Pegusa nasuta (Pallas, 1814)
Морской язык, косорот (Rus),
Blackhand sole
(Eng)
Синонимы:
Pleuronectes nasutus Pallas, 1814
Solea bleekeri Boulenger, 1898
Solea laskaris nasuta (Pallas, 1814) Морской язык, косорот (Rus),
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Морской язык — Solea lascaris nasuta (Pallas, 1811)
Спинной плавник берет начало впереди глаз. Передняя ноздря слепой стороны тела наибольшая и заметно расширенная. Боковая линия прямая. D 67—76. А 53—59. Р 7—10. Глазная сторона тела серо-буроватая с темными пятнами и более крупными отметинами неправильной формы. На грудном плавнике глазной стороны есть четкое черное пятно. Слепая сторона светлая. Длина тела не превышает 30 см. Обитает на дне Черного моря.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
МОРСКОЙ ЯЗЫК — SOLEA LASCARIS NASUTA (PALLAS)
Условия размножения. Нерест морского языка в Черном море происходит летом. В районе Севастополя нахождение зрелых рыб было отмечено в июне; личинки длиной 7 мм были пойманы в июле (Зернов, 1913). По наблюдениям Борча (1933), нерест морского языка у румынского побережья происходит с середины апреля до конца июля.
Икринки были впервые обнаружены в планктоне в районе Новороссийской бухты в конце июля и августе (Водяницкий, 1930а). Дальнейшие наблюдения показали, что икринки встречаются в этом районе с середины июня до конца сентября (Косякина, 1938). Личинки и мальки морского языка, по данным 3. М. Пчелиной (1940), обнаруживались в планктоне редко, единичными экземплярами, с конца июля до конца сентября. В районе Карадага нахождение икринок и личинок отмечено с июня — июля по август — сентябрь (Виноградов, 1948; Смирнов, 1959). В Одесском заливе икринки встречаются в планктоне с начала июня до середины сентября (Зайцев, 1959а). У Болгарского побережья нерест начинается в конце мая и заканчивается в конце сентября.
Размножение происходит в узкой прибрежной зоне при температуре воды 15—25° (Георгиев и др., I960). По наблюдениям Ю. П. Зайцева (1959а), икринки Solea lascaris nasuta встречаются при более значительных колебаниях температуры воды — от 10 до 24,5°. Колебания солености в местах нахождения икринок в Черном море также довольно велики — от 13,5 до 18,6‰ по нашим данным (Дехник и Павловская, 1950) и от 10,55 до 18,57‰ — по наблюдениям Ю. П. Зайцева (1959а).
Нерест Solea lascaris (Rissо) в Средиземном море происходит, по-видимому, весной (Padoa, 1956з). Икринки и ранние личинки встречаются здесь очень редко. Икринки и вылупившиеся личинки описаны Холтом (1891, 1899) от берегов Ирландии и Марселя.
В Северном море этот вид встречается очень редко. Размножается предположительно с мая — июня и по июль — август (Ehrenbaum, 1905—1909; Padoa, 1956з).
Таким образом, в разных районах ареала Solea lascaris нерестится в весенне-летний период года. Нахождение нормально развивающихся икринок при значительных колебаниях температуры (10—25°) и очень большом диапазоне солености (от 10,5—13,5‰ в Черном море до 38—39‰ в Средиземном) свидетельствует об эвригалинности и эвритермности как взрослых форм, так и икринок и личинок этого вида.
Строение икринок. Икринки морского языка пелагические, сферической формы, прозрачные, с мелко сегментированным желтком. Перивителлиновое пространство очень маленькое. Они легко отличаются от других икринок наличием многочисленных мелких жировых капель в желтке, расположенных группами (рис. 63). Величина икринок Solea lascaris nasuta, по данным различных авторов, колеблется от 1,09 до 1,35 мм (Водяницкий, 1930а; Водяницкий и Казанова, 1954; Зайцев, 1959а; Георгиев и др., 1960; наши измерения).
Икринки этого вида из Средиземного и Северного морей, по данным Холта (1899), имеют диаметр 1,36—1,38 мм. Эмбрион очень рано окрашивается многочисленными разветвленными желтыми хроматофорами. Меланофоры очень редки или отсутствуют совсем (Радоа, 1956з). В Виллафранкском заливе Средиземного моря диаметр икринок Solea lascaris в среднем равен 1,34 мм (Thomopoulos, 1956).
