Trachurus japonicus (Temminck & Schlegel, 1844)
Японская ставрида, китайская популяция японской ставриды, корейская популяция японской ставриды (Rus),
Japanese jack mackerel
(Eng)
Синонимы:
Caranx trachurus japonicus Temminck & Schlegel, 1844
Trachurus argenteus Wakiya, 1924
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Trachurus japonicus (Temminck et Schlegel) - японская ставрида (рис.16)

Описание. D I,VIII, I+30-34, наичаще 30-32, в среднем 31,1; A II, I+26-31, наичаще 27-29, в среднем 27,3. L.l. 69-80, наичаще 70-73, в среднем 72,1. Голова составляет 23,6-30,0% длины тела. Верхняя (спинная) ветвь боковой линии оканчивается под основанием первого-четвертого ветвистого луча второго спинного плавника. Жаберных тычинок на первой дуге 48-54, в среднем 52,0; на нижней части дуги 35-41, в среднем 39,0. Щитки боковой линии очень крупные. Высота наиболее высоких щитков передней (кривой) части боковой линии 21,6-30,0, в среднем 24,4% длины головы; высота наиболее высоких щитков задней (прямой) части боковой линии 22,7-29,2, в среднем 25,6% той же длины. Наиболее высокие щитки задней части боковой линии обычно несколько выше щитков в ее передней части, или равны им по высоте, или, реже, слегка ниже их. Кили на щитках хорошо развиты, шипы на килеватых щитках крепкие. Прямая часть боковой линии, несущая киль, длинная; хорда дуги кривой части боковой линии 1,3-1,6 раза укладывается в длине ее прямой части. Ответвления каналов боковой линии на верхней стороне головы и на спине хорошо развиты. Тело сравнительно высокое, наибольшая его высота 20,2-25,8, в среднем 22,5% длины тела. Глаз крупный, его диаметр 26,1-33,1, в среднем 30,1% длины головы. В процентах длины тела: расстояние от вершины рыла до вертикали начала основания брюшных плавников 26,3-32,0, в среднем 28,9; расстояние от вершины рыла до вертикали начала основания первого спинного плавника 29,5-32,6, в среднем 31,1; расстояние от конца рыла до вертикали основания первого луча анального плавника 46,0-50,0, в среднем 47,6. Последние лучи второго спинного и анального плавников увеличены сравнительно с предпоследними и отставлены от остальной части плавника; расстояние между основаниями последнего и предпоследнего лучей анального плавника в 1,2-1,8 раза превосходит расстояние между основаниями предпоследнего и предшествующего ему лучей того же плавника. Грудные плавники сравнительно длинные, у взрослых заметно заходят за вертикаль основания первого луча анального плавника. Спина темная, бока и брюхо серебристые; грудные и спинные плавники темно-серые; хвостовой - желто-серый, прочие плавники светлые, с желтым оттенком; на жаберной крышке черное пятно. Длина до 45 см [Алеев, 1957а].
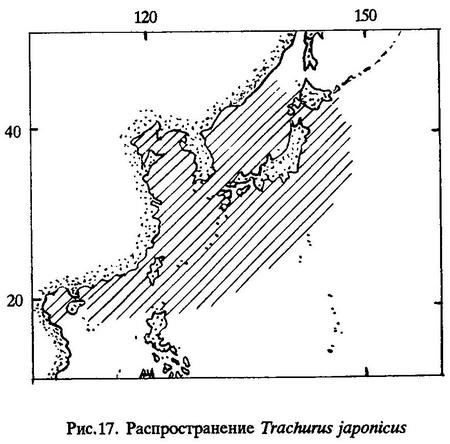
Распространение. Японская ставрида - теплолюбивый субтропический вид, ведущий пелагический образ жизни. Ее ареал охватывает Японское, Желтое, Восточно-Китайское моря и воды Тихого океана у берегов Южной Японии, от Приморья до Гонконга и Тайваня. Однако, насколько известно, наиболее многочисленна эта ставрида в Восточно-Китайском море и в прибрежных районах Южной Японии и п-ва Корея [Wakiya, 1924; Солдатов, Линдберг, 1930; Таранец, 1937; Линдберг, 1947; Румянцев, 1947] (рис.17).
Сравнительные замечания. Описанная из японских вод Trachurus argenteus Wakiya является синонимом Trachurus japonicus (Temminck et Schlegel). Согласно Вакии [Wakiya, 1924], эта форма отличается от близких форм Trachurus иным соотношением диаметра глаза и длины предглазничного расстояния, а также тем, что у нее щитки кривой части боковой линии по высоте равны щиткам прямой ее части. Однако соотношение диаметра глаза и предглазничного расстояния у видов рода Trachurus заметно меняется с возрастом, так как по мере роста диаметр глаза относительно уменьшается, а предглазничное расстояние, наоборот, относительно, увеличивается, что видно из данных, которые приводит Вакия.
Следует также заметить, что этот признак вообще описан Вакией неудовлетворительно. Так, в определительной таблице говорится, что у Trachurus japonicus диаметр глаза длиннее рыла, а в описании этого вида указано обратное. О Trachurus argenteus сказано, что длина рыла равна диаметру глаза или слегка превосходит его, тогда как из приведенных в таблице данных видно, что у крупных рыб она явно больше диаметра глаза, но бывает и меньше его или равна ему. Ю.Г.Алеев (1957а) установил, что у мелких особей Trachurus japonicus диаметр глаза может быть равен длине рыла или даже превосходить ее. У более крупных экземпляров диаметр глаза, как правило, короче длины рыла. Таким образом, следуя Вакии (1924), для разграничения Trachurus japonicus и Trachurus argenteus мы должны крупных рыб отнести к Trachurus argenteus, а сеголетков - к Trachurus japonicus. Не случайно В.К.Солдатов и Г.У.Линдберг (1930), использовавшие определитель Вакии, смогли отнести к Trachurus japonicus только малька длиной 7 см, а более крупный экземпляр вынуждены были посчитать за Trachurus argenteus. Что касается соотношения высоты щитков в кривой и прямой частях боковой линии, то у просмотренных нами 41 экз. Trachurus japonicus первые или ниже последних, или равны им по высоте, или, реже, несколько выше их. Следует отметить, что у некоторых других видов рода Trachurus, например, у Trachurus mediterraneus, Trachurus trachurus, Trachurus delagoa, Trachurus picruratus также встречаются особи, у которых щитки прямой части боковой линии по высоте равны щиткам ее кривой части, как это указано Вакией (1924) для Trachurus argenteus. Это говорит о том, что различие в высоте щитков в передней и задней частях боковой линии не может иметь диагностического значения.
Таким образом, отличительные признаки, указанные Вакией для Trachurus argenteus, частью имеют характер возрастных отличий (соотношение диаметра глаза и предглазничного расстояния), частью же не связаны с возрастом (соотношение высот щитков прямой и кривой частей боковой линии), но свойственны нескольким формам рода.
Другой вид, указанный Вакией [Wakiya, 1924] для японских вод - Trachurus declivis (non Jenyns, 1842), также является синонимом Trachurus japonicus (Temminck et Schlegel). В качестве Trachurus declivis Вакия описал, по-видимому, высокотелые экземпляры Trachurus japonicus: такие высокотелые экземпляры, у которых длина головы равна высоте тела, имеются в коллекциях Зоологического института РАН (N 22923). Что же касается высоты щитков боковой линии, то эти признаки у видов Trachurus, как уже было указано выше, значительно варьируют. Число щитков в боковой линии для Trachurus declivis указано 75-76. Но у Trachurus japonicus, как известно, оно варьирует от 69 до 80. Однако такой важный диагностический признак, как место окончания спинной ветви боковой линии, Вакия оставил без внимания.
А.Б.Стефенсон и Д.А.Робертсон [Stephenson, Robertson, 1977] показали идентичность новозеландской ставриды Trachurus novaezelandiae (прежнее название Trachurus maccullochi) и Trachurus japonicus, причем последнее название нужно считать младшим синонимом, ими переведенным в синоним Trachurus novaezelandiae.
Биология. Японская ставрида обитает преимущественно в шельфовой зоне. В Восточно-Китайском море находится в зависимости от ее размеров: мелкая ставрида длиной 15-23 см, придерживается относительно холодных вод, средняя и крупная (свыше 23 см) - в более теплых водах; самые крупные особи ловятся на юге ареала. Размерный состав ставриды даже в одном скоплении неоднороден. В японских уловах она представлена особями длиной от 4 до 41 см [Nakashima, Hotta, 1976].
Японская ставрида обитает в широком диапазоне температуры воды на поверхности - от 14° до 22°С, однако оптимальная температура, при которой наблюдаются наибольшие уловы, находится в пределах 16-18°С.
Японская ставрида обычно держится у дна, но во время интенсивного питания может подниматься к поверхности воды. Подобно другим теплолюбивым рыбам, она совершает сезонные миграции в меридиональном направлении, перемещаясь летом на север для нереста и нагула. В заливе Петра Великого до 1938 г. эта ставрида ежегодно наблюдалась с середины июня до середины октября и служила предметом небольшого промысла. С 1938 до 1946 г. она здесь не появлялась, что было связано, по-видимому, с похолоданием в этот период Японского моря. В эти же годы отмечено отсутствие у берегов Приморья и многих других теплолюбивых рыб, которые до 1938 г. были здесь обычны.
У берегов Японии ставрида ловится главным образом весной и осенью, т.е. во время весенней и осенней миграции. Зимует она в водах Южной Японии. Икра выметывается в поверхностных слоях. Плодовитость японской ставриды до 100 тыс. икринок. Икра пелагическая, с жировой каплей. Сеголетки в Японском море осенью (в сентябре) имеют длину от 5,6 до 11,0 см, что свидетельствует о растянутости нереста. Годовалые особи японской ставриды в Японском море имеют длину в среднем около 9-12 см, двухгодовалые - около 14-16 см, трехгодовалые - около 16-18 см. Встречаются экземпляры в возрасте до 12 лет; в этом возрасте экземпляр из Желтого моря имел длину 38,5 см. Длина самых крупных особей - до 42 см. В промысловых уловах обычны рыбы длиной 18-30 см. Молодь питается зоопланктоном, личинками рыб, креветками и др.[Ермаков, 1986].
В летний период косяки ставриды мигрируют из Восточно-Китайского моря в Желтое и Японское моря вдоль берегов до залива Ольги и о-ва Хоккайдо. Наиболее устойчивые скопления этой рыбы ежегодно наблюдаются в районе Корейского пролива и о-вов Гото (рис.18).
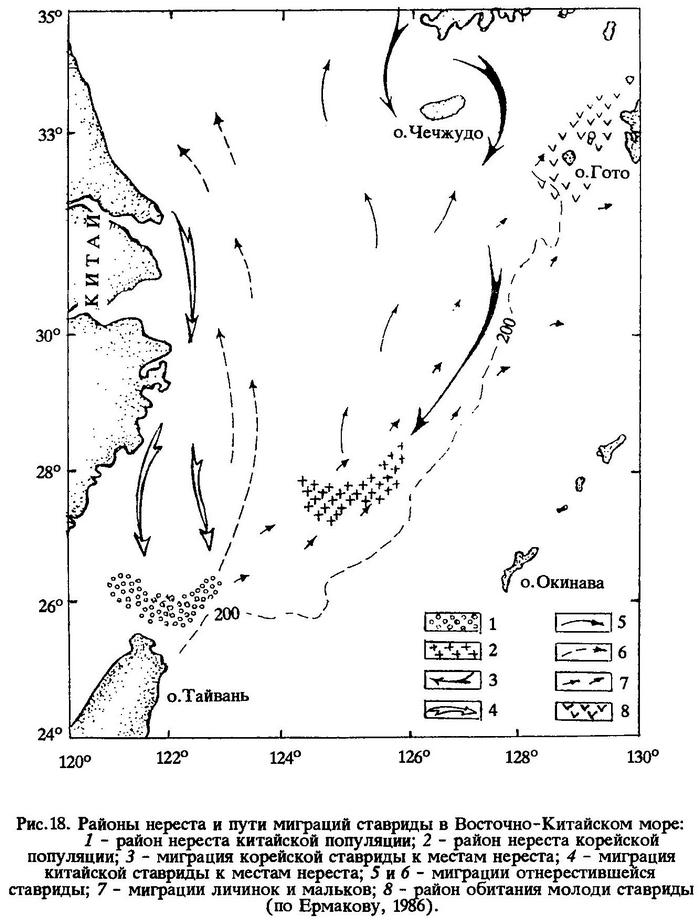
В результате отечественных исследований в Восточно-Китайском море выделены две популяции ставриды - китайская и корейская, существование которых связывают с двумя центрами нереста и различиями в его сроках. Корейская популяция из районов мелководья у п-ова Корея и о-ва Чечжудо совершает нерестовую миграцию в центральную часть Восточно-Китайского моря. С охлаждением шельфовых вод перемещается на юг, придерживаясь присвальной части шельфа. Другая популяция японской ставриды - китайская - нерестится значительно южнее [Чигиринский, 1969]. С началом охлаждения шельфовых вод она мигрирует вдоль побережья Китая в юго-западную часть моря, где нерестится в январе - марте (пик нереста приходится на конец января - начало февраля). Основной район нереста - воды вблизи о-ва Тайвань у входа в Тайваньский пролив. Выклюнувшиеся личинки выносятся течением к юго-западным берегам Японии.
Корейская популяция ставриды нерестится в центральной части моря между 27° и 29°с.ш. Пик нереста наблюдается в апреле, а в целом нерест продолжается с марта по июнь. Миграции ставриды к нерестилищам в центральную часть моря происходят вдоль материкового склона, где охлаждение вод до оптимальных нерестовых температур благодаря влиянию Куросио происходит значительно позже, чем в западной части моря. Личинки выносятся течением к юго-западному побережью Японии.
Отнерестившаяся ставрида китайской популяции с потеплением воды мигрирует в район нагула на север вдоль западного побережья Восточно-Китайского моря. Ставрида корейской популяции мигрирует после нереста в северном и северо-восточном направлении к берегам Кореи, в район Цусимского пролива, к о-вам Чечжудо, Гото, Дандзе, где происходит ее нагул.
Молодь обеих популяций держится в одном районе - у о-вов Гото и Дандзе. Этот район характеризуется высокой биологической продуктивностью, что обеспечивает благоприятные кормовые условия для молоди. По мере созревания молодая ставрида мигрирует к западному берегу Восточно-Китайского моря и в северную часть ареала вдоль Японских островов. Считают возможным [Новиков, 1986], что отдельная популяция ставриды обитает, по всей видимости, и у южного тихоокеанского побережья Японии. Судя по крайним южным местам обнаружения ее личинок в Тихом океане, ареал японской ставриды ограничивается на юге 30°с.ш. Массовый нерест происходит в шельфовой зоне у южного побережья о-вов Хонсю, Сикоку, Кюсю (в среднем около 33°с.ш.). Мигрирующие вдоль тихоокеанского побережья о-ва Хонсю скопления японской ставриды достигают на севере прибрежья префектуры Аомори (40°с.ш.), где и держатся до времени их обратного перемещения на юг. В отдельные годы молодь японской ставриды наблюдается вблизи о-ва Кунашир (44°с.ш.). Таким образом, ареал японской ставриды простирается от 30° до 44°с.ш., а протяженность сезонных перемещений нерестовой части популяции ограничена 33-41°с.ш. [Новиков, 1986].
Японские исследователи [Hotta, Nakashima, 1971] на основании анализа строения чешуи и отолитов различают у западного побережья Японии не менее трех популяций японской ставриды. Ими установлено, что период максимума нереста четко дифференцирован для каждой популяции: для позднезимненерестующей - февраль-март, весенненерестующей - апрель-май, зимненерестующей - январь, причем нерестилища каждой популяции обособлены.
Для изучения возраста и роста были проанализированы отолиты от 6173 рыб [Nakashima, 1982]. При определении возраста лишь третье кольцо принимали за годовое, а затем каждое кольцо учитывали как событие "год жизни". Получены следующие результаты:

В опытах по выращиванию японской ставриды в аквакультуре к концу первого года жизни рыбы достигали в среднем длины 17,4 см и массы 67 г [Ochiai et al., 1983].
Соотношение полов в преднерестовых скоплениях японской ставриды примерно 1:1. Половозрелой она становится на втором году жизни при длине 16-18 см [Семенов, 1966].
Японская ставрида относится к рыбам с асинхронным ростом ооцитов, обеспечивающим созревание нескольких порций икры. Это в свою очередь обусловливает растянутость нереста. В Восточно-Китайском море нерест ставриды, по многолетним наблюдениям, начинается в январе [Чигиринский, 1964; Семенов, 1966]. В водах о-ва Кюсю (южное и тихоокеанское побережья) на протяжении всего года можно встретить особей ставриды со зрелыми гонадами, но пик нереста приходится здесь на март-май [Чигиринский, 1969].
Обращает на себя внимание то обстоятельство, что у всех ставрид рода Trachurus, несмотря на порционность икрометания, в той или иной мере выражен пик размножения, т.е. наблюдается период наиболее массового нахождения икры в местах нереста даже в тех случаях, когда икрометание происходит в течение длительного времени. Такое явление А.И.Чигиринский объясняет выметыванием одной резко доминирующей, обычно первой - порции икры в наиболее благоприятный период.
В Восточно-Китайском море средняя плодовитость японской ставриды длиной 30,5-31,0 см составляет 191,5 тыс.икринок, длиной 34,5-37,5 см - 280 тыс.икринок [Чигиринский, 1970].
Икра японской ставриды пелагическая. Диаметр икринок от 0,8 до 1,0 мм, диаметр жировой капли - от 0,2 до 0,3 мм. Развитие эмбриона длится примерно 40 ч. при температуре воды 20°С. Только что выклюнувшиеся личинки имеют длину 2,3-2,5 мм. На четвертый день после выклева длина личинок достигает 3 мм. В течение последней постличиночной стадии формируются первый спинной и брюшные плавники, тело удлиняется, достигая в среднем длины 13 мм на 29-й день после выклева.
После достижения мальковой стадии преоперкулярные колючки становятся меньше и почти исчезают к концу этой стадии. Мелкие чешуйки развиваются вдоль латеральной линии при достижении длины примерно 19 мм. Чешуя на остальных участках тела развивается у особей, достигших длины более 26,5 мм. На поздней стадии малька формируются лучи в грудных и хвостовом плавниках. Мальковая стадия заканчивается при достижении длины около 55 мм через 60 дней после выклева [Ochiai et al., 1982, 1983].
В составе пищи личинок ставриды науплии копепод и копеподиты длиной менее 1 мм составляют до 95 %. Основной пищей взрослых особей японской ставриды служит мезо- и макропланктон, из которого в наибольшем количестве потребляются копеподы, эуфаузииды, гиперииды, а также личинки и молодь рыб, мелкие рыбы и кальмары. В целом в питании японской ставриды отмечено около 60 видов планктонных беспозвоночных организмов и несколько видов рыб и кальмаров. В суточном ритме питания ставриды наблюдаются два пика. Наиболее интенсивно питание днем в 12-14 ч. и вечером в 19-24 ч. Существенных различий в составе пищи рыб на разных стадиях зрелости и у разноразмерных особей не отмечено. Однако по мере роста ставриды объектами ее питания становятся все более крупные организмы. Во время нереста ставрида не прекращает питаться, хотя интенсивность питания в этот период снижается [Kozasa, 1974].
Хозяйственное значение. Японская ставрида - важная промысловая рыба. Ее вылов в северо-западной части Тихого океана в 1960-1966 гг. составлял 400-600 тыс.т. В последние годы численность ее заметно сократилась, что привело к снижению вылова до 200-300 тыс.т. Около 90% всего вылова приходится на долю Японии. Кроме Японии ставриду добывают рыбаки Южной и Северной Кореи, Китая и Тайваня. В последние годы в российском промысле японская ставрида самостоятельного значения не имеет. Чаще всего она попадается при лове скумбрии кошельковыми неводами и тралами у берегов Японии и Китая.
Основными районами образования промысловых скоплений японской ставриды является восточный проход Цусимского пролива, район о-вов Гото и центральный район (27-32°с.ш.), где ее скопления наблюдаются на глубинах до 150 м. Наилучшие уловы здесь получают в преднерестовый и нерестовый периоды, когда рыбы собираются в плотные косяки и становятся более доступными для облова кошельковыми неводами. В годы высокой численности за один замет японцы вылавливали до 100 т ставриды. Значение указанных районов промысла в разное время года не одинаково. С начала нереста ставриды и до апреля основные районы японского кошелькового промысла находятся в центральной и юго-западной частях Восточно-Китайского моря. С конца апреля скопления ставриды формируются в районе о-вов Гото, Чечжудо и в Цусимском проливе. Летом этот район становится основным, хотя промысел в нем менее результативен.
У тихоокеанского побережья Южной Японии в уловах наблюдаются два заметных пика - весной (апрель-июнь) и осенью (сентябрь-ноябрь). Промысел в первый сезон более эффективен, чем во второй, и в течение обоих сезонов базируется на рыбах длиной более 20 см.
Косяки японской ставриды обычно объединяются в скопления, вытянутые узкой полосой на многие мили вдоль течения, что следует учитывать при выполнении поисковых работ и выборе курса тралений. В частности, траления по течению считаются наиболее результативными. При поиске скоплений японской ставриды даже с помощью самых современных приборов необходимо постоянно следить за температурой воды и периодически выполнять контрольные траления. Поиск значительно эффективнее, если его вести одновременно несколькими судами. При поиске скоплений следует учитывать суточные вертикальные миграции японской ставриды, так как в дневное время часть косяков держится в нижних слоях воды или непосредственно у грунта, а ночью они поднимаются в верхние слои, хотя незначительное количество рыб и остается у дна. Однако ставрида может образовывать скопления у грунта и ночью, и, таким образом, придонные траления при поиске следует производить как в светлое, так и в темное время суток. Важной особенностью поведения японской ставриды является положительная реакция на подводные и надводные искусственные источники света. Наиболее сильно эта реакция проявляется в период нереста, затем ослабевает и в летнее время практически отсутствует. Кроме того, в летне-осенний период косяки ставриды очень подвижны и разреженны, а сама рыба пуглива; все это вместе отрицательно сказывается на эффективности промысла.
Главным, но не единственным орудием лова японской ставриды в течение всех сезонов года и во всех районах промысла служит высокостенный кошельковый невод. С ноября по февраль возможен траловый промысел ставриды в Восточно-Китайском море. Существует также ручной удебный промысел. Он ведется в основном в прибрежных водах Японии с июня до октября. Техника удебного лова основывается на привлечении ставриды искусственным источником света. Наиболее результативен промысел в темные ночи. Благоприятными районами для удебного промысла являются мелководные рифовые участки с глубиной 30-50 м [Журавлев, 1967; Nakashima, Hotta, 1976].
Подавляющая часть вылова японской ставриды приходится на долю Японии. Наибольшие уловы наблюдались в период с конца 50-х до конца 60-х годов, когда ставрида входила в число 15 основных промысловых видов, обеспечивающих свыше 80% общего вылова рыбы Японией. С 1962 по 1966 г. ее вылов был максимальным и отличался высокой стабильностью. Однако затем началось неуклонное снижение уловов, и в 1980 г. было добыто всего 56,2 тыс.т против 553,3 тыс.т в 1965 г. При этом наблюдалось и некоторое сокращение ареала ставриды. В последние годы вновь отмечена тенденция к увеличению вылова (табл.36).
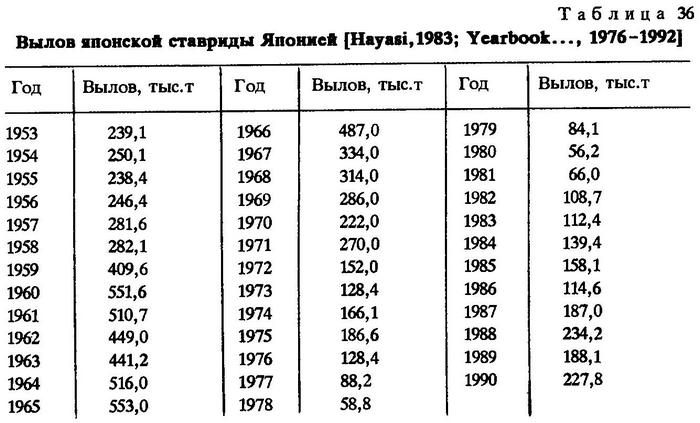
Кроме Японии промысел японской ставриды ведет Южная Корея - до 40 тыс.т в 1988 г. Промысел СССР был незначителен - 1,1-2,3 тыс.т в год, и в 1979 г. прекратился [Yearbook..., 1976, 1985, 1992].
С.Танака [Tanaka, 1983] отмечает, что изменчивость общего вылова тепловодных планктоноядных рыб в японском рыболовстве гораздо меньше, чем изменчивость вылова каждого вида в отдельности. Хотя численность отдельных видов колеблется в значительной степени, редко случается, чтобы многие виды сохраняли очень высокие или низкие уровни численности одновременно. Снижение численности одного вида обычно компенсируется увеличением численности некоторых других видов, так что продуктивность моря, очевидно, относительно постоянна. Известно, что изменчивость запасов пелагических рыб зависит от естественных изменений окружающей среды. В частности, высокий или низкий уровень пополнения определяет изменение всего запаса вида. Почти нет доказательств, что промысел играет важную роль в этой общей изменчивости. Сокращение пополнения случается и тогда, когда нерест бывает довольно эффективным. В то же время есть примеры увеличения запасов, происходящего при очень низкой численности икры на нерестилищах (Tanaka, 1983).
Японской ставриде свойственны значительные долгопериодные колебания численности. Уловы в начале 60-х годов были на уровне 0,5 млн.т. Тенденция снижения уловов стала очевидной с 1967 г.; тогда же увеличилась численность скумбрии. С.Танака (1983) считает, что ставрида является конкурентом иваси и скумбрии, поэтому в связи с наметившейся тенденцией снижения численности последних можно ожидать восстановления прежней высокой численности японской ставриды в ближайшие годы. Это предположение подтверждается значительным увеличением в последние годы численности молоди ставриды.
Содержание жира в мясе японской ставриды колеблется от 0,6 до 9,7 %, составляя в среднем 4,0%; белка - от 19 до 24%, золы - 1,4-2,2 %. Установлено, что в ноябре в более северных районах моря японская ставрида более жирная.
Окисление тканевого (подкожного) жира как мороженой, так и соленой ставриды проходит довольно интенсивно при доступе воздуха. Поэтому мороженую рыбу при хранении до трех месяцев при температуре -18°С следует обязательно глазировать; соленую рыбу надо сохранять под тузлуком.
На дегустациях крупная и средняя японская ставрида признана хорошим сырьем для приготовления кулинарных блюд и копченостей. Мясо ее - белое и нежное - при варке или обжарке и при горячем копчении обладает приятным специфическим вкусом [Мельникова, 1963].
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
В Тихом океане, у азиатских берегов,
живет японская ставрида (Trachurus japonicus),
особенно многочисленная в водах Южной
Японии, Кореи и Восточно-Китайского
моря. В осеннее время она встречается
спорадически и у берегов Приморья.
На юг она доходит до Южно-Китайского
моря.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 3. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1969 г.
1. Trachurus japonicus (Temminck et Schlegel, 1842) — Японская ставрида (рис. 202).
Caranx trachurus japonicus Temminck et Schlegel, Fauna Japonica, Poiss., 1842 : 109, Tab. 59, Fig. (Япония).
Trachurus japonicus, Wakiya, Ann. Carneg. Mus., 15, 2—3, 1924 : 144. — Солдатов и Линдберг, Обзор ..., 1930 : 120. — Алеев, Тр. Севастоп. биолог, станции, 9, 1957 : 220.
Trachurus argenteus, Солдатов и Линдберг, Обзор..., 1930 : 121.
Trachurus argenteus, Wakiya, Ann. Carneg. Mus., 15, 2—3, 1924 : 145.
Trachurus declivis, (nec Jenyns) Wakiya, 1. c. 1924 : 146.
Зал. Петра Великого. 3 X 1950. ТИНРО. 1 экз.
16961. р. Туманган. IX 1913. А. И. Черский. 1 экз.
17007. р. Туманган. 7 IX 1913. А. И. Черский. 1 экз.
17021. р. Туманган. 27 IX 1913. А. И. Черский. 1 экз.
22463. Нагасаки. 9 III 1901. П. Ю. Шмидт. 2 экз.
22923. Цуруга. 1—5 IX 1917. В. Рожковский. 5 экз.
31376. Желтое море. Дальний. 4—10 IX 1946. В. Г. Гнездилов. 1 экз.
D VIII, I 30—34; А II, I 26—31; щитков 70—75 (33—39 + 35—39); жаберных тычинок 52—54 (13—14 + 39—40). Позвонков (с уростилем) 24 (10+14) — по рентгенограммам 10 экз.
У рыб из нашей коллекции количество щитков в боковой линии часто бывает различно на правой и левой сторонах тела, но это расхождение не превышает 3.
Сведения по биологии японской ставриды приведены в ряде работ: Алеев, 1954, 1957; Чигиринский, 1964; Hotta a. Tamura, 1957; Suzuki,1965.
Длина наших экземпляров от 88 до 410 мм (абсолютная длина) и 76—360 мм без хвостового плавника.
Распространение. В Японском море изредка встречается спорадически осенью у берегов Приморья (Румянцев, 1947 : 48); зал. Петра Великого, IX 1913; зал. Ольги, IX 1925 (Солдатов и Линдберг, 1930 : 120); зал. Рында, бухта Пластун, сентябрь 1936 г. (Таранец, 1937 : 88); указывается для всех берегов п-ова Корея (Mori, 1952 : 98); по японскому побережью на север до Отару и Хакодатэ (Snyder, 1912 : 411), на юг вдоль обоих побережий Японии (Matsubara, 1955 : 555). Известен в Желтом море (Линдберг, 1949 : 186) и в зал. Вохай (Чжан и др., 1957 : 104). На юг от Японии доходит до Южно-Китайского моря (Чжу и др., 1962 : 392).
[Trachurus argenteus Wakiya, 1924).
Диаметр глаза равен или почти равен длине рыла. Наиболее высокие щитки в передней изогнутой части боковой линии равны по высоте наиболее высоким щиткам в ее задней прямой части. При жизни спина зеленоватого цвета. Юго-восточная Япония (Сикоку, Кюсю) и о. Тайвань (Wakiya, 1924 : 147). Для Японского моря не указан.