Anarhichas lupus Linnaeus, 1758
Полосатая зубатка, обыкновенная зубатка, беломорская полосатая зубатка, баренцовоморская полосатая зубатка, английская полосатая зубатка (Rus),
Atlantic wolffish, Wolffish, Catfish, Seacat, Seawolf, Woolfish, Wauffs, Wuffs, Swinefish, Rockfish
(Eng)
Синонимы:
Anarhichas lupus Linnaeus, 1758
Anarhichas lupus marisalbi Barsukov, 1956 Беломорская полосатая зубатка (Rus),
Anarhichas strigosus Gmelin, 1789
Anarhichas vomerinus Agassiz, 1867
Anarrhichas lupus Linnaeus, 1758
Lupus marinus Jоnstоn, 1793
Lupus marinus nostras Schonevelde, 1624
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Зубатка полосатая — Anarhichas lupus (Linnaeus, 1758)
Как и у других представителей рода, нет брюшных плавников. Тело прогонистое с длинными спинным и анальным плавниками, в сумме в них порядка 100 лучей. Задние укороченные лучи спинного плавника жесткие. D LXIX—LXXIX. А 42—48. Р 19—20. Окраска тела песочно-желтая с 8—13 темными поперечными полосами, которые часто распадаются на ряд тесно лежащих пятен. Длина тела 1,2 м (обычно до 80 см). Обитает на дне морей северной части Атлантики, у нас — в Белом и Баренцевом морях.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
К семейству зубатковые (Anarrhichadidae) принадлежит очень хищная полосатая, или обыкновенная зубатка (Anarrhichas lupus). У нее вытянутое тело, рудиментарная чешуя и широкий рот с сильно развиты челюстями и бугорковидными губами. Длинный спинной плавник обособлен от хвостового. Брюшных плавников нет. Достигает в длину до 2 м. Сильными зубами способна дробить твердые раковины моллюсков. Живет зубатка в более холодных морях и известна даже у берегов Гренландии; держится на песчаных отмелях и в расщелинах скал. При плавании тело извивается подобно телу угря. Рыба ловится из-за вкусного мяса и прочной кожи, которая идет на изготовление сумок.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Полосатая, или обыкновенная, зубатка
(Anarhichas lupus) вооружена сильно
развитыми бугорковидными зубами. На
нижней челюсти они далеко сдвинуты назад,
что облегчает раздавливание раковин. Это
напоминает рычаг второго рода, где
укорочение плеча сопровождается увеличением
силы. Сверху на раковину давят зубы,
расположенные на нёбе; наиболее
крупные бугорковидные зубы сидят здесь
в сошниковом ряду, и последний тоже
простирается далеко назад за ряды
зубов на нёбных костях. По этому
признаку полосатая зубатка особенно
хорошо отличается от пятнистой и синей
зубаток. Она немного мельче, чем два
этих вида, и до сих пор не встречалась
длиной более 125 см и весом более 21 кг.
Бурые поперечные полосы на сером или
буровато-сером фоне хорошо маскируют
обыкновенную зубатку в зарослях
водорослей, где она любит держаться летом.
Давно замечено, что даже отдыхающая
в расщелинах скал зубатка нередко
медленно извивается на месте, что усиливает
сходство ее полос с водорослями,
колеблемыми токами воды. На глубинах, куда
полосатая зубатка уходит зимой, в ее
окраске появляются желтоватые оттенки,
а полосы бледнеют и часто становятся
почти совсем незаметными.
Полосатая зубатка обычна в западной
части Балтийского моря до островов Рюгена и Борнхольма, в Северном море
севернее 53° с. ш., у Шетландских и
Фарерских островов, у берегов Норвегии, в
Кольском и Мотовском заливах, у
северных берегов Кольского полуострова, у
острова Медвежьего, у западных берегов
Шпицбергена, вокруг Исландии (где
особенно многочисленна), у Гренландии и у
атлантического побережья Северной
Америки от полуострова Лабрадор до мыса
Код. В Баренцевом море она не
встречается севернее возвышенности Персея,
Центральной возвышенности и Новоземельской банки. Только этот вид зубаток
обитает в Белом море (у Карельского и
редко Терского берегов), образуя здесь
особый подвид.
Передвижения в основном
ограничиваются выходом к берегам и отходом на
глубины (до 450 м).
В южных районах чаще зимой, а в
северных летом самки полосатой зубатки
выметывают вблизи берегов от 600 до
40 тыс. (при длине рыбы 30—100 см)
крупных, 5—7 мм в диаметре, икринок,
которые прочно склеиваются в шарообразную
кладку, прилипающую ко дну. Самцы,
возможно, охраняют кладку, но только
в самом начале, так как развитие
эмбрионов длится долго, в течение нескольких
месяцев, и вылупляются они обычно весной,
будучи очень крупными (17—25 мм).
Вскоре после вылупления молодь начинает
подниматься над дном, нередко достигая
поверхности моря. При длине около 6—7 см все мальки переходят к жизни у дна
и в толще воды ловятся редко. От питания
планктоном они переходят к питанию
донными организмами, прежде всего
моллюсками и раками-отшельниками, а также
офиурами, морскими звездами и
морскими ежами, крабами и т. п. Полосатая
зубатка редко хватает рыбу. Моллюски —
лучшая наживка при лове ее на крючки.
Полосатую зубатку раньше били
острогой у берегов (любители на Белом море
бьют еще и поныне). Затем, как и других
зубаток, ловили главным образом
ярусами, а в настоящее время добывают в
основном тралом.
Мясо полосатой зубатки вкусное и в
ухе, и жареное, а также копченое и
соленое. Конечно, вкус его, как и у других
рыб, не всегда одинаков, зависит от того,
как и чем питалась зубатка перед
выловом. Недавно посоленная икра зубатки
мало уступает по вкусу кетовой икре.
Печень зубатки скандинавские рыбаки
считают деликатесом. Желчь в древности
употреблялась вместо мыла, а головой,
плавниками и костями исландцы кормили
овец и особенно коров, что улучшало
качество молока.
Фауна СССР. Рыбы. Том V, вып. 5. Семейство Зубаток (Anarhichadidae). В.В.Барсуков 1959 г.
4. Anarhichas lupus Linne — Обыкновенная или полосатая зубатка.
(Lupus — волк; на языке нескольких народов называется морским волком (см. ниже).
Anarhichas lupus Linne, Syst. nat., ed. X, 1758 : 247 («Oceano Septentrionalis Angliae»).
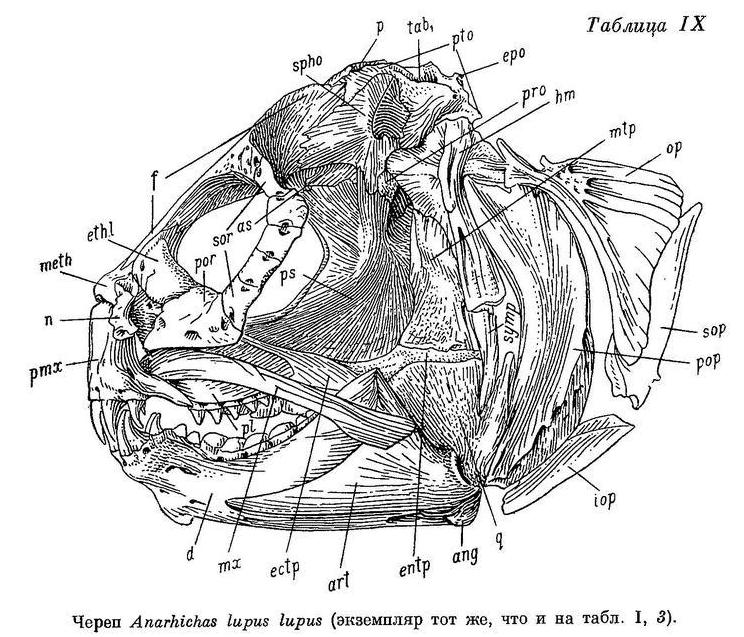
D (69) 70—77 (79?), A (42) 43—48, P 18—22, С 23—26, позвонков 72—77 (78) (туловищных 24—27, хвостовых 46—52), лучей жаберной перепонки 6—7, окологлазничных косточек с praeorbitale (5) 6—7. Наибольшая высота тела у взрослых 4.0—5.7 раз в его длине, у мальков длиной 6—10 см 4.7—7.2 раза. Антеанальное расстояние у взрослых 45.1—49.9% длины тела, у мальков 44.1—48.1%. Антедорсальное расстояние у взрослых 17.3—20.7%, у мальков 20.5—22.1%. Длина головы от конца рыла до заднего угла operculum у взрослых 17.9—21.6%, у мальков 21.1—22.5%. Расстояние от начала спинного плавника до верхнего края жаберной щели от 1/3 до 1/2 длины головы (у взрослых 31.4—43.3%, у мальков 27—38%). Ширина основания грудного плавника от 40 до 60% высоты тела у начала анального плавника. Хвостовой плавник несколько более закруглением у Anarhichas minor. Нижняя поверхность верхней губы покрыта неправильными складками, ее толщина здесь почти равна толщине нижней губы. Сошниковый ряд зубов длиннее нёбного ряда, простирается назад за нёбные ряды всегда дальше, чем вперед. У мальков зубы на нёбных костях расположены обычно в два ряда, иногда не продолжающихся назад далее зубов на сошнике. Длина сошникового ряда зубов у взрослых в 1.3—2.0 раза больше длины нёбного ряда, у мальков в 1.2—2.0 раза. Зубы внешнего ряда на нижней челюсти (исключая клыки) и зубы внутреннего ряда на нёбных костях округлые или тупоконические, часто сильно стертые. Внешний ряд зубов на нижней челюсти образует перерыв в области пережима нижнечелюстной кости. Наибольшие давящие зубы расположены далеко за пережимом. Жевательные мышцы развиты сильно, у взрослых составляют 10.3—14.8% веса головы с плечевым поясом и 4.2—6.0% веса тела. Сухожилие, идущее от переднего края порции А1 к передней части maxillare, отсутствует. Frontale позади заглазничных отростков у особей длиной более 20—30 см сужено в виде киля, в образовании которого pteroticum участия не принимает. Sphenoticum опущено незначительно: расстояние от вершины гребней на sphenoticum до верхнего края pteroticum в несколько раз меньше длины самих гребней. Верхний край гребней на sphenoticum расположен далеко позади вертикали переднего края parietalia. Крыша черепа довольно плоская (табл. XVII, 3, 4). Задний угол глазницы находится примерно на вертикали заглазничных отростков frontale. Бугорки на opistoticum сдвинуты вверх и не образуют часть контура неврокраниума (вид сзади). Кости прочные. Обонятельные нервы тоньше зрительных. Селезенка сдавлена с одной из сторон, образуя несколько заостренный край. В мышцах содержится около 80% воды.
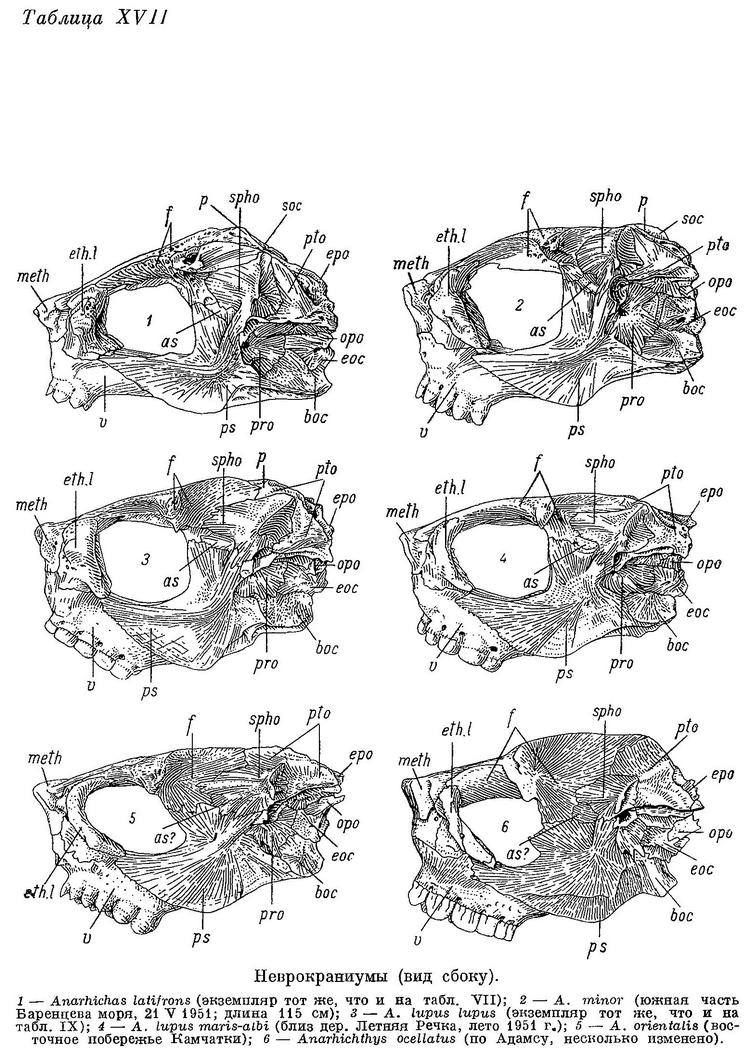
Общий фон окраски от желтоватого или светло-серого до коричневато-серого и даже темно-коричневого. Спина и бока несут 8—13 поперечных бурых полос, состоящих из неясных, сливающихся мелких пятен неправильной формы и продолжающихся на спинной плавник. Полосы нередко сливаются с общим фоном и становятся очень плохо заметными, что чаще наблюдается у взрослых особей. На спине и сверху головы иногда бывают мелкие, не превышающие диаметра зрачка пятна, более темные, чем полосы.
Придерживается наименьших глубин по сравнению с другими атлантическими видами и каменистых грунтов.
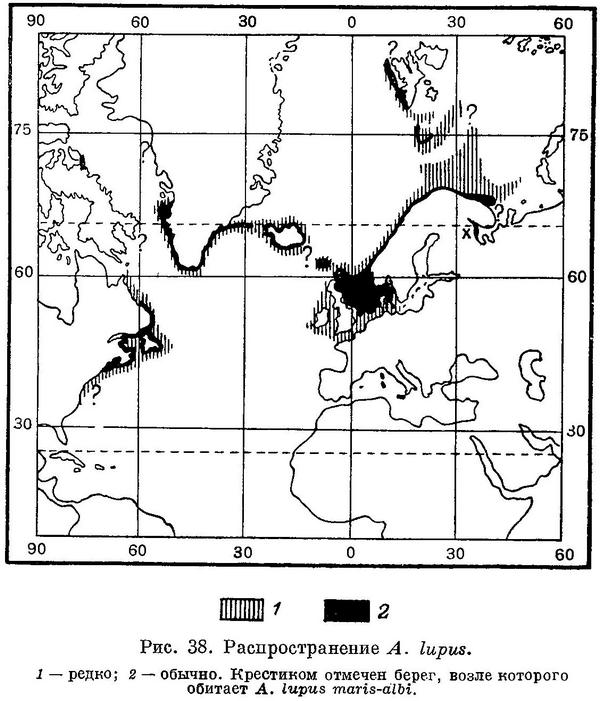
Распространение. Северная часть Атлантического океана (рис. 38). От северо-западных берегов Франции до западного побережья Шпицбергена. Скагеррак и Каттегат, Северное, Баренцево и Белое моря. Вокруг Исландии, у Гренландии. Обычна у американских берегов на юг до мыса Код и изредка встречается до мыса Гаттерас.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДВИДОВ
1 (2). Нижняя челюсть спереди обычно не утолщена. Зубов на обеих нёбных костях у особей длиной более 20 см в среднем не менее 14, обычно не менее 15. 4а. Anarhichas lupus lupus Linne.
