Anarhichas minor Olafsen, 1772
Пятнистая зубатка, пестрая зубатка, баренцевоморская пятнистая зубатка, баренцевоморская пестрая зубатка (Rus),
Spotted wolffish, Spotted sea-cat
(Eng)
Синонимы:
Anarhichas karrak Bonnaterre, 1788
Anarhichas leopardus Agassiz, 1831
Anarhichas maculatus Bloch & Schneider, 1801
Anarhichas minor Olafsen, 1772
Anarhichas scansor Bloch & Schneider, 1801
Anarhichas steenstrupii Gill, 1861
Anarrhichas egerti Steenstrup, 1842
Anarrhichas pantherinus Zuiew, 1784
Lycichthys minor Jordan, Evermann & Clark, 1930
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Пятнистая зубатка (Anarhichas minor) во многих отношениях занимает
промежуточное положение между полосатой
и синей зубатками.
Бугорковидные зубы у нее развиты не
так сильно, как у полосатой зубатки, и
сошниковый ряд зубов не выдается назад
за нёбные ряды.
Пятнистая зубатка крупнее полосатой,
но, должно быть, мельче синей; достигает
длины 144 см при весе более 30 кг.
Только у мальков пятнистой зубатки на
теле хорошо заметны поперечные полосы,
широкие и черные. После перехода к жизни у дна они разбиваются на отдельные
пятна; эти пятна хорошо обособлены друг
от друга, а в полосы группируются далеко
не так ясно, как у полосатой зубатки.
Пятнистая зубатка обычно ловится в
тех же районах, что и полосатая, но в
южных районах реже, а в северных,
напротив, чаще. У побережья Норвегии ее
можно встретить лишь севернее Бергена,
в Северное море она заходит очень редко,
а в Балтийском не появляется совсем.
У берегов Исландии в среднем ловят
примерно одну пятнистую зубатку на 20
полосатых. У атлантических берегов
Северной Америки уже вблизи залива
Массачусетс пятнистая зубатка очень редка;
зато на север вдоль западного побережья
Гренландии она идет гораздо дальше (до
Туле), чем полосатая зубатка (до залива
Диско). Обитает она, как и другие виды
зубаток, в основном на материковой
отмели, но так близко к берегам, как
полосатая зубатка, не подходит и зарослей
водорослей избегает, предпочитая в средцем
большие глубины (до 550 м). Как и все
другие зубатки, летом держится в общем
на меньших глубинах, чем зимой;
передвигается при этом на несколько большие
расстояния, чем полосатая зубатка.
Самки пятнистой зубатки выметывают
летом 12—50 тыс. икринок (при длине рыб
87—120 см), сходных по размерам с
икринками полосатой зубатки и тоже
слипающихся в кладку. Однако эти кладки
находят дальше от берегов, на
большей глубине (свыше 100 м), чем
кладки полосатой зубатки, дальше от берегов
держится и вылупившаяся молодь; у
самой поверхности моря она встречается
реже. К жизни у дна мальки переходят при
сходных размерах, но все же вначале
поднимаются выше над дном, чем мальки
полосатой зубатки; таким образом,
переход к жизни у дна происходит не столь
быстро и резко.
Пища пятнистой зубатки в общем
похожа на пищу полосатой, но включает
меньше моллюсков, а иглокожих — офиур,
морских звезд и морских ежей — заметно
больше.
Ловят пятнистую зубатку в основном
в Баренцевом море и у Гренландии. По
вкусу мяса пятнистая зубатка не
уступает полосатой или даже превосходит ее.
Шкура пятнистой зубатки чаще
употребляется на всякого рода поделки — верха
для легкой обуви, сумочки, переплеты для
книг и т. д. Это древнее занятие (еще в
XVIII в. в Гренландии из шкур зубаток
делали сумочки для сбора ягод) ныне
угасает со снижением стоимости и ростом
качества синтетических материалов.
Фауна СССР. Рыбы. Том V, вып. 5. Семейство Зубаток (Anarhichadidae). В.В.Барсуков 1959 г.
3. Anarhichas minor Olafsen — Пятнистая или пестрая зубатка.
(Minor — меньший; названа так потому, что была впервые описана по молодым особям небольших размеров (Olafsen, 1772 : 592; Fabricius, 1780 : 139).)
Flekstenbid (норвежцы); fläckstenbit, tigerfläckade hafkatten, fläckig havkatt (шведы); tigerplettet havkat (датчане); hlyri, steinbitsbrödir (исландцы); gefleckter Seawolf, gefleckter Katfisch, Tiger-Katze (немцы); spotted sea-cat (англичане), meripantteri (финны), kerak (гренландцы).
Anarrhichas minor Olafsen, Reise giennem Island, 1, 1772 : 592, pl. XLII (Исландия; цит. по немецкому переводу, 1774 : 314—315). — Richardson, Fauna Воr.-Amer., III, 1836 : 95—96 (Гренландия). — Steenstrup, Vidensk. Meddel. Naturh. Foren., (1876), 1877 : 161, 200, pl. III, fig. 2 (череп, зубы, сравнение с Anarhichas lupus). — Jordan a. Gilbert, Synops. fish. N. Amer., 1882 : 781. — Lilljeborg, Sverig. Norg. Fiskar, I, 1891 : 540 (описание, синонимия). — Young а. Сlarke, Ann. Scottish Nat. Hist., 1, 1892 : 26—29, pl. 1 (Эбердин; длина 124 см, вес 10.6 кг; фотогр.). — Smitt, Scand. fish., I, 1893 : 236, pl. XIII, fig. 1 (цветн. рис.). — Collett, Rep. Norw, Fish. Mar. Invest., (1909), II, 2, 1905 : 113—114 (западнее Фарерских островов на глуб. до 456 м; о. Медвежий, окраска молоди длиной 85—145 мм). — Книпович, Определитель, 1926 : 94, fig. 59 (Баренцево море; длина до 134.5 см.). — Lühmann, Verhandl. Anat. Ges., Ergänzungsheft zum 97 Band, 1951 : 241—242 (смена зубов). — Lühmann, Zeitschr. Zellforsch. mikrosk. Anat., 40, 5, 1954 : 470—509 (смена зубов, гистология). — Kampfe u. Kittel, Wiss. Zeitschr. Martin-Luther Univ. Halle—Wittenberg, II, 8, mathem.-naturwiss. Reihe 4, 1952—1953 Abb. 5, 8 (фотогр.). — Lühmann, Ber. Deutsch. wiss. Komm. Meeresforsch., XIII, 4, 1954 : 310—326, Abb. 1, 4, 5, 10, 12 (окраска, череп, жевательные мышцы, зубной аппарат, время нереста и смены зубов).
Anarhichas minor Müller, Zool. Dan., 1776 : 40 (Исландия). — Fabricius, Fauna Groenl., 1780 : 139—140 (Гренландия). — Jordan a. Evermann, Fish. N. a. M. Amer., III, 1898 : 2 446 (синонимия). — Pietschmann, Ann. К. K. Naturh. Hofmus., XXII, 1907—1908 : 305—306 (Баренцево море; измерения, окраска). — Вrеdеr, Field-book mar. fish. Labr. Texas, 1929 : 275, 276 (опред. табл.) — Saemundsson, Zool. Iceland, IV, 72, 1949 : 31—34 (Исландия; синонимия, экология). — Андрияшев, Рыбы сев. морей СССР, 1954 : 222—224, фиг. 111 (2), 113
(синонимия). — Барсуков, Докл. АН СССР, XCV, 4, 1954 : 897—899 (смена зубов). — Walters, Bull. Amer. Mus. Nat. Hist., 106, 5, 1955 : 306 (боковая линия). — Mакушок. Тр. Зоолог. инст. АН СССР, XXV, 1958 : 122, рис. 22, А (скелет хвостового плавника).
Anarrhichas pantherinus Zоuiеw, Acta Acad. Sci. Petrop., I, (1781), 1784 : 271— 272, 289, 230, 235, 292—293, pl. VI (неправильно указывается для Белого моря: описание и рисунок по чучелу пятнистой зубатки, сделанному Лепехиным). — Gmelin, Linn. Syst. Nat., ed. XIII, I, 3, 1788 : 1144 (sec. Zouiew). — Esmark, Forh. Skand. Naturf., 10, (1868), 1869 : 524—525 (Берген). — Lütken, Revised cat. fish. Greenl., 1875 : 119 (синонимия). — Collett, Norg. Fiske, 1875 : 70 (Берген, длина 126 см).
Anarhichas karrak Bonnaterre, Tabl. Encycl., Ichthyol., 1788 : 38—39 (Гренландия).
Anarhichas karrak + Anarhichas pantherinus Lacepède, Hist. nat. poiss., IV, 1801 : 44—47 (sec. Zouiew a. Bonnaterre).
Anarhichas maculatus Вlосh, fed. Schneider, Syst. Ichth., 1801 : 496.
Anarhichas pantherinus Pallas, Zoogr. Rosso-Asiat., III, 1811 : 76 (сев. побережье России, по чучелу, сделанному Лепехиным).
Anarrhichas leopardus Agassiz in Spix, Selecta Gen. Species Pise. Bras., 1829 : 92—93, 51 (описан по чучелу из сборов Спикса). — Cuvier et Valenciennes, Hist. nat. poiss., XI, 1836 : 493 (отрицают принадлежность к сем. зубаток). — Steenstrup, Vidensk. Meddel. Naturh. Foren., 1877—1878 : 109—114 (идентичность с Anarhichas minor).
Anarhicas lupus (non Linne) Faber, Naturgesch. Fisch. Islands, 1829 : 70—75 (частью; связь строения с питанием).
Anarrhichas lupus (non Linne) Cuvier et Valenciennes, Hist. nat. poiss., XI, 1836 : 473 (частью; критическое обозрение предыдущих работ). — Günther, Cat. fish., III, 1861 : 208—211 (частью: «american variety»).