Эмбриональное развитие морского языка прослежено от начала обрастания желтка бластодермой. Икринки в начале III этапа развития (рис. 63, а) встречаются в планктоне в ранние утренние часы. Следовательно, вымет икры, как и у подавляющего большинства других летнеразмножающихся видов, происходит в ночное время. Формирование зародыша вначале идет довольно медленно. Края бластодиска уже заходят за среднюю линию желтка, а зародышевая полоска не имеет четких очертаний. С одного края бластодермы видно лишь широкое расплывчатое зародышевое утолщение (рис. 63, б). Головной конец зародыша слабо намечается, когда на вегетативном полюсе остается небольшая желточная пробка (рис. 63, в).
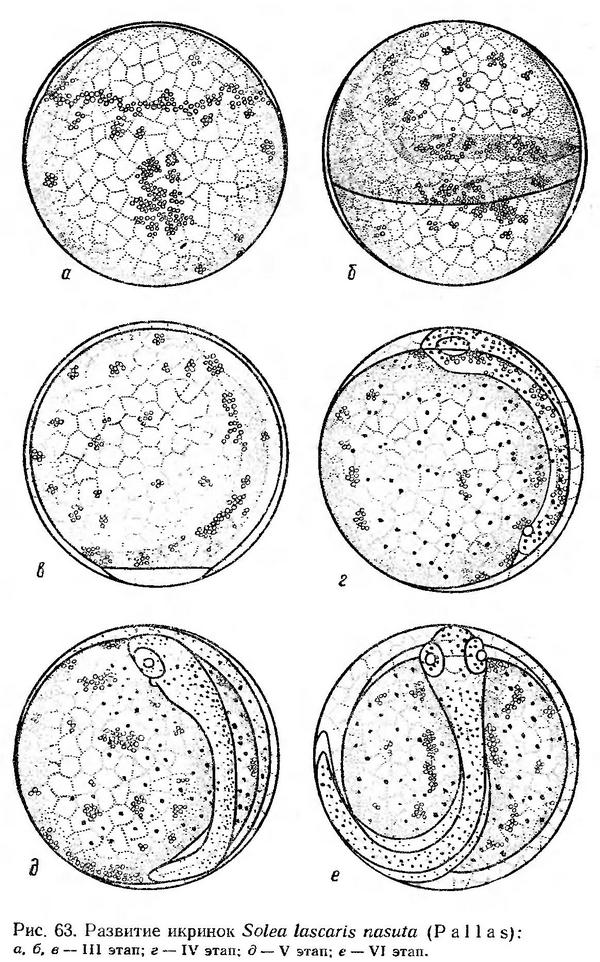
Процесс обрастания желтка и гаструляция при температуре воды 24,4—25,0° продолжается около 7 ч. После замыкания бластопора начинается быстрый органогенез. Образуются глаза, формируются мозговые пузыри, закладывается хорда, дифференцируются туловищные миотомы. По всему телу эмбриона и на желтке появляются довольно крупные точечные меланофоры (рис. 63, г). К началу отделения хвоста от желтка в глазах намечаются хрусталики, образуются слуховые капсулы, зачатки сердца и других внутренних органов. Намечается плавниковая кайма (рис. 63, д).
Когда хвост эмбриона заходит за середину желтка, хвостовой отдел начинает слабо, но довольно часто подергиваться (через 7—9 сек) и одновременно начинается пульсация сердца. По мере развития движения эмбриона становятся более энергичными. Он начинает изгибаться и переворачиваться в оболочке. Ко времени выклева зародыш охватывает около 3/4 поверхности желтка (рис. 63, е). Выклев происходит с головного конца.