2 (1). Нижняя челюсть спереди обычно утолщена. Зубов на обеих нёбных костях у особей длиной более 20 см в среднем менее 14, обычно менее 13. 4б. Anarhichas lupus maris-albi Barsukov.
4a. Anarhichas lupus lupus Linne — Обыкновенная или полосатая зубатка, называемая иногда синей зубаткой.
(Очевидно, последнее название пошло от беломорских рыбаков, называющих так свою беломорскую зубатку. В данном случае совпадение русских названий Anarhichas lupus и Anarhichas latifrons совершенно случайно, так как по внешнему виду они резко отличаются друг от друга.)
Stenbit, havkat, graassteinbit, hafskuse (норвежцы); havskatt (шведы); havkat, blåfisk (датчане); zeewolf (голландцы); Seewolf, gestreife Seewolf, Katfisch, gestreifter Katfisch, Steinbeisser, Karbonadenfisch, Austernfisch, Klippenstör (немцы); catfish, seacat, wolffish, seawolf, woolfish, wauffs, wuffs, swinefish, rockfish (англичане); merikissa (финны); steinbit(u)r (исландцы); kigutilik (гренландцы); loup (французы); zebacz (поляки).
Anarhichas lupus Linnaeus, Syst. nat., ed. X, 1758 : 247 («Oceano Septentrionalis Angliae»). — Müller, Zool. Dan., 1776 : 40 (Северное море). — Fabricius, Fauna Groenl., 1780 : 138—139 (Гренландия, экология). — Вlосh, Oekonom. Naturg. Fish. Deutschl., III, 1784 : 18—23 (Северное море; получена из Санкт-Петербурга). — Donovan, Nat. hist. Brit. fish., I, 1802, pl. XXIV (синонимия, использование). — Flemming, Hist. British anim., 1828 : 208 (длина в Северном море 2—3 фута). — Yarrell, Hist. Brit. Fish., I, 1836 : 247 (длина 6—7 футов). — Schlеgel, De Dieren Nederl., 1862 : 67—69 (Северное море). — Smitt, Scand. Fish., I, 1893 : 232—235, pl. XII, fig. 2 (морфология, экология, использование). Jordan a. Evermann, Fish. N. a. M. Amer., III, 1898 : 2446 (атлантическое побережье Сев. Америки). — Bigelow a. Welsh, Fish. Gulf Maine, 1925 : 370—375, tig. 186—188 (личинки, миграции). — McCormick, Trans. Roy. Canad. Inst., XV, 33, 1926 : 75 («основной» островок Лангерганса). — Вreder, Field-book mar. fish. Labr. Texas, 1929 : 275 (опред. табл.). — Jordan, Evermann a. Clark, Check list fish. N. a. M. Amer., 1930 : 472 (синонимия). — Есипов, в изд.: Пром. рыбы СССР, 1949 : 611, атлас, табл. 187 (сводка, цветн. рис.). — Андрияшев, Рыбы сев. морей СССР, 1954 : 224—227, фиг. 110, 3—4, 111 (3), 114 (синонимия). — Наlmе, Pohjolan kalat, 1954 : 114, tab. 47 (1) (цветн. рис.). — Барсуков, Докл. АН СССР, XCV, 4, 1954 : 897—899 (смена зубов). — Барсуков, Докл. АН СССР, 103, 4, 1955 : 715—716 (число лучей в плавниках у личинок и взрослых). — Макушок, Тр. Зоолог. инст. АН СССР, XXV, 1958 : 122, рис. 6, Д (число свободных хвостовых позвонков; кишечник).
Anarrhichas lupus Gronovius, Museum Ichth., 1754 : 16 (Северное море). — Andre, Philosoph. Trans. Royal Soc., LXXIV, II, 1784 : 274—277, pl. 11 (строение зубов). — Сuvier, Lepons anat. сотр., III, 1805 : 113, pl. 32, fig. 7 (строение зубов). — Кuhl, Beitr. Zool. vergl. Anat., 1820 : 171—173, 203—211, pl. IX (строение зубов, анатомия). — Сuvier, Anim. kingd., 10, 1834 : 234—235 (нет пилорических придатков и плавательного пузыря). — Cuvier et Valenciennes, Hist. nat. poiss., 11, 1836 : 472—492 (частью; критический обзор работ предыдущих авторов). — Retzius, Mullers Arch. Anat. Physiol., 1937 : 528—532 (строение зубов). — Parnell, Wernerian Nat. Hist. Soc., VII, 1838 : 239 (лучи спинного плавника). — Krøyer, Danmarks Fiske, I, 1838 : 369—381 (измерения; описание скелета и внутренних органов; созревание икринок, паразиты). — Owen, Odontography, I, 1840—1845 : 157—161, pl. 60—61 (строение зубов). — Dеkаy, Zool. New-York, III, 1842 : 158—159, pl. 16, fig. 43 (лучи хвостового плавника). — Crisp, Ann. Mag. Nat. Hist., (2), XI, 1853 : 463—466 (строение зубов, внутренние органы). — Günther, Catal. fish., III, 1861 : 208 (частью: «european variety», строение черепа). — Malm, Goteb. Bohusl. Fauna, 1877 : 468—469 (длина до 116 см). — Steenstrup, Vidensk. Meddel. Naturh. Foren., (1876), 1877 : 167, 200, pl. III, fig. 1 (череп, зубы, сравнение с Anarhichas minor; питание). — Bean, Proc. U. S. Nat. Mus., (1879), II, 1880 : 215 (измерения). — Collett, Christ. Vidensk.-Selsk. Forhandl., (1879), 1, 1880 : 50, 52 (измерения). — Retzius, Gehörorg. Wirbeltiere, 1881 : 66, pl. 10, fig. 5, 6 (внутреннее ухо). — Sørensen, Lydorgan. Fisk., 1884 : 68— 69 (скелет и мускулатура спинного плавника). — Dresel, Proc. U. S. Nat. Mus., VII, 16, 1884 : 247—248 (Дэвисов пролив; число лучей в плавниках и зубов). — McIntosh, Ann. Mag. Nat. Hist., (5), XV, 1885 : 432 (строение яичников, сравнение с Zoarces). — McIntosh a. Prince, Trans. Royal Soc. Edinb., XXXV, III, 1890 : 671—673, 781, 874—928, pl. I, fig. 25, pl. XX, figs 1—8, pl. XXI, figs 1—5a, pl. XXII, figs 2—3, pl. XXIII, figs 1—6, pl. XXIV, figs 1—7, pl. XXV figs 1—7, pl. XXVI, figs 1—4, pl. XXVII, figs 1—6 (подробное описание развития личинок в аквариуме). — Lilljeborg, Sverig. norg. Fisk., I, 1891 : 530—540 (описание скелета, измерения). — Маlmе, Bihang К. Svensk. Vet.-Acad. Handl., 17, IV, 3, 1891 : 27—28, pl. II, fig. 27 (строение мозга). — Mоreau, Manuel ichth. frang., 1892 : 151—152 (побережье Франции). — Gооd a. Bean, Ocean. Ichth., 1895 : 299, fig. 269 (измерения). — Haus, Intern. Monatschr. Anat. Physiol., XIV, 1897 : 42— 51, pl. V, fig. 1—7 (морфология и гистология кишечника). — Rennie, J. Anat. Physiol., XXXVII, 4, 1903 : 375, 378 («основной» островок Лангерганса). — Rennie, Quart. Journ. Microsc. Sci., N. S., 48, 191, 1905 : 384, 390, pl. 28, fig. 5 (островки Лангерганса, гистологическое строение «основного» островка). — Scott, 24-th ann. rep. Fish. Board Scotl., III, 1906 : 59, pl. III, figs 40—43, pl. V, fig. 21 (отолиты). — Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 170, figs 1, 2, 11, 12, pl. 17, 19, 23—26, 28 (рис. плечевого пояса, остальные рис. заимствованы). — Gemmill, Teratol. fish., 1912 : 1, 2, 30 (уродство). — Meek, Migr. fish., 1916 : 383—384, pl. XII (фотогр. кладки икры и молодой зубатки). — Lissner, Wiss. Meeresunters., Helgol., XIV, 1918 : 151, pl. V, fig. 15—17 (строение мозга). — Grenholm, Uppsala univ. arsskr., matem. naturvet., 2, 1923 : 189—190 (мышцы плавников). — Книпович, Определитель, 1926 : 93, фиг. 58. — Ehrenbaum, Handb. Seefischer. N. Europas, II, 1936 : 138—139 (промысел). — Van Dоbben, Arch. Neerl. Zool., II, 1937: 39—41 (строение челюстного аппарата). — Nybelin, in: Andersson, Fiskar Fiske Norden, I, 1942 : 103—104, pl. 29 (цветн. рис; нерест). — Ясвоин, Докл. АН СССР, LXXIV, 4, 1950 : 815 (строение дентина). — Lühmann, Verhandl. Anat. Ges., Erganzungsheft z. 97 Band, 1951 : 241—242 (смена зубов). — Kämpfe u. Kittel, Wiss. Zeitschr. Marton-Luther Univ. Halle—Wittenberg, II, 8, mathem.- naturwiss. Reihe, 4, 1952—1953 : 468 (довольно тонкие стенки кишечника не повреждаются обломками пищевых организмов). — Kittel u. Kämpfe, там же : 474 (относительный вес сердца, печени, селезенки и мозга). — Schmidt, Zeitschr. Zellforsch. mikrosk. Anat., 40, 1,1954 : 25—48 (строение и развитие зубов, поражение их паразитом «Mycelites ossifragus»). — Lühmann, Zeitschr. Zellforsch. mikrosk. Anat., 40, 5, 1954 : 470—509 (смена зубов, гистология). — Lühmann, Ber. Deutsch. wiss. Komm. Meeresforsch., XIII, 4, 1954 : 310—326, Abb. 1—3, 7, 8, 11, 16 (окраска, череп, жевательные мышцы, зубной аппарат; сроки нереста и смены зубов). — Schmidt, Natur u. Volk, 85, 2, 1955 : 58—59, Bild. 1—2 (поражение зубов микроорганизмом «Mycolites ossifragus»).
Anarhicas lupus Bonnaterre, Tabl. Encyol., Ichthyol., 1788 : 38, pl. 26, fig. 87 (форма фронтальных разрезов тела). — Retzius, Fauna Suec., 1800 : 315 (синонимия). — Faber, Naturgesch. Fisch. Islands., 1829 : 70—75 (Исландия; частью; связь строения зубов с питанием, зависимость окраски от грунта).
Anarchicas lupus Smith, Nat. hist. fish. Massachusets, 1833 : 249—251 (размеры).
Anarrhicas lupus Gill, Proc. Acad. Nat. Sci. Phila., 1861 : 46 (nomen nudum).
Anarrhichas strigosus Gmelin, Linn. Syst. nat., ed. XIII, 1, 3, 1788 : 1144 (синонимия).
Anarrhichas vomerinus Agassiz in: Storer, Mem. Amer. Acad. Arts Sci., V, 1, 1853 : 265—266, pl. XVlII, fig. 1 (рис. головы спереди; подход к берегам зимой; на каменистом грунте). — Gregory, Fish skulls, 1933 : 376, fig. 254c, 255 (рис. черепа).
Annarhichas (Anarrhichas) lupus Baron, Zeitschr. wiss. Zool., 146, 1, 1935 : 4, 29—31, 34, Abb. 19 (островок Лангерганса и цимогенная ткань поджелудочной железы).
? Anarrhichas sp. Collett, Christiania Vidensk.-Selsk. Forhandl., (1879), 1, 1880 : 58 (личинка 24 мм у Варде).
? Anarrhichas latifrons (non Steenstrup) Dannevig, Canad. Fish Exp. 1914—1915, Ottawa, 1918 : 16 (зал. Св. Лаврентия; личинки 21—25 мм).
Anarhichas denticulatus (non Krøyer) Pietschmann, Meddel. Granl., 92, 3, 1932 : 53—55 (Гренландия; молодой экземпляр, измерения, окраска в спирте).
Anarrhichas minor (non Olafsen) Schnakenbeck, Zool. Anz., CII, 7—8, 1933 : 191—193, fig. 1 (личинки 22—23 мм у м. Черный). — Перцева, Тр. Всесоюзн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., IV, 1939 : 460 (личинки 22—23 см в Мотовском заливе). — Расс, Тр. Всесоюзн. научно-исслед. инст. морск. рыбн. хоз. и океаногр., XVII, 1949, рис. 15 а.(малёк).
Wolf-fish Pennant, Brit. Zool., III, 1769 : 119, fig. 7 (измерения; связь ширины кишки с питанием; экология). — Low, Fauna Orcad., 1813 : 187—188 (движения ноздрей).
До Линнея:
Anarrhichas, Scansor Gеsnеr, Fischbuch, 1598 : 63 (привязанность к каменистому грунту; ноздри, зубы). — Аrtеdi, Synon. nomin. pise., ichth., ed. II, IV, 1793 : 38 (синонимия).
Lupus marinus nostras Schonevelde, Ichth. nomen. anim., 1624 : 45, pl. V (у Гельголанда; описание формы тела, зубов, окраски).
Lupus marinus Schoneveldii Jоnstоn, pl. 47, fig. 2 (цит. по: Artedi, 1793).
Latargus Klein, Hist. Pisc. nat., 4, 1740 : 16—17 (ископаемые зубы).
Lupus marinus nostras et Schonefeldii Willoughby a. Ray, Hist, pise., 1743 : 130 (описание формы тела и внутренних органов).
