, мы уже не вправе рекомендовать промышленности вылавливать всех взрослых рыб в ЮВТО-ЮЗТО, так как это образование ставрид в океанической зоне не является биологическим тупиком (рис.25).
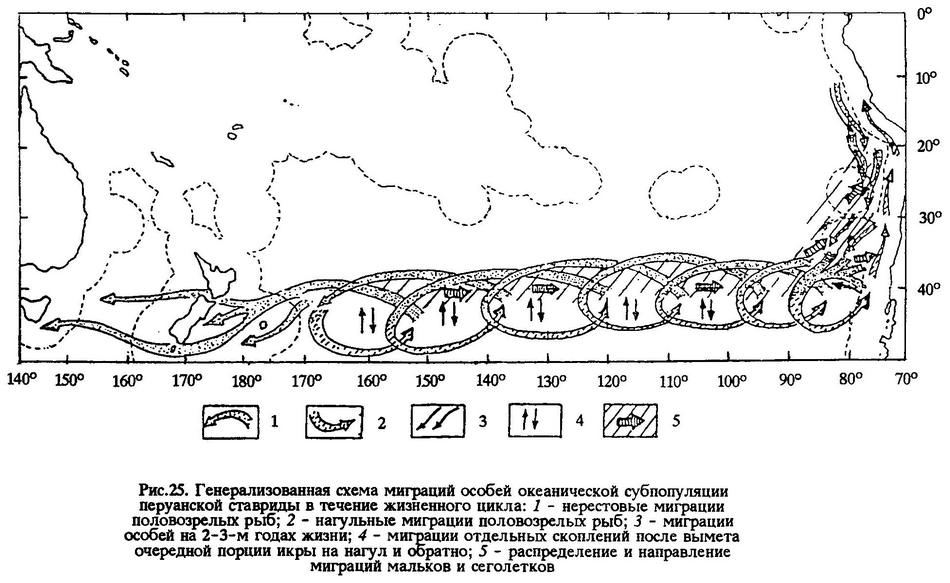
Думается, что этих рассуждений достаточно для того, чтобы не отказываться от попыток обосновывать величины оптимального вылова перуанской ставриды в открытых водах океана с целью рационального ведения промышленного рыболовства.
Одной из основных задач, стоящих перед специалистами по сырьевым ресурсам, является определение оптимальной интенсивности вылова. Известно, что отношение величины улова к промысловому запасу года дает представление о степени использования запаса промыслом (интенсивность вылова). Однако, не зная, каковы естественная смертность рыб и прирост массы в разном возрасте, а также каков предельный возраст рыб, нельзя решить и вопрос о том, с какой интенсивностью и с какого возраста следует вылавливать данную рыбу, чтобы ее улов по массе (а не по счету) был наибольшим из возможных.
Но если получение данных о годовых приростах массы рыб не представляет никаких затруднений, то оценить величину естественной смертности несравненно сложнее. Коэффициенты естественной смертности чаще всего весьма условны, потому что они получены не на основании фактических данных о предельном возрасте рыб, а путем подсчетов, основанных на теории вероятности и не учитывающих биологические особенности рыб. К тому же часто принимается один и тот же коэффициент для всех возрастов, тогда как на самом деле размеры естественной смертности в разном возрасте неодинаковы.
Основное препятствие в определении норм естественной убыли промысловых рыб - это промысел, нарушающий естественное соотношение возрастных групп в стаде. Но для суждения о том, каковы могут быть возрастной состав перуанской ставриды в открытой части океана и максимальный возраст ее, мы располагаем уникальной возможностью. Мы воспользовались определениями возраста рыб, сделанными на девственном стаде, т.е. на стаде, которое еще не эксплуатировалось промыслом. Такой материал был нами собран в ЮЗТО в 1982 г., т.е. до начала промысла перуанской ставриды в этом районе. Кроме того, здесь еще не успело сказаться возможное влияние промысла ставриды в ЮВТО, так как облавливаемые здесь в 1979-1980 гг. поколения еще не подошли к 146°з.д., где брались пробы на возраст.
Оценка естественной смертности перуанской ставриды в ЮЗТО сделана на основании 1730 определений возраста. Для суждения о возрастной структуре скоплений послужили данные о длине тела 20465 экз. рыб (табл.53).
