(Dinophyta Dillon, 1963) Тип Динофиты, Динофлагелляты, Динофитовые водоросли, Перидинеи, Панцирные жгутиконосцы, Phylum Dinophyta Dillon, 1963 = Dinoflagellata Bütschli, 1885 = Peridinea Ehrenberg, 1830 (Dinoflagellates) 3 класса
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Динофиты, Динофлагелляты, Динофитовые водоросли, Перидинеи, Панцирные жгутиконосцы (Dinophyta Dillon, 1963, синонимы Dinoflagellata Bütschli, 1885, Peridinea Ehrenberg, 1830) — крупная группа протистов из надтипа альвеолят (Alveolata), которой традиционно присваивают ранг типа. Известно около 4000 ископаемых и более 2500 современных видов, из которых 90 % обитает в морях, остальные — в пресных водах. Около половины представителей — свободноживущие фотосинтезирующие организмы, однако известны и бесцветные гетеротрофные формы, и паразитические динофлагелляты. Некоторые виды являются симбионтами коралловых полипов и двустворчатых моллюсков. Наличие пигмента пирофилла объясняет наличие другого названия — «пирофитовые водоросли».
Первые организмы, в настоящее время относимые к динофлагеллятам, были описаны в 1753 году английским натуралистом Генри Бейкером в трактате «Employment for the microscope» как «крошечные животные, которые вызывают свечение морской воды»; речь шла о ночесветках. В 1773 году датский натуралист Отто Фредерик Мюллер описал в своей книге два вида пресноводных динофлагеллят, дав им имена Bursaria hirundinella и Vorticella cincta (ныне известны соответственно как Ceratium hirundinella и Peridinium cinctum). В 1830-х годах немецкий естествоиспытатель Христиан Готфрид Эренберг, изучив множество проб морской и пресной воды, описал в статье «Beitrage zur Kenntnis der Organisation der Infusorien und ihrer geographischer Verbreitung, besonders in Sibirien» и ряде последующих работ несколько выделяемых и ныне родов динофлагеллят, среди которых Peridinium, Prorocentrum и Dinophysis. По имени первого из них представителей рассматриваемой группы стали именовать перидинеями.
В 1885 году немецкий учёный Отто Бючли выделил данную группу организмов в отряд простейших Dinoflagellata (в типифицированном варианте — Dinoflagellida; данное название таксона длительное время использовалось протозоологами). Название образовано от древнегреческого слова δινος (dinos) — «вращение» и латинского слова flagellum — «жгутик». В то же время альгологи Эугениус Варминг (1890) и Адольф Энглер (1892) поместили динофлагеллят в царство растений. Немецкий ботаник Адольф Пашер в своей системе водорослей (1914) выделил динофлагеллят в класс Dinophyceae в составе отдела Pyrrophyta (буквально «огненные растения» — от древнегреческих слов πυρ (pyr) — «огонь» и φυτον (phyton) — «растение»; название дано из-за способности некоторых видов к биолюминесценции). В позднейших ботанических классификациях динофлагеллят обычно либо трактовали как класс в составе Pyrrophyta, либо выделяли в самостоятельный отдел Dinophyta.
В 1950—1960-е годы появились первые подробные системы ископаемых динофлагеллят; наметившийся разрыв между классификациями современных и ископаемых динофлагеллят был в основном преодолён в 1990-е годы, чему способствовали более тщательное изучение морфологии ископаемых представителей (ставшее возможным с развитием электронной микроскопии) и жизненных циклов современных видов.
Динофлагелляты представлены преимущественно одноклеточными монадными формами, подвижные клетки снабжены двумя неравными жгутиками. Клеточные покровы представлены амфиесмой (текой), состоящей из везикул, которые залегают под плазмалеммой. Митохондрии с трубчатыми кристами. Хлоропласты у разных представителей имеют различное происхождение и характеризуются различными фотосинтетическими пигментами. Обычно они окрашены в жёлто-бурый цвет, поскольку, кроме зелёных хлорофиллов, имеют дополнительные пигменты — например, перидинин. Геном хлоропластов часто фрагментирован на короткие кольцевые хромосомы. У многих видов имеется стигма (глазок) различного строения. Как правило, осморегуляцию осуществляют уникальные органеллы — пузулы. Нередко имеются стрекательные структуры (трихоцисты и другие). Пресноводные формы запасают крахмал, морские — липиды и стеролы.
Хромосомы динофлагеллят обычно конденсированы в течение всего клеточного цикла и лишены типичных эукариотных гистонов. Митоз закрытый, метафазная пластинка не образуется. Отличительной особенностью динофлагеллят, не встречающейся среди других эукариот, является частичное замещение тимина в их ядерной ДНК другим азотистым основанием — 5-гидроксиметилурацилом.
Размножение чаще всего происходит продольным делением клетки надвое. Бесполое размножение — зооспорами и апланоспорами. У некоторых видов описан половой процесс. У большинства представителей жизненный цикл гаплобионтный с зиготической редукцией, но есть и исключения. Многие виды способны образовывать покоящиеся формы (цисты).
Некоторые виды способны к биолюминесценции. Кроме того, часто в клетках динофлагеллят образуются токсины — например, гониатоксин. Данный токсин имеет свойство накапливаться в тканях моллюсков, ракообразных, рыб, что приводит к отравлению животных, питающихся ими, а также человека, если он употребит их в пищу.
Динофлагелляты играют важную роль в биоценозах морей и океанов, выступая, наряду с диатомеями, в качестве основных первичных продуцентов. Фототрофные динофлагелляты способны вызывать цветение воды, а массовые вспышки их численности в прибрежных водах ответственны за возникновение «красных приливов». Паразитические динофлагелляты иногда служат причиной эпизоотий в популяциях рыб и ракообразных.