Anarrhichas Egerti Steenstrup, Forh. Scand. Naturf., III, 1842 : 647 (nomen nudum).
Anarrhicas Steenstrupii Gill, Catal. fish. N. Amer., 1861 : 46 (nomen nudum).
Lycichthys minor Jordan, Evermann a. Clark, Check list fish. N. a. M. Amer., 1930 : 472 (сев. часть Атлантического океана, оба побережья).
Распространение, биология. Good a. Bean, Bull. Essex. Inst., 11, 1879: 11 (Истпорт, Глостер). — Collett, Christ. Vidensk. Selsk. Forhandl., (1879), I, 1880 : 45—46 (м. Нордкап, Порсангер-фиорд, молодь в желудках трески и пикши). — Gооd, Fish. a. Fish. Industr. U. S., I, 1884 : 249 (залив Фёнди). — Collett, , Nyt Mag. Naturvidensk., (1879), 29, 2, 1885 : 66 (обильна у м. Нордкап). — Günther, Rep. sci. resalts voyage Challenger, XXII, 1887 : 70 (43°52' N, 59° W). — Книпович, Ежег. Зоолог. муз. Акад. наук, II, 1897 : 148 (ошибочно указывается частой в Белом море). — Ehrenbaum, Fauna Arctica, Fische, II, 1901 : 95—96 (Шпицберген, о. Медвежий). — Книпович, Эксп. научно-пром. исслед. Мурмана, I, 1902 : 55 и сл., фиг. 3, 4, 54 (многочисленные местонахождения у берегов Мурмана; длина до 134.5 см, фотогр; мальки у о. Медвежьего). — Брейтфус, там же, II, 1903. — Книпович, там же, II, 1, 1904 (местонахождения в Баренцевом море). — Ehrenbaum, Wiss. Meeresunters., Helgol., VII, 1, 1906 : 54 (от о. Медвежьего до южной оконечности Шпицбергена; лучи в плавниках). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1906; II, 1908 (местонахождения в Баренцевом море). — Atkinsоn, Journ. Mar. Biol. Ass., VIII, 2, 1908 : 96 (питание). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1912; II, 1915 (местонахождения в Баренцевом море). — Thielemann, Wiss. Meeresunters., Helgol., XIII, 2, 1921 : 206, Karte 5 (экология, питание, распространение в южной части Баренцева моря). — Wollebaek, Norg. fisk., 1924 : 210 (сев. побережье Норвегии, летом во множестве на глуб. 200—400 м). — Bigelow a. Wеlsсh, Fish. Gulf Maine, 1925 : 375, fig. 189 (на юг до залива Массачусетс). — Jenkins, Fish. Brit. Island, 1925 : 110 (Эбердин в зап. Шотландии). — Jеnsеn, Rap. Proc.-Verb., XXXIX, 1926 : 99 (Дэвисов пролив). — Saemundsson, Synops. Fish Iceland, 1927 : 18 (Исландия, на глуб. 25—400 м). — Dunker u. Моhr, Tierwelt N. Ostsee, XII, 1928 : 91—92, fig. 3 (к западу от Хаугесунда, 59°20' с. ш., апрель 1908 г.). — Броцкая, Докл. 1-й сессии Гос. океаногр. инст., 4, 1931 : 11, 13, 50—51, рис. 6, 28, 1 (питание). — Lundbeck, Wiss. Meeresunters., Helgol., XVIII, 3, 1932 : 13—16, 18, 21—22. — Смapагдова, Тр. Гос. океаногр. инст., III, 2, 1933 : 99—123 (уловы в южной части Баренцева моря). — Merriman, Copeia, 3, 1935 : 143 (Ново-Шотландские банки; фотогр.). — Rass, Int. Rev. gesamt. Hydrobiol. Hydrogr., XXXIII, 3—4, 1936 : 262—263, fig. 3 (икра и личинки зубаток в Баренцевом море, Anarhichas lupus и Anarhichas minor?). — Еhrеnbaum, Handb. Seefischer. N. Europas, II, 1936 : 140 (промысел). — Lundbeck, Ber. Deutsch. wiss. Komm. Meeresforsch., N. F., VIII, 2, 1936 : 100—103, tab. 4, 19 (уловы в разных частях ареала; вес, длина и возраст). — Bigelow a. Schroeder, Bull. Bureau Fisher., XLVIII, 20, 1936 : 337 (в заливе Мэн редка). — Есипов, Пром. рыбы Баренц. моря, 1937 : 51, фиг. 6 (Баренцево море на восток до Гусиной банки). — McKenzie a. Homans, Proc. Nova Scotian inst. sci., XIX (1936—1937), 3, 1938 : 279 (Ново-Шотландские банки). — Nybelin, in: Andersson, Fiskar Fiske Norden, I, 1942 : 104 (до Бергена, на глуб. 200—400 м). — Маслов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., VIII, 1944 : 156 (Баренцево море; темп роста, плодовитость, промысел). — Jensen, Spolia Zool., Mus. Haun., IX, 1948 : 118—121 (многочисленные местонахождения у западного побережья Гренландии; питание). — Есипов в изд.: Пром. рыбы СССР, 1949 : 614, атлас, табл. 188 (сводка, цветн. рис.). — Травин, в сб.: Пром. рыбы Баренц. и Бел. морей, 1952 : 174—175 (сводка, данные по распространению и биологии). — Kämpfe u. Kittel, Urania, XI, 11, 1952 : 416—420, fig. 9—11 (фотогр.). — Bigelow a. Schroeder, Fish. Gulf Maine, 1953 : 507—508 (Ново-Шотландские банки; в заливе Мэн очень редка; для залива Св. Лаврентия, Нью-Фаундлендских банок и внешнего Лабрадора неизвестна). — Полянский, Тр. Зоолог. инст. АН СССР, XIX, 1955 : 50, 59, 61, 63, 64, 70, 78, 88, 99, 106, 116 (паразиты). — Østvedt, Ann. Biol., Cons. perm, intern. l'Expl. Mer, (1954), 11, 1956 : 29 (результаты мечения в Баренцевом море). — Константинов, Тр. Полярн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., X, 1957 : 85—86 (результаты мечения в Баренцевом море). — Барсуков, Вопр. ихт., 8, 1957 : 34—41, рис. 3—8, 11, 2, 12, 2, 4, 5, 13, 2 (распределение в Баренцевом море). — Kotthaus u. Krefft, Ber. Deutsch. wiss. Komm. Meeresforsch., XIV, 3, 1957 : 179 (места поимки у побережья Гренландии и Исландии).
D 74—78 (80), А 44—48, Р 20—23 (24), С 21—22, позвонков 76—79 (туловищных 27—29, хвостовых 48—51), лучей жаберной перепонки (6) 7, окологлазничных косточек (с praeorbitale) 6—7. Наибольшая высота тела у взрослых 3.8—4.9 раза в его длине, у мальков длиной 10—19 см 5.2—6.6 раза. Антеанальное расстояние взрослых 47.1—54.0% длины тела, у мальков 44.1—48.9%. Антедорсальное расстояние у взрослых 15.3—18.6%, у мальков 18.9—21.5%. Длина головы от конца рыла до заднего угла operculum у взрослых 17.4—20.9%, у мальков 19.7—23.8%. Расстояние от начала спинного плавника до верхнего края жаберной щели от 1/3 до 1/2 длины головы (у взрослых 41.2—50.0%, у мальков 33.4—43.2%). Ширина основания грудного плавника от 40 до 60% высоты тела у начала анального плавника. Хвостовой плавник более или менее усеченный. Нижняя поверхность верхней губы перед клыками покрыта неправильными складками, ее толщина здесь почти равна толщине нижней губы. Сошниковый ряд зубов длиннее нёбного ряда или равен ему; простирается назад почти также далеко, как и нёбные ряды или (очень редко) слегка заходит за них. У мальков длиной 7—10 см зубы на нёбных костях обычно мелкие, сидят разбросанно в один ряд, продолжающийся назад несколько далее зубов на сошнике, очень высоких и расположенных обычно в два неправильных ряда. Длина сошникового ряда зубов составляет у взрослых 1.0—1.2 длины нёбного ряда, у мальков 1.1—1.8. Зубы внешнего ряда на нижней челюсти (за исключением клыков) и зубы на нёбных костях остроконические или со слабо стертыми вершинами. Внешний ряд зубов на нижней челюсти не образует перерыва в области пережима нижнечелюстной кости. Жевательные мышцы у взрослых составляют 7.8—8.7% веса головы с плечевым поясом и 2.2—3.0% веса тела. Сухожилие, идущее от переднего края порции А1 к передней части maxillare, имеется. Frontale позади заглазничных отростков не сужена в виде киля, ширина ее здесь у взрослых несколько меньше ширины между орбитами. Sphenoticum не опущено (см. Anarhichas latifrons). Верхний край гребней на sphenoticum расположен примерно на вертикали переднего края parietalia. Крыша черепа довольно плоская (табл. XVII, 2). Задний угол глазницы находится примерно на вертикали заглазничных отростков frontale. Бугорки на opistoticum не сдвинуты или лишь слегка сдвинуты вверх. Кости прочные. Обонятельные нервы тоньше зрительных. Селезенка округлая. В мышцах содержится около 80% воды.
