Diplodus annularis (Linnaeus, 1758)
Ласкирь, морской карась (Rus),
Annular seabream
(Eng)
Синонимы:
Sargus annularis (Linnaeus, 1758)
Sparus annularis Linnaeus, 1758
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Карась морской (ласкирь) — Diplodus annularis (Linnaeus, 1758)
Тело довольно высокое, сильно сжатое с боков. Зубы крупные и широкие, передние имеют вид резцов. Чешуя крупная, плотно сидящая. D X—XI 11—12. А III 10—11. Окраска тела коричневато-оливковая с темным округлым пятном на хвостовом стебле. Длина тела обычно 15 см. Обычная прибрежная рыба Черного моря.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
МОРСКОЙ КАРАСЬ — DIPLODUS ANNULARIS (LINNE)
Условия размножения. Многими исследователями отмечено нахождение икры и личинок карася в планктоне в разные месяцы летнего сезона — с июля до середины сентября (Водяницкий, 1930а; Пчелина, 1936, 1940; Косякина, 1938; Виноградов, 1948; Дехник и Павловская, 1950; Павловская, 1950, Водяницкий и Казанова, 1954; Овен, 1959; Дука, 1959; Салехова, 1966а; Дехник и др., 1970).
Массовый нерест карася начинается с конца июня и продолжается до конца июля — начала августа. Нерест происходит повсеместно в незначительном удалении от берегов на небольших глубинах. В районе наших наблюдений в пределах 2,5—10-мильной прибрежной зоны отмечается заметное уменьшение количества икры и личинок в планктоне в направлении от берега к открытому морю.
Нерест карася в Черном море ограничен небольшими колебаниями температуры. В местах нахождения икринок температура воды колеблется от 18 до 25°. Личинки появляются обычно позже, при температуре воды не ниже 19°. Нерест карася в аквариумах Л. П. Салехова (1966а) наблюдала при температуре воды 17—17,6°.
Икра и личинки обычно встречаются в местах с соленостью не ниже 16—17‰. В отдельных случаях отмечено нахождение икринок при солености 12,8‰ (Дехник и Павловская, 1950). По наблюдениям Л. П. Салеховой (1966а), искусственно оплодотворенная икра морского карася в воде с соленостью ниже 16,2‰ погружается на дно аквариума и погибает. В опресненном Одесском заливе икра и личинки морского карася в планктоне не найдены (Зайцев, 1959а). У побережья Болгарии в водах с неустойчивым гидрологическим режимом не отмечено нахождение в планктоне ни взрослых особей со зрелыми половыми продуктами, ни икринок и личинок этого вида (Георгиев и др., 1960). Однако в Азовском море в районе Казантипского маяка при солености воды 11,55‰ и температуре 24,4° найдено большое количество нормально развивающейся икры и личинок этого вида.
В Средиземном море нерест морского карася происходит в близких к Черному морю температурных условиях и солености до 37—38‰. В Неаполитанском заливе особи со зрелыми половыми продуктами встречаются с апреля по июнь (Lo Bianco, 1908—1909; Ranzi, 1933). В северной часта Адриатического моря этот вид нерестится в апреле — мае (D'Ancona, 1949). Биологический анализ морских карасей, произведенный Л. П. Салеховой (19666) из уловов у берегов Албании, показал, что в мае гонады самок наряду с созревающими овоцитами содержат зрелые прозрачные икринки, готовые к вымету. Анализ карасей, пойманных в июле и сентябре, свидетельствовал о давно законченном ими нересте.
Температура поверхностного слоя воды с мая по июнь в прибрежных районах Адриатического моря повышается в среднем с 18 до 22°. Соленость на большей площади не снижается ниже 37—38‰, и только в северной части Адриатического моря она опускается до 34‰ (Богданова, неопубликованные данные).
Приведенные данные свидетельствуют о том, что нерест морского карася в морях Средиземноморского бассейна происходит в условиях сравнительно стабильного температурного режима, но при большом диапазоне солености воды. Следовательно, по отношению к солености этот вид, как и многие другие средиземноморские виды, натурализовавшиеся в Черном море, проявляет значительную лабильность, которая сохраняется во все периоды жизненного цикла. Икринки и личинки нормально развиваются во взвешенном состоянии при солености от 11,55 до 37—38‰.