Вегетативная стадия жизненного цикла динофлагеллят преимущественно представлена монадными формами с дорсовентральным строением, гораздо реже встречаются ризоподиальные и коккоидные клетки. Однако другие стадии жизненного цикла могут быть представлены пальмеллоидными, амебоидными, нитчатыми и даже многоядерными формами. Так, паразитическая Pfiesteria piscicida на некоторых этапах своего жизненного цикла трансформируется в амебоидную форму. Другие динофлагелляты тоже могут менять свой облик. В частности, некоторые представители способны принимать форму «голого», лишённого жгутиков шара (пальмеллы). Делясь, одноклеточная пальмелла даёт начало плотному скоплению клеток. Динофлагелляты, обитающие в кораллах как зооксантеллы, всегда имеют форму пальмелл.
Форма динофлагеллят разнообразна: шаровидная, эллипсовидная, яйцевидная, обратнояйцевидная, грушевидная, звёздчатая, палочковидная, в виде полумесяца или тетраэдра. Клетки, как правило, уплощённые, в поперечном сечении имеют вид эллипса или пластинки, реже тело округлое и неуплощённое. Размеры клеток варьируют от 6 до 2000 мкм. Наиболее крупные динофлагелляты обитают в морях, а среди пресноводных представителей самые крупные клетки характерны для видов рода Ceratium, их размер составляет до 450 мкм.
Выросты на панцирях динофлагеллят могут иметь форму рогов, крыльев, чешуй, они значительно увеличивают поверхность клетки. Показано, что некоторые динофлагелляты способны сбрасывать панцирь при некоторых условиях, например, при охлаждении до 4 °С, однако при переносе сбросивших панцирь клеток в обычные условия панцирь восстанавливается. Пластинки, образующие панцирь, играют ключевую роль в определении панцирных динофлагеллят. Они составляют так называемую формулу теки, где определённые пластинки обозначаются цифрами и буквами. В качестве систематических признаков используют также форму, количество, размещение пластинок и наличие выростов.
У различных динофлагеллят жгутики могут быть прикреплены по-разному. Если два неравных жгутика отходят от апикального конца клетки, то такой тип называется десмоконтным (например, у Prorocentrum); такой вариант отхождения жгутиков считается примитивным. В этом случае при плавании один жгутик направлен вперёд, а другой — перпендикулярно первому. Однако у большинства динофлагеллят имеет место более эволюционно продвинутый вариант, при котором оба неравных жгутика отходят от вентральной стороны клетки (диноконтный тип отхождения жгутиков). У этих динофлагеллят клетки разделены на эпивальву и гиповальву, причём поперечный жгутик лежит в поперечной бороздке, а продольный — в продольной.
Среди фотосинтезирующих одноклеточных организмов динофлагелляты самые быстрые пловцы, их скорость передвижения составляет 200—500 мкм/с. Впрочем, они всё-таки уступают Mesodinium — организму, представляющему собой эндосимбиоз инфузории и криптофитовой водоросли. Клетки Lingulodinium polyedrum плавают с линейной скоростью 250 мкм/с при температуре 20 °С, а клетки Gyrodinium при той же температуре развивают линейную скорость 319 мкм/с.
Для митоза динофлагеллят характерен ряд особенностей: ядерная мембрана сохраняется (закрытый митоз), не формируется метафазная пластинка. Такой тип закрытого митоза получил название диномитоз. В качестве примера наиболее продвинутого типа митоза у динофлагеллят можно рассмотреть митоз Syndinium. Начало деления знаменуется удвоением оснований жгутиков с двух до четырёх. В течение этой стадии ядро увеличивается в размерах и можно разглядеть много Y- и V-образных хромосом. В профазе митоза происходит продольное расщепление хромосом и образуются хроматиды, спирально закрученные вокруг друг друга. Продолжительность профазы у динофлагеллят в значительной мере определяется длиной хромосом: чем хромосомы длиннее, тем больше времени занимает их раскручивание.
Оболочка ядра остаётся интактной во время митоза, и во время митоза возникают её глубокие впячивания (у некоторых динофлагеллят ядерная оболочка растворяется). Некоторые из них образуют сквозные туннели, проходящие через всё ядро насквозь (образуется от 1 до 15 туннелей). Эти туннели заполнены цитоплазмой, и в них начинают собираться микротрубочки, которые, однако, не соединены с нетронутой ядерной оболочкой. Микротрубочки собираются в пучки по 25—40 сразу после разделения хромосом на хроматиды. У Amphidinium carterae в ядре образуется 2—4 туннеля, у других видов их больше.
Хлоропласты динофлагеллят дисковидные, однако они могут иметь пластинчатую, лентовидную или другую форму. В клетке они могут располагаться около границы клетки, а могут радиально расходиться от центра клетки. Количество хлоропластов в клетке варьирует от одного до множества.
В отличие от других эукариот, у динофлагеллят процессы приобретения хлоропластов в ходе эндосимбиоза и их утери протекают сравнительно легко и являются обычным явлением. Этим, вероятно, и объясняется огромное разнообразие хлоропластов у динофлагеллят. Для большинства их разновидностей характерно наличие оболочки из трёх мембран, в состав которой не входит эндоплазматический ретикулум, тилакоиды собраны в стопки по три. Среди предполагаемых древних и недавно приобретённых эндосимбионтов динофлагеллят обнаружены:
цианобактерии: например, у Ornithocercus magnificus, Citharistes regius;
красные водоросли: у всех динофлагеллят, содержащих хлоропласты с тремя мембранами и перидинином, например, у видов рода Ceratium;
гаптофитовые водоросли: например, у Gymnodinium mikimotoi, Gymnodinium breve и Gymnodinium galatheanum;
криптофитовые водоросли: например, у Gymnodinium eucyaneum, Gymnodinium acidotum;
празинофициевые водоросли: например, у Lepidodinium viride, Gymnodinium chlorophorum;
диатомовые водоросли: например, у Peridimium balticum, Peridimium foliaceum;
предполагаемые силикофлагелляты: например, у Podolampas bipes.