Постэмбриональное развитие. Только что выклюнувшиеся личинки имеют длину 2,1—2,5 мм. Желточный мешок большой, передний край его немного выступает за линию головы. Жировые капли расположены группами по всей поверхности желточного мешка. Туловищный отдел несколько длиннее хвостового (54—55% длины тела). Голова плотно прижата к желточному мешку. Плавниковая кайма над головой образует выступ, который, по-видимому, как и у Arnoglossus, выполняет защитную функцию. Грудные плавники зачаточные. На спинной стороне тела густо расположены мелкие точечные меланофоры. По всему телу, на плавниковой кайме и желточном мешке разбросаны также ветвистые желтые пигментные клетки (рис. 64, а). Сердце пульсирует очень быстро — 100—120 раз в минуту. Несмотря на большой желточный мешок, личинки очень подвижны. Они плавают в горизонтальном положении брюшной стороной вверх. Через короткие промежутки времени (7—15 сек) личинки винтообразно поднимаются к поверхности или двигаются в горизонтальной плоскости, не меняя положения в воде.
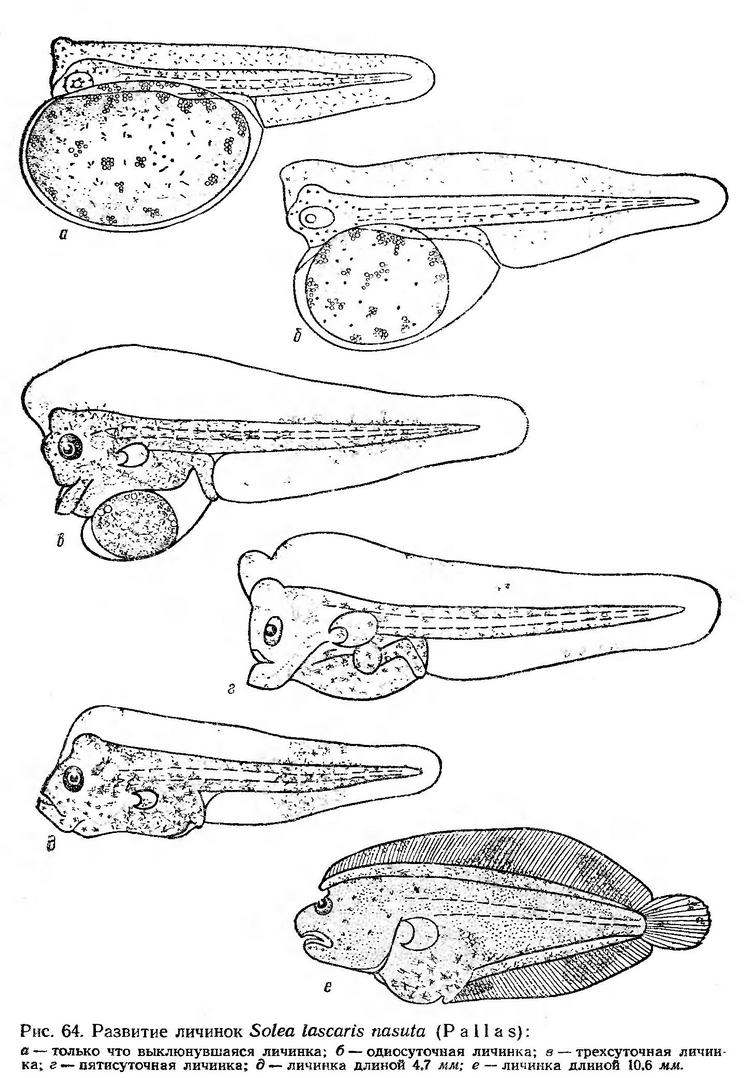
К суточному возрасту длина личинок увеличивается до 2,8—3,1 мм. В области среднего мозга образуется выпуклость и в этой же части имеется выпячивание плавниковой каймы. Образуется ротовая ямка. В глазах появляется черный пигмент. Желточный мешок уменьшается и принимает сферическую форму. Зачатки грудных плавников несколько увеличиваются. Желтый пигмент принимает диффузный характер. Меланофоры начинают смещаться к брюшной стороне, (рис. 64, б). Личинки очень активны. Они двигаются при помощи быстрых вибрирующих изгибов хвоста в горизонтальной плоскости или поднимаются вверх, преодолевая расстояние 5—10 см, затем пассивно падают вниз на дно аквариума. В покое личинки могут занимать любое положение: лежат на боку, подвешиваются вертикально или косо головой вниз или вверх, иногда принимают нормальное горизонтальное положение.
