Gaidropsarus mediterraneus (Linnaeus, 1758)
Средиземноморский трехусый морской налим, галея (Rus),
Shore rockling
(Eng)
Синонимы:
Gadus argenteolus Montagu, 1818
Gadus fuscus Risso, 1810
Gadus jubatus Pallas, 1814
Gadus mediterraneus Linnaeus, 1758
Gadus mustella Risso, 1810
Gadus tricirratus Brünnich, 1768
Gaidropsarus mustellaris Rafinesque, 1810
Gaidropsarus zernovi Gratzianow, 1907
Merlangus communis Costa, 1844
Motella communis Costa, 1844
Motella fusca Moreau, 1881
Motella maculata Moreau, 1881
Motella mediterranea Lütken, 1882
Motella tricirrata Nilssоn, 1832
Onos fusca Risso, 1826
Onos maculata Risso, 1826
Onos mediterraneus Collett, 1885
Onos mustella Risso, 1826
Onos sellai Cipria, 1938
Onos tricirratus Smitt, 1893
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Налим морской — Gaidropsarus maditerraneus (Linnaeus, 1758)
Тело прогонистое, сильно сужающееся к хвосту. Два спинных плавника, из которых первый имеет вид одного длинного луча с тянущейся за ним короткой кожной складкой (обычно погружена в желобок на спине рыбы). Второй спинной и анальные плавники длинные. Есть три усика — подбородочный и пара, начинающаяся у передних ноздрей. D1 1. D2 53—58. А 44—49. Окраска тела желтовато-оливковая. Длина тела 50 см. Прибрежная рыба, обитающая на мелководьях у дна в Черном море.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
СРЕДИЗЕМНОМОРСКИЙ ТРЕХУСЫЙ МОРСКОЙ НАЛИМ — GAIDROPSARUS MEDITERRANEUS (LINNE)
Условия размножения. Период размножения налима в Черном море значительно растянут и приходится в основном на холодное время года. Согласно данным различных исследователей (Водяницкий, 1930а; Косякина, 1938; Виноградов, 1948, 1949; Дука, 1958; Зайцев, 1959а; Георгиев и др., 1960), нерест налима начинается в сентябре при температуре воды 19,5—19,8° и заканчивается в апреле — мае. Наиболее интенсивный нерест происходит в октябре — декабре при температуре 10,5—15,0°. В Одесском заливе развивающиеся икринки налима встречались в сентябре — декабре при температуре 9—16° и солености 14,36—17,89‰ (Зайцев, 1959а).
В Средиземном море морской налим размножается также в холодное время года. По наблюдениям Рафаэле (1888) и Ло Бианко (1888, 1908 —1909), в Неаполитанском заливе икринки встречаются в фаопланктоне с ноября по март, личинки — с ноября по апрель. Мальки держатся среди плавающих водорослей и встречаются в марте. Более поздними исследованиями (Aboussouan, 1964; Lee, 1966) подтверждено нахождение икринок и личннок этого вида в Марсельском заливе с января по апрель. У берегов Ирландии и в районе Плимута морской налим нерестится летом — в июне — августе (Holt, 1893).
Приведенные данные свидетельствуют о том, что нерест и развитие икринок и личинок морского налима происходят при значительных колебаниях температуры (9—20°) и солености (14—38‰).
Строение икринок. Икринки морского налима пелагические, сферической формы, мелкие, с одной жировой каплей (рис. 7). Оболочка икринок очень тонкая, гладкая, совершенно прозрачная. Перивителлиновое пространство узкое — от 2,5 до 9,75% диаметра икринки (Дука, 1958). По данным В. А. Водяницкого (1930а), диаметр икринок колеблется от 0,70 до 0,85 мм, диаметр жировой капли равен 0,15 мм. Ю. П. Зайцев (1959а) приводит соответствующие размеры: диаметр икринок 0,75—0,83 мм, средний размер 0,78 мм, жировой капли — 0,15—0,18 мм (в среднем 0,16 мм). Размеры икринок налима у Болгарского побережья колеблются от 0,76 до 0,89 мм, жировой капли — от 0,15 до 0,19 мм (Георгиев и др., 1960). По данным Л. А. Дука, икринки налима несколько крупнее — 0,85—0,95 мм в диаметре. Размер живых икринок, по нашим измерениям, 0,78—0,90 мм (в среднем 0,81), диаметр жировой капли 0,13—0,19 мм (в среднем 0,15 мм). Икринки этого вида из Средиземного моря мельче. Их диаметр колеблется от 0,66 до 0,72 мм, жировой капли — от 0,15 до 0,19 мм (Aboussouan, 1964).