У некоторых динофлагеллят, например, у Dinophysis spp. и Amphidinium poecilchroum, имеет место клептопластия: в их клетках временно содержатся хлоропласты, заимствованные у фотосинтезирующей добычи. Для вида Dinophysis acuminata характерна двойная клептопластия: представители данного вида заимствуют пластиды у поедаемых ими инфузорий Myrionecta rubra, а те, в свою очередь, накапливают хлоропласты поглощаемых ими криптофитов вида Teleaulax amphioxeia.
Все изученные динофлагелляты демонстрируют фототаксис в ответ на одну и ту же часть спектра, причём для волны длиной 450 нм степень проявления фототаксиса максимальна. Стигма (глазок) для фототаксиса необязательна, и лишённые стигмы клетки тоже могут реагировать на свет. Рецепторы, реагирующие на свет, определённо находятся не в стигме и, вероятно, связаны с плазматической мембраной.
По типу питания динофлагелляты могут быть автотрофами, миксотрофами и гетеротрофами. По-видимому, автотрофии в строгом смысле среди динофлагеллят практически не бывает, зато чрезвычайно широко распространена миксотрофия. Известно только менее 10 чисто автотрофных видов, не нуждающихся в готовых органических соединениях. Большая часть динофлагеллят, способных к фотосинтезу, ауксотрофна по витаминам группы В (B1 (тиамин), В12 (цианокобаламин) и Н (биотин)) и вынуждена получать их извне в готовом виде. Кроме того, известны фотосинтезирующие динофлагелляты, способные к фаготрофии.
Около половины известных видов динофлагеллят — облигатные гетеротрофы, лишённые пластид и поглощающие пищу осмотрофным или фаготрофным путём. Фаготрофия имеет более широкое распространение у динофлагеллят. Фаготрофные организмы поглощают твёрдые пищевые частицы и переваривают их в пищеварительных вакуолях. Они питаются другими водорослями, в том числе и другими динофлагеллятами, нематодами, личинками полихет, икрой рыб. Для захвата пищевых частиц они используют разнообразные специальные органеллы, среди которых стебельки (педункли), щупальца (тентакли), поршни (пистоны) и вуали (паллиум).
Стебельки — небольшие подвижные пальцевидные выросты, которые располагаются в борозде у основания жгутиков. Вероятно, они выполняют сенсорные функции и обеспечивают прикрепление к субстрату.
Щупальца — более удлинённые образования, служащие для захвата пищи и направления её к клеточному рту (цитостому). Щупальца описаны у Nocticula, Pronocticula, Pavillardia и др.
Поршень представляет собой полый цилиндрический вырост, подтягивающий пищу к цитостому. Он может быстро удлиняться, причём его длина может в 15 раз превышать длину тела, расширяться, а потом быстро сужаться и сокращаться. Поршень проделывает отверстие в клетке-жертвы и высасывает из неё цитоплазму. Такой поршень характерен для Gymnodinium fungiforme, зооспор Pfiesteria piscicida, а также фотосинтезирующей динофлагелляты Amphidinium cryophilum.
Вуаль — псевдоподиальная мембрана, выходящая из борозды вблизи основания жгутика и имеет вид ловчей сети, по размеру превышающей клетку. К этой сети прилипают отдельные крупные клетки и колониальные и диатомеи, далее они обволакиваются и их содержимое всасывается. Такой акт внеклеточного пищеварения длится 20—30 минут. Вуаль описана у некоторых видов родов Protoperidinium, Oblea и Diplopsalis. Первые питаются в основном диатомеями и динофлагеллятами, а все остальные могут ловить не только диатомей, но и гаптофитовых, празинофициевых и криптофитовых водорослей.
В клетках динофлагеллят может присутствовать большая везикула, содержащая останки переваренных органелл (тельце накопления). Вероятно, она соответствует везикулам с теми же функциями, имеющимся у криптофитовых водорослей и других жгутиконосцев. Тельце накопления особенно характерно для симбиотических динофлагеллят.
Динофлагелляты имеют стрекательные структуры (экструсомы) трёх типов: трихоцисты, нематоцисты и мукоцисты. Трихоцисты располагаются перпендикулярно поверхности клетки по её периферии. Трихоциста состоит из тела и шейки и окружена одной мембраной. Внутри трихоцист находятся кристаллические белковые фибриллы длиной несколько микрометров (кор), которые закручены по спирали. Во внутренней части трихоцисты треть кора представлена короткими трубчатыми элементами, чуть выдающимися вперёд. На самом верху трихоцисты группа из 20—22 фибрилл выдаётся из кора в сторону закрывающей мембраны, и более тонкие фибриллы далее соединяют их с апикальной частью мембраны трихоцисты.
В пределах закрывающей мембраны трихоцисты находится тонкое непрозрачное кольцо. Внешняя часть мембраны трихоцисты прикрепляется к плазматической мембране между текальными везикулами или к текальным везикулам, расположенным под округлыми, тонкими областями текальных пластинок, которые формируют специальные поры для трихоцист.
Трихоцисты формируются в аппарате Гольджи и, вероятно, покидают его в виде сферических везикул, которые потом приобретают веретеновидную форму и развиваются в трихоцисты. При возбуждении «заряженная» трихоциста превращается в «незаряженную» в течение нескольких миллисекунд — возможно, из-за быстрого притока воды. «Незаряженные» трихоцисты представляют собой прямые конусовидные палочки, во много раз длиннее «заряженных» трихоцист (до 200 мкм у Prorocentrum). Трихоцисты имеются у большинства динофлагеллят, но есть и исключения (Gymnodinium neglectum, Aureodinium pigmentosum, Woloszynskia tylota, Symbiodinium microadriaticum). Конкретная польза (если она есть) от трихоцист неясна. Они могут служить для быстрого отскакивания клетки в направлении, противоположном выбрасыванию нити, или для непосредственного поражения врага.