Суточный ритм размножения. Нерест морского карася в природных условиях происходит в сумеречные часы. Икринки на этапе дробления встречаются в промежутке между 18 и 04 ч следующих суток (табл. 22). Встречаемость икринок на последующих стадиях развития соответственно сдвигается во времени. В. А. Водяницкий (1930а) наблюдал нерест морского карася в море в 18 ч. По данным Е. Г. Косякиной (1938), икрометание карася начинается около 19 ч.
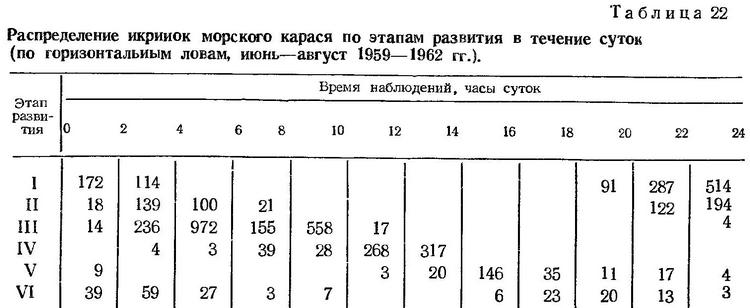
Четкий суточный ритм установлен Л. П. Салеховой (1966б) по наблюдениям за состоянием половых желез в течение суток. На большом материале показано, что самки с текучей икрой встречаются в уловах только в сумеречные часы. По наблюдениям этого же автора, в аквариуме морские караси нерестятся обычно с 20 ч до 22 ч 30 мин. Ни в какие другие часы суток вымет икры не наблюдался. Следовательно, и в неволе рыбы сохраняли свойственный им в природных условиях суточный ритм размножения.
Такая же стабильность суточного ритма размножения в природных и экспериментальных условиях была установлена для султанки (Овен, 1961а, 1962а).
Строение икринок. Икринки морского карася пелагические, сравнительно мелкие, с одной маленькой жировой каплей. Оболочка икринок совершенно прозрачная, желток гомогенный, перивителлиновое пространство очень узкое (рис. 26). По мере развития эмбриона перивителлиновое пространство несколько увеличивается. Диаметр икринок колеблется от 0,75 до 0,97 мм, жировой капли — от 0,15 до 0,20 мм (Водяницкий, 1930а; Косякина, 1938; Водяницкий и Казанова, 1954; Салехова, 1966а; наши данные). Средний диаметр икринок по нашим измерениям в разные сроки нерестового сезона составляет 0,89 мм, жировой капли — 0,18 мм. Икринки этого вида из Средиземного моря не описаны.
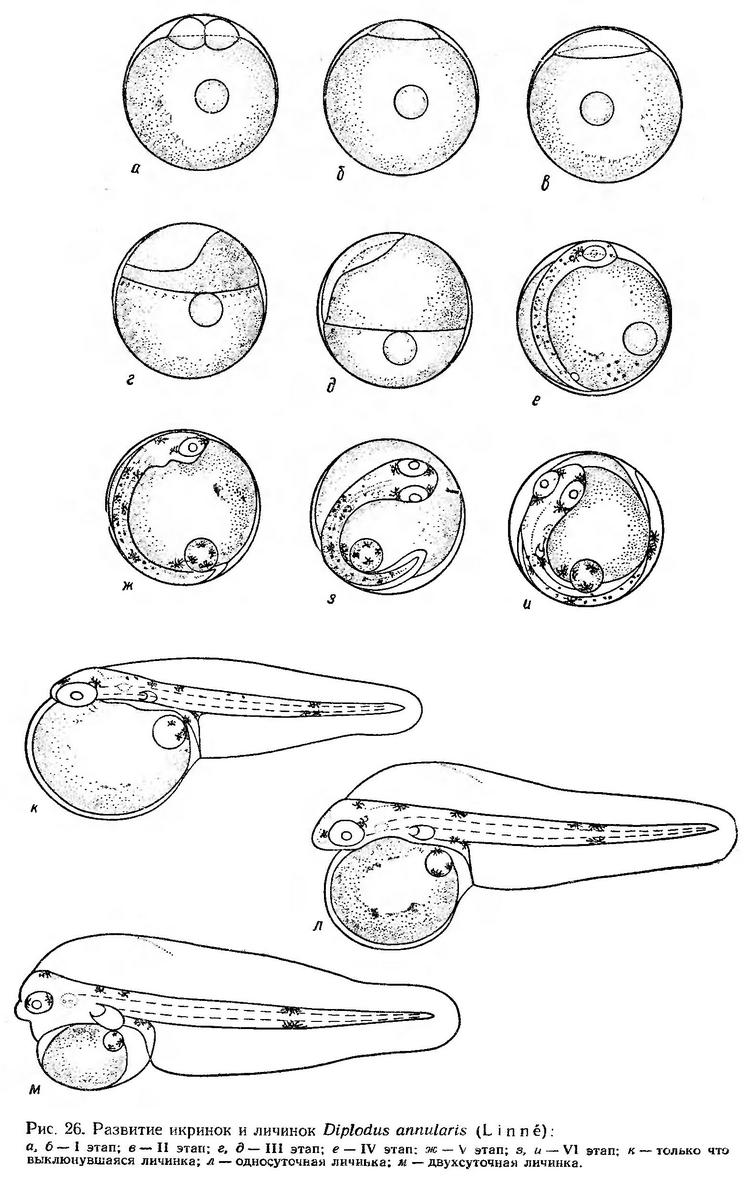
Эмбриональное развитие морского карася детально изучено Л. П. Салеховой (1966а). Наши опыты по эмбриональному и постэмбриональному развитию этого вида были поставлены на икринках, выловленных в море в промежуток времени от 20 до 21 ч, когда они находились на самых ранних стадиях развития.
Температура воды в опытах колебалась от 19,6 до 21,0°.
При температуре воды 19,6° первый этап — дробление — длился около 4 ч. Длина основания зародышевого диска составляет в среднем 0,43 мм, высота — 0,10 мм (рис. 26, а, б, табл. 23).