Распространение, биология. Olafsen, Reise durch Island, 1774 : 314—315 (Исландия; прибрежный промысел, использование). — Озерецковский, Описание Колы и Астрахани, 1804 : 36 («Архангелогородцы, для красы, привешивают... к потолокам...» зубаток, привезенных, очевидно, из Кольского залива). — Smith, Nat. hist. fish. Massachusets, 1833 : 249—251 (промысел). — Thompson, Proc. Zool. Soc. London, III, 1835 : 80 (Ирландия). — Fries a. Exstrom, Scand. Fish., 1836 : 39—42 (экология). — Yerrill, Amer. Nat., V, 1871 : 400 (питание). — McIntosh, Mar. invert. fish. St. Andrews, 1875 : 175 (питание). — Couch, Hist. fish. Brit. Isl., II, 1877 : 242—244 (не остается на берегу после отлива); — Good a. Bean, Bull. Essex. Inst., 11, 1879 : 11 (атлантич. побережье Америки). — Wilson, Nature, XIX, 1879 : 556—557 (питание). — Winthеr, Prodr. ichth. Dan. Mar., 1879 : 22 (вертикальное распределение). — Day, Fish. Great. Brit. Irel., I, 1880—1884 : 194—197, tab. LVIII (экология). — Olsen, Pise, atlas North Sea, 1883 : 25 (промысел, цит. по Gill, 1911 : 183). — Kent, Handbook mar. freshw. fish. Brit. Isl., 1883 : 56—58 (поведение в аквариуме). — Good, Fish a. fish. industr. U. S., I, 1884 : 248 (молодь с глубины более 180 м). — Brook, 4-th ann. rep. Fish. Board Scotl., III, 1886 : 245 (нерест). — McIntosh, Rep. St. Andrews Mar. Labor, IV, 1886 : 356—357 (икринки; цит. по: Gill, 1911 : 186). — McIntosh, 4-th ann. rep. Fish. Board Scotl., III, 1886 : 213 (молодь). — McIntosh, 5-th ann. rep. Fish. Board Scotl., III, 1887 : 356 (находка кладки с эмбрионами). — Smith 7-th ann. rep. Fish. Board Scotl., III, 1889 : 231, 252 (питание). — Ewart a. Fulton, там же: 197, 212 (зрелые самки в августе и феврале). — Smith, 8-th ann. rep. Fish. Board Scotl., III, 1890 : 231, 244—245, 251 (питание). — Fulton, там же : 161—163, 176, 183, 201, 204, 269 (предположительные размеры при созревании, экология, личинки). — Fulton, 9-th ann. rep. Fish. Board Scotl., III, 1891 : 244, 248, 253—254 (плодовитость; быстрый переход из III в IV стадию зрелости). — Smith, там же : 232, 235, 239, 240, 253 (питание). — Smith, 10-th ann. rep. Fish. Board Scotl., III, 1892 : 229 (питание). — Fulton, там же : 239, tabl. V (созревание). — Brenner, Meddel. Soc. Fauna Flora Fenn., 22—23, 1896 : 59 (2 экз. около 40 см у Хельсинки, по рассказу). — Knipowitsch, Ежег. Зоолог. муз. Акад. наук, II, 1897 : 148 (Мурман, Белое море, обычна). — McIntosh a. Masterman, Life-hist. mar. food-fish., 1897 : 200—205, pl. VIII, fig. 1—2 (развитие личинок в аквариуме). — Fulton, 16-th ann. rep. Fish. Board. Scotl., III, 1898 : 89, 91 (созревание икринок). — Milrоy, там же : 143 (содержание воды в икринке). — McIntosh, Resources of the sea, 1899 : 51—52, 174—175, 178—179, tab. 6, 7, 10—13, 15, 17, 22 (кладка с эмбрионами; уловы по месяцам; приуроченность к каменистым грунтам). — Ehrenbaum, Fauna Arctica, II, 1901 : 95—96 (Шпицберген, о. Медвежий). — Scott, 20-th ann. rep. Fish. Board Scotl., III, 1902 : 500 (питание). — Книпович, Эксп. научно-пром. исслед. Мурмана, I, 1902 : 64 и сл. (многочисленные местонахождения у берегов Мурмана). — Книпович, Ежег. Зоолог. муз. Акад. наук, VIII, 1903 : 153 (у Шпицбергена только на западном побережье). — Fulton, 21-st ann. rep. Fish. Board. Scotl., III, 1903 : 23, 63—65, 80, 82—96, 98, 99 (миграции; молодь придерживается каменистых грунтов и поэтому избегает трала). — Scott, там же : 220 (питание). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1903. — Книпович, там же, II, 1, 1904 (местонахождения в Баренцевом море). — Thompson, North Sea Fish. Invest. Comm., 1905 : 344—346 (уловы в Северном море). — Fulton, 23-rd ann. rep. Fish. Board Scotl., III, 1905 : 251—254 (созревающие самки в августе). — Ehrenbaum, Eier u. Larven, 1905—1909 : 91, fig. 41 (икра, личинки). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1906 (местонахождения в Баренцевом море). — Ehrenbaum, Wiss. Meeresunters., Helgol., VII, I, 1905 : 54 (юго-зап. Шпицберген и о. Медвежий). — Todd, North sea fish, invest, comm., rep. 2, (1902—1903), 1905 : 233 (питание). — Соrnish, Rep. Dept. Mar. Fisher. Canada (1902—1905), 39, 1907 : 87—88 (питание). — Todd, North Sea Fish. Invest. Comm., rep. 2, (1904—1905), 1907 : 58 (питание). — Thompson, там же, rep. 3, (1904—1906), 1908 : 272—276, figs 76—81 (траловые уловы в Северном море по месяцам). — Lеbоur, Northumberl. Fish. Comm. Rep. Sci. Invest., 1908 : 25 (питание). — Carr, там же : 71 (питание). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1908 (местонахождения в Баренцевом море). — Fulton, 27-th ann. rep. Fish. Board Scotland, III, 1909 : 157, 162, 166,169, 172, 174, 178 (уловы). — Брейтфус, Эксп. научно-пром. исслед. Мурмана, II, 1912; II, 1915 (местонахождения в Баренцевом море). — Дерюгин, Фауна Кольского залива, 1915 : 592 (Кольский залив). — Thielemann, Wiss. Meeresunters., Helgol., XIII, 2, 1921 : 205—206, karte 5 (Баренцево море). — Wоllebaek, Norg. fisk., 1924 : 210, fig. 234 (икра, личинки). — Воrley а. Тhursby-Реlham, Fish. invest., ser. II, IX, 4, 1926 : 5, charts 7—8 (в Северном море практически отсутствует к югу от 53° N). — Saemundsson, Synops. Fish. Iceland, 1927 : 17 (обычна вокруг Исландии). — Dunker u. Mohr, Tierwelt Nord.-Ostsee, XII, 1928 : 90—91 (длина до 125 см; биология). — Расс, Докл. 1-й сессии Гос. океаногр. инст., 5, 1931 : 3, 4, рис. 1, 2 (кладки зубаток в Баренцевом море в июле-августе; Anarhichas lupus?). — Базикалова, в: Сборн. научно-пром. работ Мурмана, 1932 : 137, 143—147 (паразиты). — Lundbeck, Wissensch. Meeresunters., Helgol., XVIII, 3, 1932 : 13—16, 18 (миграции). — Смapагдова, Тр. Гос. океаногр. инст., III, 2, 1933 : 99—123 (уловы в южной части Баренцева моря). — Merriman, Copeia, 3, 1935 : 143 (Ново-Шотландские банки; фотогр.). — Rass, Intern. Rec. ges. Hydrobiol. Hydrogr., 33, 3—4, 1936 : 262—263, fig. 3 (кладки и личинки зубаток в Баренцевом море; Anarhichas lupus и Anarhichas. minor?). — Lundbeck, Ber. Deutsch. wiss. Komm. Meeresforsch., VIII, 2, 1936 : 100—103, tab. 1, 3, 4, 6, 7, 19 (уловы в разных частях ареала; вес, длина и возраст в уловах; в табл. 1, 3, 6, 7 Anarhichas lupus и Anarhichas minor приведены вместе). — Есипов, Пром. рыбы Баренц. моря, 1937 : 50, фиг. 15 (на восток до Гусиной банки). — Jensen, Rapp. Proces-Verb., СП, 6, 1937 : 7 (у Борнхольма, по Otterstrøm, Flora och Fauna, 1930 : 45). — MсКenzie a. Homans, Proc. Nova Scotian Inst. Sci., XIX (1936—1937), 3, 1938 : 279 (кладка икры с эмбрионами). — Маслов, Тр. Полярн. научно-исслед. инст. рыбн. хоз. и океаногр., VIII, 1944 : 154 (темп роста, плодовитость). — Jensen, Spolia Zool. Mus. Haun., IX, 1948 : 117—118 (Гренландия). — Saemundssоn, Zool. Iceland, IV, 72, 1949 : 31—32 (Исландия). — Bigelow a. Schroeder, Fish. Gulf Maine, 1953 : 502—507 (окраска, размеры; в эстуариях не встречается; питание; личинки и мальки; уловы; обычна в заливе Мэн). — Morrow, Copeia, 3, 1957 : 240—241 (Лонг-Айленд-Саунд — южный предел распространения за последние 55 лет; самка длиной 80 см поймана 17 V 1956 в оттертрал). — Полянский, Тр. Зоолог. инст. АН СССР, XIX, 1955 : 45, 46, 49, 59, 61, 63—65, 82, 88, 90, 94, 97, 99, 109, 117 (паразиты). — Барсуков, Вопр. ихт., 1957 : 29—34, рис. 1, 2, 11, 1, 12, 1, 13, 1 (распределение в Баренцевом море). — Kotthaus u. Krefft, Ber. Deutsch. wiss. Komm. Meeresforsch., XIV, 3, 1957 : 178 (места поимки у побережья Гренландии и Исландии; экземпляр длиной 5 см из желудка трески). — Полянский, в сб.: Основные пробл. паразитолог. рыб, 1958 : 83 (у берегов до верхних горизонтов сублиторали).
D (69) 70—77 (79?), А (42) 43—48, Р 18—22, С 23—26, позвонков 74—77 (78) (туловищных 25—27, хвостовых 47—52), лучей жаберной перепонки (6) 7, окологлазничных косточек (с praeorbitale) 6—7. Нижняя челюсть спереди не утолщена. Зубы мельче и многочисленнее, чем у Anarhichas lupus maris-albi. Зубов на обеих нёбных костях, у особей длиной более 20 см 12—20, в среднем около 16, а у особей более 60 см около 18. Всех зубов на межчелюстных, нижнечелюстных, нёбных костях и сошнике 58—94, в среднем около 70, а у особей более 60 см — около 80. Зубов на глоточных костях 163—286 (2 экз.). Posttemporale обычно несколько наклонена вниз, образуя тупой угол с передним краем supracleithrum.
Молодь длиной 6—7 см по расположению зубов на нижнечелюстных, межчелюстных, нёбных костях и сошнике близка к взрослым особям: зубы на нёбных костях и сошнике располагаются в два ряда, сошниковые ряды простираются назад значительно далее нёбных (табл. I, 3а).
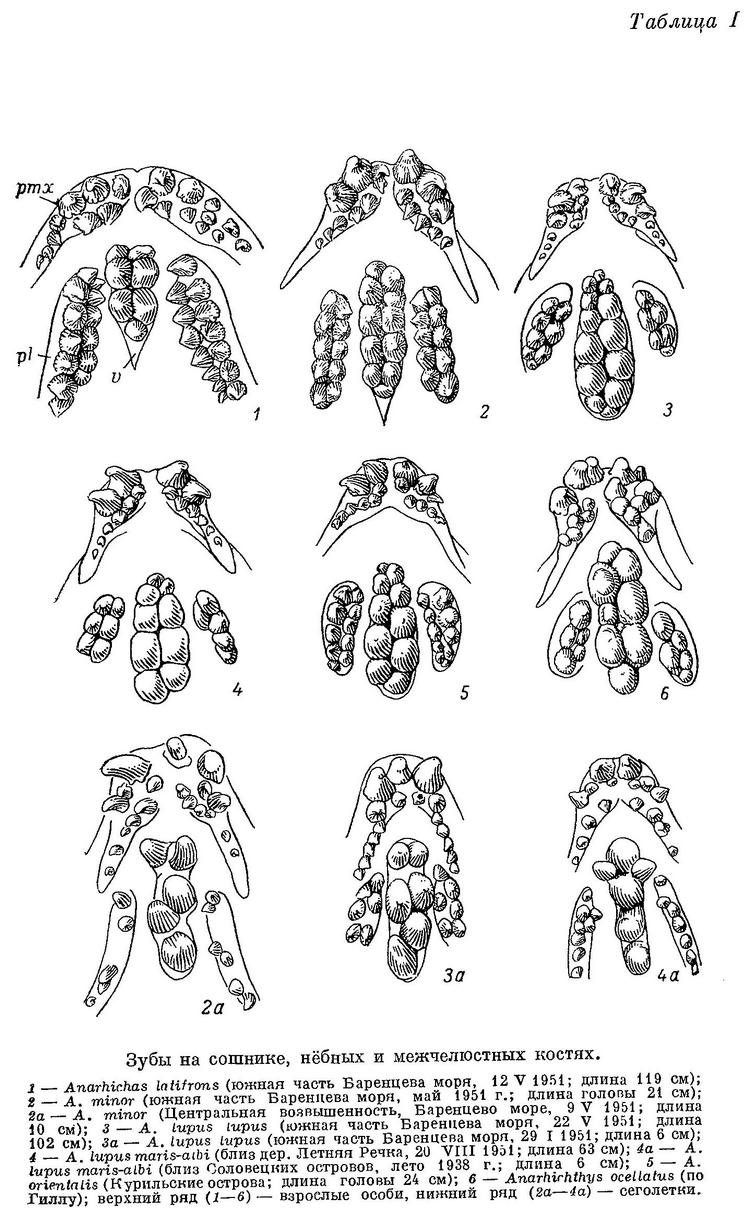
В общем фоне окраски имеются желтоватые или голубоватые тона. Полосы у молоди бледнее, менее отчетливы, чем у следующего подвида; пятен того же цвета, что и полосы, между ними нет. По Бигелоу и Шредеру (Bigelow a. Schroeder, 1953 : 505), у мальков длиной 3—4 см бока имеют серебристый оттенок, пропадающий при фиксации.
Описание дано по 200 экз. из Баренцева моря; позвонки и лучи в плавниках подсчитаны у 35—56 экз., зубы у 34 экз., череп и жевательные мышцы описаны (см. Anarhichas lupus) по 3—8 экз.
Длина до 120—125 см; зубатка длиной 120 см имела вес 20.5 кг (50 фунтов — Ehrenbaum, 1936 : 139). Встречающиеся в литературе указания на длину 150, а тем более 180 см основаны, судя по всему, на неточных данных. Так, по Бредеру (Breder, 1929 : 275), зубатка длиной 5 футов (150 см) имела вес всего 30 фунтов (12.3кг): такое соотношение длины и веса возможно разве что у Anarhichthys, но не у Anarhichas. Были указания и на еще большую длину полосатой зубатки — 10—15 и даже 18 футов (Willoughby, 1743 : 130; Donavan, 1802 : XXIV, и др.); ошибочность этих данных несомненна, неясно лишь их происхождение. Длина наибольшего экземпляра Anarhichas lupus у берегов Швеции 116 см (Malm, 1877 : 468), в Баренцевом море 115 см; зубатка длиной 102 см имела вес 9.1 кг. В уловах обычна от 20 до 90 см (особей длиной 70—80 см в 1946—1950 гг. вылавливалось несколько больше, чем длиной 50—60 и 60—70 см). В Северном море, у побережья Исландии и Северной Америки особи длиной более 90 см также редки. Максимальная длина у побережья Гренландии в 1955 г. 83 см (Kotthaus a. Krefft, 1957 : 178; по 386 экз.). Средний размер в траловых уловах в южной части Баренцева моря 64 см, у о. Медвежьего 58 см, у Лофотенских островов и западного побережья Шпицбергена 39 см (1941—1950 гг.). В 1928—1931 гг. средняя длина в Баренцевом море 69.8 см, у о. Медвежьего 70.5 см, у берегов Исландии 71.2 см, в Северном море 62.0 см, в проливе Каттегат 56.3 см; средний вес, соответственно — 2.4, 2.8, 2.3, 1.73, 1.42 кг (Lundbeck, 1936, tab. 19). В Северном море средняя длина зрелых самцов 74 см (33 экз.), зрелых самок 70 см (26 экз.; Fulton, 1890 : 349-350).