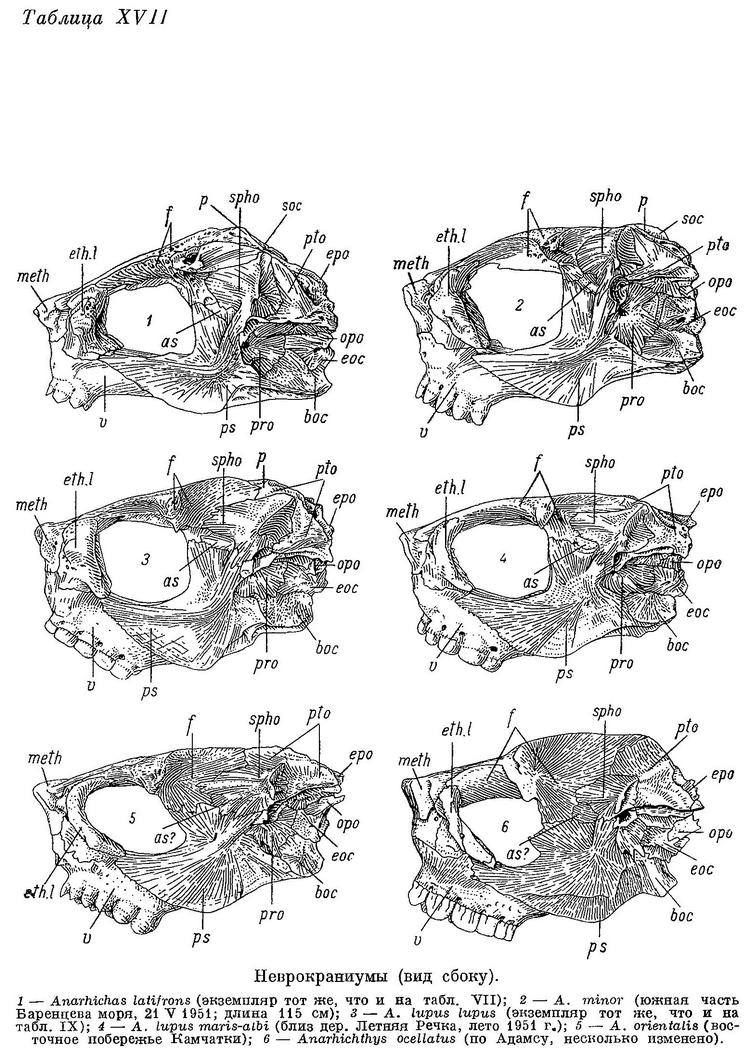
Общий фон окраски от желтоватого до серовато-коричневого или даже темно-коричневого. Верх головы до переднего края глаз, щеки, спинной плавник, спина и бока покрыты многочисленными округлыми коричневато-черными четкими пятнами, иногда неясно группирующимися в поперечные полосы. Многие из пятен превышают диаметр зрачка. Мальки имеют на боках тела 7—11 темных широких поперечных полос, которые в дальнейшем разбиваются на пятна. Количество пятен с возрастом увеличивается.
Описание дано по 200 экз. из Баренцева моря; позвонки и лучи в плавниках подсчитаны у 22—34 экз., череп описан по 20 экз., жевательные мышцы — по 4 экз.
Длина особей, добытых в Баренцевом море, до 135 см (Книпович, 1902; материалы ПИНРО), на Ново-Шотландских банках до 134 см (McKenzie a. Homans, 1938 : 279), у западного побережья Гренландии до 122 см (по 67 экз.; Jensen, 1948 : 118—120). Есть указания на длину до 180—200 см (Esmark, 1869 : 524; Saemundsson, 1943 : 33, и др.), основанные, по-видимому, на неточных данных. В Баренцевом море обычна в траловых уловах длиной от 20 (а в южной части от 60) до 120 см; преобладающие размеры 90—110 см при весе 7—17 кг. Средняя длина особей в южной части Баренцева моря 90 см, на юго-западном углу Центральной возвышенности — 76 см, у о. Медвежьего — 67 см, у западного побережья Шпицбергена — 56 см (1946—1951 гг.). В 1928—1931 гг. средняя длина в Баренцевом море 98.8 см; у о. Медвежьего 97.6 см; у берегов Исландии — 80.3 см (преобладающие размеры 70—90 см, — Lundbeck, 1936, tab. 19); средний вес, соответственно, — 7.8, 7.0 и 3.4 кг. В северо-западной части Баренцева моря (сентябрь 1950 г.) средняя длина самцов 78.2 см (56 экз.), самок 71.1 см (46 экз.); в южной части (май 1951 г.) — самцов 102 см (17 экз.), самок 91 см (12 экз.); максимальный размер самцов 122 см, самок 124 см.
Распространение. Северная часть Атлантического океана (рис. 36). В Баренцевом море на север до возвышенности Персея, на северо-восток и восток до губы Крестовой на Новой Земле и района о. Вайгач (добыта на 70°05' с. ш. и 65°15' в. д.). Входит в Кольский и Мотовской заливы. В Белом море не встречается. Зуев (Zouiew, 1784 : 271), впервые описавший пятнистую зубатку из Белого моря, ссылается на экземпляр, находившийся в таком состоянии, что сделать описание по нему было невозможно; описание было дано по экземпляру, добытому Лепехиным, очевидно, в Баренцевом море (Pallas, 1811 : 76—77). Кроме того, имеется указание Левандера (Levander, 1888—1889 : 231; без описания) на два экземпляра Anarhichas minor, добытых в районе Соловецких островов. В этом районе в 1951 г. мной было исследовано около 2000 зубаток; среди них не оказалось ни одной Anarhichas minor. Местные рыбаки, промышляющие и в Баренцевом море, а поэтому прекрасно различающие Anarhichas lupus и Anarhichas minor, единодушно утверждают, что последний вид, пятнистая зубатка, в Белом море не встречается. В Гренландском и Норвежском морях распространена по крайней мере от о. Принца Карла (западное побережье Шпицбергена) на севере до Бергена на юге. Вдоль норвежских берегов изредка встречается еще южнее (у Хаугесунда). У северного и восточного побережий Шпицбергена и в Стурфиорде не обнаружена (Книпович, 1903 : 153, 155). Фарерские и Шетландские острова, подводный хребет между Фарерскими островами и Исландией. Однажды найдена у Эбердина (Шотландия; Young a. Clark, 1892 : 26—29). Обычна вокруг Исландии, но реже встречается на ее южном и западном побережьях (Lundbeck, 1936 : 102; Saemundsson, 1949 : 32). На западном побережье Гренландии отмечена к северу пока что до фиорда Упернавик, на восточном — до фиорда Ангмагссалик. Дэвисов и Датский проливы (Jensen, 1948 : 117; Kotthaus u. Kreflt, 1957 : 179). У американских берегов в районе Ново-Шотландских банок и на юг до залива Массачусетс (очень редко; редка и в заливе Мэн).
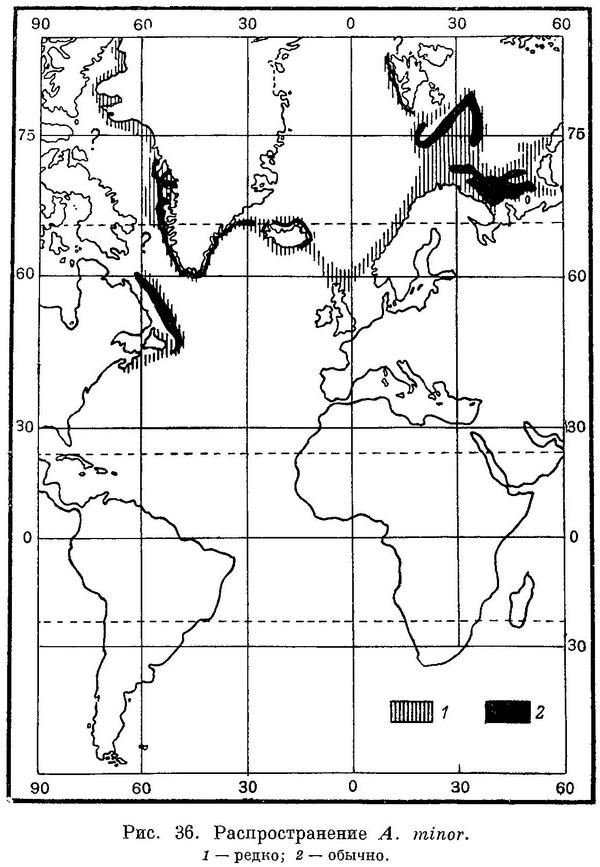
Биология. В Баренцевом море встречается на глубинах от 51 до 475 м, у побережья Исландии от 25 до 478 (500) м, у побережья Гренландии от 25 до 450 м, у побережья Северной Америки от 63 до 550 м. В Баренцевом море и у берегов Исландии (Saemundsson, 1949 : 32) чаще всего вылавливается на глубинах 100—200 м (на западном склоне Медвежинской банки иногда образует скопления на глубинах 300—400 м), у берегов Гренландии — на 50—250 м (Jensen, 1948 : 118—120), у берегов Норвегии — на 200—400 м (Wollebaek, 1924 : 210). В Баренцевом море на наибольших глубинах держится в середине зимы, затем вплоть до начала следующей зимы выходит на меньшие глубины. Молодь придерживается несколько больших глубин, чем взрослые особи (Барсуков, 1957 : 38); очевидно, и в других частях ареала ведет себя сходно. Так, у западного побережья Гренландии молодь длиной 37—79 см вылавливалась на глубинах 68—398 м, а взрослые особи длиной 80—122 см — на глубинах 25—288 м; наибольший улов молоди наблюдался на глубине 225—315 м, взрослых — на глубине 126 м (Jensen, 1948 : 118—120). В июне-октябре 1955 г. молодь у побережья Гренландии ловилась тралом также в общем на больших глубинах, чем взрослые особи (Kotthaus u. Krefft, 1957 : 179).
Встречается на илистых и илисто-песчаных грунтах, причем уловы часто совпадают с попаданием в трал камней. ЭНПИМ в 1898—1905 гг. не встретила Anarhichas minor, как и Anarhichas latifrons, при солености менее 34.51‰. Однако, судя по тому, что пятнистая зубатка у Мурманского побережья концентрируется более близко к берегам, чем Anarhichas latifrons, и, в отличие от последней, заходит в Мотовской и Кольский заливы, можно полагать, что требовательность ее к солености несколько ниже.