Эпителиальная бластула при температуре 19,6—21,0° формируется в течение 2,5—3 ч. Длина основания бластодиска заметно увеличивается; его края почти вплотную примыкают к оболочке (рис. 26, в). Одновременное обрастанием желтка бластодермой происходит гаструляшя, и вскоре образуется зародышевое утолщение (рис. 26, г).
Через 2,5—3 ч от начала обрастания края бластодиска достигают экватора; зародышевая полоска принимает четкие очертания (рис. 26, д). Перед закрытием бластопора в возрасте 14—15 ч намечаются первые туловищные миотомы. Головной конец зародыша расширен, обозначены глаза, нервный тяж, сформировался купферов пузырек. На спинной стороне зародыша и на желтке появляются мелкие меланофоры. При температуре 21,0—19,8° третий этап длился 7,5—8 ч.
После замыкания бластопора на IV этапе развития появляются слуховые капсулы, хрусталики, формируются мозговые доли. Намечаются характерные заглазничные скопления желтого пигмента; появляются мелкие клетки желтого пигмента на теле (рис. 26, е). В конце IV этапа в туловищном отделе насчитывается 16—17 сегментов. При температуре 19,8—20,6° IV этап длился 4,5—5 ч.
С момента отделения хвоста от желтка начинается быстрый рост хвостового отдела и одновременно дифференцировка хвостовых миотомов. Появляются зачатки сердца, печени, кишечника, намечается плавниковая складка (рис. 26, ж).
В возрасте 23—24 ч начинается слабая пульсация сердца. К этому времени хвост эмбриона незначительно заходит за середину желтка (рис. 26, з). Эмбрион изредка подергивается. Незадолго до выклева сердце пульсирует 67—70 раз в минуту. Намечаются зачатки грудных плавников. Зародыш принимает характерную пигментацию. На спине и боковых поверхностях разбросаны мелкие точечные меланофоры. Большие парные скопления желтого пигмента сосредоточены за глазами, менее крупные скопления находятся перед глазами и за слуховыми капсулами. Пятна желтого пигмента расположены также впереди ануса и в средней части хвостового отдела на вентральной стороне. Желтый пигмент появляется также на жировой капле и на желтке (рис. 26, и).
В возрасте 30—32 ч начинается выклев эмбрионов. К этому времени эмбрион охватывает 2/3 поверхности желтка. Процесс выклева при температуре 20,4° длится несколько минут. Выклев начинается с головного конца.