Нематоцисты обнаружены у немногих динофлагеллят (у родов Nematodinium, Warnowia, Proterythropsis и некоторых представителей рода Polykrikos, причём у последних нематоцисты заметно отличаются по своей морфологии). Они крупнее трихоцист, достигают 20 мкм в длину; иногда их сравнивают с книдоцитами стрекающих.
Мукоцисты — простые мешки, выделяющие слизь на поверхность клетки; они близки к аналогичным структурам эвгленовых. Такие мешки найдены у некоторых динофлагеллят в области брюшного поля (пластинок продольной борозды) под клеточной мембраной.
В условиях, которые не позволяют популяции увеличиваться дальше (например, при низком содержании питательных веществ), динофлагелляты могут образовывать неподвижные покоящиеся споры, или цисты. Цисты динофлагеллят (диноцисты) у большинства видов морфологически отличаются от вегетативных клеток. Они достигают 30—70 мкм в диаметре, имеют гладкую или шероховатую поверхность. Новообразованные цисты вида Scrippsiella trochoidea содержат в 10 раз больше углеводов, чем вегетативные клетки; при этом скорость окислительных процессов у них составляет 1,5 % от вегетативной стадии. Клеточные стенки цист отличаются высокой стойкостью и содержат диноспорин, который химически близок к спорополленину, покрывающему пыльцу высших растений. Процесс инцистирования, или образования покоящихся спор, регулируется сложным взаимодействием внешних факторов, в числе которых длина дня, температура и концентрация питательных веществ. В ходе инцистирования содержание мелатонина повышается в несколько раз, что может предотвращать окисление липидов цисты.
После своего образования циста перемещается на дно озера или океана, где сохраняет жизнеспособность многие годы. Океанические течения могут переносить эти бентосные цисты в другие места. Когда условия становятся благоприятными, цисты могут прорастать, давая начало живым плавающим клеткам. Образование цист, их перемещение и прорастание объясняют многие аспекты экологии и географии ядовитого цветения воды, вызванного динофлагеллятами. Этим объясняется тот факт, что цветения воды не обязательно происходят каждый год в одном и том же месте, а также то обстоятельство, что цветение воды связано с переизбытком питательных веществ в океане, вызванным, например, попаданием в него сточных вод или стоков с агрикультурных предприятий. Кроме того, из-за цист места цветения воды могут сменять друг друга каждый год.
Цисты можно опознать по отсутствию хлоропластов, наличию микрогранулярной коричневой цитоплазмы и красного глазка (если вегетативная стадия имеет глазок). У некоторых видов обызвествление осуществляется путём откладывания кристаллов карбоната кальция в узком промежутке между клеточной стенкой и плазматической мембраной. Цисты Ceratium hirundinella имеют наружный кремниевый слой.
Динофлагелляты могут размножаться вегетативным, бесполым и половым путём. При вегетативном размножении клетка делится надвое в продольном или наклонном направлении, при этом оболочка исходной клетки по-разному участвует в создании оболочек дочерних клеток. При бесполом размножении динофлагелляты образуют зооспоры и апланоспоры. При недостатке азота и изменении температуры воды некоторые виды динофлагеллят приступают к половому размножению. У динофлагеллят описаны три типа полового процесса: изогамия, анизогамия и хологамия.
Планктонные формы динофлагеллят, обитающие во всех океанах мира, обладают способностью к люминесценции (часто её ошибочно называют фосфоресценцией), которую часто можно видеть летом ночью в потревоженной океанской воде. Люминесцирующие динофлагелляты населяют в основном поверхностные воды. Среди фотосинтезирующих протистов динофлагелляты — единственная группа, способная к биолюминесценции; всего зарегистрировано около 85 фотосинтезирующих и гетеротрофных видов морских динофлагеллят, обладающих такой способностью (пресноводные виды ею не обладают). Возбуждённые клетки испускают вспышки сине-зелёного цвета (с максимумом излучения на длине волны 475 нм) длительностью обычно от 0,05 до 5 с.
Люминесценция воды, возникающая при движении корабля, и свечение тропических заливов обусловлены в основном видами класса Dinophyceae; в умеренных широтах свечение моря обычно вызывает ночесветка. Словно бы пылающая морская вода с древних времён привлекала внимание мореплавателей, и долгое время этому сиянию воды приписывали сверхъестественное происхождение, пока в 1830 году не была экспериментально доказана биологическая природа данного свечения. Немецкий естествоиспытатель Курт Тезинг дал такое описание картины свечения моря, которую наблюдал в Неаполитанском заливе: «То вблизи, то вдали вспыхивали светлые, синеватые искры, вспыхивали и тотчас же исчезали. Всё сильнее и больше становился блеск моря, каждая волна при падении рассыпалась огненным дождём… Казалось, что мы плывём по лучезарному потоку; все вещи, опущенные в воду, казались залитыми жидким серебром. Когда мы поздно ночью стояли на молу, вся гавань и всё море, насколько хватал глаз, казались залитыми мягким огнём. Это было неописуемое зрелище, и лишь тот, который видел его во всей красе, может понять то неизгладимое впечатление, которое оно производит на каждого человека».
Наиболее сильное свечение характерно для Pyrocystis, который обусловливает свечение морей в экваториальных районах. Клетка Pyrocystis испускает в 1000 раз больше света, чем особи рода Lingulodinium, и в 100 раз больше, чем клетка Ceratium fusus, Peridinium pentagonium и Pyrodinium bahamense.
Существует две теории относительно адаптивного значение биолюминесценции у динофлагеллят.
Гипотеза «сигнала тревоги». Согласно этой гипотезе, при появлении беспозвоночного, питающегося динофлагеллятами, динофлагелляты испускают свечение и указывают на нахождение этого животного. Этот сигнал воспринимается хищником, питающимся поедающим динофлагеллят беспозвоночным.
Гипотеза «испуга». Согласно этой гипотезе, вспышка света, которую испускает динофлагеллята при механическом раздражении, вызываемым поедателем (например, веслоногим рачком), пугает рачка и заставляет его уплыть прочь.
Среди свободноживущих динофлагеллят встречаются как морские (около 90 % всех динофлагеллят обитают в морских водах, где входят в состав как планктона, так и бентоса), так и пресноводные формы. Наряду с диатомовыми водорослями они составляют основную часть морского фитопланктона, причём в тропических водах доминируют динофлагелляты, а в полярных и приполярных — диатомеи. Динофлагелляты занимают второе место после диатомей как продуценты первичной продукции в прибрежных морских водах. Наибольшее разнообразие и обилие динофлагеллят наблюдаются в неритической зоне океанов, где за счёт смывов с берегом содержится больше органики, однако они обитают и в пелагиали (открытая вода) в тропиках и субтропиках.
Среди бентосных динофлагеллят выделяются две основные экологические группы: псаммофильные виды, обитающие в морском песке, и эпифитные виды, живущие на морских водорослях и травах (к эпифитным динофлагеллятам, в частности, относятся известные своей токсичностью представители родов Prorocentrum и Gambierdiscus).
В пресноводном фитопланктоне динофлагелляты представлены значительно беднее (здесь наиболее часто встречаются виды Ceratium и Peridinium).
Судя по всему, большинство динофлагеллят — космополиты. Они встречаются в северных, умеренных и южных широтах, хотя наиболее обильны в тёплых водах, где их численность высока в течение всего года. В умеренных областях наибольшая численность динофлагеллят достигается только поздней весной и летом. Как в морских, так и в пресных водах перед массовым развитием динофлагеллят происходит массовое развитие диатомей. Существуют криофильные динофлагелляты, которые обитают в снегу и придают ему красную окраску. Среди динофлагеллят эпифиты и типичные бентосные формы гораздо более редки, чем планктонные.
Динофлагелляты способны за короткий срок давать большую биомассу, а потому служат важным звеном в цепи питания рыб, китообразных, коловраток, различных простейших и других гидробионтов — как непосредственно, так и после предварительного поедания представителями зоопланктона.
Нередко динофлагелляты выступают как симбионты (зооксантеллы) других организмов — морских беспозвоночных (губок, рифообразующих кораллов, медуз, плоских червей, моллюсков) и протистов (радиолярий, фораминифер, инфузорий). В каждом из случаев симбиоза хозяину достаточно, проглотив динофлагелляту, инкорпорировать её, не переваривая в свои ткани; после этого симбионт неоднократно делится и начинает синтезировать углеводы, которыми снабжает организм хозяина. Степень зависимости хозяина от такого симбиоза варьирует весьма значительно: так, актиния Anemonia вполне может прожить без зооксантелл, а некоторые кораллы без симбионтов-динофлагеллят обойтись не могут. Известно по крайней мере 5 видов динофлагеллят (4 из которых относятся к роду Symbiodinium), которые могут выступать в роли зооксантелл. Зооксантеллы покрыты тонкой оболочкой, способны к вегетативному размножению и образованию гимнодиниумподобных спор. Лучше всего изучена ассоциация зооксантелл и рифообразующих кораллов. Она представляет собой случай мутуализма, и оба организма извлекают из него выгоду. Зооксантеллы отдают своему хозяине до 50 % фиксированного углерода, большая часть которого поступает в виде глицерина, меньшая — в виде глюкозы, аланина, органических кислот. Показано, что кораллы с зооксантеллами быстрее обызвествляются. Коралл же снабжает водоросль некоторыми органическими соединениями (мочевина, глицерол-3-фосфат, аминокислоты) и факторами роста.
Часто динофлагелляты сами являются хозяевами для многих симбиотических водорослей, как эндо-, так и экзосимбионтов. Так, цианобактерии Synechocystis и Synechococcus живут на поверхности клеток бесцветных динофит Citharister, Histioneis, Ornithocercus и Parahistioneis. Эндосимбионтами динофлагеллят могут выступать криптофитовые, зелёные и золотистые водоросли, а также бактерии. Так, в водах Новой Гвинеи встречаются зелёные Noctiluca, которые, однако, лишены хлоропластов, но содержат от 6 до 12 тысяч маленьких клеток зелёной водоросли Pedinomonas.
Около 20 видов динофлагеллят являются паразитами беспозвоночных (в частности, мелких ракообразных), рыб и нитчатых водорослей. Широко распространённая динофлагеллята Dissodinium pseudolunula — эктопаразит яиц копепод, имеет хлоропласты. Эктопаразиты, лишённые хлоропластов, паразитируют на рыбах, яйцах копепод, диатомеях и других водорослях (представители родов Chytriodinium, Paulsenella и др.). Два рода эндопаразитических динофлагеллят, обитающих в яйцах копепод — Blastodinium и Schizodinium — имеют хлоропласты. Эндопаразиты, не имеющие хлоропластов, обитают в других динофлагеллятах, сифонофорах и копеподах. Грань между симбионтами и паразитами иногда бывает размыта; так, виды рода Symbiodinium обычно выступают как симбионты кораллов и других беспозвоночных, но некоторые их виды ведут себя скорее как паразиты и способны вызывать массовую гибель кораллов.
Ряд видов динофлагеллят, размножаясь в значительных количествах, способны вызывать цветение воды. В частности, вспышка численности морских динофлагеллят в прибрежных водах, происходящая обычно в тёплое время года, приводит к явлению, получившему название «красный прилив» (в морях России красные приливы наблюдаются на Дальнем Востоке). Во время красного прилива морская вода из-за наличия в ней микроскопических водорослей, содержащих фотосинтетические пигменты, приобретает необычный цвет — обычно красный, но иногда вишнёвый, коричневый или жёлтый. Появлению красных приливов способствуют свойства воды: тёплые поверхностные слои, высокое содержание питательных веществ, низкая солёность (что часто бывает после длительного периода дождей) и отсутствие сильного волнения. Если виды, вызывающие красный прилив, являются токсинообразующими, то результатом может быть массовая гибель рыб, моллюсков и ракообразных, питающихся планктоном. Стоит отметить, что многие токсинообразующие динофлагелляты не изменяют окраску воды. Так, в конце августа 1987 года на западном побережье Флориды наблюдался самый сильный красный прилив за последние 150 лет, и пляжи были завалены сотнями тысяч мёртвых рыб. За это оказалась ответственной динофлагеллята Gymnodinium breve.
В пресноводных водоёмах цветение воды обычно вызывают водоросли, не относящиеся к динофлагеллятам; однако в некоторых местностях последние (особенно представители родов Peridinium, Ceratium и Peridiniopsis) играют ведущую роль. Так, Ceratium hirundinella при массовом размножении придаёт воде буровато-белесую окраску (что можно наблюдать летом в заводях и пойменных озёрах рек Украины). Другой известный пример: наблюдавшееся до 1964 года в отдельные солнечные дни (в период с июля по сентябрь) окрашивание в интенсивный кроваво-красный цвет вод горного озера Лаго ди Товель, лежащего на северо-западе провинции Тренто, Италия. Длительное время полагали, что за такое окрашивание ответственны виды рода Glenodinium, однако позднее было установлено, что окрашивание вызывали представители описанного в 2006 году вида Tovellia sanguinea (они встречаются в озере и ныне, но в небольшом количестве: преобладание перешло к другой динофлагелляте Baldinia anauniensis, имеющей зелёную окраску).
С размножением пресноводных динофлагеллят обычно связывают и явление «Красного Нила». Так называют вторую фазу летне-осеннего половодья в нижнем течении Нила, во время которого река выходит из берегов, оставляя затем на прилегающих полях очередной слой плодородного ила (во время первой фазы — «Зелёного Нила» — река благодаря присутствующим в ней микроскопическим водорослям принимает зелёный цвет, а во время второй — тёмно-красный). Конкретные виды динофлагеллят, ответственные за данное явление, пока не выявлены (некоторые авторы выдвигали предположения, по которым причиной красной окраски нильских вод могли быть цианобактерия Oscillatoria rubescens, эвгленоидея Euglena sanguinea или же зелёная водоросль Haematococcus pluvialis, но надёжного обоснования эти гипотезы не получили).
Ископаемые остатки динофлагеллят известны со среднего триаса; в то же время остатки двух родов, Arpylorus и Palaeodinophysis, предположительно относящихся к динофлагеллятам, датируются соответственно поздним силуром и девоном. В раннем мезозое разнообразие динофлагеллят резко возросло, возникли их основные морфологические типы. В мелу динофлагелляты достигли наивысшего морфологического разнообразия, но с неогена происходит его уменьшение.
В ископаемом состоянии динофлагелляты представлены преимущественно диноцистами, которые благодаря своей стойкости хорошо сохраняются в древних осадочных породах. Находки диноцист известны с перми, а для стратификации мезозоя и кайнозоя они имеют большое значение, поскольку по ним разработаны локальные стратифические шкалы. Представляют они ценность и для палеоэкологических исследований. Ископаемые цисты динофлагеллят называются гистрихосферидами (гистрихоспорами) и рассматриваются в составе акритархов — похожих на цисты структур ископаемых одноклеточных водорослей. При этом уже у раннекембрийских акритархов (520 млн лет назад) найдены диностеролы и 4α-метил-24-этилхолестен, специфичные для динофлагеллят; если предположения о связи акритархов и динофлагеллят верны, то эволюционную историю последних можно продлить до раннего кембрия и даже более раннего периода (800 млн лет назад).
Гистрихосфериды были открыты независимо разными палеонтологами и классифицируются как отдельная таксономическая группа, представленная исключительно ископаемыми видами; многие покоящиеся споры динофлагеллят идентичны кайнозойским гистрихосферидам, так что в этом случае одна структура имеет два названия. С перми известны и ископаемые остатки панцирей динофлагеллят, но они встречаются значительно реже.
Значение динофлагеллят в биосфере весьма велико. В экосистеме коралловых рифов фототрофные динофлагелляты являются наиболее важными первичными продуцентами, а среди первичных морских продуцентов в целом они уступают только диатомеям. Гетеротрофные же морские динофлагелляты оказываются важными компонентами так называемой микробной петли, помогая направить значительные потоки энергии в планктонные пищевые сети.
Места с повышенной численностью динофлагеллят (места «цветения») являются одними из важнейших естественных хранилищ жиров (например, хранилище в Северном море). Ископаемые породы, содержащие отмершие клетки динофлагеллят, также служат хранилищами накопленных в клетках жиров. Крупнейшее из таких хранилищ — сланцы Киммеридж (Kimmeridge Clay) в Англии. В месторождениях нефти и древних осадочных пород также встречается 4α-метилстирольные углеводороды, которые, вероятно, произошли от 4α-метилстиролов динофлагеллят.
Паразитические динофлагелляты способны вызывать эпизоотии в популяциях рыб и ракообразных. Многие динофлагелляты весьма чувствительны к присутствию в воде органических загрязнений, а поэтому используются как биоиндикаторы при санитарно-биологическом обследовании водных ресурсов. Ископаемые цисты динофлагеллят служат геологам для оценки возраста предполагаемых месторождений нефти.
Некоторые виды динофлагеллят способные образовывать опасные токсины; среди видов токсинообразующего фитопланктона на долю динофлагеллят приходится около 75—80 %, причём выделяемые динофлагеллятами токсины относятся к числу наиболее мощных из известных биотоксинов. Известно около 60 видов токсинообразующих динофлагеллят; более половины из них относятся к родам Prorocentrum, Alexandrium и Dinophysis, в меньшей степени к Gambierdiscus, Ostreopsis и Karenia.