Anarhichas minor встречается в Баренцевом море при температурах от —1.32 до +5°. Наибольшие концентрации ее здесь наблюдаются при температуре от —1 до 0° летом и от 0 до +2° зимой. Взрослые особи весной и летом, кроме упомянутых выше концентраций при отрицательных температурах, образуют также скопления при положительных температурах — от 2 до 4°, резче выраженные в северо-западной части моря, чем в южной, и наблюдающиеся у мелких особей позднее, чем у крупных. По-видимому, эти скопления связаны с периодом созревания половых продуктов (Барсуков, 1957 : 41). У побережья Гренландии в июне-сентябре 1955 г. ловилась при температурах от 1 до 5.9°; у побережья Исландии в июне-июле — от 1.1 до 6.8° (Kotthaus u. Krefft, 1957 : 179).
Миграции, места и сроки нереста точно неизвестны. Материалы по распределению Anarhichas minor в Баренцевом море приводят к выводу, что взрослые особи, скапливающиеся зимой в южной части моря, мигрируют летом частью на северо-запад для нереста, частью на восток для нагула; передвижения же молоди летом на север, а зимой на юг, как правило, не выходят за пределы северо-западной части моря (Барсуков, 1957 : 39). Мечение пятнистой зубатки, пока еще немногочисленное, как будто подтверждает вывод о ее миграциях, сделанный на основании распределения уловов. В апреле-октябре зубатки двигаются на восток, по струе Мурманского теплого течения; в апреле-июне часть зубаток идет на запад (Константинов, 1957 : 85—86, рис. 5). Весенне-летние миграции меченых пятнистых зубаток из восточных частей Баренцева моря на банки Финмаркена обнаружены также Остведтом (Ostvedt, 1956 : 29), отмечающим нерестовый характер этих миграций. Остведт считает, что нерест происходит в июне-июле, что вполне вероятно; но принятие банок Финмаркена за места нереста сомнительно (см. ниже). Скорее всего, на банках Финмаркена ловятся не нерестящиеся особи, а продвигающиеся к местам нереста. Следует, однако, заметить, что большое число меченых зубаток было выловлено вблизи мест выпуска по истечении очень продолжительного срока (до 2 лет с лишним). Надо полагать, лишь небольшая часть этих особей могла оказаться вблизи мест выпуска уже после совершения кормовых либо нерестовых миграций, основная же часть не совершала значительных передвижений. Имеются, очевидно, индивидуальные различия в протяженности и сроках миграций, перемежаемых определенными периодами оседлости. Эти периоды, возможно, более характерны для сравнительно молодых экземпляров; к сожалению, длина выловленных меченых зубаток не указывается. Кроме того, тенденция к оседлости могла значительно усилиться в результате травмы, нанесенной при поимке в трал и при мечении. Из ярусных уловов было помечено 156 зубаток, из них выловлено вторично только 5; малый возврат особей объясняется повышенной смертностью зубаток в результате повреждений, нанесенных крючком (Ostvedt, 1956 : 29).
Самки баренцевоморской Anarhichas minor, по Милинскому (1944), созревают при длине от 60 см и выше, основная масса — при длине 80—105 см. Диаметр зрелых икринок, по Рассу (1941 : 31), 5.5—6.0 мм. Плодовитость 12 баренцевоморских самок колебалась от 12 430 икринок у самки длиной 87 см до 50 000 икринок у самки длиной 120 см (по данным Сахно, см.: Маслов, 1944 : 156).
В работах ряда авторов (Книпович, 1902 : 109, 111, 141, 428; Брейтфус, 1903 : 63, 1906 : 96; Pietschmann, 1907-1908 : 306; Thielemann, 1921, tabl. L—XCVII) отмечено нахождение в Баренцевом море «созревающих», «почти зрелых» и «зрелых» самок Anarhichas minor с конца марта по конец июля. Как в этих работах, так и в станционно учетных карточках ПИНРО не приводятся ни диаметр икринок, ни отношение веса гонад к весу тела. Шкала стадий зрелости, разработанная специально для зубаток, отличающихся очень крупной икрой, до сих пор отсутствовала. Поэтому следует отнестись с осторожностью к сведениям из указанных работ и к материалам ПИНРО. Они намечают скорее всего лишь конец нереста, но не его начало. Продолжительность инкубационного периода у зубаток неизвестна. К тому же никто не указывает надежных признаков, с помощью которых можно было бы отличить кладки икры и личинок Anarhichas minor от таковых же Anarhichas lupus lupus. Поэтому сообщения ряда авторов (Schnackenbeck, 1933 :191; Перцева, 1939 : 460; Есипов, 1949 : 614, и др.) о находках кладок и личинок якобы Anarhichas minor имеют малую ценность для установления мест и сроков ее нереста.
Из 87 самок Anarhichas minor, исследованных мной, 20 самок длиной 9—49 см имели I стадию зрелости, 33 самки длиной 49—66 см — I, I—II и II стадии, 9 самок длиной 67—75 см — только II стадию, 25 самок длиной 76 см и более — II, II—III, III и VI стадии. Все самки III (9 экз.) и VI (1 экз.) стадий были добыты в конце мая, самки же, добытые в сентябре, имели только II и II—III стадии зрелости. Отношение веса гонад к весу тела у самок и самцов приведено в табл. 18.
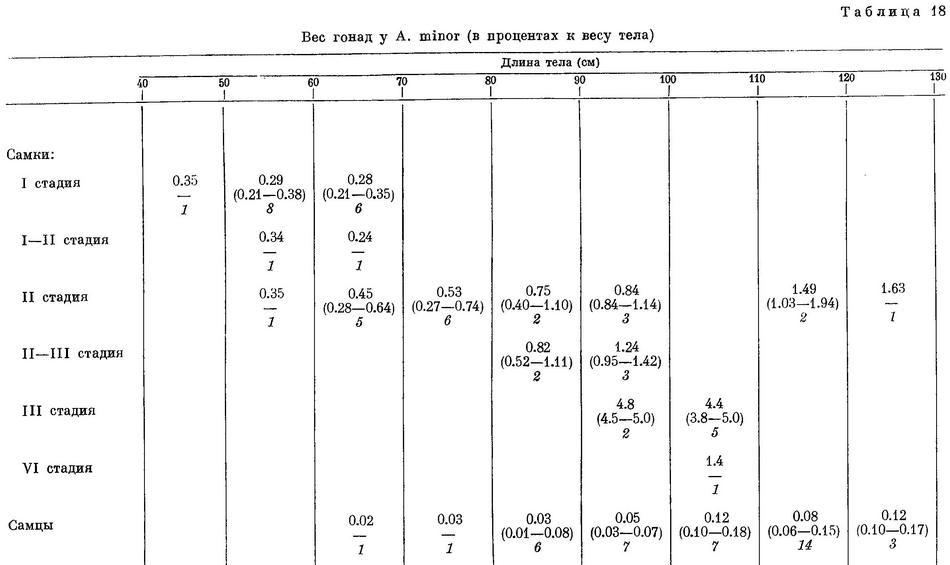
У самок III стадии, добытых в конце мая, диаметр икринок колебался от 3.0 до 3.5 мм, в среднем 3.2 мм. Anarhichas lupus maris-albi имела в среднем приблизительно такой же диаметр икринок (при несколько большем отношении веса гонад к весу тела) в конце июня, а отнерестовала в июле начале августа. По аналогии с Anarhichas lupus maris-albi можно судить, что упомянутые выше самки III стадии могли отнерестовать большей частью примерно через месяц, т. е. в конце июня — начале июля. В музее ПИНРО хранятся яичники Anarhichas minor IV стадии зрелости, близкой к вымету. Эта самка добыта в июле 1939 г; ее длина 68.5 см, место поимки не указано.
Приведенные наблюдения дают возможность полагать, что нерест Anarhichas minor происходит в основном не в марте-апреле (Есипов, 1949 : 614), а позднее, примерно в июне-июле. В мае нерестятся, по-видимому, лишь сравнительно немногие особи; притом неизвестно, не остаются ли крупные икринки в яичниках самок VI стадии еще с прошлого года. По Люману (Lühmann, 1954b : 318), у побережья Исландии Anarhichas minor нерестится также летом (к сожалению, этот вывод не сопровождается описанием состояния половых продуктов у исследованных особей, не указано и их количество).
Сеголетки Anarhichas minor размером от 7 см и более на Центральной возвышенности, у о. Медвежьего и на возвышенности Персея иногда вылавливаются обычными промысловыми тралами; в трал Петерсена здесь попадают мальки еще меньших размеров (4—7 см). Выше уже указывалось, что мальки зубаток длиной более 4.4 см в толще воды не встречались еще никем. Личинки Anarhichas minor при выходе из икринки, по всей вероятности, имеют длину около 2 см — судя по сходству с Anarhichas lupus в размере зрелых икринок. Малек в течение первого года жизни достигает 9.6—18.1 см (см. ниже), т. е. вырастает за год примерно на 8—16 см. Из сопоставления этих данных видно, насколько коротким должен быть пелагический период жизни у личинок и мальков, если он вообще существует. Не более того времени, за которое малек успеет вырасти на первые 1/4—1/8 своего годового прироста. При этом следует учесть, что годовой прирост часто определяется всего лишь несколькими месяцами нагула и что в начале нагульного периода скорость линейного роста мальков может быть значительно большей, чем в конце. Милинский (1944) предполагает, что развитие и рост личинок происходит в придонных слоях воды. Но Енсен (Jensen, 1948 : 121), напротив, считает, что молодь держится у поверхности.
Молодь Anarhichas minor длиной менее 25 см у побережья Исландии и Гренландии неизвестна (Saemundsson, 1949 : 33; Jensen, 1948 : 118—120; Kotthaus u. Krefft, 1957 : 179), а у побережья Норвегии и в южной части Баренцева моря редка (длиной менее 20 см она и здесь не встречается, если не считать одной находки в кишечниках трески и сайды у мыса Нордкап — Collett, 1885 : 66).