Постэмбриональное развитие. Личинки карася выклевываются слабо развитыми с большим, чуть выдающимся за голову, желточным мешком и плотно прижатой к нему головой. Жировая капля находится в задней верхней части желточного мешка. Грудные плавники зачаточные (рис. 26, к). Длина только что выклюнувшихся личинок колеблется от 2,00 до 2,20 мм. Анус открывается в край плавниковой складки сразу же за желточным мешком. Антеанальное расстояние составляет 44—45% длины тела, высота — около 7%. У выклюнувшихся личинок сохраняется эмбриональная пигментация.
Личинки плавают в вертикальном или несколько наклонном положении, головой вниз; в аквариумах объемом 15 л они размещаются во всей толще. Периоды покоя значительно продолжительнее моментов движения (до 1,5—2 мин). Большой, почти шаровидный, желточный мешок затрудняет движения. Личинки вращаются вокруг оси тела и изредка винтообразно перемещаются на небольшое расстояние. На прикосновение постороннего предмета почти не реагируют.
Длина односуточных личинок в среднем 2,53 мм. Желточный мешок у них сокращается. Зачатки грудных плавников увеличиваются. В передней части спинной плавниковой каймы образуется оводненная полость (рис. 26, л). Положение личинок в воде не изменяется. В покое они медленно погружаются головой вниз. Жировая капля, расположенная у заднего края желточного мешка, занимает верхнее положение. Личинки перемещаются на короткое расстояние винтообразно вверх или скачками в стороны, вверх и вниз.
К двухсуточному возрасту голова освобождается от желточного мешка, намечается ротовая ямка. Грудные плавники к этому времени довольно большие. Глаза приобретают сероватый пигмент. Тело личинок становится более прогонистым и стройным за счет удлинения хвостового отдела и соответственно относительного уменьшения туловища (рис. 26, м). В этом возрасте личинки уже способны передвигаться на сравнительно большие расстояния (разовое перемещение на расстояние до 2—3 см). Иногда они делают резкие скачки, быстро реагируя на приближение какого-либо предмета.
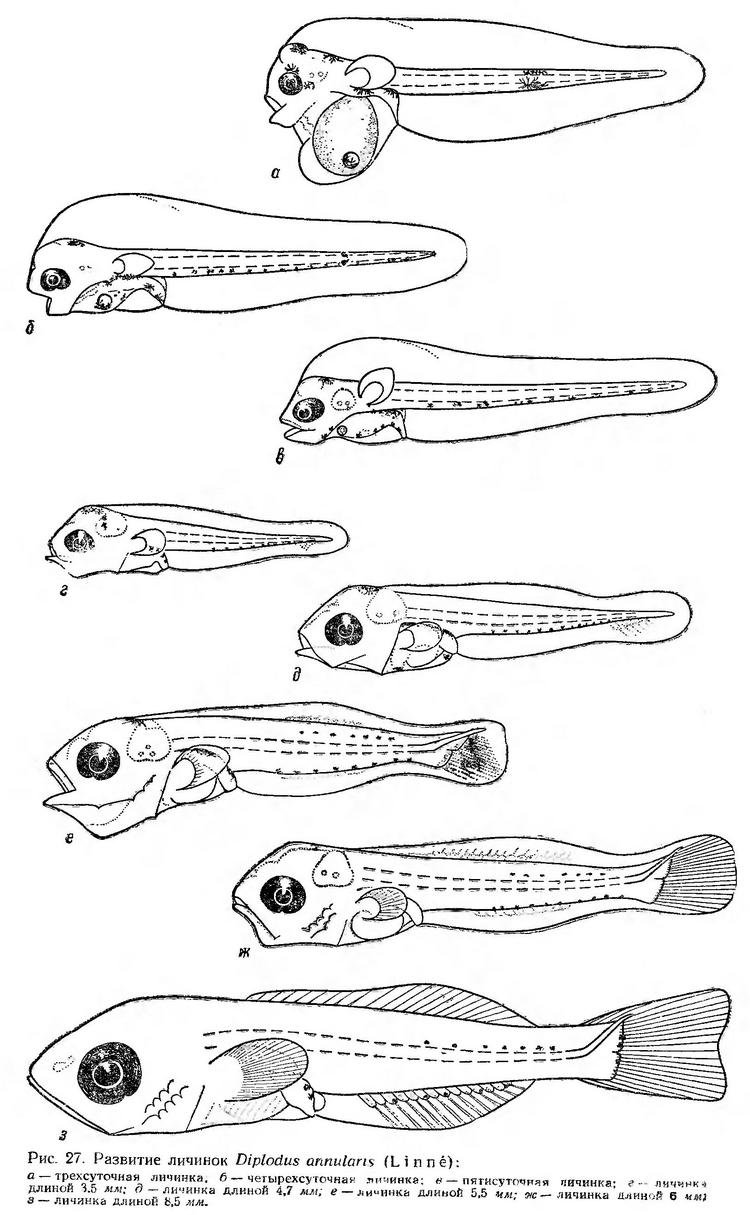
В возрасте трех суток прорезается рот и почти сразу же наблюдается его слабая аритмичная подвижность. Глаза интенсивно пигментируются (рис. 27, а). Большие грудные плавники приобретают подвижность. Это дает возможность личинкам менять направление движения, делать повороты. По мере рассасывания желточного мешка увеличивается полость в передней части спинной плавниковой каймы.