Эмбриональное развитие. Эмбриональное и постэмбриональное развитие налима изучено в лабораторных условиях. Опыты были поставлены на икринках, собранных в море, и параллельно на искусственно оплодотворенных икринках в январе — феврале 1966 г. Температура воды в опытах колебалась от 11 до 16,6°.
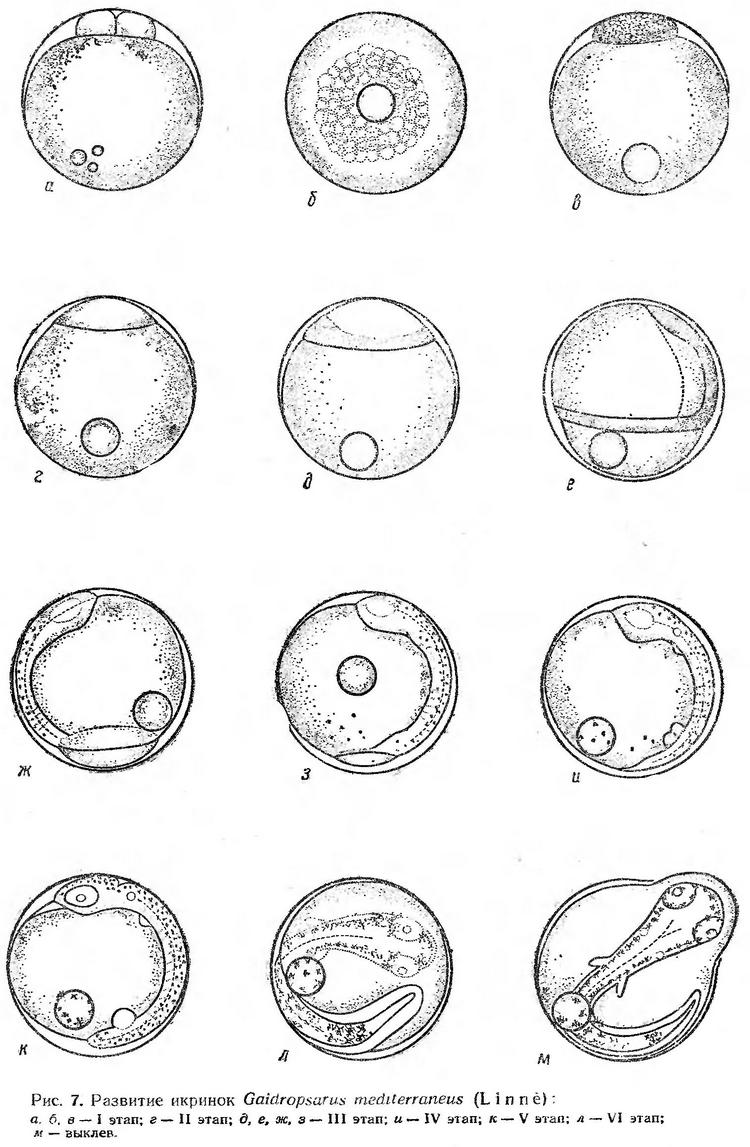
В момент вымета икринки содержат большое количество (около двух десятков) мелких жировых капель, разбросанных по поверхности желтка. Через час после оплодотворения оболочка разбухает, образуется узкая перивителлиновая полость. При температуре воды 13,5° через 2,5 ч после оплодотворения образуется два крупных бластомера. Жировые капли стягиваются к вегетативному полюсу и сливаются. Ко времени образования двух бластомеров в желтке остается 3—7 жировых капель разных размеров (рис. 7, а).
При дальнейшем развитии, еще до образования крупноклеточной морулы, в желтке остается одна жировая капля, которая всегда занимает верхнее положение, перемещаясь при повороте икринки по поверхности желтка кверху. Процесс дробления при температуре 13,5—14,0° продолжался 12—14 ч. Бластодиск на стадии мелкоклеточной морулы занимает очень небольшую часть поверхности желтка. Диаметр бластодиска составляет 0,35—0,37 мм, высота — 0,10—0,12 мм (рис. 7, б, в, табл. 10).