В Баренцевом море основная масса молоди Anarhichas minor длиной до 60—70 см держится в северо-западной части моря, от о. Медвежьего и западного побережья Шпицбергена на западе до возвышенности Персея и Центральной возвышенности на востоке (Барсуков, 1957 : 36). Очевидно, нерест баренцевоморских взрослых особей происходит именно в этих районах. Судя по местам нахождения мальков минимальных размеров (от 4 до 19 см), нерест имеет место на северных окраинах Западного желоба, от западного склона Медвежинской банки до юго-западного угла Центральной возвышенности. Кладка, найденная 26 V 1945 на Центральной возвышенности (глубина 240 м, температура +0.4°), может принадлежать пятнистой зубатке, как и кладка с развивающимися эмбрионами, обнаруженная у о. Медвежьего 23 VII 1901 (глубина 140 м, температура —0.5°, — Collett, 1905 : 113—114). Однако в марте-апреле скопления взрослых особей наблюдаются не здесь, а в южной части моря, что подтверждает высказанные выше предположения о более позднем нересте. Тем не менее значительная часть взрослых особей остается и летом в южной части моря, продвигаясь на восток: по всей вероятности, это особи, не нерестящиеся в данном году. Наблюдения ПИНРО над состоянием гонад у зубаток, к сожалению, производившиеся лишь от случая к случаю, показывают, что и в июне-июле (Мурманское мелководье и северная часть Новоземельского, 7 экз. длиной 70— 125 см — из станционно-учетных карточек ПИНРО.) и в феврале (Центральный желоб, 15 II 1948; зубатка составляла 50% уловов — из отчета за рейс № 3 РТ-65) самки Anarhichas minor имели в южной части моря лишь II или II—III стадии зрелости.
Упомянутые выше 9 самок III стадии зрелости были добыты на Рыбачьей, Финмаркенской и Нордкинской банках. Эти самки, по-видимому, продвигались к нерестилищам у о. Медвежьего вдоль северных берегов Норвегии. Как указывает Эренбаум (Ehrenbaum, 1936 : 141), Anarhichas minor у северных берегов Норвегии весной и летом в большом числе попадает на яруса, а у мыса Нордкап иногда до сотни экземпляров на один ярус (Collett, 1885 : 66).
В кишечниках Anarhichas minor в Баренцевом море найдены офиуры — Ophiura sarsi, Ophioglypha sarsii, Ophiopleura borealis, Gorgonocephalus, морские звезды — Ctenodiscus crispatus, Solaster, Asterias, морские ежи — Strongylocentrotus droebachiensis, морские лилии — Heliometra glacialis, голотурии — Psolus phantapus, двустворчатые моллюски — Pecten groenlandicus, Cardium, Cyprina islandica, Astarte, Mytilus edulis, брюхоногие моллюски — Buccinum, Trophonopsis, Natica и др., ракообразные — Hyas araneus, Eupagurus pubescens, Idotea, Pandalus borealis, пикногоны — Colossendeis, трубочки червей, мшанки, губки, актинии Hormatia nodosa, рыба — мойва, сайка, молодь трески и пикши, пинагор, камбала-ерш, бычки Icelus, сельдь, молодь морского окуня, отбросы с траулеров. Резко преобладают иглокожие. Ракообразные и моллюски встречаются реже; моллюски обычно не с очень толстой раковиной. Обычна рыба, но немногим менее обычны и отбросы с траулеров; возникает тот же вопрос, что и в отношении Anarhichas latifrons: какую часть рыбы Anarhichas minor ловит, а какую подбирает выброшенной с траулеров. Несомненно, что часть рыбы она ловит, так как попадает иногда на поддев (Брейтфус, 1915 : 350). У берегов Исландии Anarhichas minor также питается в основном иглокожими, особенно офиурами, не пренебрегая и отходами рыбы с траулеров (Saemundsson, 1949 : 33). Стенструп (Steenstrup, 1877 : 195) там же в желудках Anarhichas minor нашел немного мелких Cyprinae, средней величины Pectines и очень много морских ежей — Spatangus, офиур, морских звезд — Asterias rubens и др., крабов — Hyas araneus и раков-отшельников — Pagurus bernhardus и Pagurus pubescens. У западного побережья Гренландии в кишечниках Anarhichas minor обнаружены крабы, трубки червей, морские ежи, голотурии Psolus, морские звезды, двустворчатые и брюхоногие моллюски, водоросли и песок (Jensen, 1948 : 121).
В сентябре Anarhichas minor питается интенсивно. В это время (1950 г.) был произведен полевой анализ питания у 102 особей из района Медвежинской банки и возвышенности Персея. У всех в кишечниках была найдена пища. Вес содержимого желудка колебался от 0.1 до 21% веса тела, в среднем 4.9%, вес содержимого кишки — от 2.5 до 9.2%, в среднем 6.5%. В кишечниках Anarhichas minor, добытых в районе Медвежинской банки, преобладали морские ежи, а на возвышенности Персея — морские лилии; эти же организмы в указанных местах преобладали и в прилове трала. В конце сентября 1950 г. на 76°43'—77°01' с. ш. и 31°58'—82°58' в. д. наблюдались большие скопления мойвы. В кишечниках всех Anarhichas minor (а также трески и других рыб) встречалась здесь почти исключительно мойва, причем наполнение желудков по отношению к весу тела было наибольшим.
В мае 1951 г. было вскрыто 28 особей Anarhichas minor в районе Рыбачьей, Финмаркенской и Нордкинской банок. Интенсивность питания оказалась значительно меньшей, чем в сентябре: 10 особей (36%) имели пустые кишечники, у 10 в кишечниках были найдены лишь отбросы с траулеров, у остальных 8 особей пищи в кишечниках было мало. Вес содержимого желудка колебался от 0.1 до 6.3% веса тела, в среднем 1.6%, вес содержимого кишки — от 0.1 до 7.2%, в среднем 1.8%, причем наибольший вес в обоих случаях давали отбросы с траулеров. В сентябре эти отбросы в кишечниках зубаток встречались очень редко вследствие того, что вокруг нашего корабля не было тогда скоплений промысловых траулеров. Молодь Anarhichas minor на юго-западном углу Центральной возвышенности питалась в мае морскими лилиями, а также офиурами.
Снижение интенсивности питания в мае по сравнению с сентябрем сказалось на весе печени особей обоих полов, исключая самок III стадии зрелости (табл. 19).

Темп роста в Баренцевом море, по данным П. З. Сахно (Маслов, 1944 : 155), дополненным мной для первых лет жизни Anarhichas minor, ниже, чем у Anarhichas latifrons, но выше, нежели у Anarhichas lupus (табл. 20).
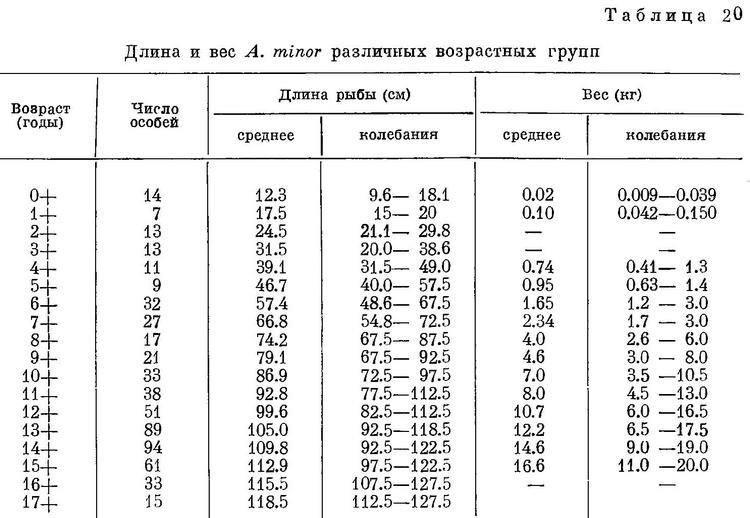
Зубатка наибольшего возраста, 21-годовалая, имела длину 122 см. По Лундбеку (Lundbeck, 1936, Tab. 19), в уловах на Баренцевом море встречаются пятнистые зубатки до 40-годовалого возраста, преимущественно от 8 до 25-годовалого, у берегов Исландии от 5 до 30-годовалого, но главным образом от 7 до 20-годовалых.
На кривых Сунда, вычерченных по данным ПИНРО за 1946—1950 гг., у Anarhichas minor, как и у Anarhichas latifrons, заметны две «волны» поколений повышенной численности (рис. 24).
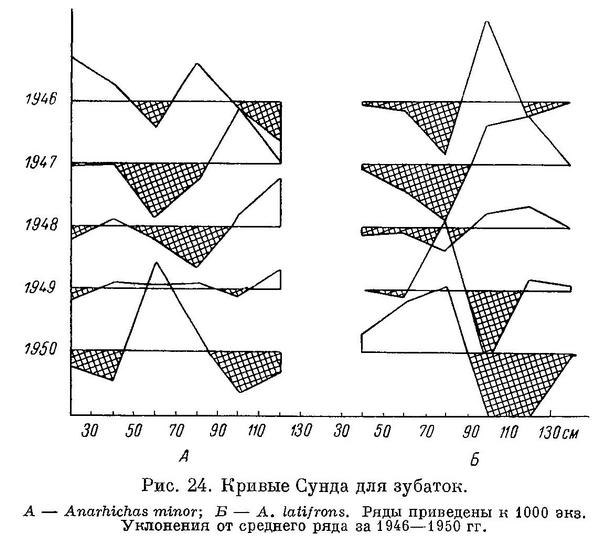
Anarhichas minor была встречена мной в кишечнике полярной акулы (Somniosus microcephalus). Выше уже было указано, что молодь Anarhichas minor была однажды найдена в кишечниках трески и сайды.