Четырехсуточные личинки достигают в среднем 2,75 мм длины. У них еще сохраняется небольшой остаток желтка и маленькая жировая капля. Оформляются челюсти, рот ритмично подвижен. Грудные плавники большие, широко расставленные, непрерывно вибрируют. Желтый пигмент исчезает. Появляется ряд мелких меланофоров вдоль вентральной стороны хвостового отдела и звездчатые меланофоры на кишечнике (рис. 27, б). В покое личинки находятся в перевернутом горизонтальном или вертикальном положении, но периоды покоя очень короткие. Движения личинок становятся быстрыми, порывистыми. Время от времени они стремительно поднимаются к поверхности вертикально головой вверх, затем поворачиваются головой вниз и опускаются ко дну аквариума. В этом возрасте личинки переходят на внешнее питание. По наблюдениям В. И. Синюковой (1964), в пищевом комке личинок на этапе смешанного питания насчитывается до 10 видов планктонных организмов. Основным объектом питания личинок морского карася размером 2,2—2,5 мм являются науплиусы Copepoda, которые составляют свыше 80% веса пищевого комка. Сравнительно большое значение в питании личинок играют инфузории, в частности Metacylis mediterranea (свыше 15% веса пищевого комка). Кишечники личинок, перешедших на внешнее питание, в светлое время суток всегда заполнены пищей, что свидетельствует об их способности отыскивать и захватывать необходимые пищевые организмы.
У личинок в возрасте пяти суток желток полностью рассасывается, сохраняется лишь маленькая жировая капелька. Тело у них в этот период прогонистое, стройное. Оводненная полость сокращается, и плавниковая кайма несколько уменьшается в высоте. Кишечник петлеобразно изгибается (рис. 27, в). Личинки находятся почти в непрерывном движении и принимают в воде нормальное положение.
Дальнейшее описание личинок морского карася дано по фиксированному материалу.
Личинки длиной 3,5 мм имеют большую голову, короткое широкое туловище и длинный хвостовой отдел. В хвостовом плавнике начинают закладываться лучи (рис. 27, г). Личинки питаются в основном науплиусами Copepoda, которые составляют в среднем 73% веса пищевого комка (Дука, Синюкова и др., 1970). По мере развития плавниковая кайма постепенно редуцируется и одновременно дифференцируются непарные плавники, в результате чего возрастает скорость движения личинок.
Личинки длиной 4,7 мм (рис. 27, д) продолжают питаться мелкими формами зоопланктона (науплиусами), но в их питании некоторое значение уже приобретают взрослые формы Oithona minuta, Podon polyphemoides, Paracalanus parvus и Penilia avirostris (Дука, Синюкова и др., 1970).
У личинок длиной 5,5 мм (рис. 27, е) хвостовой плавник заметно отчленяется от плавниковой каймы, уростиль загнут кверху. В спинном и анальном плавниках закладываются лучи.