Распространение. Северная часть Атлантического океана. На юг распространяется далее других видов, изредка встречаясь даже у северо-западных берегов Франции (по-видимому, лишь в особо холодные годы). Обычна в Северном море (на юг до 53° с. ш.), у Шетландских и Фарерских островов, у берегов Норвегии, в Кольском и Мотовском заливах и у берегов Кольского полуострова вплоть до мыса Святой Нос; заходит ли в воронку Белого моря, неизвестно. В южной части Баренцева моря распространяется на север до юго-западного угла Центральной возвышенности, на восток до Гусиной банки, а в наиболее теплые годы, судя по данным Г. И. Милинского, почти до Новоземельской банки. В северозападной части моря обычна на западном склоне Медвежинской банки и у западного побережья Шпицбергена по крайней мере до о. Принца Карла; по склонам Медвежинской банки распространяется до о. Надежды и возвышенности Персея, где редка. Известна в западной части Балтийского моря до островов Рюгена и Борнгольма, но отдельные экземпляры проникают — по-видимому, очень редко — и до Финского залива (Bloch, 1784 : 23; Brenner, 1896 : 59). Вокруг Ирландии редка. Вокруг Исландии обычна, по большей части ловится на западном и южном побережьях. У Гренландии по западному побережью отмечена на север до Диско-фиорда, по восточному — до фиорда Ангмагссалик. Дависов и Датский проливы (Jensen, 1948 : 117; Kotlhaus u. Krefft, 1957 : 178). Обычна у американских берегов от залива Хамильтон (Лабрадор) на юг до мыса Код; изредка встречается до Нью-Джерси, а по данным некоторых авторов, и до мыса Гаттерас (может быть, в особо холодные годы?). За последние 50 лет перед 1953 г. не встречалась южнее Вайньярд-Саунда (Bigelow a. Schroeder, 1953 : 506); лишь в мае 1956 г. была добыта несколько южнее, на Лонг-Айленд-Саунд (Morrow, 1957 : 241). На Нью-Фаундлендских банках распространена широко.
Биология. В Баренцевом море ловится в тралы и на яруса на глубинах от 18 до 380 м, у побережья Исландии — от 10 до 430 (450) м, у побережья Гренландии — от 14 до 420 (450) м, у побережья Северной Америки — от 2 до 435 м. По Г. И. Милинскому, в Баренцевом море встречается и у самых берегов, на совсем незначительных глубинах. Это подтверждается зараженностью Anarhichas lupus lupus паразитом Podocotyle atomon; что является верным свидетельством питания либо на литорали (Полянский, 1955 г 162), либо в верхних горизонтах сублиторали (Полянский, 1958 : 83). У других атлантических видов зубаток Podocotyle atomon не найден. То же самое наблюдается у побережья Гренландии (Fabricius, 1780 : 139) и Норвегии (Bloch, 1785 : 22; Donovan, 1802 : XXIV, и др.), где ее бьют или били острогою. На глубины 1—2 м, к самым берегам, подходит обычно ранней весной, летом же концентрируется на глубинах от одного до нескольких десятков метров. В начале, а в теплые годы в середине зимы, начинается массовый отход на глубины 100—200 м и более. В разных частях ареала наблюдаются значительные отклонения от этой схемы, да и в одном и том же районе зубатки ведут себя по-разному. Например, летом и в начале осени 1955 г. у побережья Гренландии, кроме обычных скоплений на глубине в несколько десятков метров (58—70 экз. за полчаса траления на глубинах 58—70 м), наблюдались довольно значительные скопления на глубинах 200—350 м (около 30 экз. за полчаса траления, — Kotthaus u. Krefft, 1957 : 178). Кроме индивидуальных, имеются возрастные различия: в Баренцевом море молодь и на мелководья, и на глубины выходит реже и в меньшем числе, чем взрослые особи (Барсуков, 1957 : 32). У побережья Исландии молодь длиной 8—10 см на мелководьях также не наблюдалась (Saemundsson, 1949 : 32); у побережья Норвегии на больших глубинах отмечены особо крупные экземпляры (Lilljeborg, 1891 : 537), — судя по всему, и здесь предельные глубины у молоди различаются меньше, чем у взрослых.
Всюду предпочитает каменистые грунты; особенно привязана к ним молодь.
Требовательность к солености, очевидно, ниже, чем у других атлантических зубаток, однако в эстуариях не встречается (Bigelow a. Welsh, 1925 : 372; Bigelow a. Schroeder, 1953 : 504).
В Баренцевом море ловится при температурах от —0.97 до +7.4°; наибольшие уловы тралом наблюдались при 4—6°. У побережья Гренландии в июне-октябре 1955 г. встречалась при температурах от 1.3 до 5.5°; у побережья Исландии в июне-сентябре — от 1.8 до 7.1° (Kotthaus u. Krefft, 1957 : 178). Во всех этих районах у самых берегов, где не захватывается тралением, обитает, возможно, при более высоких температурах. У берегов Шотландии ловилась в феврале-мае при 6—8°, а в сентябре — при 9—10° (Fulton, 1891 : 40). Предельные температуры, при которых встречалась в заливе Мэн, примерно от —1 до +11° (Bigelow a. Schroeder, 1953 : 504).
У берегов активна только ночью; днем отстаивается в укрытиях дна. Как ведет себя во время полярного дня, не выяснено.
Миграции почти неизвестны. Тем не менее некоторые данные позволяют предполагать не столь значительную протяженность миграций, как у других атлантических видов: к выводам о малой протяженности миграций полосатой зубатки склоняется ряд авторов, наблюдавших ее у побережья Скандинавии, Исландии и Северной Америки (Nillsson, 1855 : 211; Smitt, 1893 : 235; Bigelow a. Welsch, 1925 : 373; Saemundsson, 1949 : 32; Bigelow a. Schroeder, 1953 : 504). При передвижениях в Северном море Anarhichas lupus, по-видимому, не выходит за его пределы. По распределению уловов можно предполагать, что с июня-июля начинается подход к берегам, на каменистые грунты, неудобные для траления. Этот подход наиболее интенсивен в сентябре-декабре, в силу чего уловы снижаются вплоть до января, когда начинается отход от берегов в открытое море, причем наибольшие скопления наблюдаются в средней части моря, перед проливом Скагеррак (Thompson, 1905 : 344—345; 1908 : 202—274; Lundbeck, 1954 : 15). Молодь же длиною менее 30 см постоянно держится на каменистых грунтах и поэтому в тралы попадает редко (Fulton, 1903 : 64). Предполагаются также сезонные миграции полосатой зубатки с севера на юг весной и ранним летом (Borley a. Thursby-Pelham, 1926 : 5). В Баренцевом море в местах, охваченных тралением, в течение всего года концентрируется в одних и тех же районах (Восточно-Прибрежный район, западный и южный склоны Медвежинской банки).
Размеры, при которых созревают самки, в разных частях Баренцева моря, очевидно, различны. Самки I—II стадии начинают встречаться у Лофотенских островов при длине 15 см, у о. Медвежьего — при 22 см, в южной части моря — при 24 см; самки II стадии — у Лофотенских островов при длине 17 см, в южной части моря — при 36 см. Самок более крупных мной исследовано очень мало; из них самки II—III стадии встречались при длине не менее 41 см, самки III стадии — не менее 50 см. По Г. И. Милинскому (1944), Anarhichas lupus созревает в Баренцевом море при длине не менее 40—45 см, в возрасте более 5 лет. В Северном море и у берегов Исландии созревает при длине не менее 50—60 см (Fulton, 1890: 161; Saemundsson, 1949 : 32).
Плодовитость самок, по Сахно (Маслов, 1944 : 156), от 2560 (длина тела 43 см) до 23 580 (81 см) икринок (Баренцево море); у более крупных особей — до 40 000 (Северное море, — McIntosh a. Masterman, 1897 : 200). Диаметр зрелых икринок обычно 5.5—6.0 см (Северное и Баренцево моря, Ново-Шотландские банки), реже до 6.5 см, в Баренцевом море иногда 7.0 мм (Wollebaek, 1924 : 210; Книпович, 1926 : 94; собственные наблюдения). Жировая капля одна, диаметром 1.75 мм, постоянно находится на верхнем полюсе икринки. Оболочка икринки довольно толстая, 36—41 μ, очень плотная, пронизана тонкими канальцами; к периоду вылупления становится менее прочной (McIntosh a. Prince, 1890 : 672). Выметанные икринки склеиваются так крепко, что их лишь с трудом можно оторвать друг от друга; в кладке они сохраняют шарообразную форму, вследствие чего между икринками остаются промежутки, необходимые для аэрации. Кладка имеет форму довольно правильного шара различных размеров (от кулака до футбольного мяча), выметывается на дно, среди камней и водорослей. Глубины различны: у берегов Шотландии кладка (назовем ее кладка № 1) была найдена на глубине 9—11 м (McIntosh a. Prince, 1890 : 672), у Мурманского побережья (кладка № 2) — на глубине 78—84 м (данные ПИНРО).
Кладка № 2 была ошибочно определена как кладка Anarhichas minor, очевидно, из-за больших размеров (с футбольный мяч). Однако сравнение числа лучей в плавниках у вылупившихся из ее икринок эмбрионов с числом лучей у взрослых Anarhichas lupus, Anarhichas minor и Anarhichas latifrons с несомненностью доказывает, что кладка принадлежит Anarhichas lupus (табл. 23). Вряд ли можно сомневаться в том, что у зубаток кладка состоит из икры, выметанной лишь одной самкой. Колебания плодовитости самок у Anarhichas lupus и Anarhichas minor сильно перекрываются, следовательно, поскольку диаметр икринок примерно одинаков, должны перекрываться и колебания размеров кладок. До сих пор это почему-то не учитывалось при неоднократных попытках определить видовую принадлежность кладок по их размерам.

Кройер (Krøyer, 1838 : 380) установил, что икринки Anarhichas lupus в Северном море в январе не больше макового зерна, и лишь через несколько месяцев достигают 2''' (5 мм). До работы Макинтоша и Принца (McIntosh a. Prince, 1890 : 672—673) считалось, что нерест происходит весной, в мае-июне (Fabricius, 1780 : 139; Bloch, 1784 : 22; Mitchill, 1818 : 242; Faber, 1829 : 75, и др.). В результате наблюдений над созреванием гонад у полосатой зубатки Макинтош и Принц пришли к выводу, что она нерестится в Северном море предположительно с ноября по январь. Но, по всей вероятности, сроки нереста здесь колеблются еще сильнее. Так, зрелая самка с диаметром икринок 6 мм была обнаружена в августе 1904 г. (Fulton, 1905 : 252), а вместе с ней еще несколько с диаметром икринок 3—4 мм. Самки с такими икринками довольно близки к переходу в очень непродолжительную IV стадию зрелости, за которой следует нерест. Две августовских самки, исследованные Макинтошем и Принцем (McIntosh, 1885 : 432; McIntosh a. Prince, 1890 : 874) имели такой же диаметр икринок. Почти зрелая самка была найдена в августе 1898 г. (Ewart a. Fulton, 1889: 197, 212), одна зрелая в июле и две в сентябре 1889 г. (Fulton, 1890:269). Учитывая небольшое общее число исследованных самок, нерест в августе и сентябре никоим образом нельзя признать исключением, как это делает Гилл (Gill, 1911 : 176). С другой стороны, зрелая самка с диаметром икринок 5 мм была найдена в феврале (McIntosh, 1885 : 472). Макинтош и Принц (McIntosh a. Prince, 1890 : 672) считают этот случай патологическим, однако в феврале же были обнаружены еще три зрелых и одна почти зрелая самка (Ewart a. Fulton, 1889 : 197, 212). Очевидно, сроки неоеста Anarhichas lupus в Северном море очень растянуты.
У побережья Исландии наибольшие икринки (5.0—5.5 мм) в яичниках Anarhichas lupus lupus наблюдаются в августе (Saemundsson, 1949 : 32; других сроков Семундсон не дает). Казалось бы, поэтому нерест у берегов Исландии растянут гораздо меньше, чем в Северном море, и происходит в августе-сентябре, а не в январе-феврале, как предполагает Семундсон по встречаемости молоди. Однако и для Исландии имеются указания (как и в отношении Anarhichas minor, без описания материала) на нерест Anarhichas lupus lupus в зимние месяцы (Lühmann, 1954b : 310), а именно с ноября по февраль, с максимумом в декабре-январе. Наблюдения эти основаны, по-видимому, на траловом лове. Траулеры же облавливают большие глубины обитания Anarhichas lupus lupus, на которых могли держаться запоздавшие с нерестом особи. Не исключена возможность, что начало и разгар нереста у основной массы исландских особей Anarhichas lupus lupus наблюдается раньше.
Очень немногочисленные наблюдения над состоянием гонад у баренцевоморской Anarhichas lupus lupus дают возможность полагать, что нерест происходит здесь примерно в июле—сентябре. 20 IV добыта самка III стадии с довольно мелкими икринками (экспонат из музея ПИНРО). Конец мая 1951 г.: одна самка II—III стадии, две в III стадии (диаметр икринок 3.0—3.5 мм, вес яичников 3.7% веса тела), одна в VI стадии с двумя остаточными икринками диаметром 7 мм (остались от прошлого года?). 21—25 VII: две самки III стадии с крупными икринками, одна в III—IV стадии (экспонат из музея ПИНРО). Сентябрь 1950 г.: одна самка III стадии, близкая к переходу в IV, две — в VI—II стадии.
У берегов Шотландии кладку зубатки (№ 1) выловили тралом 16 I 1886; внутри икринок находились эмбрионы, часто и сильно двигавшиеся. Через несколько дней после помещения кладки в аквариум они вылупились, но были недоразвиты (см. ниже). В Восточно-Прибрежном районе Баренцева моря кладку (№ 2) выловили тралом 13 II 1950, другую (№ 3), надо полагать, ошибочно считаемую кладкой Anarhichas minor — 10 IV 1931 (Schnakenbeck, 1933: 191). Из обеих кладок личинки начали вылупляться сразу после того, как они были подняты из воды. Личинки вылупляются хвостом вперед (McIntosh a. Prince, 1890 : 875; McKenzie a. Homans, 1938 : 280; собственные наблюдения). На Ново-Шотландских банках кладку с вылупляющимися эмбрионами нашли 19 II 1937 (Mc-Kenzie a. Homans, 1938 : 279). Во всех этих районах нерест Anarhichas minor весьма мало вероятен, так как здесь совершенно не встречается ее молодь меньших размеров, к тому же и взрослые особи, за исключением Восточно-Прибрежного района Баренцева моря, очень редки. Очевидно, все упомянутые выше кладки принадлежат Anarhichas lupus.