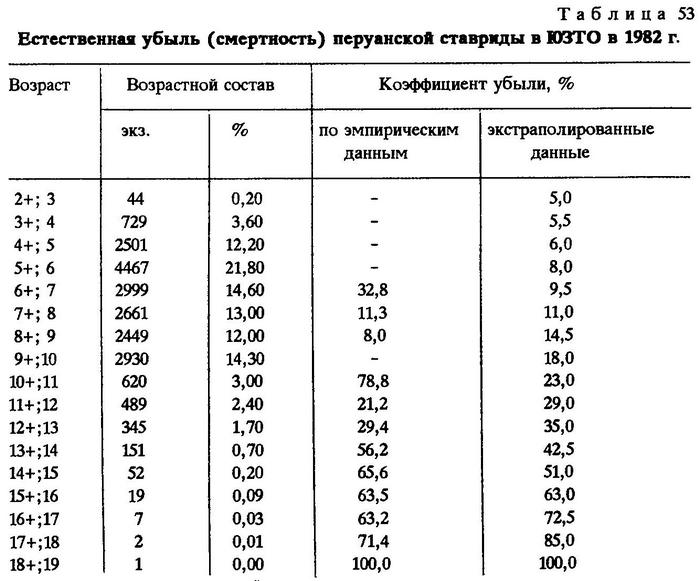
Так как материал был собран до начала промысла, то можно считать, что уменьшение численности перуанской ставриды старших возрастов в необлавливаемой популяции в основном обусловлено ее естественной смертностью.
Разницу между численностью данной и предшествующей возрастных групп начиная с трехлетнего возраста можно рассматривать как величину, характеризующую естественную убыль в этом возрасте. Например, шестилеток было 4467, а семилеток - 2999, следовательно, численность перуанской ставриды на седьмом году уменьшилась на 1468 экз., или, по отношению к численности шестилеток, на 32,8%.
Некоторый разброс показателей смертности смежных возрастных групп обусловлен, очевидно, недостаточностью материала. Однако, несмотря на небольшое отклонение, общий характер кривой смертности выявляется вполне отчетливо: с возрастом смертность повышается (рис.26).
Известно, что перуанскую ставриду вылавливают сейчас практически с годовалого и двухгодовалого возраста. Однако при определении оптимальной интенсивности вылова данные естественной смертности ее двухгодовиков и более молодых особей интереса не представляют, поскольку вопрос о недопустимости лова ставриды, не достигшей половой зрелости, ясен. Трехгодовалая ставрида уже считается промысловой рыбой. По крайней мере все особи в этом возрасте имели возможность отнереститься, что является, по П.В.Тюрину (1962), критерием возможного начала промысла, поэтому знать естественный отход на четвертом году ее жизни необходимо. Вот почему мы прибегли к методу экстраполяции кривой смертности, начиная с возраста трехлеток (2+). Такой метод в рыбохозяйственной науке вполне применим [Бойко, 1964]. К тому же известно, что с наступлением половой зрелости естественная смертность должна несколько повышаться. У трехлеток условно естественную смертность мы приняли за 5%.
При получении способом осреднения соседних точек кривой были расчислены путем экстраполяции недостающие показатели смертности перуанской ставриды в ранних и старших возрастных группах, а также откорректированы показатели смертности промежуточных возрастов. Эта исправленная кривая и принята нами для характеристики норм естественной смертности перуанской ставриды на 3-19-м годах жизни. Влияние поколений различной урожайности на соотношение возрастных групп при таком положении, естественно, сглаживается.
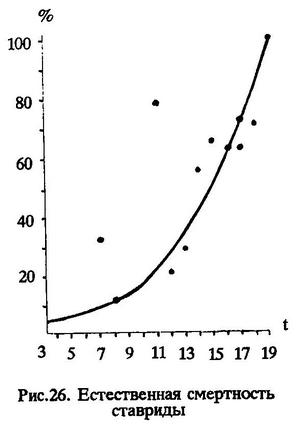
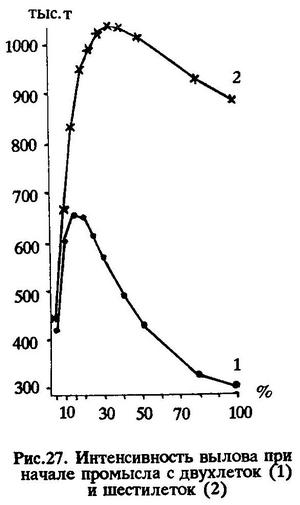
Итак, в необлавливаемой популяции перуанской ставриды естественная смертность от старости начинает проявляться после трех лет жизни и с возрастом увеличивается. Расчеты абсолютных величин этих потерь при разной интенсивности промысла (вылова), дают следующие результаты.
В ЮВТО и ЮЗТО ежегодно вылов составляет около 2 млрд. экз. Расчеты были произведены начиная с двухлеток (1+), так как с наступлением этого возраста ставрида вылавливается.
Разница между запасом и уловом есть остаток, часть которого погибает естественной смертностью (убыль) во все сезоны. Убыль от естественной смертности перуанской ставриды учитывалась с возраста трехлеток. Нормы естественной убыли в каждом возрасте сдвинуты на год вперед. Разница между остатком и убылью - есть запас к следующему году, когда перуанская ставрида станет на год старше. Интенсивность вылова принималась за 5, 10, 15, 20, 25, 30, 40, 50, 80 и 100%.