Ю. И. Полянский (1955) обнаружил у баренцевоморских Anarhichas minor (8 экз.) следующих паразитов: Podocotyle reflexa (кишечник), Neophasis lageniformis (желудок, кишечник), Steringophorus furciger (кишечник), Fellodistomum fellis (желчный пузырь), Fellodistomum agnotum (желчный пузырь, кишечник), Diphterostomum microacetabulum (кишечник), Lepidophyllum steenstrupii (мочевой пузырь), Derogenes varicus (желудок), Scolex polymorphus (кишечник), Ascarophis sp. (желудок), Platybdella anarrhichae (жабры, кожа), Clavellodes rugosa (жабры). Из 28 Anarhichas minor, вскрытых мной в мае 1951 г. в южной части Баренцева моря, по крайней мере 20 особей оказались зараженными сосальщиком Fellodistomum fellis. Цвет желчи у таких экземпляров варьировал от темно-зеленого, почти черного, до молочно-белого с желтым оттенком; у здоровых зубаток желчь обычно светло-зеленая. Значительных различий в общем весе тела, в относительном весе печени и гонад у зараженных и незаражениых особей одинаковой длины не замечено.
Хозяйственное значение в Баренцевом море больше, чем у других видов зубаток, в то время как уловы зубаток у берегов Исландии и Северной Америки преимущественно, а в Северном море исключительно состоят из полосатой зубатки. Наши уловы пятнистой зубатки достигали в довоенные годы почти 33000 ц (табл. 21). Основные уловы дает траловый промысел в южной части моря, где средний размер пятнистой зубатки наиболее высок. Наибольшие уловы на 1 час траления наблюдаются обычно в марте и апреле, наибольший процент к общему улову — в феврале.
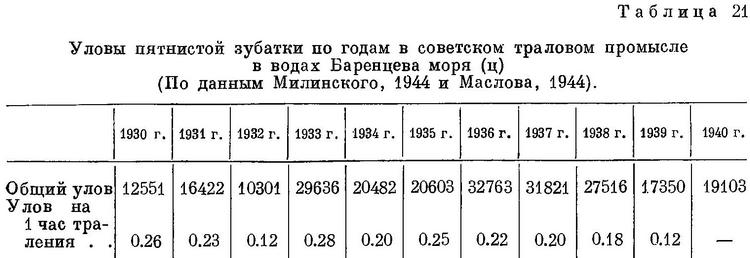
Мясо на Мурмане преимущественно засаливается, а кроме того, используется в копченом, вяленом и свежем виде. Считается, что вкусовые качества мяса пятнистой зубатки выше, чем других видов; оно содержит 76—80% влаги, 13—17% белка и 3—7% жира (Травин, 1952 : 175); по данным ПИНРО, жир в основном концентрируется в печени (23—42%) и костях (12—26%). Икра также употребляется в пищу. Шкура идет на различного рода изделия.
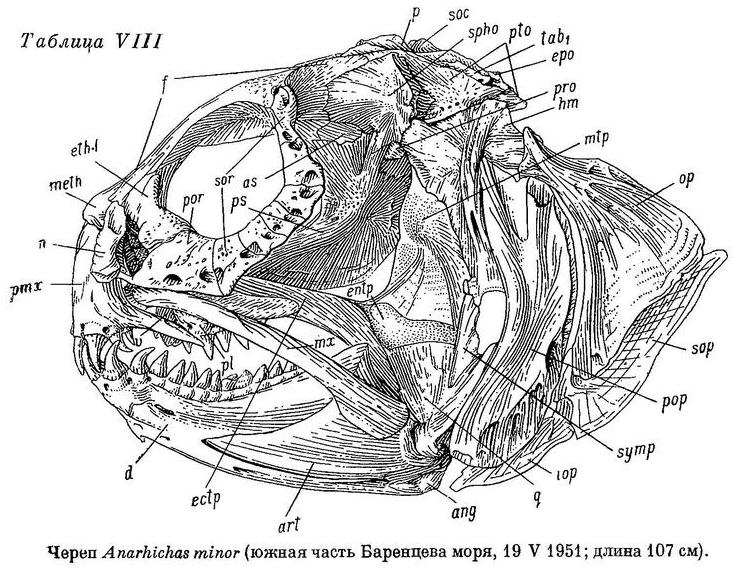
У Anarhichas minor передние клыки нижней челюсти входят в промежутки между передними клыками верхней. Клыки, как и все другие зубы, крупнее, чем у Anarhichas latifrons; сильнее стираются к концу лета; соединение их с цоколями прочнее ввиду большего развития складчатости оснований и уменьшения полости пульпы (табл. VI, 2).

Правая и левая площадки на обеих челюстях сближены сильнее, чем у Anarhichas latifrons, что обеспечивает лучший захват цепляющегося за субстрат или прикрепляющегося к нему организма. Большая часть зубов на нёбных костях (а иногда и все), зубы внешнего ряда и задней части внутреннего на нижнечелюстных костях высокие, конические, как и у Anarhichas latifrons. Эти зубы образуют такие же «зубчатые ножницы», с той лишь разницей, что вследствие большего развития пережима на нижнечелюстных костях зубы нёбных костей располагаются частью кнаружи от внешнего ряда зубов нижнечелюстных костей и отодвинуты от клыков назад. Наибольшие давящие нижнечелюстные зубы оттесняются с площадки также назад, располагаясь в области пережима и сразу же за нею (табл. II, 2).
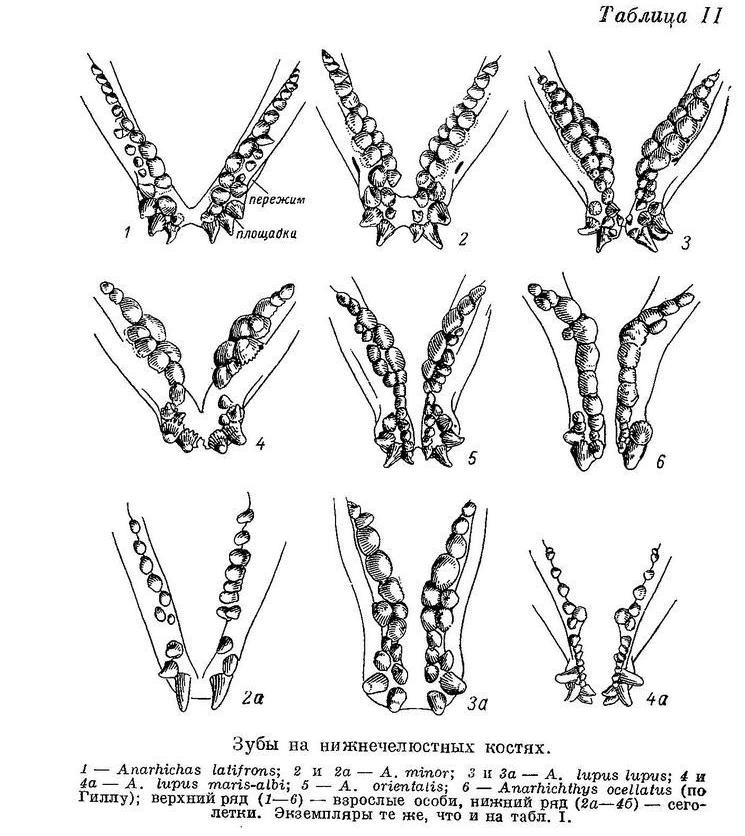
Соответственно удлиняются и ряды зубов на сошнике (табл. I, 2), простираясь теперь назад сходно с рядами зубов на нёбных костях (последние кончаются почти там же, где и внутренний ряд зубов на нижнечелюстных костях, что остается неизменным для всех видов зубаток без исключения).
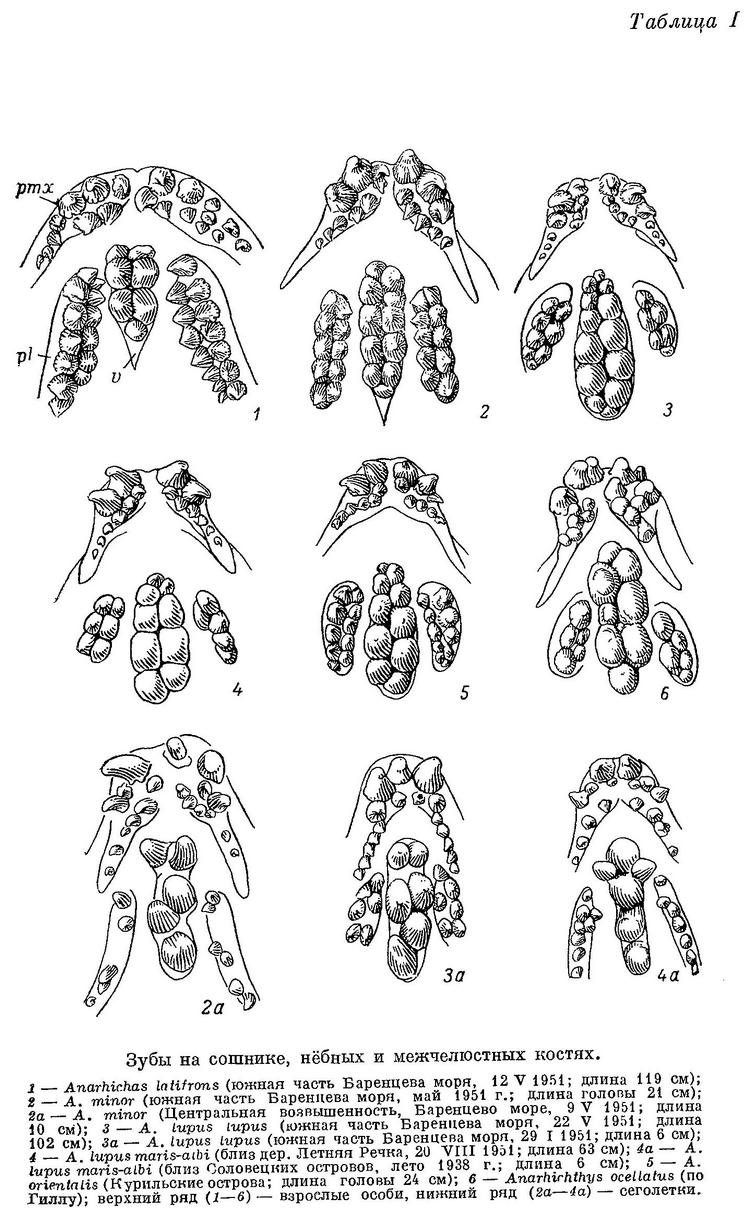
Увеличение давящих зубов по сравнению с размерами соседних, если и заметно, то не в переднем конце сошника, а посередине. Пережим на нижнечелюстных костях развит сильнее (табл. II, 2), опускание нижнечелюстных давящих зубов вниз и наклон их внутрь хорошо выражены. Задние зубы внутреннего ряда на нёбных костях обычно ниже передних и располагаются частью напротив, а частью внутрь от задних нижнечелюстных давящих, работая в паре с ними и подкрепляя таким образом действие сошника с боков. Следовательно, у Anarhichas minor, как и у Anarhichas latifrons, часть зубов, составляющих «зубчатые ножницы», начинает переходить к давящей функции, соответственно изменяя свою форму, причем выражено это в общем лучше и наблюдается у большего числа особей. Изменения в строении зубного аппарата Anarhichas minor явно вызваны приспособлением к питанию моллюсками большего размера и с более прочной раковиной: отрыванию их от субстрата (клыки) и раздавливанию (давящие). Питание иглокожими сохраняет свое значение: «зубчатые ножницы» на боках рта (конические зубы нижнечелюстных и нёбных костей) развиты сильно. Что касается питания более подвижными организмами, то возможность его должна быть несколько снижена изменением в форме и расположении клыков, особенно в период перед сменой, когда они затупляются.