Личинки и мальки зубаток размером 17—25 мм встречаются в Северном море, у берегов Исландии и в заливе Мэн с января по март, в Баренцевом море и в заливе Св. Лаврентия с февраля по июнь-июль (Faber, 1829 : 75; Cuvier et Valenciennes, 1836 : 473; Fulton, 1890 : 269; McIntosh a. Masterman, 1897 : 202; Dannevig, 1919 : 16; Bigelow a. Welsch, 1925 : 375; Pacc, 1936 : 263; Перцева, 1939 : 260; Saemundsson, 1949 : 32). Лишь однажды личинка зубатки была поймана в ноябре (Collett, 1879 : 58).
Как видно из вышеизложенного, личинки и мальки зубаток с июля по ноябрь нигде не встречаются, а с ноября по январь весьма редки. Точно так же в тех районах, где нерест других видов зубаток, кроме Anarhichas lupus, почти совершенно исключен, ранее января-февраля не найдены и кладки икры с развивающимися или вылупляющимися эмбрионами. Между тем, судя по состоянию гонад, Anarhichas lupus нерестится в этих районах, по крайней мере частью, уже в июле-августе. Все это позволяет предполагать длительный инкубационный период в несколько месяцев, к чему склоняются также Бигелоу и Велш (Bigelow a. Welsch, 1925 : 374), Бигелоу и Шредер (Bigelow a. Schroeder, 1953 : 505).
Минимальные размеры личинок, ловившихся у берегов Шотландии, 17 мм, у берегов Исландии — примерно 18 мм, в заливе Мэн — 21 мм, в Мотовском заливе (Баренцево море) — 22 мм (во всех этих районах, кроме Исландии, нерест других видов зубаток весьма мало вероятен). Нигде не встречены вылупившиеся в море личинки, которые имели бы длину 11—12 мм и большой желточный мешок. Однако именно такими были личинки Anarhichas lupus, вылупившиеся в аквариуме из кладки № 1. Их желточный мешок значительно превышал размеры головы (рис. 39, А), лишь через две недели он стал несколько меньше головы, а совершенно рассосался только к началу мая — середине июня. Личинки из кладки с Ново-Шотландских банок длиной 17—18 мм также несли большой желточный мешок. Между тем личинки из кладки № 2 имели длину 24—25 мм, небольшой желточный мешок и были вообще гораздо более сформированы (рис. 39, Б). Отсюда следует, что личинки Anarhichas lupus lupus вылупляются в море значительно более крупными и более развитыми, чем в аквариуме, как это предполагали и сами наблюдатели (McIntosh a. Prince, 1890 : 875); возможно, и в море вылупление происходит на разных стадиях развития, в зависимости от условий среды. Между прочим, личинки, вылупившиеся на палубе (кладка № 2), также могли быть недоразвитыми, хотя и в гораздо меньшей степени, чем вылупившиеся в аквариуме.
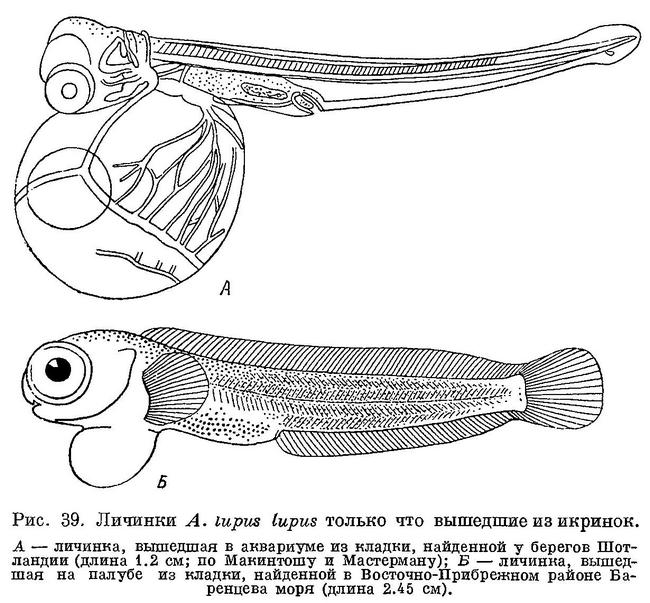
Личинок из кладки № 3 (см. выше) Шнакенбек причислил к Anarhichas minor. К этому его побудила большая величина (22 мм) и сформированность личинок, в частности отсутствие у них желточных мешков. Однако, как выясняется, вылупляющиеся личинки Anarhichas lupus могут иметь еще большую длину и быть не менее сформированными. Что же касается желточных мешков, то они могли просто оторваться (происходит это очень легко, как я мог убедиться на личинках из кладки № 2). Это тем более вероятно, что, судя по приведенному рисунку, личинки были доставлены Шнакенбеку в плохом состоянии. Впрочем, Шнакенбек и сам не был уверен в своем определении.
Приводимое Рассом (1939) различие в размерах между личинками Anarhichas lupus (17 мм) и личинками Anarhichas minor (22—23 мм) есть различие между наименьшими из личинок Anarhichas lupus, известных у берегов Шотландии (McIntosh a. Masterman, 1897 : 202—205) и личинками Шнакенбека, по-видимому, тоже принадлежащими Anarhichas lupus. Что же касается личинок Anarhichas minor, то они по существу неизвестны (поскольку ни один из авторов не приводит более никаких признаков, отличающих их от личинок Anarhichas lupus), как неизвестны и личинки Anarhichas latifrons.
Интересно поведение личинок, вылупившихся в аквариуме (кладка № 1). Сразу после вылупления начали работать грудные плавники, и первое время они находились в постоянном движении. Пока желточный мешок был большой, личинки лишь временами подпрыгивали извивающимся движением на несколько дюймов от дна и вновь садились обратно. С рассасыванием желточного мешка становились подвижнее и плавали более устойчиво. После того как мешок рассосался, стали много плавать, значительно окрепли и, по-видимому, мало страдали, оставаясь несколько дней в очень небольшом объеме морской воды.
Личинки и мальки зубаток длиной 17—44 мм изредка ловятся и у дна и вблизи поверхности. В Северном море личинка длиной 17 мм с рассосавшимся желточным мешком была выловлена 21 I у дна (McIntosh a. Masterman, 1897 : 202), другая, длиной 22 мм, 21 II у поверхности (Fulton, 1890 : 269). В заливе Мэн в толще воды было добыто около 20 личинок длиной 21—44 мм, причем из восьми уловов только два были у поверхности, тогда как остальные на глубине 54—108 м (Bigelow a. Welsh, 1925 : 375). Бигелоу и Велш пришли к выводу, что пелагическая стадия у личинок Anarhichas lupus коротка, протекает в основном в толще воды у дна и личинки не подвержены длительным пассивным миграциям; позднее Бигелоу и Шредер (Bigelow a. Schroeder, 1953 : 506) придерживались того же самого мнения. Милинский (1944) считает, что после рассасывания желточного мешка личинки Anarhichas lupus питаются населяющими морское дно мелкими организмами. На свету они интенсивно темнеют (McIntosh, 1890 : 288; по словам А. С. Бараненковой, окраска добытых в толще воды мальков зубаток была темной, а у одного почти черной), что, казалось бы, естественно для донных личинок и мальков и мало подходит для пелагических. Назначение серебристого оттенка на боках мальков длиной 3—4 см остается неясным. По всей вероятности, личинки Anarhichas lupus живут обычно на дне, поднимаясь зачастую в придонные слои воды, а иногда и к самой поверхности. Привязанность к дну в дальнейшем становится, очевидно, сильнее: молодь длиной более 4.4 см в толще воды никем еще не поймана. Сеголетки длиной 5—7 см и больше ловятся в промысловые тралы и в южной части Баренцева моря, и в районе Лофотенских островов, и на Медвежинской банке. Судя по их распределению, а также по находкам кладок и личинок (см. выше), Anarhichas lupus нерестится во всех этих районах, в отличие от Anarhichas minor и Anarhichas latifrons, которые в южной части моря и у Лофотенских островов вряд ли нерестятся (Барсуков, 1957 : 36—37, 42). Кладки зубаток, найденные в южной части моря — в Восточно-Прибрежном районе, на Финмаркенской банке, в Мотовском заливе, могут принадлежать Anarhichas lupus, хотя и приписываются Есиповым (1949 : 614) пятнистой зубатке.
В кишечниках Anarhichas lupus lupus в Баренцевом море найдены брюхоногие и двустворчатые моллюски — Buccinum undatum, Neptunea despecta, Astarte sp., Pecten islandicus sp., Сardium cilliatum и др., раки-отшельники — Pagurus pubescens, крабы — Lithodes, Hyas, креветки — Crangon, Pandalus и др., морские звезды — Solaster papposus, Solaster endeca, Ctenodiscus crispatus, морские ежи, офиуры — Ophiopholis aculeata, Ophiura sarsi, полихета Nereis, актинии, голотурии, Balanus sp., мшанки, капшак, рыба — мелкая треска, Gadus esmarki и отбросы с траулеров. В Северном море: моллюски — Buccinum undatum, Natica catena, Littorina littorea, Nassa incrassata, Triton fuscus, Fusus antiquus, Fusus gracilis, Pecten opercularis, Pecten tigrinus, Cardium echinatum, Trochi sp., Mya sp., Modiola modiolus, Mytilus edulis, Astarte compressa, Solen siliqua, Venus lincta, Echinocardium sp., ракообразные — Pagurus sp., Hyas arenaria, Hyas coarctatus, Lithodes maja, Stenorynchus rostratus, Nephrops sp., Portunus sp., Crangon sp., Galathea sp., Cancer pagurus, Eupagurus bernhardus, Atelecyclus septendentatus, иглокожие — Echinus esculentus, Ophiura ciliaris, Ophiocoma sp., Ophioglipha sp., Ophiotrix fragilis, черви — Aphrodite aculeata, рыба (сельдь). У берегов Исландии: моллюски — Modiolus papuanus, Cyprina islandica, Pecten islandicus, Mytilus major, Cardium ciliatum, Macoma calcarea, крабы — Hyas aranea, морские звезды — Asterias papposus, морские ежи, рыба (песчанка). У берегов Гренландии: моллюски — Mytilus и др., ракообразные, икра рыб. У берегов Северной Америки: моллюски, двустворчатые и брюхоногие — Buccinum undatum, Lunatia sp., Chrysodomus sp., Fusus sp., Mactra sp., Polynices sp., Sipho sp. (предпочтение отдается брюхоногим, — Bigelow a. Schroeder, 1953 : 505); раки-отшельники, крабы, морские ежи — Euryechinus droebachiensis, морские звезды. Всюду преобладают моллюски, притом с более прочной раковиной, чем используемые Anarhichas minor. Очень обычны ракообразные, особенно раки-отшельники и крабы. Нередки иглокожие, но их роль в питании гораздо меньше, чем у двух предыдущих видов зубаток. Реже, чем у последних, встречается рыба и отбросы с траулеров. У берегов Северной Америки Anarhichas lupus чаще ловится удочкой, наживленной моллюсками,чем на яруса, наживленные рыбой, а именно сельдью (Bigelow a. Schroeder, 1953 : 505). И в Баренцевом море на яруса попадает реже, чем два предыдущих вида.
Вес печени в мае у самки I стадии 1.6, у самца 2.5, у самки VI стадии 2.6, у самки III стадии 4.1% веса тела (собственные наблюдения). В июне вес печени у 3 экз. Anarhichas lupus lupus длиной 41—51 см равнялся 1.57—1.72% веса тела (Баренцево море; Kittel u. Kampfe, 1952—1953 : 474).
Темп роста в Баренцевом море, по данным Сахно (Маслов, 1944 : 155), дополненным мной, ниже, чем у других видов (табл. 24).

В Баренцевом море перед 1941 г. встречалась до 17-годовалого возраста, преобладали в уловах 3—8-годовалые (по Милинскому, 1944); в 1928— 1931 гг. — до 18-годовалого, преобладали 7—15-годовалые. Тогда же у берегов Исландии встречалась до 20-годовалого возраста, преобладали 7—15-годовалые, в Северном море — до 18-годовалого, преобладали 7— 12-годовалые (Lundbeck, 1936, tab. 19).
Макинтош и Мастерман (McIntosh a. Masterman, 1897 : 205) считают, что молодь Anarhichas lupus через 6—7 месяцев после вылупления вырастает примерно до 6 дюймов (15 см), в следующий февраль до 8.5—10 дюймов (21— 25 см), а на второй год до 18—20 дюймов (46—51 см) — темп роста необычайно быстрый. К сожалению, авторы не указывают, что послужило материалом для данного сообщения. Возможно, это было наблюдение над ростом личинок в аквариуме и речь идет не о дюймах, а о линиях, — тогда мы имеем дело с чрезвычайно замедленным ростом.
Молодь Anarhichas lupus lupus иногда встречается в желудках трески.
Полянский (1955 : 117) обнаружил у баренцевоморских Anarhichas lupus lupus (15 экз.) следующих паразитов: Prosorhynchus squamatus (кишечник), Plagioporus idonea (кишечник), Podocotyle atomon (кишечник), Neophasis lageniformis (кишечник), Steringophorus furciger (кишечник), Fellodistomum fellis (желчный пузырь), Fellodistomum agnotum (желчный пузырь, кишечник), Zoogonoides viviparus (кишечник), Diphterostomum microacetabulum (кишечник), Lepidophyllum steenstrupii (мочевой пузырь), Steganoderma spinosa (кишечник), Bothriocephalus scorpii (кишечник), Ascarophis sp. (кишечник), Contracoecum aduncum (желудок, кишечник), Anisakis sp. larvae (поверхность печени, полость тела, мезентерий), Echinorhynchus gadi (кишечник), Platybdella anarrhichae (плавник, кожа) и Clavellodes rugosa (жабры). Описано также поражение зубов микроорганизмом «Myceliles ossifragus», {Schmidt, 1954; поражение, по-видимому, такого же рода встречено мной у баренцевоморской Anarhichas lupus длиной 105 см). При этом зубы, пронизанные ходами паразита, теряют свою естественную окраску, становятся белыми, как мел, и быстро снашиваются. Особенно велика интенсивность заражения паразитом Neophasis lageniformis, при довольно большом проценте (53.3). Зараженность паразитом Fellodistomum fellis — стопроцентная, что отмечает также Базикалова (1932 : 143). Интенсивность заражения велика. Это указывает на значительную роль промежуточного хозяина данного паразита — Ophiura sarsi — в питании баренцевоморской полосатой зубатки.