Переход к эпителиальной бластуле, как и у хамсы, сопровождается появлением пузыревидных образований в перибласте, которые по мере развития сползают к вегетативному полюсу или, вернее, поднимаются, поскольку икринки в плавучем состоянии всегда обращены анимальным полюсом вниз. Образование эпителиальной бластулы при температуре 13,2—14,0° продолжается около 6 ч (рис. 7, г). Почти одновременно с обрастанием желтка бластодиском происходит подворачивание его краев и вскоре образуется зародышевый бугорок (рис. 7, д). Когда бластодиск охватывает примерно 2/3 поверхности желтка, зародышевая полоска четко выражена. Головной конец заметно расширен. Прослеживается нервный тяж, купферов пузырек (рис. 7, е). Через 12—14 ч после начала гаструляции непокрытой остается небольшая часть желтка. Образуются зачатки глаз, начинается сегментация туловищного отдела, на спинной стороне появляются точечные черные пигментные клетки, которые вскоре распространяются на желток (рис. 7, ж, з). Ко времени замыкания бластопора головной конец зародыша достигает анимального полюса; в туловищном отделе насчитывается 6—7 сегментов. Глаза четко обозначены. Намечается зачаток сердца, кишечник, грудные плавники, слуховые капсулы. Усиливается меланинная пигментация дорсальной стороны тела (рис. 7, и). Процесс обрастания желтка бластодиском и образования зародышевой полоски при температуре 13,0—14,5° продолжался 28—30 ч.
После замыкания бластопора начинается быстрая сегментация тела, происходит дальнейшее формирование заложенных зачатков органов. Тело эмбриона интенсивно пигментируется. Меланофоры располагаются двумя рядами вдоль спинной стороны тела и между рядами разбросаны более мелкие пигментные клетки. Интенсивно пигментирована верхняя часть головы; появляется пигмент на жировой капле — на той части ее поверхности, которая обращена к зародышу. В плавучем состоянии пигментированная сторона зародыша и соответственно жировая капля обращены вниз.
Через 12—15 ч после замыкания бластопора появляется хвостовая почка, начинается отделение хвоста от желтка и рост хвостового отдела (рис. 7, к). К этому времени эмбрион охватывает чуть больше половины поверхности желтка. Когда от желточного мешка отчленяется около 10 сегментов, начинаются очень слабые подергивания эмбриона и медленная, еле уловимая, пульсация сердца — 36—37 раз в минуту (рис. 7, л). Промежутки между движениями эмбриона составляют 45—50 сек.
По мере развития эмбриона паузы сокращаются. К началу выклева подергивания эмбриона наблюдаются через каждые 10—30 сек. К этому времени эмбрион охватывает 2/3 поверхности желтка (рис. 7, м). Пульсация сердца учащается до 42—44 раз в минуту. Перед выклевом голова плотно прижимается к оболочке и выпячивает ее. Затем в этом месте оболочка разрывается и через разрыв постепенно выталкивается голова вместе с желточным мешком. Процесс выклева продолжается несколько минут. Эмбриональное развитие морского налима при температуре 13,0—14,5° происходит в среднем в течение 4 суток (табл. 10).
Постэмбриональное развитие. Выклюнувшиеся личинки имеют длину 1,77—1,85 мм. Они снабжены большим яйцевидным желточным мешком. Жировая капля расположена у заднего края желточного мешка. Анальное отверстие открывается непосредственно за желточным мешком, как у личинок всех тресковых, сбоку плавниковой складки. Антеанальное расстояние составляет около 55% длины тела. Желточная оболочка и плавниковая кайма характеризуются своеобразным рисунком, образованным выпуклыми пузыревидными клетками эпителия. Выклюнувшиеся личинки, как и эмбрионы, имеют только черный пигмент. Звездчатые меланофоры расположены на голове, спине, переходят на боковые стороны и в средней части хвостового отдела размещены на вентральной стороне так, что образуется пигментный пояс (рис. 8, а). Личинки держатся в поверхностном слое в горизонтальном перевернутом положении (спинной стороной вниз). Длительное время они находятся в состоянии покоя, лишь изредка (1—2 раза в минуту) совершая короткие передвижения при помощи изгибов хвоста.