У личинок длиной 6 мм непарные плавники довольно четко обособлены. Лучи в хвостовом плавнике сформированы (рис. 27, ж). Личинки переходят на питание более крупными формами зоопланктона. В пищевом комке преобладают взрослые формы Oithona minuta, Paracalanus parvus, Podon polyphemoides и Penilia avirostris, которые вместе составляют 70—100% веса пищевого комка. При длине личинок 8,5 мм (рис. 27, з) заканчивается формирование непарных плавников. Личинки питаются только крупными зоопланктерами. В светлое время суток кишечники у них всегда заполнены пищей (Дука, Синюкова и др., 1970).
Распределение и численность икринок и личинок. Летом икра и личинки морского карася встречаются вдоль всего побережья Крыма и Кавказа (Салехова, 1966а). Особенно большие концентрации их наблюдаются в Каркинитском заливе (Павловская, 1950; Смирнов, 1951). О нахождении икры и личинок карася у Анатолийского побережья сведения отсутствуют.
Распределение икринок карася, как и всех других тепловодных видов, ограничено слоем температурного скачка. Основные концентрации наблюдаются в приповерхностном слое моря. Однако и на горизонте 10 м могут быть обнаружены сравнительно большие концентрации икринок (табл. 24)
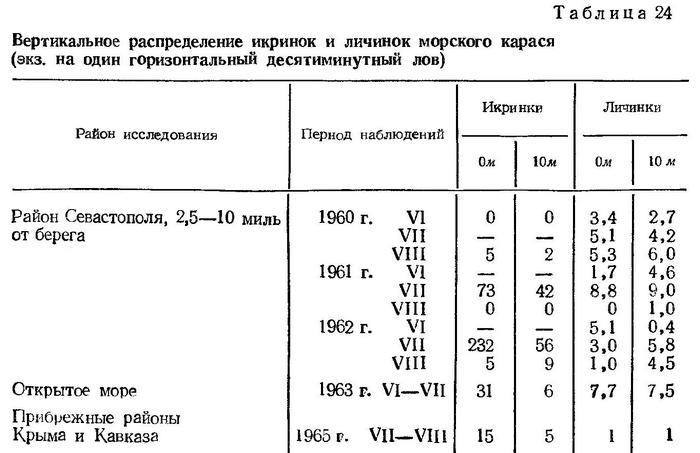
В вертикальном распределении личинок карася не выявлено четкой приуроченности к какому-либо определенному слою, как это показано для других видов. В одних случаях личинки карася концентрируются в поверхностном слое, в других — их численность выше на горизонте 10 м (табл. 24).
Поскольку приведенные в таблице данные (по району Севастополя) получены на основании стационарных многосуточных наблюдений, можно полагать, что такая разобщенность распределения личинок не носит случайного характера, а вполне закономерна для этого вида.
Численность икринок и личинок карася в планктоне подвержена значительным колебаниям (табл. 25). Икринки обычно встречаются в самых прибрежных районах, и уже на расстоянии 2,5 миль от берега количество их в планктоне резко уменьшается. Личинки распределяются на более широком пространстве, в основном до 10 миль от берега. Численность их в планктоне изменяется, по нашим данным, от 1 до 3 экз/м2 и в отдельных случаях достигает более высоких значений (до 10 экз/м2). Какой-либо зависимости изменения численности икринок и личинок карася от других массовых форм ихтиопланктона из приведенных наблюдений не установлено. В районе Севастополя в нерестовый сезон 1962 г., когда в планктоне были отмечены наиболее высокие концентрации икринок и личинок массовых видов (хамсы, ставриды и султанки), численность икринок и личинок морского карася была в среднем даже несколько ниже, чем в другие периоды наблюдений (табл. 25).

В течение всего нерестового сезона в планктоне значительно преобладают мелкие личинки карася — 2—4 мм длиной. Средний размер личинок в разные периоды нерестового сезона и в разные годы наблюдений почти постоянен (преобладают личинки размером 3—3,5 мм). Личинки карася длиной более 6 мм очень редко встречаются в планктоне; нахождение личинок размером более 8 мм в наших сборах не отмечено. Л. П. Салехова (1966а) считает, что личинки карася опускаются в придонные слои достигая длины 10—11 мм, в возрасте примерно двух недель. По данным Ранци (Ranzi, 1933), молодь карася переходит к придонному образу жизни достигая длины 11 — 13 мм. Наши наблюдения позволяют предположить, что придонный период жизни начинается раньше, при длине 8—9 мм.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
В Черном море
широко распространен ласкирь, или морской карась (Diplodus annularis).