К трехсуточному возрасту длина личинок увеличивается незначительно (3,1—3,3 мм). Туловищный отдел заметно укорачивается. Прорезывается рот, но остается пока неподвижным. Грудные плавники большие, веслообразные, широко расставлены в стороны, быстро вибрируют. Глаза окрашиваются в интенсивно черный цвет. В спинной плавниковой кайме образуется оводненная полость. Меланофоры принимают паутинообразную форму, покрывают все тело личинки и желточный мешок. В средней части хвостового отдела они переходят на плавниковую кайму. Желтый пигмент расположен по всему телу личинки и желточному мешку (рис. 64, в). Личинки очень подвижны. Они плавают в нормальном положении — спиной кверху. Движения в основном круговые. Массивный головной конец, широкий туловищный отдел, сравнительно большой еще желточный мешок не позволяют личинкам производить быстрых стремительных движений. Периоды покоя очень короткие (2—3 сек). В покое личинки лежат на боку.
В возрасте пяти суток у личинок сохраняется еще небольшой желточный мешок. Длина их в условиях эксперимента не изменяется. Нижняя челюсть выдается вперед за верхнюю. Рот приобретает подвижность (рис. 64, г).
Личинка Solea lascaris nasuta длиной 4,7 мм имеет совершенно симметричное тело. Туловищный отдел сравнительно короткий (40—42% длины тела), кишечник массивный. По фоновой желтоватой окраске густо расположены крупные ветвистые меланофоры. В средней части хвостового отдела желтый пигмент и меланофоры переходят на спинную и брюшную части плавниковой каймы (рис. 64, д). В аквариуме объемом воды 10 л личинки держатся преимущественно у дна, плавают в разных направлениях, быстро делают повороты, часто производят скачки. Очень четко выражен поисковый инстинкт: плавая у дна, личинки захватывают различные осевшие там частицы. У личинок длиной 10,6 мм хорошо выражен метаморфоз. Левый глаз смещается на правую сторону. Нижняя челюсть укорачивается. Лучи в непарных плавниках сформированы. По пигментации обе стороны тела пока не отличаются (рис. 64, е). По данным З. М. Пчелиной (1940), личинки длиной 9,9—11 мм имеют уже ассиметрично расположенные глаза, но пигментированы еще с обеих сторон.
Распределение и численность икринок и личинок. Икринки морского языка летом встречаются в различных районах Черного моря: у берегов Крыма, Кавказа, в северо-западной части в Одесском заливе, у побережья Болгарии (Водяницкий, 1930а; Косякина, 1938; Виноградов, 1948; Дехник и Павловская, 1950; Дука, 1959; Зайцев, 1959а; Овен, 1959; Георгиев и др., 1960).
Во всех районах Черного моря икринки этого вида встречаются обычно в небольшом количестве, чаще единичными экземплярами. По наблюдениям Ю. П. Зайцева (1959а), в северо-западной части моря вертикальное распределение икринок морского языка определяется плотностью воды. В годы с высокой плотностью икра плавает у самой поверхности. При средней плотности воды она распространяется равномерно во всей толще до слоя температурного скачка; во время опреснения икра находится в основном непосредственно над слоем скачка.
Наши наблюдения в районе Камышовой бухты показали, что икринки морского языка наиболее часто встречаются в планктоне в июле. Их численность в это время достигает 2—3 экз. под 1 м2 поверхности моря. В июне и августе икринки Solea lascaris nasuta вылавливаются обычно единичными экземплярами. В открытом море икринки этого вида в планктоне не найдены.
Личинки морского языка длиной 4,9—11 мм встречались в планктоне Новороссийской бухты и перед входом в нее с конца июля до конца сентября (Пчелина, 1940). В наших сборах было обнаружено 11 личинок (в июне, июле и августе) длиной 2,7—10,6 мм. По горизонтам лова личинки распределялись следующим образом: на 0 м выловлено 5 личинок (2 лова), на 10 м — 3 (3 лова), на 20 м — 1 (1 лов), в слое 25—0 м — 2 личинки (2 лова).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
У нас в Черном и Азовском морях живет
солея морской язык, или косорот (Solea laskaris nasuta). Окрашен косорот сверху
в серовато-бурый цвет с черными
пятнами или с мраморным рисунком. На
грудных плавниках имеется по черному глазку.
Он достигает длины 30 см.
Косоротов промышляют повсюду, где
они водятся. Мясо многих видов вкусное,
и они ценятся, несмотря на малую
величину рыб.