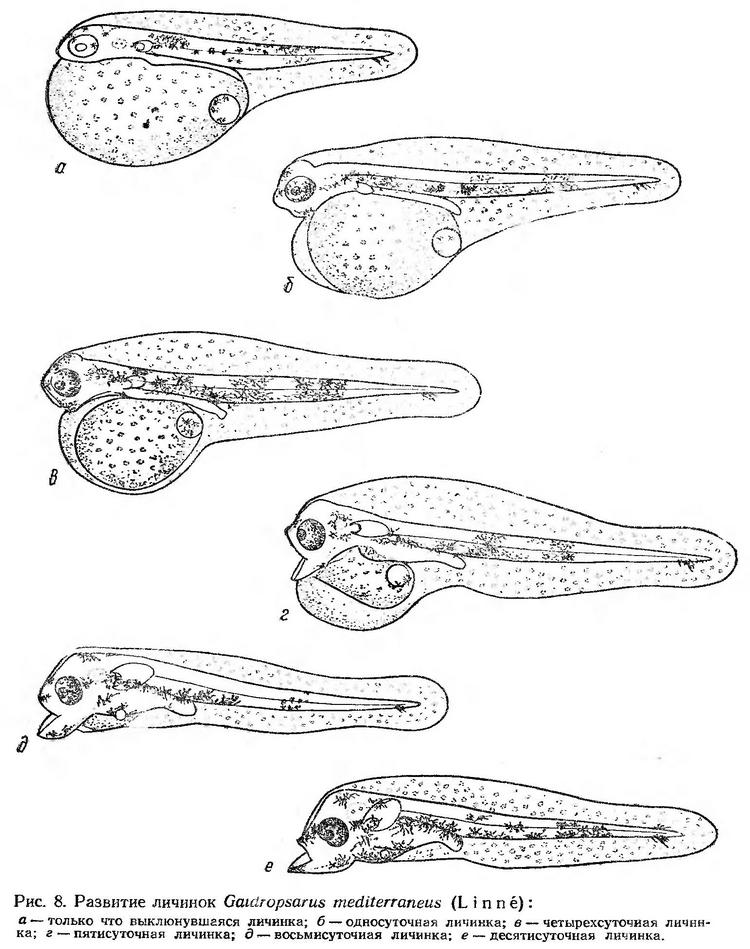
Односуточные личинки налима достигают 2,05—2,15 мм. Они по-прежнему пассивно парят в воде, сохраняя перевернутое горизонтальное положение. Голова отделяется от желточного мешка, намечается ротовая ямка. Пигментация усиливается. Наблюдается перемещение меланофоров на вентральную сторону. В хвостовом отделе выделяются два пигментных пояска — сразу за анусом и в средней части. Начинается пигментация глаз (рис. 8, б). Личинки длительное время находятся в покое, почти касаясь желточным мешком поверхностной пленки. Периоды покоя продолжаются 35—40 сек. Кратковременные перемещения продолжительностью 2—3 сек производятся за счет быстрых изгибов хвостового отдела. При этом личинки, сохраняя перевернутое горизонтальное положение, двигаются в одной плоскости или вращаются вокруг оси тела.
В возрасте трех суток личинки налима во время движения быстро принимают нормальное положение — спинной стороной вверх, затем при наступлении паузы снова медленно переворачиваются. В этом возрасте ротовая ямка четко обозначена, пигментация глаз усиливается. Грудные плавники увеличиваются, но еще неподвижны. В одном и том же опыте (икра получена от одной самки) личинки налима существенно различаются по пигментации. Тело многих личинок интенсивно пигментировано крупными ветвистыми меланофорами, густо расположенными в туловищном отделе и на голове и сгруппированными в два пигментных пояса в постанальном отделе. Некоторые личинки имеют точечную меланинную пигментацию. Пигментные пояса в таких случаях выражены не четко, туловище и голова пигментированы значительно менее интенсивно.
Длина четырехсуточных личинок увеличивается до 2,27—2,40 мм. Желточный мешок значительно сокращается в размере. Образуется ротовое отверстие, оформляются челюсти. Хвостовой отдел относительно длины тела увеличивается, а высота тела уменьшается (рис. 8, в). В плавниковой кайме в области головы образуется полость. Подобная полость свойственна, как считает С. Г. Крыжановский (1953), личинкам всех морских пелагофильных рыб, которые развиваются из чрезвычайно оводненной икры. Она образуется за счет избыточной воды в желточном мешке, которая по мере его рассасывания перемещается в нее. Периоды движения личинок также непродолжительны (до 2—3 сек), периоды покоя в отдельных случаях достигают 3—4 мин, обычно 20—45 сек. Пульсация сердца составляет 40—60 ударов в минуту.