Некоторые токсины, вырабатываемые динофлагеллятами, летучи, и известны случаи, когда после вдыхания или попадания на кожу они вызывали отравление у учёных, изучавших динофлагеллят. Токсины динофлагеллят могут быть водо- и жирорастворимыми, оказывают цитолитическое, гепатотоксическое и нейротоксическое действие.
Нередко (особенно во время красных приливов) эти токсины накапливаются в тканях двустворчатых моллюсков, питающихся фитопланктоном, не причиняя им особого вреда, однако их уровень в тканях оказывается токсичным (зачастую летальным) для поедающих моллюсков позвоночных — для рыб или человека.
Некоторые динофлагелляты, выделяющие токсины, способные поражать рыб, млекопитающих и птиц, имеют большое хозяйственное значение, поскольку наносят вред хозяйствам, занимающимся разведением морских животных. К числу таких видов относится Pfiesteria piscicida, отличающаяся своим необычным жизненным циклом. Токсины, выделяемые этой динофлагеллятой, столь сильны, что рыбы, питающиеся ими (например, атлантическая сельдь, южная камбала), погибают уже через несколько минут. За массовую гибель рыб во время красных приливов у атлантического побережья США несёт ответственность именно этот вид динофлагеллят.
Динофлагеллята Crypthecodinium cohnii используется для получения в промышленных масштабах докозагексаеновой (цервоновой) кислоты, относящейся к важнейшим для организма человека незаменимым омега-3-полиненасыщенным жирным кислотам.
На протяжении почти всего XX века классификация динофлагеллят разрабатывалась протозоологами и ботаниками-альгологами практически независимо. Протозоологи трактовали таксон как отряд Dinoflagellida Butschli, 1885 в составе типа Protozoa; альгологи — либо как класс Dinophyceae Pascher, 1914 в составе отдела Pyrrophyta (включавшего также класс Cryptophyceae, а иногда — ещё и класс Chloromonadophyceae), либо как самостоятельный отдел Dinophyta Dillon, 1963. При этом внутреннюю классификацию динофитовых водорослей ботаники строили на основе типа организации таллома, выделяя порядки Peridiniales (монадные формы), Dinamoebidiales (ризоподиальные формы), Gloeodiniales (пальмеллоидные формы), Dinococcales (коккоидные формы), Dinothrichales (нитчатые формы). В конце XX века эти ранние классификации полностью устарели.
Современные исследования в области систематики динофлагеллят основаны на анализе их филогении, проводимом с учётом как морфологических, так и молекулярных данных. Согласно данным филогенетического анализа (2014 год), основывавшегося на сопоставлении нуклеотидных последовательностей РНК 76 видов альвеолят и гетероконтов (представители последних играли роль внешней группы), динофлагелляты в пределах альвеолят представляют собой монофилетическую группу, сестринскую апикомплексам, причём общий предковый таксон динофлагеллят и апикомплексов был сестринским по отношению к инфузориям. В пределах динофлагеллят из участвовавших в анализе видов наиболее рано отделился Perkinsus marinus (по другим классификациям, перкинсей и родственные им группы включают не в состав динофлагеллят, а в состав сестринской по отношению к ним группы Perkinsozoa), вслед за ним — Oxyrrhis marina, ещё позже — клада синдиней, а оставшиеся группы динофлагеллят (динокариоты) образуют сестринскую по отношению к ней кладу. Внутренние взаимоотношения представителей динокариот ещё не ясны, а имеющиеся данные во многом противоречивы.
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ОТРЯД ПАНЦИРНЫЕ ЖГУТИКОНОСЦЫ, ИЛИ ПЕРИДИНЕИ (DINOFLAGELLATA, ИЛИ PERIDINEA)
Панцирные жгутиконосцы — обширная группа простейших, широко распространенная в морях и пресных водах. Подавляющее большинство видов этого отряда относится к планктонным формам.
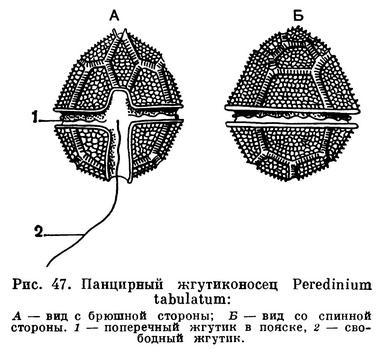
Характерной особенностью панцирных жгутиконосцев является своеобразное строение жгутикового аппарата (рис. 47). Имеющиеся у них два жгута берут начало рядом друг с другом на одной стороне тела простейшего, которую условно называют брюшной. Один жгутик направлен назад и свободно выдается в окружающую среду. Начальная часть его лежит в небольшом желобке, идущем вдоль тела. Второй жгутик, называемый поперечным, опоясывает все тело в экваториальной плоскости и расположен в довольно глубоком желобе, называемом поясом (cingulum), который опоясывает все тело. Поперечный жгут производит характерные волнообразные движения, что создает ложное впечатление, будто в поперечной борозде расположен ряд ресничек.
Вторая характерная особенность большинства (но не всех) панцирных жгутиконосцев — наличие у них оболочки, состоящей из клетчатки. Эта оболочка складывается из строго определенного числа закономерно расположенных отдельных пластинок, соединенных друг с другом швами.
Цитоплазма жгутиконосцев обычно бывает окрашена в буроватый цвет, который обусловлен наличием в ней большого количества мелких дисковидных хроматофоров, содержащих хлорофилл и некоторые дополнительные пигменты. Продуктом ассимиляции является крахмал, зерна которого рассеяны в цитоплазме.
Рассмотрим представителей двух наиболее широко распространенных родов динофлагеллят.
На рисунке изображен часто встречающийся в пресных водах перидиний (Peridinium tabulatum). У него отчетливо выражены упомянутые выше пластинки. Форма тела более или менее правильно яйцевидная. Иногда бурное развитие перидиний вызывает «цветение» воды.
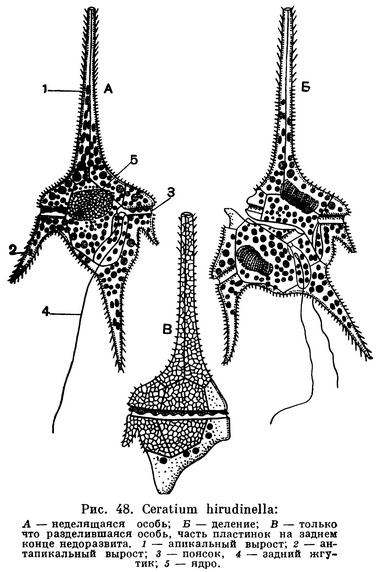
Широко распространены как в пресной, так и в морской воде многочисленные виды рода церациум (Ceratium). Характерной особенностью представителей этого рода являются длинные выросты. У пресноводного Ceratium hirudinella (рис. 48) один вырост (апикальный) направлен вперед и три (антапикальных) — назад. Особенно сильно эти выросты выражены у морских видов Ceratium, у которых они вторично ветвятся (рис. 49). Для морских же видов характерно образование временных колоний-цепочек (рис. 50), которые возникают в результате того, что разделившиеся особи остаются некоторое время связанными друг с другом. Длинные отростки у морских видов Ceratium представляют собой приспособление к планктонному образу жизни. Отростки увеличивают поверхность тела, что способствует «парению» в воде. Аналогичные приспособления мы уже видели у радиолярий.
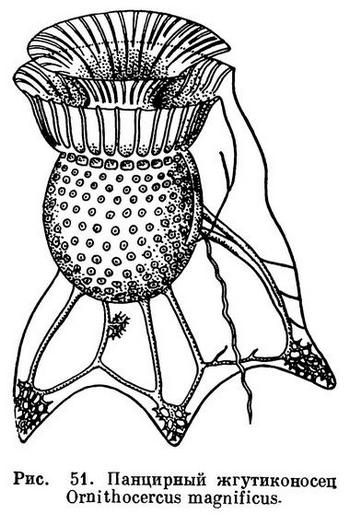
Особенно причудливую форму приобретает панцирь у представителей рода Ornithocercus, где по краям борозд образуются длинные крыловидные выросты (рис. 51). Это тоже приспособление к «парению» в воде.


В планктоне теплых и тропических морей очень часто, и нередко в огромных количествах, встречается жгутиконосец ночесветпка (Noctiluca miliaris). В летнее время это простейшее обычно развивается и в Черном море.
Ночесветка имеет шаровидное тело до 2 мм в диаметре. В отличие от других панцирных жгутиконосцев, она не имеет оболочки из клетчатки и лишена хроматофоров (рис. 52). Поэтому ночесветка не способна к фотосинтезу. В отличие от большинства Dinoflagellata, ночесветке свойственно животное (анимальное) питание. Она заглатывает и переваривает различные оформленные пищевые частицы: мелкие водоросли, мелкие виды простейших.
На одной стороне шаровидного тела ночесветки имеется углубление, соответствующее продольному желобку типичных панцирных жгутиконосцев. В глубине его помещается отверстие, выполняющее функцию рта. Через него и происходит поглощение пищи. В области этого же углубления помещаются два жгута. Один из них короткий и толстый. Он скорее напоминает щупальце. Этот жгут совершает относительно очень медленные колебательные движения (не более 10 в минуту), способствующие направлению пищевых частиц к ротовому отверстию. Второй, более короткий и тонкий жгутик также расположен в области ротового углубления (рис. 52, 4).
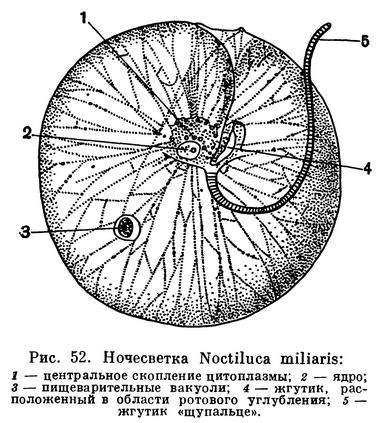
Очень своеобразно строение эндоплазмы ночесветки. Непосредственно под ротовым отверстием располагается центральное скопление цитоплазмы, от которого к периферии во всех направлениях отходят тонкие тяжи. Вся же остальная масса тела заполнена жидкостью и представляет собой систему вакуолей. Ядро расположено в центральном скоплении цитоплазмы. В цитоплазматических тяжах имеются многочисленные жировые включения.
Описанное выше своеобразное строение цитоплазмы следует рассматривать как одну из форм приспособления к планктонному образу жизни. Сильная вакуолизация цитоплазмы и наличие жировых включении ведут к уменьшению удельного веса и, следовательно, способствуют «парению» в толще воды.
У ночесветки есть еще одна интересная особенность, за которую она и получила свое название. При раздражении (механическом, химическом и т. п.) ночесветка ярко «вспыхивает». Это явление незаметно днем, при ярком свете, но очень отчетливо выражено в темноте, ночью. Ночесветка — это один из организмов, вызывающих явление свечения, столь характерное для теплых морей. Всякий, кому приходилось в теплую летнюю ночь плыть по Черному морю в лодке или на пароходе, вероятно, имел возможность наблюдать это эффектное явление. При ударе весел по воде, при падении капель воды с весел в море, при вращении пароходного винта и т. п. вода начинает светиться слабым фосфорическим, но вполне отчетливым светом. При этом видны бывают отдельные вспышки, напоминающие искорки.
Свечение наступает в результате окисления жировых включений, о наличии которых уже говорилось выше.
Свечение имеет, вероятно, защитное значение (отпугивает хищников).
Некоторые виды панцирных жгутиконосцев являются симбионтами радиолярий. Недавно выяснено, что они в большом количестве поселяются и в мягких тканях коралловых полипов.