Не менее интересна возрастная изменчивость в строении зубной системы.
У сеголеток Anarhichas minor длиной 10—12 см большие клыки на межчелюстных костях по размерам сильно превосходят малые; малых клыков на нижнечелюстных часто вообще не бывает. Внешний ряд нижнечелюстных зубов короток, зубов в нем очень немного (табл. II, 2а). Зубы на нёбных слабые, короткие и расположены чаще всего в один ряд (табл. I, 2а). Зубы внутреннего ряда на нижнечелюстных высокие, конические. Сильно развиты зубы на сошнике — конические, очень длинные (табл. V, 2), в несколько раз превышающие по размерам зубы нёбных костей.
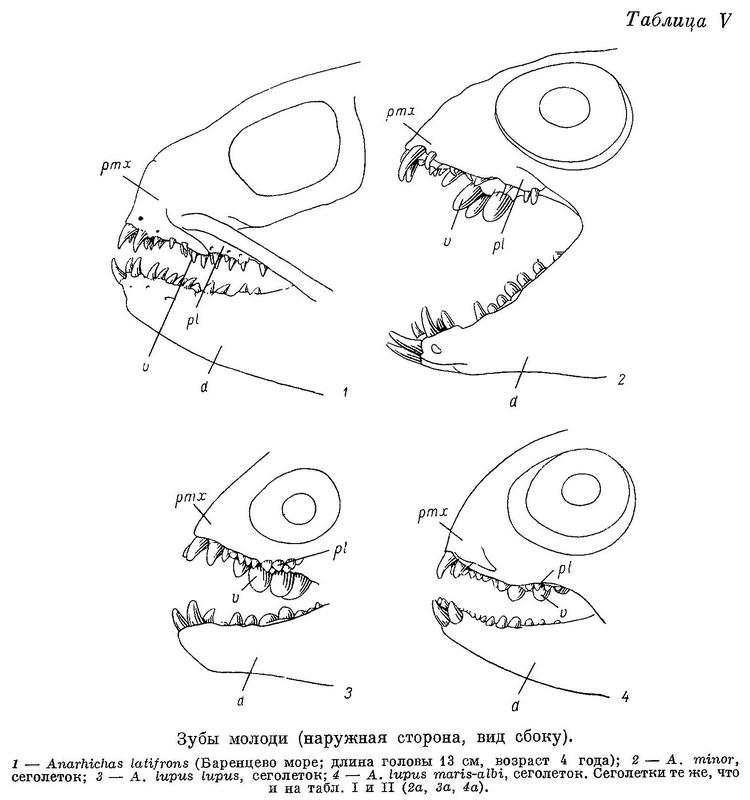
Таким образом, роль «зубчатых ножниц», как и у молоди Anarhichas latifrons, исполняют зубы сошника и внутренние ряды нижнечелюстных зубов, но ряды клыков по бокам челюстей не развиты. В дальнейшем роль «зубчатых ножниц» берут на себя удлиняющийся назад внешний ряд нижнечелюстных зубов и зубы на нёбных костях, быстро увеличивающиеся в числе и размерах. Передние же зубы внутреннего ряда на нижнечелюстных (считая от клыков) и зубы сошника приобретают округлую форму, характерную для давящих зубов. Судя по строению зубов, у Anarhichas minor уже сеголетки питаются в основном иглокожими, а в следующие годы в рацион все больше включаются моллюски.
Окраска
У молоди Anarhichas minor длиной менее 20 см бока и спина опоясаны темно-бурыми, почти черными поперечными полосами. На боках 7—11 полос, на спине 5—8; последние, как у Anarhichas lupus, сдвинуты по отношению к первым. На полосах можно различить более темные участки — будущие пятна, между полосами — неяркие серые пятна. На спинном плавнике напротив полос на спине от 1 до 7 крупных черных пятен, обычно по одному напротив каждой из полос, начиная с первой от головы; крупное черное пятно против нее всегда имеется. С возрастом полосы, начиная с передней, распадаются на отдельные пятна. Количество пятен на спинном плавнике, на теле, а также на верхней части головы (от затылка до переднего края глаз) и на щеках в дальнейшем возрастает, очевидно, за счет бледных и неясных пятен, увеличивающихся в размерах, приобретающих яркость и четкость. Однако группировка наиболее крупных пятен в полосы часто остается заметной и у взрослых особей. У некоторых особей, обычно самых крупных, в центре больших пятен образуется иногда светлое поле. Появляются также пятна в основаниях грудного и хвостового плавников. У молоди имеются неясные серые пятна на анальном плавнике, с возрастом исчезающие. Пятна на теле Anarhichas minor, очевидно, имитируют мелкие неровности грунта в местах ее обитания. Вероятно, указанные неровности довольно постоянны по размерам; поэтому размеры пятен, а вследствие появления новых и расстояния между ними с ростом тела зубаток увеличиваются медленнее, чем размер всего тела. Широкие полосы на теле молоди похожи на крупные пятна. Интенсивность бурых тонов в окраске может сильно измениться в течение немногих часов: крупный живой экземпляр, помещенный мной рядом с радиатором парового отопления, быстро и очень сильно побурел. Тем не менее эта интенсивность, как правило, связана с количеством пятен, меняющимся лишь с ростом: обычно зубатки с темно-бурым общим фоном имеют много пятен, а со светлым, имеющим желтовато-зеленоватые тона — мало. Различаются также особи с крупными и мелкими пятнами (Книпович, 1902 : 62, рис. 3 и 4). Общий фон окраски у взрослых обычно несколько темнее, чем у молоди, как и у предыдущего вида. Различие между более темными спиной и боками и более светлым брюхом у взрослых особей обоих видов сохраняется, но становится менее резким, чем у молоди.
Вариации окраски исландских Anarhichas lupus lupus и Anarhichas minor (Lühmann, 1954b: 313—315) весьма сходны с описанными выше вариациями окраски баренцевоморских.
Дополнение к разделу «Anarhichas minor». Возле побережья Лабрадора, по данным ПИНРО, Anarhichas minor встречена к западу до 60°14' с. ш. и 61°50' з. д. Отмечены случаи поимки возле Туле (P. М. Нansеn og F. Hermann, Fisken og havet ved Grønland, København, 1953 : 81). Обычна вдоль всего западного побережья Шпицбергена, где обнаружена сотрудниками ПИНРО вплоть до 79°50' с. ш. (тралений вдоль северного побережья не было).
В марте 1959 г. самки III стадии не встречались; обнаружена самка II—III стадии с несколькими икринками, оставшимися в яичниках от предыдущего нереста. Пелагическая молодь, по данным А. С. Бараненковой и Н. С. Хохлиной, ловилась в июне-июле вдоль южных окраин Западного желоба и далее к востоку вплоть до Финмаркенской банки; очевидно, наблюдениями были охвачены лишь южные окраины области распространения молоди Anarhichas minor. Тем не менее нерестилища этого вида должны быть распределены на несколько большей акватории, чем предполагалось ранее.
3а. Anarhichas minor Olafsen х Anarhichas lupus Linne.
Lühmann, Ber. Deutsch. wiss. Komm. Meeresforsch., XIII, 4, 1954 : 310—326, 17 Abb.
В уловах, привезенных в Киль из районов к северо-западу и западу от Исландии, Люманом обнаружены рыбы, несущие ряд промежуточных черт между Anarhichas lupus lupus и Anarhichas minor — в окраске, строении черепа и зубного аппарата. В уловах из районов к северо-востоку от Исландии таких особей нет. Число позвонков, а также лучей в спинном, анальном и грудном плавниках у них не промежуточное, а крайнее, немного превышающее это число у Anarhichas minor, что рассматривается как атавизм. Передний конец мезэтмоида вздут сильнее, а межчелюстная кость резче расширяется к цоколям зубов, чем у обоих видов. В той или иной степени нарушается периодичность смены зубов, их форма и расположение. Чаще и резче всего это наблюдается у зубов, испытывающих наибольшую нагрузку, но иногда захватываются даже глоточные зубы. Встречаются также повреждения суставов — прежде всего artuculare—quadratum, степень которых прямо зависит от степени нарушений в зубном аппарате и от выраженности промежуточных черт в окраске. Люман считает этих особей гибридами Anarhichas minor х Anarhichas lupus, вероятнее всего, от скрещивания самок Anarhichas minor с самцами Anarhichas lupus. По всем промежуточным признакам гибриды стоят ближе к Anarhichas minor, чем к Anarhichas lupus, и связаны с Anarhichas minor постепенными переходами. Предполагается, что гибриды плодовиты и возможно их повторное скрещивание с Anarhichas minor, но не с Anarhichas lupus.
Подобный же экземпляр с нарушенной сменой, формой и расположением зубов, что сильнее всего было выражено на сошнике, встречен мной в Баренцевом море на 70°28' с. ш. и 32°05' в. д. 16 V 1951. Его длина 111.5 см. Форма hyomandibulare, а также верхних частей metapterigoideum и praeoperculum с левой стороны ненормальна: создается впечатление, что нёбная дуга в этом месте была когда-то сломана, изогнувшись наружу, а затем срослась.
Питание у таких особей, по Люману, затрудняется лишь в случае глубоких нарушений зубного аппарата. По упитанности они мало отличаются от родительских видов. Относительно особенностей роста ничего определенного сказать нельзя: Люман считает, что Anarhichas minor и Anarhichas lupus растут одинаково быстро, но это вряд ли верно, по крайней мере для особей из Баренцева моря.
Не исключена возможность, что в данном случае мы имеем дело не с гибридизацией, а с какой-то болезнью, сопровождающейся нарушением обмена веществ.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1. Anarhichas minor Olafsen — Пятнистая, или пестрая, зубатка (рис. 113).
Anarrhichas minor Olafsen, Reise gjennem Island., I, 1772 : 592, tab. XLII (Исландия) (Цит. по: Collett, Rep. Norw. Mar. Fish. Invest., II, 3, 1905 : 113. В нашем распоряжении был лишь французский перевод книги Олафсена (v. 3, 1802 : 338).). — Steenstrup, Vidensk. Meddel. Naturh. Foren., (1876), 1877 : 161, 200, tab. III, fig. 2 (череп, зубы, сравнение с Anarhichas lupus). — Lilljeborg, Sverig. Norg. Fiskar, I, 1891 : 540 (описание, синонимия). — Smitt, Scand. Fish., I, 1893 : 236, pl. XIII, fig. 1 (цветн. рис.). — Книпович, Определитель, 1926 : 94, фиг. 59.
? Anarrhichas pantherinus Zоuiеw, Acta Ac. Sci. Petropol., V, 1, (1781), 1784 : 271, tab. VI (Архангельск; D 67, A 44, P 20). — Gmelin, Linn. Syst. Nat., ed. XIII, I, 3, 1788 : 1144 (sec. Zouiew). — Pallas, Zoogr. Rosso-Asiat., III, 1811 : 76 (Архангельск). — Collett, Norges Fiske, 1875 : 70 (Берген; дл. 126 см).
Anarrhichas lupus (non Linne) Günther, Cat. Fish., III, 1861 : 208 (частью: «american variety»).
Распространение, биология. Collett, Christ. Vidensk.-Selsk. Forhandl., (1879), 1, 1880 : 45 (Нордкап, Порсангер-фиорд; juv.). — Collett, там же, 1, 1902 : 91 (биология). — Книпович, Эксп. научно-пром. иссл. Мурмана, I, 1902 : 55 и сл., фиг. 2, 3 (многочисленные местонахождения у берегов Мурмана; дл. до 134.5 см, фотогр.). — Ehrenbaum, Wiss. Meeresunters. (Helgol.), VII, 1, 1905 : 54 (от о. Медвежьего до юго-зап. Шпицбергена; лучи в плавниках). — Collett, Rep. Norw. Mar. Fish. Invest., II, 3, 1905 : 113 (западнее Фарерск. о-вов на глуб. до 456 м; окраска мальков). — Jenkins, Fish. Brit. Island, 1925 : 110 (Эбёрдин в зап. Шотландии). — Thielemann, Wiss. Meeresunters. (Helgol.), XIII, 2, 1921 : 206 (экология, питание). — Дерюгин, Баренц. м. по Кольск. меридиану, 1924 : 74 (Баренц. м. по Кольск. меридиану под 71—72°30' с. ш.). — Bigelow a. Welsh, Fish. Gulf Maine, 1925 : 375, fig. 189 (на юг до зал. Массачусетс). — Jensen, Rap. Proc.-Verb., XXXIX, 1926 : 99 (Дэвисов прол.). — Saemundsson, Synops. Fish. Iceland, 1927 : 18 (вокруг Исландии; глуб. 25—400 м). — Duncker a. Mоhr, Tierwelt Nord u. Ostsee, XIIg, 1928 : 91, fig. 3 (близ Ставангер-фиорда, 59°20' с. ш.). — Зенкевич, Докл. 1-й сессии Гос. Океаногр. инст., 4, 1933 : 50 (питание). — Sсhnakеnbесk, Zool. Anz., CII, 7—8, 1933 : 191, fig. 1 (вост. Мурман; пелагическая личинка длиной 22 мм). — Merriman, Copeia, 3, 1935 : 143 (Ново-Шотландск. банки; фотогр.). — Rass, Int. Rev. Gesamt. Hydrobiol. Hydrogr., XXXIII, 3—4, 1936 : 262 (икра и личинки зубаток в Баренц. м.). — Есипов, Промысл. рыб. Баренц. м. 1937 : 51, фиг. 16 (Баренц. м. на восток до Гусиной банки). — Перцева, Тр. Всес. Научно-иссл. инст. морск. рыбн. хоз. океаногр., IV, 1939 : 460 (пелагические личинки в Мотовск. зал.). — Маслов, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 1944 : 156 (Баренц. м.; темп роста, плодовитость, промысел). — Есипов в изд.: Промысл. рыбы СССР, 1949 : 614, атлас табл. 188 (цветн. рис.).

D 74—80, А 45—47, Р 21—22 (По Лилльеборгу Р 24.), позвонков 78—79.
Голова с круто обрывающимся к рылу профилем. Длина сошникового ряда зубов равна длине нёбного ряда зубов или немного превышает ее. Все три ряда простираются назад почти на одинаковое расстояние, или сошниковые зубы слегка заходят за нёбные. Зубы менее уплощенные, чем у полосатой зубатки. Спинной плавник в конце с глубокой выемкой, образованной сильно укороченными колючими лучами, которые соединяются с основанием хвостового плавника. Окраска варьирует от желтоватого до серовато-коричневого тона, на котором отчетливо выступают многочисленные коричневато-черные округлые пятна, покрывающие голову, спинной плавник, спину, а также бока тела. Молодые экземпляры (8—10 см) имеют на боках тела 7—9 темных поперечных полос, которые с возрастом разбиваются на отдельные пятна. Длина в Баренцовом море до 134.5 см и вес до 20 кг, возможно достигает и несколько больших размеров, но указания на длину до 180—200 см (Смитт, Дункер и Мор и др.), вряд ли основаны на точных измерениях. В Баренцовом море чаще ловятся зубатки длиной 75—120 см и весом от 3 до 17 кг.

Распространение. Северная часть Атлантического океана и прилежащие к северу воды. Особенно многочисленна в Баренцовом море, встречаясь в значительных количествах на восток до Гусиной банки и на северо-западе до о. Медвежьего и юго-западных берегов Шпицбергена, но в северо-восточных районах моря, повидимому, отсутствует. Вдоль норвежских берегов доходит на юг до Бергена и изредка несколько южнее (район Ставангер-фиорда), далее до западных берегов Шотландии (Эбёрдин), Шетландских и Фарерских островов, вокруг Исландии. Реже встречается у западных берегов Гренландии и американского побережья, где известна на юг до зал. Массачусетс. Указания для Белого моря весьма сомнительны.
Биология. Пятнистая зубатка встречается на глубинах от 25 до 460 м, в Баренцовом море — от 53 до 425, но чаще, как и в других морях, на глубине 100—250 м. У Фарерских островов и зап. Гренландии ловится на несколько больших глубинах (350—456 м). Чаще встречается на илистых и илисто-песчаных грунтах; в отношении температурных и солевых условий, повидимому, не сильно отличается от Lycichthys denticulatus (Kröyer). Плодовитость пятнистой зубатки в Баренцовом море колеблется, по подсчетам П. 3. Сахно, от 15 180 до 20 250 икринок у самок длиной до 1 м и от 20 190 до 50 000 икринок у самок длиной более 1 м (Маслов). Нерест имеет место преимущественно на значительных глубинах в различных районах Баренцова моря, повидимому, до Новой Земли (Крестовая губа). Кладки представляют собой крупные шаровидные массы из слипшихся икринок диаметром 5—6 мм каждая (у вост. Мурмана такая кладка была обнаружена 10 апреля на глубине 110 м). Из икры выходят крупные (21—22 мм) сформированные личинки с рассосавшимся желточным пузырем, что отличает этот вид от Anarhichas lupus. Планктонные личинки появляются у берегов Мурмана в феврале—апреле, но встречаются и позднее. За первый год жизни мальки достигают 15—20 см, к 2 годам — 22.5—27.5, к 4 годам — 32.5—37.5, к 6 годам — 52.5—67.5 см (Маслов, по данным П. 3. Сахно); дальнейший рост в Баренцовом море происходит следующим образом:

Питание пятнистой зубатки составляют иглокожие (офиуры, Ctenodiscus, Strongylocentrotus и др.), крабы (Hyas), раки-отшельники, различные моллюски, отчасти рыбы (треска, камбала-ёрш).
Хозяйственное значение в Баренцовом море имеет наибольшее из других видов зубаток, в то время как в промысле Северного моря, Исландии и Сев. Америки преобладает полосатая зубатка. Наши уловы зубаток в Баренцовом море (преимущественно Anarhichas minor) достигали в довоенные годы (1936—1939) почти 40 тыс. ц. Основные уловы дает траловый промысел в западных районах Баренцова моря (весной) и центральных районах (июнь—июль). Мясо на Мурмане преимущественно засаливается, также употребляется в копченом, вяленом и свежем виде. Используется также икра и шкура зубаток.