Длина шестисуточных личинок и личинок старшего возраста (8—10-суточных) не изменяется или даже несколько уменьшается (в эксперименте). У личинок в возрасте шести суток (рис. 7, г) сохраняется небольшой остаток желточного мешка. Рот и грудные плавники приобретают подвижность. Кишечник утолщается и несколько изгибается. Туловище укорачивается и соответственно удлиняется хвостовой отдел. Личинки становятся очень подвижными. Периоды покоя значительно сокращаются (до 15—20 сек). Продолжительность разовых перемещений увеличивается до 15 сек. Благодаря подвижности грудных плавников личинки начинают плавать в разных направлениях. Рот при движении постоянно открыт. Морфологически личинка подготовлена к переходу на внешнее питание, хотя поисковый инстинкт в условиях опыта не выражен. В периоды покоя личинки держатся в косом положении, головой вниз, чему способствует заднее положение жировой капли. Меланофоры почти полностью перемещаются на вентральную сторону. Следовательно, изменение положения личинки в воде (косое в покое и нормальное горизонтальное при движении) приводит к перераспределению черного пигмента таким образом, что пигментированной оказывается в основном та сторона тела, которая обращена вниз.
Восьмисуточные личинки (рис. 8, д) почти непрерывно двигаются, периоды покоя измеряются несколькими секундами. Грудные плавники большие, вертикально поставленные, при движении быстро вибрируют. Сохраняется небольшой остаток желтка, жировая капля значительно уменьшается в размере. Нижняя челюсть несколько выдается за верхнюю, рот подвижен. Головной синус сокращается.
Десятисуточные личинки (рис. 8, е) в условиях опыта находятся в угнетенном состоянии. Длительный покой сменяется резкими стремительными бросками, после чего снова наступает период покоя, личинки переворачиваются на бок и как бы замирают. В этом возрасте желток почти полностью израсходован, сохраняется лишь небольшой остаток жировой капли. Рот ритмично подвижен; но поисковый инстинкт не выражен, хотя в аквариум внесен разнообразный корм (фитопланктон и мелкие формы зоопланктона). Личинки погибают, так и не начав питаться в опытных условиях.
Распределение и численность икринок и личинок. Икринки и личинки налима встречаются в планктоне Черного моря обычно в небольшом количестве (Водяницкий, 1930а; Зайцев, 1959а; Георгиев и др., 1960). Отмечая большие уловы икринок налима в Новороссийской бухте, Е. Г. Косякина (1938), однако, не приводит конкретных данных. В Севастопольской бухте, по наблюдениям Л. А. Дука (1958), наибольшее количество икры налима в разгар нереста в 1952 г. достигало 218 экз/м2, в 1953 г.— 52 экз/м2.
В наших материалах икринки налима встречались зимой в прибрежных районах Крымского и Кавказского побережья в количестве до 48 экз/м2. Распределение икринок ограничено приповерхностным слоем. При ловах на разных горизонтах икра налима отмечена в основном в горизонтальных поверхностных и вертикальных ловах. В планктоне личинки встречаются очень редко. За длительный период наблюдений в районе Новороссийской бухты З. М. Пчелина (1940) отметила нахождение только двух личинок налима. По данным Л. А. Дука (1958), в течение двух нерестовых сезонов (1952 и 1953—1954 гг.) в районе Севастопольской бухты было выловлено 17 недавно выклюнувшихся личинок и одна личинка размером 4,73 мм. В наших уловах личинки налима не обнаружены. Судя по интенсивной пигментации личинок, можно предположить, что они держатся преимущественно у дна в мелководных прибрежных районах, где их темная окраска имеет приспособительное значение.
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
* 3. Gaidropsarus mediterraneus (Linne) — Средиземноморский трехусый морской налим, галея (табл. I, XXII, LXVIII).
— Gadus mediterraneus Linne, Syst. nat., ed. X, 1758 : 255 (Oceano Europeo, II D 54, A 44, P 15).
— Gadus tricirratus Brünnich, Ichth. Massil., 1768 : 22 (II D 54, A 45, P 16).