Хозяйственное значение. Уловы полосатой зубатки в Баренцевом море значительно уступают уловам пятнистой и синей зубаток. Полосатая зубатка добывается главным образом в Северном море, у Фарерских островов, у побережий Исландии и Северной Америки (залив Мэн). В Северном море наибольшие уловы наблюдаются в мае (а в некоторых участках нередко в марте-апреле, или, напротив, позднее, в июне и даже июле); затем вплоть до декабря уловы снижаются, а с января вновь начинают возрастать (McIntosh, 1899 : 174—179; Thompson, 1905 : 344—346, 1908 : 272—274). По Томпсону, у Фарерских островов наиболее многочисленна с февраля по июнь или июль, у Исландии же наблюдается два максимума уловов — в апреле и сентябре. Мясо вкусное, употребляется в пищу в соленом, копченом и свежем виде. Содержит 75—82% влаги, 3—7% жира, 14—18% белка (Травин, 1952 : 175). Как и у пятнистой зубатки, жир концентрируется в основном в печени и костях. Кроме мяса используется икра и шкура.
Дополнение к разделу «Anarhichas lupus lupus». У побережья Лабрадора Anarhichas lupus lupus встречена сотрудниками ПИНРО до 60°14' с.ш. и 61°50' з. д. Обычна вдоль всего западного побережья Шпицбергена: по тем же данным на 79°50' с. ш. и 10°30' в. д. было выловлено 14 экз. Anarhichas lupus lupus длиной 24—64 см (сентябрь 1958 г., глубина 350—324 м). Экземпляр Anarhichas lupus lupus в августе 1956 г. был найден на берегу пролива Югорский Шар; ранее местные рыбаки зубаток здесь никогда не видели (В. Д. Лебедев, Зоолог. журн., XXXVIII, 4, 1959 : 636).
В марте 1959 г. самки III стадии не встречались. Пелагические мальки, по данным А. С. Бараненковой и Н. С. Хохлиной, ловились в Баренцевом море в апреле-июле (длина 19—33 мм), в основном в тех же местах, где и придонная молодь длиной 7 см и более: распределение последней в 1951—1958 гг. мало чем отличалось от ее распределения в 1941—1950 гг., как и у придонной молоди Anarhichas minor. На западном и южном склонах Медвежинской банки и в Зюйдкапском желобе пелагические мальки Anarhichas lupus lupus пойманы в июне-сентябре (длина 24—45 мм), у побережья Исландии — в мае-июне (длина 28—30 мм). Малек длиной 47 мм выловлен в открытом океане 24 VI 1955 г. (74°30' с. ш. и 3°25' в. д. ).
4б. Anarhichas lupus maris-albi Barsukov — Беломорская зубатка, на Белом море — синяя зубатка.
(Maris-albi — беломорский.)
Anarrhicas minor (non Olafsen) Levander, Meddel. Soc. Fauna Flora Fennica, XV, 1888—1889 : 231 (2 экз. у Соловецких островов).
Anarrhichas lupus + Anarrhichas minor Knipowitsch, Ежег. Зоолог. муз. Акад. наук, II, 1897 : 148 (Белое море). — Шульман-Альбова, Учен. зап. Карело-Финск. унив., IV, 3, 1952 : 82, 84—86, 88, 91—93 (паразиты).
Anarchichas lupus + Anarhichas minor Зборовская, Тр. Карело-Финск. отд. Всесоюзн. научно-исслед. инст. озерн. речн. рыбн. хоз., III, 1951 : 197—210 (Гридинская губа, с мая по сентябрь; длина не более 50 см).
Anarrhichas lupus Дерюгин, Фауна Белого моря, 1928 : 342 (1 экз. с тони у Соловков).
Anarhichas lupus Шульман и Шульман-Альбова, Паразиты рыб Белого моря, 1953 : 9, 15, 22, 41, 45—48, 56—58, 67, 68, 71—74, 76, 83, 84, 131—132, 178—179. — Барсуков, Зоолог. журн., XXXII, 6, 1953 : 1211—1216 (размножение).
Anarhichas lupus maris-albi Барсуков, Докл. АН СССР, XCV, 4, 1954 : 897— 899 (смена зубов). — Барсуков, Вопр. ихт., 6, 1956 : 129—135 (характеристика, подвида).
D 71—75, А 43—46 (48), Р 18—21, С 23—26, позвонков 72—76 (туловищных 24—27, хвостовых 46—51), лучей жаберной перепонки 6—7, окологлазничных косточек, с praeorbitale, (5) 6—7. Нижняя челюсть спереди утолщена. Зубы массивнее, чем у Anarhichas lupus lupus таких же размеров. Зубов на обеих нёбных костях 8—16, в среднем около 12 (у особей длиной более 20 см). Всех зубов на межчелюстных, нижнечелюстных, нёбных костях и сошнике 51—72, в среднем около 60. Зубов на глоточных костях 82—105, в среднем 92 (3 экз.). Posttemporale изогнута только внутрь, но не вниз, расположена на одной линии с передним краем supracleithrum при виде сбоку. У молоди длиной 6—7 см зубы на нёбных костях и сошнике располагаются в основном в один ряд, и лишь на передних частях костей группируются в два, а изредка и в три ряда; сошниковый ряд зубов не простирается назад далее нёбных рядов или лишь незначительно выдается за них.
Желтоватых и голубоватых тонов в общем фоне окраски нет. У молоди между полосами пятна того же цвета, что и полосы; мелкие пятна, более темные, чем полосы, расположенные на спине и на основании спинного плавника, встречаются чаще, чем у предыдущего подвида.
В уловах ставных ловушек встречаются особи от 21 см и выше; преобладают размеры от 29 до 41 см. Длина самцов до 66 см, самок — до 64 см, вес, соответственно, до 2.3 и 1.8 кг. Средний размер 36.7 см: самцов 37.0, самок 36.3 см. Средний вес 0.4 кг.
Описание дано по 2000 экз. с Карельского берега Белого моря (дер. Летняя речка), исследованных летом 1951 г.; лучи плавников и позвонки подсчитаны у 30—60 экз., число зубов у 35—36 экз.; отпрепарировано 80 черепов.
Распространение. Карельский берег Белого моря, от станции Кемь и Соловецких островов примерно до Кандалакши.
Биология. Держится у самых берегов, на глубине в несколько метров, со второй половины мая, когда побережье очищается от льда, до осенних заморозков, т. е. примерно до конца сентября. С мая по сентябрь уловы ее здесь снижаются: по-видимому, часть особей уходит на глубины по мере их прогревания. Максимальные глубины, на которых встречается, неизвестны; можно предполагать, что обычно не опускается ниже слоя воды с положительной температурой. Места зимовок неизвестны.
Предпочитает каменистые грунты с зарослями бурых водорослей. На илистых салмах, заросших зостерой, не встречается.
По словам рыбаков, у берегов активна только ночью, днем отстаивается в укрытиях у дна. Но вряд ли это полностью справедливо и для периода коротких ночей.
У самок I стадия зрелости длится до 5 лет, II — примерно от 2 до 6 лет (до первого нереста), III — от 1 до 3 месяцев, III—IV и IV — по-видимому, лишь несколько дней, VI — предположительно, 1—2 месяца. Отношение веса гонад к весу тела на I стадии около 0.2%, на I—II — 0.13—0.35, на II — 0.3—1.0, на II—III — 0.4—3.0, на III — 2.2—18.5, на III—IV — 9.0—12.5 (2 экз.), на VI — 0.9—2.0, на VI—II — 0.9—1.5%. Размер мелких белых икринок, встречающихся в яичниках на всех стадиях зрелости, начиная со II (следовательно, и после первого нереста их развитие длится не менее 2 лет), — 0.4—1.7 мм. У самок, нерестящихся в данном году, примерно вторая часть икринок начинает с весны увеличиваться; при диаметре их около 2.2 мм становится ясно различимой III стадия зрелости. В течение III стадии зрелости все эти икринки увеличиваются почти одинаково, сохраняя очень большое сходство в размерах. Переход из III в IV стадию у разных самок происходит при различном диаметре указанных икринок (от 3.5 до 5.5 мм). Диаметр зрелых икринок 5.7—6.5 мм. Жировая капля одна, диаметром 1.75 мм.
Вес гонад у самцов 0.03—0.27, в среднем 0.14% веса тела.
Длина впервые нерестящихся самок колеблется от 30 до 44 см, вес от 195 до 690 г, возраст — от 5 до 11 лет. При длине тела 37 см и возрасте 8+ зрелые самки по количеству начинают преобладать над незрелыми. Большая часть самцов, по-видимому, созревает несколько раньше самок, при длине 34—36 см.
Показательно, что уловы заколов, ставящихся всегда у самых берегов, состоят в основном из созревающих самок; самок I стадии по сравнению с ними очень немного. Прогрев воды у берегов наибольший. Следовательно, развитие яичников, начиная со II стадии, происходит при высоких температурах и, надо думать, для него требуются такие температуры.
Плодовитость колеблется от 260 (Длина рыбы 40 см. В яичниках этой особи было много дегенерирующих икринок. Наименьшее число икринок у самки с нормальными яичниками 650 (длина 30 см).) до 7000 (64 см) икринок и, как обычно, в среднем возрастает с размерами самок. Средняя плодовитость, с учетом размерного состава зрелых самок, 2260 икринок. Во второй половине июля крупные самки III стадии имели в среднем меньшую плодовитость, чем во второй половине июня (рис. 40), причем у самок, близких к переходу в IV стадию, иногда наблюдалось чрезвычайно резкое истощение при нормально развитых яичниках, или же при нормальном весе тела большая часть икринок дегенерировала. Отставали в развитии гонад, по-видимому, хуже питавшиеся самки. Так, самки III стадии в конце июня по среднему весу тела (без гонад) примерно на 10% превышали самок II стадии той же длины, а по среднему весу печени — приблизительно на 50%. К осени вес тела у не метавших икру самок II стадии повысился, у нерестовавших, напротив, снизился, как и вес печени (Барсуков, 1953 : 1214).

Наблюдения над состоянием яичников у беломорской зубатки показали, что основная масса самок отнерестилась в 1951 г. в июле — начале августа; лишь немногие отметали икру раньше, в июне. Очевидно, нерест происходил на некотором удалении от берегов, на несколько больших глубинах, чем те, на которых стоят ловушки. Об этом можно судить по очень редкой встречаемости в уловах самок III—IV и IV стадий, а также по снижению в течение июля количества самок III стадии относительно незрелых вплоть до полного исчезновения в начале августа. Основные сроки нереста мной относились ранее (Барсуков, 1953 : 1211) к концу июля—началу августа. Указанный факт снижения в уловах зрелых особей относительно незрелых в течение всего июля был недооценен из-за незначительной встречаемости в это время самок VI стадии. Однако они могли подойти к беретам только во второй половине августа вместе с позднее отнерестившимися самками. Судя по снижению среднего веса семенников, часть зрелых самцов в июле отходила от берегов вместе с самками (у самцов длиной 25—30 см 18 VI—3 VII семенники и среднем составляли 0.12% от веса тела, 5—25 VII — 0.10%, 17—30 VIII — 0.09%; у самцов длиной 30—37 см, соответственно, — 0.15, 0.13 и 0.10%, длиной 37—44 см — 0.17, 0.16 и 0.13%, 44—65 см — 0.15 и 0.13%). В отличие от пинагора, не наблюдалось ни одного случая вымета икры в ловушке. Текучие самки не попадались.
Отнерестившиеся самки во второй половине августа вновь появились у берегов. Самки II стадии, наоборот, к этому времени стали ловиться в совсем малом числе. У 10% из них были обнаружены оставшиеся после нереста, крупные икринки, обычно не более 2—3. У самок, больших по размерам, такие икринки встречались чаще. Вес гонад был, напротив, меньше, чем у мелких самок (табл. 25), притом большая часть крупных самок находилась не в VI, а в VI—II стадии зрелости. По-видимому, крупные самки в общем отнерестились раньше мелких.

В это время у самок II—III стадии более крупные икринки приобрели неправильную форму, а оранжевый пигмент сконцентрировался в центре икринки. Начавшееся было развитие икринок прекратилось, по-видимому, в связи с наступлением заморозков, вызвавшим понижение температуры воды. На дальнейший этап развития яичников последнее, по всей вероятности, уже не оказывает такого резкого влияния: у самки III стадии зрелости (23 VIII; длина 46 см, вес гонад 31.0 г), имевшей икринки диаметром 3.0 мм, признаков дегенерации икринок обнаружено не было. Отсутствие у берегов самок IV стадии и малое число самок III—IV стадии также может свидетельствовать о снижении потребности в высоких температурах в конце преднерестового развития гонад.
В отличие от самок, большая часть взрослых самцов после нереста к берегам не возвратилась: в июне и июле самцы по количеству несколько преобладали над самками, в августе же наблюдалось резкое преобладание самок. Возможно, самцы остались охранять икру.
Нерест ежегоден, судя по тому, что самки II и II—III стадии не встречались среди особей длиной более 44 см (113 экз.), в возрасте более 11 лет. Самки I стадии отмечены лишь среди особей в возрасте менее 5 лет. Остается предположить, как это и было сделано выше, что II стадия у некоторых самок длится до 6 лет, поскольку неежегодность нереста молодых самок, в отличие от старых, мало вероятна. Впрочем, самки II и II—III стадии старшего возраста могли проходить в свое время I стадию в течение большего срока, чем самки II стадии младшего возраста, следовательно, II стадия у первых могла занять несколько меньшее время, чем это предполагается. Следует считаться с возможностью периодических колебаний в скорости созревания основной массы особей, охватывающих ряд лет.
Сеголетки длиной 6—7 см были пойманы в бимтрал в 1938 г. около Соловецких островов на глубине 32 м при температуре +7.5° и солености около 27‰. Молодь меньших размеров пока неизвестна.
В кишечниках беломорской зубатки из районов Летней Речки (собственные наблюдения) и Гридинского (данные Чудновой, см.: Шульман-Альбова, 1949) обнаружены двустворчатые моллюски — Mytilus edulis, Modiolus modiolus, Modiolaria discors, Cardium sp., брюхоногие — Littorina saxatilis, Margarita groenlandica, Acmaea testudinalis, Tellina edulis, Natica sp., Buccinum sp., крабы — Hyas araneus; иглокожие — Ophiura robusta и Ophiopholis, асцидии, рыба — навага, корюшка, камбала, сельдь, бычки, колюшка, молодь сига, молодь трески, бельдюга, зубатка, икра пинагора, фукус и зостера. Основные объекты питания — мидии и брюхоногие. Крабы часто встречаются в кишечниках, но почти всегда лишь по одному экземпляру. Асцидии играют небольшую роль в питании. Иглокожими в районе Летней Речки зубатка совсем не питалась. Рыбу и икру пинагора хватала большей частью, уже будучи в ловушке. Фукус и зостера, по-видимому, захватываются случайно вместе с пищей, чаще тоже в ловушке. С возрастом роль мидий в питании увеличивается, а мелких брюхоногих, особенно Acmaea, уменьшается. Будучи в ловушке, крупные зубатки чаще пожирают рыбу, чем мелкие.
В течение всего периода наблюдений питание зубаток обоих полов при всех стадиях зрелости было интенсивным. Вряд ли можно сомневаться
в том, что нагульный период у беломорской зубатки значительно короче, чем у баренцевоморской. К тому же у взрослых особей беломорской зубатки преднерестовое развитие гонад, угнетающее рост, растягивается на большую часть указанного периода. Вероятно, поэтому беломорская зубатка сильно отстает в темпе роста (табл. 26).

В 1951 г. в уловах преобладали особи в возрасте 6+.
Шульман и Шульман-Альбова (1953 : 132—133) обнаружили у беломорской зубатки следующих паразитов: Myxoproteus elongatus (мочевой пузырь), Mixidium incurvatum (желчный пузырь), Podocotyle atomon {кишечник), Neophasis lageniformis (кишечник), Diphterostomum microacetabulum (задняя кишка), Zoogonoides viviparus (задняя кишка), Lepidophyllum steenstrupii (мочевой пузырь), Fellodistomum fellis (желчный пузырь), Steringophorus furciger (кишечник), Lecithaster gibbosus (кишечник), Lecithaster confusus (кишечник), Derogenes crassus (кишечник), Genarches mülleri (кишечник), Tocotrema sp. larvae (жабры), Pseudophyllidea gen. sp. larvae I (стенка кишечника), Sсоlex polymorphus (кишечник), Anisakis sp. larvae (полость тела), Terranova decipiens larvae (полость тела), Contracoecum aduncum (кишечник), Contracoecum aduncum larvae (полость тела), Echinorhynchus gadi (кишечник), Corynosoma semerne (полость тела). Наиболее распространенными паразитами являются Diphterostomum microacetabulum и Neophasis lageniformis (степень зараженности до 3000 экз.), связанные с питанием моллюсками. Высокий процент зараженности (46.6—49.9) дает кишечная форма Contracoecum aduncum, указывающая на питание рыбой, но степень зараженности не очень велика. Fellodistomum fellis, связанный с питанием иглокожими (Ophiura sarsi), дает большой процент зараженности (86.6) в Чернореченской губе и малый в Гридинской (10.2). Степень зараженности мала (1—27 паразитов в Чернореченской губе и 1—6 в Гридинской), что указывает на значительно меньшую роль Ophiura sarsi в питании Anarhichas lupus maris-albi по сравнению с Anarhichas lupus lupus. Остальные паразиты встречаются редко (кроме Terranova decipiens l.) и в малом числе.
Хозяйственное значение. Сведений об уловах нет, так как промысел имеет потребительское, чисто местное значение. Ловится вместе с другими рыбами в ставные ловушки со второй половины мая до сентября, причем уловы снижаются в течение всего этого периода (табл. 27).

В мае бьют острогой у берегов; этот промысел носит скорее всего любительский характер. Мясо и икра отличаются хорошим вкусом. На приемных пунктах засаливается. Шкура не используется.
У Anarhichas lupus lupus передние клыки еще длиннее, а разница между ними и малыми клыками еще больше; площадки сближены сильнее. Давящие крупнее, еще сильнее стираются к концу лета, как и другие зубы. В тех частях зубного аппарата, которые несут наибольшую нагрузку, зубы иногда стираются почти целиком и даже захватываются их цоколя (Lühmann, 1954b : 322 и собственные наблюдения). Складчатость их оснований увеличивается, а полость пульпы уменьшается по сравнению с Anarhichas minor (табл. VI, 3).

Наибольшие давящие нижней челюсти отодвигаются еще дальше назад, располагаясь всегда за пережимом (табл. II, 3). Соответственно удлиняются и ряды зубов на сошнике (табл. I, 3), простираясь теперь далее назад, чем зубы на нёбных костях. Ширина зубов сошника заметно нарастает спереди назад, вплоть до предпоследней пары. Пережим на нижнечелюстных костях развит еще сильнее, опускание нижнечелюстных давящих вниз и наклон их внутрь заметно больше, чем у Anarhichas minor. Зубы нёбных костей по отношению к клыкам нижнечелюстных отодвинуты еще дальше назад, а по отношению к коническим и давящим — кнаружи. Почти все зубы внешнего ряда на нёбных располагаются кнаружи от внешнего ряда на нижнечелюстных, а зубы внутреннего ряда — напротив или немного внутрь от внешнего ряда на нижнечелюстных. Внутренние зубы на нёбных и задние зубы внутреннего ряда на нижнечелюстных превращаются в давящие. Зубы внешнего ряда на нижнечелюстных, исключая клыки, хотя и закладываются как тупоконические, но очень скоро стираются под одним углом с давящими внутреннего ряда. От клыков они отделяются, в отличие от предыдущих видов, небольшим перерывом в области пережима: начинается оттеснение их назад, подобно зубам на нёбных костях. Зубы внешнего ряда на нёбных стираются значительно меньше, но все же сильно.
Зубной аппарат Anarhichas lupus lupus еще больше специализируется к питанию моллюсками, притом более крупными и прочными, чем используемые Anarhichas minor. Вследствие этого успешность питания не только подвижными организмами, но и иглокожими (морскими звездами и офиурами) уменьшается, так как «зубчатые ножницы» на боках рта редуцируются. Составляющие их зубы меняют рвущую функцию на дробящую, поддерживая основной механизм дробления — сошниковые зубы и внутренние ряды зубов нижней челюсти — в области наибольших давящих и сдвигаясь вместе с последними назад. Их побочная роль ограничителей, предупреждающих выскальзывание моллюска вбок, резко снижается, компенсируясь сильно развившимся наклоном нижнечелюстных давящих внутрь.
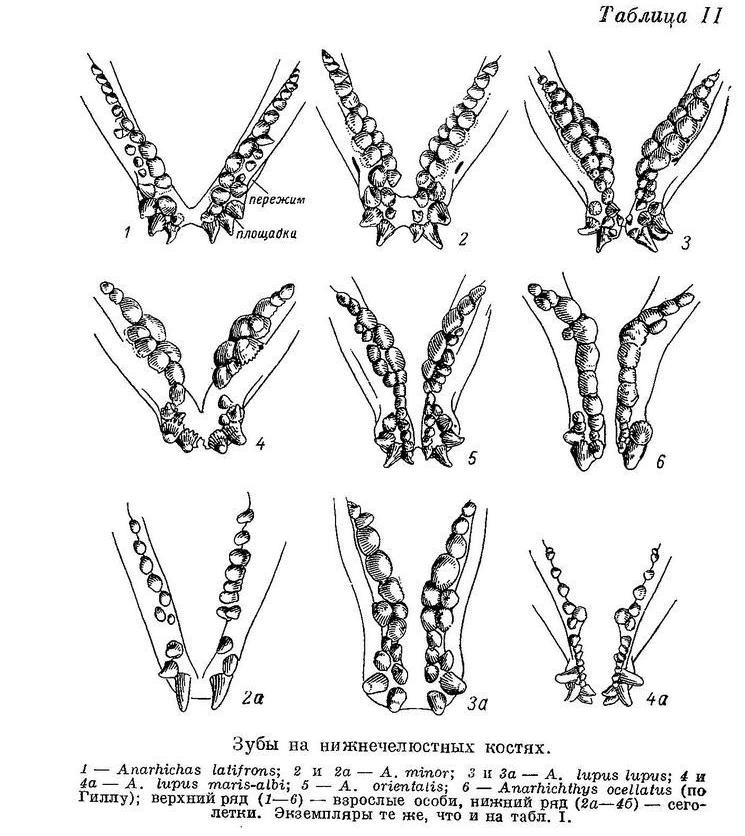
У Anarhichas lupus maris-albi расположение зубов в общем сходно с таковым у Anarhichas lupus lupus. Однако заметно дальнейшее возрастание прогиба внутреннего ряда нижнечелюстных давящих вниз и внутрь и наклона этих зубов внутрь. Большие клыки и давящие заметно шире (табл. II, 4). Складчатость на основаниях давящих усиливается, полость пульпы уменьшается (табл. VI, 4). Зубы на нёбных отодвигаются еще дальше от клыков. Специализация к питанию моллюсками у этого подвида выражена еще сильнее. Передние клыки на нижней челюсти меньше наклонены вперед, чем у Anarhichas lupus lupus: вероятно, беломорской зубатке чаще приходится брать пищу не со дна, а с боковой поверхности камней.
Не менее интересна возрастная изменчивость в строении зубной системы.
Личинки Anarhichas lupus lupus, выходящие из икры, имеют уже около 50 зубов (не считая глоточных). Все зубы острые. Внешний ряд нижнечелюстных зубов еще не продолжается за площадку (пережим уже имеется). На сошнике 4—5 зубов (табл. IV), из них передний иногда значительно крупнее остальных.
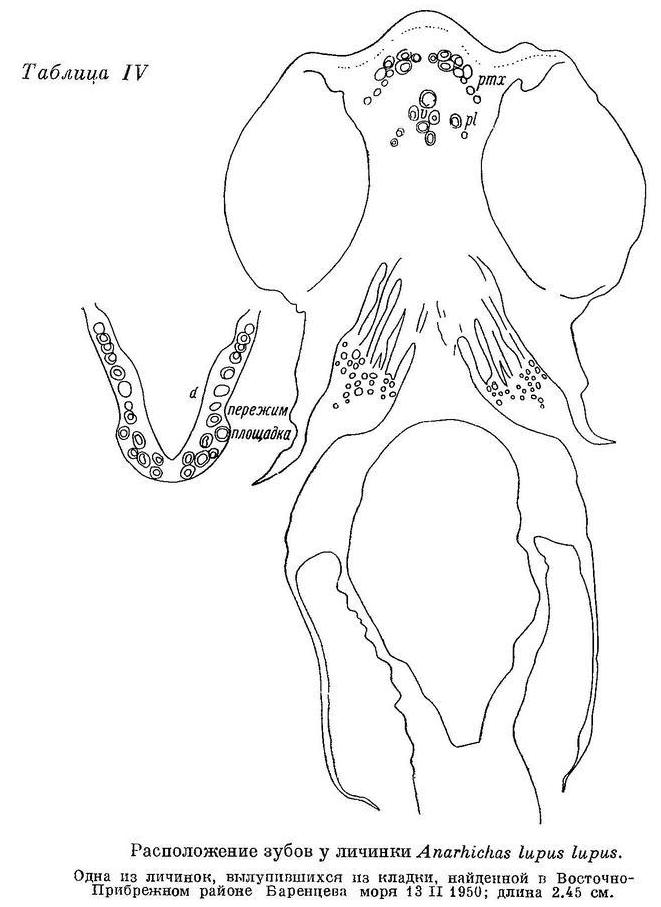
На нёбных костях всего по 2 очень маленьких зуба, расположенных на уровне задних зубов сошника. У сеголеток длиной 7 см зубы по форме и расположению близки к взрослым особям, но наибольшие нижнечелюстные давящие дальше сдвинуты назад (табл. II, 3а), а зубы сошника относительно зубов на нёбных развиты сильнее, чем у взрослых (табл. V, 3). Позднее давящие зубы на нёбных костях увеличиваются в числе и размерах несколько быстрее, чем на сошнике, принимая на себя большую долю работы при раздавливании, однако зубы сошника всегда, играют основную роль. Судя по строению зубов, у Anarhichas lupus lupus уже сеголетки питаются в основном моллюсками.
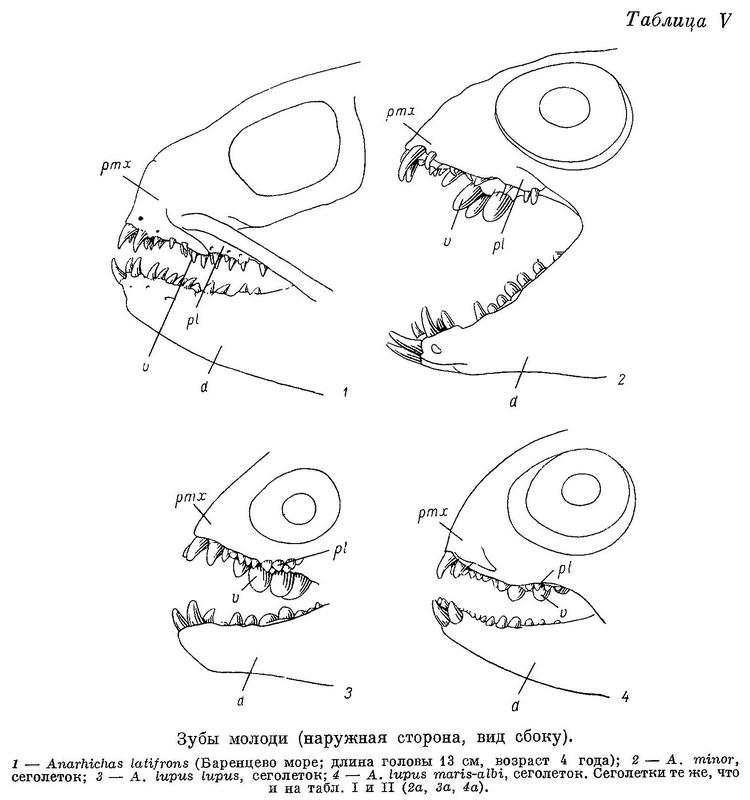
По существу то же самое можно сказать о сеголетках Anarhichas lupus maris-albi длиной 6—7 см (с той разницей, что по расположению зубов они стоят ближе к личинкам) и сеголетках Anarhichas orientalis длиной 15 см.
Окраска
У Anarhichas lupus lupus расположение и число поперечных полос такое же, как и у Anarhichas lupus maris-albi; они также состоят из сливающихся неясных пятен, у взрослых особей лучше заметных. Пятна на продолжении полос на спинном плавнике у молоди Anarhichas lupus lupus такие же, как и у молоди Anarhichas lupus maris-albi, исчезают в том же порядке, примерно при той же длине тела. Остальные плавники обычно без пятен и полос. Мелкие черные пятна на основании спинного плавника и на спине ниже него также бывают. Но общий фон окраски Anarhichas lupus lupus обычно несколько светлее, с желтовато-зеленоватыми или голубоватыми оттенками; полосы сильнее сливаются с общим фоном и часто даже у молоди плохо заметны. Пятен того же цвета, что и полосы, расположенных между полосами, не наблюдается.
Следует заметить, что окраска Anarhichas lupus maris-albi описана по рыбам, добывавшимся с глубины нескольких метров, тогда как окраска Anarhichas lupus lupus — по экземплярам, ловившимся на глубинах в несколько десятков и даже сотен метров. Требуется еще выяснить, не свойствены ли особям обоих подвидов сходные изменения окраски при переходе с мелководий на глубины и обратно.
Вариации окраски исландских Anarhichas lupus lupus и Anarhichas minor (Lühmann, 1954b: 313—315) весьма сходны с описанными выше вариациями окраски баренцевоморских.
Глаза. У личинок Anarhichas lupus lupus, выходящих из икры, диаметр глаза составляет около половины длины головы и около 0.7 ее высоты через середину глаза. Плоскости роговиц почти параллельны друг другу. Глаза выдаются вбок дальше любой из точек тела. Их верхний край находится на уровне верхнего края головы, а передний — почти на уровне ее переднего края. Поля зрения должны быть очень обширными и при движении глаз, по-видимому, охватывают все пространство сверху, спереди, с боков и снизу головы, в значительной мере даже и сзади нее. По всей вероятности, зрение у личинок служит в основном для предупреждения о приближающемся враге, который может находиться и снизу личинок в то время, когда они поднимаются над дном (Mcintosh a. Prince, 1890 : 880): для зубаток такого размера могут быть опасны даже морские звезды — их будущая жертва. Челюсти и жевательные мышцы в это время развиты очень слабо, что и делает возможным наличие такого обширного поля зрения.
В дальнейшем происходит увеличение относительных размеров челюстей и связанных с ними мышц, сопровождающееся изменениями их формы, а также и формы неврокраниума. Эти процессы происходят особенно быстро в начале онтогенеза, постепенно замедляясь к его концу. Их связь с изменениями относительных размеров, формы и расположения глаз несомненна. Вполне очевидно, что при увеличении челюстей и связанных с ними мышц, при сужении заглазничного отдела черепа сохранение прежнего размера глаз относительно всей головы и прежнего положения плоскости роговиц невозможно без коренного нарушения обтекаемости тела; значение же последней в силу увеличения подвижности возрастает. В действительности уже на первом году жизни происходит быстрое сокращение относительных размеров глаз, а также поворот их роговиц вперед и вверх. Ввиду уменьшения относительных размеров глаз возможность расширения поля зрения с помощью их движений снижается. Поля зрения резко сокращаются снизу и сзади, но увеличивается возможность бинокулярного зрения вверх и вперед. Однако особо важно направление его вперед и вниз, а не вверх и вперед, что доказывается изменением формы наружных частей глаза, включая радужину и зрачок, из круглой в продолговатую, несколько вытянутую вперед и вниз. Отсюда следует, что у Anarhichas lupus бинокулярность зрения особенно важна при добывании пищи. Такая довольно редкая форма наружных частей глаза интересна в другом отношении: она показывает, что положение окологлазничного кольца, определяемое во многом мощностью и расположением челюстей и жевательных мышц, не обеспечивает надлежащего направления поля зрения при обычной круглой форме наружных частей глаза.
3а. Anarhichas minor Olafsen х Anarhichas lupus Linne.
Lühmann, Ber. Deutsch. wiss. Komm. Meeresforsch., XIII, 4, 1954 : 310—326, 17 Abb.
В уловах, привезенных в Киль из районов к северо-западу и западу от Исландии, Люманом обнаружены рыбы, несущие ряд промежуточных черт между Anarhichas lupus lupus и Anarhichas minor — в окраске, строении черепа и зубного аппарата. В уловах из районов к северо-востоку от Исландии таких особей нет. Число позвонков, а также лучей в спинном, анальном и грудном плавниках у них не промежуточное, а крайнее, немного превышающее это число у Anarhichas minor, что рассматривается как атавизм. Передний конец мезэтмоида вздут сильнее, а межчелюстная кость резче расширяется к цоколям зубов, чем у обоих видов. В той или иной степени нарушается периодичность смены зубов, их форма и расположение. Чаще и резче всего это наблюдается у зубов, испытывающих наибольшую нагрузку, но иногда захватываются даже глоточные зубы. Встречаются также повреждения суставов — прежде всего artuculare—quadratum, степень которых прямо зависит от степени нарушений в зубном аппарате и от выраженности промежуточных черт в окраске. Люман считает этих особей гибридами Anarhichas minor х Anarhichas lupus, вероятнее всего, от скрещивания самок Anarhichas minor с самцами Anarhichas lupus. По всем промежуточным признакам гибриды стоят ближе к Anarhichas minor, чем к Anarhichas lupus, и связаны с Anarhichas minor постепенными переходами. Предполагается, что гибриды плодовиты и возможно их повторное скрещивание с Anarhichas minor, но не с Anarhichas lupus.
Подобный же экземпляр с нарушенной сменой, формой и расположением зубов, что сильнее всего было выражено на сошнике, встречен мной в Баренцевом море на 70°28' с. ш. и 32°05' в. д. 16 V 1951. Его длина 111.5 см. Форма hyomandibulare, а также верхних частей metapterigoideum и praeoperculum с левой стороны ненормальна: создается впечатление, что нёбная дуга в этом месте была когда-то сломана, изогнувшись наружу, а затем срослась.
Питание у таких особей, по Люману, затрудняется лишь в случае глубоких нарушений зубного аппарата. По упитанности они мало отличаются от родительских видов. Относительно особенностей роста ничего определенного сказать нельзя: Люман считает, что Anarhichas minor и Anarhichas lupus растут одинаково быстро, но это вряд ли верно, по крайней мере для особей из Баренцева моря.
Не исключена возможность, что в данном случае мы имеем дело не с гибридизацией, а с какой-то болезнью, сопровождающейся нарушением обмена веществ.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
2. Anarhichas lupus Linne — Обыкновенная, или полосатая зубатка (рис. 114).
Anarhichas lupus Linne, Syst. Nat., ed. X, 1758 : 247 («Oceano Septentrionalis Angliae»). — Yarrell, Hist. Brit. Fish., I, 1836 : 247 (дл. до 6 или 7 футов). — Günther, Cat. Fish., III, 1861 : 208 (частью: «european variety», строение черепа). —Steenstrup, Vidensk. Meddcl. Naturh. Foren., (1876) 1877 : 167, 200, tab. III, fig. 1 (череп, зубы, сравнение с Anarhichas minor). — Collett, Christ. Vidensk.-Selsk. Forhandl., (1879), 1, 1880 : 50, 52 (измерения). — Bean, Proc. U. S. Nat. Mus., I, 1879 : 215 (Массачусетс, Ново-Шотландск. банки, южн. Норвегия; измерения). — Smitt, Scand. Fish., I, 1893 : 232, fig. 62, pl. XII, fig. 2 (описание, биология, рис.). — Gооdea. Bean, Ocean. Ichth., 1895 : 299, fig. 269. — Gill. 1911 : 170, fig. 1, 2, 11, 12, pl. 17, 19, 23—26, 28 (рис. плечевого пояса; остальн. рис. заимствованные). — Книпович, Определитель, 1926 : 93, фиг. 58. — Есипов в изд.: Промысл. рыбы СССР, 1949 : 611, атлас табл. 187 (сводка, цветн. рис.).
Anarrhichas vomerinus Agassiz in: Stоrer, Hist. Fish. Massach., 1867 : 265, pl. 18, fig. 1 (между Бостоном и м. Энн; цит. по: Jordan a. Evermann, 1898). — Gregory, Fish Skulls, 1933 : 376, fig. 254c, 255 (рис. черепа).
Распространение, биология. Dresel, Proc. U. S. Nat. Mus., VII, 1884 : 247 (Дэвисов прол.). — Fulton, 10-th Ann. Rep. Fish. Board Scotl., 1892 : 239 (соотношение и средние размеры самцов и самок); Smith, там же : 229 (питание). — Knipowitsch, Ежег. Зоолог. муз. Акад. Наук, II, 1897 : 148 (Мурман, Белое м.; обычна). — Книпович, Эксп. научно-пром. иссл. Мурмана, I, 1902 : 64 и сл. (многочисленные местонахождения на Мурмане). — Ehrenbaum, Wiss. Meeresunters. (Helgol.), VII, 1, 1905 : 53 (о. Медвежий, юго-зап. Шпицберген). — Ehrenbaum, Eier und Larven, 1905—1909 : 91, fig. 41 (икра, личинки). — Lebоur, Northumberl. Fish. Comm. Rep. Sci. Invest., 1908 : 25 (питание). — Carr, там же : 71 (питание). — Thielemann, Wiss. Meeresunters. (Helgol.), XIII, 2, 1921 : 205, Karte 5 (Мурман). — Дерюгин, Баренц. м. по Кольск. меридиану, 1924 : 74 (Баренц. м. по Кольск. меридиану под 70°—71° с. ш.). — Bigelow a. Welsh, Fish. Gulf Maine, 1925 : 370, fig. 186—188 (зал. Мэн; биология, промысел). — Saemundsson, Synops. Fish. Iceland, 1927 : 17 (обычна вокруг Исландии). — Dunсker a. Mohr, Tierwelt Nord- u. Ostsee, XIIg, 1928 : 90, ng. 1—2 (дл. до 125 см, биология). — Ehrenhaum, Handb. Seefischer. N. Enropas, II, 1936 : 138, fig. III (биология, промысел). — Есипов, Промысл. рыбы Баренц. м., 1937 : 50, фиг. 15 (на восток до Гусиной банки). — Берг, Рыбы Финск. зал., 1940 : 26 (Финск. зал., на восток до о. Рюгена и Борнгольма). — Andеrssоn, Fiskar Fiske Norden, I, 1942 : 103, pl. 29 (цветн. фотогр.). — Macлов, Тр. Полярн. научно-иссл. инст. морск. рыбн. хоз. океаногр., 1944 : 155 (темп роста, плодовитость). — Saemundsson, Zool. Iceland, IV, 72, 1949 : 31 (биология).
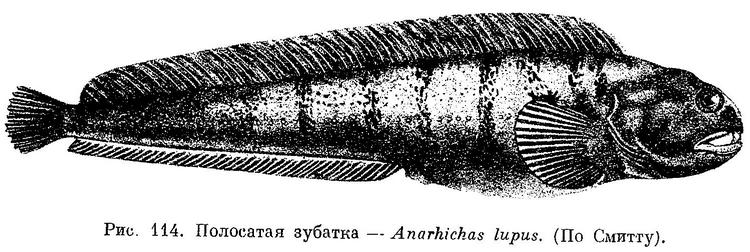
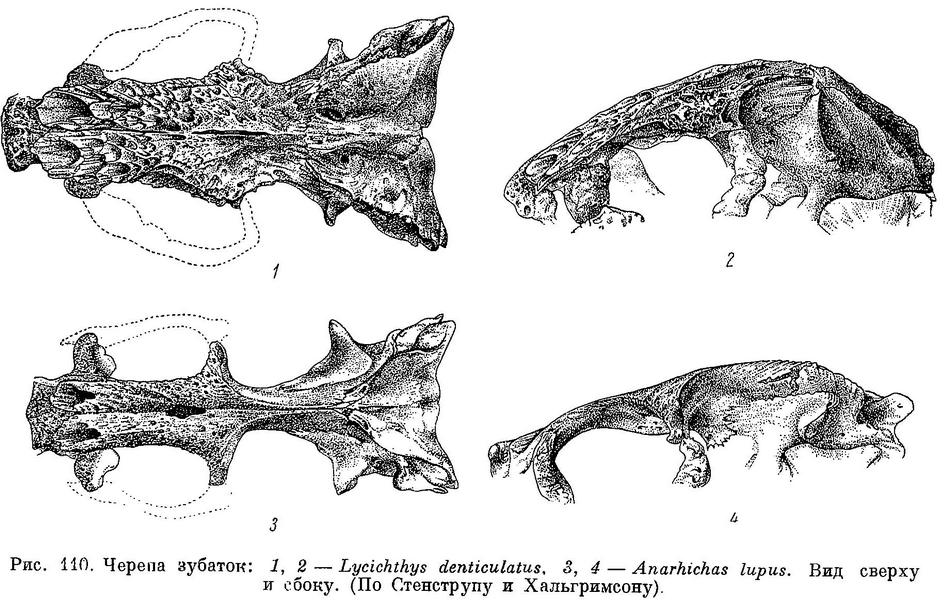
D 69—79, А 42—48, Р 18—20, позвонков 74—76.
Зубы более массивные и уплощенные, чем у других видов. Сошниковый ряд зубов простирается назад значительно дальше нёбных; его длина в 1.5—1.75 раза превышает длину нёбного ряда зубов. Задние (колючие) лучи спинного плавника обычно равномерно укорочены. Грудной плавник сравнительно большой, 13—16% абс. длины тела. По внешнему ппду полосатая зубатка напоминает пятнистую, но тело ее окрашено и коричневато- или голубовато-серый цвет с 9—12 темными поперечными полосами, состоящими из частично сливающихся мелких черных пятен, переходящих и на спинной плавник. Беломорские зубатки, возможно, представляют особую форму. Длина в Баренцовом море до 102 см и вес до 6.5 кг, чаще встречаются экземпляры 30—70 см длиной и весом до 4 кг. У берегов Норвегии и в Северном море достигает длины до 120— 125 см, у берегов Канады изредка до 1.5 м , и веса 13.5 кг. Часто встречающиеся в литературе указания (Книпович и др.) на большие размеры (до 180 см и более) основаны, повидимому, на неточных данных старых авторов.

Распространение. Северная часть Атлантического океана и прилежащие к северу моря. Широко распространена в Баренцовом море, хотя уступает здесь по численности другим видам зубаток. Обычна в Белом море. В Баренцовом море на восток доходит до Гусиной банки, а в северо-западной части моря — до о. Медвежьего и юго-западных берегов Шпицбергена. Имеет более широкое распространение, чем другие атлантические виды, доходя на юг до северо-западных берегов Франции. Обычна у берегов Великобритании и Ирландии, в Северном море и у берегов Норвегии; в Балтийском море встречается реже, но отдельные экземпляры доходят до Финского залива. Вокруг Исландии, у Гренландии (Дэвисов пролив); обычна у американских берегов на юг до м. Код и изредка до Нью-Джерси.
Биология. Живет на умеренных глубинах от 15—20 до 150—200 м или немного глубже. В Баренцовом море встречается до глубины 200—260 м и очень редко глубже (до 350 м). Чаще попадается на каменистых грунтах и при положительных температурах. Плодовитость в Баренцовом море колеблется от 2 560 до 23 580 икринок [по 12 экз. длиной 43—81 см (Маслов); для английской зубатки указывается большая плодовитость]. Нерест в европейских водах и у берегов Сев. Америки с ноября по январь. Икра откладывается на значительной глубине между камнями в виде крупных шаров из слипшихся желтоватых икринок диаметром 5.5—6.0 мм (жировая капля 1.75 мм). Выклевывающиеся через 2—3 месяца личинки длиной около 12 мм имеют большой желточный пузырь, который рассасывается лишь через 3.5 месяца (по аквариальным наблюдениям), когда личинки достигают 17—20 мм. До рассасывания желточного пузыря личинки живут на дне и лишь при длине 20—44 мм обитают в толще воды, редко, однако, встречаясь близ поверхности. К концу первого года мальки достигают 10—15 см. Дальнейший их рост в Баренцовом море, по данным П. 3. Сахно (Маслов), показан ниже:

Питание в основном состоит из моллюсков, раковины которых обычно раздробляются мощными зубами (Buccinum, Chrysodomus, Pecten, Cardium, Mactra, Fusus, Astarte и мн. др.); поедает также иглокожих (ежи, офиуры, звезды) и донных ракообразных (Hyas, Eupagurus, Crangon и др.). Рыбы в желудках полосатой эубатки встречаются сравнительно редко.
Хозяйственное значение. Второстепенная промысловая рыба европейского севера. Промышляется главным образом в Северном море и прилежащих водах, а также у берегов Исландии и Сев. Америки. В Баренцовом море по величине уловов значительно уступает пятнистой зубатке. Общий улов европейских стран не превышал 186 тыс. ц. Мясо полосатой зубатки отличается хорошими вкусовыми качествами и употребляется в пищу в соленом, копченом й свежем виде. Используется также икра и шкура.