Оказалось, что при промысле начиная с двухлеток (а именно с этого возраста и ведется промысел в ЮВТО) оптимальный объем вылова обеспечивается при интенсивности промысла 15%. При этом уровне интенсивности получается наибольшая масса вылова - 658,9 тыс.т, а потери значительно ниже - 474,2 тыс.т, тогда как при интенсивности 10% уловы составляют 604,9% тыс.т, а потери - 845,6% тыс.т. При интенсивности вылова 20%, хотя потери и снижаются до 287,8 тыс.т, величина его уже ниже, чем при 15%, и составляет 652,6 тыс.т. При еще большей интенсивности эксплуатации масса вылова резко снижается.
Следовательно, расчеты показывают, что современная интенсивность вылова ставриды в ЮВТО является оптимальной (рис.27).
В связи с изложенным кажется, что рекомендовать что-либо иное с целью обеспечения оптимального промысла не представляется возможным.
И тем не менее, оказывается, можно значительно увеличить вылов перуанской ставриды в открытой части Тихого океана на основании рационального ведения промысла.
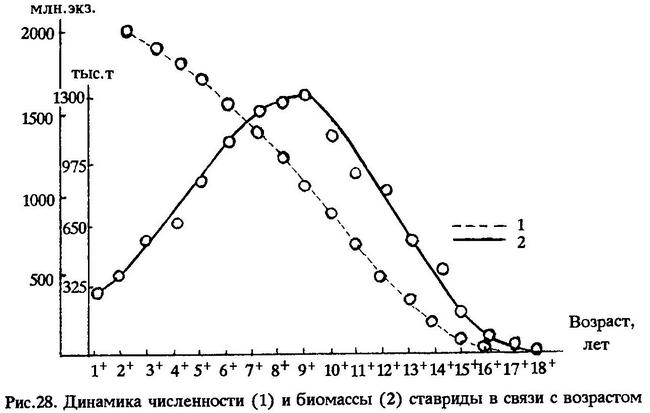
Для этого необходимо вести промысел ставриды начиная с возраста шестилеток, т.е. рыб длиной 36 см и более. Известно, что с увеличением возраста рыбы увеличивается масса каждой особи. Поэтому с возрастом, несмотря на уменьшение численности особей одного поколения, к середине жизненного цикла биомасса этого поколения значительно увеличивается (рис.28).
Соответствующие расчеты показывают, что лов шестилеток и рыб более старших возрастных групп даже при интенсивности вылова 10% более целесообразен, чем промысел двухлеток и старших особей при любой интенсивности вылова. Однако в этом случае слишком велики потери - 1220,7 тыс.т. Если вести промысел шестилеток и старше с интенсивностью вылова 15%, то последний может значительно превысить возможности, которые доступны при промысле начиная с двухлеток. Правда, и потери будут примерно на таком же уровне. Оптимальным промысел перуанской ставриды с шестилетнего возраста окажется при интенсивности вылова 35%, в этом случае можно выловить 1035,4 тыс.т при потерях 331,0 тыс.т (табл.54) (см.рис.27).
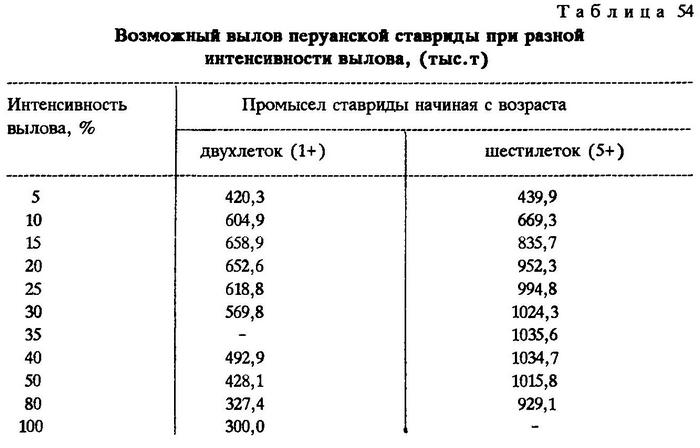
Таким образом, если ныне существующую интенсивность вылова, равную 15%, при начале промысла с двухлеток принять за 100%, то рекомендуемая нами степень эксплуатации перуанской ставриды начиная с шестилетнего возраста на уровне 35% может составить 157% вылова. Иными словами, при современном промысловом запасе перуанской ставриды в открытых водах океана и при рациональном ведении промысла вылов можно увеличить в 1,5 раза.
Промысел крупноразмерной перуанской ставриды в возрасте шестилеток и старше при разумной эксплуатации существующей сырьевой базы целесообразен не только с биологической точки зрения, но и из-за возможности направления ее на производство высокоделикатесной продукции благодаря повышенной жирности особей. Известно, что крупная ставрида в пищевом отношении является более ценным сырьем - она жирнее. Эмпирическим путем установлено, что ставрида с жирностью мяса более 4% имеет ожирение внутренностей, легко определяемое визуально и равное примерно 2 баллам. В апреле - августе в ЮЗТО у всей перуанской ставриды длиной более 37 см отмечается ожирение внутренностей в среднем 2 балла и более. Следовательно, в этот период всю рыбу крупнее 37 см можно направлять на изготовление деликатесной продукции.