— Gadus mustella, Gadus fuscus Risso, Ichth. Nice, 1810 : 215 (II D 52—54, A 44, P 14—18).
— Gaidropsarus mustellaris Rafinesque, Ind. ittiol. Sicil., II, 1810 : 51.
— Gadus jubatus Pallas, Zoogr. rosso-asiat., III, 1811 : 202 (Черное море).
— Gadus argenteolus Montegu, Mem. Wern. Nat. Hist. Soc., II, 1818 : 449 (P 16—18, juv.).
— Onos mustella, Onos maculata, Onos fusca Risso, Hist. nat. Eur. merid., III, 1826 : 215, 216 (II D 52—58, A 44—48, P 14—18).
— Motella tricirrata Nilssоn, Prodr. ichth. Scand., 1832 : 48 (по Bloch'y, 1795; II D 56, A 46, P 18).
— Motella communis Canestrini, Arch. Zool., Anat. Fisiol., II, 1862 : 369 (II D 52—55, A 44—47, P 17, V 6, Средиземное море).
— Motella vulgaris Steindachner, Sitzungsber. Akad. Wiss. Wien., math.-naturw. CI., LVII, 1, 1868 : 710 (ex parte).
— Motella tricirrata, Motella maculata, Motella fusca Moreau, Hist. nat. poiss. France, III, 1881 : 268, 270, 272.
— Motella mediterranea Lütken, Vidensk. Medd. naturhist. Foren. Kbnhvn., (1881), 1882 : 235 (II D 55—56, A 47—49, P 16, V 6—7).
— Motella tricirrata Day, Fish. Gr. Brit. Irel., I, 1884 : 317, pl. LXXXVIII, f. 1 (II D 55—57, A 48—50, P 16—18, библиография, распространение).
— Onos mediterraneus Collett, Nyt Mag. Naturv., XXIX, 1885 : 93.
— Onus mediterraneus Collett, Christ. Vidensk.-Selsk. Forh., (1891), 1892 : 6 (сводка).
— Onos mediterraneus, Onos fuscus et Onos tricirratus (ex parte) Smitt, Scand. fish., I, 1893 : 357, f. 131 (библиография, измерения).
— Gaidropsarus zernovi Грацианов, Опыт обзора рыб, 1907 : 463 (Черное море, Стрелецкая бухта).
Распространение, биология. Nordman, Faune Pontique, 1840 : 531, pl. XXVI, f. 1 (Черное море); Кесслер, Рыбы Арало-касп.-понт. обл., 1877 : 235 (Черное море); Пчелина, Тр. Новоросс. биол. ст., II, 1, 1936 : 33 (личинки); Дренски, Спис. Българск. Акад. Наук., XXV, 1923 : 109 (Черное море у берегов Болгарии); Косякина, Тр. Новоросс. биол. ст., II, 2, 1938 : 26 (нерест в районе Новороссийска); Пчелина, Тр. Новоросс. биол. ст., II, 3, 1940 : 71 (мальки в районе Новороссийской бухты); Виноградов, Тр. Карадагск. биол. ст., 4, 1931 : 142 (мальки у Карадага).
I D 1, II D 54—58, А 44—49, Р 17—19, V (5) 6 (По Collett'y (1892), II D 55—56, A 47—49, Р 17 (16), V 6. Lütken указывает для V 6 (7).), sp. br. 7—10, арр. pyl. 13—14 (По Day, до 20.), vert. 48—49. Жаберные тычинки в виде бугорков с небольшими шипиками на вершине. Плавательный пузырь есть, небольшой и с тонкими стенками (Указания на отсутствие его неверны.). Голова небольшая, 5.0—5.5 раз (18.0—19.6%) в длине тела. Глаза небольшие, 6.1—7.7 раз (13.0—16.4%) в длине головы, меньше длины луча 1-го спинного плавника и длины рыла. В процентах к длине тела: антеанальное расстояние 41.4—46.7, антедорсальное расстояние 16.8—17.9, длина грудного плавника 11.0—13.5, длина брюшного плавника 9.2—13.5, длина основания 1-го спинного плавника 10.5—12.4, основания 2-го спинного 55.2—58.0, длина основания анального плавника 39.5—43.5, длина луча 1-го спинного плавника 4.1—5.8, высота хвостового стебля 4.7—5.7, длина хвостового стебля 4.2—5.8. В процентах к длине головы: длина рыла 23.2—26.3, длина верхней челюсти 41.3—46.0, длина нижней челюсти 50.7—56.6, межглазничный промежуток 9.1—12.8 (только твердая костная часть его; с кожистой складкой — 18.5—19.2). Окраска в зависимости от места обитания сильно вариирует. Спина чаще всего коричневая, более или менее темная; бока и брюхо светлее. Голова, все тело и плавники с неправильными, округлой формы многочисленными пятнами и пятнышками. В пелагической постларвальной стадии спина голубая, бока и брюшко серебристые. Длина до 50 см.

Распространение. Наиболее обычен у берегов Великобритании и Ирландии, юго-западных берегов Европы, в Средиземном и Черном морях. На север вдоль берегов Норвегии до Олезунда, на восток до северной части Каттегата. Экспедицией «Travailleur» и «Talisman» найден у берегов Марокко и Судана.
Биология. Живет обычно на небольшой глубине у берегов на каменистых грунтах с водной растительностью. Реже встречается и на больших глубинах и на других грунтах. Нерест, повидимому, растянут: в Черном море у Новороссийска происходит с сентября по март, массовое количество икринок обнаружено в октябре, ноябре и декабре, в 1934 и 1935 гг. также и в марте; пелагические личинки длиной 40—53 мм пойманы в мае, личинки длиной 2.1 мм пойманы в ноябре 1934 г. перед входом в Новороссийскую бухту, личинка длиной 3 мм — в конце марта 1938 г. в средней части бухты в придонном слое. В районе Новороссийска малек длиной 53 мм пойман в открытом море на глубине 60 м в июне 1933 г. В Галацкой бухточке малек длиной 40 мм пойман в марте 1938 г., 2 малька длиной 49 мм — в мае того же года.
Хозяйственного значения почти не имеет.
Сравнительные заметки. Описание дано по экземплярам из Черного моря. Кроме того, просмотрены экземпляры из Средиземного моря и Атлантического океана (Палермо, Ницца, Мадейра). Некоторые из приведенных в синонимах видов описаны на основании различий только в окраске. Различия в размерах зубов, указываемые для некоторых видов, обусловлены возрастной изменчивостью. Об отношении к близкому Gaidropsarus vulgaris см. сравнительные заметки к роду. Motella tricirrata Nilsson следует считать синонимом этого вида, а не Gaidropsarus vulgaris, так как Nilsson приводит его по Bloch'y, дающему формулу лучей в плавниках, ближе подходящую для Gaidropsarus mediterraneus. Сюда же следует отнести Gadus argenteolus Montegu (Р 16—18, juv.). Формула лучей в плавниках у Onos tricirratus Smitt [II D (55) 58—60 (64), A (45) 47—49 (54), P 19—21] целиком охватывает пределы колебаний их у Gaidropsarus mediterraneus и Gaidropsarus vulgaris, так как в нее включены количества лучей Motella vulgaris Lütken [= Gaidropsarus vulgaris (Yarr.)] и Motella tricirrata Day [= Gaidropsarus mediterraneus (L.)]. Steindachner объединяет оба эти вида в один.
Как видно из определительной таблицы, взятой без изменений из Collett (1892) и De Buen (l. c.), различия между видами этого рода основываются на количестве лучей в грудном и брюшном плавниках и в соотношениях в размерах глаз, межглазничного промежутка, головы и луча 1-го спинного плавника. Между тем, если принять во внимание количество лучей в спинном и анальном плавниках, то различия между двумя наиболее распространенными видами этого рода — Gaidropsarus mediterraneus и Gaidropsarus vulgaris — будут несколько иными. Ниже приведены количества лучей в спинном (D), анальном (А), грудном (Р) и брюшном (V) плавниках у обоих этих видов по нашим данным и данным ряда других авторов (По экземплярам из Черного моря; такое же количество лучей имеют просмотренные экземпляры и из Средиземного моря и Атлантического океана. При подсчете лучей в грудном плавнике, как и в остальных плавниках, кожные покровы удалялись.):

Gaidropsarus reinhardti (Collett) очень близок к Gaidropsarus mediterraneus и Gaidropsarus vulgaris, отличаясь от первого несколько большим количеством лучей в грудном плавнике, от второго — меньшим количеством лучей во II D и в А, как это видно из нижеследующего:
